
Submitted:
19 June 2024
Posted:
20 June 2024
You are already at the latest version
Abstract
Astrocytes are considered to possess a noticeable role in brain metabolism and as a partners in neuron-glia cooperation contribute to synthesis, bioconversions and regulation of the flux of substrates for neuronal metabolism. With the aim to investigate to which extend are human astrocytes metabolizing amino acids and by which compounds are they enriching their surrounding we employed the metabolomics analysis of their culture media by 1H-NMR. In addition, we compered the composition of media with either 5 mM or 25 mM glucose. The quantitative analysis of culture media by 1H-NMR revealed that astrocytes readely dispose from their milieu glutamine, branched-chain amino acids, and pyruvate with significantly high rate, while they enrich the culture media with lactate, branched-chain keto acids, citrate, acetate, ketone bodies, and alanine. The hyperglycemia suppressed the capacity of astrocytes to release branched-chain amino acids while stimulating the generation of ketone bodies. Our results highlight the active involvement of astrocytes in metabolism of several amino acids, and the regulation of key metabolic intermediates. The observed metabolic activities of astrocytes provide valuable insights into their roles in supporting neuronal function, brain metabolism, and intercellular metabolic interactions within the brain. Understanding the complex metabolic interactions between astrocytes and neurons is essential for elucidating brain homeostasis and the pathophysiology of neurological disorders.
Keywords:
astrocyte
; proton-nuclear magnetic resonance
; amino acids
; branched-chain 2-oxo acid
; branched-chain amino acid
; leucine
; isoleucine
; valine
; euglycemia
; hyperglycemia
; ketone body
; metabolomics
Introduction
Astrocytes are central to brain metabolism, performing essential tasks including energy production, neurotransmitter regulation, detoxification, and maintaining homeostasis [1,2,3,4,5]. These diverse roles ensure that the brain’s metabolic demands are met, neuronal function is supported, and the brain environment remains stable and protected from damage. The brain relies on a limited number of substances imported from the bloodstream to support its metabolism. The specificity and capacity of transporters expressed at the blood-brain barrier restrict the availability of polar and ionic substrates for brain metabolism [6]. Among these substrates, glucose is the primary energy source, with several glucose transporters maintaining its import capacity, surpassing that of all other compounds [7].
Recent data suggest that, in addition to glucose, amino acids can also serve as fuel molecules in energy production, playing a significant role in sustaining brain metabolism under various pathological and experimental conditions. Among the amino acids, neutral amino acids have a high importing capacity through specific transporters [8,9,10], implicating their involvement in brain metabolism [11,12,13,14,15,16]. Specifically, leucine, isoleucine, and valine are the most effectively imported into the human brain parenchyma. These three essential amino acids, known as branched-chain amino acids (BCAAs), have a common structural motif and serve several critical functions in the brain. BCAAs act as monomers for protein synthesis, nitrogen donors for de novo synthesis of glutamate and GABA, and balance the glutamate-glutamine cycle. Additionally, BCAAs can serve as alternative sources of acetyl-CoA and propionyl-CoA, contributing to brain anaplerotic, anabolic, and oxidative metabolism [11,12,13,14,15,16,17,18].
The biochemical aspects of astrocytic metabolism have been studied using animal models, animal cell cultures, and human cancer cell lines. Studies with cultured rat astrocytes have shown that these cells can catabolically convert BCAAs into several molecules [19,20,21], including ketone bodies [22]. The metabolic capacity of rat astrocytes correlates well with the expression of enzymes specific to BCAA catabolic pathways, observed in both cultured astrocytes and in situ [23,24,25].
Hyperglycemia, a major comorbidity of untreated diabetes, can affect brain metabolism [26,27,28], including the metabolism of amino acids [29,30]. It is hypothesized that hyperglycemia-induced changes in amino acid levels in the brain parenchyma may result from altered astrocytic metabolism. To investigate the extent to which increased glucose levels affect human astrocyte metabolism, we employed 1H-NMR analysis. This technique allowed us to identify changes in the ability of cultured human astrocytes to utilize components from their culture media and subsequently enrich their milieu with metabolic products. Our results revealed that human astrocytes, regardless of glucose levels, remove glutamine, BCAAs, and pyruvate from their medium. Additionally, they enrich the culture medium with several metabolic intermediates, including lactate, citrate, acetate, alanine, branched-chain keto acids (BCKAs), and ketone bodies. Experimental hyperglycemia influenced the ability of cultured human astrocytes to release BCKAs and ketone bodies into the culture medium. Estimating the ratio of BCKA to BCAA, which reflects the rate for BCKA entry into the irreversible part of BCAA catabolism, revealed that hyperglycemia supports the catabolism of carbon skeletons originating from BCAAs. Given that BCAAs and their cognate BCKAs play roles in both the synthesis of glutamate and the glutamate/glutamine cycle, the hyperglycemic effect on BCAA catabolism may contribute to the molecular changes underlying the pathoneurological aspects of hyperglycemia.
Materials and Methods
Chemicals
Nicotinamide adenine dinucleotide (NAD+), and 3-hydroxybutyrate dehydrogenase from Rhodobacter sphaeroides were purchased from Roche (Basel, Switzerland). DC Protein assay kit was from Bio-Rad Laboratories, Hercules, CA, USA. All other chemicals, namely glycine, hydrazine, lactate, 3-hydroxybutyrate, sodium 3-(trimethylsilyl)propionic-2,2,3,3-D4, and bovine serum albumin were purchased from Sigma (Sigma St. Louis, MO, USA).
Cell Culture
In this study, we used normal human astrocytes (NHA) obtained from Lonza Group (Basel, Switzerland, Cat. No. CC-2565). The human astrocyte cells were cultured in a Dulbecco’s Modified Eagle Medium (Gibco), supplemented with 10% (v/v) One Shot Fetal Bovine Serum (Gibco), 1% (v/v) N-2 Supplement, 100 U/ml penicillin, and 0,1 mg/ml streptomycin sulfate, which is a specific medium for astrocytes growth, at 37 °C, in an incubator containing 5% CO2. The culture medium was renewed every two days. The trypsinization were used for passaging after cells had reached approximately confluency of 80 – 90%, using 0,5 % trypsin solution.
