
Submitted:
19 June 2024
Posted:
20 June 2024
You are already at the latest version
Abstract
The prospect of drinking water serving as a conduit for gut bacteria, artificially selected by disinfection strategies and lack of monitoring at the point of use, is concerning. Certain opportunistic pathogens, notably some nontuberculous mycobacteria (NTM), often exceed coliform bacteria levels in drinking water, posing safety risks. NTM and other microbiota resist chlorination and thrive in plumbing systems. When inhaled, opportunistic NTM can infect the lungs of immunocompromised or chronically ill patients, and the elderly, primarily postmenopausal women. When ingested with drinking water, NTM often survive stomach acidity, reach the intestines, migrate to other organs using immune cells as vehicles, potentially colonizing tumor tissue, including in breast cancer. The link between the microbiome and cancer is not new, yet the recognition of intratumoral microbiomes is a recent development. Breast cancer risk rises with age, and NTM infections emerged as a concern among breast cancer patients. In addition to studies hinting at a potential association between chronic NTM infections and lung cancer, NTM have also been detected in breast tumors at levels higher than normal adjacent tissue. Evaluating the risks of continued ingestion of contaminated drinking water is paramount, especially given the ability of various bacteria to migrate from the gut to breast tissue via entero-mammary pathways. This underscores a pressing need to revise water safety monitoring guidelines and delve into hormonal factors, which includes addressing the disproportionate impact of NTM infections and breast cancer on women and examining the potential health risks posed by the cryptic and unchecked microbiota from drinking water.
Keywords:
Drinking water
; microbiota
; nontuberculous mycobacteria
; entero-mammary pathways
; breast cancer
1. Microbiology of Drinking Water
The provision and accessibility of clean drinking water stands as a major achievement in public health [1]. Nonetheless, achieving universal access to safe drinking water remains a daunting challenge for the 21st century [2]. Globally, water quality confronts an array of obstacles, from pollution and toxins to microplastics and pharmaceutical contaminants, including antibiotics, which contribute to the proliferation of antimicrobial resistance (AMR) [3]. The dissemination of waterborne pathogens presents a significant concern, particularly pronounced in low-income countries yet prevalent in high-income ones as well. While water disinfection methods are standard practice in high-income countries, they do not offer an infallible safeguard for drinking water safety. Consequently, populations are continuously exposed to waterborne opportunistic pathogens despite these efforts. The consequences of regularly consuming water contaminated with a cryptic microbiota are largely unknown, although evidence suggests that the drinking water microbiota, selected by the conditions of the water plumbing distribution system and the disinfectants used, can have a significant impact on the structure of the gut microbiome [4].
In the late 19th century, outbreaks of cholera and typhoid fever underscored the risk posed by sewage-contaminated water. Identification of the responsible pathogens confirmed the grave risks associated with the fecal-oral route in water safety. Subsequent public health endeavors throughout the following century were dedicated to thwarting fecal-oral transmission, culminating in the establishment of modern water quality standards. These standards rely on the detection of fecal contamination using culture-based bacterial indicators and water disinfection methods.
While these initiatives have significantly curtailed waterborne diseases, it has become apparent that not all waterborne pathogens adhere to the fecal-oral transmission route [1]. Consequently, respiratory illnesses, ear infections, and dermatological issues have surged in prevalence [5]. The emergence of pathogens like Legionella pneumophila, Pseudomonas aeruginosa, and Mycobacterium spp. (nontuberculous mycobacteria, NTM) within drinking water distribution systems perpetuates ongoing public health hazards. These opportunistic pathogens defy conventional water quality metrics and disinfection protocols [6].
The effects of water purification processes on the microbiota inhabiting water treatment and distribution systems, and consequently on the microbiological integrity of drinking water, have been elucidated through advanced high-throughput DNA sequencing techniques. The composition of the microbiome within drinking water is predominantly influenced by several key factors, including the initial quality of the water source, the methods employed for treatment, and the infrastructure of plumbing distribution systems. Moreover, environmental variables such as temperature, pH levels, and the materials used in plumbing systems can further influence these microbial communities [7,8].
The potabilization process is a standardized procedure typically involving coagulation, filtration, and disinfection, commonly employing chlorine-based methods. Despite this process gradually diminishing the abundance and diversity of microorganisms [7], a diverse microbiota, estimated between 106 to 108 cells per liter, may persist, encompassing potentially pathogenic species [9]. The selection of specific water treatment steps is contingent upon the quality of the source water [10], with each stage of treatment exerting an impact on the water microbiome [7]. Furthermore, it's noteworthy that biofilters harbor a microbial community capable of disseminating throughout the downstream water system [11,12].
As water traverses through the distribution system, a myriad of factors shape microbial communities and their persistence, namely the distance from the treatment point, contact duration, physicochemical parameters, and local environmental conditions, such as maintenance procedures (e.g., corrosion control) or plumbing material and biofilms [13]. Notably, the microbiome residing within plumbing biofilms contributes significantly more to overall biodiversity than the initial bulk water present in the treatment plant post-potabilization [14]. The task of delineating a specific group of taxa characteristic of drinking water is arduous due to the plethora of variables that fluctuate across different stages of the treatment and influence the drinking water microbiota [14].
Furthermore, a diverse range of bacterial taxa, notably Pseudomonadota, Planctomycetota, Actinomycetota, Acidobacteriota, Bacteroidota, and Chloroflexota, are frequently identified within these distribution systems [7,13]. Interestingly, the presence and abundance of Archaea and Eukarya appear to be influenced by the use of disinfectants [15]. Moreover, free-living amoebae are ubiquitous in distribution systems and serve as potential reservoirs for amoeba-resistant bacteria [16].
Biofilm communities within treatment and water distribution systems exhibit distinct differences from planktonic communities found in bulk water. However, they do share several taxa that rank among the most abundant in both environments [17]. Studies have pinpointed dominant genera within biofilms, including Pseudomonas, Mycobacterium, Methylobacterium, Sphingomonas, among others, although the prevalence of these genera may fluctuate [18-21] [17,22,23].
The final phase of water treatment, prior to distribution to consumers, involves disinfection, during which a residual amount of disinfectant, such as chlorine or chloramine, is maintained throughout the system. In Northwestern European nations, due to the high quality of source water and robust treatment methods, the need for disinfectant residuals is eliminated. However, despite these strategies, microbial growth within the plumbing and distribution system is inevitable [7,15]. Numerous studies have noted a decline in microbial richness and evenness attributed to residual disinfection. Conversely, distribution systems lacking a residual disinfectant tend to exhibit greater microbial diversity and abundance, albeit with fewer pathogens. Nonetheless, conflicting evidence persists regarding the abundance of Mycobacterium spp. and other potential pathogens in disinfected systems [23,24] albeit levels of bacteria such as the genera Mycobacterium, Methylobacterium, Sphingomonas, Pseudomonas, and Legionella increase following disinfection [14,18,23,25]. Coliforms are rarely detected within the core microbiome of treated water. Occasionally, a few non-classical genera are identified at low abundance, indicating their limited presence under typical water system conditions [14]. Classical waterborne pathogens like Vibrio, Salmonella, Shigella, and Escherichia coli are highly susceptible to chlorine and are typically absent [6]. Therefore, their presence in water systems under normal operational conditions is unlikely.
Once water exits the distribution system, it enters premise plumbing, comprising pipelines, water heaters, fixtures, and faucets in private residences, public establishments, hospitality venues and healthcare facilities [26]. The conditions prevailing within premise plumbing, characterized by diminishing disinfectant residuals, extensive surface areas, relatively elevated temperatures, and irregular water flow patterns, foster microbial proliferation [27]. At this stage of the plumbing network, bacteria may have withstood rigorous water treatment, developed resistance to filtration and disinfection, formed biofilms that bolster their resilience, exhibited the ability to thrive within free-living amoebae, and flourished in oligotrophic environments [6]. Mycobacterium, Methylobacterium, Sphingomonas, Bradyrhizobium, Sphingobium, and Nitrospira, along with cyanobacteria and others, can endure the challenging conditions within premise plumbing. Nonetheless, research on premise plumbing often concentrates on the potential of tap water to serve as a reservoir for waterborne infections, driven by the presence of opportunistic premise plumbing pathogens (OPPPs) like Mycobacterium, Methylobacterium, Citrobacter, Pseudomonas, Stenotrophomonas, Legionella, and Acinetobacter [6]. These pathogens are also frequently encountered in showerheads, shower curtains, faucets, washing machines, and other endpoint water devices [28].
Additionally, Enterococcus and Escherichia can be detected in water-related apparatus at the endpoint, despite their limited presence in upstream distribution systems [28,29]. Respiratory infections have been more frequently associated with Legionella and Mycobacterium, bacteremia with Aeromonas, and dermal infections with Pseudomonas from domestic premise plumbing water [28]. Notably, OPPPs have been identified in hospital plumbing, contributing to approximately 21% of all documented cases of hospital-acquired infections [30]. Exposure can occur via aerosolized water droplets generated by showerheads, faucets, and other endpoint devices, or through ingestion or direct contact with contaminated tap water [31].
Water from showerheads, along with biofilms and shower curtains, often harbors elevated levels of Mycobacterium [27,32,33]. Notably, Mycobacterium can be transmitted from water to indoor air during showering [32]. Studies have revealed a correlation between the species detected in the homes of patients with NTM infections and those found within the patients in roughly 35% of cases [34]. Regions with high levels of potentially pathogenic NTM in showerheads often coincide with areas where lung disease is prevalent [33]. However, it was also observed that NTM were significantly more common in showerheads supplied with municipal water compared to those supplied with well water, emphasizing the importance of the source water and treatment methods. Furthermore, households in the United States exhibited a higher abundance of NTM compared to those in Europe [33].
2. Nontuberculous Mycobacteria: Environmentally Versatile Opportunistic Pathogens
Nontuberculous mycobacteria are environmental bacteria that are commonly found in tap water, leading to continuous human exposure throughout life. Their innate resistance to common disinfectants gives them a competitive edge over other bacteria present in water. While a few Mycobacterium species have been associated with opportunistic infections to different degrees, the full extent of chronic exposure's impact on human health is still not fully understood. Therefore, understanding their unique physiology, metabolism, behavior and adaptation to water distribution systems is vital for accurately assessing the risks associated with inadequate disinfection methods, which can lead to the proliferation of these bacteria in water presumed safe for human consumption.
The genus Mycobacterium encompasses over 200 formally described species of acid-fast aerobic or microaerophilic bacilli, characterized by long-chain mycolic acids in their cell walls [35]. Currently classified within the family Mycobacteriaceae of the phylum Actinomycetota, this genus was established by Lehmann and Neumann in 1896, delineated by features observed in the type strain Mycobacterium tuberculosis, including its growth as fungus-like pellicles on liquid media [36]. In addition to M. tuberculosis and closely related species, the agents of tuberculosis, and M. leprae, which is responsible for leprosy, this large genus encompasses over 190 additional environmental species referred to as nontuberculous mycobacteria (NTM), some of which like M. abscessus or M. avium and others, can be dangerous opportunistic pathogens [37]. A proposal to divide the Mycobacterium genus into five distinct genera [38], backed by genomic evidence was initially controversial due to potential misinterpretations in clinical microbiology, but the original (basonym) name Mycobacterium remains valid [39], and will be used throughout this article.
NTM possess a versatile metabolism and a distinctive lipid-rich cell wall, enabling them to thrive in nutrient-poor environments and withstand immune and drug pressures. Their lipid-rich outer membrane contributes to their slow growth, impermeability, and hydrophobicity, rendering them capable of forming aerosols and resistant to disinfectants and antibiotics. In addition to their oligotrophic metabolism, they exhibit tolerance to low pH, high temperatures, and desiccation [40]. Most mycobacteria display microaerophilic behavior and are capable of thriving under hypoxic conditions [41], such as those encountered in lung granulomas, organized tissue structures triggered by infection and immune response, characterized by the accumulation of immune cells, predominantly macrophages, surrounded by lymphocytes [42,43]. Interestingly, granulomas share several structural similarities with solid tumors, both of which recruit immune cells and experience oxygen deprivation [44]. NTM are capable of surviving and reproducing within protozoans, particularly free-living amoeba, providing added protection in harsh environments [40,45,46]. Furthermore, certain NTM engage in the exchange of genetic material through plasmid-mediated horizontal gene transfer, a process facilitated within biofilms, and which enhances their resistance to antibiotics and metals [47]. NTM flourish in diverse environments, spanning natural waters, hot springs, soils, and dust, as well as artificial settings like disinfected water supply networks, tap and showerhead water, and peat-rich potting soil. Such proliferation can significantly heighten human exposure [40].
