
Submitted:
19 June 2024
Posted:
20 June 2024
You are already at the latest version
Abstract
Clostridioides difficile is a Gram-positive pathogen known for its toxin production and spore formation. It is primarily responsible for most cases of antibiotic-associated diarrhea. Bacterial persisters are a small subset of the population that exhibits transient tolerance to bactericidal substances, and they are of significant medical concern due to their association with the emergence of antibiotic resistance and difficult-to-treat chronic or recurrent infections. Vancomycin, the predominant antibiotic utilized in the management of C. difficile infection, is extensively applied in the realm of clinical practice. Previous studies have demonstrated a persister-like phenotype with treatments involving this antibiotic. However, the mechanism in C. difficile remains largely unknown, primarily due to the challenge of isolating this small population at any given time. To better characterize C. difficile persister cells, we present a study that enables the enrichment and characterization of persister cells from bacterial cultures in both exponential and stationary phases. Moreover, we could differentiate between triggered (induced by antibiotics, such as vancomycin) and spontaneous (stochastic) persister cells. Additionally, we observed the involvement of toxin-antitoxin systems and Clp proteases in persister cell formation.
Keywords:
Clostridiodes difficile
; Persisster cells
; Antibiotic tolerance
Introduction
Clostridioides difficile, an anaerobic Gram-positive pathogen [1], is notorious for causing a spectrum of gastrointestinal ailments, including diarrhea, toxic megacolon, and pseudomembranous colitis [8]. In healthcare environments, C. difficile infection (CDI) is strongly correlated with antibiotics, which disrupt the normal colonic bacterial population, leading to dysbiosis. This disruption affects the metabolism of bile salts, particularly taurocholate, which is crucial for C. difficile spore germination [2,3,4,5,6,7]. The mortality rate associated with CDI typically hovers around 5%, but during severe outbreaks, it can spike to 20% [5]. Furthermore, CDI recurrence poses a substantial challenge in management, with up to 35% of patients experiencing a recurrence after their initial episode [11], and the likelihood of recurrence escalating with subsequent episodes. Recurrence may involve either the relapse of the same strain or reinfection with a different strain, occurring in approximately 38-56% of cases [12,13]. A key contributor to recurrent disease is C. difficile's capacity to re-sporulate and persist in the gastrointestinal tract, even in the presence of antibiotics [11]. Current guidelines and FDA approval lists offer multiple treatment options (vancomycin, fidaxomicin, metronidazole) and possible adjunctive therapies (anti-toxin antibodies, FMT, among others, [54] but vancomycin and fidaxomicin are the current standard treatments for CDI [55]. This antibiotic, belonging to the class of tricyclic glycopeptides, inhibits cell wall synthesis by binding to the D-alanyl-D-alanine ends of cell wall precursors [14].
Recent investigations suggest that the outcomes of CDI may not solely correlate with virulence characteristics or sporulation levels, indicating the involvement of other biological processes.[8] The bacterium's persistence in biofilms could underlie colonization and relapse, potentially facilitating adherence to colonic epithelial cells [16,17,18,19], in which sporulation and persister cell formation could be different mechanisms for pathogen persistence occurring at different stages of the infectious cycle, serving different purposes. Sporulation is a complex and gradual process that produces a resilient morphotype, the spore, adapted for transmission between hosts or as a long-term reservoir between recurrent infections [2,3,4,5,6,7]. In contrast, persister cells are generated more rapidly, which is particularly suited for antibiotic tolerance during biofilm formation [15,20,21,22,23,24,25,26].
On the other hand, it has been hypothesized that two types of persister cells exist. Triggered persister cells (or Type I) are induced by environmental stimuli, such as antibiotic use, while spontaneous (or Type II) persister cells arise stochastically during bacterial growth; these persister cells accumulate during growth and are typically found in the stationary phase [23,24,25,26]. Since Balaban defined persistence to antibiotics in 2019, triggered persistence has been observed in bacteria as a response to stress signals like starvation, where cells enter a persistent state even after the stress is removed. Various stressors can trigger this persistence, including nutrient limitation and antibiotic exposure. Spontaneous persistence, on the other hand, happens without an obvious trigger during stable growth conditions and remains constant [52]. Despite recent advances in the field, the mechanisms responsible for persister cell formation in C. difficile are still largely unknown. This lack of understanding can be attributed, to a significant extent, to the challenges associated with isolating the exceedingly small fraction of the population that finds itself in this state at any given point in time, as well as the anaerobic characteristics of this pathogen. In this study, we present, to our knowledge, the first investigation that enables the enrichment of C. difficile persister cells and allows for their thorough characterization. We have successfully differentiated between two distinct types of persister cells, namely triggered persister cells induced by vancomycin during both the exponential and stationary phases and spontaneous persister cells induced by stochastic processes. Additionally, we identified RNA metabolic-decreased persister cells in C. difficile. In addition to these findings, we have also observed overexpression of toxin-antitoxin systems and Clp protease genes, which are known to play pivotal roles in forming persister cells in several bacteria [30,31,32,33,34].
Methods
Bacterial Culture
The growth of Clostridioides difficile R20291 [36,37] and Δspo0A strains were performed in BHIS (BD difco), containing 0.2% of taurocholate (Merck), 0.1% of fructose (Merck), and 0.1% glucose (Merck). This growth occurred at 37 °C under anaerobic conditions within the BACTRON EZ anaerobic chamber provided by Shellab, USA, as shown in Figure S1. To obtain cultures in the exponential and stationary phases, an overnight (16 h) culture was utilized. For exponential growth, 1% v/v of this culture was used until OD600 0.5 or OD600 0.9 for stationary culture. To isolate persister cells, vancomycin Sigma-Aldrich (100 μg/mL), ceftriaxone Sigma-Aldrich (50 μg/mL), pefloxacin Sigma-Aldrich (50 μg /mL), or rifampicin Sigma-Aldrich (10 μg/mL) (10X MIC for each antibiotic) were administered separately for 1 h.
Enrichment of Persister Cells
This procedure involves subjecting exponential or stationary cultures to a lysis solution (1 mM NaOH, Merck, and 0.005% SDS, Merck) for 10, 20, 30, 60 min [23] followed by 4 mg/mL of lysozyme (Sigma-Aldrich) for 15 min (0.5X MIC for C. difficile) [52]. Subsequently, aliquots were plated at 2, 10, 15, 30, and 60 min. The identical experimental procedure is also applied to cultures that have undergone antibiotic treatment, as previously described, but with a duration of 2 h. Plated aliquots were then collected at 2, 10, 30, and 60 min after the lysis treatment (see Figure S2).
Staining of Persister Cells
As previously described, under an anaerobic chamber, the culture enriched in persister cells was through two washes using 1× TE buffer [37,40]. To this culture, Propidium Iodide Sigma-Aldrich (40 ng/mL) and Thioflavin-T (Thermoscientific) (5 µg/mL, 0.008X of MIC for C. difficile determined for this work) were added and incubated for 15 min in complete darkness. Thioflavin-T only stained intact bacteria with metabolically active RNA but no lysed bacteria (data not shown). Two additional washes using 1× TE buffer were performed following the incubation period. Subsequently, the washed cell suspension that had been stained was observed using an Olympus™ epifluorescent microscope equipped with FITC (fluorescein isothiocyanate with a center wavelength of 475 nm and FWHM of 35 nm) and TRITC (Tetramethylrhodamine Isothiocyanate with a center wavelength of 542 and FWHM of 20 nm) filters and Qimaging Retiga 6 camera.
Flow Cytometry
Flow cytometry was performed with the BD FACSymphony ™ A1 Cell Analyzer (BD Biosciences). For every sample, unstained cultures containing 1% dimethyl sulfoxide (DMSO, Sigma-Aldrich) were used to determine the autofluorescent signal considering size (FSC), complexity (SSC), and fluorescence. Cell viability was determined with Propidium iodide (PI- Sigma-Aldrich, 0.2 µg/mL), while metabolic activity was measured with Thioflavin-T (Thio-T Thermoscientific, 30 µg/mL). A total number of 100,000 events were analyzed if not specified otherwise. Data were analyzed with FlowJo (version 10.7.2, FlowJo LLC, Ashland, OR, USA) software.
RNA Extraction and Quantitative Real-Time PCR
For gene expression, antibiotic treatment was performed for 15 min in the exponential or stationary phase. The extraction of total RNA and qRT-PCR was carried out using the method previously outlined [41]. Briefly, C. difficile cultures were treated with phenol acid to extract total RNA from the culture. Subsequently, 1 µg of total RNA was converted into cDNA through reverse transcriptase and random primers (Promega). These cDNAs, in turn, were employed to carry out the qRT-PCRs reaction in the following manner: a 10 µL reaction containing 25 ng cDNA, 5 µL of Brilliant II SYBR Green QPCR Master Mix (Agilent Technologies), 0.25 µM of each primer (Table S1), and water. The PCR conditions consisted of 50 °C for 2 min, 95 °C for 10 min, followed by 95 °C for 10 s, 60 °C for 30 s, and 72 °C for 30 s for a total of 40 cycles. Melting curves were performed by increasing the temperature by 1 °C within 60 °C to 95 °C. The data derived from the Real-Time PCR were analyzed, employing 16s rRNA and dnaK levels for normalization, following a previously detailed methodology [41]. The generation of the graphics was accomplished through the utilization of GraphPad Prism 7 software. The experiment was conducted with three biological replicates, each with three technical replicates.