1H-NMR Analysis
Astrocyte cells were cultured in 6-well plates to confluency. For experiment, the medium was replaced with a Dulbecco´s Modified Eagle Medium (DMEM) with 25 mM (Sigma, D6429) or 5 mM (Sigma, D6046) glucose level, which were enriched with 10 % fetal bovine serum and cultured at 37 °C for 24 hours. After incubation for 24 h, the medium was collected, centrifuged at 10 000 x g and stored at -80 °C untill the analysis.
For 1H-NMR analysis, 500 µL of culture medium was mixed with 100 µL of NMR solution (consisting from 200 mM phosphate buffer with pH 7.4 in D2O, enriched with [sodium 3-(trimethylsilyl)propionic-2,2,3,3-D4] (TMS-D4) to level 0.2 mM) and transferred to NMR tube. The 1H-NMR spectra were recorded on Bruker Avance III, 600 MHz ecquiper with TCI cryoprobe and subsequenlty processed as already described [31]. The concentration of metabolites in the culture medium before incubation was determined using external standards, where measured concentration was calculated through linear relation to the signals of known concentration.
Protein Estimation
For measurement of total protein concentration in all samples the commercially available DC Protein assay kit (Bio-Rad Laboratories, Hercules, CA, USA) was used. Bovine serum albumin was use as an standard. The measurement of absorbance was performed according to the manufacturer´s instructions using BioTek Synergy H4 hybrid microplate reader (Bio Tek, Winooski, VT, USA).
Cell Survival
The lactate dehydrogenase (LDH) enzymatic assay was performed to calculate the total and specific activity of LDH.
The reaction buffer 1 (RB1) consisted of a mixture of 0.1 M glycine/NaOH buffer with pH 9 with addition of 0.05 M hydrazine, 5 mM NAD+ and 10 mM lactate. For the assay, 30 μl of centrifuged medium or centrifuged lysate was mixed with 270 μl of RB1 in the 96-well plate. NADH formation in time was recorded at absorbance (λ = 340 nm). The measured data were used to calculate the enzymatic activity of LDH in the medium or lysates.
The cell viability was calculated as the ratio of LDH activity in lysates to total LDH activity according to method previosly described [32]. The total LDH activity is the sum of LDH activities estimated in lysates and media.
Enzymatic Estimation of 3-Hydroxybutyrate Level
The measurement of amount of released 3-hydroxybutyrate (3-OHB) in culture medium was performed as already described [31]. Briefly, the reaction buffer (RB2) consisted of a mixture of 0.1 M glycine/NaOH buffer with pH 9 with supplementation of 0.05 M hydrazine, 5 mM NAD+, and 3 µg/mL of purified 3-hydroxybutyrate dehydrogenase from Rhodobacter sphaeroides. A total of 30 μl of clarified supernatant prepared from denaturated and centrifuged culture medium was mixed with 270 μl of RB2 and the solution was immediately used for measurement of generated NADH, which was monitored at 340 nm by Synergy H4 microplate reader (Bio Tek, Winooski, VT, USA). The absorbance was monitored until the measured values reached plateau lasting at least 5 min. The calibration curve from increasing concentrations of the 3-OHB solution in phosphate buffer was used to quantify the 3-OHB released to culture medium. Subsequently, the apability of astrocyte cells to release the 3-OHB into culture medium was determined by considering the molar amount of released 3-OHB per mg of cellular proteins over a precisely defined incubation duration (24 hours).
Statistical Analysis
The results represent the mean ±SD, or ±SEM of six independent experiments. One-way analysis of variance with post hoc comparisons using the Student-Newman-Keuls test was performed to test for differences between test groups. Statistical analysis was performed using InStat software (GraphPad Software, USA).
Results
To demonstrate the effect of higher (25 mM) and normal (5 mM) glucose concentrations in culture medium on cultured human NHA astrocytes cells, the culture medium were subjected to metabolic analysis after a 24 hours incubation. We found, that the concentration of 5 and 25 mM glucose in the culture medium had no significant effect on total protein content of lysates from NHA cells, cell survival, and specific activity of lactate dehydrogenase in lysates (Table 1). The cell survival was calculated as the ratio of activity the lactate dehydrogenase in the culture medium released from dead cells to the sum of the activities of lactate dehydrogenase in the medium and lysates.
To asses the capability of astrocyte cells to metabolize amino acids and other metabolites from their culture medium, the culture medium with 25 mM or 5 mM glucose concentration was collected after a 24 hours incubation and lately subjected to 1H-NMR analysis. The spectra obtained from the 1H-NMR chromatogram (Figure 1) were used to calculate the residual concentrations of the metabolites composing the medium, and metabolites released into the culture medium (Table 2). The concentration of metabolites were determined by measuring their levels in the culture medium before incubation with cells against external standards. To estimate the specific uptake and release of some metabolites from or into the culture medium, the data obtained from 1H-NMR analysis were standardized to total protein content and incubation time (nmol*mg-1*h-1; Figure 2). Our data shown, that branched chain amino acids (BCAA) are mostly removed from culture medium compared to other amino acids (Table 2) and the significant changes of uptake of BCAA from culture media caused by higher glucose level were no observed (Figure 2A). Simultaneously, the presence of the peaks specific for branched chain keto acids (BCKA, Figure 1B,C) among obtained spectra points out the capability of cultured astrocytes cells to release BCKA into the culture medium (Figure 2B). The incubation of cells with 25 mM glucose revealed a significant decrease in the release of two BCKA, namely α-ketoisocaproate (KIC), and α-keto-methylvalerate (KMV), into the culture medium compare to 5 mM glucose. In addition, the appearance of specific peak for acetone (Figure 1D), ketone body, was identified on the spectra with higher intensity in the culture medium with 25 mM glucose (Figure 2C). The calculated ratios of released KIC to taken up leucine, KMV to isoleucine, and BCKA to BCAAs indicate that 50% ±10% of leucine molecules and 46% ±7% of isoleucine molecules taken up from the culture medium, were released as KIC, respectively KMV in the case of 5 mM glucose concentration in the culture medium, in additionally were significantly increased compared to 25 mM glucose level, KIC/Leu 35% ±4%; KMV/Ile 33% ±7% (Figure 2D). Indeed, the higher glucose concentration (25 mM) significantly increased the ratio of BCKA to BCAA, 48% ±8%, in comparison to normal (5 mM) glucose concentration in the culture medium, 64% ±11% (Figure 2D). Therefore, it could be assumed that in the case of higher glucose levels, a large amount of BCAA molecules undergo further intracellular metabolism.