NTM can cause both pulmonary and extrapulmonary disease, which encompass a range of diseases involving the skin, skeleton, soft tissues, as well as the urinary and gastrointestinal tracts, and even the central nervous system [48,49]. Extrapulmonary conditions can result in substantial morbidity, particularly in cases of healthcare-associated infections stemming from open-wound procedures or insertion of invasive medical devices [50]. Pulmonary manifestations make up a significant proportion of NTM-related diseases, accounting for approximately 77% to 90% [51-53]. Globally, the incidence of these diseases has been progressively rising at an average rate of 4.1% (3.2–5) annually for prevalent species such as M. avium and related strains, as well as for M. abscessus [54]. Other studies also found a consistent annual incidence rate of extrapulmonary NTM disease at 1.5 cases per 100,000 population [55]. These patients had a lower median age than pulmonary NTM patients, and fast-growing NTM species appear more common in extrapulmonary cases than in pulmonary cases. These data suggest that NTM are adept at spreading throughout the human body, which may be facilitated if they are continuously ingested at abnormally high numbers such as those observed in different studies in recent decades (see below).
The diagnosis of NTM lung disease poses challenges due to the frequently nonspecific symptoms and the requirement for extensive laboratory analysis. Treatment entails prolonged administration of multiple antibiotics tailored to the specific infecting strain and disease severity, aiming for a minimum of 6-12 months of culture negativity [56]. Treatment often involves harsh and protracted effects, potentially leading to patient discontinuation or non-adherence [57]. On average, culture conversion (two consecutive pathogen free sputum cultures) rates hover around 60–70%, with recurrence rates reaching 50% [58]. Hence, it is imperative to optimize treatment strategies and explore novel, effective, and well-tolerated medications [59].
NTM infections are contracted from environmental sources through ingestion, dermal contact, or inhalation of NTM-laden aerosols emitted from waters and soils [40]. The precise risk factors for NTM disease remain incompletely understood; however, repeated exposure is deemed a main factor, especially for individuals with compromised immune systems, advanced age, or underlying lung conditions like bronchiectasis or cystic fibrosis (CF) [60]. Women, particularly postmenopausal women, are more susceptible to NTM infections [61,62]. This increased susceptibility may be partially due to immunosenescence phenomena, specifically the decline in competence of innate immune system cells [63], although hormonal factors may be at play. Certain host phenotypes and genetic variations, such as low body mass index, thoracic skeletal anomalies (referred to as Lady Windermere syndrome in women), and congenital disorders affecting IL12/IL23-INF-γ mediated immunity, may heighten susceptibility to NTM infection [64]. Genome-wide association studies have identified single nucleotide polymorphisms (SNPs) associated with susceptibility to NTM disease caused by members of the M. avium complex (MAC) across Japanese, Korean, and American populations (e.g., rs109592 and rs849177) [64,65].
NTM lung disease clinical case definition for diagnostic and treatment purposes that is endorsed by the main scientific and clinical organizations in respiratory medicine was established 25 years ago and last updated in 2020 [56]. The diagnostic criteria were developed based on the most common pathogens such as M. avium complex and M. abscessus, for the majority of NTM the applicability of the diagnostic criteria is not established. Uncertainty about diagnosis, disease progression, correct time to initiate antimycobacterial therapy is considerable. Furthermore, direct human-to-human transmission of NTM appears to be uncommon, contributing to the disease’s non-notifiable status, consequently epidemiological understanding relies on local and regional surveillance mechanisms. Lack of consensus on outcome parameters leads to the use of varying case definitions for monitoring infection rates and identifying risk factors which results in imprecise incidence data [54,66]. In the USA, estimated prevalence rates for NTM pulmonary disease (NTM-PD) have been on the rise, increasing from 6.8 per 100,000 in 2008 to 11.7 per 100,000 in 2015 [67]. In 2020, the annual prevalence of NTM disease in some European countries ranged from 6.1 to 6.6 per 100,000. This marks a notable contrast with East Asian nations, notably Japan, where the prevalence stood at 24.9 per 100,000 population [68]. According to [69], the prevalence of NTM-PD in East Asian countries was 7.5% higher than in other nations, consistent with studies indicating increased susceptibility to NTM disease in Asian populations. In Australia, mycobacterial infections are subject to mandatory reporting, with 25.9 cases per 100,000 population reported in 2015. Research suggests substantial regional disparities in the incidence and frequency of isolation of common pathogens. The most frequently encountered pathogens include strains of the M. avium complex (MAC) and M. abscessus complex (MABC). Mycobacterium xenopi is more prevalent in Croatia, Czech Republic, and Serbia while M. kansasii is dominant in Poland and Spanish regions and M. malmoense in Scotland and the Netherlands [54].
In addition to infections linked to environmental sources, including gardening soil, soil dust, and water distribution and plumbing systems in community and healthcare settings [70-72], NTM disease has also been linked to showerheads and bathroom fixtures, hot tubs, indoor swimming pools, public baths, and contaminated ink in tattoo parlors [34,73-80]. Healthcare-associated outbreaks have been linked to exposure to NTM-contaminated water and inadequate disinfection or sterilization procedures associated with various medical procedures such as dental procedures, the use of heater-cooler devices during cardiac surgery, and the utilization of invasive medical devices [81-85]. Apart from the individual risk factors and environmental exposures mentioned earlier, broader environmental factors have also been examined. These factors, generally associated with water, climate, and soil, impact entire populations and contribute to the variation in NTM disease risk across different geographic locations. However, assessing their correlation with NTM infection incidence is intricate due to the prolonged incubation period [72]. Several studies have noted seasonal upticks in NTM levels in drinking water systems during warmer periods [19,86,87]. Rainfall also appears to influence incidence rates, with varying effects depending on the region's dryness [88]. Regions characterized by a higher proportion of land covered by surface water and elevated potential mean daily evapotranspiration levels are linked to an increased risk of NTM lung disease [89]. Additional research has also connected the concentrations of trace metals in water sources, such as molybdenum, vanadium, copper, and soil sodium levels, to heightened risks of NTM disease [89-91]. Associations have been identified between NTM isolation and exposure to water-saturated soils and, to a lesser extent, acidic soils (pH < 5.5), as well as shallow soil depths in agricultural regions [88,92].
Although it was believed that NTM only infected immunocompromised individuals, it became evident that immunocompetent individuals are also targeted [93]. Combining the high levels of NTM ingested with tap water in comparison to those inhaled from aerosols [94], with their apparent ability to travel between organs within the human body, tap water could hypothetically also be an alternative source of lung infections.
Mycobacteria in point-of-use tap water have been reported at counts ranging from 10 to 700,000 CFU/L (colony forming units per liter) in studies conducted in both the USA and throughout Europe [94-97]. Current guidelines for assessing the microbiological quality of drinking water fail to address NTM or other abundant microbiota that multiply within the plumbing system [94,98,99]. Standardized procedures for assessing these bacteria are lacking, despite their prevalence being significantly higher than that immediately downstream of treatment plants. This oversight suggests that the proliferation of microorganisms within the plumbing system results in their unintended ingestion by the population in significant quantities, raising uncertainties about potential health effects [94]. The lack of standardization in culture medium, incubation times, and temperature, leads to discrepancies, especially on NTM isolation, quantification, and analysis. Standardization of protocols for these records is therefore urgently needed, along with prompt regulation by public health authorities of microbiological assessment of drinking water safety. Suggested approaches with significant promise for drastically lowering NTM levels in drinking water have been put forward and validated [100]. Yet, their adoption within communities necessitates intervention from health authorities.
Considering an average of 1L to 2.5 L of daily consumption of drinking water in Europe per person [101] and 1.1 L in the USA [102], it is possible that individuals are ingesting NTM at levels significantly much higher than suspected on a daily basis, and for years. Although not part of the core gut microbiome, NTM can be detected in the intestine and in stool samples [103,104].
3. Breast Cancer: Epidemiology, Biology, and Pathology
Breast cancer stands as the primary cause of cancer-related fatalities among women globally. The year 2020 alone saw approximately 2.3 million new cases diagnosed, culminating in 685,000 deaths [105]. Breast cancer inflicts substantial physical, emotional, social, and economic burdens, constituting around 30% of female cancer cases worldwide and carrying a mortality rate of 15% [106,107]. Despite considerable progresses and breakthroughs in breast cancer treatment, it remains a formidable threat globally. Recent years have witnessed a decrease in mortality rates, notably in Western demographics, particularly among younger age cohorts [108]. The continued expansion of access to top-tier prevention, early detection, and treatment services for all women holds promise in further driving down mortality rates [109].
Approximately 10% of breast cancer cases are linked to genetic predisposition or family history, with variations among countries and ethnicities. The most prevalent germline mutations linked to breast cancer occur in the BRCA1 and BRCA2 genes, vital for DNA repair, carrying an average cumulative lifetime risk of approximately 70% [110]. A substantial portion of breast cancer cases can be attributed to factors related to pregnancy, hormone therapy, and lifestyle choices such as obesity, physical inactivity, alcohol consumption, low-fiber diet, and smoking [111]. The potential association between hormonal contraceptives and breast cancer risk has long been debated, with the absolute risk being small and not linked to an increased mortality [112]. Menopausal hormone therapy has been more definitively associated with increased breast cancer risk in women [113]. In recent years, attention has shifted towards exploring the association between bacteria and breast cancer, a topic that will be discussed in the following sections.
Histologically, the most prevalent form of breast cancer is invasive ductal carcinoma, often referred to as "no special type," affecting 50%-75% of patients. This is followed by invasive lobular carcinoma, observed in 5-15% of patients, characterized by mutations in epithelial cadherin (CDH1) and a distinctive growth pattern. Breast cancer exhibits high heterogeneity and is clinically categorized into five intrinsic subtypes based on the expression of estrogen receptor (ER), progesterone receptor (PR), epidermal growth factor 2 (ERBB2), and the Ki67 proliferation marker protein (MKI67). Estrogen receptor alpha (ERα), expressed in approximately 70% of invasive breast cancer cases, functions as a steroid hormone nuclear receptor and a transcription factor that, when activated by estrogen, initiates oncogenic pathways in breast cancer cells. The presence of the related steroid hormone progesterone receptor (PR) is also indicative of ERα signaling. Targeting ER signaling with endocrine agents constitutes the primary systemic therapy for ER-positive or PR-positive breast cancer.
The second major molecular target in breast cancer is epidermal growth factor 2 (ERBB2, previously known as HER2 or HER2/neu), a transmembrane receptor tyrosine kinase belonging to the epidermal growth factor receptor family. ERBB2 is amplified or overexpressed in around 20% of breast cancer cases and is associated with a poor prognosis without systemic therapy [114]. Patients with ERBB2-overexpressing breast cancer benefit from targeted therapy, such as anti-ERBB2 antibodies. Triple-negative breast cancer (TNBC), constituting approximately 15% of all breast tumors, lacks expression of the molecular targets ER, PR, or ERBB2 and have a heightened risk of distant relapse within the initial 3-5 years post-diagnosis [115]. About 15–20% of TNBC cases are linked to germline mutations in BRCA1 or BRCA2. High-risk, HER2-negative, hormone-receptor-positive breast cancer is correlated with germline mutations in BRCA1 or BRCA2 in about 10–15% of cases [116]. Apart from variations in the expression of targetable receptors, these subtypes also exhibit differences in their immune profiles, including variations in PD-L1 expression, tumor-associated antigens, tumor mutational burden, and the quantity and composition of tumor-infiltrating lymphocytes within the tumor immune microenvironment [117].
Breast cancer can metastasize to various organs, including the liver, lungs, brain, bone and other organs like adrenal glands or skin, through the bloodstream or lymphatic system. The pattern of metastatic spread varies based on the breast cancer subtype, stage, and individual patient characteristics.
Chronic inflammation, attributed to bacterial infections, is suggested to play a prominent role in the metastasis of breast cancer to other organs like the lungs. Bacterial infections can alter the immune environment of affected organs, promoting the colonization of tumor cells and facilitating metastasis by recruiting tumor-promoting MHCIIhi neutrophils by differential expression of specific cytokines and chemokines [118]. Breast cancer patients with concomitant NTM infection have circulating exosomes containing proteins that promote epithelial to mesenchymal transition, a mechanism involved in tumor progression with metastatic expansion and immune modulation achieved by altering the expression of various cytokines and chemokines, potentially heightening susceptibility to NTM disease [119,120].
