
Submitted:
19 June 2024
Posted:
19 June 2024
You are already at the latest version
Abstract
Animal personalities play a crucial role in invasion dynamics during alien species invasions. The behavioral strategies adopted by native species varied among personalities when facing an invasion by alien species, just as the invasive species exhibited variations in behavior strategies across different personalities during the invasion process. However, the effects of personality interactions between native species and invasive species on behavior and growth are rarely illustrated. We used the globally invasive freshwater turtle Trachemys scripta elegans and the native freshwater turtle Mauremys reevesii to investigate how the personality combinations of native and invasive turtles would impact the foraging strategy and growth of both species. We found that M. reevesii exhibited bolder and more exploratory personalities than T. scripta elegans. The foraging strategy of M. reevesii was mainly affected by the personality of T. scripta elegans, while the foraging strategy of T. scripta elegans was influenced by both their own personality and M. reevesii's personality. In addition, we did not found that the personality combination would affect the growth of either T. scripta elegans or M. reevesii. Difference in foraging strategy may be due to the competitive advantage of invasive species and variations in the exploration-exploitation foraging strategy among different personalities. Overall, our results reveal the mechanisms of personality interaction effects on the foraging behavior of both native and invasive species during alien species invasion and provide empirical evidence to help understand the effects of personality in invasion dynamics.
Keywords:
animal personality
; invasion dynamics
; interspecific competition
; foraging strategy
; morphology traits
; freshwater turtles
1. Introduction
The invasion of non-native species is considered a key threat to the integrity of natural habitats and ecosystems worldwide, leading to biodiversity losses [1,2]. Invasive species are known to generate severe economic and environmental harm within native ecosystems [3]. Determining the mechanisms of invasion dynamics and the ecological consequences of alien species invasion is a crucial question for themes such as invasion ecology, population ecology, and global change ecology [4]. Growing evidence shows that invaders are often not a random draw from a population [5,6,7]. Instead, invasive decisions were associated with various ecologically important phenotypic traits (e.g., morphology, physiology, behavior), which may generate variations in fitness consequences [5,7,8]. Additionally, invasive alien species have been recognized as drivers of behavioral changes in native species (e.g., foraging, defending, and growth), impacting the fitness of native species [9,10,11,12,13].
Animal personality, which refers to an individual’s variety of consistent behavioral differences across time and contexts in a broad range of taxa [14], plays a crucial role in the invasion dynamics of invasive alien species [15,16,17]. Specifically, some individuals (e.g., bolder, asocial, more explorative, and active individuals) will colonize empty landscape patches sooner, increasing the probability of an invasive population spreading further, and are more likely to successfully invade new habitats [18,19,20,21]. Moreover, invasive individuals with different personalities respond differently to native species. Some personality types consistently show a greater tendency to compete with native species than others [22]. Further, behavioral differences exist not only when invasive species respond to native species, but also when native species face biological invasion. Individuals with different personalities exhibit differences in behavioral strategies and physiological states in a post-invasion context [23,24]. However, the interaction effects between invasive species and native species in the post-invasion period have not been thoroughly studied. The behavioral and physiological change models of invasive species and native species with different personalities remain unclear.
The red-eared slider turtle (Trachemys scripta elegans) is native to the southern United States and northern Mexico. It is now becoming invasive in many countries, including China, posing a serious threat to the survival of native turtle species [25,26]. The red-eared slider is listed as one of the 100 worst invasive alien species in the world and has been reported to compete with native freshwater turtle species worldwide, leading to the extinction of their native habitats, including Mauremys sp. [26,27,28]. The Chinese pond turtle (Mauremys reevesii) was once the most abundant freshwater turtle in China [29]. However, its wild populations have significantly declined in recent decades, and it is currently considered an endangered species [30]. The competition with exotic introduced turtles, mainly the red-eared slider turtle, is one of the major factors responsible for this decline [31,32]. Previous studies have observed that the T. scripta elegans has a greater swimming speed and an advantage in feeding kinematics, such as shorter feeding times, compared to the M. reevesii [33,34]. These advantages may give the red-eared slider turtle a competitive edge in obtaining food and basking spots [26,28]. However, the interaction between the behavioral and fitness consequences of different personalities in invasive T. scripta elegans and native M. reevesii has not been fully studied.
In this study, we experimentally tested whether there were differences in behavior change and growth of two species when M. reevesii with different personalities cohabitated with different personalities of T. scripta elegans. Our aim was to investigate the interaction and influence of personality traits between invasive species and native species on behavior and fitness consequences in a post-invasion context. We also aimed to elucidate the relationships among personality, ecological and fitness consequences, and the underlying mechanisms of invasion biology.
2. Materials and Methods
2.1. Animals
Juveniles of M. reevesii and T. scripta elegans were approximately 14 months old and were purchased from a natural pond-rearing turtle hatchery in Baoying County, Jiangsu Province, China. The turtles were transported to the laboratory and reared in tanks (640 × 430 × 310 mm3) with a density of 18 individuals per tank, and aerated with tap water. All turtles were fed a commercial diet (Yahua Ltd., China) once a day at 9:00 a.m. and any remaining food pellets were removed after 1 hour. All turtles were housed at a temperature of 28 ± 1°C, and the photoperiod was based on natural sunlight. The acclimation period lasted for more than three months.
2.2. Experimental Procedures
After an acclimation period, we measured the boldness and exploration personalities of M. reevesii (n = 49) and T. scripta elegans (n = 52). And in order to make our data more objective, we selected the relatively bold-exploration (n = 16 for each species) and relatively shy-avoidance turtles (n = 16 for each species) as our focal experimental samples. Each of the four turtles were raised in a tank (520 × 380 × 230 mm³) and were marked with non-toxic paint on its shell for individual identification after an acclimatization period of 4 weeks. All turtles were living with their conspecifics and divided into 4 groups: (1) BE-BE group, where two focal bold-exploration turtles lived with two bold-exploration turtles; (2) BE-SA group, where two focal bold-exploration turtles lived with two shy-avoidance turtles; (3) SA-BE group, where two focal shy-avoidance turtles lived with two bold-exploration turtles; and (4) SA-SA group, where two focal shy-avoidance turtles lived with two shy-avoidance turtles. After acclimation, we measured the foraging behavior and morphological data of all turtles during the non-invasion stage, where focal turtles were living with two conspecific turtles. Then, two M. reevesii and two T. scripta elegans were reared together in a tank (invasion stage) according to their personality combination groups developed during the non-invasion stage, with a 4-week period of acclimation. After acclimation, we measured their foraging behavior and morphological data again to compare the variations in behavior and growth changes among species and groups.
2.3. Ethical Statement
All experiments involving animals complied with current laws on animal welfare and research in China and the Guide for the Care and Use of Laboratory Animals (8th edition). The experiments were conducted according to the standards of the Animal Ethical and Welfare Committee of Nanjing Normal University (Approval No. IACUC-20230247). We strived to maximize the health of the animals and reduce their suffering.
2.4. Personality Assay
2.4.1. Boldness
We refer to Kashon & Carlson (2018) and Roth et al. (2020) methods to test turtles’ boldness personality. We placed each turtle in the center of a 600 × 600 mm2 plastic tank and used a soft rubber rod to gently tap the shell of the turtles to ensure that the head and body retracted into the shell. Researchers then retreated to the test room and monitored the turtles via an overhead camera for 5 minutes. We recorded the time elapsed from turtles retracting into the shell to (1) head emergence, defined as extending the head from the shell such that the eyes surpassed the anterior margin of the carapace; (2) body emergence, defined as extending the limbs from the shell; and (3) movement, when the turtle began walking.
