
Submitted:
19 June 2024
Posted:
20 June 2024
You are already at the latest version
Abstract
Obesity is a major risk factor for the development of life-threatening malignant ventricular tachyarrhythmias (VT) and sudden cardiac death (SCD). Risks may be highest for patients with high levels of the proinflammatory cytokine interleukin (IL)-6. We used our guinea pig model of high-fat diet (HFD) induced Ventricular arrhythmias, that exhibit heightened proinflammatory-like pathology which is also observed in human obesity VT, as well as immunofluorescence, and confocal microscopy approaches to evaluate the pathological IL-6 trans-signaling function and explore the underlying mechanisms. Using blind-stick and electrocardiogram (ECG) techniques, we tested the hypothesis that heightened IL-6 trans-signaling exhibit enhanced Ventricular arrhythmias /SCD incidence and underlying arrhythmia substrates. Remarkably, compared to low-fat (LFD) diet fed controls, HFD promotes phosphorylation of the IL-6 signal transducer and activator of transcription 4 (STAT4) leading to its activation and enhanced nuclear translocation of pSTAT4/STAT4 more than LFD controls and pSTAT3/STAT3 nuclear expression. Overactivation of IL-6 trans-signaling in guinea pigs prolonged the QT interval that resulted in greater susceptibility to VT/SCD with isoproterenol challenge, as also observed with the downstream Janus kinase (JAK) 2 activator. These findings may have potentially profound implications for more effective VT therapy in the vulnerable obese patient population.
Keywords:
Signal transducer and activator of transcription 4
; Interleukin-6 trans-signaling
; Guinea pig
; Ventricular arrhythmias
Introduction
Ventricular tachycardia (VT) accounts for the electrophysiological events leading to sudden cardiac death (SCD) in 60-80% of pathologies [1]. At the cellular level, pathological modulation of expression and/or electrophysiological function of major cardiac ion channel subunits, with a subsequent prolongation of the ventricular action potential duration, underlies an elevated risk for prolonged QT interval linked life-threatening VT [2,3]. Obesity increases VT/SCD risk particularly under conditions of repolarization disorder and prolonged QT interval [4]. The pathology of obesity related arrhythmias is associated with abnormal accumulation of lipids (lipotoxicity), that induces an increase in proinflammatory cytokines, including IL-6, elevating the risk for adverse ventricular electrical remodeling [5]. Preventing such lipotoxic effects is a promising direction for therapeutic intervention in the progression of VT, and ultimately prevention of SCD.
IL-6 is a powerful predictor of the severity of heart diseases [6]. Classical IL-6 signaling occurs through its membrane-bound receptor (IL-6Rα)- glycoprotein 130 (gp130 receptor complex), and mediates homeostasis and regenerative functions [7]. The soluble IL-6 receptor (sIL-6R) is generated by extracellular shedding or alternative processing of the mRNA encoding the IL-6R. IL-6 proinflammatory effects are mediated via trans-signaling, whereby IL-6 binds to the sIL-6R, leading to activation of the downstream Janus kinase (JAK)-signal transducer and activator of transcription (STAT) pathway [7]. STATs upon phosphorylation dimerize and translocate to the nucleus, where they can induce the expression of genes involved in proliferation and differentiation. While there is overwhelming evidence supporting the presence of inflammation in heart disease and proinflammatory cytokine channelopathies [5,8], there are still unanswered questions in the arrhythmia field as to whether/how IL-6 trans-signaling remodels ventricular pathology and VT predisposition in obese heart and the molecular mechanisms involved.
Here, we investigated the effect of high-fat diet (HFD) feeding on IL-6 trans-signaling in guinea pig hearts and determined its role in ventricular arrhythmogenesis. Our data show IL-6 activated IL-6 receptor (IL-6R) and the JAK2/STAT4 pathway to be pathologically overexpressed in HFD-lipotoxic guinea pig ventricular hearts. Moreover, hyper-IL-6 (a potent and selective activator of IL-6-trans-signaling[7]) and a JAK2 activator directly induced a severely prolonged QT interval that resulted in increased susceptibility to VT in guinea pigs and that this can be prevented with the IL-6 trans-signaling inhibitor, olampkicept.
Our data reveal a novel inflammatory channelopathy as a risk factor for clinical obesity QT interval prolongation, whereby IL-6 trans-signaling-STAT4-mediated inhibition of the delayed rectifier channels may increase vulnerability to QT prolongation, cause significantly more ventricular arrhythmias, and increased risk for SCD than low fat diet (LFD)-fed or untreated control guinea pigs.
Materials and Methods
Animals, Low-Fat-Diet, High-Fat-Diet (Palmitic-Acid (PA) Diet), and Oleic-Acid Diet (OAD) Feeding
Guinea pigs (male/female; 300–350 g) were obtained from Charles River Laboratories (Wilmington, MA, USA). The control guinea pigs were fed, ad libitum, a LFD (Research Diets Inc., New Brunswick, NJ, USA) containing (in kcal%): 10 fat, 70 carbohydrates, 20 protein, and 2300 corn starch. The HFD/PA-diet group was fed a diet (in which most of the soybean was replaced with 1598% lard or 315 kcal% palm oil) containing 45%/10% of kilocalories from fat, 35%/70% from carbohydrates, and 20% from protein. The monounsaturated OAD group was challenged with a diet in which the soybean was substituted for 198% safflower oil and contained (in kcal%): 45% fat, 35% carbohydrates and 20% protein. The HFD/PA-rich diet contained saturated and unsaturated free fatty acids (FFAs), which provided 31.6%/48.4% and 35.5%/36.8% of the fat-derived calories, while the LFD provided 23.5% and 29.7% and OAD provided 46.2% and 41.4%. Guinea pig groups were challenged with specific diets for a duration of 100 days (~14 weeks).
Electrocardiogram (ECG)
Surface ECG was recorded in slightly anesthetized guinea pigs using a Dual Animal BioAmp amplifier PowerLab (LabChart 8/s, AD instruments, Colorado Springs, CO, USA) and analysis system (LabChart v8.1.2, AD instruments, Colorado Springs, CO, USA). Guinea pigs were placed on a warm pad and subjected to anesthetic inhalation, using a table-top isoflurane (3–5%) vaporizer (Harvard Apparatus, Holliston, MA). A cone mask was used to maintain anesthesia with 1–2% isoflurane (mix of isoflurane and 700 mL O2/minute). Anesthesia depth from isoflurane was monitored by respiratory rate and toe pinch response. Electrodes were positioned on the sole of each guinea pig foot. After a 10 minute stabilization period, 2 boluses of cytokines, and coumermycin were injected at 15-minute intervals. Electrical signals were recorded at 1200 Hz, stored on a computer hard disk, and analyzed off-line using the average of five representative consecutive beats. Tracings were analyzed and calculated for QTc interval by Bazett’s formula where QTc = QT/√RR.
Estimation of Interstitial Fibrosis in Ventricular Tissue Slices form Guinea Pig
Tissue was fixed in 4% paraformaldehyde and then dehydrated with the standard gradient of 70% to 100% ethanol concentration and then cleared with Citrisolv prior to paraffinization. Tissue was sectioned with a HistoCore BIOCUT Microtome (Leica Biosystems, Wetzlar, Germany) and samples were adhered to slides and stained using an automated Masson’s trichrome protocol on the Dako Artisan Link Pro (Agilent, Santa Clara, CA). After initial dehydration of the samples, all sectioning and processing was performed at Associated Regional and University Pathologists Inc. (ARUP, Salt Lake City, UT). Slides were then imaged on an EVOS XL Core microscope (Invitrogen, Carlsbad, CA) with a 20x objective at room temperature (20–25 °C) and quantification of fibrotic area was performed using MATLAB R2023a (RRID:SCR_001622).
Enzyme-Linked Immunosorbent (ELISA) Assay
Guinea pig serum and cardiac tissue cytokine levels were measured by ELISA kits (R&D systems, Minneapolis, MN) according to the manufacturer’s instructions.
Preparation of Bovine Serum Albumin (BSA)-Conjugated FFA Solutions
PA stock solution was prepared as previously described [9]. Briefly, fatty-acid-free bovine serum albumin (BSA, Roche) (20%) was dissolved in Dulbecco Phosphate Buffered Saline (DPBS) and filtered to sterilize. The saturated PA (Sigma-Aldrich, St Louis, MO, USA) was dissolved in ethanol to make a 0.2 M fatty acid (FA) stock solution. BSA (20%) and PA (0.2 M) were mixed in a 20:1 volumetric ratio. FA stock solutions (~10 mM) were directly added to M199 cardiomyocyte culture medium to a final concentration of 0.5 -1 mM.
Guinea Pig Ventricular Myocyte Isolation
Adult male and female Hartley guinea pigs were deeply anesthetized with isoflurane in accordance with the guidelines of the Declaration of Helsinki and approved by the Institutional Review Board (or Ethics Committee) of the University of Utah (Protocol #: 21-09006) Animal Care and Use Committees and conform to NIH guidelines. Primary myocyte isolation procedures have been previously described [8,10,11].
LipidSpot Lipid Droplet Staining
Guinea pig ventricular myocytes were pre-exposed (2 hr.) to either BSA alone or PA-BSA (0.5 mM) and were subsequently incubated with a solution containing paraformaldehyde (4%) supplemented with glucose (4%) for 10 minutes at room temperature (23–25 °C). Fixed ventricular myocytes were labeled with Lipid Spot 610 (#70069; Biotium, Fremont, California) to stain aggregated lipid droplets and was excited with a 561 nm laser and emission collected at 592-638 nm. Immunohistochemistry was performed using a 1:100 dilution of anti-IL6Ra (H-7) mouse mAb (#sc-373708, Santa Cruz Biotechnology, Dallas, Texas, RRID:AB_10947248) and a 1:100 dilution of anti-IL-6 [1] mouse mAb (#sc-130326, Santa Cruz Biotechnology, Dallas, Texas, RRID:AB_2127744). Donkey anti-Mouse IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor 488 (#A-21202, ThermoFisher Scientific, Waltham, MA, RRID:AB_141607) was used and excited with a 488 nm laser and emission was collected at 491–610 nm. Two dimensional images were acquired on a Leica SP8 confocal microscope using a GaAsP-HyD detector and a 40x or 63x oil immersion lens (numerical aperture 1.2) with a 0.1×0.1 μm pixel size at room temperature. All samples were imaged with identical imaging parameters. Sequential framing approaches were used to avoid simultaneous excitation of fluorophores and minimize cross-talk. Images were processed for noise reduction and background correction, then visualized with the same intensity ranges for comparison. Lipid droplet size and count were quantified as described previously described [12] using MATLAB R2023a (RRID:SCR_001622).
Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
Total RNA from guinea pig ventricular tissue was extracted with RNeasy® Plus Mini Kits (74134, Qiagen, Hilden, Germany). The quantity, purity, and integrity of RNA and cDNA samples were determined by spectrophotometry (NanoDrop Lite Plus, Thermo Fisher Scientific, Madison, WI). cDNA synthesis was carried out using the iScript cDNA Synthesis Kit (1708891, BioRad, Hercules, CA). All assays were performed according to the manufacturer’s supplied protocol. qRT-PCR assays were performed using PowerUpTM SYBR™ Green Master Mix (A25742, Applied Biosystems, Vilnius, Lithuania) and a CFX Opus 96 Real-Time PCR System (#12011319; Bio-Rad, Hercules, California). All primer sequences for genes of interest are listed in Table 1. Relative expression values and fold changes were calculated using the ∆∆Ct analysis method relative to GAPDH (endogenous control) and LFD as the control sample, and are presented as mean ± SEM. Primers were designed to span exon–exon regions to avoid amplification of genomic DNA and primer specificity was verified by blasting all sequences using the NCBI Primer-BLAST tool (http://www.ncbi.nlm.nih.gov/tools/primer-blast/ ; RRID:SCR_003095).
Immunofluorescence, Confocal Imaging, and Image Analysis
Guinea pig ventricular tissues were fixed in 4% paraformaldehyde and sectioned using a vibratome (#VT1200, Leica, Deer Park, Illinois) at a 20-μm thickness. Immunohistochemistry was performed using the following primary antibodies: 1:100 dilution of anti-Phospho-STAT3 (Tyr705) (D3A7) XP® rabbit mAb (#9145 Cell Signaling, Danvers, Massachusetts, RRID:AB_2491009); 1:100 dilution of anti-STAT3 (F-2) mouse mAb (#sc-8019, Santa Cruz Biotechnology, Dallas, Texas, RRID:AB_628293); a 1:100 dilution of anti-IL6Ra (H-7) mouse mAb (#sc-373708, Santa Cruz Biotechnology, Dallas, Texas, RRID:AB_10947248); 1:100 dilution of anti-IL-6 [1] mouse mAb (#sc-130326, Santa Cruz Biotechnology, Dallas, Texas, RRID:AB_2127744); 1:200 dilution of anti-hERG1b rabbit pAb (#ALX-215-051-R100, Enzo, Farmingdale, NY, RRID:AB_2051587); 1:200 dilution of anti-hERG1a (NT) rabbit (#ALX-215-050-R100, Enzo, Farmingdale, NY, RRID:AB_2051586); 1:50 dilution of anti-phospho-Stat4 (Tyr693) rabbit pAb (#5267, Cell Signaling, Danvers, Massachusetts, RRID:AB_10545446); 1:50 dilution of anti-Stat4 (C46B10) rabbit pAb (#2653, Cell Signaling, Danvers, Massachusetts, RRID:AB_22551556); 1:50 dilution of anti-IL-18 (D2F3B) rabbit pAb (#54943, Cell Signaling, Danvers, Massachusetts, RRID:AB_2909592); and 1:100 dilution of anti-TGFβ rabbit pAb (#21898-1-AP, ProteinTech, Rosemont, IL, RRID:AB_2811115). Donkey anti-Rabbit IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor™ 647 (#A-31573 ThermoFisher Scientific, Waltham, MA, RRID:AB_2536183) or Donkey anti-Mouse IgG (H+L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor™ 647 (#31571, ThermoFisher Scientific, Waltham, MA, RRID:AB_162542) were used at a dilution of 2μg/mL. Wheat germ agglutinin (WGA) conjugated to Alexa Fluor 488 (#W11261, ThermoFisher Scientific, Waltham, MA) and DAPI (#D1306, ThermoFisher Scientific, Waltham, MA) were used following the instructions of the manufacturer. Two dimensional images were acquired on a Leica SP8 confocal microscope using a GaAsP-HyD detector and a 40x oil immersion lens (numerical aperture 1.2) with a 0.1×0.1μm pixel size at room temperature. DAPI was excited with a 405 nm laser and emission collected at 410–550nm. WGA was excited with a 488 nm laser and emission collected at 491–610nm. Alexa Fluor 647 conjugated antibody was excited with a 633nm laser and emission collected at 638–775nm. All samples were imaged with identical imaging parameters including pinhole size (1 Airy unit). Sequential framing was used to limit simultaneous excitation of fluorophores and minimize cross-talk. Images were processed for noise reduction and background correction, then visualized with the same intensity ranges for comparison. Fluorescence colocalization was quantified in MATLAB R2023a (RRID:SCR_001622).