4. Bacteria and Cancer
For decades, the potential link between bacteria and cancer has intrigued researchers. While initially suggested as far back as 1884 [121], it wasn't until relatively recently that concrete evidence emerged. The pivotal moment emerged when Helicobacter pylori was identified as a Group 1 carcinogen for gastric adenocarcinoma during the 1994 National Institutes of Health Consensus Conference [122]. Despite this landmark discovery, subsequent research has not led to the inclusion of other bacteria in the Group 1 list by the International Agency for Research on Cancer of the World Health Organization. Nonetheless, in recent years, there has been a surge of interest in the relationship between bacteria and cancer, driven largely by advancements in -omics sciences, particularly microbiome research. It has become increasingly apparent that bacteria play significant roles in the tumorigenesis of various cancers [123-125].
The link between the gut microbiome and cancer, especially breast cancer, has been well-established [126]. This association is attributed to the production of potentially carcinogenic toxins that may reach breast tissue via circulation, as well as the generation of metabolites that could potentially impede its progression [127,128]. Gut microbes have also been observed to produce enzymes that deconjugate excreted estrogen leading to its reabsorption into circulation and thus to increased circulating estrogen levels. Additionally, gut microbes also synthesize several estrogen-like compounds or estrogen mimics from dietary sources, such as enterolactone or enterodiol, which can influence systemic estrogen levels, induce proliferation of ER positive breast cancer cell lines, increase cell viability and their clonogenic potential, thereby impacting breast carcinogenesis [129]. As approximately 70% of all breast cancers are ER positive subtype, the imbalance of estrogen and of estrogen mimics levels can impact breast carcinogenesis [129].
Alterations in the healthy gut microbiota, known as dysbiosis, can significantly impact host immunity. Furthermore, the gut microbiome can influence cancer immunotherapy by encompassing various microbes that can either bolster or hinder the therapeutic efficacy [128]; [130]. This is supported by recent findings, indicating that cancer patients undergoing checkpoint inhibitors’ immunotherapy, who received antibiotics before or during treatment, exhibited poorer clinical outcomes compared to those who did not receive antibiotics [131]. Moreover, experiments with mice have shown that inoculation with exogenous bacteria can compromise tumor chemotherapy and accelerate tumor growth and metastatic progression [132,133].
Research is currently exploring the association between the gut microbiome and cancer for therapeutic applications through various approaches, such as modulation of the gut microbiota via specific diets and probiotics, as well as the utilization of bacteriophages [128,134]. Additionally, fecal microbiota transplantation from healthy donors has shown effectiveness in some studies [135]. However, the connection between microbes and cancer may extend beyond the influence of the gut microbiome alone.
Gut dysbiosis can compromise the integrity of the intestinal barrier, allowing bacteria and microbial products to escape into circulation, which can trigger pro-inflammatory pathways, disrupting immune balance and fostering tumor development [136]. Pathogens are detected through pathogen-associated molecular patterns (PAMPs) by Toll-like receptors (TLRs), initiating signaling pathways that activate genes associated with immune response and inflammation. Additionally, PAMPs prompt the differentiation of various immune cells, such as T cells, B cells, and CD4 T cells, into regulatory T cells (Treg) and Th17 cells, influencing both gut and systemic immunity [132] [127,128]. Irrespective of bacterial route to reach tissues, their colonization of tumors is facilitated by the permeable vasculature and immunosuppressed environment characteristic of tumors. Not rarely, bacterial transport appears to be carried out by immune cells that migrate from the gut to other parts of the body [137,138].
5. Entero-Mammary Pathways and the Intratumoral Microbiome
A concept that has gained momentum in recent years is the notion of a gut-to-tumor route for bacterial migration. Studies comparing tumor microbiome of metastatic melanoma from patients who responded to immune checkpoint inhibitor therapy to the tumor microbiome of non-responders found that the patterns of differentially abundant taxa between the two groups found in melanoma samples matched the patterns that had been previously reported for gut microbiome data when comparing responders and non-responders [139,140]. Furthermore, several reports suggest that the majority of bacteria found in tumor tissue, including breast cancer, are located intracellularly, primarily within CD45+ immune cells, suggesting that both cancerous and host cells may serve as vehicles for bacterial transport to the tumor and normal adjacent tissue [126,141-143]. This migration mode appears to occur naturally and gains particular significance during pregnancy and lactation, where a greater variety of bacteria can be detected in peripheral blood mononuclear cells compared to non-pregnant and non-lactating women [144]. In mice, bacterial translocation to mesenteric lymph nodes was significantly increased during the perinatal period and followed by bacterial presence in the breast shortly after delivery. Within 24 hours postpartum, fewer animals have detectable bacteria in their mesenteric lymph nodes, but most women have viable bacteria in their mammary tissue [144].
Mounting evidence suggests that bacteria play integral roles within tumor tissues across various cancer types, challenging the conventional notion of tumors as sterile environments and introducing the concept of an intratumoral microbiome [139,145]. The breast harbors a microbiome that seems to be able to maintain immune responses that can combat breast tumor development and progression. Some bacterial virulence factors have been directly implicated in tumorigenesis [123]. Breast dysbiosis can foster tumor progression, which may further disrupt the mammary microbiome, suggesting that bacterial dysbiosis is an early event in breast tumor formation [146]. Therefore, also in the breast, the interaction between the microbiome and cancer cells appears to be bidirectional.
Breast cancer appears to exhibit the most abundant and diverse microbiome in this context. The presence of bacteria within breast tumors may not be surprising when considering that both breast tissue and breast milk harbor unique microbiotas indicating an effective physiological route for microbes to access the breast [126,147,148]. It has been proposed that bacteria from the skin and oral cavity may use the nipple as an entry point to reach the breast ducts, potentially establishing a distinct microbiome within the breast tissue [147]. However, this route alone does not fully explain the presence of various gut-associated strict anaerobes, such as Faecalibacterium, Roseburia, Bifidobacterium, Blautia, Bacteroides, and Parabacteroides, in breast milk, as demonstrated by several studies. This strongly suggests the translocation of gut bacteria to the breast tissue and milk [148,149]. Concerning this route, also known as the entero-mammary pathway, mounting evidence suggests that bacterial migration from the gut to breast tissue and other organs may be a common phenomenon facilitated by innate immune cells emerging from the gut [137,138,150]. Interestingly, menopause-associated immune senescence has been proposed to result in increased cytokine and chemokine production and macrophage recruitment, but reduced cytotoxicity and phagocytosis in macrophages, which seem to become impaired in bacterial clearance, potentially facilitating NTM to exit the gut [63].
The microbiome of breast tumors differs from that of healthy breast tissue [139,145]. However, direct comparison between tumor and normal adjacent tissue reveals significant differences in the abundance of certain bacteria namely of the genera Tepidimonas, Lactococcus, and Streptococcus, as well as some Bacteroidia and Prevotelacea, while the genera Enterococcus, Lactobacillus, and Bacillus were found to be differentially abundant in the lung. Moreover, when analyzing beta-diversity within and across tumor types (breast, ovary, bone, glioblastoma multiforme (GBM), melanoma, pancreas, and lung), it became apparent that microbiomes within the same tumor type exhibit greater similarity compared to those in different tumor types [139]. For instance, in breast cancer, distinct subtypes categorized by estrogen receptor (ER), progesterone receptor (PR), and HER2 status display variations in the prevalence of specific taxa [139,146]. The genera Granulicatella and Dyadobacter were enriched in HER2+ tumors, Actinomyces, Alkanindiges, Lautropia, Sphingomonas were enriched in ER- tumors and Corynebacterium was enriched in ER+ tumors [139]. Another study found distinct microbial signatures associated with different breast cancer types some of which considered opportunistic premise plumbing pathogens, for example Legionella in ER+ tumors [151]. Additionally, the microbial load in breast cancer tumors was found to be tenfold higher than in paired normal tissue, with a decrease in bacterial load observed during disease progression from Stage 1 to 3 [152].
While further research on the composition of the breast cancer-associated microbiome is warranted, certain taxa have been proposed to be enriched in human breast tumors, namely Enterococcus, Streptococcus, Lactobacillus, Staphylococcus, Bacillus, Enterobacteriaceae, and Fusobacterium nucleatum [139,142,145]. Notably, Staphylococcus epidermidis and Escherichia coli strains isolated from breast cancer patients have been found to induce DNA double-stranded breaks in HeLa cells, suggesting a potential oncogenic mechanism for breast tissue colonization by specific bacteria [145]. The presence of F. nucleatum is of particular interest, as it has been demonstrated to promote tumorigenesis and protect tumors from immune cell action in colorectal cancer. These effects likely extend to breast cancer, as evidenced by a study showing that intravascular administration of F. nucleatum leads to colonization of breast tumors in a mouse model, resulting in exacerbated tumor growth and metastatic progression [133].
6. Mycobacteria and Cancer: Focus on Breast Cancer
Despite variations in the proposed pathways bacteria utilize to infiltrate breast tumor tissue and establish specific intratumoral microbiomes, the detection of NTM in breast tumors has been reported in some studies [153,154]. Mycobacteria naturally resist the acidic environment of the human stomach. Additional research has shown that clinical isolates of mycobacteria not only withstand pH 2.2 for 2 hours but can also prolong their survival to 24 hours when pre-adapted in water before exposure to acidic conditions [155]. There is historical evidence of M. avium infection in AIDS patients occurring through the gastrointestinal tract [156]. Immunocompromised individuals are susceptible to M. avium infections through the intestinal tract, where the bacteria can invade epithelial cells causing disseminated disease [157]. Recent studies further confirmed the presence of NTM in the gut and stool samples [103]. Interestingly, the transfer of the opportunistic pathogen Pseudomonas aeruginosa from the gut to the lungs has been documented in certain patients [158], indicating that aerosols may not be the sole pathway for lung infections. This observation prompts consideration of traditional avenues for NTM infection transmission.
Given the association between M. tuberculosis and tuberculosis (TB), a substantial volume of literature has arisen regarding the molecular mechanisms underlying this disease, including its possible involvement in carcinogenesis. The Mycobacterium oncogenic hypothesis has garnered increasing attention in recent years [159-161]. These studies delve into the intuitive association between TB and lung cancer, drawing on numerous epidemiological investigations that suggest a possible link between M. tuberculosis infection and various malignant tumors, particularly lung cancer. Despite conflicting findings, most research indicates a significant elevation in lung cancer risk associated with TB. Moreover, the hypothesis is supported by the up-regulation of at least 18 genes related to cell cycle regulation, checkpoint control, and apoptosis, which are commonly implicated in both lung cancer and tuberculosis [161]. These genes include BRCA1, whose mutations are well-known contributors to breast cancer.
NTM infections have also been associated to aerodigestive cancers, including lung cancer [162]. Among patients with NTM lung infections, 2–8.5% also present with lung cancer, highlighting the latter as a significant comorbidity in this population. The diagnosis of NTM infection in cancer patients may be delayed or overlooked because of overlapping symptoms and radiographic features, such as lung masses, cavities, and nodules as well as weight loss, cough, and hemoptysis. It's not uncommon for computed tomography (CT) and positron emission tomography (PET) imaging findings to be unable to differentiate between the two conditions [163], as they frequently display similar heterogeneous features. Certain case reports have even documented the simultaneous presence of NTM and carcinoma within the same lung tumor [164].
Chronic inflammation of lung tissue has been suggested as an underlying driver for the potential contribution of M. tuberculosis and M. avium infections to the development of lung cancer [160]. Similarly, M. ulcerans has been associated with skin carcinogenesis, potentially through oncogene mutations that may induce malignant transformations in host cells through lateral gene transfer and by stimulating the release of inflammatory mediators known to promote cancer [160]. Additionally, mycobacteria-induced reactive oxygen species have been proposed to inflict damage on host cell DNA, potentially leading to cancer development. Mycobacterium was also identified as one of the enriched genera in the responsive group when comparing two cohorts of melanoma patients based on their response to checkpoint inhibitor immunotherapy [139].
