
Submitted:
19 June 2024
Posted:
19 June 2024
You are already at the latest version
Abstract
Medicinal mushrooms, especially Ganoderma species hold immense promise for production of a wide range of bioactive compounds with various effects. The biochemical potential of indigenous fungal strains, specific to a region, could play a critical role in the continuous search for novel strains with superior activities on a global scale. This research focused on the ethanolic (EtOH) and hot-water (H2O) extracts of fruiting bodies of two wild-growing Ganoderma species: G. pfeifferi and G. resinaceum with an aim to assess its nutritional (total carbohydrate content-TCC), and mineral composition in relation to bioactive properties: antioxidant, antiproliferative and lipid-lowering. Atomic absorption spectrophotometry (AAS) revealed that G. pfeifferi is a promising source of minerals that are essential for numerous physiological functions in the human body like bone health and muscle and nerve function, with calcium (Ca 4.55 ± 0.41 mg/g d.w.) and magnesium (Mg 1.33 ± 0.09 mg/g d.w.), being the most abundant macro element present. Zinc (Zn), manganese (Mn), and chromium (Cr) were particularly notable, with concentrations ranging from 21.49 to 41.70 mg/kg d.w.
The EtOH extract of G. pfeifferi demonstrated significantly elevated levels of TCC, essential macromolecules for energy and structural functions in the body, with higher quantities of all three standard carbohydrates detected in this type of extract. Similar to revealed composition the same species, G. pfeifferi stood out as the most prominent antioxidant agent, with H2O extract being stronger than EtOH in ABTS assay (86.85 ± 0.67 mg TE/g d.w.), while EtOH extract displayed the highest anti OH• radical scavenging ability (IC50 = 0.18 ± 0.05 μg/mL) as well as the most notable reducing potential among all. The highest antiproliferative effect against the breast cancer cell line (MCF-7), demonstrated the H2O extracts from G. resinaceum with the most pronounced activity after 24 hours (IC50 = 4.88 ± 0.50 μg/mL) which surpasses that of the standard compound, ellagic acid (IC50 = 33.94 ± 3.69 μg/mL). Administration of both Ganoderma extracts mitigated diabetic lipid disturbances and exhibited potential renal and hepatic protection in vivo on white Wistar rats by preservation of kidney function parameters in G. resinaceum H2O pre-treatment (urea: 6.27 ± 0.64 mmol/L, creatinine: 50.00 ± 6.45 mmol/L) and the reduction of ALT levels (17.83 ± 3.25 U/L) com-pared to diabetic control groups treated with saline (urea: 46.98 ± 6.01 mmol/L, creatinine: 289.25 ± 73.87 mmol/L, and ALT: 60.17 ± 9.64 U/L). These results suggest that pre-treatment with G. resinaceum H2O extracts may have potential antidiabetic properties.
In summary, detected microelements are vital for maintaining overall health, supporting metabolic processes, and protecting against various chronic diseases. Further research and dietary assessments could help determine the full potential and applications of two underexplored Ganoderma species native to Serbia in nutrition and health supplements.
Keywords:
Ganoderma
; natural products
; health benefits
; fungal extracts
; anticancer activity
; liver protection
; kidney protection
; mineral composition
1. Introduction
The integration of traditional knowledge with modern scientific research continues to uncover valuable natural compounds with potential health benefits. The rising incidence of chronic diseases due to aging and lifestyle changes underscores the need for effective treatments [1]. Fungi, with their rich nutritional profile and therapeutic properties, offer promising avenues for addressing global health concerns, particularly oxidative stress. Leveraging ancient folk remedies, many of which form the basis of modern medications, can enhance our approach to managing and preventing these prevalent health issues [2]. For instance, recent research has focused on natural products for the prevention and treatment of metabolic syndrome, with particular attention to medicinal mushrooms [3,4]. Among these, the Ganoderma species, known for their extensive use in traditional medicine, have shown promising potential [5,6]. Ganoderma species, such as G. resinaceum, have been documented to possess various bioactive compounds, including nortriterpenoids (lucidone A–F, H, I–K, ganosineniol B–C, ganoderense F, and ganosineniol), which exhibit a wide range of beneficial biological activities [7]. These activities include anti-inflammatory, antioxidant, and antidiabetic effects, which are particularly relevant to combating the multifaceted aspects of metabolic syndrome [5,7].
Species belonging to the genus Ganoderma (P. Karst.) have been utilized as a natural resource in Traditional Chinese Medicine (TCM) for over two hundred years [2,8]. Its utilization has primarily relied on observations and accounts of its efficacy in cancer prevention and treatment, combating infections, modulating the immune system, and regulating blood pressure, among other medicinal benefits [5,6]. The medicinal properties of Ganoderma spp. primarily stem from the abundance of various bioactive compounds they produce, notably triterpenoids like ganoderic acids, which have been reported to exhibit anti-hypercholesterolemic, anticancer, hepatoprotective, antioxidant, anti-inflammatory, antimicrobial, and hypoglycemic properties [9,10]. G. resinaceum extracts have shown inhibitory effects against enzymes such as acetylcholinesterase, tyrosinase, α-amylase, and α-glucosidase [11,12]. Also, nortriterpenoids extracted from Ganoderma species display a diverse range of biological activities, such as antitumor, anti-inflammatory, neurotrophic, hepatoprotective, and anti-HIV-1 protease effects [7]. These properties are noteworthy due to their structural diversity and potential as models in pharmaceutical research. Additionally, polysaccharides such as β-glucans and phenolic compounds play significant roles, exhibiting anti-inflammatory, antimicrobial, antioxidant and antiproliferative [5,6,11,12,13,14,15,16,17], while ergosterol peroxide has been shown to induce tumor cell death [18]. Wang et al. [8] proposed that the bioactive compounds from G. pfeifferi and other Ganoderma species may exert a synergistic effect, indicating that their medicinal properties typically result from the combination of multiple ingredients.
Ganoderma species, renowned for their medicinal properties, have been extensively studied for their various health benefits [5,6,12,15,16,19,20,21]. In 1889, Patouillard expanded the genus to include 48 species, while data from 2022 indicate that the genus now comprises 181 species [22]. Among them, G. lucidum is globally recognized, owing to its use in TCM as the “Mushroom of Immortality” [23]. The genus is widely distributed across the globe, particularly in tropical and subtropical areas spanning Africa, America, Asia, and Europe [24]. In Europe, the genus consists of seven species [20], with G. pfeifferi and G. resinaceum being among the less studied species compared to G. lucidum, G. applanatum, and G. adspersum, among others.
Regarding taxonomy of these species, Rašeta et al. [12] claimed that G. pfeifferi and G. resinaceum, along with G. subamboinense and three G. lucidum strains from the United States and Taiwan, form a single monophyletic cluster based on phylogenetic analysis. This group is distinguished by its ability to produce chlamydospores in culture. Two selected species, G. pfeifferi and G. resinaceum are from genus Ganoderma, which is the major genus of the Ganodermataceae family (Polyporales, Basidiomycota), known for producing bioactive compounds such as triterpenoids, polysaccharides, sterols and phenolics [6,20].
Studies have also highlighted the efficacy of Ganoderma extracts in improving insulin sensitivity, regulating lipid metabolism, and reducing inflammation, as key factors in the management of metabolic syndrome [25,26,27,28]. The bioactive compounds in Ganoderma species can modulate multiple metabolic pathways in the body [29], making Ganoderma a promising candidate for the prevention and treatment of various chronic diseases [30].
As of today, the focus on antioxidant, antimicrobial, and enzyme inhibition including in vitro antidiabetic activity [11,12,13,14,16,17,19,31,32] has focused on new medicinal species G. pfeifferi, originated from Serbia but also presents an autochthonous European species, and G. resinaceum, distributed in Asia, and North Africa as well.
In our previous study, we presented findings on the use of (EtOH) and hot-water (H2O) extracts in suspension form derived from G. pfeifferi and G. resinaceum, which showed potential for application in diabetes [16]. Hence, the objective of this study was to conduct a thorough comparative assessment of the in vitro antiproliferative and in vivo lipid-lowering, and hepato- and nephroprotective characteristics of G. pfeifferi and G. resinaceum, an area notably deficient in current literature.
2. Materials and Methods
2.1. Fungal Material and Extracts Preparation
Fruiting bodies of two Ganoderma fungal species, namely G. pfeifferi and G. resinaceum, were gathered in September 2010 from Nature Park Begečka Jama and the University of Novi Sad Campus in Serbia, respectively. The determination and identification of the collected specimens took place at the Department of Biology and Ecology, University of Novi Sad and voucher specimens have been archived in the ProFungi Laboratory, Department of Biology and Ecology, University of Novi Sad (Serbia), under the numbers 12-00723 and 12-00722.
All experiments utilized ethanol (EtOH) and hot-water (H2O) extracts, prepared according to previously established methods [15,16]. Both extracts were dissolved in distilled water (dH2O), achieving a final concentration of 100 mg/mL dry weight (d.w.) and were stored at temperatures of −20 °C before undergoing analysis.
2.2. Mycochemical Characterization
2.2.1. Quantification of Macro- and Microelements by Using Atomic Absorption Spectrophotometry (AAS)
Macro- (Ca and Mg) and microelements (Cu, Ni, Cd, Pb, Cr, Mn, Fe and Zn) were analyzed in powdered fungal samples following the procedure outlined by Kebert et al. [33] using a flame technique within Atomic Absorption Spectrophotometry (AAS) [model FS AAS240/GTA120, Agilent] (Santa Clara, California, USA). Approximately 0.3 g of dried material (oven-dried at 70 °C for 24 hours) was ground, homogenized, and digested in a mixture of 10 mL of nitric acid and 2 mL of 30% (w/v) hydrogen peroxide using a microwave-assisted digestion system (D series; Milestone, Bergamo, Italy) at 180 °C (900 W) for 45 minutes. The resulting homogenates were filtered and diluted to 25 mL with deionized water. The prepared samples were analyzed by using with the acetylene/air burner flame technique for Cr, Cu, Mn, Fe, and Zn and the nitrous oxide (N2O)–acetylene flame for Ca content determination. Concentrations of each element were determined using single-element hollow-cathode lamps at specific wavelengths and expressed as mg/kg dry weight (d.w.) of fungal material.
2.2.2. Total Carbohydrate Content (TCC)
The total carbohydrate content (TCC) of the fungal extracts was determined using the phenol–sulfuric acid method outlined in Rašeta et al. [16]. Initially, 50 μL of each fungal extract or glucose was mixed with 150 μL of concentrated H2SO4 (or dH2O for correction) and shaken for 30 minutes at room temperature in an incubator shaker (IKA KS 4000i control, Staufen, Baden-Württemberg, Germany). Subsequently, 30 μL of 5% phenol in water was added, and the mixture was heated for 10 minutes at 70 °C in the same incubator shaker. After allowing the microplate (Thermo Fisher Scientific, Waltham, Massachusetts, USA) to cool to room temperature for 5 minutes in a water bath, it was dried, and absorbances (Thermo Fisher Scientific, Waltham, Massachusetts, USA) were taken at 490 nm for glucose (hexose), 750 nm for sucrose as a disaccharide, and 480 nm for xylose (pentoses). A calibration curves using glucose, sucrose and xylose (13.59–2173.91 μg/mL), were prepared to determine the total carbohydrate content of the fungal extracts, expressed as mg glucose equivalents (mg GluE), mg sucrose equivalents (mg SucE), and mg xylose equivalents (mg XylE), per gram of dry weight (d.w.).