Statistical Analyses
Data are reported as means ± S.E.M. Statistical differences were determined using two-tailed unpaired t test for comparisons between groups and considered significant at P < 0.05. All experiments were blinded to experimenters and those doing the analyses. Data points were discarded as outliers only if they are greater than three standard deviations from the group mean.
Results
Effect of HFD on Cardiac Fibrosis in Guinea Pig
Male and female guinea pigs were challenged, ad libitum, for 14 weeks with LFD and HFD that we have previously shown to promote significant changes in total cholesterol and total triglycerides [13]. Compared to LFD, HFD feeding increased change (Δ) in body weight (ΔBW) (33 or 402.4±57.6g, n=8; vs. 435.6±34g; n=15, P=0.62). In other experiments, ΔBW was Δ34.6 (or 402.4±57.6g, n=8; vs. 437.05±47.5g; n=8, P=0.65) and Δ (-)1.47 for the OAD challenged group when compared to LFD and the HFD guinea pigs respectively. The HFD group showed a marked increase in % interstitial fibrosis in guinea pig ventricles (Figure 1A,B), and this can be reduced with OAD feeding (Figure 1C) to LFD control levels. After 14 weeks, % fibrosis was increased by 501% in the HFD group but the extent of fibrosis in OAD group was similar to LFD fed controls (Figure 1D), respectively. Our data show a significant increase (from 1% to about 6%) in interstitial fibrosis in HFD hearts when compared to LFD challenged controls. However, while this is a large increase in the amount of interstitial fibrosis it is not severe.
Furthermore, confocal imaging of LFD and HFD ventricular tissue was used to determine whether the transforming growth factor beta (TGF-β) pathway expression, known to be profibrotic, was affected. In 5 separate experiments using HFD guinea pig ventricular tissue, TGF-β protein expression was significantly increased (by 56%, Figure 1E,F) compared to the LFD controls. These data demonstrate the fibrosis-promoting effect of chronic HFD feeding in guinea pigs and that OAD may be associated with blunted % fibrosis in the heart that could lead to beneficial ventricular remodeling and reduced risk for arrhythmias.
HFD feeding is associated with upregulated cardiac IL-6 linked inflammation in guinea pig ventricular tissue.
There have been previous reports of an increased release of the immunomodulatory proinflammatory cytokine IL-6 in obese patients [14,15,16], leading to a state of chronic inflammation [7,17,18]. We also measured cardiac (or local) and systemic IL-6 levels following HFD feeding. Our data revealed HFD-challenged guinea pigs showed a significant increase in serum (Figure 2A) and cardiac tissue (Figure 2B) IL-6 levels. On average, serum, and ventricular tissue IL-6 significantly (*p<0.05) increased, by 29.1% (Figure 2A) and 33.8% (Figure 2B), respectively, in HFD-fed guinea pigs compared to LFD controls. A similar picture emerged with IL-1β (Figure 2C,D, 35.3% vs. 23.7%) and TNF-α (Figure 2E,F, 49.8% vs. 44.7%), and further establishes our HFD guinea pig as an inflammatory model. Moreover, compared to serum levels, HFD induced a larger increase in local cardiac IL-6 but this difference was not seen with IL-1β and TNF-α suggesting that the overactivated IL-6 signaling pathway may elicit a greater inflammatory response and exacerbate cardiac dysfunction.
Overactivated IL-6 Trans-Signaling Causes Pathological Guinea Pig Heart Electrophysiology and Increased Arrhythmic Risk
Obesity increases VT risk [19,20,21,22], particularly under conditions of repolarization disorder and prolonged heart rate corrected QT interval (QTc, an established risk factor for VT) [4,23,24]. We have discovered, in the surface electrocardiogram (Figure 3A), HFD feeding is accompanied by a prolonged QTc interval (Figure 3B, Table 2), and this can be prevented with OAD feeding (Table 2). We have further found that HFD guinea pigs displayed increased risks for spontaneous ventricular arrhythmias (sinus rhythm with abnormal conduction in the ventricular conduction system leading to altered QRS
morphology and T waves suggesting bundle branch block. Figure 3C) compared to LFD-fed controls, consistent with increased risk markers of VT/SCD in HFD linked obesity. Given the established role of heightened IL-6 signaling in obesity, endogenous IL-6R and JAK2 transcripts, which are part of the pro-inflammatory IL-6 pathway, were next assessed in LFD and HFD hearts using qRT-PCR assays. Compared to LFD, our data revealed that HFD guinea pigs had increased expression levels of ventricular IL-6R and JAK2 genes (Figure 3D), substantiating our observation of HFD-linked overactivation of IL-6 signaling (Figure 2).
To test the hypothesis that increased IL-6 levels in obesity directly contribute to an increased risk of VT/SCD and may represent a novel therapeutic target, adult guinea pigs were exposed to activators of IL-6 trans-signaling (recombinant IL-6-sIL-6R) through the cranial vena cava and right atrium (Figure 4A, as we have previously [8] established), and then changes in QTc and susceptibility to ventricular arrhythmias were monitored. Figure 4B, shows typical ECG traces measured in guinea pigs exposed to IL-6-sIL-6R. Compared to vehicle-injected controls, QTc and change in QTc interval (ΔQTc) was significantly greater (Table 3) in IL-6-sIL-6R guinea pigs. To further uncover overactivated IL-6 trans-signaling-linked electrophysiological defects, we did a provocative test with isoproterenol (ISO, 0.5 mg/kg) to reproduce sympathetic stimulation. Shown in Figure 4C,D are examples of ventricular arrhythmias induced by an ISO challenge in IL-6-sIL-6R guinea pigs. On average, IL-6-sIL-6R-ISO challenged guinea pigs show a higher total occurrence of arrhythmias (Figure 4C,D 5/5/100% guinea pigs), and ultimately SCD (2/5/40% guinea pigs) compared to controls (1/4/25%, and 0/4/0% respectively), consistent with the signature high-risk proarrhythmic effect that underlies fatal arrhythmias of ventricular origin in patients. Compared to LFD-fed guinea pig ventricles, HFD severely depressed expression of genes encoding for KCNQ1 (Figure 4E) and KCNE1 (Figure 4F) channel subunits consistent with the ability of IKs to limit action potential duration (APD) during sympathetic stimulation and its potential involvement in VT/SCD risk in obese patients with heightened inflammatory response particularly during exercise [21].
As a complementary approach, guinea pigs were challenged with designer hIL-6 [25,26,27,28] (25ng/ml/kg), a highly potent and selective (independent of classic signaling) activator of IL-6 trans-signaling. At baseline conditions, ECGs were recorded from all sixteen initially unchallenged guinea pigs, which served as their own controls (Figure 5A). On average, hIL-6 (Figure 5B) alone or the JAK2 agonist, coumermycin (C-A1[29], 100μg/kg, Figure 5C) significantly (*p<0.05) prolonged QTc interval and profoundly increased ΔQTc (Table 3, or by Δ34.8±8.15ms and Δ38.4±10.9ms respectively vs. Δ21.66±6.12ms with IL-6-sIL-6R), compared to guinea pigs challenged with the first natural and specific inhibitor of IL-6 trans-signaling, olamkicept (2.5mg/kg [30], Table 3) or to saline control. We also discovered that hIL-6 or coumermycin directly triggered arrhythmias (Figs. 5D,E). For example, In Figure 5D, ventricular complexes III and V are preceded by a P-wave, but with inconsistent PR intervals. Furthermore, ventricular complexes I and IV have a similar shape as III and V, but no visible P-wave, suggesting complexes of ventricular origin. Complexes II and VI display a narrow QRS complex, but without a P-wave consistent with arrhythmias of ventricular origin, likely from the conduction system. Furthermore, in Figure 5E, complex V is preceded by a P-wave but the PR interval is shorter than the four previous beats. In addition, the QRS is wider in complex V reminiscent of a premature ventricular beat (or premature ventricular complex, PVC). Complex VI is preceded by a P wave and the PR interval seems to be similar as complex IV and suggests a sinus beat. Similarly, complexes I and III are similar to VI, which could mean those are also sinus beats. Complex II is different and could be a ventricular premature beat likely originating from the conduction system. Taken together, our data suggest that IL-6 trans-signaling may be mediating inflammation ventricular arrhythmogenesis, in part via IL-6R/JAK2 overactivation.
IL-18 Enhances IL-6 Trans-Signaling Effects on Guinea Pig Ventricular Cardiac Electrophysiology
Despite the advantages of selectively investigating a role for IL-6 trans-signaling in obesity-linked VT, our data (Figure 5) revealed that while overactive IL-6-trans-signaling ΔQTc was statistically significant, it was less remarkable than the effects of HFD, suggesting that other cytokines (IL-1β, TNFα, IL-18, Figure 2 ), likely contribute to the HFD-linked ΔQTc. While the effects of IL-1β and TNF-α on cardiac electrophysiology have been studied [5,31], the role of IL-18 is under-explored. IL-18 has increased expression in obesity [32,33,34], and can induce the production of copious amounts of IL-6, serving as an amplifier of IL-6 effects. We hypothesized IL-18 may heighten IL-6 effects on ventricular electrophysiology and arrhythmia risks. To test this hypothesis, we challenged guinea pigs with a combination of recombinant IL-6-sIL-6R and IL-18 (10μg/kg [35,36,37]). Table 3, shows that IL-18 significantly increased QTc interval due to IL-6-sIL-6R. Furthermore, in the presence of IL-18, IL-6-sIL-6 guinea pigs displayed first degree atrioventricular (AV) block that transitions into complete AV dissociation in 4/6 guinea pigs (Figure 6). Panel B shows 2:1 AV block (with distinct T-wave after the pause). Panels C and D shows Wenckebach AV block (prolong PR interval until AV conduction fails) with a shortened QT interval as well as the shape of the T-wave. Panel E, shows complete AV block (second half of the trace). Our data suggests a potential role for IL-18 in IL-6 trans-signaling-linked pathological ion channel remodeling, action potential phenotypes, and increased propensity for heart block.
Proinflammatory Cytokines Induce Dramatic Ventricular Electrophysiology Remodeling during Adverse Sympathetic Regulation and is Associated with VT in Guinea Pig
During high β-adrenergic activity, IKs becomes larger and displays faster activation kinetics [38]. Thus, IKs becomes critical for ventricular action potential shortening [39] during tachycardia. Therefore, IKs is another key component of “repolarization reserve” in the ventricles [40], suggesting that impaired IKs function would be expected to be proarrhythmic in the setting of chronically elevated key obesity components (sympathetic activation, heightened proinflammatory cytokine levels). In our study, we observed a significant prolongation of QTc and ΔQTc in guinea pigs treated with the chromanol 293B (2mg/kg), an IKs blocker [41], as compared with untreated guinea pigs (Table 3). Moreover, a similar picture emerged in chromanol 293B guinea pigs subsequently exposed to ISO alone (10 min) or with a cytomix (containing IL-6-sIL-6R+IL-1β+TNFα+IL-18, 10-30 min, Table 3). Under these conditions, we further observed a severe induction of ventricular arrhythmias which lasted for ~20 s (Figure 7A,B), suggesting that reductions in IKs may exacerbate ventricular electrical remodeling and contribute to exercise induced VT in obese conditions.
ERG1a Protein Expression is Reduced in HFD Hearts
Ion channel dysfunction remains one of the crucial factors in VT initiation, and in most cases of acquired arrhythmias, IKr remains the dominant channel of clinical cardiotoxicity concern; and the FDA and European Medicines Agency requires screening against the human ether-à-go-go related gene (hERG) for all new drugs being evaluated [42,43]. At the cellular level, decreases in IKr function underlies an elevated risk for VT [8]. Next, we hypothesized ERG1 subunit expression changes in response to HFD feeding may occur at the posttranslational level. To test this hypothesis, we assessed surface ERG1a protein expression using confocal imaging of ventricular slices (Figure 8). We have found ERG1a membrane expression is severely reduced in HFD ventricular slices compared to LFD controls (Figure 8A,B). Thus, we investigated if HFD alters IL-18 and IL-6R protein expression as a readout for a ventricular inflammatory state and found that it did. HFD guinea pig ventricular slices revealed that both IL-18 (Figure 8C) and IL-6R (Figure 8D) protein expression are significantly upregulated more than LFD-fed controls. Furthermore, IL-18 slices displayed a greater fluorescence (215%, Figure 8E) labelling of the sarcolemma and t-tubules in HFD compared to IL-6R (106%, Figure 8F). Together, our data suggests IL-18 effects on IL-6 trans-signaling linked QTc (Table 3) could be the result of severe depression of IKr likely due to enhanced defective ERG channel subunit functional expression at the cardiomyocyte cell-surface.
HFD Feeding is Associated with Increased STAT4 Expression in Guinea Pig Heart
IL-6 proinflammatory effects are mediated via trans-signaling, whereby IL-6 binds to the sIL-6R [44,45], and engages gp130 on target cells leading to activation of downstream JAK-STAT [44,46,47,48]. STATs upon phosphorylation dimerize and translocate to the nucleus, where they can induce the expression of genes involved in proliferation and differentiation [49,50,51,52]. Although there is evidence that STAT4 is involved in cardiovascular diseases [53,54,55], the potential role for STAT4 in obesity VT risk is unknown. We investigated whether STAT4 was functionally active in our HFD guinea pig model. In agreement with overactive IL-6 and JAK2 activity, our data revealed for the first time HFD increased phosphorylation of STAT4, consistent with enhanced inflammation, leading to its activation and enhanced nuclear translocation of pSTAT4/STAT4 compared to LFD control ventricular tissue slices (Figure 9A-C), and pSTAT3/STAT3 nuclear expression (Figure 9D-F), suggesting a minimal role for activated STAT3 in our HFD model. Our results demonstrate that HFD-induced inflammation increases cardiac STAT4 activity, supporting the pathological role of IL-6 trans-signaling in the obese heart.