Recently, NTM have been linked also to breast cancer. An increase in the abundance of M. fortuitum and M. phlei, both opportunistic pathogens, was observed in breast cancer tissue compared to adjacent normal tissue [165]. Similarly, Mycobacterium was identified as a common genus across all breast cancer subtypes when compared to normal breast tissue controls. Additionally, other studies have noted an increased prevalence of NTM in breast cancer tissue compared to normal breast tissue, with further elevation observed in HER2+ breast cancer tissue compared to HER2- counterparts [139,151]. Taken together, these findings suggest that the presence of NTM in cancer may not be coincidental and that they might even play a role in carcinogenesis and metastasis mechanisms.
More recently, research has expanded the characterization of the breast microbiota to include male samples, revealing Mycobacterium as one of the apparent genera enriched in both male and female breast cancer samples compared to normal tissue [166]. Additionally, NTM were found to be enriched in the gut microbiome of breast cancer patients with low levels of tumor-infiltrating lymphocytes (TILs) compared to those with high levels of TILs in their breast tumors, suggesting a potential association with poorer outcomes and treatment efficacy, particularly in the context of immune checkpoint inhibitor therapy [167]. Overall, while certain NTM species possess immunomodulatory properties [168], their specific effects on the gut immune system remain poorly understood. These effects may vary depending on the bacterial species, virulence determinants, and the context of exposure, highlighting the need for further research in this area.
While investigating the correlation between breast cancer and bronchiectasis from NTM lung disease in women diagnosed with both conditions, it was found that while breast cancer diagnosis typically precede NTM disease in the majority of cases, there were instances where the sequence of events was reversed, with some women developing NTM infection before the onset of breast cancer [120]. While this observation alone does not establish NTM infection as oncogenic, it is noteworthy that in a subsequent study, the researchers identified several somatic mutations in cancer-predisposing genes among NTM patients, regardless of whether they had a concurrent breast cancer diagnosis [120].
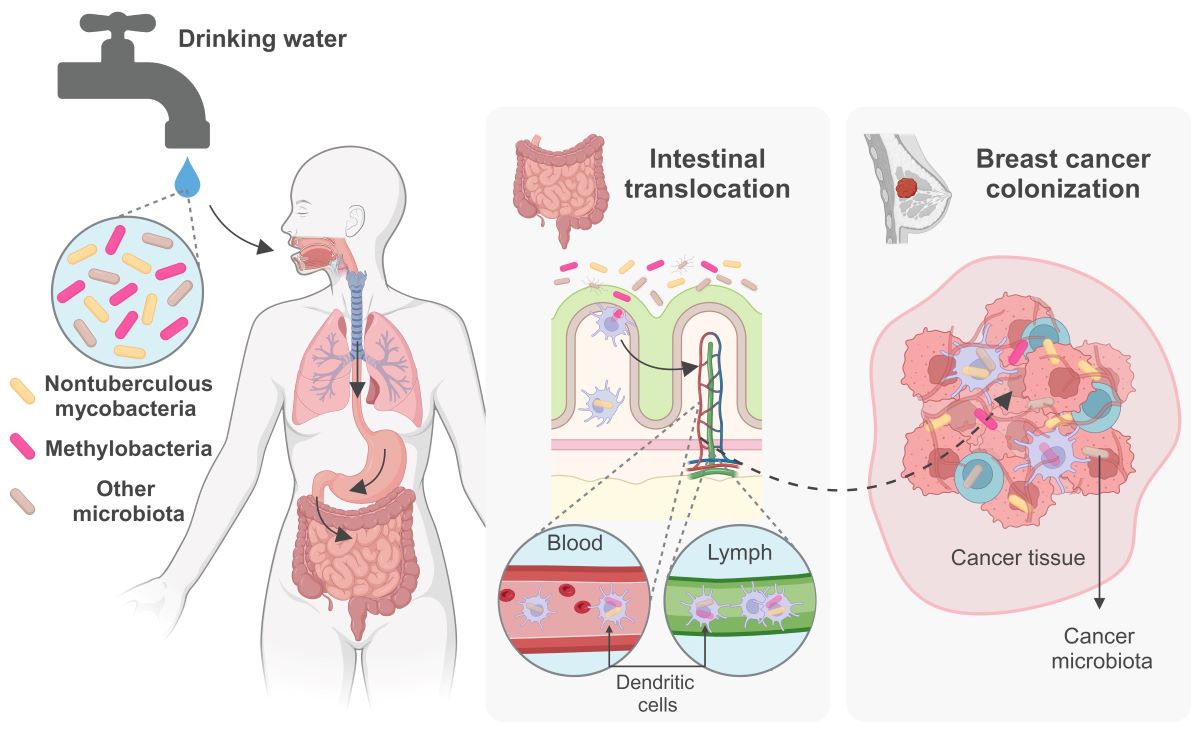
While the involvement of NTM in tumorigenesis processes remains speculative, these bacteria pose formidable challenges. They exhibit resilience against water disinfection methods, infiltrate drinking water sources unchecked, and are ingested continuously and in undetermined levels by aging populations, compounded by a rise in chronic illnesses and therefore progressively vulnerable. Moreover, NTM can withstand the acidic conditions of the stomach, traversing to the intestines, where they can be incorporated by immune cells and disseminated to distant bodily sites, including breast and tumor tissues. This complex interplay underscores the critical need for immediate attention from public health authorities to address the unregulated microbiological quality of water at the point of consumption.
7. Concluding Remarks
In addition to various other microbes present in drinking water, nontuberculous mycobacteria (NTM) are commonly detected in municipal water distribution systems, posing a potential health risk to populations that depend on these sources for their daily water consumption. Some NTM species can cause lung chronic infections, especially in individuals with compromised immune systems, chronic illnesses, the elderly, or those with underlying lung conditions. Recent studies have indicated a possible association between NTM lung infection and specific types of lung cancer, highlighting the complex relationship between microbial colonization and lung carcinogenesis. Despite previously being classified as contaminants there is growing recognition of the potential role of certain bacteria in cancer development and progression. Emerging evidence suggests that NTM may also integrate the intratumoral microbiome, interact with cancer cells, particularly in breast cancer. In this context, chronic ingestion of NTM with tap water may play a significant yet undetermined role, as they may be able to migrate from the gut to the breast, like various other bacteria of intestinal origin found in breast tissue and human milk. However, the mechanisms and implications for breast cancer biology remain subjects of ongoing and future research. Investigating the intratumoral microbiome poses significant technical challenges due to its inherently low biomass. These include managing sample and database contamination, addressing batch effects, refining analytical pipelines, and rectifying problematic data processing methods, all of which could compromise study outcomes. Therefore, rigorous research with robust controls and suitable analytical tools for low-biomass microbiome analysis are imperative. Integrating sequencing data with imaging and culturing techniques, along with utilizing cellular and animal models, is essential for confirming causation and elucidating molecular mechanisms. Studying the diversity, epidemiology, and prevalence of the diverse microbiota including NTM in drinking water, and their impact on lung infections and the gut-breast axis, as well as the potential involvement in lung and breast cancer pathogenesis, is a dynamic and evolving research frontier (Figure 1). This field harbors considerable potential for deepening our comprehension of both public health and cancer biology, paving the way for groundbreaking insights into disease origins and the exploration of pioneering preventive and therapeutic approaches aimed at safeguarding population’s health.
Author Contributions
Conceptualization, resources, and funding acquisition NE; Investigation, AM, SA, DN-C, PC, NE; Writing-original draft, AM, DN-C, IM-M, IR, PC, OC, NE. Writing-review and editing, AM, DN-C, PC, NE. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fundação para a Ciência e a Tecnologia projects, PTDC/BIA-MIC/0122/2021, 2022.06809.PTDC, UIDB/04539/2020, UIDP/04539/2020, LA/P/0058/2020.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgements
AM and DN-C acknowledge project PTDC/BIA-MIC/0122/2021 for contract and BPD grant, respectively. IM-M is supported by contract under Cure Parkinson’s UK (CP:SC01). IR and SA are supported by PhD scholarship SFRH/BD/145135/2019 and contract DL57/2016-SFRH/BPD/108299/2015, respectively.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- NRC. Indicators for Waterborne Pathogens; National Research Council (NRC); Division on Earth and Life Studies; Board on Life Sciences; Water Science and Technology Board; Committee on Indicators for Waterborne Pathogens: Washington, DC, 2004; p. 328. [Google Scholar]
- WHO. Guidelines for drinking-water quality Fourth edition incorporating the first and second addenda. 2022.
- Manaia, C.M.; Aga, D.S.; Cytryn, E.; Gaze, W.H.; Graham, D.W.; Guo, J.; Leonard, A.F.C.; Li, L.; Murray, A.K.; Nunes, O.C.; et al. The Complex Interplay Between Antibiotic Resistance and Pharmaceutical and Personal Care Products in the Environment. Environ. Toxicol. Chem. 2022. [Google Scholar] [CrossRef]
- Vanhaecke, T.; Bretin, O.; Poirel, M.; Tap, J. Drinking Water Source and Intake Are Associated with Distinct Gut Microbiota Signatures in US and UK Populations. The Journal of Nutrition 2022, 152, 171–182. [Google Scholar] [CrossRef]
- IOM. Global Issues in Water, Sanitation, and Health: Workshop Summary; Institute of Medicine The National Academies Press: Washington, DC, 2009; p. 328. [Google Scholar]
- Falkinham III, J.O.; Pruden, A.; Edwards, M. Opportunistic Premise Plumbing Pathogens: Increasingly Important Pathogens in Drinking Water. Pathogens 2015, 4, 373–386. [Google Scholar] [CrossRef]
- Abkar, L.; Moghaddam, H.S.; Fowler, S.J. Microbial ecology of drinking water from source to tap. Sci. Total Environ. 2023, 908, 168077. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Li, W.; Chen, J.; Zhou, Y.; Wei, Z.; Gong, L. Microbial diversity in full-scale water supply systems through sequencing technology: a review. RSC Adv. 2021, 11, 25484–25496. [Google Scholar] [CrossRef] [PubMed]
- Hammes, F.; Berney, M.; Wang, Y.; Vital, M.; Koster, O.; Egli, T. Flow-cytometric total bacterial cell counts as a descriptive microbiological parameter for drinking water treatment processes. Water Res. 2008, 42, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhang, Y.; van der Mark, E.; Magic-Knezev, A.; Pinto, A.; van den Bogert, B.; Liu, W.; van der Meer, W.; Medema, G. Assessing the origin of bacteria in tap water and distribution system in an unchlorinated drinking water system by SourceTracker using microbial community fingerprints. Water Res. 2018, 138, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Dinkla, I.J.T.; Muyzer, G. Microbial ecology of biofiltration used for producing safe drinking water. Appl. Microbiol. Biotechnol. 2022, 106, 4813–4829. [Google Scholar] [CrossRef]
- Pinto, A.J.; Xi, C.; Raskin, L. Bacterial community structure in the drinking water microbiome is governed by filtration processes. Environ. Sci. Technol. 2012, 46, 8851–8859. [Google Scholar] [CrossRef]
- Bruno, A.; Agostinetto, G.; Fumagalli, S.; Ghisleni, G.; Sandionigi, A. It’s a Long Way to the Tap: Microbiome and DNA-Based Omics at the Core of Drinking Water Quality. Int. J. Env. Res. Public Health 2022, 19, 7940. [Google Scholar] [CrossRef]