2.3. In Vitro Examination of Biological Activities
2.3.1. Antioxidant Activity
The antioxidant activity was assessed through the ABTS radical scavenging activity assay [34], OH radical scavenging activity assay [35], while the reducing power of fungal extracts was determined using the ascorbate equivalent antioxidant capacity (A.E.A.C.) assay [36]. The scavenging activity in the case of ABTS assay and reducing power of the extracts were determined based on the standard curve equation of Trolox and ascorbic acid, respectively, while results of scavenging OH assay was expressed as IC50 values (concentration that inhibits 50% of hydroxyl radicals). The results were quantified as milligrams of Trolox equivalents per gram of dry weight (mg TE/g d.w.) and milligrams of ascorbic acid equivalents per gram of dry weight (mg AAE/g d.w.).
2.3.2. Antiproliferative Activity
The antiproliferative activity of the analyzed fungal extracts was assessed using the estrogen-dependent breast cancer cell line (MCF-7) following the method outlined by Mosmann [37]. Ellagic acid was utilized as positive control agent. Cancer cell viability was monitored over a 24-hour (acute) and 72-hour (chronic) incubation period for extract concentrations ranging from 50 to 250 µg/mL. Cell cytotoxicity was determined as the IC50, which represents the concentration that inhibits 50% of cell growth extrapolated from concentration–response curves.
2.4. In Vivo Procedures and Assays
2.4.1. Laboratory Animals
In vivo part of the research was conducted on white Wistar rats of both genders, obtained from the Military Medical Academy of Belgrade, Republic of Serbia. The rats, weighing between 210 and 340 grams and aged up to four months, were accommodated in UniProtect airflow cabinets (Ehret GmbH, Emmendingen, Germany) with standard plexiglass cages. The housing conditions maintained a constant room temperature of 22 ± 1 °C, 55% ± 1.5% humidity, and a regular circadian rhythm (12-hour day/night cycle). Throughout the entire experiment, the rats were provided with unrestricted access to tap water and standard pelleted laboratory rodent feed from the Veterinary Institute Subotica, Serbia. All experimental procedures adhered to the guidelines of the European Directive (2010/63/EU) for animal experiments and were subject to review and approval by the Ethics Committee for the Protection and Welfare of Experimental Animals at the University of Novi Sad, Serbia.
2.4.2. Experimental Procedures
The animals were randomly allocated into ten groups, each comprising six individuals. Among these, five groups underwent no alloxan pre-treatment (normoglycemic), while the remaining five groups were subjected to alloxan-induced hyperglycemia (diabetic). In order to induce hyperglycemia alloxan was dissolved in saline and applied intraperitoneally at a dose of 100 mg/kg. Hyperglycemia was confirmed 48 hours following the application of alloxan and animals with a glycemia higher than 15 mmol/L were selected for subsequent experiment. Both, normoglycemic and diabetic, control groups of animals were treated with saline at a dose of 1 mg/mL. Similarly, experimental groups of normoglycemic and diabetic animals were subjected to identical treatment, receiving an oral aqueous suspension (1 mg/mL) containing the EtOH and H2O extracts of the two analyzed fungal species. The extract suspensions were administered perorally by nasogastric probe over a 5-day period. Two hours after the last dose of fungal extracts or saline, the rats were anesthetized using a 25% solution of urethane at a dose of 5 mL/kg via intraperitoneal injection. Once the righting reflex was lost, the animals were exsanguinated through intracardial puncture to obtain blood and tissue samples for further analysis.
2.4.3. In Vivo Biochemical Parameters Analysis
The concentration of lipids (lipid status) was determined in the serum of animals. The concentrations of total cholesterol, total triglycerides and high-density lipoprotein (HDL) cholesterol and low-density lipoprotein (LDL) were measured using clinical biochemistry methods.
The enzymatic activity of aspartate aminotransferase (AST) and alanine aminotransferase (ALT), as well as the concentrations of urea and creatinine, were assessed in order to monitor hepatic and renal function using serum of the examined animals.
All analyses were performed using standard spectrophotometric methods on an automatic chemical analysis system, Olympus AU 400 (Hamburg, Germany).
2.4. Statistical Analysis
The study utilized an array of statistical methods, including descriptive statistics, one-way Analysis of Variance (ANOVA), t-tests, Principal Component Analysis (PCA), dendrogram hierarchical clustering, and Pearson correlation analysis. The differentiation among analyzed fungi species in the one-way ANOVA was assessed using the Fisher (F) test, with statistical significance denoted by “p-values.” The results of the t-tests were depicted with box-plot diagrams. All statistical analyses were performed using the R programming environment. The “rstatix” R package was employed for descriptive statistics, two-way ANOVA, and t-tests, while dendrogram clustering was carried out with the “dendextend” R package. Various data visualizations were created using the “ggplot2” R package [38,39,40].
3. Results and Discussion
3.1. Mycochemical Characterization
3.1.1. AAS Quantification of Macro- and Microelements
AAS analysis was utilized to evaluate the multi elemental composition of both G. pfeifferi and G. resinaceum, encompassing a total of 11 metals. These included macroelements such as Ca, K and Mg, and microelements like Cu, Ni, Cd, Pb, Cr, Mn, Fe and Zn. The summarized results can be found in Figure 1a,b.
Figure 1a,b display the outcomes of the mineral composition analysis, highlighting the prevalence of three major macroelements, namely Mg, Ca and K. In G. pfeifferi, the predominant macroelements were Mg and Ca, constituting a substantial portion of the overall mineral composition [32], which aligns with previous studies highlighting the essential role of Mg and Ca in various physiological processes, including the immune regulating actions of Mg and its crucial role in regulating inflammation and immune response to infectious agents and malignancies [41,42]. Contrastingly, G. resinaceum exhibited a distinctive macroelement profile, with a notable abundance of K. The significance of potassium in cellular activities, particularly in maintaining osmotic balance and regulating enzyme functions, suggests potential therapeutic implications associated with G. resinaceum consumption [43,44].
Beyond macroelements, the AAS analysis also unveiled the presence of essential microelements in both Ganoderma species (Figure 1b). It is noteworthy that elements such as Zn and Cu were detected in appreciable amounts in both tested species, with higher concentration of Zn in G. pfeifferi (41.70 ± 1.11 mg/kg d.w.), in contrast to G. resinaceum, where Cu was a more dominant microelement (22.22 ± 0.48 mg/kg d.w.). Moreover, the concentration of Cu in G. resinaceum from Poland was two times lower (11.00 ± 3.00 mg/kg d.m.) [45] compared to concentration determined in this study, while Zn was not quantified. These microelements play crucial roles in enzymatic activities, oxidative stress defense, and overall metabolic processes [46,47].
The comparative assessment of mineral composition between G. pfeifferi and G. resinaceum unveils species-specific variations and prevalence of tested elements in G. pfeifferi. This aligns with the previous report where Marek et al. [45] suggested that among tested group of Ganoderma species, fruit bodies of wild growing G. resinaceum and cultivated G. pfeifferi were characterized by a higher level of all elements jointly than the other analyzed Ganoderma species. Rašeta et al. [48] highlighted that the mineral content of edible fungi after consumption is influenced by cooking or processing methods, often resulting in mineral leakage into water or brine. While these minerals offer nutritional benefits, excessive intake may pose risks, especially considering fungus’ ability to accumulate toxic elements and radionuclides. Therefore, it’s crucial to assess metal content in wild fungi and consume them in moderation.
Considering this information and as depicted in Figure 1b, it can be inferred that G. pfeifferi and G. resinaceum exhibited negligible accumulation of toxic elements such as the examined Cd and Pb (ranging from 2.84 to 4.93 mg/kg and 2.84 to 3.45 mg/kg d.w., respectively). Such distinctions may influence the therapeutic potential of these fungi, as the interplay between different elements could contribute to their observed biological activities.
In summary, the metal composition analysis of G. pfeifferi and G. resinaceum indicated significant concentrations of biogenic metals alongside minor levels of heavy metals, potentially attributable to bioaccumulation processes, which is in accordance with Yalcin et al. [32]. Consequently, there arises a necessity for the regulated cultivation of these fungi with promising medicinal attributes. Moreover, understanding the mineral composition of Ganoderma species is pivotal for unraveling the potential therapeutic benefits associated with these fungal species.
3.1.2. Total Carbohydrate Content (TCC)
Regarding the TCC analysis, it is important to highlight that EtOH extracts of G. pfeifferi demonstrated a notably higher concentration of glucose and sucrose (303.87 ± 54.80 mg GluE/g d.w. and 44.51 ± 9.49 mg SucE/g d.w., respectively) compared to the same extract of G. resinaceum, indicating a substantial disparity in their carbohydrate composition (Figure 2). Conversely, G. resinaceum EtOH exhibited increased xylose content (233.18 ± 11.37 mg XylE/g d.w.), whereas all hot-water extracts displayed statistically significant lower TCC in both species (Figure 2).
These findings align with our earlier investigation, wherein EtOH extracts demonstrated superiority in TCC compared to water extracts, accompanied by a higher TCC content measured in glucose equivalents in G. pfeifferi [12]. On the other hand, higher TCC was determined in the study of G. resinaceum from Serbia, where 52.1 ± 3.2 g GluE/100 g was quantified in hot water extract [31]. This discrepancy in carbohydrate levels between the two species and type of solvent used, suggest that extraction method, together with geographical origin i.e., ecological factors, may have implications for their nutritional value and potential applications, as suggested for other fungal species [49]. Quantification of monosaccharide content in analyzed species represent valuable report, since generally fungal polysaccharides are composed of glucose, galactose, and mannose, but other carbohydrates can also be found (e.g., xylose, arabinose, fucose, ribose) [50], as demonstrated in this study as well. Since ratio of monosaccharide composition in fungi is very important and it has been shown that polysaccharides from G. lucidum, have positive effects as hypoglycemic agents [51], future research should be based on detailed identification of polysaccharides including monosaccharide composition from the analyzed two Ganoderma species as well.
3.2. Biological Activities of Examined Extracts
3.2.1. Antioxidant Activity
The antioxidant activity of Ganoderma species was assessed through ABTS, OH, and A.E.A.C assays, revealing varying degrees of efficacy across the tested species (Figure 3).
Results from the ABTS assay indicated potent antioxidant capacity in G. pfeifferi H2O extracts (86.85 ± 0.67 mg TE/g d.w.), while G. resinaceum exhibited a comparatively lower neutralization of ABTS radical. Furthermore, there was no statistically significant distinction observed in the neutralization of this radical between the EtOH and H2O extracts of G. resinaceum. This similarity is also evident in the extracts of G. pfeifferi, albeit in the neutralization of the OH radical, where extracts demonstrated notable antioxidant potential in comparison with the analyzed standard compound (PG) (Figure 3). Interestingly, the A.E.A.C assay also highlighted G. pfeifferi as possessing the highest reduction power activity, compared to reduction ability of G. resinaceum extracts.