Lipotoxicity Promotes Overactivation of IL-6 Trans-Signaling in Guinea Pig Ventricular Myocytes via Lipid Droplet Accumulation
Because of the well-established link between HFD, lipotoxicity and cardiac inflammation, we next tested directly the role of lipotoxicity in HFD-linked overactivation of cardiomyocyte IL-6 trans-signaling. To mimic the HFD condition in vitro, we challenged ventricular myocytes with PA (a potent inducer of lipotoxicity [8]) and then assessed the expression of IL-6R. Specifically, we found that abnormal lipid droplet count (Figure 10A-C) and size (Figure 10D), a classic hallmark of cardiac lipotoxicity [56,57,58,59,60], can be achieved by treating guinea pig ventricular myocytes with PA (0.5 mM, 2hr), compared to BSA alone-treated control. IL-6R expression was increased by 110% following lipid droplet accumulation (Figure 10E-G), supporting a role of lipid droplet remodeling in overactive IL-6 trans-signaling-induced cardiac inflammation.
Discussion
Obesity is associated with a heightened proinflammatory cytokine response, elevating the risk for adverse arrhythmias [5]. Preventing such pathological effects is a promising direction for therapeutic intervention in the progression of heart disease, and ultimately prevention of VT/SCD risk. In the present study, we have found that our previously developed HFD-induced lipotoxic guinea model[8,9,61], animals display significant development of interstitial fibrosis, prolonged QT interval and are highly susceptible to spontaneous ventricular arrhythmias, compared to LFD controls, similar to changes observed in obese patients. HFD feeding further revealed over-activation of proinflammatory IL-6 signaling in the ventricles of these guinea pigs and is associated with a novel activation and enhanced nuclear translocation of downstream signal transducer and activator of transcription 4 (STAT4), thus providing a unique opportunity to reveal novel cellular proarrhythmic inflammatory mechanisms of obesity VT. Our data show IL-6/JAK2 elicits a proarrhythmic electrical response highlighting the important role of proinflammatory cytokines in the pathogenesis of obese heart. To our knowledge, our study will be the first to show a potential role for pathological remodeling of IL-6 signaling, possibly by way of overactivated STAT4 on VT/SCD risk, in obese heart. Our findings further suggest suppression of STAT4 activation may protect against ventricular pathology, representing a novel therapeutic approach for VT/SCD predisposition in obesity.
The Canakinumab Anti-Inflammatory Thrombosis Outcomes Study (CANTOS) has shown that canakinumab, a human therapeutic monoclonal antibody targeting interleukin (IL)-1β, significantly reduces major adverse cardiovascular events (MACE) without affecting lipid levels in patients with a history of acute myocardial infarction with elevated C-reactive protein (CRP). However, patients remain at an increased risk for recurring cardiovascular events, particularly among those with the highest levels of IL-6 [62]. Thus, IL-6 signaling could play a fundamental role in cardiovascular prognosis. Intriguingly, in obesity, excess dietary fat in adipose tissue stimulates the release of immunomodulatory cytokines [7,17,18], leading to a state of chronic inflammation in patients. Obese epicardial adipose tissue, a rich local source of IL-6 [63,64,65], may predispose to increased VT/SCD risk [66,67,68], highlighting its utility as an anti-arrhythmic target in a model of obesity VT. Therefore, we focused on IL-6 signaling in this study.
Our study suggests that blockade of IL-6 trans-signaling in concert with other proinflammatory cytokines may provide a more favorable clinical outcome than targeting IL-6 trans-signaling alone. For example, IL-1β or IL-18, both of which requires the NLRP3 inflammasome for activation [69], can induce the production of copious amounts of IL-6, serving as an amplifier of IL-6 effects. While the effects of IL-1β and TNF-α on cardiac electrophysiology have been studied, the role of IL-18 is under-explored. IL-18 has increased expression in obesity and heart failure [32,33,34], contributes to heart rhythm disorders [70], and VT in mice [71], our data establish the novel finding that IL-18 may heighten cytokine effects on proarrhythmic ion channel functional phenotypes and VT risks.
Channelopathies and ion channel dysfunction remain one of the crucial factors in VT initiation. At the cellular level, the normal ventricular cardiac action potential (AP) is defined by Phase 0, due to a large inward sodium current (INa), followed by currents due to voltage-gated L-type calcium (Ca) (ICa,L) and the Na-Ca exchanger (INCX) channels [72]. Repolarization is controlled by the delayed rectifier current (IK), comprised of the rapid (IKr) and slow (IKs) components. The resting membrane potential is controlled by the inward rectifier K current (IK1) [73]. Thus, decreases in outward currents [8,10,74] or increases in depolarizing mechanisms [75] delay repolarization, resulting in prolongation of QT interval, which predisposes to fatal VT.
We have previously [8,76] demonstrated negative modulation by IL-6 of IKr or IKs density in guinea pig ventricular myocytes. Other studies have shown IL-6 [77] increases ICa,L (which increases calcium load in myocytes), and can be associated with VT [78]. TNF-α has been shown to decrease IKr [79], and the transient outward current is also inhibited by IL-1β [80] and IL-18 [71], while IL-1β [81] increases ICa,L, altogether leading to prolongation of the APD. Moreover, we have previously [76] shown that inward sodium current (INa) density measured in guinea pig ventricular myocytes, is not altered in the presence of acute (40-min) exposure to IL-6. However, we observed marked bradycardia followed by a complete atrioventricular dissociation, heart block, and asystole in guinea pigs challenged with in vivo applications of IL-6-sIL-6R-IL-18, suggesting an involvement of a decrease in peak INa. Although other mechanisms cannot be ruled out, our data supports the hypothesis that IL-18, which by impacting SCN5A channel biophysics, may contribute to VT/SCD risk by impacting AP upstroke velocity, conduction velocity, and QT interval, highlighting the importance of multi-ion channel analyses that may inform the rational development of safer (reduced cardiotoxic effects), anti-arrhythmic monotherapy and polytherapy approaches for patients.
Here we have further investigated in HFD ventricles the functional expression of two major repolarizing mechanisms, hERG (IKr ) and KCNQ1-KCNE1 (IKs), robustly expressed in humans and guinea pig, and found severe channel modulation manifested as depression of subunit surface expression (hERG1a) and gene (KCNQ1-KCNE1) regulation. Therefore, the inhibitory effect of HFD feeding and/or overactive IL-6-trans-signaling on hERG1 and KCNQ1-KCNE1 channels may occur, at least in part, both at the protein trafficking and transcriptional level. Notably, our findings demonstrated that HFD shows preferential signaling through STAT4. Accordingly, we show that HFD guinea pigs showed significantly increased pSTAT4/STAT4 expression (62%) compared to LFD-fed controls or pSTAT3/STAT3 (8%). This means that STAT4 may have the potential to significantly regulate the transcription of K channels in obese heart with implications for clinically actionable targets for prevention of VT/SCD risk. This will be a first step in determining the mechanisms of how major cardiac ion channels may be regulated in the presence of key obesity components (lipotoxicity-cytokines-sympathetic overactivity) and how that affects cardiac function and homeostasis.
Tocilizumab (TCZ, which globally blocks IL-6 activities through IL-6R-α and sIL-6R) [82,83], when used in treating rheumatoid arthritis is associated with adverse cardiovascular events [84] and an increase in serum cholesterol [85], which enhances cardiovascular risk [86,87]. Thus, effective therapeutic interventions for prevention of obesity VT/SCD risk would likely require chronic administration, which would render TCZ ineligible for the long-term treatment of chronic metabolic diseases. However, the potential therapeutic benefits and efficacy of selectively targeting IL-6 trans-signaling (olamkicept) for prevention of VT in obesity is unknown. Interestingly, we have found that QT prolongation due to overactivated IL-6 trans-signaling can be prevented with olamkicept, highlighting an emerging and important anti-arrhythmic role for olamkicept in obesity arrhythmias. Notably, olamkicept has shown encouraging results in phase II clinical studies for inflammatory bowel disease [7,88,89,90,91], and thus further provides the rationale to refine olamkicept for the development of the next generation of proinflammatory cytokine inhibitors as an antiarrhythmic in the setting of obesity.
Study Limitations
It is possible with longer dietary intervention, there may be dispersion in weight differences and we are currently investigating this possibility as part of a separate study. However, we have found that the HFD and OAD fed guinea pigs displayed similar change in BW when compared to the LFD controls. We speculated that this may be because the OAD is a variation of the HFD and may still be able to induce weight gain despite supplementation of the lard (1598/198, HFD vs. OAD) with Safflower oil (0/198, HFD vs. OAD). Our focus for this study is not about an increase in body weight but to highlight the importance of differences in dietary fat. OAD-linked inflammatory signaling and its role in arrhythmias is part of an on-going study in our lab. However, as part of this study, we have introduced the ability of OAD feeding to rescue HFD linked fibrosis and QT interval, highlighting the beneficial effects of monounsaturated fatty acids. Although not investigated in this study, Oleic acid administration has been shown to lower levels of proinflammatory cytokines, including IL-6 in a model of HFD challenged Wister albino rats [92].
We have not investigated or measured visceral fat in our model. Epicardial adipose tissue (EAT) is a rich local source of IL-6 signaling in obesity [65,93] and a key contributor to a higher-risk for cardiac events in patients [63,94], and therefore likely contributes to cardiac tissue levels. In this regard, EAT has been shown to increase dramatically in guinea pigs and the amount of EAT is closely correlated to the amount of other types of visceral fat [95]. Therefore, we suspected that EAT is likely to adequately mirror local IL-6 levels defined in pathophysiological conditions, contribute to augmented cellular arrhythmogenesis and increased malignant VT/SCD risk.
Conclusion
To our knowledge, we are not aware of any studies that have mechanistically established the link between IL-6 trans-signaling, ion channel regulation and VT in a HFD guinea pig that show high vulnerability to arrhythmias. Specifically, our data revealed over-activation of IL-6 signaling is associated with a novel activation and enhanced the nuclear translocation of downstream signal transducer and activator of transcription 4 (STAT4). We have further found a larger accumulation of interstitial fibrosis, increased TGFβ expression in HFD hearts and direct stimulation of JAK2 induced arrhythmias. At the cellular level, our data show cardiac lipotoxicity overactivated IL-6 trans-signaling and this occurred by way of lipid droplet accumulation.
Overall, the present study is the first to demonstrate that pathologically overactive IL-6 trans-signaling or JAK2 activation leads to dramatic pathology and are directly associated with VT. IL-6 and IL-18 are emerging as relevant cytokines involved in the inflammatory process with severe ventricular electrophysiological consequences [8,71]. Thus, this represents an important step prior to establishing whether unique or common mechanisms underlie the complex ventricular arrhythmia phenotypes associated with individual or multiple combinations of proinflammatory cytokines. The significance of our study is further underscored by the identification of STAT4 as a likely final effector that may promote impaired IKr/IKs biophysics (channel subunit transcription regulation, surface expression and trafficking mechanisms). This could help establish the principle that STAT4 drives obesity-linked overactive IL-6 trans-signaling leading to IKr/IKs dysfunction, and VT/SCD risk, the concept of which should then be developed with other pro-inflammatory cytokines and ion channels.
Clinical Perspectives
IL-6 is a pleiotropic cytokine (downstream of IL-1β action) [17], and is a powerful predictor of the severity of heart diseases [6]. Crucially, IL-6 trans-signaling inhibition has the potential to enhance the efficacy and even exceed the beneficial effects (anti-arrhythmic) of anti-inflammatory drugs currently in clinical trials. While anti-inflammatory (including non-steroidal anti-inflammatory [96,97], and corticosteroids [98,99]) drugs have shown beneficial anti-arrhythmic properties in experimental atrial fibrillation (AF) [100], all have shown an increased risk of AF in patients, likely because our knowledge of cytokine mechanisms is incomplete. Understanding of the link between IL-6-STAT4 and obesity VT may have broad clinical implications for many cardiac rhythm disorders. Thus, future efforts that interrogate whether therapeutic manipulation of IL-6-STAT4 efficiently prevents proarrhythmic signatures (ion channel function, APD phenotypes, QT prolongation, VT/SCD vulnerability) in diseased HFD/obese ventricular tissues/myocytes, thus counteracting the redundancy of cytokines and undesired side-effects elicited by breaching the essential homeostatic role of classical IL-6 signaling are likely to be rewarded with novel therapeutic perspectives and beneficial translational implications in patients.
Author Contributions
A.C., and K.A.A. performed experiments, analyzed data, undertook manuscript editing, approval and finalized the manuscript. A.S.A. obtained funding, conceived of, and designed experiments, analyzed results, and wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Nora Eccles Harrison Treadwell Foundation and the NIH (R01 HL147044 to A.S.A.).
Institutional Review Board Statement
The adult male and female Hartley guinea pigs were deeply anesthetized with isoflurane in accordance with the guidelines of the Declaration of Helsinki and as approved by the Institutional Review Board (or Ethics Committee) of the University of Utah (Protocol #:2020-01-10-AA01; approved 1/10/2020) Animal Care and Use Committees, conforming to NIH guidelines.
Data Availability Statement
All the relevant data are included within the paper itself.
Acknowledgments
We thank Abbie Saccomanno, and Kamrul Hasan Chowdry for technical assistance. We also thank an anonymous reviewer for providing insightful interpretations of the complex arrhythmia signatures presented in this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Nonstandard Abbreviations and Acronyms | |
|
ERG CVC FFA QTc HFD hERG hIL-6 IL-6 IL-1β JAK2 LFD OAD PA IKr IKs STAT4 sIL-6 TGF-β TNF-α |
Ether-à-go-go related gene Cranial vena cava Free fatty acid Heart rate corrected QT interval High-fat diet Human ether-à-go-go related gene Hyper-IL-6 Interleukin-6 Interleukin-1 beta Janus kinase 2 Low-fat diet Oleic acid diet Palmitic acid Rapidly activating delayed rectifier K current Slowly activating delayed rectifier K current Signal transducer and activator of transcription 4 Soluble interleukin-6 receptor Transforming growth factor beta Tumor necrosis factor alpha |
References
- Marsman RF, Tan HL, and Bezzina CR. Genetics of sudden cardiac death caused by ventricular arrhythmias. Nature Reviews Cardiology 11: 96-111, 2014. [CrossRef] [PubMed]
- Kallergis EM, Goudis CA, Simantirakis EN, Kochiadakis GE, and Vardas PE. Mechanisms, Risk Factors, and Management of Acquired Long QT Syndrome: A Comprehensive Review. The Scientific World Journal 2012: 212178, 2012.