- Thom, C.; Smith, C.J.; Moore, G.; Weir, P.; Ijaz, U.Z. Microbiomes in drinking water treatment and distribution: A meta-analysis from source to tap. Water Res. 2022, 212, 118106. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Sevillano-Rivera, M.C.; Calus, S.T.; Bautista-de Los Santos, Q.M.; Eren, A.M.; van der Wielen, P.; Ijaz, U.Z.; Pinto, A.J. Disinfection exhibits systematic impacts on the drinking water microbiome. Microbiome 2020, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Delafont, V.; Brouke, A.; Bouchon, D.; Moulin, L.; Hechard, Y. Microbiome of free-living amoebae isolated from drinking water. Water Res. 2013, 47, 6958–6965. [Google Scholar] [CrossRef] [PubMed]
- Ling, F.; Hwang, C.; LeChevallier, M.W.; Andersen, G.L.; Liu, W.T. Core-satellite populations and seasonality of water meter biofilms in a metropolitan drinking water distribution system. ISME J. 2016, 10, 582–595. [Google Scholar] [CrossRef]
- Revetta, R.P.; Gomez-Alvarez, V.; Gerke, T.L.; Curioso, C.; Santo Domingo, J.W.; Ashbolt, N.J. Establishment and early succession of bacterial communities in monochloramine-treated drinking water biofilms. FEMS Microbiol. Ecol. 2013, 86, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Revetta, R.P.; Gomez-Alvarez, V.; Gerke, T.L.; Santo Domingo, J.W.; Ashbolt, N.J. Changes in bacterial composition of biofilm in a metropolitan drinking water distribution system. J. Appl. Microbiol. 2016, 121, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Diversity and Antibiotic Resistance Patterns of Sphingomonadaceae Isolates from Drinking Water. Appl. Environ. Microbiol. 2011, 77, 5697–5706. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Wang, W.; Liu, Y.; Liu, S.; Lou, L.; Cheng, D.; He, X.; Zhou, X.; Qiu, S.; Fu, L.; et al. Pyrosequencing analysis of bacterial communities in biofilms from different pipe materials in a city drinking water distribution system of East China. Appl. Microbiol. Biotechnol. 2015, 99, 10713–10724. [Google Scholar] [CrossRef]
- Aggarwal, S.; Gomez-Smith, C.K.; Jeon, Y.; LaPara, T.M.; Waak, M.B.; Hozalski, R.M. Effects of Chloramine and Coupon Material on Biofilm Abundance and Community Composition in Bench-Scale Simulated Water Distribution Systems and Comparison with Full-Scale Water Mains. Environ. Sci. Technol. 2018, 52, 13077–13088. [Google Scholar] [CrossRef]
- Waak, M.B.; Hozalski, R.M.; Hallé, C.; LaPara, T.M. Comparison of the microbiomes of two drinking water distribution systems—with and without residual chloramine disinfection. Microbiome 2019, 7, 87. [Google Scholar] [CrossRef]
- Bautista-de los Santos, Q.M.; Schroeder, J.L.; Sevillano-Rivera, M.C.; Sungthong, R.; Ijaz, U.Z.; Sloan, W.T.; Pinto, A.J. Emerging investigators series: microbial communities in full-scale drinking water distribution systems – a meta-analysis. Environ. Sci.: Water Res. Technol. 2016, 2, 631–644. [Google Scholar] [CrossRef]
- Chiao, T.-H.; Clancy, T.M.; Pinto, A.; Xi, C.; Raskin, L. Differential Resistance of Drinking Water Bacterial Populations to Monochloramine Disinfection. Environ. Sci. Technol. 2014, 48, 4038–4047. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Bédard, E.; Prévost, M.; Camper, A.K.; Hill, V.R.; Pruden, A. Methodological approaches for monitoring opportunistic pathogens in premise plumbing: A review. Water Res. 2017, 117, 68–86. [Google Scholar] [CrossRef] [PubMed]
- Feazel, L.M.; Baumgartner, L.K.; Peterson, K.L.; Frank, D.N.; Harris, J.K.; Pace, N.R. Opportunistic pathogens enriched in showerhead biofilms. Proc. Natl. Acad. Sci. 2009, 106, 16393–16399. [Google Scholar] [CrossRef] [PubMed]
- Hayward, C.; Ross, K.E.; Brown, M.H.; Bentham, R.; Whiley, H. The Presence of Opportunistic Premise Plumbing Pathogens in Residential Buildings: A Literature Review. Water 2022, 14, 1129. [Google Scholar] [CrossRef]
- De Sotto, R.; Tang, R.; Bae, S. Biofilms in premise plumbing systems as a double-edged sword: microbial community composition and functional profiling of biofilms in a tropical region. J. Water Health 2020, 18, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Hayward, C.; Brown, M.H.; Whiley, H. Hospital water as the source of healthcare-associated infection and antimicrobial-resistant organisms. Curr. Opin. Infect. Dis. 2022, 35, 339–345. [Google Scholar] [CrossRef]
- Logan-Jackson, A.R.; Batista, M.D.; Healy, W.; Ullah, T.; Whelton, A.J.; Bartrand, T.A.; Proctor, C. A Critical Review on the Factors that Influence Opportunistic Premise Plumbing Pathogens: From Building Entry to Fixtures in Residences. Environ. Sci. Technol. 2023, 57, 6360–6372. [Google Scholar] [CrossRef]
- Shen, Y.; Haig, S.-J.; Prussin, A.J., II; LiPuma, J.J.; Marr, L.C.; Raskin, L. Shower water contributes viable nontuberculous mycobacteria to indoor air. PNAS Nexus 2022, 1. [Google Scholar] [CrossRef]
- Gebert, M.J.; Delgado-Baquerizo, M.; Oliverio, A.M.; Webster, T.M.; Nichols, L.M.; Honda, J.R.; Chan, E.D.; Adjemian, J.; Dunn, R.R.; Fierer, N. Ecological Analyses of Mycobacteria in Showerhead Biofilms and Their Relevance to Human Health. mBio 2018, 9, 10–1128. [Google Scholar] [CrossRef]
- Thomson, R.M.; Tolson, C.; Carter, R.; Coulter, C.; Huygens, F.; Hargreaves, M. Isolation of Nontuberculous Mycobacteria (NTM) from Household Water and Shower Aerosols in Patients with Pulmonary Disease Caused by NTM. J. Clin. Microbiol. 2013, 51, 3006–3011. [Google Scholar] [CrossRef] [PubMed]
- Martin, I.; Pfyffer, G.E.; Parrish, N. Procedures. In Manual of Clinical Microbiology, 12th ed.; Carroll, K.C., Pfaller, M.A., Eds.; ASM Press: Washington, D.C, 2023. [Google Scholar]
- Etymologia: Mycobacterium. Emerg. Infect. Dis. 2008, 14, 377. [CrossRef]
- Fedrizzi, T.; Meehan, C.J.; Grottola, A.; Giacobazzi, E.; Fregni Serpini, G.; Tagliazucchi, S.; Fabio, A.; Bettua, C.; Bertorelli, R.; De Sanctis, V.; et al. Genomic characterization of Nontuberculous Mycobacteria. Sci Rep 2017, 7, 45258. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.S.; Lo, B.; Son, J. Phylogenomics and Comparative Genomic Studies Robustly Support Division of the Genus Mycobacterium into an Emended Genus Mycobacterium and Four Novel Genera. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Meehan, C.J.; Barco, R.A.; Loh, Y.-H.E.; Cogneau, S.; Rigouts, L. Reconstituting the genus Mycobacterium. Int. J. Syst. Evol. Microbiol. 2021, 71, 004922. [Google Scholar] [CrossRef] [PubMed]
- Falkinham III, J.O. Nontuberculous mycobacteria in the environment. Tuberculosis 2022, 137, 102267. [Google Scholar] [CrossRef]
- Magee, J.G.; Ward, A.C. Mycobacterium. In Bergey's Manual of Systematics of Archaea and Bacteria; 2015; pp. 1-84.
- Drapal, M.; Wheeler, P.R.; Fraser, P.D. Metabolite analysis of Mycobacterium species under aerobic and hypoxic conditions reveals common metabolic traits. Microbiology 2016, 162, 1456–1467. [Google Scholar] [CrossRef]
- Kalia, N.P.; Singh, S.; Hards, K.; Cheung, C.-Y.; Sviriaeva, E.; Banaei-Esfahani, A.; Aebersold, R.; Berney, M.; Cook, G.M.; Pethe, K. M. tuberculosis relies on trace oxygen to maintain energy homeostasis and survive in hypoxic environments. Cell Reports 2023, 42, 112444. [Google Scholar] [CrossRef]
- Ashenafi, S.; Brighenti, S. Reinventing the human tuberculosis (TB) granuloma: Learning from the cancer field. Front. immunol. 2022, 13. [Google Scholar] [CrossRef]
- Shi, Y.; Queller, D.C.; Tian, Y.; Zhang, S.; Yan, Q.; He, Z.; He, Z.; Wu, C.; Wang, C.; Shu, L. The Ecology and Evolution of Amoeba-Bacterium Interactions. Appl. Environ. Microbiol. 2021, 87, e01866–01820. [Google Scholar] [CrossRef]
- Pereira, A.C.; Ramos, B.; Reis, A.C.; Cunha, M.V. Non-Tuberculous Mycobacteria: Molecular and Physiological Bases of Virulence and Adaptation to Ecological Niches. Microorganisms 2020, 8, 1380. [Google Scholar] [CrossRef] [PubMed]
- Blanc, S.M.; Robinson, D.; Fahrenfeld, N.L. Potential for nontuberculous mycobacteria proliferation in natural and engineered water systems due to climate change: A literature review. City Environ. Interact. 2021, 11, 100070. [Google Scholar] [CrossRef]
- Brown-Elliott, B.A.; Philley, J.V. Rapidly Growing Mycobacteria. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Holt, M.R.; Kasperbauer, S. Management of Extrapulmonary Nontuberculous Mycobacterial Infections. Semin. Respir. Crit. Care Med. 2018, 39, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Costa, D.; Alarico, S.; Dalcolmo, M.P.; Correia-Neves, M.; Empadinhas, N. The looming tide of nontuberculous mycobacterial infections in Portugal and Brazil. Tuberculosis 2016, 96, 107–119. [Google Scholar] [CrossRef]
- Omori, K.; Kitagawa, H.; Yamaguchi, K.; Sakamoto, S.; Horimasu, Y.; Masuda, T.; Miyamoto, S.; Nakashima, T.; Iwamoto, H.; Fujitaka, K.; et al. Clinical characteristics of extrapulmonary nontuberculous mycobacteria infections in comparison with pulmonary infections: A single-center, retrospective study in Japan. J. Infect. Chemother. 2023, 29, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Grigg, C.; Jackson, K.A.; Barter, D.; Czaja, C.A.; Johnston, H.; Lynfield, R.; Vagnone, P.S.; Tourdot, L.; Spina, N.; Dumyati, G.; et al. Epidemiology of Pulmonary and Extrapulmonary Nontuberculous Mycobacteria Infections at 4 US Emerging Infections Program Sites: A 6-Month Pilot. Clin. Infect. Dis. 2023, 77, 629–637. [Google Scholar] [CrossRef]
- Cassidy, P.M.; Hedberg, K.; Saulson, A.; McNelly, E.; Winthrop, K.L. Nontuberculous Mycobacterial Disease Prevalence and Risk Factors: A Changing Epidemiology. Clin. Infect. Dis. 2009, 49, e124–e129. [Google Scholar] [CrossRef]
- Prevots, D.R.; Marshall, J.E.; Wagner, D.; Morimoto, K. Global Epidemiology of Nontuberculous Mycobacterial Pulmonary Disease: A Review. Clin. Chest. Med. 2023, 44, 675–721. [Google Scholar] [CrossRef]
- Ricotta, E.; Adjemian, J.; Blakney, R.; Lai, Y.L.; Kadri, S.; Prevots, D.R. Extrapulmonary Nontuberculous Mycobacteria Infections in Hospitalized Patients, United States, 2009–2014. Emerg. Infect. Dis. 2021, 27, 845. [Google Scholar] [CrossRef]