In samples of G. resinaceum obtained from Turkey as well Zengin et al. [11] determined lower antioxidant potential in comparison with results of this study, underscoring the diverse antioxidant properties among Ganoderma species. Results from this study are in accordance with the study of Yalcin et al. [32], where extracts of G. pfeifferi from Turkey showed higher reduction potential compared to neutralization of ABTS radical. Moreover, compared to our results, H2O extracts of G. pfeifferi exhibited two times higher ability of ABTS radical neutralization (170.32 ± 3.17 mg TE/g) [32]. Recently, Sułkowska-Ziaja et al. [19] conducted antioxidant analyses on both studied species. In comparison with our findings, methanolic extracts from mycelial cultures (Lublin, Poland) exhibited lower activity in the ABTS assay (9.77 ± 0.13 mg TE/g for G. pfeifferi and 11.60 ± 0.36 mg TE/g for G. resinaceum, respectively), while the reduction potential of the samples analyzed by Sułkowska-Ziaja et al. [19] was also lower (ranging from 8.10 to 31.78 mg TE/g). This suggests that the choice of extract solvent and preparation method could influence experimental outcomes.
PCA analysis was conducted to revile a connection between the antioxidant activity and the detected TCC, alongside our previous research (Table S1) [16], where the phenolic profile was determined in the tested extracts. PCA analysis revealed 60.40% variance of PC1 and 25.97% of PC2, while distinct clustering may be observed among two analyzed Ganoderma species (Figure 4).
Evidently, G. resinaceum separated in the positive quadrant of both PCs, opposite to G. pfeifferi extracts and all quantified compounds, suggesting negative correlation between phenolics (Table S1), TCC and antioxidant properties of G. resinaceum extracts [16] (Figure 4). On the contrary, in G. pfeifferi extracts clustered together with TCC and all identified phenolics, suggesting they role as strong antioxidative agents in this species. Moreover, this is in accordance with correlation analysis where strong positive correlation between antioxidant properties except in the case of neutralization of OH radical and phenolic compounds is evident (Figure 5). Also, this assumption was confirmed in previous research as well [11,12,13,15,16,49]. However, separation of G. resinaceum extracts in PCA graph, opposite to phenolics (Table S1), could be related to lower levels of these compounds and thus lower antioxidant properties observed.
3.2.2. Antiproliferative Activity
The MTT assay was utilized to assess the in vitro antiproliferative activity of the two analyzed crude extracts and standard compound (ellagic acid). According to data from Table 1, it is evident that all extracts demonstrated antiproliferative effects during subacute incubation (after 24 hours), with the most potent activity observed in the H2O extract of G. resinaceum species (IC50 = 4.88 ± 0.50 μg/mL), comparable to the activity of standard compound (IC50 = 33.94 ± 3.69 μg/mL for ellagic acid). In general, EtOH extracts showed less inhibition of MCF-7 cells, except for the G. pfeifferi EtOH extract, which exhibited significant subacute inhibition (IC50 = 154.05 ± 12.92 μg/mL).
PCA analysis was performed to establish a link between the antiproliferative activities observed in this study and detected TCC, together with our prior research findings (Table S1) [16] where phenolic profile was determined in the tested extracts. The PCA analysis demonstrated distinct clustering patterns among the Ganoderma species based on their antiproliferative activity. The initial pair of principal components, PC1 and PC2, explained 86.37% of the overall variance, demonstrating a significant portrayal of the dataset. Specifically, G. resinaceum which exhibited the highest antiproliferative activity, separated in the positive quadrant of both PCs, opposite to G. pfeifferi and all quantified compounds [16] (Figure 4). This is in accordance with obtained results, since lower levels of phenolic compounds (Table S1) and TCC were observed in G. resinaceum EtOH and H2O extracts. On the other hand, correlation matrix revealed positive correlation among xylose and antiproliferative activity (Figure 5), suggesting that maybe carbohydrate compounds are important for cytotoxic effect. In contrast, extracts from G. pfeifferi exhibited diminished antiproliferative activity, yet demonstrated elevated levels of TCC (Figure 4) and phenolic content presented in Table S1, clustering together within the I and III quadrants. Moreover, correlation matrix showed that only protocatechuic acid, one of the most abundant phenolic in G. pfeiferi extracts (Table S1), has a significant positive correlation with antiproliferative activity (Figure 5).
Rašeta et al. [15] summarized that extensive research conducted in recent decades has identified a broad spectrum of bioactive compounds extracted from Ganoderma species, including phenolic acids, isoflavones, polysaccharides, triterpenes, sterols, nucleosides, proteins, and polysaccharide-protein complexes, all with potential antiproliferative effects. Considering the analysis of the MCF-7 cell line with unpurified extracts, it is hypothesized that interactions among various biomolecules present in the tested extracts may synergistically demonstrate antiproliferative activity, potentially mitigating the toxicity of individual components. This suggests that the interaction between different biomolecules could enhance the therapeutic efficacy of Ganoderma crude extracts [15]. However, drawing on previously reported data, it is theorized that the polysaccharides present in the analyzed hot-water extracts are responsible for the antiproliferative activity, while terpenoids potentially present in the EtOH extracts contribute primarily to a proliferative effect on MCF-7 cells [15]. Consequently, it can be inferred that this effect is linked to the direct antiproliferative treatment against tumor cells [52]. The same group claimed that high-molecular weight fungal compounds such polysaccharides and polysaccharide–protein complexes are significant for the exhibition of antitumor activity, due to their increased solubility in water [52]. The most active polysaccharides of Ganoderma belong to β-(1-3)-D-glucans, well known in promotion antitumor activity in animals and humans by acting as immune modulators – biological response modifiers, because they promote natural and acquired immunity of host organism itself [15].
In contrast, our results indicate that identified phenolic compounds (Table S1) and TCC are probably not responsible for detected antiproliferative activity, especially in G. resinaceum. This suggests that terpenoid compounds could play the main role in this activity [10], since various extracts from Ganoderma lucidum enriched with triterpenoids inhibit the growth of hepatoma cells by suppressing protein kinase C and activating mitogen-activated protein kinases [53]. Also, various types of ganoderic acids, including ganoderic acid T and its C-3 epimer compound, isolated from G. orbiforme, showed cytotoxic effect on MCF-7 cells [54], indicating that in this study triterpenoids could be responsible for high antiproliferative activity of G. resinaceum, as well. The results of the study of Rikame et al. [55] support this assumption, since terpenoids were quantified as major components in G. resinaceum extracts and exhibited cytotoxic activity against human colon HCT 116 cancer cells, while ganoresinoid A from this species alleviated LPS-induced apoptosis as described by Kou et al. [56], but further research should be conducted.
3.2.3. Lipid-Lowering Properties
Diabetes mellitus was induced in the animals, leading to dyslipidemia characteristic for this disease, as best observed in the group receiving a physiological solution for five days after diabetes induction. In this group and based on Table 2, a drastic increase in serum total triglyceride concentration (4.19 mmol/L) is evident, which is significantly higher compared to the total triglyceride values of the control group (1.14 mmol/L) and all other experimental groups treated with fungal extracts. Another laboratory parameter supporting diabetic dyslipidemia is the elevated level of LDL cholesterol fraction, significantly higher in the group of animals with diabetes treated with physiological solution compared to all other investigated groups.
The animal model of alloxan-induced diabetes effectively mirrors lipid metabolism disturbances in humans with hyperglycemia. This primarily leads to an increase in blood triglycerides as an alternative energy source, resulting in the elevation of very-low-density lipoproteins (VLDL) and subsequently the highly atherogenic LDL fraction. Therefore, considering the clinical therapeutic goal, the focus is primarily on lowering LDL cholesterol and secondarily on total triglycerides [57].
The valuable nutritional profile and benefits of using fungi in hyperglycemic conditions have been demonstrated in our previous studies [16,58,59]. In this study, it is evident that treatment with extracts from both fungal species of the genus Ganoderma prevented the disturbance in lipid status common in diabetes mellitus. There is even a tendency towards an increase in HDL cholesterol in certain groups of diabetic animals treated with fungal extracts, although unfortunately, statistical significance was not reached.
Furthermore, clinical studies have indicated a connection between dyslipidemia and the risk of cancer development, emphasizing that anything favorably impacting the lipid profile can be beneficial in prevention. In addition to showing antiproliferative potential, the examined Ganoderma species demonstrate a positive effect on in vivo lipid status, representing their additional pharmacological value [60,61].
In addition to lipid metabolism disturbances in diabetes, changes in renal and hepatic tissues also occur, as observed in this study. Serum levels of urea and creatinine, biochemical indicators of kidney function, were significantly elevated in the group of diabetic animals treated with physiological solution compared to the control group of healthy animals (urea – 46.98:7.40 mmol/L; creatinine – 289.25:52.25 mmol/L). In all normoglycemic animals treated with fungal extracts, the serum concentration of urea and creatinine remained at the level of the control group values, which was not the case for diabetic animals. Neither the H2O or EtOH extract of G. pfeifferi succeeded in preventing the increase in kidney function parameters; however, both extracts of G. resinaceum achieved this. Although it has been demonstrated that meroterpenoids from fungi of the genus Ganoderma exhibit renoprotective effects [62,63], we consider that the treatment duration with extracts in this study is too short for a direct impact on renal tissue. Nevertheless, it is possible that indirectly, through the regulation of glycemia, as shown in our previous research [16], there has been a positive influence on serum levels of urea and creatinine in the case of the G. resinaceum.
Regarding liver function, the ANOVA test indicated no statistically significant differences in AST values when comparing all groups of animals. In contrast to AST, the values of ALT, which is a more specific indicator of liver function, differ significantly. It is observed that serum ALT values are lower in all groups of animals treated with the extract of the G. resinaceum compared to all G. pfeifferi groups, as well as the control groups of normoglycemic and diabetic animals. Additionally, the ALT concentration in groups of diabetic animals treated with a physiological solution or G. pfeifferi extracts is significantly lower compared to the control normoglycemic group. However, it cannot be confidently claimed that a hepatoprotective effect has been demonstrated, only by considering the result that compared to normoglycemic control in the most groups of animals treated with fungal extracts a decrease in ALT values was evident. In our previous study, using the same model and experimental design, we showed that there were no noticeable histo-morphological changes in liver tissue indicating damage. The same study described that treatment with G. pfeifferi and G. resinaceum fungal extracts has a positive effect on liver biochemical parameters, with lipid peroxidation in the first place [16].
PCA analysis and correlation matrix for antidiabetic activity were also conducted to determine the correlation levels among this activity and determined bioactive compounds. PCA total variance was (76.07%), where PC1 was 58.66% and PC2 was 17.41% (Figure 6).