- El-Sherif N, Turitto G, and Boutjdir M. Acquired Long QT Syndrome and Electrophysiology of Torsade de Pointes. Arrhythm Electrophysiol Rev 8: 122-130, 2019. [CrossRef] [PubMed]
- Powell-Wiley TM, Poirier P, Burke LE, Després J-P, Gordon-Larsen P, Lavie CJ, Lear SA, Ndumele CE, Neeland IJ, Sanders P, and St-Onge M-P. Obesity and Cardiovascular Disease: A Scientific Statement From the American Heart Association. Circulation 143: e984-e1010, 2021.
- Ali A, Boutjdir M, and Aromolaran AS. Cardiolipotoxicity, Inflammation, and Arrhythmias: Role for Interleukin-6 Molecular Mechanisms. Front Physiol 9: 1866, 2018.
- Ridker PM, and Rane M. Interleukin-6 Signaling and Anti-Interleukin-6 Therapeutics in Cardiovascular Disease. Circulation Research 128: 1728-1746, 2021. [CrossRef] [PubMed]
- Rose-John S, Jenkins BJ, Garbers C, Moll JM, and Scheller J. Targeting IL-6 trans-signalling: past, present and future prospects. Nat Rev Immunol 1-16, 2023.
- Chowdhury MKH, Martinez-Mateu L, Do J, Aromolaran KA, Saiz J, and Aromolaran AS. Macrophage-Dependent Interleukin-6-Production and Inhibition of I(K) Contributes to Acquired QT Prolongation in Lipotoxic Guinea Pig Heart. Int J Mol Sci 22: 2021.
- Aromolaran AS, Colecraft HM, and Boutjdir M. High-fat diet-dependent modulation of the delayed rectifier K(+) current in adult guinea pig atrial myocytes. Biochem Biophys Res Commun 474: 554-559, 2016. [CrossRef] [PubMed]
- Puckerin A, Aromolaran KA, Chang DD, Zukin RS, Colecraft HM, Boutjdir M, and Aromolaran AS. hERG 1a LQT2 C-terminus truncation mutants display hERG 1b-dependent dominant negative mechanisms. Heart Rhythm 13: 1121-1130, 2016. [CrossRef]
- Martinez-Mateu L, Saiz J, and Aromolaran AS. Differential Modulation of IK and ICa,L Channels in High-Fat Diet-Induced Obese Guinea Pig Atria. Front Physiol 10: 1212, 2019. [CrossRef] [PubMed]
- Al-Kofahi Y, Lassoued W, Lee W, and Roysam B. Improved automatic detection and segmentation of cell nuclei in histopathology images. IEEE Trans Biomed Eng 57: 841-852, 2010. [CrossRef]
- Aromolaran AS, Colecraft HM, and Boutjdir M. High-fat diet-dependent modulation of the delayed rectifier K(+) current in adult guinea pig atrial myocytes. Biochem Biophys Res Commun 474: 554-559, 2016. [CrossRef]
- Roytblat L, Rachinsky M, Fisher A, Greemberg L, Shapira Y, Douvdevani A, and Gelman S. Raised interleukin-6 levels in obese patients. Obes Res 8: 673-675, 2000. [CrossRef]
- Kern L, Mittenbühler MJ, Vesting AJ, Ostermann AL, Wunderlich CM, and Wunderlich FT. Obesity-Induced TNFα and IL-6 Signaling: The Missing Link between Obesity and Inflammation-Driven Liver and Colorectal Cancers. Cancers (Basel) 11: 2018.
- El-Mikkawy DME, El-Sadek MA, El-Badawy MA, and Samaha D. Circulating level of interleukin-6 in relation to body mass indices and lipid profile in Egyptian adults with overweight and obesity. Egyptian Rheumatology and Rehabilitation 47: 7, 2020. [CrossRef]
- Ridker PM, Libby P, MacFadyen JG, Thuren T, Ballantyne C, Fonseca F, Koenig W, Shimokawa H, Everett BM, and Glynn RJ. Modulation of the interleukin-6 signalling pathway and incidence rates of atherosclerotic events and all-cause mortality: analyses from the Canakinumab Anti-Inflammatory Thrombosis Outcomes Study (CANTOS). Eur Heart J 39: 3499-3507, 2018. [CrossRef]
- Battineni G, Sagaro GG, Chintalapudi N, Amenta F, Tomassoni D, and Tayebati SK. Impact of Obesity-Induced Inflammation on Cardiovascular Diseases (CVD). Int J Mol Sci 22: 2021.
- Pietrasik G, Goldenberg I, McNitt S, Moss AJ, and Zareba W. Obesity as a risk factor for sustained ventricular tachyarrhythmias in MADIT II patients. J Cardiovasc Electrophysiol 18: 181-184, 2007. [CrossRef]
- Sabbag A, Goldenberg I, Moss AJ, McNitt S, Glikson M, Biton Y, Jackson L, Polonsky B, Zareba W, and Kutyifa V. Predictors and Risk of Ventricular Tachyarrhythmias or Death in Black and White Cardiac Patients: A MADIT-CRT Trial Substudy. JACC Clin Electrophysiol 2: 448-455, 2016.
- Sabbag A, Sidi Y, Kivity S, Beinart R, Glikson M, Segev S, Goldenberg I, and Maor E. Obesity and exercise-induced ectopic ventricular arrhythmias in apparently healthy middle aged adults. Eur J Prev Cardiol 23: 511-517, 2016. [CrossRef] [PubMed]
- Remme CA. Sudden Cardiac Death in Diabetes and Obesity: Mechanisms and Therapeutic Strategies. Can J Cardiol 38: 418-426, 2022. [CrossRef] [PubMed]
- Mukerji R, Terry BE, Fresen JL, Petruc M, Govindarajan G, and Alpert MA. Relation of left ventricular mass to QTc in normotensive severely obese patients. Obesity (Silver Spring) 20: 1950-1954, 2012. [CrossRef]
- Lazzerini PE, Laghi-Pasini F, Bertolozzi I, Morozzi G, Lorenzini S, Simpatico A, Selvi E, Bacarelli MR, Finizola F, Vanni F, Lazaro D, Aromolaran A, El Sherif N, Boutjdir M, and Capecchi PL. Systemic inflammation as a novel QT-prolonging risk factor in patients with torsades de pointes. Heart 2017.
- Rakemann T, Niehof M, Kubicka S, Fischer M, Manns MP, Rose-John S, and Trautwein C. The designer cytokine hyper-interleukin-6 is a potent activator of STAT3-dependent gene transcription in vivo and in vitro. J Biol Chem 274: 1257-1266, 1999.
- Fischer M, Goldschmitt J, Peschel C, Brakenhoff JP, Kallen KJ, Wollmer A, Grotzinger J, and Rose-John S. I. A bioactive designer cytokine for human hematopoietic progenitor cell expansion. Nat Biotechnol 15: 142-145, 1997.
- Rose-John S. Interleukin-6 signalling in health and disease. F1000Res 9: 2020.
- Gatsiou A, Tual-Chalot S, Napoli M, Ortega-Gomez A, Regen T, Badolia R, Cesarini V, Garcia-Gonzalez C, Chevre R, Ciliberti G, Silvestre-Roig C, Martini M, Hoffmann J, Hamouche R, Visker JR, Diakos N, Wietelmann A, Silvestris DA, Georgiopoulos G, Moshfegh A, Schneider A, Chen W, Guenther S, Backs J, Kwak S, Selzman CH, Stamatelopoulos K, Rose-John S, Trautwein C, Spyridopoulos I, Braun T, Waisman A, Gallo A, Drakos SG, Dimmeler S, Sperandio M, Soehnlein O, and Stellos K. The RNA editor ADAR2 promotes immune cell trafficking by enhancing endothelial responses to interleukin-6 during sterile inflammation. Immunity 56: 979-997 e911, 2023.
- Gong L, Manaenko A, Fan R, Huang L, Enkhjargal B, McBride D, Ding Y, Tang J, Xiao X, and Zhang JH. Osteopontin attenuates inflammation via JAK2/STAT1 pathway in hyperglycemic rats after intracerebral hemorrhage. Neuropharmacology 138: 160-169, 2018.
- Goumas FA, Holmer R, Egberts JH, Gontarewicz A, Heneweer C, Geisen U, Hauser C, Mende MM, Legler K, Röcken C, Becker T, Waetzig GH, Rose-John S, and Kalthoff H. Inhibition of IL-6 signaling significantly reduces primary tumor growth and recurrencies in orthotopic xenograft models of pancreatic cancer. Int J Cancer 137: 1035-1046, 2015.
- Aromolaran AS, and Boutjdir M. Cardiac Ion Channel Regulation in Obesity and the Metabolic Syndrome: Relevance to Long QT Syndrome and Atrial Fibrillation. Frontiers in Physiology 8: 2017.
- Mallat Z, Heymes C, Corbaz A, Logeart D, Alouani S, Cohen-Solal A, Seidler T, Hasenfuss G, Chvatchko Y, Shah AM, and Tedgui A. Evidence for altered interleukin 18 (IL)-18 pathway in human heart failure. Faseb j 18: 1752-1754, 2004. [CrossRef]
- O’Brien LC, Mezzaroma E, Van Tassell BW, Marchetti C, Carbone S, Abbate A, and Toldo S. Interleukin-18 as a therapeutic target in acute myocardial infarction and heart failure. Mol Med 20: 221-229, 2014. [CrossRef]
- Toldo S, Mezzaroma E, O’Brien L, Marchetti C, Seropian IM, Voelkel NF, Van Tassell BW, Dinarello CA, and Abbate A. Interleukin-18 mediates interleukin-1-induced cardiac dysfunction. Am J Physiol Heart Circ Physiol 306: H1025-1031, 2014.
- Enoksson SL, Grasset EK, Hagglof T, Mattsson N, Kaiser Y, Gabrielsson S, McGaha TL, Scheynius A, and Karlsson MC. The inflammatory cytokine IL-18 induces self-reactive innate antibody responses regulated by natural killer T cells. Proc Natl Acad Sci U S A 108: E1399-1407, 2011.
- Doyle SL, Lopez FJ, Celkova L, Brennan K, Mulfaul K, Ozaki E, Kenna PF, Kurali E, Hudson N, Doggett T, Ferguson TA, Humphries P, Adamson P, and Campbell M. IL-18 Immunotherapy for Neovascular AMD: Tolerability and Efficacy in Nonhuman Primates. Invest Ophthalmol Vis Sci 56: 5424-5430, 2015.
- Chiang HY, Lu HH, Sudhakar JN, Chen YW, Shih NS, Weng YT, and Shui JW. IL-22 initiates an IL-18-dependent epithelial response circuit to enforce intestinal host defence. Nat Commun 13: 874, 2022.
- Thompson E, Eldstrom J, Westhoff M, McAfee D, Balse E, and Fedida D. cAMP-dependent regulation of I(Ks) single-channel kinetics. J Gen Physiol 149: 781-798, 2017. [CrossRef]
- Jost N, Virag L, Bitay M, Takacs J, Lengyel C, Biliczki P, Nagy Z, Bogats G, Lathrop DA, Papp JG, and Varro A. Restricting excessive cardiac action potential and QT prolongation: a vital role for IKs in human ventricular muscle. Circulation 112: 1392-1399, 2005. [CrossRef] [PubMed]
- Sarkar AX, and Sobie EA. Quantification of repolarization reserve to understand interpatient variability in the response to proarrhythmic drugs: a computational analysis. Heart Rhythm 8: 1749-1755, 2011.
- Bosch RF, Gaspo R, Busch AE, Lang HJ, Li GR, and Nattel S. Effects of the chromanol 293B, a selective blocker of the slow, component of the delayed rectifier K+ current, on repolarization in human and guinea pig ventricular myocytes. Cardiovasc Res 38: 441-450, 1998. [CrossRef]
- Hancox JC, McPate MJ, El Harchi A, and Zhang YH. The hERG potassium channel and hERG screening for drug-induced torsades de pointes. Pharmacol Ther 119: 118-132, 2008.
- Garrido A, Lepailleur A, Mignani SM, Dallemagne P, and Rochais C. hERG toxicity assessment: Useful guidelines for drug design. Eur J Med Chem 195: 112290, 2020. [CrossRef] [PubMed]
- Taga T, and Kishimoto T. Gp130 and the interleukin-6 family of cytokines. Annual review of immunology 15: 797-819, 1997.
- Fontes JA, Rose NR, and Cihakova D. The varying faces of IL-6: From cardiac protection to cardiac failure. Cytokine 74: 62-68, 2015. [CrossRef] [PubMed]
- Akira S, Isshiki H, Sugita T, Tanabe O, Kinoshita S, Nishio Y, Nakajima T, Hirano T, and Kishimoto T. A nuclear factor for IL-6 expression (NF-IL6) is a member of a C/EBP family. Embo j 9: 1897-1906, 1990.
- Akira S, Nishio Y, Inoue M, Wang XJ, Wei S, Matsusaka T, Yoshida K, Sudo T, Naruto M, and Kishimoto T. Molecular cloning of APRF, a novel IFN-stimulated gene factor 3 p91-related transcription factor involved in the gp130-mediated signaling pathway. Cell 77: 63-71, 1994.
- Naka T, Narazaki M, Hirata M, Matsumoto T, Minamoto S, Aono A, Nishimoto N, Kajita T, Taga T, Yoshizaki K, Akira S, and Kishimoto T. Structure and function of a new STAT-induced STAT inhibitor. Nature 387: 924-929, 1997. [CrossRef] [PubMed]
- Zhong Z, Wen Z, and Darnell JE, Jr. Stat3: a STAT family member activated by tyrosine phosphorylation in response to epidermal growth factor and interleukin-6. Science 264: 95-98, 1994. [CrossRef]
- Darnell JE, Jr. STATs and gene regulation. Science 277: 1630-1635, 1997. [CrossRef]
- Horvath CM, and Darnell JE. The state of the STATs: recent developments in the study of signal transduction to the nucleus. Curr Opin Cell Biol 9: 233-239, 1997.