- Griffith, D.E.; Aksamit, T.R. Diagnostic Criteria and the Decision to Treat Nontuberculous Mycobacterial Pulmonary Disease. Clin. Chest. Med. 2023, 44, 757–769. [Google Scholar] [CrossRef] [PubMed]
- Sawka, A.; Burke, A. Medications and Monitoring in Treatment of Nontuberculous Mycobacterial Pulmonary Disease. Clin. Chest. Med. 2023, 44, 815–828. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, L.E.; Waterer, G. Beyond antibiotics: recent developments in the diagnosis and management of nontuberculous mycobacterial infection. Breathe 2022, 18. [Google Scholar] [CrossRef] [PubMed]
- Alffenaar, J.-W.; Märtson, A.-G.; Heysell, S.K.; Cho, J.-G.; Patanwala, A.; Burch, G.; Kim, H.Y.; Sturkenboom, M.G.G.; Byrne, A.; Marriott, D.; et al. Therapeutic Drug Monitoring in Non-Tuberculosis Mycobacteria Infections. Clin Pharmacokinet 2021, 60, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Honda, J.R.; Virdi, R.; Chan, E.D. Global Environmental Nontuberculous Mycobacteria and Their Contemporaneous Man-Made and Natural Niches. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Chan, E.D.; Iseman, M.D. Slender, Older Women Appear to Be More Susceptible to Nontuberculous Mycobacterial Lung Disease. Gender Medicine 2010, 7, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Kang, M.J.; Han, C.H.; Lee, S.M.; Kim, C.J.; Lee, J.M.; Kang, Y.A. Prevalence, incidence, and mortality of nontuberculous mycobacterial infection in Korea: a nationwide population-based study. BMC Pulmonary Medicine 2019, 19, 140. [Google Scholar] [CrossRef] [PubMed]
- Weathered, C.; Wei, N.; Pienaar, E. Reduced macrophage killing of M. avium drives infection risk in post-menopausal patients. Tuberculosis 2023, 139, 102304. [Google Scholar] [CrossRef] [PubMed]
- Namkoong, H.; Holland, S.M. Host Susceptibility to Nontuberculous Mycobacterial Pulmonary Disease. Clin. Chest. Med. 2023, 44, 723–730. [Google Scholar] [CrossRef]
- Namkoong, H.; Omae, Y.; Asakura, T.; Ishii, M.; Suzuki, S.; Morimoto, K.; Kawai, Y.; Emoto, K.; Oler, A.J.; Szymanski, E.P.; et al. Genome-wide association study in patients with pulmonary Mycobacterium avium complex disease. Eur. Respir. J. 2021. [Google Scholar] [CrossRef]
- van Ingen, J.; Aksamit, T.; Andrejak, C.; Böttger, E.C.; Cambau, E.; Daley, C.L.; Griffith, D.E.; Guglielmetti, L.; Holland, S.M.; Huitt, G.A.; et al. Treatment outcome definitions in nontuberculous mycobacterial pulmonary disease: an NTM-NET consensus statement. Eur. Respir. J. 2018, 51, 1800170. [Google Scholar] [CrossRef] [PubMed]
- Winthrop, K.L.; Marras, T.K.; Adjemian, J.; Zhang, H.; Wang, P.; Zhang, Q. Incidence and Prevalence of Nontuberculous Mycobacterial Lung Disease in a Large U.S. Managed Care Health Plan, 2008–2015. Ann. Am. Thorac. Soc. 2020, 17, 178–185. [Google Scholar] [CrossRef]
- Schildkraut, J.A.; Gallagher, J.; Morimoto, K.; Lange, C.; Haworth, C.; Floto, R.A.; Hoefsloot, W.; Griffith, D.E.; Wagner, D.; Ingen, J.v. Epidemiology of nontuberculous mycobacterial pulmonary disease in Europe and Japan by Delphi estimation. Respir. Med. 2020, 173. [Google Scholar] [CrossRef]
- Zhou, Y.; Mu, W.; Zhang, J.; Wen, S.W.; Pakhale, S. Global prevalence of non-tuberculous mycobacteria in adults with non-cystic fibrosis bronchiectasis 2006–2021: a systematic review and meta-analysis. BMJ Open 2022, 12, e055672. [Google Scholar] [CrossRef]
- De Groote, M.A.; Pace, N.R.; Fulton, K.; Falkinham III, J.O. Relationships between Mycobacterium Isolates from Patients with Pulmonary Mycobacterial Infection and Potting Soils. Appl. Environ. Microbiol. 2006, 72, 7602–7606. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, K.; Ito, Y.; Hirai, T.; Kubo, T.; Imai, S.; Tatsumi, S.; Fujita, K.; Takakura, S.; Niimi, A.; Iinuma, Y.; et al. Environmental Risk Factors for Pulmonary Mycobacterium avium-intracellulare Complex Disease. Chest 2011, 140, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Honda, J.R. Environmental Sources and Transmission of Nontuberculous Mycobacteria. Clin. Chest. Med. 2023, 44, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Nishiuchi, Y.; Maekura, R.; Kitada, S.; Tamaru, A.; Taguri, T.; Kira, Y.; Hiraga, T.; Hirotani, A.; Yoshimura, K.; Miki, M.; et al. The Recovery of Mycobacterium avium-intracellulare Complex (MAC) from the Residential Bathrooms of Patients with Pulmonary MAC. Clin. Infect. Dis. 2007, 45, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Tzou, C.L.; Dirac, M.A.; Becker, A.L.; Beck, N.K.; Weigel, K.M.; Meschke, J.S.; Cangelosi, G.A. Association between Mycobacterium avium Complex Pulmonary Disease and Mycobacteria in Home Water and Soil. Ann. Am. Thorac. Soc. 2020, 17, 57–62. [Google Scholar] [CrossRef]
- Lande, L.; Alexander, D.C.; Wallace, R.J.; Kwait, R.; Iakhiaeva, E.; Williams, M.; Cameron, A.D.S.; Olshefsky, S.; Devon, R.; Vasireddy, R.; et al. Mycobacterium avium in Community and Household Water, Suburban Philadelphia, Pennsylvania, USA, 2010–2012. Emerg. Infect. Dis. 2019, 25, 473–481. [Google Scholar] [CrossRef]
- Griffin, I.; Schmitz, A.; Oliver, C.; Pritchard, S.; Zhang, G.; Rico, E.; Davenport, E.; Llau, A.; Moore, E.; Fernandez, D.; et al. Outbreak of Tattoo-associated Nontuberculous Mycobacterial Skin Infections. Clin. Infect. Dis. 2019, 69, 949–955. [Google Scholar] [CrossRef] [PubMed]
- CDC. Tattoo-associated nontuberculous mycobacterial skin infections--multiple states, 2011-2012. MMWR Morb Mortal Wkly Rep 2012, 61, 653–656. [Google Scholar]
- Fjällbrant, H.; Akerstrom, M.; Svensson, E.; Andersson, E. Hot tub lung: an occupational hazard. Eur. Respir. Rev. 2013, 22, 88–90. [Google Scholar] [CrossRef]
- Prevots, D.R.; Adjemian, J.; Fernandez, A.G.; Knowles, M.R.; Olivier, K.N. Environmental Risks for Nontuberculous Mycobacteria. Individual Exposures and Climatic Factors in the Cystic Fibrosis Population. Ann. Am. Thorac. Soc. 2014, 11, 1032–1038. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Kwak, S.H.; Yong, S.H.; Lee, S.H.; Leem, A.Y.; Kim, S.Y.; Lee, S.H.; Chung, K.; Kim, E.Y.; Jung, J.Y.; et al. The Association between Behavioral Risk Factors and Nontuberculous Mycobacterial Pulmonary Disease. Yonsei Med. J. 2021, 62, 702–707. [Google Scholar] [CrossRef]
- Lyman, M.M.; Grigg, C.; Kinsey, C.B.; Keckler, M.S.; Moulton-Meissner, H.; Cooper, E.; Soe, M.M.; Noble-Wang, J.; Longenberger, A.; Walker, S.R.; et al. Invasive Nontuberculous Mycobacterial Infections among Cardiothoracic Surgical Patients Exposed to Heater-Cooler Devices. Emerging Infect. Dis. 2017, 23, 796–805. [Google Scholar] [CrossRef]
- Peralta, G.; Tobin-D'Angelo, M.; Parham, A.; Edison, L.; Lorentzson, L.; Smith, C.; Drenzek, C. Notes from the Field: Mycobacterium abscessus Infections Among Patients of a Pediatric Dentistry Practice--Georgia, 2015. MMWR Morb Mortal Wkly Rep 2016, 65, 355–356. [Google Scholar] [CrossRef]
- Padoveze, M.C.; Fortaleza, C.M.C.B.; Freire, M.P.; Assis, D.B.d.; Madalosso, G.; Pellini, A.C.G.; César, M.L.V.; Neto, V.P.; Beltramelli, M.M.; Chimara, E.; et al. Outbreak of surgical infection caused by non-tuberculous mycobacteria in breast implants in Brazil. J. Hosp. Infect. 2007, 67, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Daniau, C.; Lecorche, E.; Mougari, F.; Benmansour, H.; Bernet, C.; Blanchard, H.; Robert, J.; Berger-Carbonne, A.; Cambau, E. Association of Healthcare and Aesthetic Procedures with Infections Caused by Nontuberculous Mycobacteria, France, 2012‒2020. Emerging Infect. Dis. 2022, 28, 518–526. [Google Scholar] [CrossRef]
- Shapiro, K.; Cross, S.J.; Morton, T.H.; Inaba, H.; Holland, A.; Fasipe, F.R.; Adderson, E.E. Healthcare-Associated Infections Caused by Mycolicibacterium neoaurum. Emerging Infect. Dis. 2023, 29, 1516–1523. [Google Scholar] [CrossRef]
- van der Wielen, P.W.J.J.; van der Kooij, D. Nontuberculous Mycobacteria, Fungi, and Opportunistic Pathogens in Unchlorinated Drinking Water in the Netherlands. Appl. Environ. Microbiol. 2013, 79, 825–834. [Google Scholar] [CrossRef]
- Thomson, R.M.; Carter, R.; Tolson, C.; Coulter, C.; Huygens, F.; Hargreaves, M. Factors associated with the isolation of Nontuberculous mycobacteria (NTM) from a large municipal water system in Brisbane, Australia. BMC Microbiol. 2013, 13, 89. [Google Scholar] [CrossRef]
- Thomson, R.M.; Furuya-Kanamori, L.; Coffey, C.; Bell, S.C.; Knibbs, L.D.; Lau, C.L. Influence of climate variables on the rising incidence of nontuberculous mycobacterial (NTM) infections in Queensland, Australia 2001–2016. Sci. Total Environ. 2020, 740, 139796. [Google Scholar] [CrossRef] [PubMed]
- Adjemian, J.; Olivier, K.N.; Seitz, A.E.; Falkinham III, J.O.; Holland, S.M.; Prevots, D.R. Spatial Clusters of Nontuberculous Mycobacterial Lung Disease in the United States. Am. J. Respir. Crit. Care Med. 2012, 186, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Lipner, E.M.; Crooks, J.L.; French, J.; Strong, M.; Nick, J.A.; Prevots, D.R. Nontuberculous mycobacterial infection and environmental molybdenum in persons with cystic fibrosis: a case–control study in Colorado. J. Expo. Sci. Environ. Epidemiol. 2022, 32, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Lipner, E.M.; French, J.P.; Falkinham III, J.O.; Crooks, J.L.; Mercaldo, R.A.; Henkle, E.; Prevots, D.R. Nontuberculous Mycobacteria Infection Risk and Trace Metals in Surface Water: A Population-based Ecologic Epidemiologic Study in Oregon. Ann. Am. Thorac. Soc. 2022, 19, 543–550. [Google Scholar] [CrossRef]
- DeFlorio-Barker, S.; Egorov, A.; Smith, G.S.; Murphy, M.S.; Stout, J.E.; Ghio, A.J.; Hudgens, E.E.; Messier, K.P.; Maillard, J.-M.; Hilborn, E.D. Environmental risk factors associated with pulmonary isolation of nontuberculous mycobacteria, a population-based study in the southeastern United States. Sci. Total Environ. 2021, 763, 144552. [Google Scholar] [CrossRef]