The obtained 1H-NMR spectra did not allow the identification of the signal of two remaining ketone bodies, 3-hydroxybutyrate (3-OHB) and acetoacetate, and also their quantification. For this purpose, the spectrophotometric method for quantification of 3-hydroxybutyrate was performed. The estimation of the specific release of 3-OHB was calculated in the same way as for 1H-NMR method, thus the data obtained from the spectrophotometric analysis were standardized for total protein content and incubation time (nmol*mg-1*h-1; Table 2). This analysis showed, that specific release of the 3-OHB into the culture medium was significantly increased during 24 hours incubation with 25 mM glucose in comparison to 5 mM glucose in the culture medium (Figure 2C).
Discussion
The 1H-NMR analysis on the media from primary culture of human astroglial cells revealed the capability of cells to actively modify the composition of media by concurrent elimination and enrichment with several compounds. In addition, our study demonstrates that hyperglycemia significantly influences the metabolism of BCKAs, and ketone bodies by astrocytes. These findings are relevant for deducing the metabolic role of astrocytes in human brain and furthemore to the metabolic responses of the astroglial cells to hyperglycemia.
Under experimental euglycemic conditions astrocytes efficiently removed glucose, BCAAs, glutamine, and pyruvate from the medium, while enriching it with intermediates of cellular metabolism, such as lactate, citrate, acetate, alanine, BCKAs, and ketone bodies. This metabolic profile underscores the versatility of astrocytes in utilizing multiple substrates to sustain their energy and biosynthetic demands, thereby contributing to the overall metabolic homeostasis of the brain in euglycemic conditions.
Astrocytes are recognized for their crusial role in brain energy metabolism, particulary in conversion of glucose to lactate [33,34,35,36,37,38]. Studies with animal models have demostrated that rodent astrocytes possess high specific capacity of lactate release [39], surpassing the estimated values for cultured human astroglial cells (Table 2). Nonetheless, the observed capability of human astroglial cells to release lactate supports the hypothesis of an astrocyte-neuron lactate shuttle, which supports the energy metabolism of neurons [40]. Additionally, astrocytic release of lactate may serve signaling roles in the brain [41].
Although human astrocytes can import pyruvate from the extracellular space (Table 2), the estimated quantity of its import accounts for only a fraction of the total amount of released lactate. Similar to animal astrocytes, human astroglial cells are expected to generate most of the lactate through anaerobic glycolysis of glucose [38]. Indeed, the cultured human astroglial cells readely disposed glutamine, an amino acid with several distinct functions in cellular metabolism and synthesis of signaling molecules. Glutamine’s structure allows it to contribute to the cellular metabolism of nitrogen-containing compounds. Additionally, brain glutamine is essential for sustaining the levels of glutamate and gamma-aminobutyrate in the brain parenchyma [42]. Furthermore, the carbon skeleton of glutamine can play an anaplerotic role and may enter energetic metabolism, being oxidized to carbon dioxide in mitochondrial [33].
The catabolism of glutamine can provide the amino group for transamination of pyruvate to alanine. The later may be released from astrocytes for either to dispose the superfluous nitrogen from astrocytes, to exchange the amino group with neurons or to support neuronal mitochondrial metabolism [43,44]. The analyzed media were also enriched with pyroglutamate, a compound that can be generated either spontaneously through cyclization of glutamate and glutamine or enzymatically from glutathione [45].
Moreover, human astroglial cells dispose of a substantial amount of BCAAs, and their irreversible catabolism appeard to be glucose-dependent. Hyperglycemia enhances the disappearance of BCKAs and increases release of ketone bodies into the culture medium. This suggests that human astrocytes may be capable of endogenously generating ketone bodies in the brain parenchyma. Even previous data showed that culured rat astroglial cells generate substantial amount of the ketone bodies from leucine [22] under hyperglycemic conditions the ability of astrocytes to generate ketone bodies at euglycemic conditions remained opened question.
The elevated production and release of ketone bodies under hyperglycemia highlight the metabolic plasticity of human astrocytes. Ketone bodies, typically produced during states of decreased glucose level or insulin availability, are now synthesized in response to hyperglycemia, indicating a robust metabolic adaptability of astrocytes to varying glucose levels. This ketogenesis may provide an auxiliary energy source for neighboring neurons [46], supporting neuronal function and contribute to intercellular communication between glial and neurons [46,47].
In contrast to BCAAs, the levels of three essential amino acids, namely: lysine, methionine and tyrosine; were significantly increased in media with euglycemic conditions. The levels of remaining essential amino acids did not changed significantly either at euglycemic nor hyperglycemic media (Table 2). The possibility of increased levels of some of essential amino acids in culture media from astrocytes has already been documented [22], but not studied in detail. However, it could be hypothesized, that similar to human glioma cells that also astrocytes are capable to use extracellular proteins as the putative source of amino acids [48]. An other aspect, that may affect the overall metabolism of essential amino acids, could be the proteosynthetic ratio of the cells. Since we used confluent cultres of astrocytes, the growth and dividing of the cells may be expected to be restricted by contact inhibition of proliferation [49] and therefore the uptake of those essential amino acids, which are not further catabolized may be minimal.