- Imada K, and Leonard WJ. The Jak-STAT pathway. Mol Immunol 37: 1-11, 2000.
- Koglin J, Glysing-Jensen T, Gadiraju S, and Russell ME. Attenuated Cardiac Allograft Vasculopathy in Mice With Targeted Deletion of the Transcription Factor STAT4. Circulation 101: 1034-1039, 2000. [CrossRef] [PubMed]
- Svenungsson E, Gustafsson J, Leonard D, Sandling J, Gunnarsson I, Nordmark G, Jönsen A, Bengtsson AA, Sturfelt G, Rantapää-Dahlqvist S, Elvin K, Sundin U, Garnier S, Simard JF, Sigurdsson S, Padyukov L, Syvänen A-C, and Rönnblom L. A <em>STAT4</em> risk allele is associated with ischaemic cerebrovascular events and anti-phospholipid antibodies in systemic lupus erythematosus. Annals of the rheumatic diseases 69: 834-840, 2010.
- Meinert C, Gembardt F, Böhme I, Tetzner A, Wieland T, Greenberg B, and Walther T. Identification of intracellular proteins and signaling pathways in human endothelial cells regulated by angiotensin-(1–7). Journal of Proteomics 130: 129-139, 2016. [CrossRef] [PubMed]
- Hotamisligil GS, Arner P, Caro JF, Atkinson RL, and Spiegelman BM. Increased adipose tissue expression of tumor necrosis factor-alpha in human obesity and insulin resistance. J Clin Invest 95: 2409-2415, 1995.
- Sharma S, Adrogue JV, Golfman L, Uray I, Lemm J, Youker K, Noon GP, Frazier OH, and Taegtmeyer H. Intramyocardial lipid accumulation in the failing human heart resembles the lipotoxic rat heart. Faseb j 18: 1692-1700, 2004. [CrossRef] [PubMed]
- Del Alamo JC, Lemons D, Serrano R, Savchenko A, Cerignoli F, Bodmer R, and Mercola M. High throughput physiological screening of iPSC-derived cardiomyocytes for drug development. Biochim Biophys Acta 1863: 1717-1727, 2016.
- D’Aquila T, Zembroski AS, and Buhman KK. Diet Induced Obesity Alters Intestinal Cytoplasmic Lipid Droplet Morphology and Proteome in the Postprandial Response to Dietary Fat. Front Physiol 10: 180, 2019.
- Zadoorian A, Du X, and Yang H. Lipid droplet biogenesis and functions in health and disease. Nature Reviews Endocrinology 19: 443-459, 2023.
- Martinez-Mateu L, Saiz J, and Aromolaran AS. Differential Modulation of I(K) and I(Ca,L) Channels in High-Fat Diet-Induced Obese Guinea Pig Atria. Front Physiol 10: 1212, 2019.
- Ridker PM, MacFadyen JG, Thuren T, and Libby P. Residual inflammatory risk associated with interleukin-18 and interleukin-6 after successful interleukin-1beta inhibition with canakinumab: further rationale for the development of targeted anti-cytokine therapies for the treatment of atherothrombosis. Eur Heart J 41: 2153-2163, 2020.
- Mazurek T, Zhang L, Zalewski A, Mannion JD, Diehl JT, Arafat H, Sarov-Blat L, O’Brien S, Keiper EA, Johnson AG, Martin J, Goldstein BJ, and Shi Y. Human epicardial adipose tissue is a source of inflammatory mediators. Circulation 108: 2460-2466, 2003.
- Barbarroja N, Lopez-Pedrera R, Mayas MD, Garcia-Fuentes E, Garrido-Sanchez L, Macias-Gonzalez M, El Bekay R, Vidal-Puig A, and Tinahones FJ. The obese healthy paradox: is inflammation the answer? Biochem J 430: 141-149, 2010. [CrossRef] [PubMed]
- Vyas V, Blythe H, Wood EG, Sandhar B, Sarker SJ, Balmforth D, Ambekar SG, Yap J, Edmondson SJ, Di Salvo C, Wong K, Roberts N, Uppal R, Adams B, Shipolini A, Oo AY, Lawrence D, Kolvekar S, Lall KS, Finlay MC, and Longhi MP. Obesity and diabetes are major risk factors for epicardial adipose tissue inflammation. JCI Insight 6: 2021. [CrossRef] [PubMed]
- Wu CK, Tsai HY, Su MY, Wu YF, Hwang JJ, Tseng WY, Lin JL, and Lin LY. Pericardial fat is associated with ventricular tachyarrhythmia and mortality in patients with systolic heart failure. Atherosclerosis 241: 607-614, 2015.
- Fuller B, Garland J, Anne S, Beh R, McNevin D, and Tse R. Increased Epicardial Fat Thickness in Sudden Death From Stable Coronary Artery Atherosclerosis. Am J Forensic Med Pathol 38: 162-166, 2017. [CrossRef] [PubMed]
- Packer M. Epicardial Adipose Tissue May Mediate Deleterious Effects of Obesity and Inflammation on the Myocardium. J Am Coll Cardiol 71: 2360-2372, 2018.
- Kelley N, Jeltema D, Duan Y, and He Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int J Mol Sci 20: 2019.
- Vm M, Al S, Aa A, As Z, Av K, Rs O, Im M, and Ga K. Circulating interleukin-18: Association with IL-8, IL-10 and VEGF serum levels in patients with and without heart rhythm disorders. Int J Cardiol 215: 105-109, 2016. [CrossRef] [PubMed]
- Gupta A, Fei YD, Kim TY, Xie A, Batai K, Greener I, Tang H, Ciftci-Yilmaz S, Juneman E, Indik JH, Shi G, Christensen J, Gupta G, Hillery C, Kansal MM, Parikh DS, Zhou T, Yuan JX, Kanthi Y, Bronk P, Koren G, Kittles R, Duarte JD, Garcia JGN, Machado RF, Dudley SC, Choi BR, and Desai AA. IL-18 mediates sickle cell cardiomyopathy and ventricular arrhythmias. Blood 137: 1208-1218, 2021.
- Bers DM, and Despa S. Na+ transport in cardiac myocytes; Implications for excitation-contraction coupling. IUBMB life 61: 215-221, 2009.
- Varro A, Nanasi PP, and Lathrop DA. Potassium currents in isolated human atrial and ventricular cardiocytes. Acta Physiol Scand 149: 133-142, 1993. [CrossRef]
- Aromolaran AS, Subramanyam P, Chang DD, Kobertz WR, and Colecraft HM. LQT1 mutations in KCNQ1 C-terminus assembly domain suppress IKs using different mechanisms. Cardiovasc Res 104: 501-511, 2014.
- Cheng EP, Yuan C, Navedo MF, Dixon RE, Nieves-Cintron M, Scott JD, and Santana LF. Restoration of normal L-type Ca2+ channel function during Timothy syndrome by ablation of an anchoring protein. Circ Res 109: 255-261, 2011. [CrossRef] [PubMed]
- Aromolaran AS, Srivastava U, Ali A, Chahine M, Lazaro D, El-Sherif N, Capecchi PL, Laghi-Pasini F, Lazzerini PE, and Boutjdir M. Interleukin-6 inhibition of hERG underlies risk for acquired long QT in cardiac and systemic inflammation. PLoS One 13: e0208321, 2018.
- Hagiwara Y, Miyoshi S, Fukuda K, Nishiyama N, Ikegami Y, Tanimoto K, Murata M, Takahashi E, Shimoda K, Hirano T, Mitamura H, and Ogawa S. SHP2-mediated signaling cascade through gp130 is essential for LIF-dependent I CaL, [Ca2+]i transient, and APD increase in cardiomyocytes. J Mol Cell Cardiol 43: 710-716, 2007. [CrossRef] [PubMed]
- Landstrom AP, Dobrev D, and Wehrens XHT. Calcium Signaling and Cardiac Arrhythmias. Circ Res 120: 1969-1993, 2017. [CrossRef] [PubMed]
- Wang J, Wang H, Zhang Y, Gao H, Nattel S, and Wang Z. Impairment of HERG K(+) channel function by tumor necrosis factor-alpha: role of reactive oxygen species as a mediator. J Biol Chem 279: 13289-13292, 2004.
- Monnerat G, Alarcon ML, Vasconcellos LR, Hochman-Mendez C, Brasil G, Bassani RA, Casis O, Malan D, Travassos LH, Sepulveda M, Burgos JI, Vila-Petroff M, Dutra FF, Bozza MT, Paiva CN, Carvalho AB, Bonomo A, Fleischmann BK, de Carvalho AC, and Medei E. Macrophage-dependent IL-1beta production induces cardiac arrhythmias in diabetic mice. Nat Commun 7: 13344, 2016. [CrossRef] [PubMed]
- Li YH, and Rozanski GJ. Effects of human recombinant interleukin-1 on electrical properties of guinea pig ventricular cells. Cardiovasc Res 27: 525-530, 1993.
- Kotake S, Sato K, Kim KJ, Takahashi N, Udagawa N, Nakamura I, Yamaguchi A, Kishimoto T, Suda T, and Kashiwazaki S. Interleukin-6 and soluble interleukin-6 receptors in the synovial fluids from rheumatoid arthritis patients are responsible for osteoclast-like cell formation. J Bone Miner Res 11: 88-95, 1996. [CrossRef] [PubMed]
- Rose-John S. The Soluble Interleukin 6 Receptor: Advanced Therapeutic Options in Inflammation. Clin Pharmacol Ther 102: 591-598, 2017.
- Campbell L, Chen C, Bhagat SS, Parker RA, and Ostor AJ. Risk of adverse events including serious infections in rheumatoid arthritis patients treated with tocilizumab: a systematic literature review and meta-analysis of randomized controlled trials. Rheumatology (Oxford, England) 50: 552-562, 2011.
- Smolen JS, Beaulieu A, Rubbert-Roth A, Ramos-Remus C, Rovensky J, Alecock E, Woodworth T, Alten R, and Investigators O. Effect of interleukin-6 receptor inhibition with tocilizumab in patients with rheumatoid arthritis (OPTION study): a double-blind, placebo-controlled, randomised trial. Lancet 371: 987-997, 2008. [CrossRef] [PubMed]
- Ding C, and Jones G. Anti-interleukin-6 receptor antibody treatment in inflammatory autoimmune diseases. Rev Recent Clin Trials 1: 193-200, 2006.
- Kawashiri SY, Kawakami A, Yamasaki S, Imazato T, Iwamoto N, Fujikawa K, Aramaki T, Tamai M, Nakamura H, Ida H, Origuchi T, Ueki Y, and Eguchi K. Effects of the anti-interleukin-6 receptor antibody, tocilizumab, on serum lipid levels in patients with rheumatoid arthritis. Rheumatol Int 31: 451-456, 2011. [CrossRef] [PubMed]
- Chen B, Zhang S, Wang B, Chen H, Li Y, Cao Q, Zhong J, Xie M, Ran Z, Tang T, Yang M, Guo T, Xu B, Cai Z, Ma L, Schreiber S, and Chen M. 775b Olamkicept, an IL-6 Trans-Signaling Inhibitor, is Effective for Induction of Response and Remission in A Randomized, Placebo-Controlled Trial in Moderate to Severe Ulcerative Colitis. Gastroenterology 161: e28-e29, 2021.
- Chen B, Zhang S, Wang B, Chen H, Li Y, Cao Q, Zhong J, Xie M, Ran Z, Tang T, Yang M, Guo T, Xu B, Cai Z, Schreiber S, and Chen M. DOP01 Efficacy and safety of the IL-6 trans-signalling inhibitor olamkicept: a phase 2 randomized, placebo-controlled trial in moderately to severely active Ulcerative Colitis. Journal of Crohn’s and Colitis 15: S041-S042, 2021.
- Schreiber S, Aden K, Bernardes JP, Conrad C, Tran F, Hoper H, Volk V, Mishra N, Blase JI, Nikolaus S, Bethge J, Kuhbacher T, Rocken C, Chen M, Cottingham I, Petri N, Rasmussen BB, Lokau J, Lenk L, Garbers C, Feuerhake F, Rose-John S, Waetzig GH, and Rosenstiel P. Therapeutic Interleukin-6 Trans-signaling Inhibition by Olamkicept (sgp130Fc) in Patients With Active Inflammatory Bowel Disease. Gastroenterology 160: 2354-2366 e2311, 2021.
- Schulte DM, Waetzig GH, Schuett H, Marx M, Schulte B, Garbers C, Lokau J, Vlacil A-K, Schulz J, Seoudy AK, Schieffer B, Rosenstiel P, Seeger M, Laudes M, Rose-John S, Lützen U, Grote K, and Schreiber S. Case Report: Arterial Wall Inflammation in Atherosclerotic Cardiovascular Disease is Reduced by Olamkicept (sgp130Fc). Frontiers in Pharmacology 13: 2022.
- Salama A, Amin MM, and Hassan A. Effects of oleic acid and/or exercise on diet-induced thermogenesis and obesity in rats: involvement of beige adipocyte differentiation and macrophage M1 inhibition. Res Pharm Sci 18: 219-230, 2023.
- Conte M, Petraglia L, Poggio P, Valerio V, Cabaro S, Campana P, Comentale G, Attena E, Russo V, Pilato E, Formisano P, Leosco D, and Parisi V. Inflammation and Cardiovascular Diseases in the Elderly: The Role of Epicardial Adipose Tissue. Front Med (Lausanne) 9: 844266, 2022.
- Konwerski M, Gąsecka A, Opolski G, Grabowski M, and Mazurek T. Role of Epicardial Adipose Tissue in Cardiovascular Diseases: A Review. Biology (Basel) 11: 2022.
- Swifka J, Weiss J, Addicks K, Eckel J, and Rosen P. Epicardial fat from guinea pig: a model to study the paracrine network of interactions between epicardial fat and myocardium? Cardiovasc Drugs Ther 22: 107-114, 2008.