2.4.2. Exploration
To test exploration personality, we used an open field test to measure turtles’ exploration personality. The open field (600 × 600 × 250 mm³) was divided into 4 equal-sized areas. At the beginning of the test, the turtle was placed in the center of the open field and covered by a clear plastic cage for 5 minutes. Then, we removed the cage and monitored the turtles’ behavior via an overhead camera for 10 minutes. We used the total number of area crossed and distance moved during the 10-minute assay as proxies for exploratory behavior [37,38].
All tests were measured once again two weeks later to determine the consistency and repeatability of bold-exploration personality traits in turtles.
2.5. Foraging Behavior
All turtles were given half of their food pellets (approximately 0.5% of body weight) 48 hours before the experiment [39] and fasted for 24 hours before the experiment. Then, foraging behavior was measured starting at 9:00 a.m. All turtles were given approximately 3% of their body weight in food pellets. We used an overhead camera to monitor the turtles’ behavior for 60 minutes and then removed any remaining food pellets. The foraging behavior tests were conducted three times during both the non-invasion and invasion stages, with a one-week interval between each test. We recorded the latency, frequency, and duration of foraging during the experiment.
2.6. Morphology Traits
Morphological traits including body mass, carapace height, carapace width and carapace length. Body mass was measured by an electronic balance (Sartorius Model BL 1500, ± 0.1 g). Maximum carapace height, carapace straight width and carapace straight length were measured with a digital caliper (Pro skit, PR China) to the nearest millimeter.
2.7. Statistical Analysis
2.7.1. Personality
We conducted a principal component analysis (PCA) using the “ade4” package in R to derive composite bold-exploration scores for each individual. These scores were based on five performance parameters collected during the initial bold and exploration assay [40] and were utilized as individual bold-exploration personality scores. Then, the second personality assay data were imported into the PCA model to obtain the second bold-exploration score. A Markov chain Monte Carlo (MCMC) generalized linear mixed model approach (“MCMCglmm” package in R) was used to assess the repeatability [41]. The repeatability of bold-exploration personality was assessed using linear mixed models with experiment number as a fixed effect, turtle’s ID, and tanks ID as random effects. We calculated 95% credible intervals (95% CIs) to assess the repeatability by running 1,000 permutations of each test and using the posterior distributions to calculate the repeatability as: R = Vind / (Vind + Ve). Difference in five behavioral parameters between M. reevesii and T. scripta elegans were analyzed by t-test.
2.7.2. Foraging Behavior
Normality and homogeneity of variance of the foraging behavior data were examined using Shapiro-Wilk and Levene tests, respectively. Due to the non-normal distribution of foraging behavior data and the presence of zero values, all data were normalized by taking the logarithm base 10 of (x+1). We conducted a three-way repeated measures analysis of variance (ANOVA) to examine the differences in foraging latency, foraging frequency, and foraging duration of M. reevesii and T. scripta elegans under non-invasion and invasion stages, followed by Tukey’s post hoc tests. The experiment stage was included as a within-subjects factor. The species of the turtle subjects and the groups were included as two between-subjects factors. Behavioral changes between groups and species were analyzed using a two-way ANOVA followed by Tukey’s post hoc tests.
2.7.3. Morphological Traits
Normality and homogeneity of variance of the foraging behavior data were examined using Shapiro–Wilk and Levene tests, respectively. Body mass and carapace height/width ratio were analyzed across experimental stages, species, and groups using a three-way repeated measures ANOVA, followed by Tukey’s post hoc tests. Carapace height, width, and length were analyzed between experimental stages, species, and groups using a three-way repeated measures ANOVA with body mass as the covariate, followed by Tukey’s post hoc tests.
Statistical analyses were performed using the SPSS 20.0 software package. The results were expressed as the means ± SEM, and the level of statistical significance was set at p < 0.05.
3. Results
3.1. Bold-Exploration Personality
The Principal Component Analysis (PCA) reduced the number of variables to 2 components with an eigenvalue >1 (Table 1). Together, these components explain 90.11% of the total variance. The PCA results showed that PC1 explained 70.02% of the total variance. It was positively correlated with the exploration test and negatively correlated with the latency measurements of the boldness test. Therefore, we chose to retain the transformed data obtained from PC1 in our study as bold-exploration scores. Repeatability analysis revealed that the bold-exploration personality of M. reevesii and T. scripta elegans was highly repeatable. The total repeatability was R = 0.567 (95% confidence interval 0.415 - 0.677), with R = 0.540 (95% confidence interval 0.339 - 0.744) for M. reevesii and R = 0.610 (95% confidence interval 0.367 - 0.731) for T. scripta elegans.
The M. reevesii exhibited a significantly higher bold-exploration score (1.27 ± 0.14) than T. scripta elegans (-1.19 ± 0.14) (t-test; t = -8.691, p < 0.001, as shown in Figure 1a). In terms of behavioral variables, M. reevesii exhibited a higher crossing frequency (t-test; t = -5.311, p < 0.001, Figure 1b) and covered a longer distance while moving (t-test; t = -6.568, p < 0.001, Figure 1c) compared to T. scripta elegans during exploration tests. M. reevesii showed shorter head emergence latency (t-test; t = 5.887, p < 0.001, Figure 1d), body emergence latency (t-test; t = 7.624, p < 0.001, Figure 1e), and movement latency (t-test; t = 7.525, p < 0.001, Figure 1f) compared to T. scripta elegans during boldness tests.
3.2. Foraging Behavior
3.2.1. Foraging Latency
We found a significant difference between M. reevesii and T. scripta elegans in foraging latency (F(1, 184) = 29.64, p < 0.001) and groups (F(1, 184) = 5.53, p = 0.001). However, there was no interaction effect between species and groups (F(3, 184) = 2.07, p = 0.106). The foraging latency between groups of M. reevesii showed no significant difference during the non-invasion phase (F(3, 96) = 1.59, p = 0.198, Figure 2a) . However, there was a significant difference during the invasion phase (F(3, 96) = 4.95, p = 0.003, Figure 2a) , with the SA-SA group exhibiting a longer foraging latency compared to the other groups. The foraging latency between groups of T. scripta elegans showed no significant difference during the non-invasion phase (F(3, 96) = 1.15, p = 0.333, Figure 2d). However, there was a significant difference during the invasion phase (F(3, 96) = 12.51, p < 0.001, Figure 2d), with the SA-SE and SA-SA groups exhibiting longer foraging latency compared to the BE-BE and BE-SA groups. The foraging latency showed a significant difference between non-invasion and invasion stages for M. reevesii and T. scripta elegans (F(1,64) = 4.57, p = 0.037). However, there was no significant difference between groups (F(3,64) = 2.16, p = 0.102) and no significant interaction effects between species and groups (F(3,64) = 0.54, p = 0.659).
3.2.2. Foraging Frequency
The foraging frequency showed a significant difference between M. reevesii and T. scripta elegans (F(1, 184) = 45.83, p < 0.001) and among groups (F(3, 184) = 6.86, p < 0.001). There was also a significant interaction effect between species and groups (F(3, 184) = 4.98, p = 0.002). The foraging frequency between groups of M. reevesii showed no significant difference during the non-invasion phase (F(3, 96) = 0.67, p = 0.674, Figure 2b). However, there was a significant difference during the invasion phase (F(3, 96) = 4.52, p = 0.005, Figure 2b), with the BE-BE and SA-BE groups exhibiting higher foraging frequency compared to the SA-SA group. The foraging latency between groups of T. scripta elegans showed no significant difference during the non-invasion phase (F(3, 96) = 0.75, p = 0.524, Figure 2e). However, there was a significant difference during the invasion phase (F(3, 96) = 13.33, p < 0.001, Figure 2e), with the BE-BE group exhibiting a higher foraging frequency compared to the other groups. The foraging frequency changes between non-invasion and invasion stages were significantly different between M. reevesii and T. scripta elegans (F(1,64) = 5.50, p = 0.023), but there was no significant difference between groups (F(3,64) = 1.25, p = 0.300) and no significant interaction effects between species and groups (F(3,64) = 0.50, p = 0.684).