Regarding PCA analysis, a similar pattern emerged as seen with antioxidant and antiproliferative activity (Figure 6), where extracts from G. resinaceum distinctly separated from those of G. pfeifferi, along with all previously identified phenolics (Table S1) and TCC. This implies that the potent antidiabetic activity observed in G. resinaceum extracts cannot be solely attributed to phenolics and carbohydrates, suggesting that other compounds may likely play a pivotal role. To corroborate this, findings from other authors illustrate hepatoprotective effect of terpenoid compounds isolated from Ganoderma fungi, such as ganomycin, fornicatin A, D, and F [8,63]. However, the role of phenolic compounds in antidiabetic activity should not be ignored, since correlation matrix revealed that detected phenolic compounds (Table S1) showed significant positive correlation with measured parameters regarding lipid-lowering properties of examined extracts. This is also in accordance with our previous study [16] and Prabhakar [64], suggesting the necessity of further research and investigation of synergistic effects.
4. Conclusions
By comprehending the unique chemical compositions and therapeutic potentials of G. pfeifferi and G. resinaceum, these species could offer opportunities for the creation of innovative biopharmaceutical treatments, particularly for conditions which encompass both irregular cell growth and irregular lipid metabolism. The AAS quantification of micro and macroelements in G. pfeifferi and G. resinaceum provides valuable insights into the nutritional and therapeutic aspects of these medicinal fungal species. Further research exploring the biological activities associated with specific mineral compositions is warranted, as it could enhance our understanding of the potential health benefits conferred by these fungi.
Our findings underscore the therapeutic potential of Ganoderma extracts in ameliorating dyslipidemia associated with diabetes mellitus. Moreover, these extracts did not lead to organ damage; rather, they resulted in a decrease or no change in the biochemical parameters indicative of liver and kidney function compared to the control group. Further investigation into the underlying mechanisms and long-term effects of Ganoderma extracts on lipid metabolism and organ function is warranted for comprehensive understanding and clinical translation.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: LC-MS/MS detection of phenolic compounds in examined extracts.
Author Contributions
Conceptualization, M.R.; methodology, M.R., M.K. (Marko Kebert), S.K. (Sonja Kaišarević), N.S. and S.V.; validation, M.R., M.K. (Marko Kebert), S.K. (Sonja Kaišarević), N.S. and S.V.; formal analysis, M.R., M.K. (Marko Kebert), N.S. and S.V.; investigation, M.R., J.M. and M.K. (Marko Kebert); resources, M.R., M.K. (Marko Kebert), S.K. (Sonja Kaišarević), N.S. and S.V.; data curation, M.R., M.K. (Marko Kebert), S.K. (Saša Kostić), N.S. and S.V.; writing-original draft preparation, M.R. and J.M.; writing-review and editing, M.R., J.M., M.K. (Marko Kebert), S.K. (Saša Kostić), S.K. (Sonja Kaišarević), N.S., S.V. and M.K. (Maja Karaman); visualization, M.R. and S.K. (Saša Kostić); supervision, M.R. and M.K. (Maja Karaman); project administration, M.R., N.S., and S.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science, Technological Development, and Innovation of the Republic of Serbia, grant numbers 451-03-66/2024-03/200125, 451-03-65/2024-03/200125 and 451-03-47/2023-01/200114.
Institutional Review Board Statement
The experimental procedures were carried out following the guidelines of the European Directive (2010/63/EU) on animal experiments and received approval from the Ethics Committee for Protection and Welfare of Experimental Animals at the University of Novi Sad, Serbia.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors express their gratitude to retired professor Dr. Mira Popović for her invaluable assistance, knowledge transfer, and mentorship during the doctoral dissertation of Dr. Milena Rašeta.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Shamim, M.Z.; Mishra, A.K.; Kausar, T.; Mahanta, S.; Sarma, B.; Kumar, V.; Mishra, P.K.; Panda, J.; Baek, K.; Mohanta, Y.K. Exploring edible mushrooms for diabetes: unveiling their role in prevention and treatment. Molecules 2023, 28, 2837. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Márquez, L.; Trigos, Á.; Couttolenc, A.; Padrón, J.M.; Shnyreva, A.V.; Mendoza, G. Antiproliferative and antibacterial activity of extracts of Ganoderma strains grown in vitro. Food Sci. Biotechnol. 2021, 30, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Venturella, G.; Ferraro, V.; Cirlincione, F.; Gargano, M.L. Medicinal mushrooms: bioactive compounds, use, and clinical trials. Int. J. Mol. Sci. 2021, 22, 634. [Google Scholar] [CrossRef] [PubMed]
- Ekiz, E.; Oz, E.; Abd El-Aty, A.M.; Proestos, C.; Brennan, C.; Zeng, M.; Tomasevic, I.; Elobeid, T.; Çadırcı, K.; Bayrak, M.; Oz, F. Exploring the potential medicinal benefits of Ganoderma lucidum: from metabolic disorders to coronavirus infections. Foods 2023, 12, 1512. [Google Scholar] [CrossRef] [PubMed]
- Karaman, M.; Stahl, M.; Vesic, M.; Novaković, M.; Janjić, Lj.; Matavulj, M. Bioactive properties of wild-growing mushroom species Ganoderma applanatum (Pers.) Pat. from Fruška Gora Forest (Serbia). RPMP Ethnomed. Ther. Valid. 2012, 17, 339–356. [Google Scholar]
- Karaman, M.; Čapelja, E.; Rašeta, M.; Rakić, M. Diversity, Chemistry, and Environmental Contamination of Wild Growing Medicinal Mushroom Species as Sources of Biologically Active Substances (Antioxidants, Anti-Diabetics, and AChE Inhibitors). In Biology, Cultivation and Applications of Mushrooms; Arya, A., Rusevska, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2022; Volume 8, pp. 203–257. [Google Scholar] [CrossRef]
- Chen, X.-Q.; Chen, L.-X.; Zhao, J.; Tang, Y.-P.; Li, S.-P. Nortriterpenoids from the fruiting bodies of the mushroom Ganoderma resinaceum. Molecules 2017, 22, 1073. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, J.; Zhang, J.; Li, Z.; Liu, H.; Wang, Y. Traditional uses, chemical components and pharmacological activities of the genus Ganoderma P. Karst.: a review. RSC Adv. 2020, 10, 42084–42097. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.; Fu, H.; Lu, Y.; Han, Y.; Shen, Y.; Venkatakrishnan, K.; Golovinskaia, O.; Wang, C. Triterpenoids and polysaccharide peptides-enriched Ganoderma lucidum: a randomized, double-blind placebo-controlled crossover study of its antioxidation and hepatoprotective efficacy in healthy volunteers. Pharm. Biol. 2017, 55, 1041–1046. [Google Scholar] [CrossRef] [PubMed]
- Kolniak-Ostek, J.; Oszmianski, J.; Szyjka, A.; Moreira, H.; Barg, E. Anticancer and antioxidant activities in Ganoderma lucidum wild mushrooms in Poland, as well as their phenolic and triterpenoid compounds. Int. J. Mol. Sci. 2022, 23, 9359. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Sarikurkcu, C.; Gunes, E.; Uysal, A.; Ceylan, R.; Uysal, S.; Gungor, H.; Aktumsek, A. Two Ganoderma species: profiling of phenolic compounds by HPLC-DAD, antioxidant, antimicrobial and inhibitory activities on key enzymes linked to diabetes mellitus, Alzheimer’s disease and skin disorders. Food Funct. 2015, 6, 2794–2802. [Google Scholar] [CrossRef] [PubMed]
- Rašeta, M.; Popović, M.; Beara, I.; Šibul, F.; Zengin, G.; Krstić, S.; Karaman, M. Anti-inflammatory, antioxidant and enzyme inhibition activities in correlation with mycochemical profile of selected indigenous Ganoderma spp. from Balkan region (Serbia). Chem. Biodivers. 2021, 18, e2000828. [Google Scholar] [CrossRef] [PubMed]
- Karaman, M.; Mimica-Dukić, N.; Knežević, P.; Svirčev, Z.; Matavuly, M. Antibacterial properties of selected lignicolous mushrooms and fungi from northern Serbia. Int. J. Med. Mushrooms 2009, 11, 269–279. [Google Scholar] [CrossRef]
- Karaman, M.; Jovin, E.; Malbaša, R.; Matavulj, M.; Popović, M. Medicinal and edible lignicolous fungi as natural sources of antioxidative and antibacterial agents. Phytother. Res. 2010, 24, 1473–1481. [Google Scholar] [CrossRef]
- Rašeta, M.; Karaman, M.; Jakšić, M.; Šibul, F.; Kebert, M.; Novaković, A.; Popović, M. Mineral composition, antioxidant and cytotoxic biopotentials of wild-growing Ganoderma species (Serbia): G. lucidum (Curtis) P. Karst vs. G. applanatum (Pers.). Pat. Int. J. Food Sci. Technol. 2016, 51, 2583–2590. [Google Scholar] [CrossRef]
- Rašeta, M.; Popović, M.; Čapo, I.; Stilinović, N.; Vukmirović, S.; Milošević, B.; Karaman, M. Antidiabetic effect of two different Ganoderma species tested in alloxan diabetic rats. RSC Adv. 2020, 10, 10382–10393. [Google Scholar] [CrossRef] [PubMed]
- Rašeta, M.; Mišković, J.; Čapelja, E.; Zapora, E.; Petrović Fabijan, A.; Knežević, P.; Karaman, M. Do Ganoderma species represent novel sources of phenolic based antimicrobial agents? Molecules 2023, 28, 3264. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, Q.; Bu, M.; Hu, L.; Du, W.W.; Jiao, C.; Pan, H.; Sdiri, M.; Wu, N.; Xie, Y.; Yang, B.B. Ergosterol peroxide activates Foxo3-mediated cell death signaling by inhibiting AKT and c-Myc in human hepatocellular carcinoma cells. Oncotarget. 2016, 7, 33948–33959. [Google Scholar] [CrossRef] [PubMed]
- Sułkowska-Ziaja, K.; Zengin, G.; Gunia-Krzyzak, A.; ̇Popiół, J.; Szewczyk, A.; Jaszek, M.; Rogalski, J.; Muszyńska, B. Bioactivity and mycochemical profile of extracts from mycelial cultures of Ganoderma spp. Molecules 2022, 27, 275. [Google Scholar] [CrossRef] [PubMed]
- Sułkowska-Ziaja, K.; Balik, M.; Szczepkowski, A.; Trepa, M.; Zengin, G.; Kała, K.; Muszyńska, B. A review of chemical composition and bioactivity studies of the most promising species of Ganoderma spp. Diversity 2023, 15. [Google Scholar] [CrossRef]
- Gafforov, Y.; Umar, A.; Ghosh, S.; Tomšovský, M.; Yamaç, M.; Rašeta, M.; Yarasheva, M.; Wan-Mohtar, W.A.A.Q.I.; Rapior, S. Ganoderma adspersum (Schulzer) Donk; Ganoderma applanatum (Pers.), Pat., Ganoderma lucidum (Curtis), P., Karst., *!!! REPLACE !!!*, Ganoderma resinaceum Boud. – GANODERMATACEAE. In Ethnobiology of Uzbekistan (Ethnomedicinal Knowledge of Mountain, Communities), Khojimatov, O.K., Gafforov, Y., Bussman, R.W., Eds.; Springer Nature: Basel, Switzerland, 2023, pp. 1135–1169. [Google Scholar] [CrossRef]