In conclusion, our study elucidates the dynamic role of astrocytes in maintaining brain metabolic homeostasis through substrate flexibility and adaptive metabolic responses. Our results underscore the importance of astrocytes not only as supportive glial cells but as active participants in brain metabolism, capable of responding to and modulating metabolic challenges. These findings enhance our understanding of astrocytic contributions to brain metabolism, particularly under pathological conditions such as hyperglycemia. Future research should aim to delineate the signaling pathways and regulatory mechanisms underlying these metabolic adaptations, which could uncover potential therapeutic targets for metabolic disorders affecting the brain.
Acknowledgement:
Data Availability
All data generated or analysed during this study are included in this published article. Enquiries about data availability should be directed to the authors.
Cometing interests:
The authors declare no competing interests.
Funding
This work was supported by project VEGA 1/0042/24.
References
- Beard, E.; Lengacher, S.; Dias, S.; Magistretti, P.J.; Finsterwald, C. Astrocytes as Key Regulators of Brain Energy Metabolism: New Therapeutic Perspectives. Front Physiol 2022, 12, 825816. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, M.; Allaman, I.; Magistretti, P.J. Brain Energy Metabolism: Focus on Astrocyte-Neuron Metabolic Cooperation. Cell Metabolism 2011, 14, 724–738. [Google Scholar] [CrossRef] [PubMed]
- Bonvento, G.; Bolaños, J.P. Astrocyte-Neuron Metabolic Cooperation Shapes Brain Activity. Cell Metabolism 2021, 33, 1546–1564. [Google Scholar] [CrossRef]
- Hamprecht, B.; Verleysdonk, S.; Wiesinger, H. Enzymes of Carbohydrate and Energy Metabolism. In Neuroglia; Kettenmann, H., Ransom, B.R., Eds.; Oxford University Press, 2004; p. 0 ISBN 978-0-19-515222-7.
- Verkhratsky, A.; Semyanov, A. The Great Astroglial Metabolic Revolution: Mitochondria Fuel Astrocyte Homeostatic Support and Neuroprotection. Cell Calcium 2022, 104, 102583. [Google Scholar] [CrossRef] [PubMed]
- Daneman, R.; Prat, A. The Blood–Brain Barrier. Cold Spring Harb Perspect Biol 2015, 7, a020412. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. Brain Metabolism: A Perspective from the Blood-Brain Barrier. Physiol Rev 1983, 63, 1481–1535. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.D.; Ye, M.; Levy, A.F.; Rothstein, J.D.; Bergles, D.E.; Searson, P.C. The Blood-Brain Barrier: An Engineering Perspective. Front. Neuroeng. 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Smith, Q.R. Transport of Glutamate and Other Amino Acids at the Blood-Brain Barrier. The Journal of Nutrition 2000, 130, 1016S–1022S. [Google Scholar] [CrossRef] [PubMed]
- Zaragozá, R. Transport of Amino Acids Across the Blood-Brain Barrier. Front Physiol 2020, 11, 973. [Google Scholar] [CrossRef] [PubMed]
- Yudkoff, M.; Daikhin, Y.; Nissim, I.; Horyn, O.; Luhovyy, B.; Lazarow, A.; Nissim, I. Brain Amino Acid Requirements and Toxicity: The Example of Leucine. The Journal of Nutrition 2005, 135, 1531S–1538S. [Google Scholar] [CrossRef] [PubMed]
- Conway, M.E.; Hutson, S.M. BCAA Metabolism and NH3 Homeostasis. In The Glutamate/GABA-Glutamine Cycle; Schousboe, A., Sonnewald, U., Eds.; Advances in Neurobiology; Springer International Publishing: Cham, 2016; ISBN 978-3-319-45094-0. [Google Scholar]