3.2.3. Foraging Duration
The foraging duration showed a significant difference between M. reevesii and T. scripta elegans (F(1, 184) = 63.53, p < 0.001) as well as among groups (F(3, 184) = 7.71, p < 0.001). Additionally, there was a significant interaction effect between species and groups (F(3, 184) = 2.82, p = 0.040). The foraging duration between groups of M. reevesii showed no significant difference during the non-invasion phase (F(3, 96) = 0.31, p = 0.815, Figure 2c). However, there was a significant difference during the invasion phase (F(3, 96) = 4.62, p = 0.005, Figure 2c), with the BE-BE and SA-BE groups having a longer foraging duration compared to the BE-SA group. The foraging duration between groups of T. scripta elegans showed no significant difference during the non-invasion phase (F(3, 96) = 0.49, p = 0.692, Figure 2f). However, there was a significant difference during the invasion phase (F(3, 96) = 9.16, p < 0.001, Figure 2f), with the BE-BE group exhibiting a shorter foraging duration compared to the SA-BE and SA-SA groups. The foraging duration changes between non-invasion and invasion stages were significantly different between M. reevesii and T. scripta elegans (F(1,64) = 10.97, p = 0.002), but there was no significant difference between groups (F(3,64) = 2.51, p = 0.068) and no significant interaction effects between species and groups (F(3,64) = 0.20, p = 0.099).
3.3. Morphology Traits
3.3.1. Body Mass
The body mass showed a significant difference between species (F(1, 56) = 23.89, p < 0.001, Figure 3a), with the body mass of M. reevesii being lower than that of T. scripta elegans. But there was no significant difference between groups (F(3, 56) = 0.87, p = 0.464, Figure 3a), and no interaction effect between species and groups (F(3, 56) = 0.47, p = 0.706, Figure 3a). Body mass showed a significant difference over time (F(1, 56) = 6.59, p = 0.013, Figure 3a), and there was an interaction effect between time and species (F(1, 56) = 7.11, p = 0.010). However, there were no interaction effects between time and groups (F(3, 56) = 0.41, p = 0.748), or among time, species, and groups (F(3, 56) = 0.76, p = 0.524).
3.3.2. Carapace Length
The carapace length showed a significant difference between species (F(1, 55) = 65.79, p < 0.001, see Figure 3b), with the carapace length of M. reevesii being shorter than that of T. scripta elegans. But there was no significant difference between groups (F(3, 55) = 1.03, p = 0.388, see Figure 3b) and no interaction effect between species and groups (F(3, 55) = 0.41, p = 0.747, see Figure 3b). The carapace length showed a significant difference among time points (F(1, 55) = 6.42, p = 0.014, Figure 3b). There was also an interaction effect between time and species (F(1, 55) = 5.42, p = 0.024), but no interaction effects were observed between time and groups (F(3, 55) = 0.41, p = 0.745), or among time, species, and groups (F(3, 55) = 2.36, p = 0.081).
3.3.3. Carapace Width
The carapace width showed a significant difference between species (F(1, 55) = 586.36, p < 0.001, Figure 3c), with the carapace length of M. reevesii being shorter than that of T. scripta elegans. But there was no significant difference between groups (F(3, 55) = 1.11, p = 0.354, Figure 3c), and there was no interaction effect between species and groups (F(3, 55) = 0.14, p = 0.934, Figure 3c). The carapace width showed a significant difference over time (F(1, 55) = 13.37, p = 0.001, see Figure 3c), with no interaction effects observed between time and species (F(1, 55) = 0.40, p = 0.532), time and groups (F(3, 55) = 0.43, p = 0.734), or time, species, and groups (F(3, 55) = 2.24, p = 0.094).
3.3.4. Carapace Height
The carapace height showed a significant difference between species (F(1, 56) = 34.83, p < 0.001, Figure 3d), with the carapace length of M. reevesii being shorter than that of T. scripta elegans. But there was no significant difference between groups (F(3, 55) = 1.66, p = 0.185, Figure 3d), and there was no interaction effect between species and groups (F(3, 55) = 1.27, p = 0.292, Figure 3d). The carapace height showed no significant difference over time (F(1, 55) = 2.74, p = 0.103, Figure 3d). Additionally, there were no interaction effects observed between time and species (F(1, 55) = 2.25, p = 0.139) or between time and groups (F(3, 55) = 1.06, p = 0.372). There were interaction effects between time, species, and groups (F(3, 55) = 3.36, p = 0.025) The carapace height of the BE-BE group was significantly higher than that of the SA-BE group at the non-invasion stage in M. reevesii (p = 0.014).
3.3.5. Carapace Height/Width Ratio
The carapace height/width ratio showed a significant difference between species (F(1, 56) = 794.70, p < 0.001, Figure 3e), with the carapace length of M. reevesii was shorter than that of T. scripta elegans, and there was a significant difference between the groups (F(3, 56) = 3.85, p = 0.016, Figure 3e), but no interaction effect was observed between species and groups (F(3, 56) = 0.70, p = 0.557, Figure 3e). The carapace height/width ratio showed a significant difference over time (F(1, 56) = 6.65, p = 0.013, Figure 3e), with no interaction effects between a time and species (F(1, 56) = 0.95, p = 0.333), time and groups (F(3, 56) = 0.77, p = 0.518), or time, species, and groups (F(3, 56) = 1.39, p = 0.256).
4. Discussion
Animal personality could affect the behavioral and ecological strategies during the invasion process of both invasive alien species and native species [15,16,17,42]. Variation in personality affects the strength of selection and the evolution of a species [43,44,45]. Thus, behavioral variations among native species and invasive alien species may be due to their differences in personalities. The results of this study show that the native species M. reevesii exhibited bolder and more exploratory behavior (1.27 ± 0.14) compared to the invasive species T. scripta elegans (-1.19 ± 0.24), which remained consistent over time. M. reevesii moved for longer distances and entered different zones more frequently during the exploration test. Additionally, it exhibited lower hiding behavior in response to modeled predation risk during the boldness test compared to T. scripta elegans. This variation may be due to the potential predator risks in their original habitat. Both M. reevesii and T. scripta elegans face predation by terrestrial mammals and birds. However, for T. scripta elegans, another significant threat comes from aquatic predators, such as caimans and crocodiles [46]. Therefore, it may be safer for M. reevesii to emerge from the shell earlier and escape to water because there was no aquatic predation risk in their original habitat [47]. Furthermore, this variation may also impact the difference in body size between the two species. Turtles exhibited a size-dependent hiding response, with larger individuals spending more time concealed inside their shell after an attack before reappearing [48], This behavior is attributed to the higher costs that small individuals face when hiding, including missed foraging opportunities and increased thermal expenses. Small turtles cannot afford the metabolic expenditure associated with prolonged hiding, as it would detract from foraging time and lead to greater thermal costs due to a faster thermal exchange rate [49,50]. Therefore, the M. reevesii exhibited bolder and more exploratory personalities than T. scripta elegans. This difference may be an ecological and physiological adaptation consequence and could be crucial for species fitness.