Statistical Analysis
GraphPad software was employed for graphing and analyzing the data using statistical assays such as one-way and two-way ANOVA, accompanied by Tukey, Sidak, and Bonferroni Post-hoc tests82.
Results
Enrichment of Spontaneous Persister Cells of C. difficile through Lysis Treatment
Distinguishing between triggered and spontaneous persisters has been considerably challenging, hindering our ability to assess the role of antibiotics, particularly vancomycin, in generating tolerance through the formation of persister cells [23,24,25,26,27,28,29,36]. In this context, we aim to evaluate a lysis method involving weakening the cell wall of actively dividing cells. Given that persister cells do not divide, we expect that this approach will enable us to reduce the population of non-persister vegetative cells, facilitating subsequent studies with persister cells.
In order to distinguish between triggered and spontaneous persister cells, an enrichment process was conducted utilizing lysis treatment (LT) during both the exponential and stationary growth phases (Figure 1). A Δspo0A mutant strain was used to confirm if the observed phenomena were solely due to vegetative cells, not spores [36]. In the exponential phase, the plateau of persisters was achieved with just 10 min of treatment for both the WT and Δspo0A strains (Figure 1A), resulting in a noticeable delay in cell death for both strains. Subsequently, the strains collected at the end of the period were revived and subjected to LT once again, leading to the observation of typical persister behavior replicated in resurrected bacteria. Similarly, in the stationary phase, a similar pattern was observed with a delay in bacterial death after 10 min of treatment. A sustained decrease of 1 to 1.5 orders of magnitude was observed for each strain (Figure 1B). This indicates that the lysis treatment enables enrichment of the culture with cells exhibiting a persister phenotype free from spores observed during exponential and stationary phases within a 10-minute timeframe, significantly faster than what has been observed in previous studies [36].
The key characteristics of persister cells include reduced metabolism and the ability to halt or decrease cell division [15]. Previous studies have suggested that ribosome inactivation (resulting in reduced RNA metabolism or inhibition of protein synthesis) leads to a dormant state (non-dead).[35] To assess if metabolism was diminished, we evaluated the activity of RNA metabolism (activated ribosomes) and determined the viability of these bacteria. Thioflavin-T (Thio-T) has been demonstrated to serve as an RNA-binding probe, enabling the monitoring of RNA metabolism to distinguish between normal and persister cells [37]. To evaluate the metabolic activity of the bacteria isolated using lysis treatment, Thio-T staining was employed to assess RNA metabolism, while Propidium iodide (PI) staining was utilized to determine cell viability. As depicted in Figure 2A, during the exponential phase, the bacteria isolated via lysis treatment were predominantly viable (no PI staining). Still, they exhibited decreased RNA metabolism, strongly indicating their classification as persister bacteria. Interestingly, upon examination of the bacteria isolated using lysis treatment during the stationary phase, it was observed that a significant portion of the culture displayed reduced RNA metabolism even without lysis treatment (Figure 2B), suggesting the presence of spontaneous persister cells. Notably, many of these cells were stained with PI, indicating a mixture of cells with active RNA metabolism, inactive RNA metabolism, and dead cells. However, upon application of lysis treatment, the culture exhibited inactive RNA metabolism, with no dead cells detected by PI staining, thereby confirming the enrichment of spontaneous persister cells during the stationary phase (Figure 2B). Viability of this cells was confirmed by plating at the end of experiment (Figure S3). One aspect we did not expect to observe was the decrease in the size of the bacilli observed when enriching the culture with spontaneous persister cells, which was evident in both exponential and stationary phases (Figure 2). These findings suggest that the lysis treatment effectively enriches the culture with spontaneous persister cells, characterized by reduced RNA metabolism indicative of ribosome inactivation, while maintaining cell viability.
Principio del formulario
Induction of Triggered Persister Cells of C. difficile by Vancomycin
Antibiotic therapies play a crucial role in the triggering of CDI, involving the use of cephalosporins, fluoroquinolones, and rifampicin [2]. While these antibiotics are commonly prescribed for other conditions, they inadvertently disrupt the microbial balance in many patients, predisposing them to C. difficile infection via spore transmission [3,4,5]. On the other hand, vancomycin, recognized as the first-line for treating C. difficile, effectively alleviates symptoms in most patients, although recurrence of infection may occur in some instances [8,9,10].
Therefore, to assess whether the antibiotics encountered by this pathogen during its infective cycle [2,3,4,5] would induce the formation of persister cells, we have selected antibiotics from the same classes to evaluate their ability to induce the formation of triggered persister cells. The study findings revealed that a one-hour treatment with vancomycin at 10 times the minimum inhibitory concentration (MIC) increased survival rates to lysis treatment by up to 10% compared to cases without this antibiotic (Figure 3B). Additionally, it was determined that induction with vancomycin followed by enrichment with lysis treatment did not affect the MIC against this antibiotic (Figure 3A). Viability of this cells was confirmed by plating at the end of experiment (Figure S3). This suggests a potential persister behavior.
To evaluate the persister behavior triggered by vancomycin induction, we investigated different enrichment times with lysis treatment in both the exponential and stationary phases (Figure 4). During the exponential phase, we observed a typical persister response, where induction followed by isolation halted cell death after 10 min of treatment (Figure 4A). Although the number of cells collected decreased after the lysis treatment in the procedure with revived strains, we still observed persister behavior and a cessation of death (Figure 4B).
This indicates that the lysis treatment allowed us to enrich the culture with vancomycin-induced triggered persister phenotype.
To assess whether these triggered persister cells with persister phenotype exhibited reduced metabolism and were alive [15,17], we examined RNA metabolism and membrane permeability using Thio-T and PI stains of collected cells. During the exponential phase with vancomycin, the cells revealed a mixed intensity, which persisted, albeit in reduced numbers, after the lysis treatment. This suggests that vancomycin treatment modifies the expression of stress-related genes due to the high concentrations of vancomycin, a phenomenon also observed in the stationary phase (Figure 5). Furthermore, we found that this culture enriched in triggered persister cells exhibited a mixture of sizes (long and short bacilli) in the exponential phase (Figure 5A, LT) and only long bacilli in the stationary phase (Figure 5B, LT). These results suggest that the lysis treatments effectively enrich the culture with vancomycin-induced triggered persister cells, which exhibit different sizes of bacilli (see Figure S4). Additionally, they indicate that within vancomycin-induced triggered persister cells, not all exhibit inactivated ribosomes, as evidenced by the mixed intensity of fluorescence.
Flow Cytometry Analysis of Enriched Cultures Containing Persister Cells in the Exponential Growth Phase
One of the greatest challenges thus far has been conducting measurements in the presence of oxygen with C. difficile, given its nature as an obligate anaerobe. We decided to evaluate whether the observations made with LT and Vancomycin could be replicated using flow cytometry, employing the same staining techniques. Notably, a differential behavior between the positive control and the treatments was observed during these experiments (Figure 6). Initially, populations were highly defined during the exponential phase, with more than 40% of bacteria showing reduced metabolism, as indicated by Thio-T staining. This can be observed in the lower left quadrant called DN (Figure 6A). This suggests a shift towards the induction of other populations, such as sporulation or persister cells. Remarkably, vancomycin treatment increased this phenomenon to 60%, a percentage that remained consistent when using LT (Figure 6A). Additionally, it was observed that most of the Thio-T-stained population was stained with PI, strongly suggesting that these cells had damaged walls or were dead, likely due to oxygen and/or the treatments used (Figure 6A). As shown in Figure 6C, this leads us to speculate that the population with the peak on the right of the black line will be dead or damaged cells. Finally, when comparing the double-negative population (persister cells), we identify two populations called P1 and P2, as shown in Figure 6B (upper panel). In contrast, the double-positive population (bacteria with active metabolism and damaged cell walls or dead) showed an extended population without clear differences in size and complexity (Figure 6B, lower panel). This observation is consistent with vancomycin treatment results seen under microscopy, showing both long and short bacilli (Figure 5A). Unfortunately, when replicating this procedure in the stationary growth phase, we could only observe bacteria strongly stained with PI, indicating that 99% of the culture was either dead or had damaged cell walls (data not shown).
Evaluation of Expression of Persister-Related Genes
For numerous years, the implication of specific genes in the formation of persister cells has been demonstrated, suggesting mechanisms associated with their functionality [15]. The Toxin-Antitoxin (TA) systems serve as a prime example of this phenomenon, as it has been proposed that these toxins, when overexpressed, hinder bacterial growth, thus reducing cell division and metabolism in the bacteria under investigation [30,31,32,33,34]. One mechanism for activating these toxins involves the degradation of antitoxins, facilitated by specific proteases belonging to the Lon and Clp families [38,39].