- Griffin, J.W.D.; Bradshaw, P.C. Amino Acid Catabolism in Alzheimer’s Disease Brain: Friend or Foe? Oxidative Medicine and Cellular Longevity 2017, 2017, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sperringer, J.E.; Addington, A.; Hutson, S.M. Branched-Chain Amino Acids and Brain Metabolism. Neurochem Res 2017, 42, 1697–1709. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Wu, G. Metabolism of Amino Acids in the Brain and Their Roles in Regulating Food Intake. In Amino Acids in Nutrition and Health; Wu, G., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, 2020; ISBN 978-3-030-45327-5. [Google Scholar]
- Salcedo, C.; Andersen, J.V.; Vinten, K.T.; Pinborg, L.H.; Waagepetersen, H.S.; Freude, K.K.; Aldana, B.I. Functional Metabolic Mapping Reveals Highly Active Branched-Chain Amino Acid Metabolism in Human Astrocytes, Which Is Impaired in iPSC-Derived Astrocytes in Alzheimer’s Disease. Frontiers in Aging Neuroscience 2021, 13, 553. [Google Scholar] [CrossRef] [PubMed]
- Suryawan, A.; Hawes, J.W.; Harris, R.A.; Shimomura, Y.; Jenkins, A.E.; Hutson, S.M. A Molecular Model of Human Branched-Chain Amino Acid Metabolism. The American Journal of Clinical Nutrition 1998, 68, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Fernstrom, J.D. Branched-Chain Amino Acids and Brain Function. The Journal of Nutrition 2005, 135, 1539S–1546S. [Google Scholar] [CrossRef] [PubMed]
- Bixel, M.G.; Engelmann, J.; Willker, W.; Hamprecht, B.; Leibfritz, D. Metabolism of [U-(13)C]Leucine in Cultured Astroglial Cells. Neurochem Res 2004, 29, 2057–2067. [Google Scholar] [CrossRef] [PubMed]
- Murín, R.; Mohammadi, G.; Leibfritz, D.; Hamprecht, B. Glial Metabolism of Isoleucine. Neurochemical Research 2009, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Murín, R.; Mohammadi, G.; Leibfritz, D.; Hamprecht, B. Glial Metabolism of Valine. Neurochemical Research 2009, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Bixel, M.G.; Hamprecht, B. Generation of Ketone Bodies from Leucine by Cultured Astroglial Cells. J Neurochem 1995, 65, 2450–2461. [Google Scholar] [CrossRef] [PubMed]
- Bixel, M.G.; Hamprecht, B. Immunocytochemical Localization of Beta-Methylcrotonyl-CoA Carboxylase in Astroglial Cells and Neurons in Culture. J Neurochem 2000, 74, 1059–1067. [Google Scholar] [CrossRef] [PubMed]
- Murín, R.; Verleysdonk, S.; Rapp, M.; Hamprecht, B. Immunocytochemical Localization of 3-Methylcrotonyl-CoA Carboxylase in Cultured Ependymal, Microglial and Oligodendroglial Cells. Journal of Neurochemistry 2006, 1393–1402. [Google Scholar] [CrossRef] [PubMed]
- Murín, R.; Schaer, A.; Kowtharapu, B.S.; Verleysdonk, S.; Hamprecht, B. Expression of 3-Hydroxyisobutyrate Dehydrogenase in Cultured Neural Cells. Journal of Neurochemistry 2008, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Serrano, A.M.; Duarte, J.M.N. Brain Metabolism Alterations in Type 2 Diabetes: What Did We Learn From Diet-Induced Diabetes Models? Front Neurosci 2020, 14, 229. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.J.; Jiang, L.; Hamza, M.; Rangel, E.S.; Dai, F.; Belfort-DeAguiar, R.; Parikh, L.; Koo, B.B.; Rothman, D.L.; Mason, G.; et al. Blunted Rise in Brain Glucose Levels during Hyperglycemia in Adults with Obesity and T2DM. JCI Insight 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Sonneville, R.; den Hertog, H.M.; Güiza, F.; Gunst, J.; Derese, I.; Wouters, P.J.; Brouland, J.-P.; Polito, A.; Gray, F.; Chrétien, F.; et al. Impact of Hyperglycemia on Neuropathological Alterations during Critical Illness. The Journal of Clinical Endocrinology & Metabolism 2012, 97, 2113–2123. [Google Scholar] [CrossRef]
- Sickmann, H.M.; Waagepetersen, H.S. Effects of Diabetes on Brain Metabolism – Is Brain Glycogen a Significant Player? Metab Brain Dis 2015, 30, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Bai, M.; Xie, X.; Wang, J.; Weng, C.; Dai, H.; Chen, J.; Han, F.; Lin, W. Impaired Amino Acid Metabolism and Its Correlation with Diabetic Kidney Disease Progression in Type 2 Diabetes Mellitus. Nutrients 2022, 14, 3345. [Google Scholar] [CrossRef] [PubMed]