- Sun, Y.F.; Xing, J.H.; He, X.L.; Wu, D.M.; Song, C.G.; Liu, S.; Vlasák, J.; Gates, G.; Gibertoni, T.B.; Cui, B.K. Species diversity, systematic revision and molecular phylogeny of Ganodermataceae (Polyporales, Basidiomycota) with an emphasis on Chinese collections. Stud. Mycol. 2022, 101, 287–415. [Google Scholar] [CrossRef] [PubMed]
- Loyd, A.L.; Richter, B.S.; Jusino, M.A.; Truong, C.; Smith, M.E.; Blanchette, R.A.; Smith, J.A. Identifying the “Mushroom of Immortality”: assessing the Ganoderma species composition in commercial Reishi products. Front. Microbiol. 2018, 16, 1557. [Google Scholar] [CrossRef]
- Fryssouli, V.; Zervakis, G.I.; Polemis, E.; Typas, M.A. A global meta-analysis of ITS rDNA sequences from material belonging to the genus Ganoderma (Basidiomycota, Polyporales) including new data from selected taxa. MycoKeys 2020, 75, 71–143. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.C.; Guo, W.L.; Li, L.; Yu, X.D.; Liu, B. Polysaccharide peptides from Ganoderma lucidum ameliorate lipid metabolic disorders and gut microbiota dysbiosis in high-fat diet-fed rats. J. Funct. Foods 2019, 57, 48–58. [Google Scholar] [CrossRef]
- Seweryn, E.; Ziała, A.; Gamian, A. Health-promoting of polysaccharides extracted from Ganoderma lucidum. Nutrients 2021, 13, 2725. [Google Scholar] [CrossRef] [PubMed]
- Aramabašić Jovanović, J.; Mihailović, M.; Uskoković, A.; Grdović, N.; Dinić, S.; Vidaković, M. The effects of major mushroom bioactive compounds on mechanisms that control blood glucose level. J. Fungi 2021, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Babamiri, S.; Mojani Qomi, M.S.; Shiehmorteza, M. The Efficacy of Ganoderma lucidum in overweight individuals: a randomized placebo-controlled trial. Mediterr. J. Nutr. Metab. 2022, 15, 263–271. [Google Scholar] [CrossRef]
- Ahmad, M.F.; Wahab, S.; Ahmad, F.A.; Ashraf, S.A.; Abullais, S.S.; Saad, H.H. Ganoderma lucidum: A potential pleiotropic approach of ganoderic acids in health reinforcement and factors influencing their production. Fungal Biol. Rev. 2022, 39, 100–125. [Google Scholar] [CrossRef]
- Klupp, N.L.; Kiat, H.; Bensoussan, A.; Steiner, G.Z.; Chang, D.H. A double-blind, randomised, placebo-controlled trial of Ganoderma lucidum for the treatment of cardiovascular risk factors of metabolic syndrome. Sci Rep. 2016, 6, 29540. [Google Scholar] [CrossRef] [PubMed]
- Kozarski, M.; Klaus, A.; Vunduk, J.; Jakovljevic, D.; Jadranin, M.B.; Nikšić, M.P. Health impact of the commercially cultivated mushroom Agaricus bisporus and wild-growing mushroom Ganoderma resinaceum - a comparative overview. J. Serb. Chem. Soc. 2020, 85, 721–735. [Google Scholar] [CrossRef]
- Yalcin, O.U.; Sarikurkcu, C.; Cengiz, M.; Gungor, H.; Ćavar Zeljković, S. Ganoderma carnosum and Ganoderma pfeifferi: metal concentration, phenolic content, and biological activity. Mycologia 2020, 112, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kebert, M.; Kostić, S.; Vuksanović, V.; Gavranović Markić, A.; Kiprovski, B.; Zorić, M.; Orlović, S. Metal- and organ-specific response to heavy metal-induced stress mediated by antioxidant enzymes’ activities, polyamines, and plant hormones levels in Populus deltoids. Plants 2022, 11, 3246. [Google Scholar] [CrossRef]
- Arnao, M.B.; Cano, A.; Acosta, M. The hydrophilic and lipophilic contribution to total antioxidant activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C.; Aruoma, O.I. The deoxyribose method: a simple test tube assay for determination of rate constants for reaction of hydroxyl radical. Anal. Biochem. 1987, 165, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Yen, G.C.; Chen, H.Y. Antioxidant activity of various tea extracts in relation to their antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. Ggplot2. WIREs Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Galili, T. Dendextend: an R package for visualizing, adjusting and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef] [PubMed]
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests. 2023. Available online: https://rpkgs.datanovia.com/rstatix/ (accessed on 5 May 2024).
- Anghileri, L.J. Magnesium, calcium and cancer. Magnes. Res. 2009, 22, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Ashique, S.; Kumar, S.; Hussain, A.; Mishra, N.; Garg, A.; Gowda, B.; Farid, A.; Gupta, G.; Dua, K.; Taghizadeh-Hesary, F. A narrative review on the role of magnesium in immune regulation, inflammation, infectious diseases, and cancer. J. Health Popul. Nutr. 2023, 42, 74. [Google Scholar] [CrossRef] [PubMed]
- Soetan, K.O.; Olaiya, C.O.; Oyewole, O.E. The importance of mineral elements for humans, domestic animals and plants: a review. Afr. J. Food Sci. 2010, 4, 200–222. [Google Scholar]
- Moon, D.O. Exploring the role of surface and mitochondrial ATP-sensitive potassium channels in cancer: from cellular functions to therapeutic potentials. Int. J. Mol. Sci. 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Marek, S.; Piotr, R.; Przemysław, N.; Anna, B.; Monika, G.; Kalač, P.; Agnieszka, J.; Sylwia, B.; Lidia, K.; Mirosław, M. Comparison of multielemental composition of Polish and Chinese mushrooms (Ganoderma spp.). Eur. Food Res. Technol. 2017, 243, 1555–1566. [Google Scholar] [CrossRef]
- Marreiro, D.D.N.; Cruz, K.J.C.; Morais, J.B.S.; Beserra, J.B.; Severo, J.S.; De Oliveira, A.R.S. Zinc and oxidative stress: current mechanisms. Antioxidants 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Demirci-Cekic, S.; Özkan, G.; Avan, A.N.; Uzunboy, S.; Çapanoğlu, E.; Apak, R. Biomarkers of oxidative stress and antioxidant defense. J. Pharm. Biomed. Anal. 2022, 209, 114477. [Google Scholar] [CrossRef] [PubMed]
- Rašeta, J.M.; Rakić, S.M.; Čapelja, V.E.; Karaman, A.M. Update on Research Data on the Nutrient Composition of Mushrooms and Their Potentials in Future Human Diets. In Edible Fungi: Chemical Composition, Nutrition and Health Effects; Stojković, D., Barros, L. Eds.; The Royal Society of Chemistry, Cambridge, United Kingdom, 2022; Chapter 2, pp. 27–67.
- Mišković, J.; Karaman, M.; Rašeta, M.; Krsmanović, N.; Berežni, S.; Jakovljević, D.; Piattoni, F.; Zambonelli, A.; Gargano, M.L.; Venturella, G. Comparison of two Schizophyllum commune strains in production of acetylcholinesterase inhibitors and antioxidants from submerged cultivation. J. Fungi 2021, 7, 115. [Google Scholar] [CrossRef] [PubMed]
- Vetter, J. The mushroom glucans: molecules of high biological and medicinal importance. Foods 2023, 12, 1009. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Zhang, X.; Hu, Y.S.; Wu, Y.; Wang, Q.Z.; Li, N.N.; Guo, Q.; Dong, X.C. Evaluation of in vivo antioxidant activities of Ganoderma lucidum polysaccharides in STZ-diabetic rats. Food Chem. 2009, 115, 32–36. [Google Scholar] [CrossRef]
- Shimizu, K.; Miyamoto, I.; Liu, J.; Konishi, F.; Kumamoto, S.; Kondo, R. Estrogen-like activity of ethanol extract of Ganoderma lucidum. J. Wood Sci. 2009, 55, 53–59. [Google Scholar] [CrossRef]
- Dasgupta, A.; Acharya, K. (2019). Mushrooms: an emerging resource for therapeutic terpenoids. 3 Biotech 2019, 9, 369. [Google Scholar] [CrossRef] [PubMed]
- Duru, M.E.; Çayan, G.T. Biologically active terpenoids from mushroom origin: a review. Rec. Nat. Prod. 2015, 9, 456. [Google Scholar]
- Rikame, T.N.; Ranawade, P.S.; Mittal, S.P.; Barvkar, V.T.; Borde, M.Y.; Tak, R.D. Characterization and biological studies of the terpenoids from Ganoderma resinaceum and Serpula similis (Agaricomycetes). Int. J. Med. Mushrooms 2023, 25, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Kou, R.W.; Xia, B.; Wang, Z.J.; Li, J.N.; Yang, J.R.; Gao, Y.Q.; Yin, X.; Gao, J.M. Triterpenoids and meroterpenoids from the edible Ganoderma resinaceum and their potential anti-inflammatory, antioxidant and anti-apoptosis activities. Bioorg. Chem. 2022, 121, 105689. [Google Scholar] [CrossRef] [PubMed]
- Kane, J.P.; Pullinger, C.R.; Goldfine, I.D.; Malloy, M.J. Dyslipidemia and diabetes mellitus: role of lipoprotein species and interrelated pathways of lipid metabolism in diabetes mellitus. Curr. Opin. Pharmacol. 2021, 61, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Sabo, A.; Stilinovic, N.; Vukmirovic, S.; Bukumiric, Z.; Capo, I.; Jakovljevic, V. Pharmacodynamic action of a commercial preparation of the mushroom Coprinus comatus in rats. Phytother. Res. 2010, 24, 1532–1537. [Google Scholar] [CrossRef] [PubMed]
- Stilinović, N.; Čapo, I.; Vukmirović, S.; Rašković, A.; Tomas, A.; Popović, M.; Sabo, A. Chemical composition, nutritional profile and in vivo antioxidant properties of the cultivated mushroom Coprinus comatus. R. Soc. Open Sci. 2020, 7, 200900. [Google Scholar] [CrossRef] [PubMed]
- de Jesus, M.; Mohammed, T.; Singh, M.; Tiu, J.G.; Kim, A.S. Etiology and management of dyslipidemia in patients with cancer. Front. Cardiovasc. Med. 2022, 9, 892335. [Google Scholar] [CrossRef] [PubMed]
- Neshat, S.; Rezaei, A.; Farid, A.; Sarallah, R.; Javanshir, S.; Ahmadian, S.; Chatrnour, G.; Daneii, P.; Heshmat Ghahdarijanet, K. The tangled web of dyslipidemia and cancer: is there any association? J. Res. Med. Sci. 2022, 27, 93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Qin, F.; Meng, X. , Yan, Y., Cheng, Y. Renoprotective ganodermaones A and B with rearranged meroterpenoid carbon skelotons from Ganoderma fungi. Bioorg. Chem. 2020, 100, 103930. [Google Scholar] [CrossRef]
- Peng, X.; Unsicker, S.B.; Gershenzon, J.; Qiu, M. Structural diversity, hypothetical biosynthesis, chemical synthesis, and biological activity of Ganoderma meroterpenoids. Nat. Prod. Rep. 2023, 40, 1354. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, P.K. Chapter 15 - Hypoglycemic potential of mushroom and their metabolites. New and Future Developments in Microbial Biotechnology and Bioengineering, Recent Advances in Application of Fungi and Fungal Metabolites: Applications in Healthcare 2020, pp. 197–208.