- Back M, Yin L, and Ingelsson E. Cyclooxygenase-2 inhibitors and cardiovascular risk in a nation-wide cohort study after the withdrawal of rofecoxib. Eur Heart J 33: 1928-1933, 2012.
- Schmidt M, Christiansen CF, Mehnert F, Rothman KJ, and Sorensen HT. Non-steroidal anti-inflammatory drug use and risk of atrial fibrillation or flutter: population based case-control study. BMJ 343: d3450, 2011.
- van der Hooft CS, Heeringa J, Brusselle GG, Hofman A, Witteman JC, Kingma JH, Sturkenboom MC, and Stricker BH. Corticosteroids and the risk of atrial fibrillation. Arch Intern Med 166: 1016-1020, 2006.
- De Caterina R, Ruigomez A, and Rodriguez LA. Long-term use of anti-inflammatory drugs and risk of atrial fibrillation. Arch Intern Med 170: 1450-1455, 2010.
- Granier M, Massin F, and Pasquie JL. Pro- and anti-arrhythmic effects of anti-inflammatory drugs. Antiinflamm Antiallergy Agents Med Chem 12: 83-93, 2013.
Figure 1.
Effects of high fat diet feeding on guinea pig hearts. Compared to LFD-fed male and female guinea pig ventricular samples (A), HFD animals showed a marked increase in interstitial fibrosis (B), but not in OAD-fed guinea pigs (C). D, Quantification of percent (%) fibrotic area in the ventricular samples (n= 4 LFD, 5 HFD, 3 OAD). E, Confocal immunofluorescent images of LFD (Top image) and HFD (Bottom image) guinea pig ventricular slices analyzed using TGFβ antibodies (Red), WGA (Green), and DAPI (Blue), showed significantly increased TGFβ fluorescence labeling in HFD ventricular tissue slices compared to LFD controls (F) suggesting HFD feeding promotes the overactivation of inflammatory signaling in guinea pig heart (n= 5 LFD, 5 HFD). Numbers in brackets, represent number of hearts examined. Data columns represent mean ± S.E.M, *Statistical significance at P < 0.05.
Figure 1.
Effects of high fat diet feeding on guinea pig hearts. Compared to LFD-fed male and female guinea pig ventricular samples (A), HFD animals showed a marked increase in interstitial fibrosis (B), but not in OAD-fed guinea pigs (C). D, Quantification of percent (%) fibrotic area in the ventricular samples (n= 4 LFD, 5 HFD, 3 OAD). E, Confocal immunofluorescent images of LFD (Top image) and HFD (Bottom image) guinea pig ventricular slices analyzed using TGFβ antibodies (Red), WGA (Green), and DAPI (Blue), showed significantly increased TGFβ fluorescence labeling in HFD ventricular tissue slices compared to LFD controls (F) suggesting HFD feeding promotes the overactivation of inflammatory signaling in guinea pig heart (n= 5 LFD, 5 HFD). Numbers in brackets, represent number of hearts examined. Data columns represent mean ± S.E.M, *Statistical significance at P < 0.05.
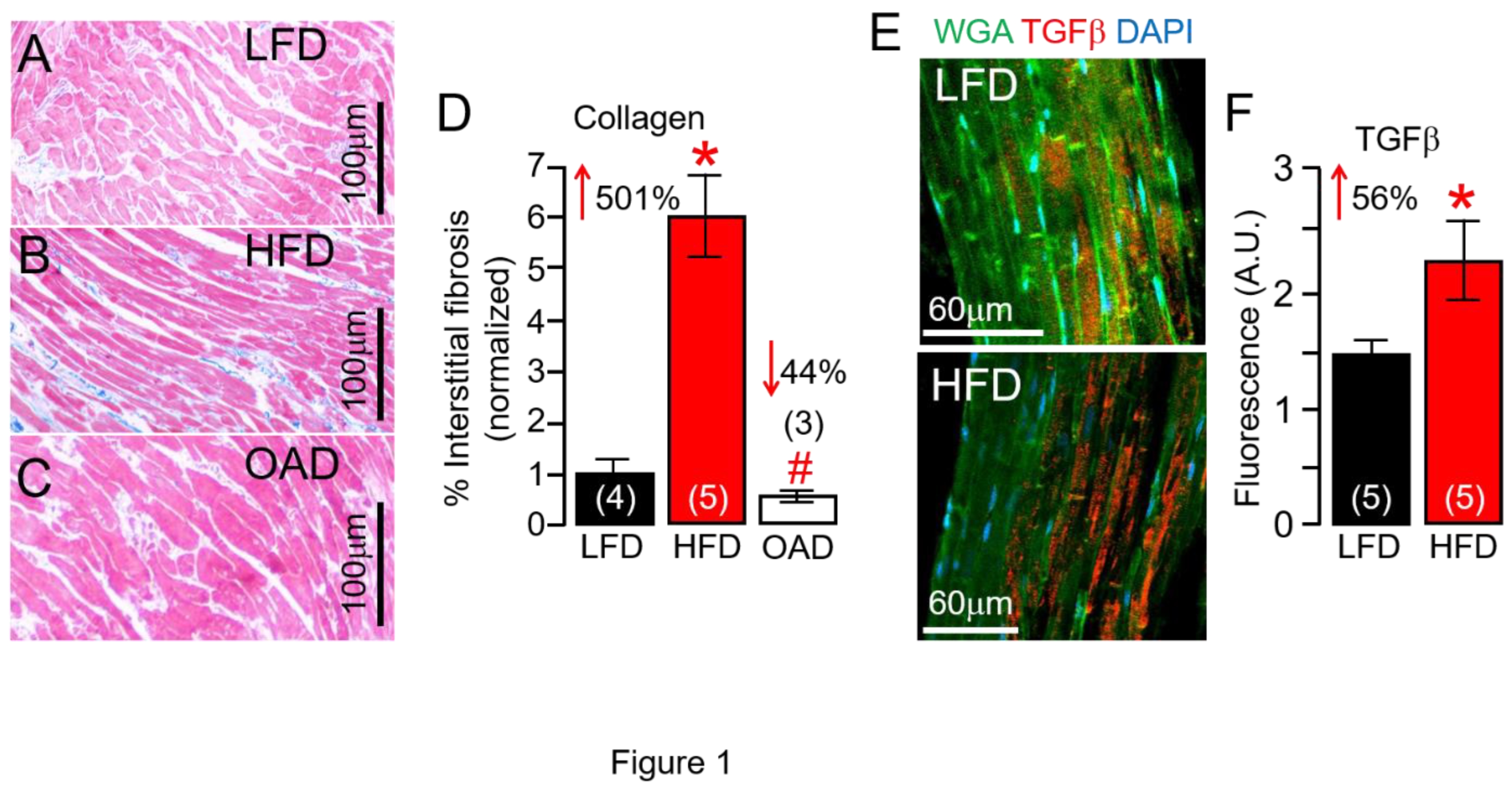
Figure 2.
High fat diet feeding is associated with overactivation of proinflammatory cytokine expression in guinea pigs. Column graphs represent ELISA assay quantification (in pg/ml), of IL-6 (A,B), IL-1(C,D), TNF (E,F) measured in LFD and HFD guinea pig serum and ventricular samples respectively. Comparison is made between serum vs. cardiac levels for a specific cytokine. Data columns are mean ± S.E.M, n = 4-7 LFD, 9-14 HFD. *Statistical significance at P < 0.05.
Figure 2.
High fat diet feeding is associated with overactivation of proinflammatory cytokine expression in guinea pigs. Column graphs represent ELISA assay quantification (in pg/ml), of IL-6 (A,B), IL-1(C,D), TNF (E,F) measured in LFD and HFD guinea pig serum and ventricular samples respectively. Comparison is made between serum vs. cardiac levels for a specific cytokine. Data columns are mean ± S.E.M, n = 4-7 LFD, 9-14 HFD. *Statistical significance at P < 0.05.
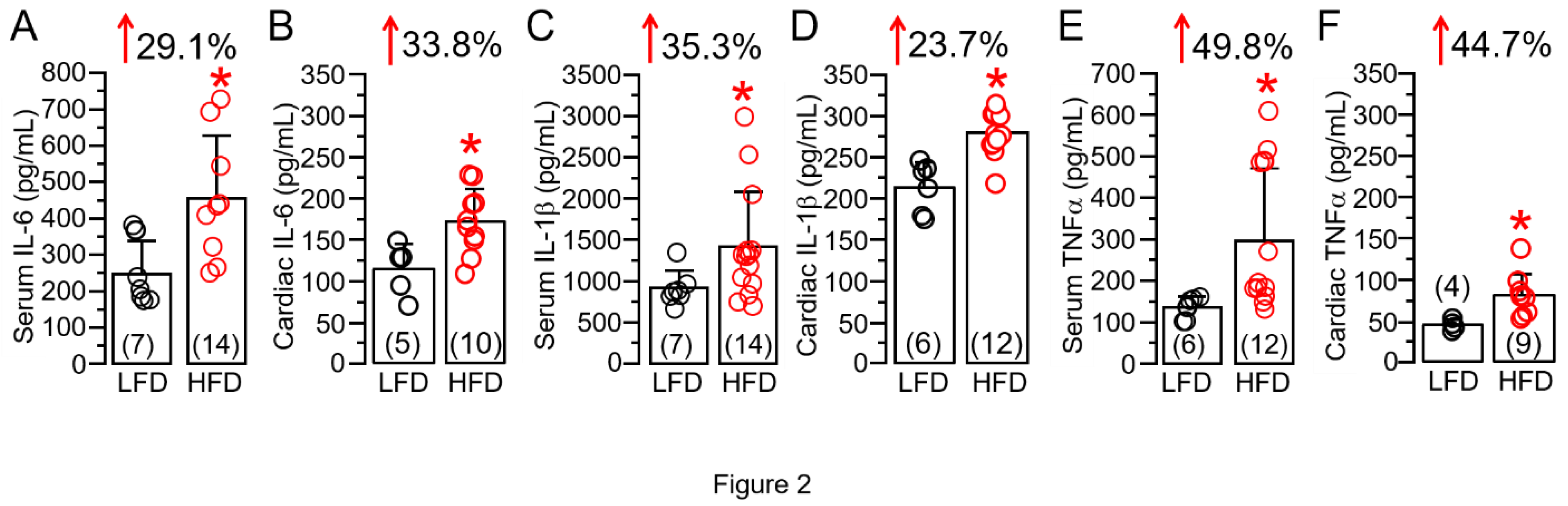
Figure 3.
High fat diet adult guinea pigs show vulnerability to spontaneous ventricular arrhythmogenesis. A, Cartoon depiction of the experimental protocol used to assess the effect of HFD feeding on QT interval and VT risk. B, Overlay of representative traces of surface ECG recorded in LFD (Black trace) and HFD (Red trace) showing HFD guinea pigs displayed prolonged QT interval compared to LFD controls. C, Exemplar ECG traces demonstrating triggered ventricular arrhythmic signatures and arrhythmias measured in HFD challenged adult guinea pigs. D, Averaged mRNA expression of IL-6R and JAK2 measured in LFD and HFD ventricular tissue samples (n = 4-5 LFD, 8 HFD). Data are expressed as the fold change in IL-6R and JAK2 expression compared with LFD after normalization to GAPDH. Data revealed HFD feeding induced profound increases in IL-6R and JAK2 expression compared to LFD-fed controls demonstrating overactivation of IL-6 trans-signaling in HFD hearts. Data columns are mean ± S.E.M, n = 4-8 separate experiments. *Statistical significance at P < 0.05.
Figure 3.
High fat diet adult guinea pigs show vulnerability to spontaneous ventricular arrhythmogenesis. A, Cartoon depiction of the experimental protocol used to assess the effect of HFD feeding on QT interval and VT risk. B, Overlay of representative traces of surface ECG recorded in LFD (Black trace) and HFD (Red trace) showing HFD guinea pigs displayed prolonged QT interval compared to LFD controls. C, Exemplar ECG traces demonstrating triggered ventricular arrhythmic signatures and arrhythmias measured in HFD challenged adult guinea pigs. D, Averaged mRNA expression of IL-6R and JAK2 measured in LFD and HFD ventricular tissue samples (n = 4-5 LFD, 8 HFD). Data are expressed as the fold change in IL-6R and JAK2 expression compared with LFD after normalization to GAPDH. Data revealed HFD feeding induced profound increases in IL-6R and JAK2 expression compared to LFD-fed controls demonstrating overactivation of IL-6 trans-signaling in HFD hearts. Data columns are mean ± S.E.M, n = 4-8 separate experiments. *Statistical significance at P < 0.05.
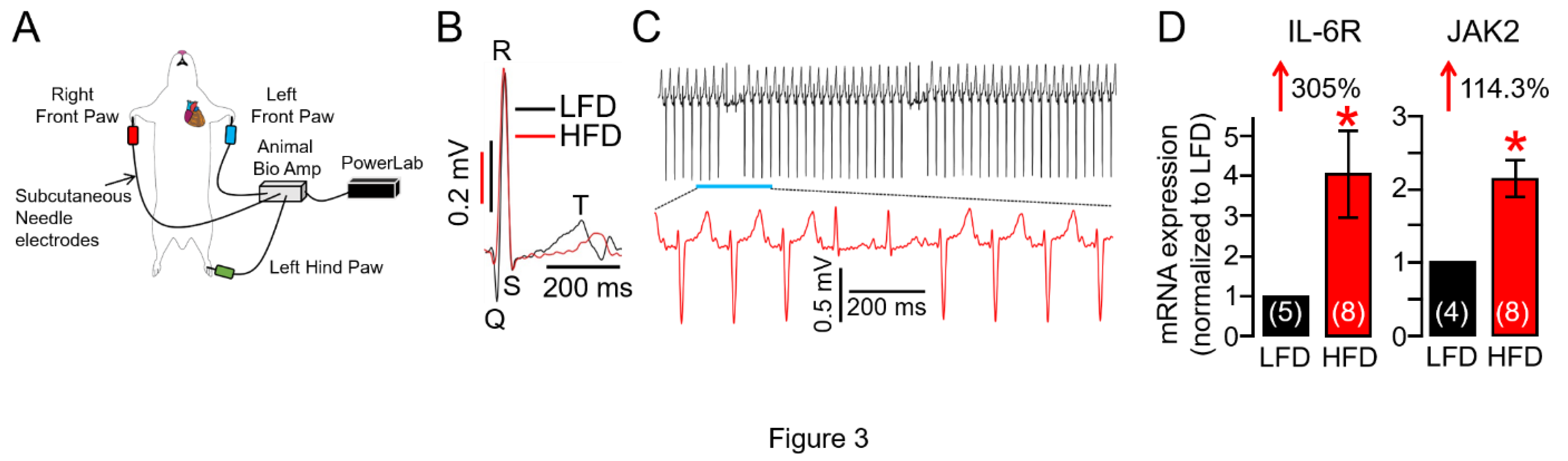
Figure 4.