- Gondáš, E.; Kráľová Trančíková, A.; Baranovičová, E.; Šofranko, J.; Hatok, J.; Kowtharapu, B.S.; Galanda, T.; Dobrota, D.; Kubatka, P.; Busselberg, D.; et al. Expression of 3-Methylcrotonyl-CoA Carboxylase in Brain Tumors and Capability to Catabolize Leucine by Human Neural Cancer Cells. Cancers 2022, 14, 585. [Google Scholar] [CrossRef] [PubMed]
- Gondáš, E.; Kráľová Trančíková, A.; Šofranko, J.; Majerová, P.; Lučanský, V.; Dohál, M.; Kováč, A.; Murín, R. The Presence of Pyruvate Carboxylase in the Human Brain and Its Role in the Survival of Cultured Human Astrocytes. Physiol Res 2023, 72, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Schousboe, A.; Westergaard, N.; Sonnewald, U.; Petersen, S.B.; Huang, R.; Peng, L.; Hertz, L. Glutamate and Glutamine Metabolism and Compartmentation in Astrocytes. Dev Neurosci 1993, 15, 359–366. [Google Scholar] [CrossRef]
- Magistretti, P.J. Neuron-Glia Metabolic Coupling and Plasticity. J Exp Biol 2006, 209, 2304–2311. [Google Scholar] [CrossRef] [PubMed]
- Magistretti, P.J.; Sorg, O.; Yu, N.; Martin, J.-L.; Pellerin, L. Neurotransmitters Regulate Energy Metabolism in Astrocytes: Implications for the Metabolic Trafficking between Neural Cells. Dev Neurosci 1993, 15, 306–312. [Google Scholar] [CrossRef]
- Magistretti, P.J.; Allaman, I. Lactate in the Brain: From Metabolic End-Product to Signalling Molecule. Nat Rev Neurosci 2018, 19, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Swanson, R.A.; Benington, J.H. Astrocyte Glucose Metabolism under Normal and Pathological Conditions in Vitro. Dev Neurosci 1996, 18, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Wiesinger, H.; Hamprecht, B.; Dringen, R. Metabolic Pathways for Glucose in Astrocytes. Glia 1997, 21, 22–34. [Google Scholar] [CrossRef]
- Walz, W.; Mukerji, S. Lactate Production and Release in Cultured Astrocytes. Neuroscience Letters 1988, 86, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Pellerin, L.; Pellegri, G.; Bittar, P.G.; Charnay, Y.; Bouras, C.; Martin, J.-L.; Stella, N.; Magistretti, P.J. Evidence Supporting the Existence of an Activity-Dependent Astrocyte-Neuron Lactate Shuttle. Dev Neurosci 1998, 20, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Horvat, A.; Zorec, R.; Vardjan, N. Lactate as an Astroglial Signal Augmenting Aerobic Glycolysis and Lipid Metabolism. Front. Physiol. 2021, 12, 735532. [Google Scholar] [CrossRef] [PubMed]
- Schousboe, A.; Scafidi, S.; Bak, L.K.; Waagepetersen, H.S.; McKenna, M.C. Glutamate Metabolism in the Brain Focusing on Astrocytes. In Glutamate and ATP at the Interface of Metabolism and Signaling in the Brain; Parpura, V., Schousboe, A., Verkhratsky, A., Eds.; Advances in Neurobiology; Springer International Publishing: Cham, 2014; ISBN 978-3-319-08893-8. [Google Scholar]
- Rabah, Y.; Francés, R.; Minatchy, J.; Guédon, L.; Desnous, C.; Plaçais, P.-Y.; Preat, T. Glycolysis-Derived Alanine from Glia Fuels Neuronal Mitochondria for Memory in Drosophila. Nat Metab 2023, 5, 2002–2019. [Google Scholar] [CrossRef] [PubMed]
- Schousboe, A.; Sonnewald, U.; Waagepetersen, H.S. Differential Roles of Alanine in GABAergic and Glutamatergic Neurons. Neurochemistry International 2003, 43, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bachhawat, A.K. Pyroglutamic Acid: Throwing Light on a Lightly Studied Metabolite. Current Science 2012, 102, 288–297. [Google Scholar]