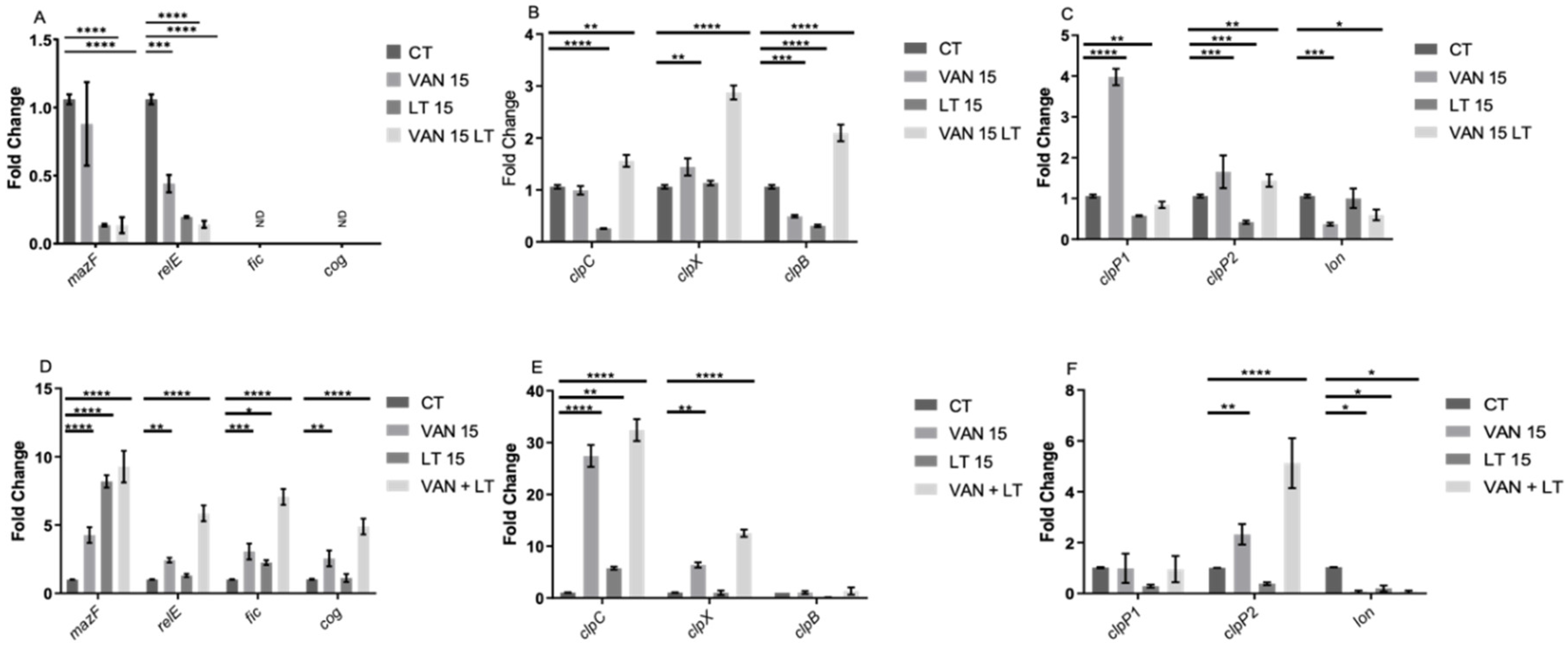
Based on the preceding information, we wondered if the evaluated treatments would alter the expression of genes associated with persister cell formation in other bacteria. Therefore, we decided to evaluate the expression of genes encoding the toxins MazF, RelE, Cog, and Fic (as described in our previous study) [37], along with the genes encoding the protease Lon, the peptidases ClpP1 and ClpP2, and the chaperones ClpC, ClpX, and ClpB. The cog and fic toxins transcripts were undetectable in the exponential phase, and the expressions of relE and mazF toxins were suppressed with the treatments evaluated (Figure 7A). The expression of chaperone genes (clpC, clpX and clpB) increased when treated with vancomycin and subsequently isolated with lysis treatment, indicating that these genes are expressed in triggered persister cells induced by vancomycin and then enrichment through lysis treatment. It is worth noting that no significant differences were found for clpX with the separate lysis treatments, but there was an increase in expression with vancomycin treatment and with the vancomycin and LT treatment, suggesting a relationship with vancomycin-induced stress; the expression of clpC decreased with lysis treatment alone, suggesting a direct involvement with induction by vancomycin. Conversely, for clpB, its induction decreased with both treatments separately, indicating a complex pattern of regulation for this gene (Figure 7B).
Regarding peptidases, clpP1 exhibited up to a fourfold increase in expression with vancomycin treatment, which decreased with lysis treatment. No significant differences were observed with lysis treatment alone. This suggests that the peptidase clpP1 is related to the vancomycin-induced stress response, but not to the formation of persister cells by this antibiotic.. As for clpP2, an increase in expression was noted with vancomycin treatment and vancomycin-induced persister cells, along with a decrease with lysis treatment alone, implying a role in the formation of persister cells induced by vancomycin. In the case of lon, a decrease in expression was observed with vancomycin treatment and vancomycin-induced persister cells, further suggesting involvement in this phenomenon (Figure 7C). Moreover, when examining the expression of the toxin genes of TA systems in the stationary phase with the treatments mentioned above, different behaviors were observed compared to those in the exponential phase, indicating a regulatory mechanism dependent on the growth phase. The expression of mazF increased during vancomycin and lysis treatment, displaying a tenfold increase in vancomycin-induced persister cells (Figure 7D). For relE, an increase was only observed with vancomycin treatment and vancomycin-induced persister cells (Figure 7D). This suggests that relE is involved in the response to vancomycin-induced stress and in the induction of persister cells by this antibiotic, but not in those enriched solely by lysis treatment. Interestingly, unlike the observations made during the exponential phase, fic and cog were detectable during the stationary phase. Significantly, fic and cog expression levels were elevated in cells induced to persist by vancomycin treatment and in cells treated exclusively with vancomycin (Figure 7D). Regarding the expression of chaperone genes clpC, clpX, and clpB, no significant differences were observed in the expression of clpB. However, clpX showed increased expression in cells treated with vancomycin and vancomycin-induced persister cells. Furthermore, clpC demonstrated a substantial up to 30-fold increase in expression in vancomycin-induced persister cells and bacteria treated solely with vancomycin (Figure 7E). It is worth mentioning that cells subjected to lysis treatment exhibited an increase in expression of up to 5-fold. The lon protease gene consistently maintained reduced expression in all three treatments, and no significant differences were observed in clpP1.
On the other hand, clpP2 displayed an increase in expression of up to 5-fold in vancomycin-induced persister cells, with a 2-fold increase noted in cells treated exclusively with vancomycin (Figure 7F). These results suggest that the toxins MazF and RelE do not directly participate in the induction of spontaneous persister enrichment, and all 3 chaperones, ClpC, ClpX, and ClpB with ClpP2 induces the formation of type I persister phenotype in the exponential phase. Moreover, the four toxins of TA systems, MazF, RelE, Fic, Cog, and chaperones ClpC and ClpX with ClpP2 participate in the induction of vancomycin-induced triggered persister phenotype, and only MazF, Fic, and ClpC induce the formation of spontaneous persisters cells in C. difficile.
Discussion
Persister cells typically accumulate during the exponential growth phase, reaching their peak expression in the stationary phase, which aligns with the characterization of spontaneous persister cells [15]. This observation was supported by research from Yang in 2016 and Cañas-Duarte in 2014, indicating that such cells arise stochastically during bacterial growth [23].
This study demonstrated the presence of both triggered persister cells induced by vancomycin and spontaneous in C. difficile. It is worth noting that the presence of taurocholate, acting as a spore germinant, initiated the germination process in the spores within the culture, leading to a decrease in the total number of spores in the medium, consistent with findings from previous research by other investigators [42]. Additionally, rapidly metabolized sugars, such as glucose and fructose, have been found to regulate approximately 18% of the C. difficile transcriptome, with half of these genes regulated by CcpA. This protein forms part of a regulon responsible for controlling sugar uptake and gene regulation [43]. Earlier studies (Antunes, 2012) have demonstrated that CcpA suppresses the expression of Spo0A and SigF, which are early regulators of sporulation [43]. Consequently, the presence of these sugars inhibits the expression of these genes, thus preventing spore formation during cultivation, which corroborates the absence of spores in all the cultures analyzed. The lysis treatment unveiled a plateau of persister cells, indicating the presence of cells with reduced metabolism and cell division [23]. A comparison with a previous study by Cañas-Duarte in 2014 revealed consistency in identifying the plateau within a similar time interval despite the lack of detail regarding the lysis treatment in that study. Unlike Cañas-Duarte, our approach involved generating a curve with revived cells, thereby resolving potential confusion between persister and resistant cells.
Thioflavin-T and Propidium Iodide stains helped us observe C. difficile cells with inactive RNA metabolism and living cells after lysis treatment. This approach provided an alternative to cell sorting methods used for aerobic bacteria and revealed unique shapes in spontaneous persister cells [23], which appeared as short bacilli. Additionally, when examining the complexity of the cells using flow cytometry, we found that most cells treated with LT had fewer complex bacilli. However, with vancomycin and vancomycin combined with LT, a new population of more complex cells emerged (Figure 6B). We believe these more complex cells are the long and short bacilli seen under the microscope. This finding offers new insights into C. difficile and adds to the growing understanding of the different shapes and electron densities of persister cells [35].
The antibiotic evaluation showed that vancomycin and pefloxacin increased the number of persister cells, with the former selected for further investigation due to its use in treating C. difficile-associated diseases. This finding supports previous studies suggesting the involvement of vancomycin in the persister cells formation [36].