Figure 1.
a) Composition of macroelements (mg/g d.w.) and b) microelements (mg/kg) in G. pfeifferi and G. resinaceum samples.
Figure 1.
a) Composition of macroelements (mg/g d.w.) and b) microelements (mg/kg) in G. pfeifferi and G. resinaceum samples.
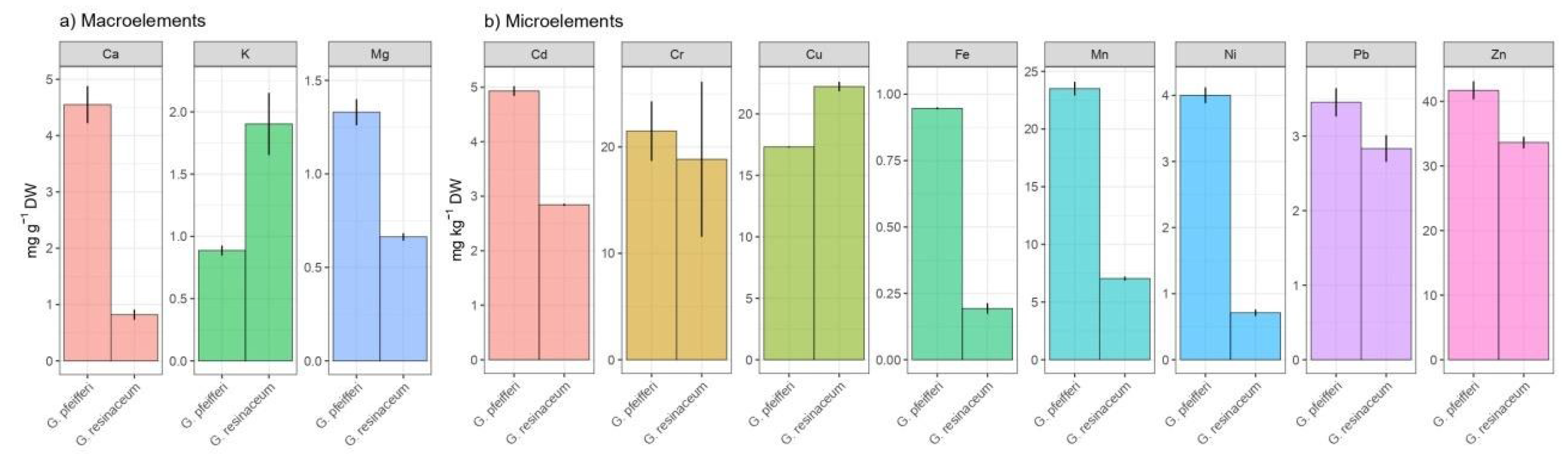
Figure 2.
Total carbohydrate content: (a) glucose, (b) sucrose, and (c) xylose. Different small letters indicate significant differences among different analyzed extracts of G. pfeifferi and G. resinaceum; Tukey’s significant difference (HSD) post hoc test (p ≤ 0.05). Data represent the mean ± standard deviation (SD).
Figure 2.
Total carbohydrate content: (a) glucose, (b) sucrose, and (c) xylose. Different small letters indicate significant differences among different analyzed extracts of G. pfeifferi and G. resinaceum; Tukey’s significant difference (HSD) post hoc test (p ≤ 0.05). Data represent the mean ± standard deviation (SD).
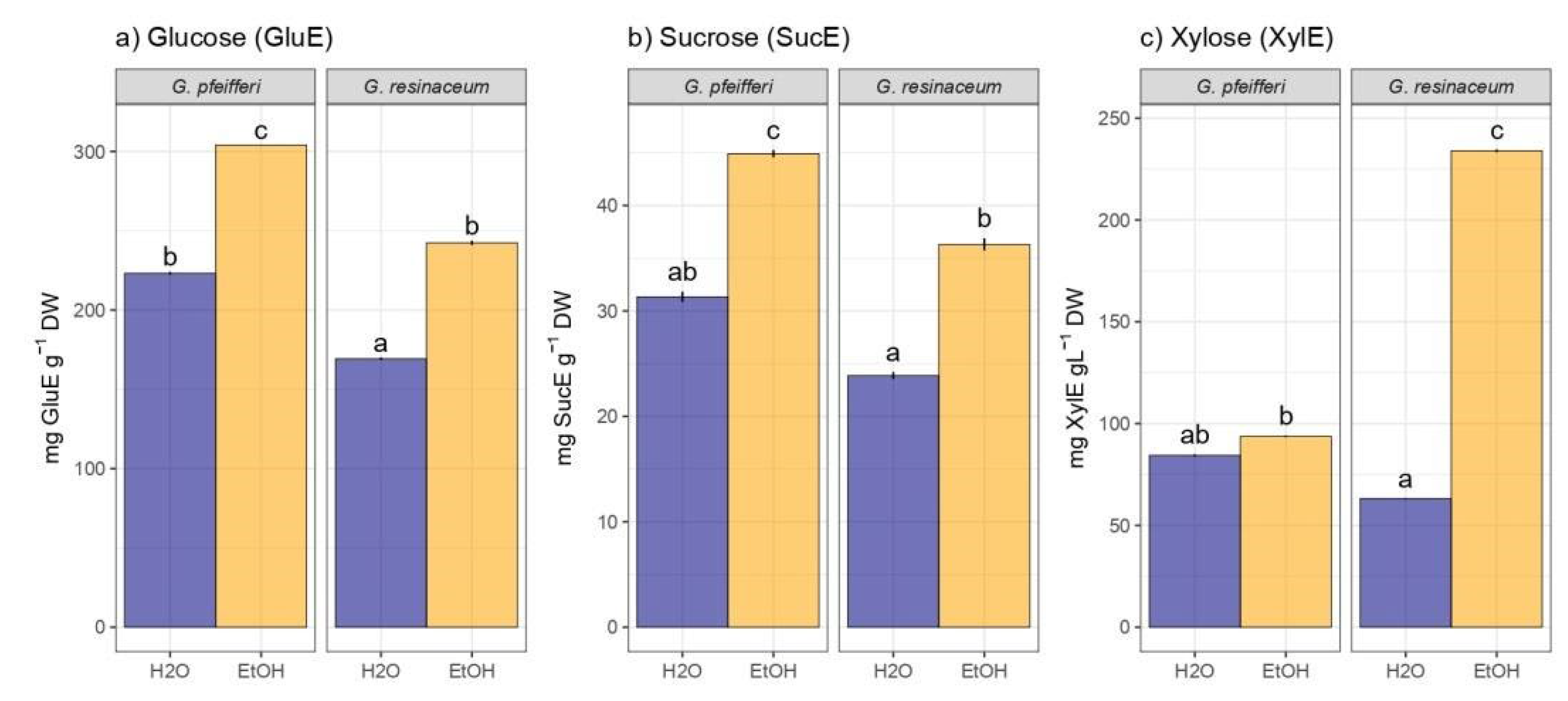
Figure 3.
Antioxidant activity in hot-water (H2O) and ethanolic (EtOH) extracts of G. pfeifferi and G. resinaceum: (a) RSC against ABTS radical (2,2′-azinobis-3-ethylbenzothiazoline-6-sulfonic acid); (b) radical scavenger capacity against OH radical; (c) ascorbate equivalent antioxidant capacity (A.E.A.C.) assay. Distinct lowercase letters denote significant differences observed among different analyzed samples including standard compound, propyl gallate (PG), as determined by Tukey’s honestly significant difference (HSD) post hoc test (p ≤ 0.05). The data are presented as the mean ± standard deviation (SD).
Figure 3.
Antioxidant activity in hot-water (H2O) and ethanolic (EtOH) extracts of G. pfeifferi and G. resinaceum: (a) RSC against ABTS radical (2,2′-azinobis-3-ethylbenzothiazoline-6-sulfonic acid); (b) radical scavenger capacity against OH radical; (c) ascorbate equivalent antioxidant capacity (A.E.A.C.) assay. Distinct lowercase letters denote significant differences observed among different analyzed samples including standard compound, propyl gallate (PG), as determined by Tukey’s honestly significant difference (HSD) post hoc test (p ≤ 0.05). The data are presented as the mean ± standard deviation (SD).
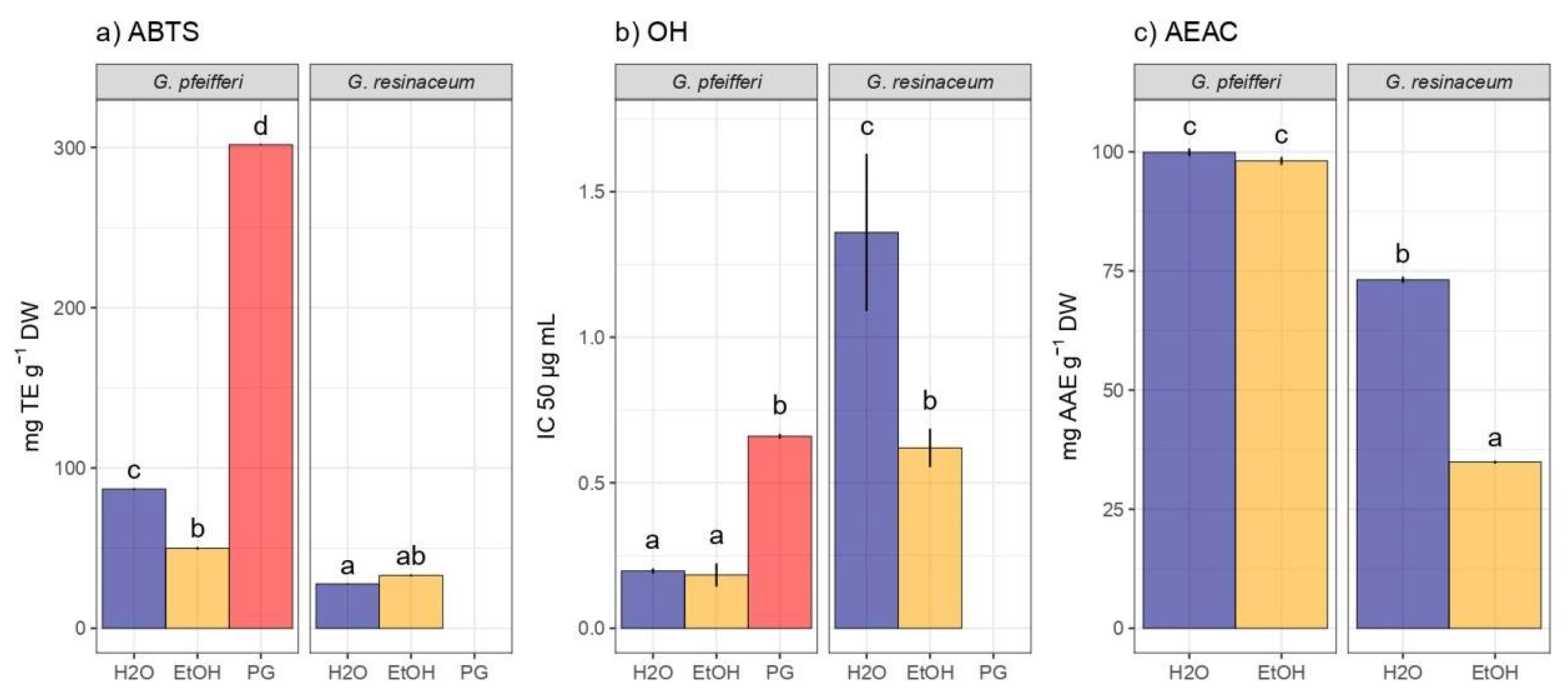
Figure 4.