Personality has been well established to have fitness consequences and to influence important life-history decisions, such as foraging strategies [51]. Our results indicate a personality-related variation in foraging behavior between two turtle species. The bolder and more exploratory M. reevesii started foraging sooner, with a higher frequency and longer durations, compared to the shyer and avoidance T. scripta elegans. Earlier and more frequent foraging may help bold-exploration individuals find more food, but it may also increase the duration of foraging and expose animals to predators [52,53]. The foraging behavior variation between M. reevesii and T. scripta elegans may result from a trade-off between potential predator risk and physiological requirements. The individuals with poorer body condition may have higher energy requirements for maintenance, which necessitates putting more effort into foraging and showing a higher motivation for it [54,55,56]. On the other hand, the lower foraging effort may indicate a higher foraging efficiency of T. scripta elegans. Foraging efficiency is known to be a major determinant of individual fitness. Higher foraging efficiencies mean individuals can gain enough energy in a shorter time, reducing the time exposed to predator risks [57,58]. The larger body size (e.g., gape size) and advantages in feeding kinematics, such as faster movement ability, shorter gape cycle time, and neck retraction time during feeding, would benefit T. scripta elegans in completing feeding more quickly and gaining an advantage in competition with other species [33,56,59].
We found that the effects of biological invasions on foraging behavior were more pronounced for native species compared to invasive species. This may be due to losing in interspecific competition with M. reevesii. Winning competitions would presumably be an attempt to increase the winner’s profit and reduce the overall foraging success of their competitors [60]. Although competition would increase the energy cost for both species, the losers have much lower energetic gains than the winners [60,61]. As a result, losers typically choose avoidance strategies that reduce foraging effort in forage competition as a way to avoid the high energetic costs and low gains of losing a competition [56,62].
Moreover, we found that the foraging behavior changes were also correlated with their personality types. Although there were no variations in foraging behavior among groups in both M. reevesii and T. scripta elegans during the non-invasion stage. During the invasion stage, both M. reevesii and T. scripta elegans decreased their foraging effort. For the native species M. reevesii, all groups showed increased foraging latency and decreased foraging frequency and duration. Interestingly, we found that the personalities of T. scripta elegans had a greater influence on foraging behavior compared to the personalities of M. reevesii. The shy-avoidance T. scripta elegans significantly reduced the foraging effort of M. reevesii compared to the bold-exploration ones. However, the foraging behavior of T. scripta elegans was influenced by both their own personality types and M. reevesii personality types. Shy-avoidant T. scripta elegans exhibited increased foraging latency and longer foraging duration, as well as decreased foraging frequency compared to bold-exploration individuals. Additionally, coexisting with shy-avoidant M. reevesii led to longer foraging latency and duration, and lower foraging frequency compared to coexisting with bold-exploration M. reevesii. This variation may be related to the competitive advantages and the exploration-exploitation foraging strategies among personalities [56,63,64,65]. Bolder and more exploratory individuals typically adopt an exploration foraging strategy, which involves exploring relatively “superficially” when foraging, while shyer and more avoidant individuals usually opt for an exploitation foraging strategy, which involves exploring relatively “thoroughly” when foraging [64,66,67,68]. The exploitation foraging strategy may increase the rate of competition for the same food resource due to their thorough foraging. This exploitation strategy may bring higher competitive pressure than exploration individuals [68]. In the context of interspecific competition, the loser needs to endure larger costs and much lower gains than the winner during foraging competition. The food available for the loser is lower when competing with shy-avoidance competitors compared to bold-exploration competitors [61,64,69,70]. From a net energy gain perspective, reducing the investment in foraging effort is a better trade-off for both bold-exploration and shy-avoidance losers. The increased competitive pressures from ‘exploitation’ (shy-avoidance) competitors may further decrease the foraging effort of the losers. Of course, winners also need to bear the costs of competition, such as a lower net rate of energy gain from foraging, which requires them to prolong the time spent foraging compared to when there are no inter-specific competitors [61,71]. Due to the intense competition and lower net energy gain rate associated with the exploitation strategy, shy-avoidant individuals need to spend much more time foraging compared to bold-exploratory individuals. Additionally, when competing with shy-avoidant individuals, the foraging time is significantly longer than when competing with bold-exploratory individuals. As competition intensifies, the victor would spend a significant amount of time expelling the losers from the food resource [61,72,73]. As a result, the shy-avoidance winners would reduce the frequency of foraging more than the bold-exploration winners, and they would also reduce the frequency of foraging more when competing with shy-avoidance competitors than with bold-exploration competitors.
Our results indicate that personality could affect inter-specific competition during invasion dynamics as personality drives foraging strategies. The personality types of native species and invasive species have an interaction effect on the foraging strategy of both species and may play a crucial role in invasion speeds and dynamics. Although our study was carried out in a relatively short time, the long-term personality effects in the relationship between native species and invasive species, as well as the population dynamics, remain unclear. Further research is needed to provide deeper insights.
5. Conclusions
We found that foraging strategies in inter-specific competition contexts were influenced by personality during alien species invasion in both native and invasive species, and were also influenced by the personality of the competitors. However, these personality interaction effects differed between M. reevesii and T. scripta elegans. The foraging strategy of M. reevesii is mainly influenced by the personality types of T. scripta elegans, while the foraging strategy of T. scripta elegans is affected by both the personality types of M. reevesii and T. scripta elegans. This result may be due to the competitive advantages of species and the foraging strategies among different personalities, which may play a crucial role in the foraging behavior of native and invasive species during invasion dynamics. Overall, our results reveal the mechanisms of personality interaction effects on the foraging behavior of both native and invasive species during alien species invasion, and provide empirical evidence to help understand the effects of personality in invasion dynamics.
Author Contributions
Conceptualization, L.G. and Y.W.; methodology, L.G.; software, L.G.; investigation, L.G., S.Z., R.Z., T.S., L.T., H.Y. and K.H.; resources, L.G. and K.H.; data curation, L.G.; formal analysis, L.G.; writing—original draft preparation, L.G.; writing—review and editing, Y.W, and K.H.; visualization, L.G.; supervision, L.G. and Y.W.; funding acquisition, Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (No. 32200701).
Institutional Review Board Statement
Not applicable.