In order to demonstrate the persister phenotype, bacteria were revived, and the protocol was replicated. This phenomenon mirrors observations conducted by other researchers using E. coli as a model, where persister cells were isolated with antibiotics (without lysis treatment) [15], and upon bacterial revival, the phenomenon recurred. Importantly, this consistency in occurrence appears independent of the antibiotic utilized, indicating that methods for isolating persister cells are generally unaffected by cells acquiring antibiotic resistance. Notably, these observations were made during the exponential phase, indicating the predominance of triggered persister cells.
Upon microscopic examination of these cultures, various bacterial morphologies were discernible. In phase contrast, elongated bacillus morphology predominated, alongside a significant presence of bacteria with short bacillus morphology.
Thioflavin-T and Propidium Iodide stains helped us observe C. difficile cells with inactive RNA metabolism and living cells after lysis treatment. This approach provided an alternative to cell sorting methods used for aerobic bacteria and revealed unique shapes in spontaneous persister cells, which appeared as short bacilli.
Examination of bacteria treated solely with vancomycin (VAN) revealed that many exhibited decreased RNA metabolisms but remained predominantly viable, with no apparent cell wall damage (Figure 6A). However, observation of vancomycin treatment followed by lysis treatment revealed a mixture of morphologies, with numerous cells displaying activated RNA metabolism, suggesting a diverse bacterial population in the isolated subculture. These populations were observed through microscopy and flow cytometry, where differences in bacillus length, RNA metabolism, and bacterial complexity were noted (Figure 5). Furthermore, when examining the complexity of the cells using flow cytometry, we found that most cells treated with LT had fewer complex bacilli. However, with vancomycin and vancomycin combined with LT, a new population of more complex cells emerged. We believe these more complex cells are the long and short bacilli seen under the microscope (Figure 5A, LT with Zoom). This finding offers new insights into C. difficile and adds to the growing understanding of the different shapes and electron densities of persister cells. Studies conducted with other models, such as Salmonella enterica, have demonstrated that the persister subculture encompasses bacteria with arrested cell division but activated metabolism, termed viable but nonculturable (VBNC) [21]. While these cells typically do not contribute to culture resurrection, they play roles in responses such as biofilm formation [44]. In the context of C. difficile, research suggests that vancomycin treatments induce biofilm formation in the stationary phase [45], which could elucidate the observed results, as biofilm formation involves alterations in gene expression. Within these cellular subcultures, division ceases, but metabolism persists to facilitate the formation of these structures [44].
In the assessment of gene expression, an unexpected finding emerged: during the exponential phase, the TA system toxin genes relE and mazF were repressed under the applied treatments, while fic and cog were undetectable. Conversely, in the stationary phase, all four toxins were overexpressed. This suggests that the behavior of these TA systems mirrors that observed in other bacteria, where they are activated or overexpressed during stress-induced persister cell formation [30,32,34,46].
In the case of chaperones, during the exponential phase, clpC and clpB were implicated in forming spontaneous persister cells, while all three chaperones played a role in developing triggered persister cells. In the stationary phase, only clpC and clpX were notable for their involvement in forming triggered persisters, with clpC implicated in both types. Regarding peptidases and proteases, clpP2 emerged as the most intriguing candidate, exhibiting altered expression under treatments in both growth phases. This finding is compelling because Clp family chaperones and peptidases are closely linked to the activation of type II TA systems [47,48,51], suggesting their joint participation, which warrants further investigation in subsequent studies. Additionally, it has been demonstrated that changes in chaperone expression influence the targets of Clp protease complexes, regulating, for example, the abundance of misfolded proteins (or total proteins) in the cell, a factor associated with persister cells [18,20].
In summary, this study not only validates the existence of persister cells in C. difficile induced by vancomycin but also sheds light on new perspectives regarding the morphology of these cells, their response to treatments, and their capacity to induce the formation of triggered persister cells. These findings enhance our comprehension of persistence of C. difficile and offer valuable insights for future research and therapeutic approaches.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Funding
This research was funded by Agencia Nacional de Investigación y Desarrollo (FONDECYT Grant 1171397) and DI-02-24/REG to F.G. and funded by ANID – Millennium Science Initiative Program –NCN17_093" to F.G. J.A.F. thanks to FONDECYT 1220584 and I.L.C. thanks to FONDECYT 1221610
References
- Zhu, D., Sorg, J. A. & Sun, X. Clostridioides difficile Biology: Sporulation, Germination, and Corresponding Therapies for C. difficile Infection. Frontiers in cellular and infection microbiology 8, 29. (2018). [CrossRef]
- Poutanen, S. M. & Simor, A. E. Clostridium difficile-associated diarrhea in adults. CMAJ : Canadian Medical Association journal = journal de l'Association medicale canadienne 171, 51-58. (2004). [CrossRef]
- Theriot, C. M., Bowman, A. A. & Young, V. B. Antibiotic-Induced Alterations of the Gut Microbiota Alter Secondary Bile Acid Production and Allow for Clostridium difficile Spore Germination and Outgrowth in the Large Intestine. 1, 10.1128/msphere.00045-00015. (2016). [CrossRef]
- Edwards, A. N. et al. Chemical and Stress Resistances of Clostridium difficile Spores and Vegetative Cells. Front Microbiol 7, 1698. (2016). [CrossRef]
- Ahmed, H. & Joshi, L. T. Clostridioides difficile spores tolerate disinfection with sodium hypochlorite disinfectant and remain viable within surgical scrubs and gown fabrics. 169. (2023). [CrossRef]
- Pike, C. M. & Theriot, C. M. Mechanisms of Colonization Resistance Against Clostridioides difficile. The Journal of infectious diseases 223, S194-s200. (2021). [CrossRef]
- Larsen, I. S. et al. Bacillus velezensis DSM 33864 reduces Clostridioides difficile colonization without disturbing commensal gut microbiota composition. Scientific Reports 13, 14941. (2023). [CrossRef]
- Mullish, B. H. & Williams, H. R. Clostridium difficile infection and antibiotic-associated diarrhoea. Clinical medicine (London, England) 18, 237-241. (2018). [CrossRef]
- Spigaglia, P. Recent advances in the understanding of antibiotic resistance in Clostridium difficile infection. Therapeutic advances in infectious disease 3, 23-42. (2016). [CrossRef]
- Polivkova, S., Krutova, M., Capek, V., Sykorova, B. & Benes, J. Fidaxomicin versus metronidazole, vancomycin and their combination for initial episode, first recurrence and severe Clostridioides difficile infection - An observational cohort study. International journal of infectious diseases : IJID : official publication of the International Society for Infectious Diseases 103, 226-233. (2021). [CrossRef]
- Song, J. H. & Kim, Y. S. Recurrent Clostridium difficile Infection: Risk Factors, Treatment, and Prevention. Gut and liver 13, 16-24. (2019). [CrossRef]
- Cole, S. A. & Stahl, T. J. Persistent and Recurrent Clostridium difficile Colitis. Clinics in colon and rectal surgery 28, 65-69. (2015). [CrossRef]
- Tsigrelis, C. Recurrent Clostridioides difficile infection: Recognition, management, prevention. 87, 347-359. (2020). [CrossRef]
- Stogios, P. J. & Savchenko, A. Molecular mechanisms of vancomycin resistance. Protein science : a publication of the Protein Society 29, 654-669. (2020). [CrossRef]
- Lewis, K. Persister cells. Annu Rev Microbiol 64, 357-372. (2010). [CrossRef]
- Normington, C. et al. Biofilms harbour Clostridioides difficile, serving as a reservoir for recurrent infection. npj Biofilms and Microbiomes 7, 16. (2021). [CrossRef]
- Soavelomandroso, A. P. et al. Biofilm Structures in a Mono-Associated Mouse Model of Clostridium difficile Infection. Front Microbiol 8, 2086. (2017). [CrossRef]
- Queraltó, C. et al. The chaperone ClpC participates in sporulation, motility, biofilm, and toxin production of Clostridioides difficile. Journal of global antimicrobial resistance 33, 328-336. (2023). [CrossRef]