Principal Component Analysis (PCA) of the observed antioxidant and antiproliferative activity along with phenolic compounds (Table S1) [16] and total carbohydrate content (TCC) in the four examined fungal extracts of G. pfeifferi and G. resinaceum. The following are the abbreviations of the examined parameters: EtOH - ethanolic extract; H2O - hot-water extract; ABTS - 2,2′-azinobis(3-ethylbenzothiozoline)-6-sulfonic acid; OH - hydroxyl radical; A.E.A.C. - ascorbate equivalent antioxidant capacity; IC50 24h/72h- antiproliferative inhibitory concentration in the case of incubation of 24h and 72h; GluE - glucose equivalents; SucE - sucrose equivalents; XylE - xylose equivalents; Tukey’s honestly significant difference (HSD) post hoc test (p ≤ 0.05). Data represent the mean ± standard deviation (SD).
Figure 4.
Principal Component Analysis (PCA) of the observed antioxidant and antiproliferative activity along with phenolic compounds (Table S1) [16] and total carbohydrate content (TCC) in the four examined fungal extracts of G. pfeifferi and G. resinaceum. The following are the abbreviations of the examined parameters: EtOH - ethanolic extract; H2O - hot-water extract; ABTS - 2,2′-azinobis(3-ethylbenzothiozoline)-6-sulfonic acid; OH - hydroxyl radical; A.E.A.C. - ascorbate equivalent antioxidant capacity; IC50 24h/72h- antiproliferative inhibitory concentration in the case of incubation of 24h and 72h; GluE - glucose equivalents; SucE - sucrose equivalents; XylE - xylose equivalents; Tukey’s honestly significant difference (HSD) post hoc test (p ≤ 0.05). Data represent the mean ± standard deviation (SD).
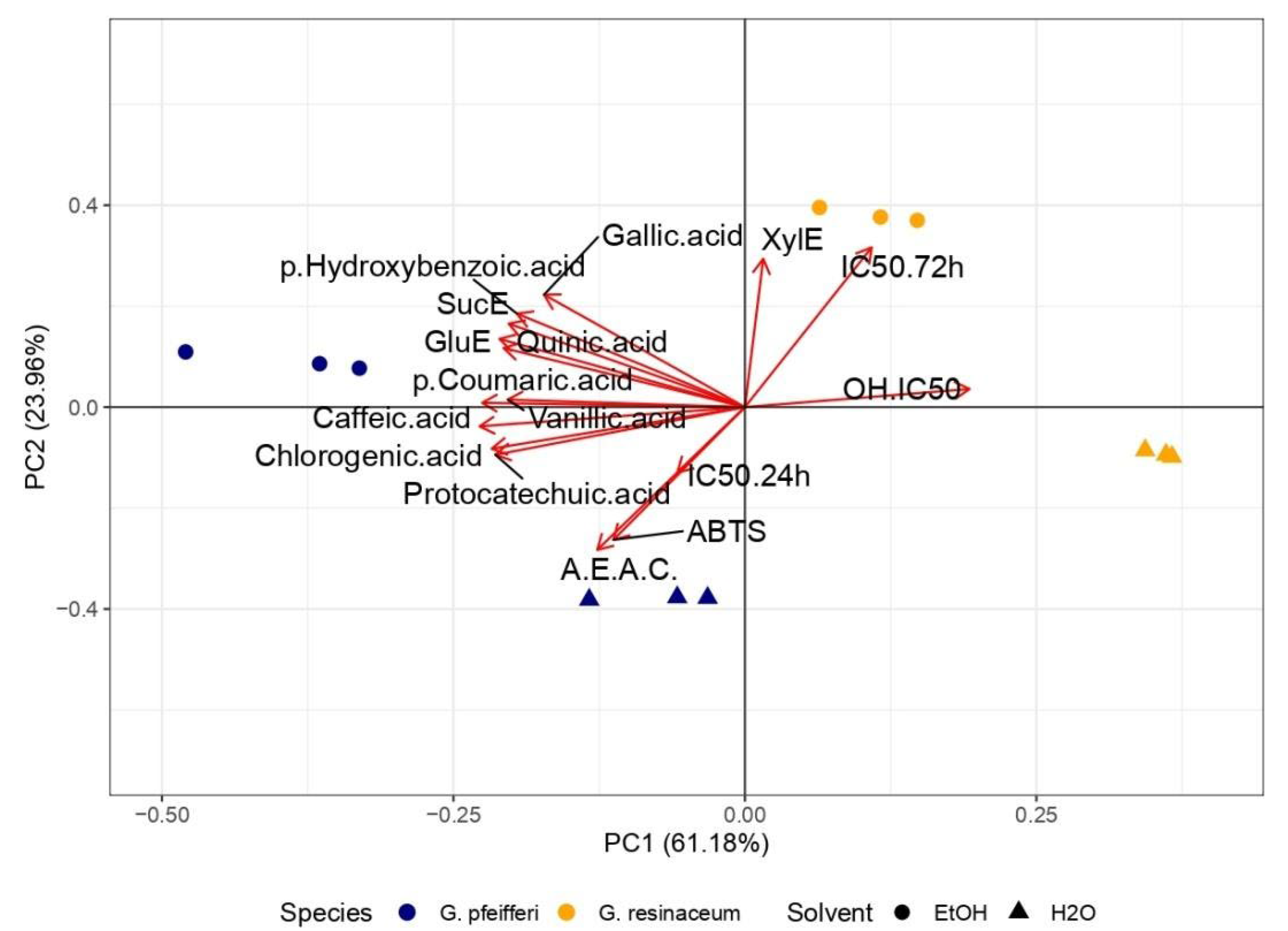
Figure 5.
Pearson’s coefficient of the correlation matrix of the examined parameters in ethanolic (EtOH) and hot-water (H2O) extracts of G. pfeifferi and G. resinaceum extracts collected on the territory of the Republic of Serbia. Blue squares represent a highly significant correlation of inspected parameters, while red squares present low interactions, assessed according to the corresponding Pearson’s coefficient. The following are the abbreviations of the examined parameters: ABTS—radical scavenger capacity against 2,2′-azinobis(3-ethylbenzothiozoline)-6-sulfonic acid, ABTS•+; OH—radical scavenger capacity against hydoxyl radical, OH•; A.E.A.C. - ascorbate equivalent antioxidant capacity; IC50 24h/72h- antiproliferative inhibitory concentration in the case of incubation of 24h and 72h; GluE - glucose equivalents content of TCC; SucE - sucrose equivalents content of TCC; XylE - xylose equivalents content of TCC; All.Sal - alloxan + saline; CHOL - cholesterol; T.TGC - total triglycerides; CREA - creatinine; ALT - alanine aminotransferase and AST - aspartate aminotransferase.
Figure 5.
Pearson’s coefficient of the correlation matrix of the examined parameters in ethanolic (EtOH) and hot-water (H2O) extracts of G. pfeifferi and G. resinaceum extracts collected on the territory of the Republic of Serbia. Blue squares represent a highly significant correlation of inspected parameters, while red squares present low interactions, assessed according to the corresponding Pearson’s coefficient. The following are the abbreviations of the examined parameters: ABTS—radical scavenger capacity against 2,2′-azinobis(3-ethylbenzothiozoline)-6-sulfonic acid, ABTS•+; OH—radical scavenger capacity against hydoxyl radical, OH•; A.E.A.C. - ascorbate equivalent antioxidant capacity; IC50 24h/72h- antiproliferative inhibitory concentration in the case of incubation of 24h and 72h; GluE - glucose equivalents content of TCC; SucE - sucrose equivalents content of TCC; XylE - xylose equivalents content of TCC; All.Sal - alloxan + saline; CHOL - cholesterol; T.TGC - total triglycerides; CREA - creatinine; ALT - alanine aminotransferase and AST - aspartate aminotransferase.
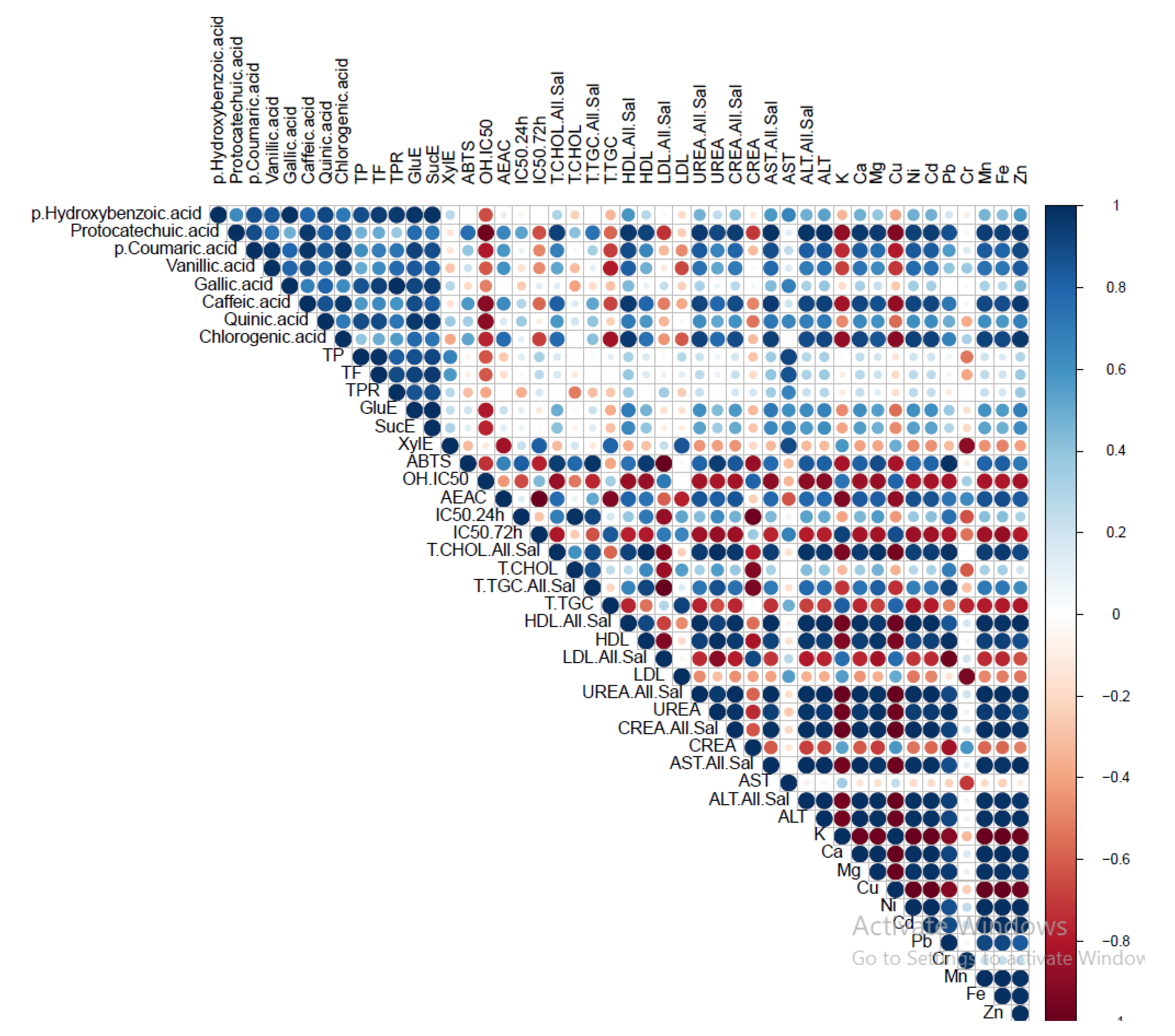
Figure 6.