Acknowledgments
We are grateful to all the members of the Reptiles Evolutionary Physiological Ecology Group for their helpful discussions.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Blackburn, T.M.; Bellard, C.; Ricciardi, A. Alien versus Native Species as Drivers of Recent Extinctions. Frontiers in Ecology and the Environment 2019, 17, 203–207. [CrossRef]
- Sala, O.E.; Stuart Chapin, F.; III; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; et al. Global Biodiversity Scenarios for the Year 2100. Science 2000, 287, 1770–1774. [CrossRef]
- Doherty, T.S.; Glen, A.S.; Nimmo, D.G.; Ritchie, E.G.; Dickman, C.R. Invasive Predators and Global Biodiversity Loss. Proceedings of the National Academy of Sciences 2016, 113, 11261–11265. [CrossRef]
- Gallardo, B.; Clavero, M.; Sánchez, M.I.; Vilà, M. Global Ecological Impacts of Invasive Species in Aquatic Ecosystems. Global Change Biology 2016, 22, 151–163. [CrossRef]
- Bowler, D.E.; Benton, T.G. Causes and Consequences of Animal Dispersal Strategies: Relating Individual Behaviour to Spatial Dynamics. Biol Rev Camb Philos Soc 2005, 80, 205–225. [CrossRef]
- Clobert, J.; Le Galliard, J.-F.; Cote, J.; Meylan, S.; Massot, M. Informed Dispersal, Heterogeneity in Animal Dispersal Syndromes and the Dynamics of Spatially Structured Populations. Ecol Lett 2009, 12, 197–209. [CrossRef]
- Cote, J.; Bestion, E.; Jacob, S.; Travis, J.; Legrand, D.; Baguette, M. Evolution of Dispersal Strategies and Dispersal Syndromes in Fragmented Landscapes. Ecography 2017, 40, 56–73. [CrossRef]
- Lowe, W.H.; McPeek, M.A. Is Dispersal Neutral? Trends Ecol Evol 2014, 29, 444–450. [CrossRef]
- Berger-Tal, O.; Blumstein, D.T.; Carroll, S.; Fisher, R.N.; Mesnick, S.L.; Owen, M.A.; Saltz, D.; St. Claire, C.C.; Swaisgood, R.R. A systematic survey of the integration of animal behavior into conservation. Conservation Biology 2016, 30, 744–753. [CrossRef]
- del Río, L.; Navarro-Martínez, Z.M.; Cobián-Rojas, D.; Chevalier-Monteagudo, P.P.; Angulo-Valdes, J.A.; Rodriguez-Viera, L. Biology and Ecology of the Lionfish Pterois Volitans/Pterois Miles as Invasive Alien Species: A Review. PeerJ 2023, 11, e15728. [CrossRef]
- Evans, M.V.; Drake, J.M.; Jones, L.; Murdock, C.C. Assessing Temperature-Dependent Competition between Two Invasive Mosquito Species. Ecological Applications 2021, 31, e02334. [CrossRef]
- Nunes, A.L.; Fill, J.M.; Davies, S.J.; Louw, M.; Rebelo, A.D.; Thorp, C.J.; Vimercati, G.; Measey, J. A Global Meta-Analysis of the Ecological Impacts of Alien Species on Native Amphibians. Proceedings of the Royal Society B: Biological Sciences 2019, 286, 20182528. [CrossRef]
- Ruland, F.; Jeschke, J.M. How Biological Invasions Affect Animal Behaviour: A Global, Cross-Taxonomic Analysis. Journal of Animal Ecology 2020, 89, 2531–2541. [CrossRef]
- Gosling, S.D. Personality in Non-Human Animals: Animal Personality. Social and Personality Psychology Compass 2008, 2, 985–1001. [CrossRef]
- Cote, J.; Fogarty, S.; Weinersmith, K.; Brodin, T.; Sih, A. Personality Traits and Dispersal Tendency in the Invasive Mosquitofish (Gambusia Affinis). Proc Biol Sci 2010, 277, 1571–1579. [CrossRef]
- Duckworth, R.A.; Badyaev, A.V. Coupling of Dispersal and Aggression Facilitates the Rapid Range Expansion of a Passerine Bird. Proceedings of the National Academy of Sciences 2007, 104, 15017–15022. [CrossRef]
- Malange, J.; Izar, P.; Japyassú, H. Personality and Behavioural Syndrome in Necromys Lasiurus (Rodentia: Cricetidae): Notes on Dispersal and Invasion Processes. acta ethol 2016, 19, 189–195. [CrossRef]
- Chapple, D.G.; Simmonds, S.M.; Wong, B.B.M. Can Behavioral and Personality Traits Influence the Success of Unintentional Species Introductions? Trends in Ecology & Evolution 2012, 27, 57–64. [CrossRef]
- Fogarty, S.; Cote, J.; Sih, A. Social Personality Polymorphism and the Spread of Invasive Species: A Model. The American Naturalist 2011, 177, 273–287. [CrossRef]
- Galib, S.M.; Sun, J.; Twiss, S.D.; Lucas, M.C. Personality, Density and Habitat Drive the Dispersal of Invasive Crayfish. Sci Rep 2022, 12, 1114. [CrossRef]
- Short, K.H.; Petren, K. Boldness Underlies Foraging Success of Invasive Lepidodactylus Lugubris Geckos in the Human Landscape. Animal Behaviour 2008, 76, 429–437. [CrossRef]
- Neumann, K.M.; Pinter-Wollman, N. Collective Responses to Heterospecifics Emerge from Individual Differences in Aggression. Behav Ecol 2019, 30, 801–808. [CrossRef]
- Santicchia, F.; Wauters, L.A.; Tranquillo, C.; Villa, F.; Dantzer, B.; Palme, R.; Preatoni, D.; Martinoli, A. Invasive Alien Species as an Environmental Stressor and Its Effects on Coping Style in a Native Competitor, the Eurasian Red Squirrel. Hormones and Behavior 2022, 140, 105127. [CrossRef]
- Winandy, L.; Denoël, M. The Aggressive Personality of an Introduced Fish Affects Foraging Behavior in a Polymorphic Newt. Behavioral Ecology 2015, 26, 1528–1536. [CrossRef]
- Pearson, S.H.; Avery, H.W.; Spotila, J.R. Juvenile Invasive Red-Eared Slider Turtles Negatively Impact the Growth of Native Turtles: Implications for Global Freshwater Turtle Populations. Biological Conservation 2015, 186, 115–121. [CrossRef]
- Polo-Cavia, N.; López, P.; Martín, J. Competitive Interactions during Basking between Native and Invasive Freshwater Turtle Species. Biol Invasions 2010, 12, 2141–2152. [CrossRef]
- 100 of the World’s Worst Invasive Alien Species: A Selection From The Global Invasive Species Database. In Encyclopedia of Biological Invasions; Simberloff, D., Rejmanek, M., Eds.; University of California Press, 2019; pp. 715–716 ISBN 978-0-520-94843-3.
- Polo-Cavia, N.; López, P.; Martín, J. Aggressive Interactions during Feeding between Native and Invasive Freshwater Turtles. Biol Invasions 2011, 13, 1387–1396. [CrossRef]
- Turtle Taxonomy Working Group; Rhodin, A.G.J.; Iverson, J.B.; Bour, R.; Fritz, U.; Georges, A.; Shaffer, H.B.; van Dijk, P.P. Turtles of the World: Annotated Checklist and Atlas of Taxonomy, Synonymy, Distribution, and Conservation Status (8th Ed.); Chelonian Research Foundation & Turtle Conservancy, 2017; ISBN 978-1-5323-5026-9.
- van Dijk, P.P. Mauremys Reevesii. The IUCN Red List of Threatened Species 2011: E.T170502A97431862. 2011.