- Ethapa, T. et al. Multiple factors modulate biofilm formation by the anaerobic pathogen Clostridium difficile. J Bacteriol 195, 545-555. (2013). [CrossRef]
- Lewis, K. Persister cells and the riddle of biofilm survival. Biochemistry. Biokhimiia 70, 267-274. (2005). [CrossRef]
- Helaine, S. et al. Internalization of Salmonella by macrophages induces formation of nonreplicating persisters. Science (New York, N.Y.) 343, 204-208. (2014). [CrossRef]
- Stapels, D. A. C. et al. Salmonella persisters undermine host immune defenses during antibiotic treatment. Science (New York, N.Y.) 362, 1156-1160. (2018). [CrossRef]
- Cañas-Duarte, S. J., Restrepo, S. & Pedraza, J. M. Novel Protocol for Persister Cells Isolation. PLOS ONE 9, e88660. (2014). [CrossRef]
- Kussell, E., Kishony, R., Balaban, N. Q. & Leibler, S. Bacterial persistence: a model of survival in changing environments. Genetics 169, 1807-1814. (2005). [CrossRef]
- Willenborg, J., Willms, D., Bertram, R., Goethe, R. & Valentin-Weigand, P. Characterization of multi-drug tolerant persister cells in Streptococcus suis. BMC Microbiology 14, 120. (2014). [CrossRef]
- Levin-Reisman, I. & Balaban, N. Q. Quantitative Measurements of Type I and Type II Persisters Using ScanLag. Methods Mol Biol 1333, 75-81. (2016). [CrossRef]
- Dörr, T., Lewis, K. & Vulić, M. SOS Response Induces Persistence to Fluoroquinolones in Escherichia coli. PLOS Genetics 5, e1000760. (2009). [CrossRef]
- Barrett, T. C., Mok, W. W. K., Murawski, A. M. & Brynildsen, M. P. Enhanced antibiotic resistance development from fluoroquinolone persisters after a single exposure to antibiotic. Nature Communications 10, 1177. (2019). [CrossRef]
- Keren, I., Kaldalu, N., Spoering, A., Wang, Y. & Lewis, K. Persister cells and tolerance to antimicrobials. FEMS Microbiology Letters 230, 13-18. (2004). [CrossRef]
- Tripathi, A., Dewan, P. C., Siddique, S. A. & Varadarajan, R. MazF-induced growth inhibition and persister generation in Escherichia coli. The Journal of biological chemistry 289, 4191-4205. (2014). [CrossRef]
- Zhang, Y. et al. MazF Cleaves Cellular mRNAs Specifically at ACA to Block Protein Synthesis in Escherichia coli. Molecular Cell 12, 913-923. (2003). [CrossRef]
- Tashiro, Y. et al. RelE-mediated dormancy is enhanced at high cell density in Escherichia coli. J Bacteriol 194, 1169-1176. (2012). [CrossRef]
- Zhao, Y., McAnulty, M. J. & Wood, T. K. Toxin YafQ Reduces Escherichia coli Growth at Low Temperatures. PLoS One 11, e0161577. (2016). [CrossRef]
- Hu, Y., Kwan, B. W., Osbourne, D. O., Benedik, M. J. & Wood, T. K. Toxin YafQ increases persister cell formation by reducing indole signalling. 17, 1275-1285. (2015). [CrossRef]
- Wood, T. K., Song, S. & Yamasaki, R. Ribosome dependence of persister cell formation and resuscitation. Journal of Microbiology 57, 213-219. (2019). [CrossRef]
- Álvarez, R. et al. Effect of antibiotic treatment on the formation of non-spore Clostridium difficile persister-like cells. The Journal of antimicrobial chemotherapy 73, 2396-2399. (2018). [CrossRef]
- Álvarez, R. et al. Evaluation of functionality of type II toxin-antitoxin systems of Clostridioides difficile R20291. Microbiological research 239, 126539. (2020). [CrossRef]
- Stapels, D. A. C. et al. <i>Salmonella</i> persisters undermine host immune defenses during antibiotic treatment. 362, 1156-1160. (2018). [CrossRef]
- Chowdhury, N., Kwan, B. W. & Wood, T. K. Persistence Increases in the Absence of the Alarmone Guanosine Tetraphosphate by Reducing Cell Growth. Scientific Reports 6, 20519. (2016). [CrossRef]
- Orman, M. A. & Brynildsen, M. P. Establishment of a method to rapidly assay bacterial persister metabolism. Antimicrobial agents and chemotherapy 57, 4398-4409. (2013). [CrossRef]
- Penaloza, D. et al. The Small RNA RyhB Homologs from Salmonella Typhimurium Restrain the Intracellular Growth and Modulate the SPI-1 Gene Expression within RAW264.7 Macrophages. Microorganisms 9. (2021). [CrossRef]
- Paredes-Sabja, D., Shen, A. & Sorg, J. A. Clostridium difficile spore biology: sporulation, germination, and spore structural proteins. Trends in microbiology 22, 406-416. (2014). [CrossRef]
- Antunes, A. et al. Global transcriptional control by glucose and carbon regulator CcpA in Clostridium difficile. Nucleic acids research 40, 10701-10718. (2012). [CrossRef]
- Li, J. & Zhao, X. Effects of quorum sensing on the biofilm formation and viable but non-culturable state. Food research international (Ottawa, Ont.) 137, 109742. (2020). [CrossRef]
- Dapa, T. & Unnikrishnan, M. Biofilm formation by Clostridium difficile. Gut Microbes 4, 397-402. (2013). [CrossRef]
- Page, R. & Peti, W. Toxin-antitoxin systems in bacterial growth arrest and persistence. Nature chemical biology 12, 208-214. (2016). [CrossRef]
- Mohiuddin, S. G., Massahi, A. & Orman, M. A. lon Deletion Impairs Persister Cell Resuscitation in Escherichia coli. mBio 13, e0218721. (2022). [CrossRef]
- Christensen-Dalsgaard, M., Jørgensen, M. G. & Gerdes, K. Three new RelE-homologous mRNA interferases of Escherichia coli differentially induced by environmental stresses. Molecular microbiology 75, 333-348. (2010). [CrossRef]
- Lee, S. et al. The structure of ClpB: a molecular chaperone that rescues proteins from an aggregated state. Cell 115, 229-240. (2003). [CrossRef]
- Leszczynska, D., Matuszewska, E., Kuczynska-Wisnik, D., Furmanek-Blaszk, B. & Laskowska, E. The Formation of Persister Cells in Stationary-Phase Cultures of Escherichia Coli Is Associated with the Aggregation of Endogenous Proteins. PLOS ONE 8, e54737. (2013). [CrossRef]
- Springer, M. T., Singh, V. K., Cheung, A. L., Donegan, N. P. & Chamberlain, N. R. Effect of clpP and clpC deletion on persister cell number in Staphylococcus aureus. Journal of medical microbiology 65, 848-857. (2016). [CrossRef]
- Kaus GM, Snyder LF, Müh U, Flores MJ, Popham DL, Ellermeier CD. Lysozyme Resistance in Clostridioides difficile Is Dependent on Two Peptidoglycan Deacetylases. J Bacteriol. 2020 Oct 22;202(22):e00421-20. PMID: 32868404; PMCID: PMC7585060. [CrossRef]
- Zhang Y. Persisters, persistent infections and the Yin-Yang model. Emerg Microbes Infect. 2014 Jan;3(1):e3. Epub 2014 Jan 8. PMID: 26038493; PMCID: PMC3913823. [CrossRef]
- Gil F, Calderón IL, Fuentes JA, Paredes-Sabja D. Clostridioides (Clostridium) difficile infection: current and alternative therapeutic strategies. Future Microbiol. 2018 Mar;13:469-482. Epub 2018 Feb 21. PMID: 29464969. [CrossRef]
- Stuart Johnson, Valéry Lavergne, Andrew M Skinner, Anne J Gonzales-Luna, Kevin W Garey, Ciaran P Kelly, Mark H Wilcox, Clinical Practice Guideline by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA): 2021 Focused Update Guidelines on Management of Clostridioides difficile Infection in Adults, Clinical Infectious Diseases, Volume 73, Issue 5, 1 September 2021, Pages e1029–e1044. [CrossRef]
Figure 1.
C. difficile persisters cells enriched with lysis treatment. The lysis treatment was conducted, after the culture were reached DO 0.5 with aliquots collected at 2, 10, 30, and 60-minute intervals. Colonies emerging from the 60-min seeding were further cultivated for 16 h, and the experiments were repeated, as depicted in Figure A. Additionally, another aliquot was taken from the overnight culture to growth until reached DO 0.9, to seed the stationary phase, and the lysis treatment was performed, with aliquots sampled at the same time intervals as before. Colonies obtained from the 60-min seeding were also regrown for 16 h, and the experiments were repeated, as shown in Figure B. The presence of persister cells reached a plateau starting from the 15-min point onwards in both cases A and B. Each experimental condition was replicated three times, resulting in three replicates for each condition. Utilizing one-way ANOVA with the Bonferroni post-hoc test, no significant differences were found when adjusting the survival values with each input.
Figure 1.
C. difficile persisters cells enriched with lysis treatment. The lysis treatment was conducted, after the culture were reached DO 0.5 with aliquots collected at 2, 10, 30, and 60-minute intervals. Colonies emerging from the 60-min seeding were further cultivated for 16 h, and the experiments were repeated, as depicted in Figure A. Additionally, another aliquot was taken from the overnight culture to growth until reached DO 0.9, to seed the stationary phase, and the lysis treatment was performed, with aliquots sampled at the same time intervals as before. Colonies obtained from the 60-min seeding were also regrown for 16 h, and the experiments were repeated, as shown in Figure B. The presence of persister cells reached a plateau starting from the 15-min point onwards in both cases A and B. Each experimental condition was replicated three times, resulting in three replicates for each condition. Utilizing one-way ANOVA with the Bonferroni post-hoc test, no significant differences were found when adjusting the survival values with each input.