- Kumar, K.; Ponnuswamy, A.; Capstick, T.G.D.; Chen, C.; McCabe, D.; Hurst, R.; Morrison, L.; Moore, F.; Gallardo, M.; Keane, J.; et al. Non-tuberculous mycobacterial pulmonary disease (NTM-PD): Epidemiology, diagnosis and multidisciplinary management. Clinical Medicine 2024, 24, 100017. [Google Scholar] [CrossRef]
- Falkinham III, J.O.; Norton, C.D.; LeChevallier, M.W. Factors Influencing Numbers of Mycobacterium avium, Mycobacterium intracellulare, and Other Mycobacteria in Drinking Water Distribution Systems. Appl. Environ. Microbiol. 2001, 67, 1225–1231. [Google Scholar] [CrossRef]
- Donohue, M.J.; Mistry, J.H.; Donohue, J.M.; O’Connell, K.; King, D.; Byran, J.; Covert, T.; Pfaller, S. Increased Frequency of Nontuberculous Mycobacteria Detection at Potable Water Taps within the United States. Environ. Sci. Technol. 2015, 49, 6127–6133. [Google Scholar] [CrossRef]
- Torvinen, E.; Suomalainen, S.; Lehtola, M.J.; Miettinen, I.T.; Zacheus, O.; Paulin, L.; Katila, M.-L.; Martikainen, P.J. Mycobacteria in Water and Loose Deposits of Drinking Water Distribution Systems in Finland. Appl. Environ. Microbiol. 2004, 70, 1973–1981. [Google Scholar] [CrossRef]
- Dantec, C.L.; Duguet, J.-P.; Montiel, A.; Dumoutier, N.; Dubrou, S.; Vincent, V. Occurrence of Mycobacteria in Water Treatment Lines and in Water Distribution Systems. Appl. Environ. Microbiol. 2002, 68, 5318–5325. [Google Scholar] [CrossRef] [PubMed]
- Loret, J.-F.; Dumoutier, N. Non-tuberculous mycobacteria in drinking water systems: A review of prevalence data and control means. Int. J. Hyg. Environ. Health 2019, 222, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Liu, T.; Men, Y.; Li, S.; Graham, N.; Yu, W. Understanding point-of-use tap water quality: From instrument measurement to intelligent analysis using sample filtration. Water Res. 2022, 225, 119205. [Google Scholar] [CrossRef]
- Norton, G.J.; Williams, M.; Falkinham III, J.O.; Honda, J.R. Physical Measures to Reduce Exposure to Tap Water–Associated Nontuberculous Mycobacteria. Frontiers in Public Health 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Säve-Söderbergh, M.; Toljander, J.; Mattisson, I.; Åkesson, A.; Simonsson, M. Drinking water consumption patterns among adults—SMS as a novel tool for collection of repeated self-reported water consumption. J. Expo. Sci. Environ. Epidemiol. 2018, 28, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Rosinger, A.Y.; Herrick, K.A.; Wutich, A.Y.; Yoder, J.S.; Ogden, C.L. Disparities in plain, tap and bottled water consumption among US adults: National Health and Nutrition Examination Survey (NHANES) 2007–2014. Public Health Nutrition 2018, 21, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Chongwe, G.; Michelo, C.; Kelly, P. Diagnostic yield of nontuberculous mycobacteria in patients booked for endoscopy at the University Teaching Hospital, Lusaka. BMC Research Notes 2017, 10, 27. [Google Scholar] [CrossRef]
- Falony, G.; Joossens, M.; Vieira-Silva, S.; Wang, J.; Darzi, Y.; Faust, K.; Kurilshikov, A.; Bonder, M.J.; Valles-Colomer, M.; Vandeputte, D.; et al. Population-level analysis of gut microbiome variation. Science 2016, 352, 560–564. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA: A Cancer Journal for Clinicians 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer Journal for Clinicians 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Luengo-Fernandez, R.; Leal, J.; Gray, A.; Sullivan, R. Economic burden of cancer across the European Union: a population-based cost analysis. The Lancet Oncology 2013, 14, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Allemani, C.; Weir, H.K.; Carreira, H.; Harewood, R.; Spika, D.; Wang, X.-S.; Bannon, F.; Ahn, J.V.; Johnson, C.J.; Bonaventure, A.; et al. Global surveillance of cancer survival 1995–2009: analysis of individual data for 25 676 887 patients from 279 population-based registries in 67 countries (CONCORD-2). The Lancet 2015, 385, 977–1010. [Google Scholar] [CrossRef] [PubMed]
- Ginsburg, O.; Bray, F.; Coleman, M.P.; Vanderpuye, V.; Eniu, A.; Kotha, S.R.; Sarker, M.; Huong, T.T.; Allemani, C.; Dvaladze, A.; et al. The global burden of women’s cancers: a grand challenge in global health. The Lancet 2017, 389, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Parmigiani, G. Meta-analysis of BRCA1 and BRCA2 penetrance. Journal of clinical oncology : official journal of the American Society of Clinical Oncology 2007, 25, 1329–1333. [Google Scholar] [CrossRef]
- Nur, U.; El Reda, D.; Hashim, D.; Weiderpass, E. A prospective investigation of oral contraceptive use and breast cancer mortality: findings from the Swedish women’s lifestyle and health cohort. BMC Cancer 2019, 19, 807. [Google Scholar] [CrossRef] [PubMed]
- Mørch, L.S.; Skovlund, C.W.; Hannaford, P.C.; Iversen, L.; Fielding, S.; Lidegaard, Ø. Contemporary Hormonal Contraception and the Risk of Breast Cancer. New Engl. J. Med. 2017, 377, 2228–2239. [Google Scholar] [CrossRef] [PubMed]
- CGHFBC. Collaborative Group on Hormonal Factors in Breast Cancer. Type and timing of menopausal hormone therapy and breast cancer risk: individual participant meta-analysis of the worldwide epidemiological evidence. The Lancet 2019, 394, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Piccart-Gebhart, M.J.; Procter, M.; Leyland-Jones, B.; Goldhirsch, A.; Untch, M.; Smith, I.; Gianni, L.; Baselga, J.; Bell, R.; Jackisch, C.; et al. Trastuzumab after Adjuvant Chemotherapy in HER2-Positive Breast Cancer. New Engl. J. Med. 2005, 353, 1659–1672. [Google Scholar] [CrossRef]
- Foulkes, W.D.; Smith, I.E.; Reis-Filho, J.S. Triple-Negative Breast Cancer. New Engl. J. Med. 2010, 363, 1938–1948. [Google Scholar] [CrossRef]
- Pohl-Rescigno, E.; Hauke, J.; Loibl, S.; Möbus, V.; Denkert, C.; Fasching, P.A.; Kayali, M.; Ernst, C.; Weber-Lassalle, N.; Hanusch, C.; et al. Association of Germline Variant Status With Therapy Response in High-risk Early-Stage Breast Cancer: A Secondary Analysis of the GeparOcto Randomized Clinical Trial. JAMA Oncology 2020, 6, 744–748. [Google Scholar] [CrossRef] [PubMed]
- Luen, S.; Virassamy, B.; Savas, P.; Salgado, R.; Loi, S. The genomic landscape of breast cancer and its interaction with host immunity. The Breast 2016, 29, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Tang, Y.; Wang, T.; Yang, Y.; Zhang, Y.; Wang, R.; Zhang, Y.; Li, Y.; Wu, M.; Tang, M.; et al. Chronic pulmonary bacterial infection facilitates breast cancer lung metastasis by recruiting tumor-promoting MHCIIhi neutrophils. Signal Transduction and Targeted Therapy 2023, 8, 296. [Google Scholar] [CrossRef] [PubMed]
- Roche, J. The Epithelial-to-Mesenchymal Transition in Cancer. Cancers (Basel) 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Philley, J.V.; Kannan, A.; Griffith, D.E.; Devine, M.S.; Benwill, J.L.; Wallace, R.J.; Brown-Elliott, B.A.; Thakkar, F.; Taskar, V.S.; Fox, J.G.; et al. Exosome secretome and mediated signaling in breast cancer patients with nontuberculous mycobacterial disease. Oncotarget 2017, 8, 18070–18081. [Google Scholar] [CrossRef] [PubMed]
- Butlin, H.T. Malignant Tumours and Parasitism. Br. Med. J. 1884, 1, 45–46. [Google Scholar] [CrossRef]
- Møller, H.; Heseltine, E.; Vainio, H. Working group report on schistosomes, liver flukes and Helicobacter pylori. Meeting held at IARC, LYON, 7–14 june 1994. Int. J. Cancer 1995, 60, 587–589. [Google Scholar] [CrossRef] [PubMed]
- El Tekle, G.; Garrett, W.S. Bacteria in cancer initiation, promotion and progression. Nature Reviews Cancer 2023, 23, 600–618. [Google Scholar] [CrossRef]
- Sepich-Poore, G.D.; Zitvogel, L.; Straussman, R.; Hasty, J.; Wargo, J.A.; Knight, R. The microbiome and human cancer. Science 2021, 371, eabc4552. [Google Scholar] [CrossRef]
- Laliani, G.; Ghasemian Sorboni, S.; Lari, R.; Yaghoubi, A.; Soleimanpour, S.; Khazaei, M.; Hasanian, S.M.; Avan, A. Bacteria and cancer: Different sides of the same coin. Life Sci. 2020, 246, 117398. [Google Scholar] [CrossRef]
- Song, X.; Wei, C.; Li, X. The Relationship Between Microbial Community and Breast Cancer. Front. Cell. Infect. Microbiol. 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, S.; Parida, S.; Lingipilli, B.T.; Krishnan, R.; Podipireddy, D.R.; Muniraj, N. Role of Gut Microbiota in Breast Cancer and Drug Resistance. Pathogens 2023, 12, 468. [Google Scholar] [CrossRef] [PubMed]
- Vitorino, M.; Baptista de Almeida, S.; Alpuim Costa, D.; Faria, A.; Calhau, C.; Azambuja Braga, S. Human Microbiota and Immunotherapy in Breast Cancer - A Review of Recent Developments. Front. Oncol. 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Parida, S.; Sharma, D. The Microbiome–Estrogen Connection and Breast Cancer Risk. Cells 2019, 8, 1642. [Google Scholar] [CrossRef] [PubMed]
- Alpuim Costa, D.; Nobre, J.G.; Batista, M.V.; Ribeiro, C.; Calle, C.; Cortes, A.; Marhold, M.; Negreiros, I.; Borralho, P.; Brito, M.; et al. Human Microbiota and Breast Cancer—Is There Any Relevant Link?—A Literature Review and New Horizons Toward Personalised Medicine. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Cren, P.-Y.; Bertrand, N.; Le Deley, M.-C.; Génin, M.; Mortier, L.; Odou, P.; Penel, N.; Chazard, E. Is the survival of patients treated with ipilimumab affected by antibiotics? An analysis of 1585 patients from the French National hospital discharge summary database (PMSI). OncoImmunology 2020, 9, 1846914. [Google Scholar] [CrossRef]
- Geller, L.T.; Straussman, R. Intratumoral bacteria may elicit chemoresistance by metabolizing anticancer agents. Molecular & Cellular Oncology 2018, 5, e1405139. [Google Scholar] [CrossRef]