Overstimulation of IL-6 trans-signaling induces QT prolongation in guinea pigs under conditions of cardiac challenge. A, Experimental protocol used to assess the effect of the overstimulation of IL-6 trans-signaling on guinea pig QT interval induced by recombinant IL-6 and sIL-6R injected in the cranial vena cava (CVC). B, Representative traces of surface ECG recorded in IL-6-sIL-6R (Top panel), and untreated control (Bottom panel) guinea pigs. The subsequent exposure to the -adrenergic receptor agonist ISO in the continued presence of IL-6-sIL-6R triggered ventricular arrhythmias (C,D). Averaged mRNA expression of KCNQ1 (E) and KCNE1 (F) measured in LFD and HFD ventricular tissue samples (n = 4 LFD, 5 HFD). Data are expressed as the fold change in expression compared with LFD after normalization to GAPDH. Our data revealed severely reduced expression of KCNQ1 and KCNE1 coding for IKs in HFD ventricular samples in agreement with dramatic ventricular electrical remodeling and increased risk for exercise induced VT. Data columns represent mean ± S.E.M from n= 4-5 guinea pig/group, *Statistical significance at P < 0.05.
Figure 4.
Overstimulation of IL-6 trans-signaling induces QT prolongation in guinea pigs under conditions of cardiac challenge. A, Experimental protocol used to assess the effect of the overstimulation of IL-6 trans-signaling on guinea pig QT interval induced by recombinant IL-6 and sIL-6R injected in the cranial vena cava (CVC). B, Representative traces of surface ECG recorded in IL-6-sIL-6R (Top panel), and untreated control (Bottom panel) guinea pigs. The subsequent exposure to the -adrenergic receptor agonist ISO in the continued presence of IL-6-sIL-6R triggered ventricular arrhythmias (C,D). Averaged mRNA expression of KCNQ1 (E) and KCNE1 (F) measured in LFD and HFD ventricular tissue samples (n = 4 LFD, 5 HFD). Data are expressed as the fold change in expression compared with LFD after normalization to GAPDH. Our data revealed severely reduced expression of KCNQ1 and KCNE1 coding for IKs in HFD ventricular samples in agreement with dramatic ventricular electrical remodeling and increased risk for exercise induced VT. Data columns represent mean ± S.E.M from n= 4-5 guinea pig/group, *Statistical significance at P < 0.05.
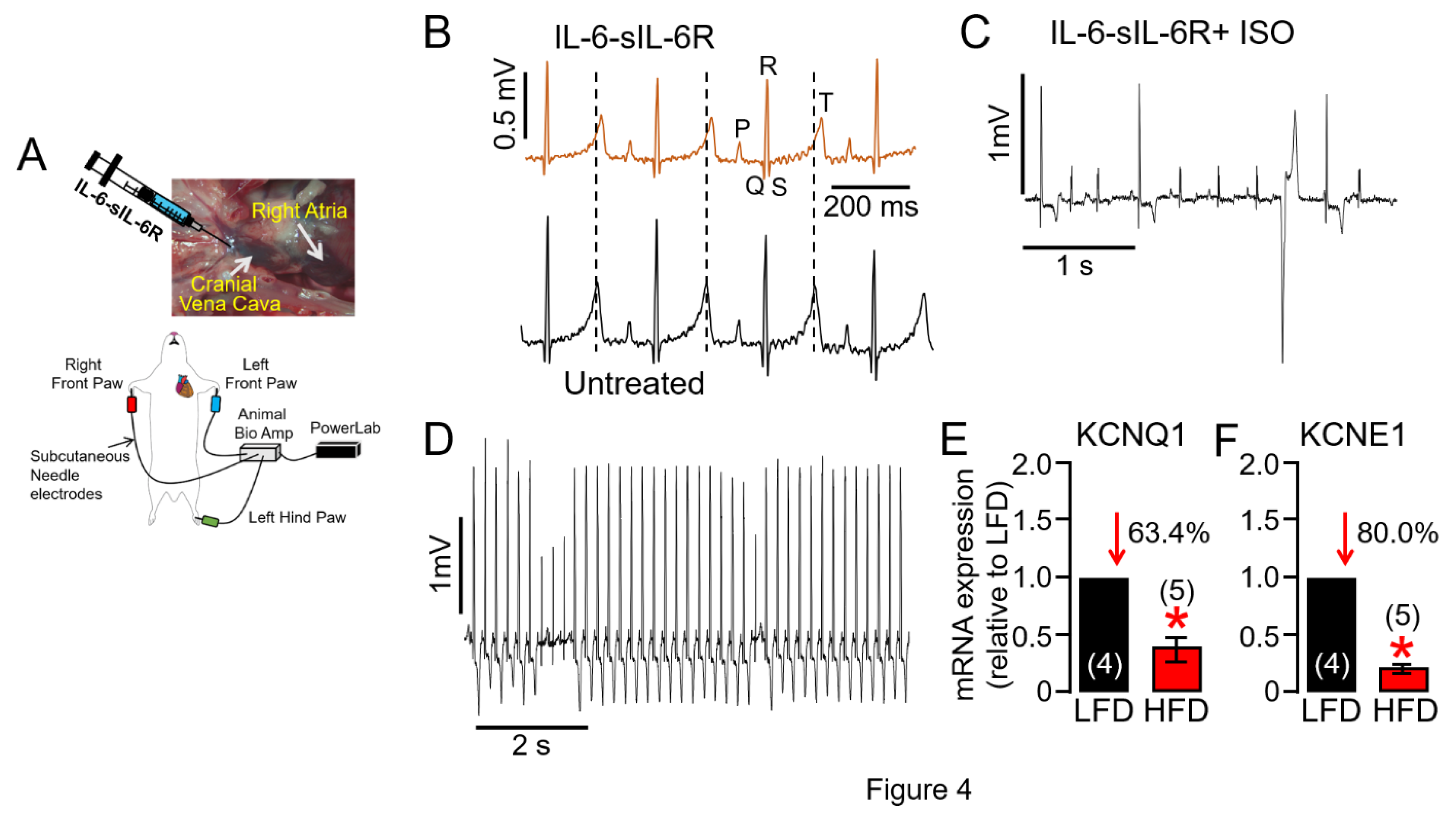
Figure 5.
Direct stimulation of IL-6 trans-signaling with hIL-6 and coumermycin induces severe arrhythmias in guinea pigs. A, Cartoon illustration of experimental protocol and arrhythmia inducibility in guinea pigs. Guinea pigs challenged with hIL-6 (B) or JAK2 activator (C) severely prolonged the QT interval and triggered ventricular arrhythmic signatures (D,E).
Figure 5.
Direct stimulation of IL-6 trans-signaling with hIL-6 and coumermycin induces severe arrhythmias in guinea pigs. A, Cartoon illustration of experimental protocol and arrhythmia inducibility in guinea pigs. Guinea pigs challenged with hIL-6 (B) or JAK2 activator (C) severely prolonged the QT interval and triggered ventricular arrhythmic signatures (D,E).
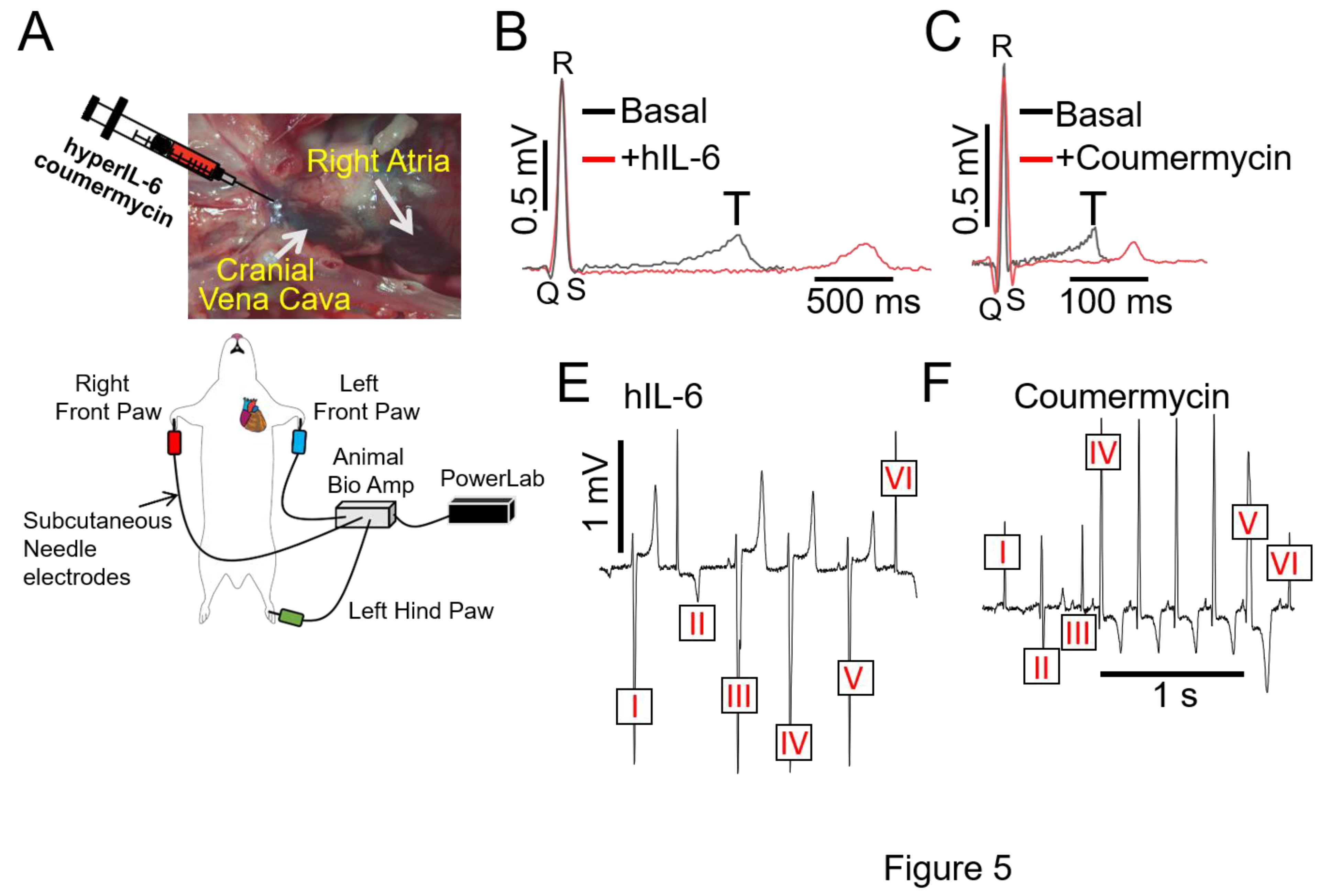
Figure 6.
IL-18 treatment increases the risk of severe arrhythmia under conditions of overactive of IL-6 trans-signaling. A, Cartoon illustration of experimental protocol and arrhythmia inducibility in conditions of cardiac hyperinflammation. IL-6-sIL-6R with IL-18 displayed conduction block and asystole (B-E) in 4/6 guinea pigs, in-line with a potential role for IL-18 to exacerbate dramatic ventricular electrical activity (ion channel remodeling, action potential phenotypes), due to overactive IL-6 trans-signaling.
Figure 6.
IL-18 treatment increases the risk of severe arrhythmia under conditions of overactive of IL-6 trans-signaling. A, Cartoon illustration of experimental protocol and arrhythmia inducibility in conditions of cardiac hyperinflammation. IL-6-sIL-6R with IL-18 displayed conduction block and asystole (B-E) in 4/6 guinea pigs, in-line with a potential role for IL-18 to exacerbate dramatic ventricular electrical activity (ion channel remodeling, action potential phenotypes), due to overactive IL-6 trans-signaling.
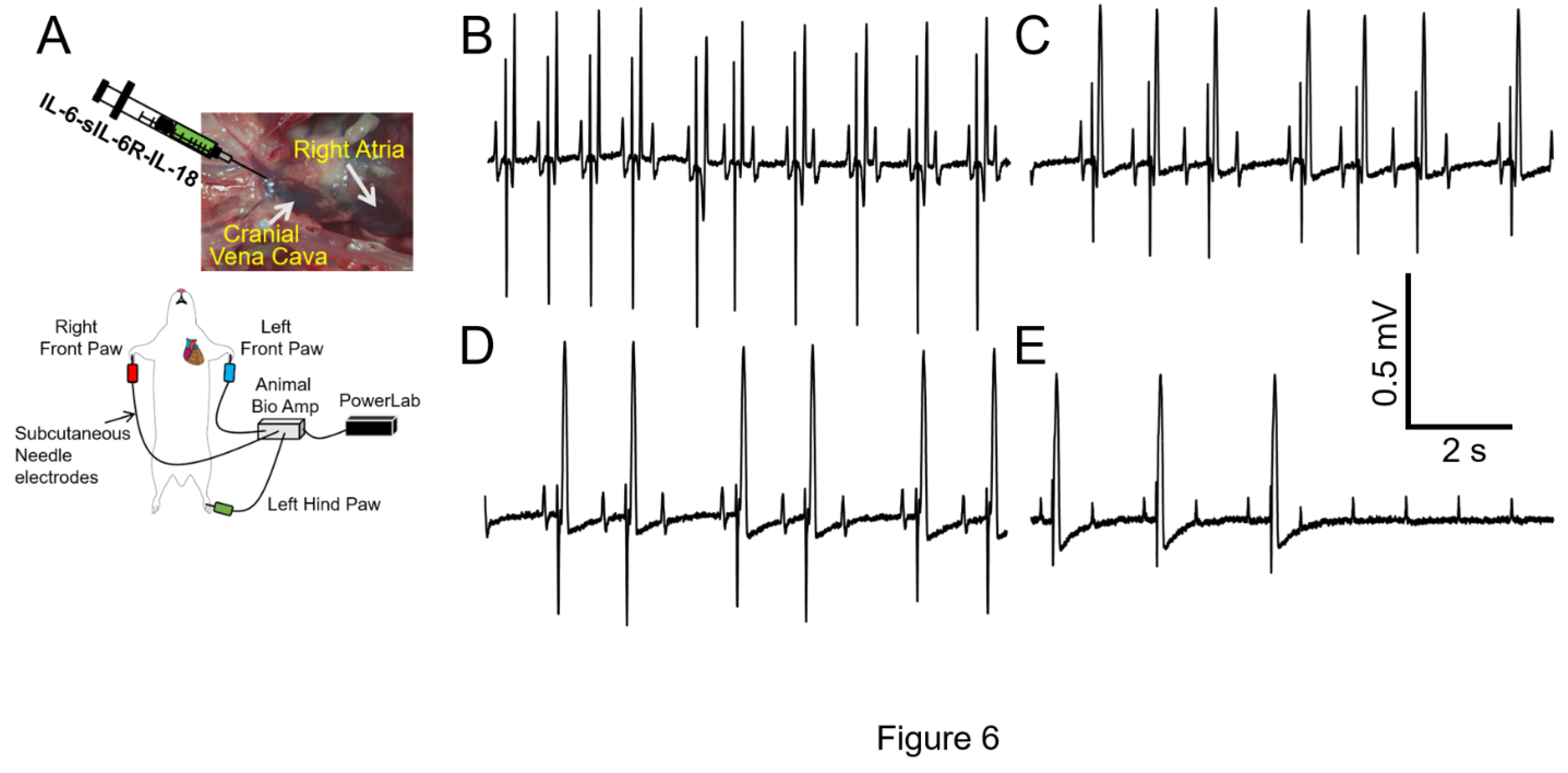
Figure 7.