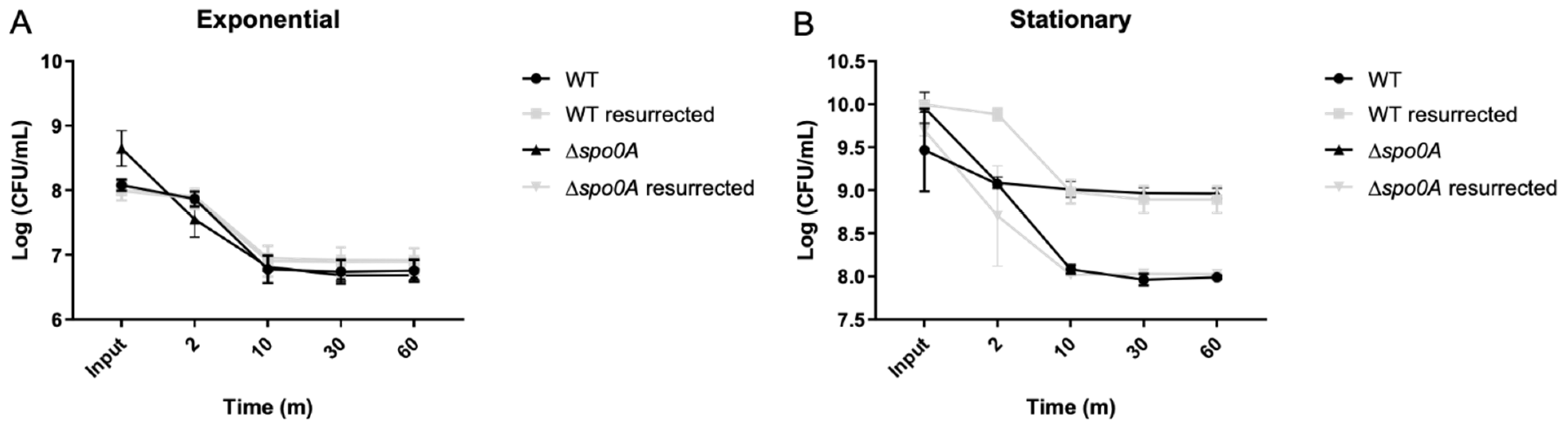
Figure 2.
RNA metabolism of enriched spontaneous persister cells. An aliquot extracted from the culture during the exponential phase (A) was used for the experiment, both untreated (CT, control treatment) and following a 15-min lysis treatment (LT, lysis treatment). The presence of green fluorescence indicated active RNA metabolism, while the absence of color indicated inactive RNA metabolism. Cell wall damage and cell death were evaluated using propidium iodide (PI) staining, shown in red. The same methodology was applied to a culture in the stationary phase (B) with both CT and LT treatments. PC: Phase contrast; Thio-T: Thioflavin-T stain; PI: Propidium iodide stain. Each experimental condition was replicated three times (n=3).
Figure 2.
RNA metabolism of enriched spontaneous persister cells. An aliquot extracted from the culture during the exponential phase (A) was used for the experiment, both untreated (CT, control treatment) and following a 15-min lysis treatment (LT, lysis treatment). The presence of green fluorescence indicated active RNA metabolism, while the absence of color indicated inactive RNA metabolism. Cell wall damage and cell death were evaluated using propidium iodide (PI) staining, shown in red. The same methodology was applied to a culture in the stationary phase (B) with both CT and LT treatments. PC: Phase contrast; Thio-T: Thioflavin-T stain; PI: Propidium iodide stain. Each experimental condition was replicated three times (n=3).
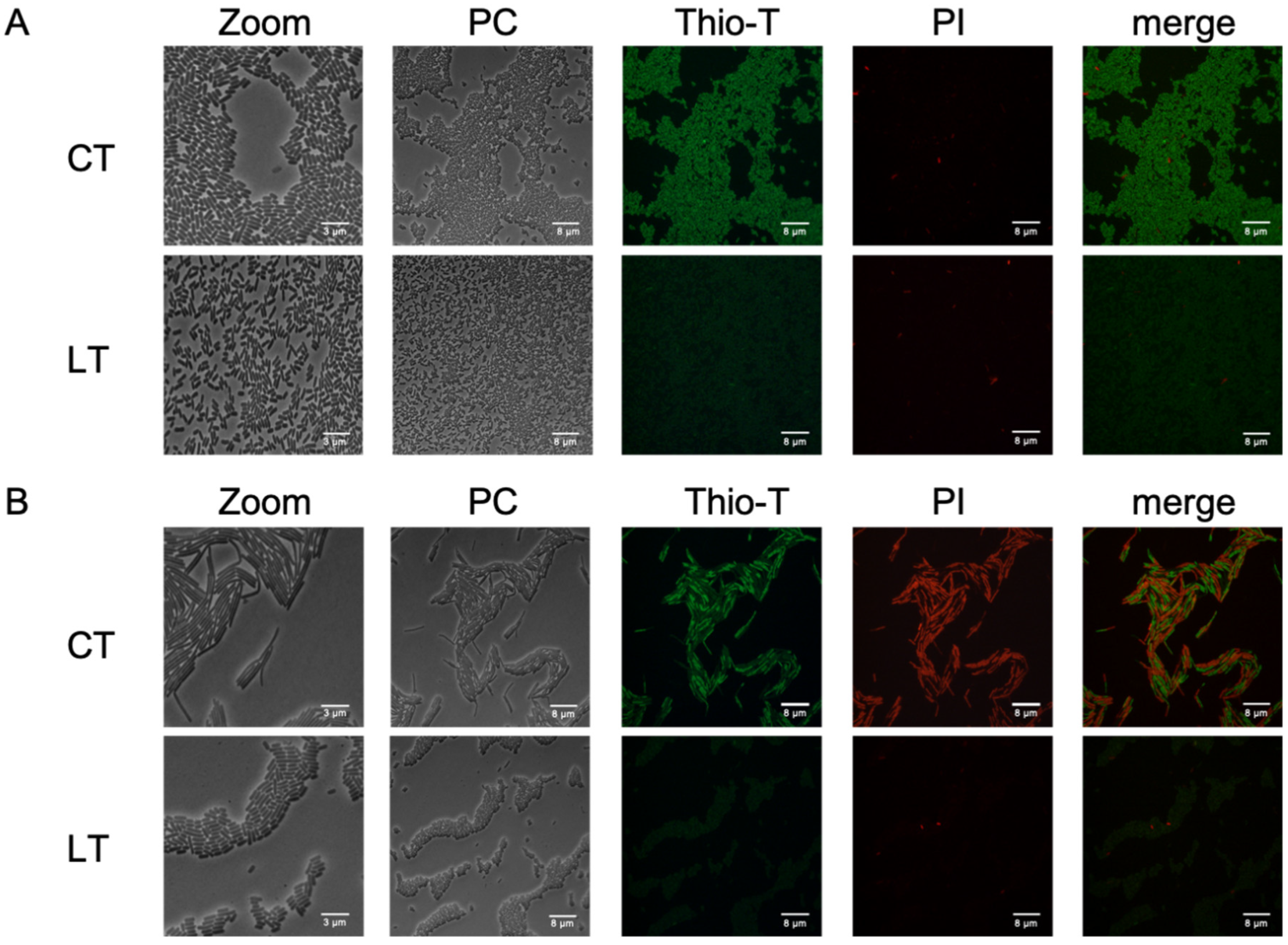
Figure 3.
Antibiotic MICs and persisters cell survival enriched with LT. A survival assay was conducted during the exponential phase, involving a 1-hour antibiotic treatment with vancomycin (VAN), ceftriaxone (CFX), pefloxacin (PEF), and rifampicin (RIF). Each antibiotic was used separately, and after the treatment, an aliquot was plated to mark the input of the experiment. Subsequently, a 15-min lysis treatment was applied. The left table presents the minimum inhibitory concentration (MIC) values before the experiment and the MIC of the resurrected bacteria post-experiment. The right graph illustrates the comparison of cell viability under each treatment during the exponential phase. The experiment was replicated three times (n=3), and statistical analysis was performed using a one-way ANOVA with Post hoc Sidak’s test (* p<0.05; ** p<0.005; *** p<0.0005; **** p<0.0001).
Figure 3.
Antibiotic MICs and persisters cell survival enriched with LT. A survival assay was conducted during the exponential phase, involving a 1-hour antibiotic treatment with vancomycin (VAN), ceftriaxone (CFX), pefloxacin (PEF), and rifampicin (RIF). Each antibiotic was used separately, and after the treatment, an aliquot was plated to mark the input of the experiment. Subsequently, a 15-min lysis treatment was applied. The left table presents the minimum inhibitory concentration (MIC) values before the experiment and the MIC of the resurrected bacteria post-experiment. The right graph illustrates the comparison of cell viability under each treatment during the exponential phase. The experiment was replicated three times (n=3), and statistical analysis was performed using a one-way ANOVA with Post hoc Sidak’s test (* p<0.05; ** p<0.005; *** p<0.0005; **** p<0.0001).