Principal Component Analysis (PCA) of the lipid-lowering properties along with phenolic compounds (Table S1) [16] and total carbohydrate content (TCC) in the four examined fungal extracts of G. pfeifferi and G. resinaceum. The following are the abbreviations of the examined parameters: EtOH - ethanolic extract; H2O - hot-water extract; Allox.Sal - alloxan + saline; CHOL - cholesterol; T.TGC - total triglycerides; CREA - creatinine; ALT - alanine aminotransferase and AST - aspartate aminotransferase; Tukey’s honestly significant difference (HSD) post hoc test (p ≤ 0.05). Data represent the mean ± standard deviation (SD).
Figure 6.
Principal Component Analysis (PCA) of the lipid-lowering properties along with phenolic compounds (Table S1) [16] and total carbohydrate content (TCC) in the four examined fungal extracts of G. pfeifferi and G. resinaceum. The following are the abbreviations of the examined parameters: EtOH - ethanolic extract; H2O - hot-water extract; Allox.Sal - alloxan + saline; CHOL - cholesterol; T.TGC - total triglycerides; CREA - creatinine; ALT - alanine aminotransferase and AST - aspartate aminotransferase; Tukey’s honestly significant difference (HSD) post hoc test (p ≤ 0.05). Data represent the mean ± standard deviation (SD).
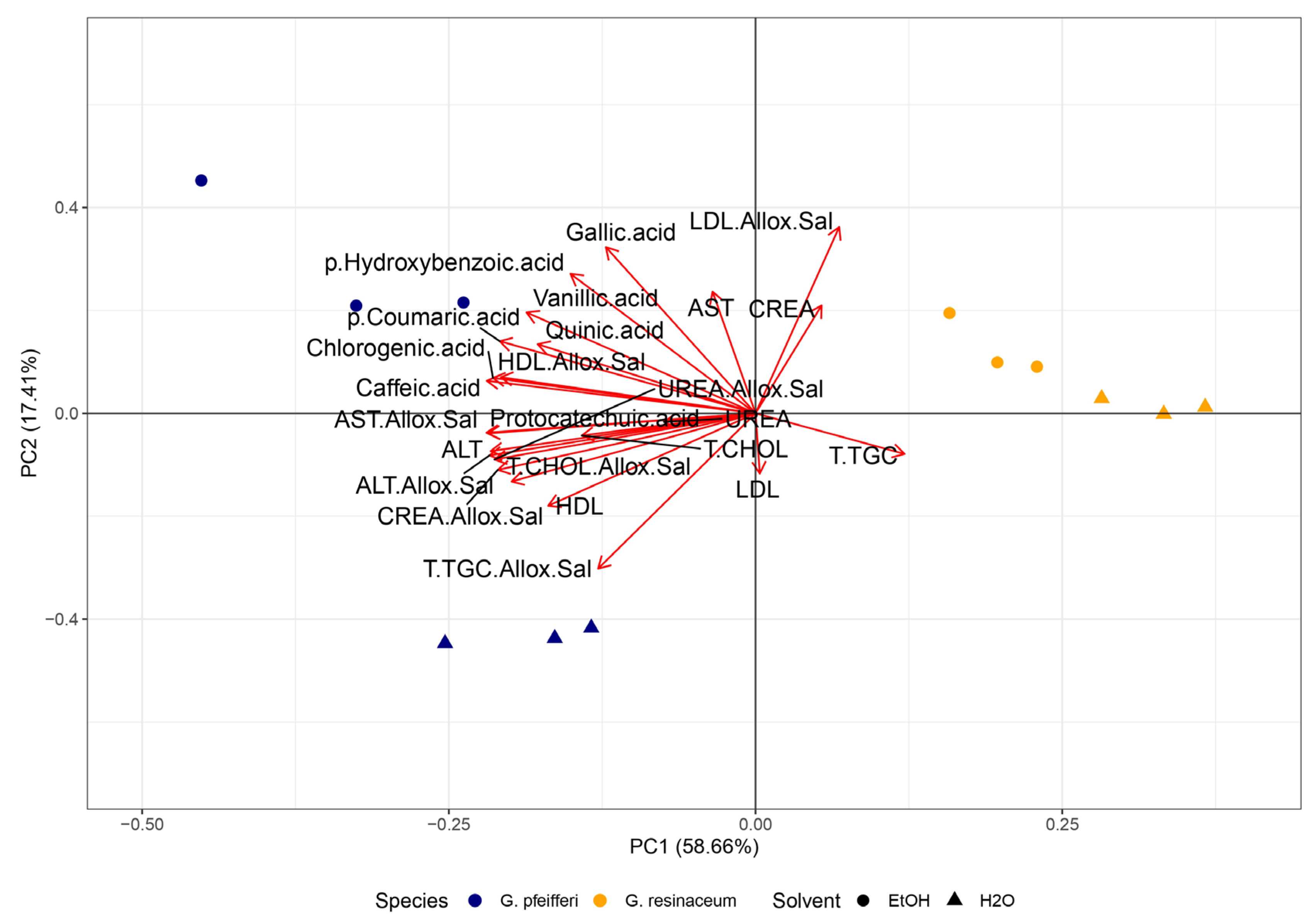

Table 1.
The antiproliferative effects of Ganoderma extracts and standard compounds on the MCF-7 cell line the MTT assay, IC50 values (μg/mL).
Table 1.
The antiproliferative effects of Ganoderma extracts and standard compounds on the MCF-7 cell line the MTT assay, IC50 values (μg/mL).
| Incubation period | Analyzed samples | ||||
|---|---|---|---|---|---|
|
G. pfeifferi EtOH |
G. pfeifferi H2O |
G. resinaceum EtOH |
G. resinaceum H2O |
Ellagic acid | |
| 24h | 154.05 ± 12.92c | 653.35 ± 10.19e | 363.87±1.51d | 4.88±0.50a | 33.94±3.69b |
| 72h | 78.33±1.89b | 49.25±1.72a | 181.07±0.21d | 113.33±0.62c | 43.06±1.22a |
IC50 – extract concentration required to inhibit cell growth by 50%. Values are expressed as mean ± SD of triplicates. a,b,c,d,e – different letters in the same row in comparison with ellagic acid as a standard compound indicate significant difference between extracts (ANOVA, Tukey post hoc, p < 0.05).
Table 2.
In vivo biochemistry parameters; The concentration of total cholesterol (mmol/L), triglycerides (TGC; mmol/L), HDL (mmol/L), and LDL (mmol/L) cholesterol, urea (mmol/L), and creatinine (mmol/L), as well as the enzymatic activity of aspartate aminotransferase (AST; U/L) and alanine aminotransferase (ALT; U/L) (mean value ± SD) in the serum of normoglycemic and diabetic (alloxan) rats treated with saline, EtOH and H2O extracts of the species G. pfeifferi and G. resinaceum.
Table 2.
In vivo biochemistry parameters; The concentration of total cholesterol (mmol/L), triglycerides (TGC; mmol/L), HDL (mmol/L), and LDL (mmol/L) cholesterol, urea (mmol/L), and creatinine (mmol/L), as well as the enzymatic activity of aspartate aminotransferase (AST; U/L) and alanine aminotransferase (ALT; U/L) (mean value ± SD) in the serum of normoglycemic and diabetic (alloxan) rats treated with saline, EtOH and H2O extracts of the species G. pfeifferi and G. resinaceum.
| Parameter | Control | Alloxan + saline | Alloxan + saline + G.p. EtOH | G.p. EtOH | Alloxan + saline + G.p. H2O | G.p. H2O |
Alloxan + saline + G.r. EtOH | G.r. EtOH | Alloxan + saline + G.r. H2O | G.r. H2O |
|
|---|---|---|---|---|---|---|---|---|---|---|---|
| Lipid status | Total cholesterol | 1.54±0.24a | 1.73±0.24a | 1.78±0.25a | 1.56±0.34a | 1.90±0.26a | 1.71±0.19a | 1.55±0.23a | 1.63±0.32a | 1.49±0.18a | 1.55±0.21a |
| Total TGC | 1.14±0.60a | 4.19±0.94b | 1.09±0.20a | 0.89±0.19a | 1.71±0.23a | 1.090.40a | 1.01±0.30a | 1.47±1.09a | 0.75±0.21a | 1.15±0.51a | |
| HDL | 0.90±0.17a | 0.75±0.17a | 1.01±0.16a | 0.90±0.21a | 0.97±0.25a | 0.98±0.15a | 0.79±0.17a | 0.78±0.10a | 0.77±0.15a | 0.740.12a | |
| LDL | 0.21±0.10a | 0.85±0.40b | 0.27±0.14a | 0.25±0.12a | 0.06±0.06a | 0.30±0.13a | 0.32±0.12a | 0.35±0.10a | 0.37±0.16a | 0.27±0.15a | |
| Renal function | Urea | 7.40±0.56a | 46.98±6.01b | 76.64±7.63c | 8.50±1.37a | 74.32±10.97c | 8.73±1.21a | 6.23±0.73a | 8.02±1.49a | 6.27±0.64a | 8.00±0.85a |
| Creatinine | 52.25±.62a | 289.25±73.87b | 379.00±79.09c | 51.00±3.03a | 395.40±97.39c | 48.50±2.43a | 48.50±4.72a | 50.40±9.02a | 50.00±6.45a | 52.83±8.47a | |
| Liver function | AST | 268.00±44.62a | 310.50±33.29a | 320.80±41.41a | 305.40±66.41a | 308.17±52.32a | 288.83±44.59a | 224.67±70.21a | 319.83±10.76a | 207.50±38.87a | 284.67±18.33a |
| ALT | 98.00±13.00c | 60.17±9.64b | 50.17±14.16b | 83.60±18.35c | 51.83±12.59b | 82.17±15.54c | 23.20±3.90a | 34.60±10.78a | 17.833.25a | 24.208.64a |
Data represent the mean ± standard deviation (SD). a, b, c Results that do not share the same superscript in the same column are statistically significant. p<0.01 (Tukey test, ANOVA).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