- Masin, S.; Bonardi, A.; Padoa-Schioppa, E.; Bottoni, L.; Ficetola, G.F. Risk of Invasion by Frequently Traded Freshwater Turtles. Biol Invasions 2014, 16, 217–231. [CrossRef]
- Wu, J.; Wu, Y.; Rao, D.; Zhou, T.; Gong, S. China’s Wild Turtles at Risk of Extinction. Science 2020, 368, 838–838. [CrossRef]
- Nishizawa, H.; Tabata, R.; Hori, T.; Mitamura, H.; Arai, N. Feeding Kinematics of Freshwater Turtles: What Advantage Do Invasive Species Possess? Zoology 2014, 117, 315–318. [CrossRef]
- Zhang, C.; Kang, C.; Huang, X.; Lu, H. Differences in swimming, righting performance and heart rate in the invasive Trachemys scripta elegans and native Mauremys reevesii hatchling. Acta Ecologica Sinica 2021, 41, 7204–7211. [CrossRef]
- Kashon, E.A.F.; Carlson, B.E. Consistently Bolder Turtles Maintain Higher Body Temperatures in the Field but May Experience Greater Predation Risk. Behav Ecol Sociobiol 2018, 72, 9. [CrossRef]
- Roth, T.C.; Rosier, M.; Krochmal, A.R.; Clark, L. A Multi-trait, Field-based Examination of Personality in a Semi-aquatic Turtle. Ethology 2020, 126, 851–857. [CrossRef]
- Miller, S.; Derenne, A.; Ellis-Felege, S.; Rhen, T. Incubation Temperature and Satiety Influence General Locomotor and Exploratory Behaviors in the Common Snapping Turtle (Chelydra Serpentina). Physiology & Behavior 2020, 220, 112875. [CrossRef]
- Reed, B.M.; Hobelman, K.; Gauntt, A.; Schwenka, M.; Trautman, A.; Wagner, P.; Kim, S.; Armstrong, C.; Wagner, S.; Weller, A.; et al. Spatiotemporal Variation of Behavior and Repeatability in a Long-Lived Turtle. Behav Ecol Sociobiol 2023, 77, 88. [CrossRef]
- Jiang, S.; Zhang, C.; Pan, X.; Storey, K.B.; Zhang, W. Distinct Metabolic Responses to Thermal Stress between Invasive Freshwater Turtle Trachemys Scripta Elegans and Native Freshwater Turtles in China. Integrative Zoology 2024, 00, 1–19. [CrossRef]
- Wauters, L.A.; Mazzamuto, M.V.; Santicchia, F.; Martinoli, A.; Preatoni, D.G.; Lurz, P.W.W.; Bertolino, S.; Romeo, C. Personality Traits, Sex and Food Abundance Shape Space Use in an Arboreal Mammal. Oecologia 2021, 196, 65–76. [CrossRef]
- Hadfield, J.D. MCMC Methods for Multi-Response Generalized Linear Mixed Models: The MCMCglmm R Package. J. Stat. Soft. 2010, 33. [CrossRef]
- Sih, A.; Cote, J.; Evans, M.; Fogarty, S.; Pruitt, J. Ecological Implications of Behavioural Syndromes. Ecology Letters 2012, 15, 278–289. [CrossRef]
- Réale, D.; Reader, S.M.; Sol, D.; McDougall, P.T.; Dingemanse, N.J. Integrating Animal Temperament within Ecology and Evolution. Biological Reviews 2007, 82, 291–318. [CrossRef]
- Réale, D.; Dingemanse, N.J.; Kazem, A.J.N.; Wright, J. Evolutionary and Ecological Approaches to the Study of Personality. Philos Trans R Soc Lond B Biol Sci 2010, 365, 3937–3946. [CrossRef]
- Wolf, M.; Weissing, F.J. Animal Personalities: Consequences for Ecology and Evolution. Trends in Ecology & Evolution 2012, 27, 452–461. [CrossRef]
- Greene, H.W. Antipredator Mechanisms in Reptiles. In: Biology of the Reptilia, Vol. 16 ( C. Gans & R. B. Huey, Eds). In Antipredator mechanisms in reptiles.; Alan R. Liss, New York, 1988; pp. 1–152.
- Polo-Cavia, N.; López, P.; Martín, J. Interspecific Differences in Responses to Predation Risk May Confer Competitive Advantages to Invasive Freshwater Turtle Species. Ethology 2008, 114, 115–123. [CrossRef]
- Ibáñez, A.; López, P.; Martín, J. Inter-Individual Variation in Antipredator Hiding Behavior of Spanish Terrapins Depends on Sex, Size, and Coloration. Ethology 2014, 120, 742–752. [CrossRef]
- Krause, J.; Loader, S.P.; McDermott, J.; Ruxton, G.D. Refuge Use by Fish as a Function of Body Length–Related Metabolic Expenditure and Predation Risks. Proceedings of the Royal Society of London. Series B: Biological Sciences 1998, 265, 2373–2379. [CrossRef]
- Polo-Cavia, N.; López, P.; Martín, J. Interspecific Differences in Heat Exchange Rates May Affect Competition between Introduced and Native Freshwater Turtles. Biol Invasions 2009, 11, 1755–1765. [CrossRef]
- Sih, A.; Bell, A.M.; Johnson, J.C.; Ziemba, R.E. Behavioral Syndromes: An Integrative Overview. The Quarterly Review of Biology 2004, 79, 241–277. [CrossRef]
- Anselme, P.; Güntürkün, O. How Foraging Works: Uncertainty Magnifies Food-Seeking Motivation. Behav Brain Sci 2018, 42, e35. [CrossRef]
- Carter, A.J.; Goldizen, A.W.; Tromp, S.A. Agamas Exhibit Behavioral Syndromes: Bolder Males Bask and Feed More but May Suffer Higher Predation. Behavioral Ecology 2010, 21, 655–661. [CrossRef]
- Ballard, G.; Dugger, K.M.; Nur, N.; Ainley, D.G. Foraging Strategies of Adélie Penguins: Adjusting Body Condition to Cope with Environmental Variability. Marine Ecology Progress Series 2010, 405, 287–302. [CrossRef]
- David, M.; Auclair, Y.; Giraldeau, L.-A.; Cézilly, F. Personality and Body Condition Have Additive Effects on Motivation to Feed in Zebra Finches Taeniopygia Guttata. Ibis 2012, 154, 372–378. [CrossRef]
- Sih, A.; Mathot, K.J.; Moirón, M.; Montiglio, P.-O.; Wolf, M.; Dingemanse, N.J. Animal Personality and State–Behaviour Feedbacks: A Review and Guide for Empiricists. Trends in Ecology & Evolution 2015, 30, 50–60. [CrossRef]
- Jeanniard-du-Dot, T.; Trites, A.W.; Arnould, J.P.Y.; Guinet, C. Reproductive Success Is Energetically Linked to Foraging Efficiency in Antarctic Fur Seals. PLoS One 2017, 12, e0174001. [CrossRef]
- Foraging: Behavior and Ecology; Stephens, D.W., Brown, J.S., Ydenberg, R.C., Eds.; University of Chicago Press: Chicago, IL, 2007; ISBN 978-0-226-77264-6.
- Zhou Z.; Cai S.; Liu Y.; Sun Y.; Luo L. Body temperature, thermal dependence of locomotor performance, compensatory growth, and the immunity of hatchling Red-eared turtles and Chinese pond turtles. Acta Ecologica Sinica 2016, 36, 7014–7022.
- Rankin, D.J.; Bargum, K.; Kokko, H. The Tragedy of the Commons in Evolutionary Biology. Trends in Ecology & Evolution 2007, 22, 643–651. [CrossRef]
- Berger-Tal, O.; Embar, K.; Kotler, B.P.; Saltz, D. Everybody Loses: Intraspecific Competition Induces Tragedy of the Commons in Allenby’s Gerbils. Ecology 2015, 96, 54–61. [CrossRef]
- Chase, I.D.; Bartolomeo, C.; Dugatkin, L.A. Aggressive Interactions and Inter-Contest Interval: How Long Do Winners Keep Winning?. Animal Behaviour 1994, 48, 393–400. [CrossRef]
- Mathot, K.J.; Wright, J.; Kempenaers, B.; Dingemanse, N.J. Adaptive Strategies for Managing Uncertainty May Explain Personality-Related Differences in Behavioural Plasticity. Oikos 2012, 121, 1009–1020. [CrossRef]
- Reader, S.M. Causes of Individual Differences in Animal Exploration and Search. Topics in Cognitive Science 2015, 7, 451–468. [CrossRef]
- Stamps, J.A.; Groothuis, T.G.G. Developmental Perspectives on Personality: Implications for Ecological and Evolutionary Studies of Individual Differences. Philosophical Transactions of the Royal Society B: Biological Sciences 2010, 365, 4029–4041. [CrossRef]
- Gharnit, E.; Dammhahn, M.; Garant, D.; Réale, D. Resource Availability, Sex, and Individual Differences in Exploration Drive Individual Diet Specialization. The American Naturalist 2022, 200, 1–16. [CrossRef]
- Li, C.; Zhang, X.; Cheng, L.; Zhang, B.; Zhang, F. Food Patch Use of Japanese Quail (Coturnix Japonica) Varies with Personality Traits. Frontiers in Zoology 2023, 20, 30. [CrossRef]
- Patrick, S.C.; Pinaud, D.; Weimerskirch, H. Boldness Predicts an Individual’s Position along an Exploration–Exploitation Foraging Trade-Off. Journal of Animal Ecology 2017, 86, 1257–1268. [CrossRef]
- Giraldeau, L.-A.; Dubois, F. Chapter 2 Social Foraging and the Study of Exploitative Behavior. In Advances in the Study of Behavior; Academic Press, 2008; Vol. 38, pp. 59–104.