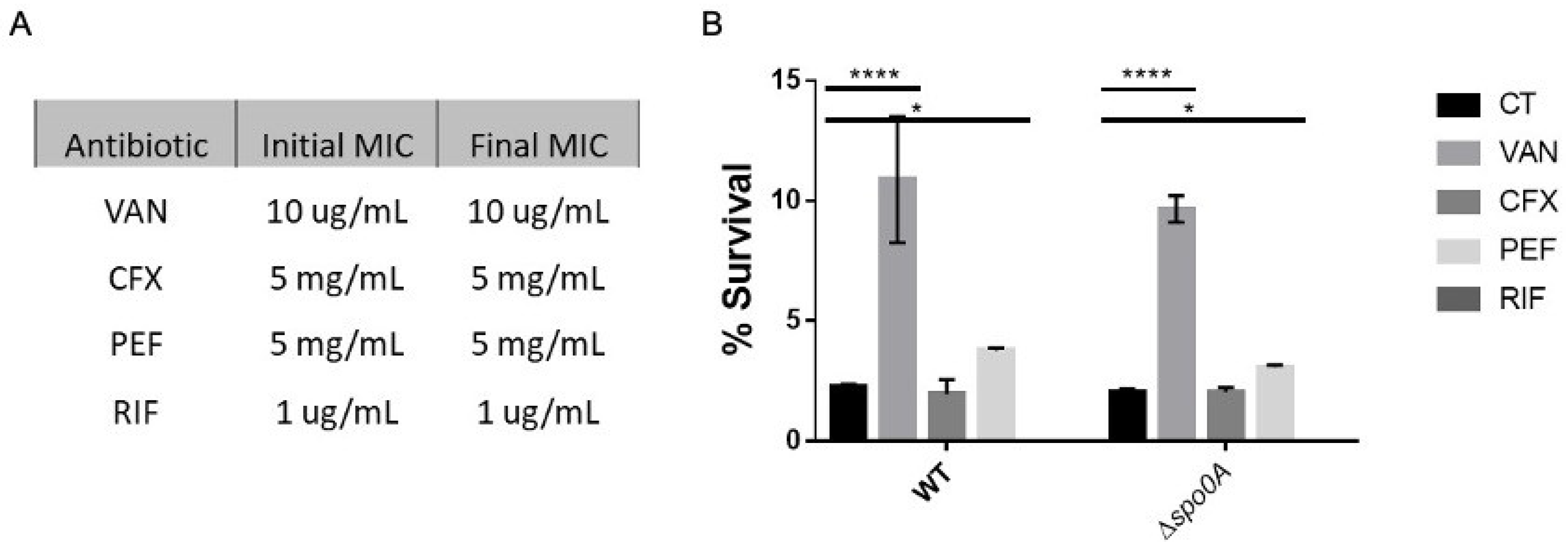
Figure 4.
Vancomycin-induced persisters cells enriched with LT. Cultures of C. difficile were grown overnight (O/N), and an aliquot was transferred to reach the exponential phase (A), following the established procedure. At the experiment’s outset, an aliquot was seeded (input), followed by a 1-hour treatment with vancomycin before seeding again. Subsequently, a 60-min lysis treatment was administered, with samples collected at 2, 10, 30, and 60 min. The same experimental setup was repeated starting from the overnight culture (B), with aliquots seeded for the input, followed by one hour of vancomycin treatment, and then subjected to 2, 10, 30, and 60 min of lysis treatment. The plateau of persister cells was reached after 10 min of treatment. Resurrected bacteria from the 60-min lysis treatment aliquot were retrieved and used to repeat the experiment. Each experimental condition was replicated three times (n=3), with one-way ANOVA analysis revealing no significant differences in survival values among the input.
Figure 4.
Vancomycin-induced persisters cells enriched with LT. Cultures of C. difficile were grown overnight (O/N), and an aliquot was transferred to reach the exponential phase (A), following the established procedure. At the experiment’s outset, an aliquot was seeded (input), followed by a 1-hour treatment with vancomycin before seeding again. Subsequently, a 60-min lysis treatment was administered, with samples collected at 2, 10, 30, and 60 min. The same experimental setup was repeated starting from the overnight culture (B), with aliquots seeded for the input, followed by one hour of vancomycin treatment, and then subjected to 2, 10, 30, and 60 min of lysis treatment. The plateau of persister cells was reached after 10 min of treatment. Resurrected bacteria from the 60-min lysis treatment aliquot were retrieved and used to repeat the experiment. Each experimental condition was replicated three times (n=3), with one-way ANOVA analysis revealing no significant differences in survival values among the input.
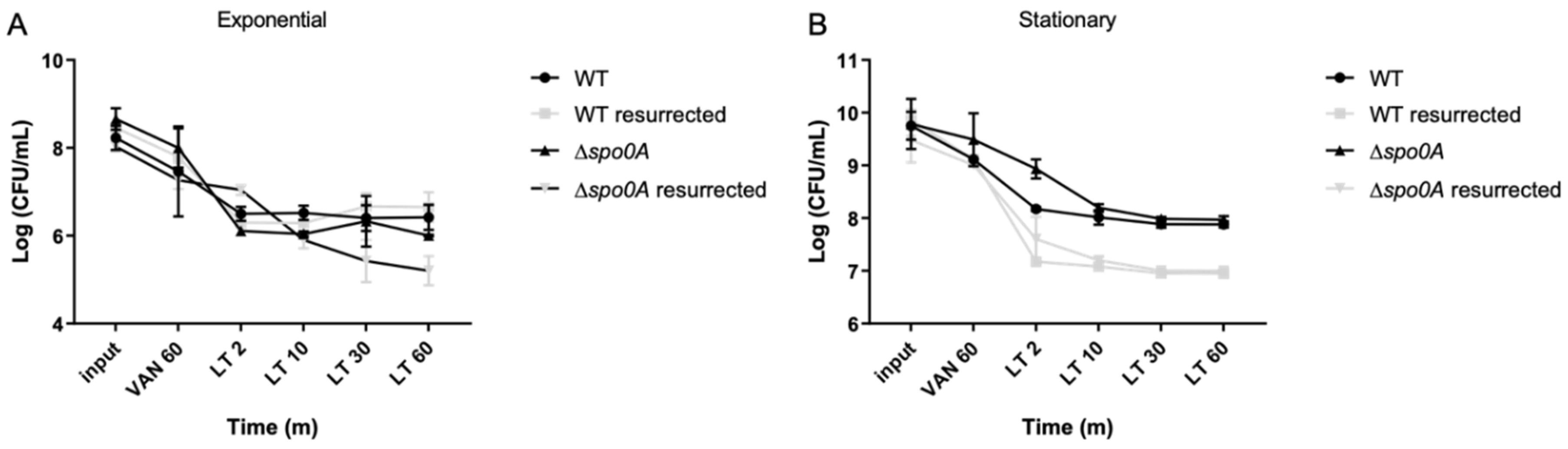
Figure 5.
RNA metabolism of enriched triggered persister cells under vancomycin treatment. Panel (A) illustrates the exponential phase growth culture treated for 1 h with vancomycin, with and without lysis treatment (LT), or control treatment (CT). A similar experiment is depicted in the stationary phase (Panel B). Thioflavin-T (Thio-T) staining was utilized to visualize active RNA metabolism (green color) or inactive metabolism (pale green staining), while propidium iodide (PI) staining was employed to observe cell wall damage or cell death, represented in red. In both experiments, a mixture of bacteria with active and inactive metabolism is observed, with the exponential phase exhibiting more cells with active RNA metabolism. Conversely, in the stationary phase, the CT condition reveals more cells with damaged cell walls or dead cells. PC: Phase contrast; Thio-T: Thioflavin-T stain; PI: Propidium iodide stain. Each experimental condition was replicated three times (n=3).
Figure 5.
RNA metabolism of enriched triggered persister cells under vancomycin treatment. Panel (A) illustrates the exponential phase growth culture treated for 1 h with vancomycin, with and without lysis treatment (LT), or control treatment (CT). A similar experiment is depicted in the stationary phase (Panel B). Thioflavin-T (Thio-T) staining was utilized to visualize active RNA metabolism (green color) or inactive metabolism (pale green staining), while propidium iodide (PI) staining was employed to observe cell wall damage or cell death, represented in red. In both experiments, a mixture of bacteria with active and inactive metabolism is observed, with the exponential phase exhibiting more cells with active RNA metabolism. Conversely, in the stationary phase, the CT condition reveals more cells with damaged cell walls or dead cells. PC: Phase contrast; Thio-T: Thioflavin-T stain; PI: Propidium iodide stain. Each experimental condition was replicated three times (n=3).