- Jensen, N.J.; Wodschow, H.Z.; Nilsson, M.; Rungby, J. Effects of Ketone Bodies on Brain Metabolism and Function in Neurodegenerative Diseases. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- García-Rodríguez, D.; Giménez-Cassina, A. Ketone Bodies in the Brain Beyond Fuel Metabolism: From Excitability to Gene Expression and Cell Signaling. Front. Mol. Neurosci. 2021, 14, 732120. [Google Scholar] [CrossRef] [PubMed]
- Buser, D.P.; Ritz, M.-F.; Moes, S.; Tostado, C.; Frank, S.; Spiess, M.; Mariani, L.; Jenö, P.; Boulay, J.-L.; Hutter, G. Quantitative Proteomics Reveals Reduction of Endocytic Machinery Components in Gliomas. EBioMedicine 2019, 46, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Pavel, M.; Renna, M.; Park, S.J.; Menzies, F.M.; Ricketts, T.; Füllgrabe, J.; Ashkenazi, A.; Frake, R.A.; Lombarte, A.C.; Bento, C.F.; et al. Contact Inhibition Controls Cell Survival and Proliferation via YAP/TAZ-Autophagy Axis. Nat Commun 2018, 9, 2961. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Representative 1H-NMR spectra of branched-chain amino acids (BCAA), leucine, isoleucine, and valine (A); branched-chain keto acids (BCKA), 2-oxoisocaproate (B), 2-oxoisovalerate, and 3-methyl-2-oxopentanonate (C); as well as of acetone (D). The spectra were recorded for DMEM/FBS media before incubation (black line) or after 24-hour incubation of astocytes in media either with 5 mM (5 mM glucose; green line) or 25 mM (25 mM glucose; blue line) glucose level. The ranges of chemichal shifts (δ) for BCAA, BCKA, and acetone are depicted.
Figure 1.
Representative 1H-NMR spectra of branched-chain amino acids (BCAA), leucine, isoleucine, and valine (A); branched-chain keto acids (BCKA), 2-oxoisocaproate (B), 2-oxoisovalerate, and 3-methyl-2-oxopentanonate (C); as well as of acetone (D). The spectra were recorded for DMEM/FBS media before incubation (black line) or after 24-hour incubation of astocytes in media either with 5 mM (5 mM glucose; green line) or 25 mM (25 mM glucose; blue line) glucose level. The ranges of chemichal shifts (δ) for BCAA, BCKA, and acetone are depicted.
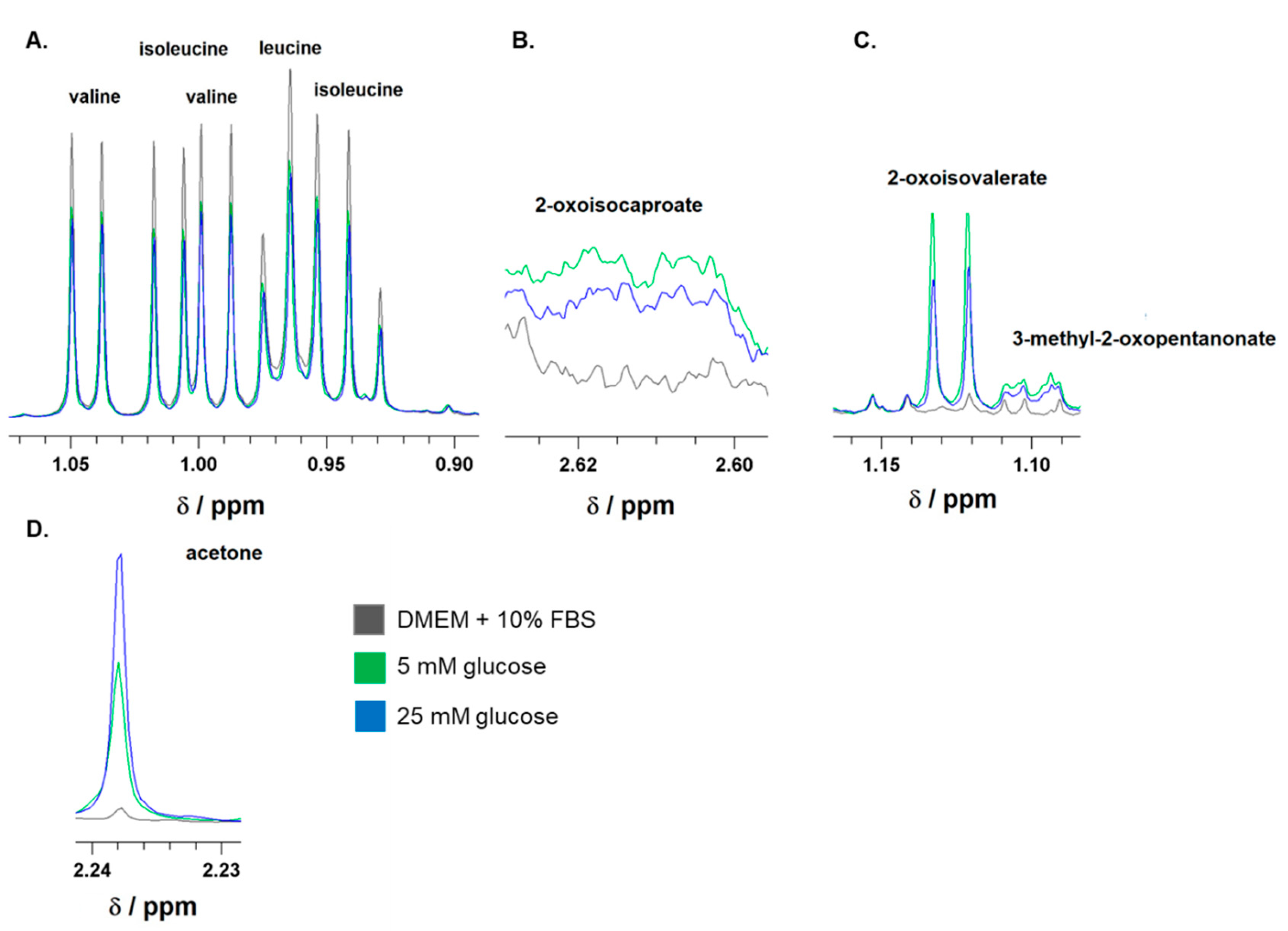
Figure 2.
The quantification of specific uptake or release of metabolites from culture medium with normoglycemia (5 mM) and hyperglycemia (25 mM) state on cultured human astrocytes by 1H-NMR analysis, except 3-hydroxybutyrate (3-OHB) that has been estimated enzymatically. The both groups were incubated for 24 hours. The specific release (negative value) and uptake of the metabolite from the culture medium were quantified by standardizing them to total cellular protein mass and incubation time (nmol*h-1*mg-1). The estimation of specific uptake of leucin (Leu), isoleucine (Ile), and valine (Val) (A) from culture medium, the specific release of BCKA, namely α-ketoisocaproate (KIC); α-keto-methylvalerate (KMV); and α-ketoisovalerate (KIV) (B), the specific release of 3-OHB, and acetone (C) into the culture medium, and calculation of KIC/leu, KMV/Ile, and BCKA/BCAA ratios (D) are depicted. All values represent the mean ±SEM from at six independent experiments and * represents the value of p ≤ 0.05, and ** p ≤ 0.01.
Figure 2.
The quantification of specific uptake or release of metabolites from culture medium with normoglycemia (5 mM) and hyperglycemia (25 mM) state on cultured human astrocytes by 1H-NMR analysis, except 3-hydroxybutyrate (3-OHB) that has been estimated enzymatically. The both groups were incubated for 24 hours. The specific release (negative value) and uptake of the metabolite from the culture medium were quantified by standardizing them to total cellular protein mass and incubation time (nmol*h-1*mg-1). The estimation of specific uptake of leucin (Leu), isoleucine (Ile), and valine (Val) (A) from culture medium, the specific release of BCKA, namely α-ketoisocaproate (KIC); α-keto-methylvalerate (KMV); and α-ketoisovalerate (KIV) (B), the specific release of 3-OHB, and acetone (C) into the culture medium, and calculation of KIC/leu, KMV/Ile, and BCKA/BCAA ratios (D) are depicted. All values represent the mean ±SEM from at six independent experiments and * represents the value of p ≤ 0.05, and ** p ≤ 0.01.