- Parhi, L.; Alon-Maimon, T.; Sol, A.; Nejman, D.; Shhadeh, A.; Fainsod-Levi, T.; Yajuk, O.; Isaacson, B.; Abed, J.; Maalouf, N.; et al. Breast cancer colonization by Fusobacterium nucleatum accelerates tumor growth and metastatic progression. Nat. Commun. 2020, 11, 3259. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.-W.; Dong, X.; Pan, P.; Chen, K.-W.; Fan, J.-X.; Cheng, S.-X.; Zhang, X.-Z. Phage-guided modulation of the gut microbiota of mouse models of colorectal cancer augments their responses to chemotherapy. Nature Biomedical Engineering 2019, 3, 717–728. [Google Scholar] [CrossRef]
- Routy, B.; Lenehan, J.G.; Miller, W.H.; Jamal, R.; Messaoudene, M.; Daisley, B.A.; Hes, C.; Al, K.F.; Martinez-Gili, L.; Punčochář, M.; et al. Fecal microbiota transplantation plus anti-PD-1 immunotherapy in advanced melanoma: a phase I trial. Nat. Med. 2023, 29, 2121–2132. [Google Scholar] [CrossRef]
- Mohseni, A.H.; Taghinezhad-S, S.; Casolaro, V.; Lv, Z.; Li, D. Potential links between the microbiota and T cell immunity determine the tumor cell fate. Cell Death & Disease 2023, 14, 154. [Google Scholar] [CrossRef]
- Galván-Peña, S.; Zhu, Y.; Hanna, B.S.; Mathis, D.; Benoist, C. A dynamic atlas of immunocyte migration from the gut. Science Immunology 2024, 9, eadi0672. [Google Scholar] [CrossRef] [PubMed]
- Rescigno, M.; Urbano, M.; Valzasina, B.; Francolini, M.; Rotta, G.; Bonasio, R.; Granucci, F.; Kraehenbuhl, J.-P.; Ricciardi-Castagnoli, P. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat. Immunol. 2001, 2, 361–367. [Google Scholar] [CrossRef]
- Nejman, D.; Livyatan, I.; Fuks, G.; Gavert, N.; Zwang, Y.; Geller, L.T.; Rotter-Maskowitz, A.; Weiser, R.; Mallel, G.; Gigi, E.; et al. The human tumor microbiome is composed of tumor type–specific intracellular bacteria. Science 2020, 368, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut microbiome modulates response to anti–PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef]
- Schorr, L.; Mathies, M.; Elinav, E.; Puschhof, J. Intracellular bacteria in cancer—prospects and debates. npj Biofilms and Microbiomes 2023, 9, 76. [Google Scholar] [CrossRef]
- Fu, A.; Yao, B.; Dong, T.; Chen, Y.; Yao, J.; Liu, Y.; Li, H.; Bai, H.; Liu, X.; Zhang, Y.; et al. Tumor-resident intracellular microbiota promotes metastatic colonization in breast cancer. Cell 2022, 185, 1356–1372. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Hu, Y. Microbiome harbored within tumors: a new chance to revisit our understanding of cancer pathogenesis and treatment. Signal Transduction and Targeted Therapy 2020, 5, 136. [Google Scholar] [CrossRef]
- Donnet-Hughes, A.; Perez, P.F.; Doré, J.; Leclerc, M.; Levenez, F.; Benyacoub, J.; Serrant, P.; Segura-Roggero, I.; Schiffrin, E.J. Potential role of the intestinal microbiota of the mother in neonatal immune education. Proceedings of the Nutrition Society 2010, 69, 407–415. [Google Scholar] [CrossRef]
- Urbaniak, C.; Gloor, G.B.; Brackstone, M.; Scott, L.; Tangney, M.; Reid, G. The Microbiota of Breast Tissue and Its Association with Breast Cancer. Appl. Environ. Microbiol. 2016, 82, 5039–5048. [Google Scholar] [CrossRef]
- Hoskinson, C.; Jiang, R.Y.; Stiemsma, L.T. Elucidating the roles of the mammary and gut microbiomes in breast cancer development. Frontiers in Oncology 2023, 13. [Google Scholar] [CrossRef]
- Urbaniak, C.; Cummins, J.; Brackstone, M.; Macklaim, J.M.; Gloor, G.B.; Baban, C.K.; Scott, L.; O'Hanlon, D.M.; Burton, J.P.; Francis, K.P.; et al. Microbiota of Human Breast Tissue. Appl. Environ. Microbiol. 2014, 80, 3007–3014. [Google Scholar] [CrossRef]
- Hunt, K.M.; Foster, J.A.; Forney, L.J.; Schütte, U.M.E.; Beck, D.L.; Abdo, Z.; Fox, L.K.; Williams, J.E.; McGuire, M.K.; McGuire, M.A. Characterization of the Diversity and Temporal Stability of Bacterial Communities in Human Milk. PLOS ONE 2011, 6, e21313. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.M.; Fernández, L.; Verhasselt, V. The Gut‒Breast Axis: Programming Health for Life. Nutrients 2021, 13, 606. [Google Scholar] [CrossRef]
- Jost, T.; Lacroix, C.; Braegger, C.P.; Rochat, F.; Chassard, C. Vertical mother–neonate transfer of maternal gut bacteria via breastfeeding. Environ. Microbiol. 2014, 16, 2891–2904. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Tian, T.; Wei, Z.; Shih, N.; Feldman, M.D.; Peck, K.N.; DeMichele, A.M.; Alwine, J.C.; Robertson, E.S. Distinct Microbial Signatures Associated With Different Breast Cancer Types. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Xuan, C.; Shamonki, J.M.; Chung, A.; DiNome, M.L.; Chung, M.; Sieling, P.A.; Lee, D.J. Microbial Dysbiosis Is Associated with Human Breast Cancer. PLoS ONE 2014, 9, e83744. [Google Scholar] [CrossRef]
- Thu, M.S.; Chotirosniramit, K.; Nopsopon, T.; Hirankarn, N.; Pongpirul, K. Human gut, breast, and oral microbiome in breast cancer: A systematic review and meta-analysis. Frontiers in Oncology 2023, 13. [Google Scholar] [CrossRef]
- Samkari, A.A.; Alsulami, M.; Bataweel, L.; Altaifi, R.; Altaifi, A.; Saleem, A.M.; Farsi, A.H.; Iskanderani, O.; Akeel, N.Y.; Malibary, N.H.; et al. Body Microbiota and Its Relationship With Benign and Malignant Breast Tumors: A Systematic Review. Cureus 2022, 14, e25473. [Google Scholar] [CrossRef]
- Bodmer, T.; Miltner, E.; Bermudez, L.E. Mycobacterium avium resists exposure to the acidic conditions of the stomach. FEMS Microbiol. Lett. 2000, 182, 45–49. [Google Scholar] [CrossRef]
- Roth, R.I.; Owen, R.L.; Keren, D.F.; Volberding, P.A. Intestinal infection withMycobacterium avium in acquired immune deficiency syndrome (AIDS). Dig. Dis. Sci. 1985, 30, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Dam, T.; Danelishvili, L.; Wu, M.; Bermudez, L.E. The fadD2 Gene Is Required for Efficient Mycobacterium avium Invasion of Mucosal Epithelial Cells. J. Infect. Dis. 2006, 193, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, R.M.; Caballero, J.D.; van der Schalk, T.E.; De Winter, F.H.R.; Shaw, L.P.; Kapel, N.; Recanatini, C.; Timbermont, L.; Kluytmans, J.; Esser, M.; et al. Gut to lung translocation and antibiotic mediated selection shape the dynamics of Pseudomonas aeruginosa in an ICU patient. Nat. Commun. 2022, 13, 6523. [Google Scholar] [CrossRef]
- Roy, D.; Ehtesham, N.Z.; Hasnain, S.E. Is Mycobacterium tuberculosis carcinogenic to humans? The FASEB Journal 2021, 35, e21853. [Google Scholar] [CrossRef] [PubMed]
- Fol, M.; Koziński, P.; Kulesza, J.; Białecki, P.; Druszczyńska, M. Dual Nature of Relationship between Mycobacteria and Cancer. Int J Mol Sci 2021, 22, 8332. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.A.; Sheikh, J.A.; Ehtesham, N.Z.; Hira, S.; Hasnain, S.E. Can Mycobacterium tuberculosis infection lead to cancer? Call for a paradigm shift in understanding TB and cancer. Int. J. Med. Microbiol. 2022, 312, 151558. [Google Scholar] [CrossRef] [PubMed]
- Conic, J.; Lapinel, N.; Ali, J.; Boulmay, B. Association between non-tuberculous mycobacterial infection and aerodigestive cancers: A case series highlighting different features, sequence and association. Respir. Med. Case Rep. 2022, 40, 101751. [Google Scholar] [CrossRef]
- Hong, S.J.; Kim, T.J.; Lee, J.-H.; Park, J.-S. Nontuberculous mycobacterial pulmonary disease mimicking lung cancer: Clinicoradiologic features and diagnostic implications. Medicine 2016, 95, e3978. [Google Scholar] [CrossRef] [PubMed]
- Taira, N.; Kawasaki, H.; Takahara, S.; Chibana, K.; Atsumi, E.; Kawabata, T. The Presence of Coexisting Lung Cancer and Non-Tuberculous Mycobacterium in a Solitary Mass. American Journal of Case Reports 2018, 19, 748–751. [Google Scholar] [CrossRef]
- Thompson, K.J.; Ingle, J.N.; Tang, X.; Chia, N.; Jeraldo, P.R.; Walther-Antonio, M.R.; Kandimalla, K.K.; Johnson, S.; Yao, J.Z.; Harrington, S.C.; et al. A comprehensive analysis of breast cancer microbiota and host gene expression. PLOS ONE 2017, 12, e0188873. [Google Scholar] [CrossRef]
- Niccolai, E.; Baldi, S.; Nannini, G.; Gensini, F.; Papi, L.; Vezzosi, V.; Bianchi, S.; Orzalesi, L.; Ramazzotti, M.; Amedei, A. Breast cancer: the first comparative evaluation of oncobiome composition between males and females. Biology of Sex Differences 2023, 14, 37. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Geng, C.; Sang, M.; Gao, W.; Li, S.; Yang, S.; Li, Z. Effect of gastrointestinal microbiome and its diversity on the expression of tumor-infiltrating lymphocytes in breast cancer. Oncol Lett 2019, 17, 5050–5056. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Fang, L.T.; Tang, A.Z.; Chen, H.L.; Xu, M.L.; Wei, X.S.; Pang, G.D.; Li, C.Q. Mycobacterium vaccae alleviates allergic airway inflammation and airway hyper-responsiveness in asthmatic mice by altering intestinal microbiota. Immunology 2024. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Hypothetical pathways of microbiota from drinking water into the gut-breast axis. Following ingestion, drinking water microbiota including NTM, all selected by artificial disinfection and the plumbing system harsh conditions, may cross the acidic gastric environment, reach and eventually traverse the intestinal epithelium, and enter immune cells, aiding their dissemination via the bloodstream or lymphatic system. These pathways offer potential routes for drinking water microbiota to reach breast tissue, where they may integrate the intratumoral microbiota. (Created with BioRender.com).
Figure 1.
Hypothetical pathways of microbiota from drinking water into the gut-breast axis. Following ingestion, drinking water microbiota including NTM, all selected by artificial disinfection and the plumbing system harsh conditions, may cross the acidic gastric environment, reach and eventually traverse the intestinal epithelium, and enter immune cells, aiding their dissemination via the bloodstream or lymphatic system. These pathways offer potential routes for drinking water microbiota to reach breast tissue, where they may integrate the intratumoral microbiota. (Created with BioRender.com).
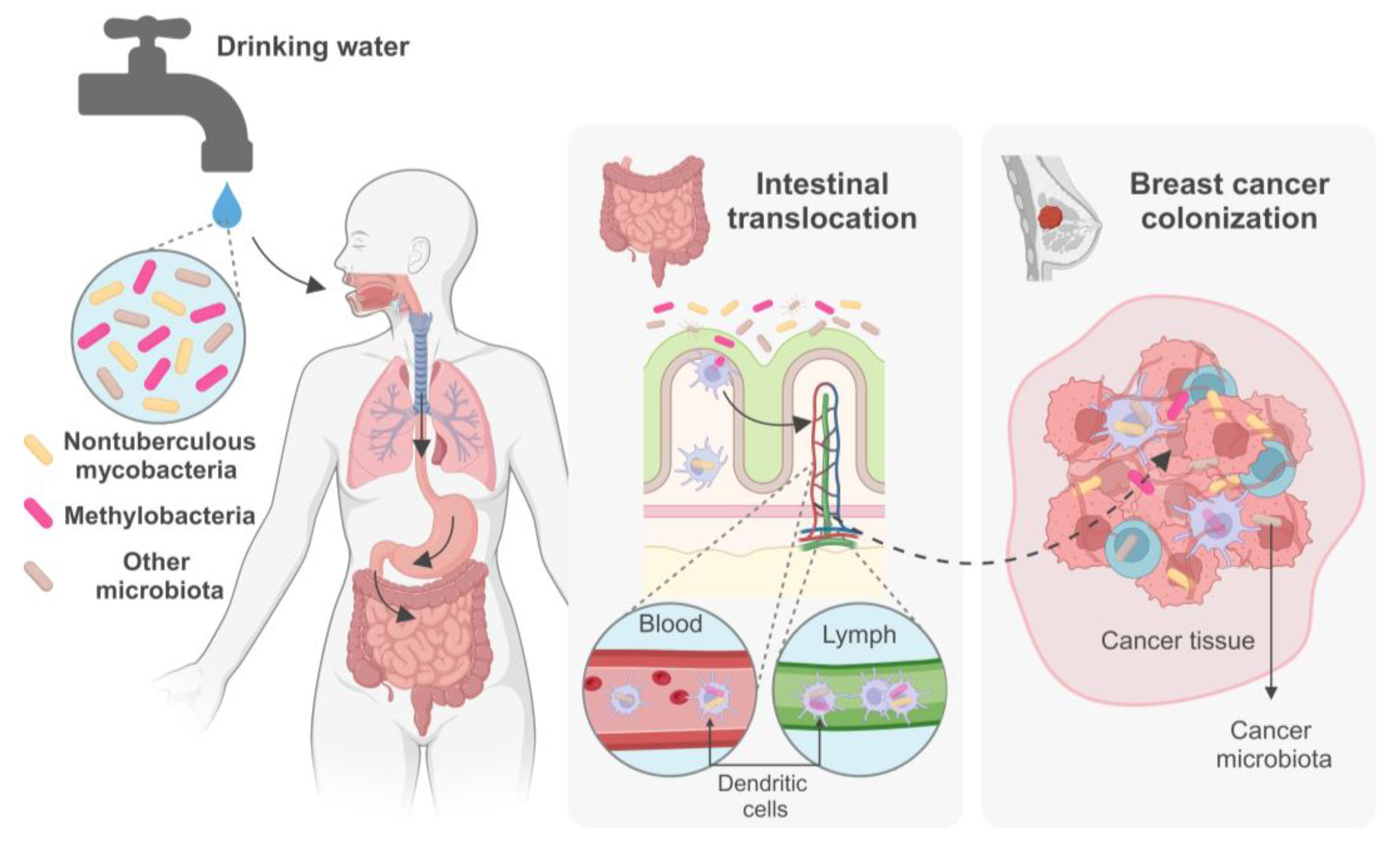
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