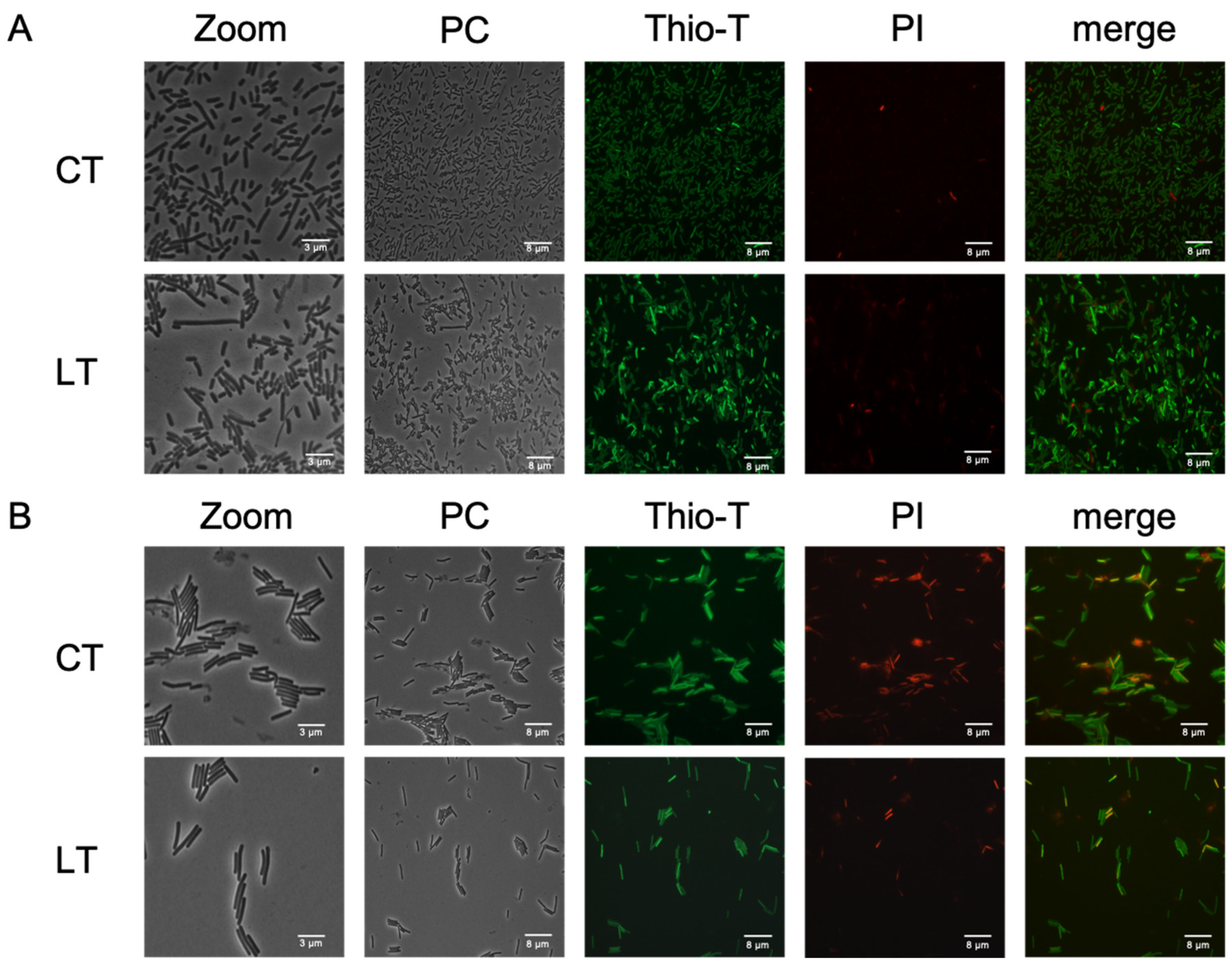
Figure 6.
Characterization of persister C. difficile cells by flow cytometry. (A) Identification of C. difficile stained with Thio-T and PI after application of the different treatments: Lysis treatment (LT), Vancomycin (VAN), Vancomycin + Lysis treatment (VAN + LT), (C+) Positive control of growth. (B) Identification of size and complexity of double negative (DN) (upper) and double positive (DP) (down) populations of C. difficile stained with Thio-T and PI. (C) A representative histogram of total Thio-T expression in C. difficile was previously treated with C+, LT, VAN, and VAN + LT. The black dotted line shows the negative signal for Thio-T to the left and the positive signal for Thio-T to the right of the line.
Figure 6.
Characterization of persister C. difficile cells by flow cytometry. (A) Identification of C. difficile stained with Thio-T and PI after application of the different treatments: Lysis treatment (LT), Vancomycin (VAN), Vancomycin + Lysis treatment (VAN + LT), (C+) Positive control of growth. (B) Identification of size and complexity of double negative (DN) (upper) and double positive (DP) (down) populations of C. difficile stained with Thio-T and PI. (C) A representative histogram of total Thio-T expression in C. difficile was previously treated with C+, LT, VAN, and VAN + LT. The black dotted line shows the negative signal for Thio-T to the left and the positive signal for Thio-T to the right of the line.
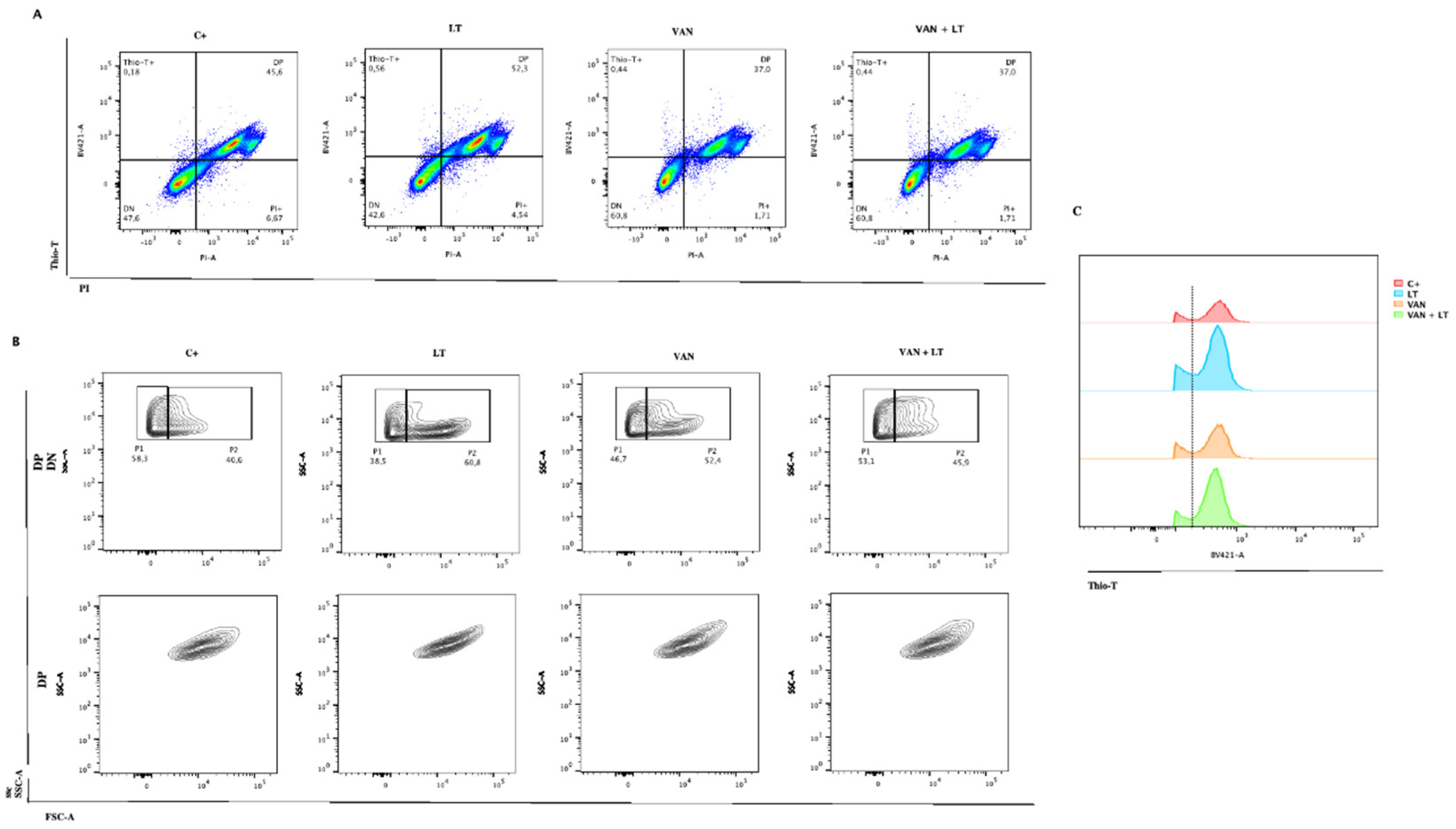
Figure 7.
Gene expression of enriched persister cells under vancomycin treatment. RNA extraction was performed during the exponential growth phase (A, B, and C) and the stationary phase (D, E, and F). In each growth phase, gene expression was evaluated under three conditions: a 15-min treatment with vancomycin (VAN 15), a 15-min lysis treatment (LT 15), and a sequential treatment involving vancomycin followed by a 15-min lysis treatment (VAN 15 LT). Expression of toxin genes from TA systems, namely relE, mazF, fic, and cog (A and D), chaperone genes from the clp family, including clpC, clpX, and clpB (B and E), as well as peptidase and protease genes, specifically clpP1, clpP2, and lon (C and F) were measured. The experiment was conducted with n=3 replicates, and statistical analysis was performed using one-way ANOVA with Post-hoc Bonferroni's test (* p<0.05; ** p<0.005; *** p<0.0005; **** p<0.0001).
Figure 7.
Gene expression of enriched persister cells under vancomycin treatment. RNA extraction was performed during the exponential growth phase (A, B, and C) and the stationary phase (D, E, and F). In each growth phase, gene expression was evaluated under three conditions: a 15-min treatment with vancomycin (VAN 15), a 15-min lysis treatment (LT 15), and a sequential treatment involving vancomycin followed by a 15-min lysis treatment (VAN 15 LT). Expression of toxin genes from TA systems, namely relE, mazF, fic, and cog (A and D), chaperone genes from the clp family, including clpC, clpX, and clpB (B and E), as well as peptidase and protease genes, specifically clpP1, clpP2, and lon (C and F) were measured. The experiment was conducted with n=3 replicates, and statistical analysis was performed using one-way ANOVA with Post-hoc Bonferroni's test (* p<0.05; ** p<0.005; *** p<0.0005; **** p<0.0001).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