- Jeffries, P.M.; Patrick, S.C.; Potts, J.R. Be Different to Be Better: The Effect of Personality on Optimal Foraging with Incomplete Knowledge. Theor Ecol 2021, 14, 575–587. [CrossRef]
- Sjerps, M.; Haccou, P. Effects of Competition on Optimal Patch Leaving: A War of Attrition. Theoretical Population Biology 1994, 46, 300–318. [CrossRef]
- Gyimesi, A.; van Rooij, E.P.; Nolet, B.A. Nonlinear Effects of Food Aggregation on Interference Competition in Mallards. Behav Ecol Sociobiol 2010, 64, 1897–1904. [CrossRef]
- Scharf, I.; Filin, I.; Ovadia, O. An Experimental Design and a Statistical Analysis Separating Interference from Exploitation Competition. Popul Ecol 2008, 50, 319–324. [CrossRef]
Figure 1.
Bold-exploration personality and behavioral variables difference between M. reevesii and T. scripta elegans: (a) personality scores; (b) cross frequency in exploration test; (c) moving distance in exploration test; (d) head emergency latency in boldness test; (e) movement latency in boldness test. *** p < 0.001.
Figure 1.
Bold-exploration personality and behavioral variables difference between M. reevesii and T. scripta elegans: (a) personality scores; (b) cross frequency in exploration test; (c) moving distance in exploration test; (d) head emergency latency in boldness test; (e) movement latency in boldness test. *** p < 0.001.
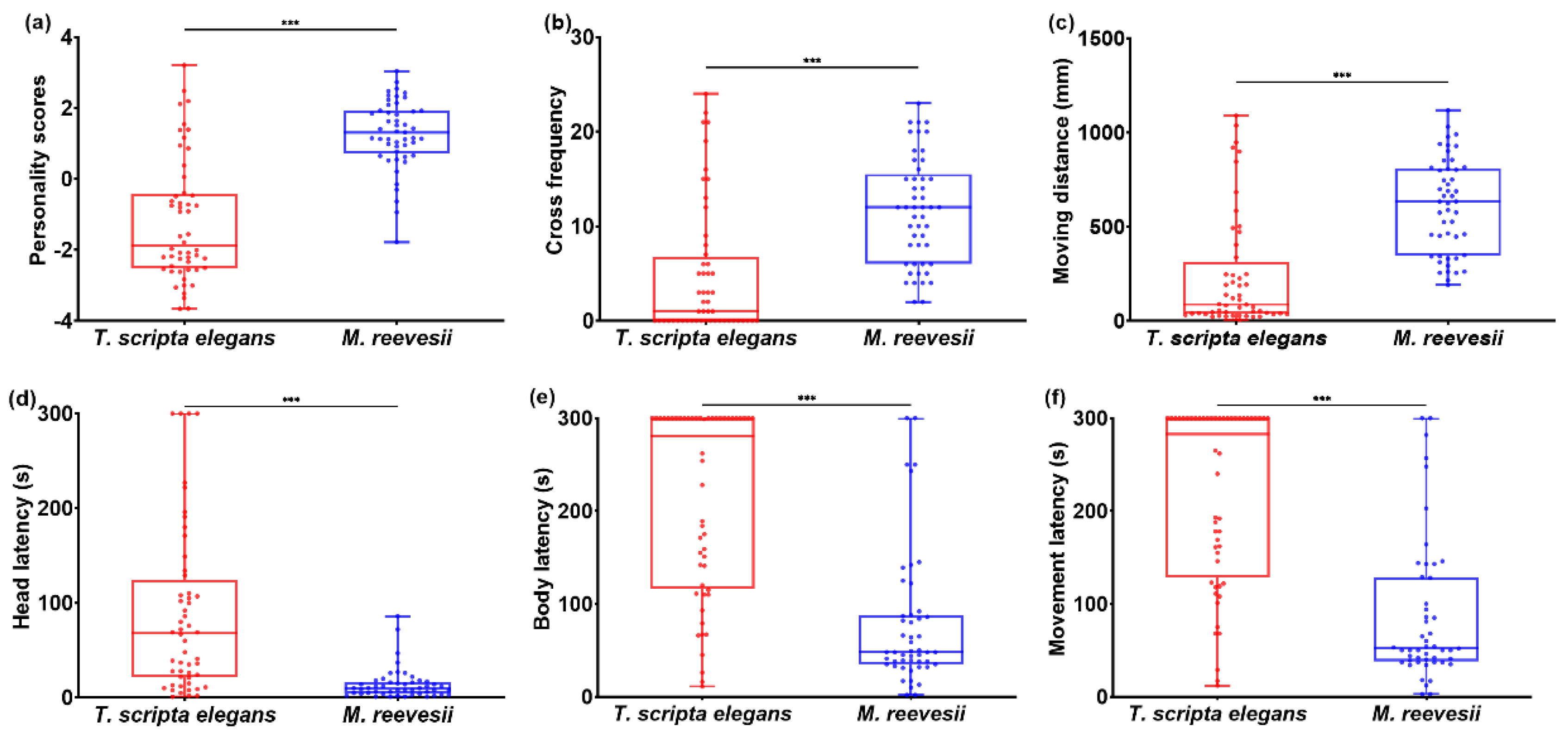
Figure 2.
Foraging behavior traits of M. reevesii and T. scripta elegans: (a) Log foraging latency of M. reevesii; (b) Log foraging frequency of M. reevesii; (c) Log foraging duration of M. reevesii; (d) Log foraging latency of T. scripta elegans; (b) Log foraging frequency of T. scripta elegans; (c) Log foraging duration of T. scripta elegans. * p < 0.05; ** p 0.01; *** p< 0.001.
Figure 2.
Foraging behavior traits of M. reevesii and T. scripta elegans: (a) Log foraging latency of M. reevesii; (b) Log foraging frequency of M. reevesii; (c) Log foraging duration of M. reevesii; (d) Log foraging latency of T. scripta elegans; (b) Log foraging frequency of T. scripta elegans; (c) Log foraging duration of T. scripta elegans. * p < 0.05; ** p 0.01; *** p< 0.001.