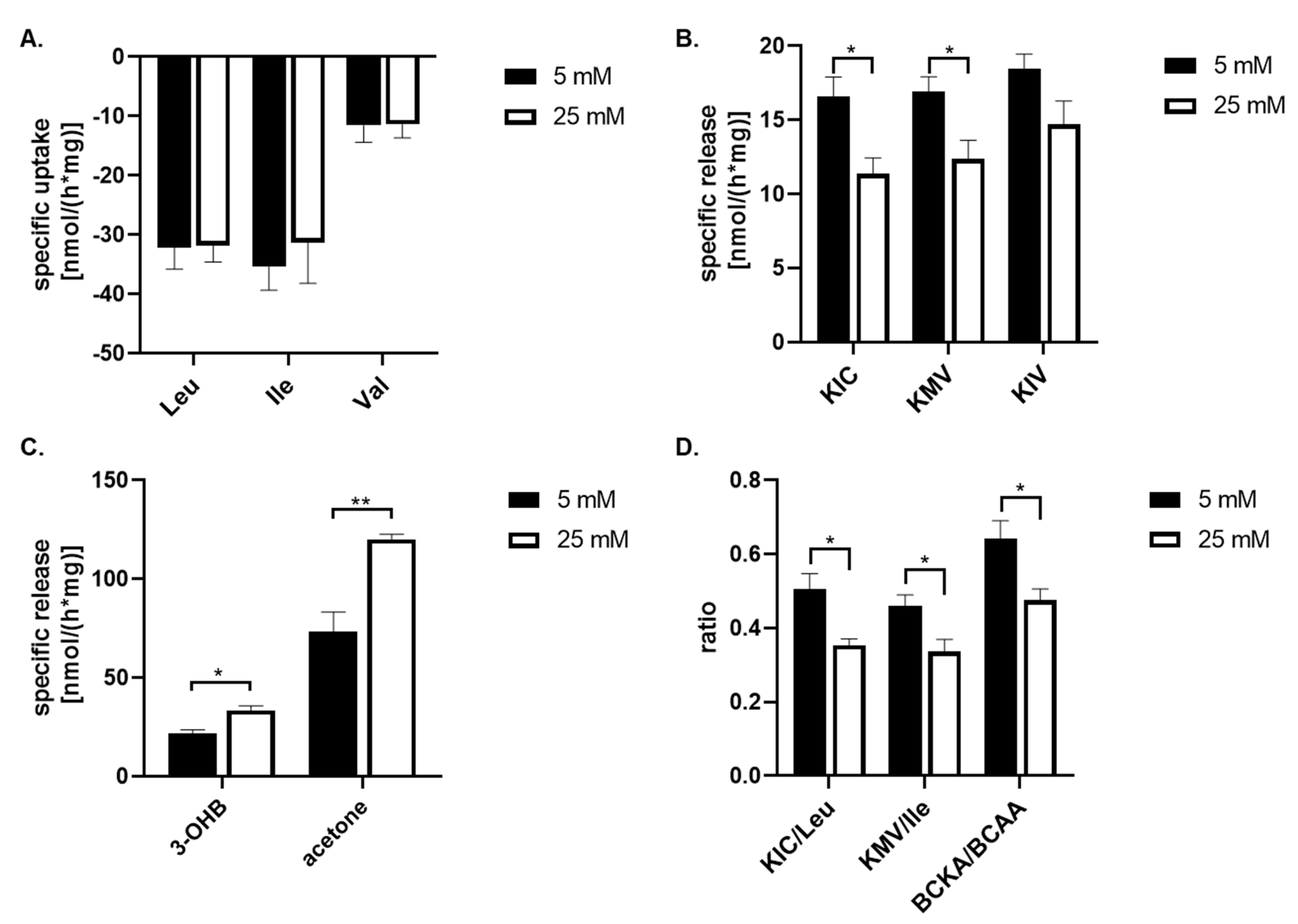

Table 1.
Effect of normoglycemia (5 mM) and hyperglycemia (25 mM) on total lysate protein, cell survival, and specific activity of lactate dehydrogenase (as LDH) on human astrocytes. The both groups were incubated for 24 hours in culture medium. All values represent mean ±SD from six independent experiments.
Table 1.
Effect of normoglycemia (5 mM) and hyperglycemia (25 mM) on total lysate protein, cell survival, and specific activity of lactate dehydrogenase (as LDH) on human astrocytes. The both groups were incubated for 24 hours in culture medium. All values represent mean ±SD from six independent experiments.
| Parameter (Unit) | Glucose | ||
|---|---|---|---|
| 5 mM | 25 mM | ||
| Protein content (mg) | 0.11 ±0.03 | 0.09 ±0.03 | |
| Cell survival (%) | 70 ±5 | 67 ±6 | |
| as [LDH] (U/g) | 266 ±29 | 231 ±50 | |
Table 2.
The level of metabolites in culture DMEM/FCS prior (Medium) or after 24 hour incubation in presence of cultured human astrocytes. To simulate euglycemia or hyperglycemia the medium at the begging of incubation contained glucose at level either 5 mM or 25 mM, respectively. The concentrations of compounds were estimated either by 1H-NMR analysis, or enzymatically. All values represent mean ±SD from six independent experiments. The statistical analysis compared 5 mM glucose concetration in the culture medium againts 25 mM and * represents the value of p ≤ 0.05, and ** p ≤ 0.01, *** p ≤ 0.001.
Table 2.
The level of metabolites in culture DMEM/FCS prior (Medium) or after 24 hour incubation in presence of cultured human astrocytes. To simulate euglycemia or hyperglycemia the medium at the begging of incubation contained glucose at level either 5 mM or 25 mM, respectively. The concentrations of compounds were estimated either by 1H-NMR analysis, or enzymatically. All values represent mean ±SD from six independent experiments. The statistical analysis compared 5 mM glucose concetration in the culture medium againts 25 mM and * represents the value of p ≤ 0.05, and ** p ≤ 0.01, *** p ≤ 0.001.
| Concentration (µM) | Medium | Glucose | |
| 5 mM | 25 mM | ||
| lactate | 801 ±38 | 2195 ±176 ** | 1862 ±60 |
| pyruvate | 917 ±26 | 341 ±27 ** | 430 ±27 |
| citrate | 18.5 ±0.5 | 25 ±1 | 26 ±1 |
| acetate | 90 ±12 | 136 ±14 | 128 ±40 |
| acetone | 29 ±2 | 174 ±78 | 253 ±98 |
| 3-hydroxybutyrate | 0 | 56 ±2 | 62 ±5 |
| glutamine | 5045 ±180 | 3878 ±136 * | 3716 ±50 |
| leucine | 793 ±15 | 714 ±18 | 727 ±7 |
| α-ketoisocaproate | 22 ±4 | 61 ±5 ** | 45 ±3 |
| isoleucine | 751 ±19 | 663 ±24 | 674 ±12 |
| α-keto-methylvalerate | 14 ±2 | 53 ±4 *** | 39 ±2 |
| valine | 773 ±10 | 745 ±14 | 748 ±16 |
| α-ketoisovalerate | 7 ±1 | 48 ±4 ** | 35 ±2 |
| alanine | 148 ±2 | 206 ±6 ** | 190 ±3 |
| histidine | 464 ±8 | 467 ±12 | 465 ±11 |
| lysine | 867 ±40 | 1024 ±21 * | 980 ±68 |
| methionine | 209 ±3 | 222 ±5 ** | 210 ±2 |
| phenylalanine | 470 ±6 | 487 ±8 | 482 ±8 |
| threonine | 1016 ±34 | 1043 ±41 | 1014 ±41 |
| tryptophan | 21 ±1 | 23 ±2 | 23 ±1 |
| tyrosine | 367 ±6 | 381 ±7 * | 371 ±6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