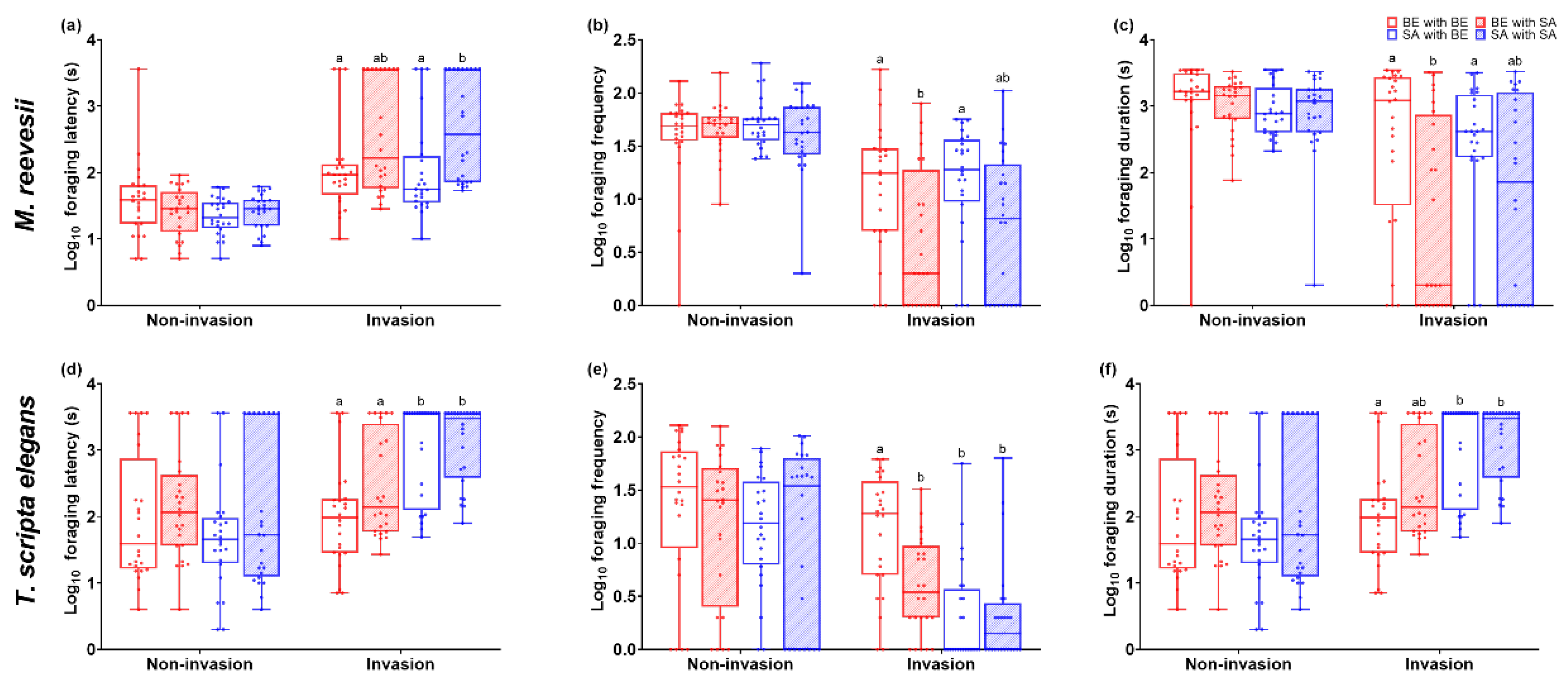
Figure 3.
Morphology traits of M. reevesii and T. scripta elegans: (a) body mass; (b) carapace length; (c) carapace width; (d) carapace height; (e) carapace height/width ratio. *** p < 0.01.
Figure 3.
Morphology traits of M. reevesii and T. scripta elegans: (a) body mass; (b) carapace length; (c) carapace width; (d) carapace height; (e) carapace height/width ratio. *** p < 0.01.
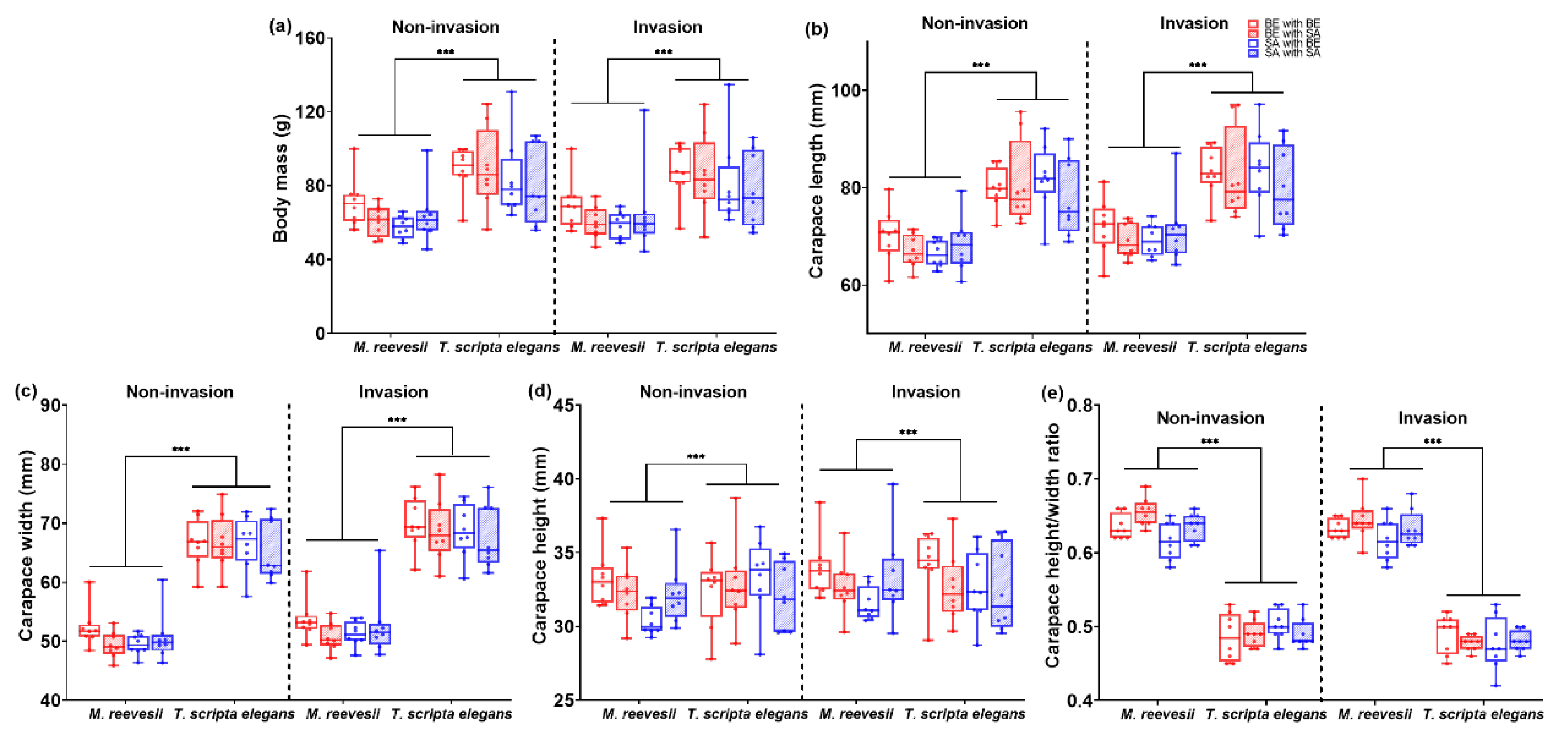

Table 1.
Correlation of each behavior observed during the boldness and exploration tests on M. reevesii and T. scripta elegans with the components of the principal component analysis (PCA).
Table 1.
Correlation of each behavior observed during the boldness and exploration tests on M. reevesii and T. scripta elegans with the components of the principal component analysis (PCA).
| Behavioral Variables | Component 1 (PC1) | Component 2 (PC 2) |
|---|---|---|
| Boldness: head latency | -0.388 | -0.470 |
| Boldness: body latency | -0.490 | -0.299 |
| Boldness: movement latency | -0.488 | -0.292 |
| Exploration: moving distence | 0.442 | -0.517 |
| Exploration: mross frequency | 0.419 | -0.580 |
| Eigenvalue | 3.50 | 1.00 |
| Total variance (%) | 70.02% | 20.09% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