Effect of multiple proinflammatory cytokines on arrhythmia risk in guinea pigs under conditions of IKs inhibition. A, Cartoon illustration of experimental protocol for arrhythmia induction. B, Exemplar ECG traces recorded from guinea pigs initially treated with the IKs blocker chromanol 293B, and subsequently challenged with cytomix (IL-6-sIL-6R+IL-1+TNF+IL-18) and ISO. Expanded view of ventricular electrical activities revealed severe arrhythmogenesis. Data were generated from three different guinea pig hearts.
Figure 7.
Effect of multiple proinflammatory cytokines on arrhythmia risk in guinea pigs under conditions of IKs inhibition. A, Cartoon illustration of experimental protocol for arrhythmia induction. B, Exemplar ECG traces recorded from guinea pigs initially treated with the IKs blocker chromanol 293B, and subsequently challenged with cytomix (IL-6-sIL-6R+IL-1+TNF+IL-18) and ISO. Expanded view of ventricular electrical activities revealed severe arrhythmogenesis. Data were generated from three different guinea pig hearts.
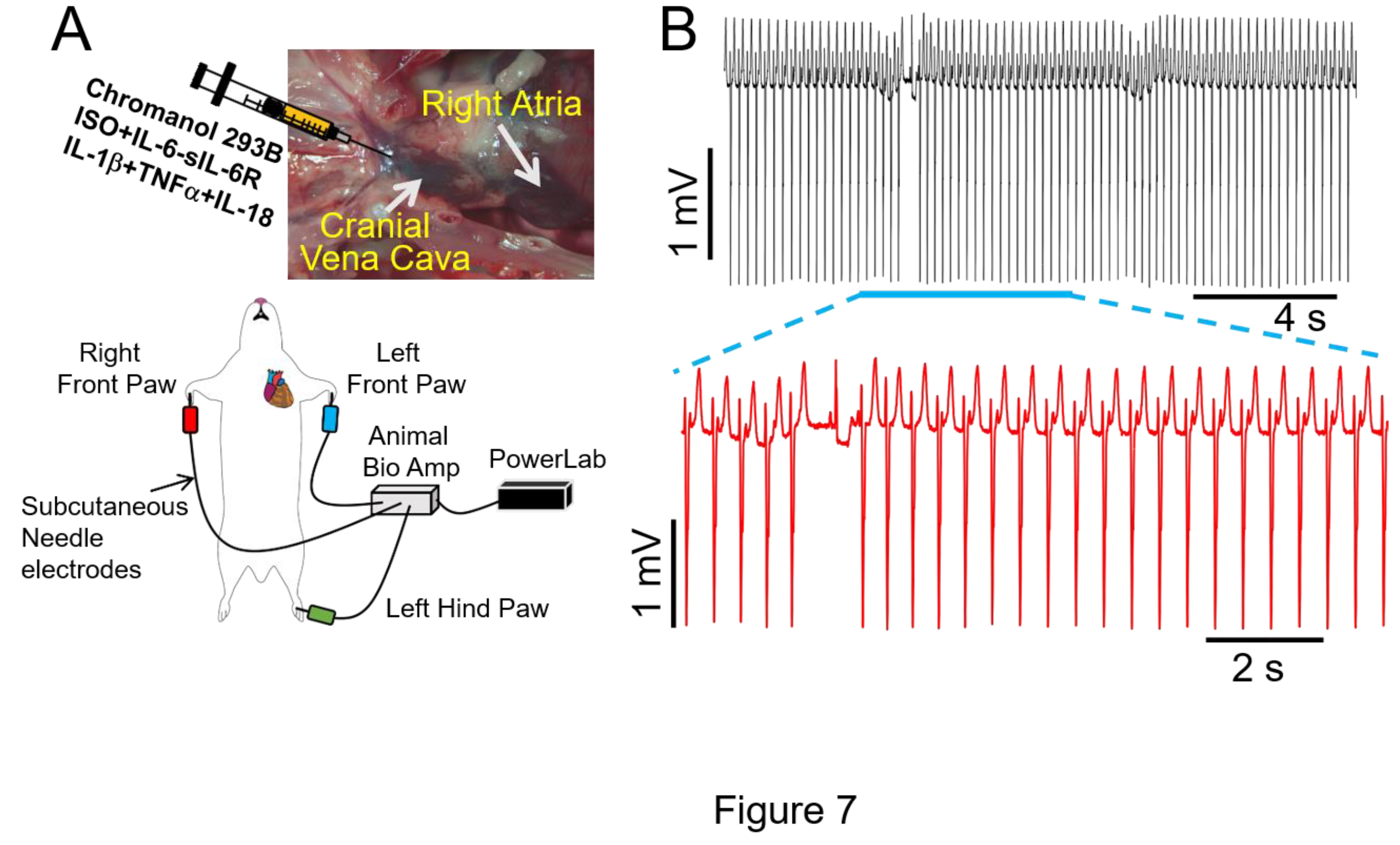
Figure 8.
Effects of high fat diet on subcellular localization of hERG1a channels, IL-18, and IL-6R in guinea pig ventricular tissue slices. A, Confocal images of permeabilized LFD and HFD adult guinea pig ventricular tissue samples showing, hERG1a (Green), WGA (Red), and DAPI (Blue), fluorescence images. B, Normalized mean fluorescence signals from yellow positive tissue samples provide an index of relative surface density of hERG1a in HFD samples (n = 4 LFD, 6 HFD). Data for IL-18 (C) or IL-6R (D) antibodies indicates increased surface expression of IL-18 and IL-6R in HFD compared to LFD-fed controls, same format as A,B (n = 3-5 LFD, 3-5 HFD). Data columns are mean ± S.E.M. hERG1a experiments had 4-8 images per animal. IL-18 and IL-6R experiments had 3-4 images and 3-7 images per animal respectively. *Statistical significance at P < 0.05.
Figure 8.
Effects of high fat diet on subcellular localization of hERG1a channels, IL-18, and IL-6R in guinea pig ventricular tissue slices. A, Confocal images of permeabilized LFD and HFD adult guinea pig ventricular tissue samples showing, hERG1a (Green), WGA (Red), and DAPI (Blue), fluorescence images. B, Normalized mean fluorescence signals from yellow positive tissue samples provide an index of relative surface density of hERG1a in HFD samples (n = 4 LFD, 6 HFD). Data for IL-18 (C) or IL-6R (D) antibodies indicates increased surface expression of IL-18 and IL-6R in HFD compared to LFD-fed controls, same format as A,B (n = 3-5 LFD, 3-5 HFD). Data columns are mean ± S.E.M. hERG1a experiments had 4-8 images per animal. IL-18 and IL-6R experiments had 3-4 images and 3-7 images per animal respectively. *Statistical significance at P < 0.05.
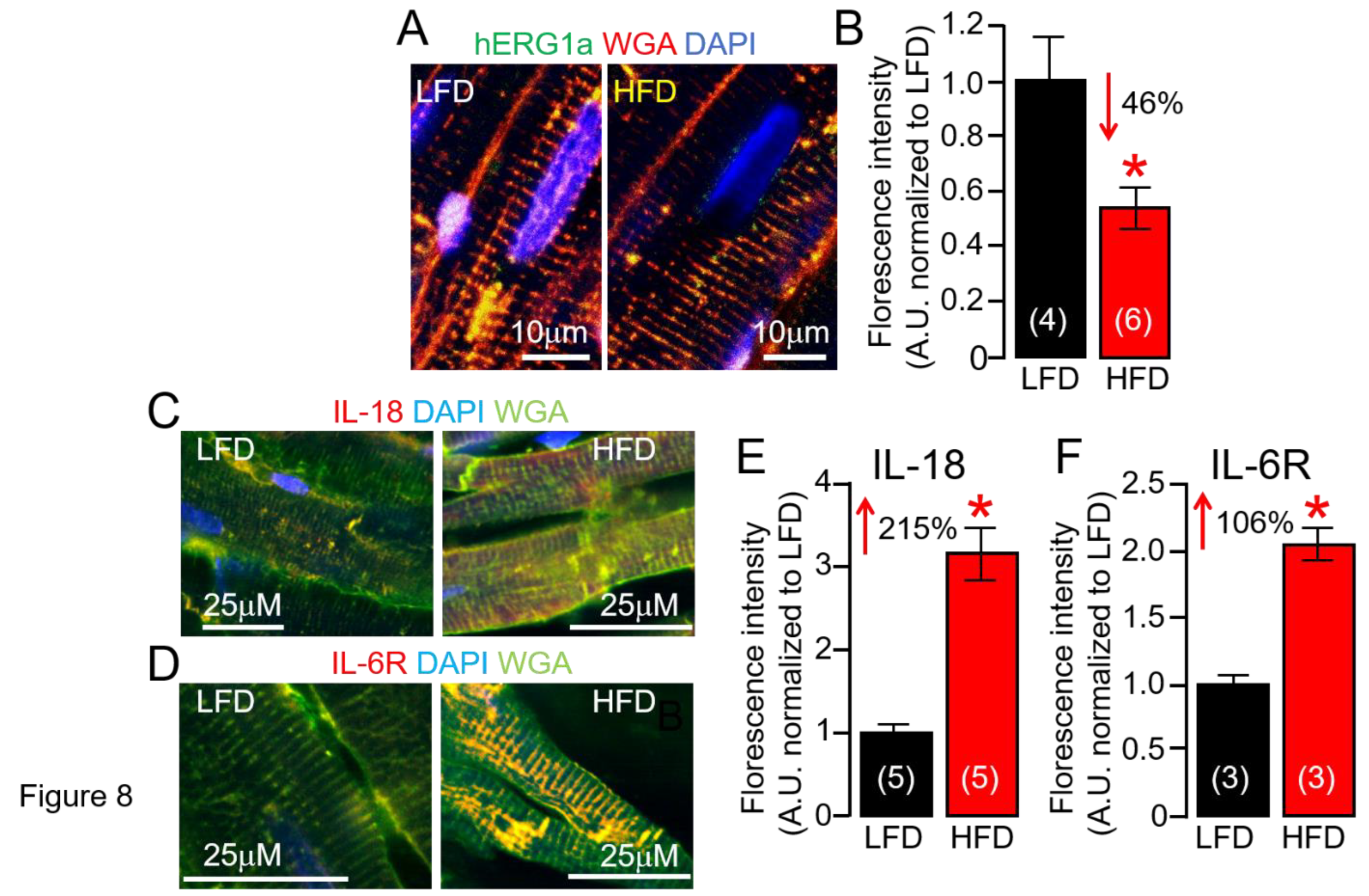
Figure 9.
High fat diet selectively induces the phosphorylation of STAT4 in guinea pig ventricular tissue slices. Confocal images of permeabilized LFD and HFD adult guinea pig ventricular tissue samples showing, STAT4 (A) or pSTAT4 (B, Red), WGA (Green), and DAPI (Blue), fluorescence images (A,B). C, Normalized mean fluorescence signals from magenta- and magenta-blue positive tissue ventricular samples provide an index of relative expression of pSTAT4/STAT4 and nuclear translocation (n = 3 LFD, 7 HFD). HFD promotes phosphorylation of STAT4 leading to its activation and enhanced nuclear translocation of pSTAT4/STAT4 more than LFD controls and pSTAT3/STAT3 nuclear expression. D-F, Data for STAT3 (D) or pSTAT3 (E), same format as A-C (n = 3 LFD, 3 HFD). Data columns are mean ± S.E.M. 3 images are included for each animal with a total number of segmented nuclei per image ranging from 8 to 24 *Statistical significance at P < 0.05.
Figure 9.
High fat diet selectively induces the phosphorylation of STAT4 in guinea pig ventricular tissue slices. Confocal images of permeabilized LFD and HFD adult guinea pig ventricular tissue samples showing, STAT4 (A) or pSTAT4 (B, Red), WGA (Green), and DAPI (Blue), fluorescence images (A,B). C, Normalized mean fluorescence signals from magenta- and magenta-blue positive tissue ventricular samples provide an index of relative expression of pSTAT4/STAT4 and nuclear translocation (n = 3 LFD, 7 HFD). HFD promotes phosphorylation of STAT4 leading to its activation and enhanced nuclear translocation of pSTAT4/STAT4 more than LFD controls and pSTAT3/STAT3 nuclear expression. D-F, Data for STAT3 (D) or pSTAT3 (E), same format as A-C (n = 3 LFD, 3 HFD). Data columns are mean ± S.E.M. 3 images are included for each animal with a total number of segmented nuclei per image ranging from 8 to 24 *Statistical significance at P < 0.05.
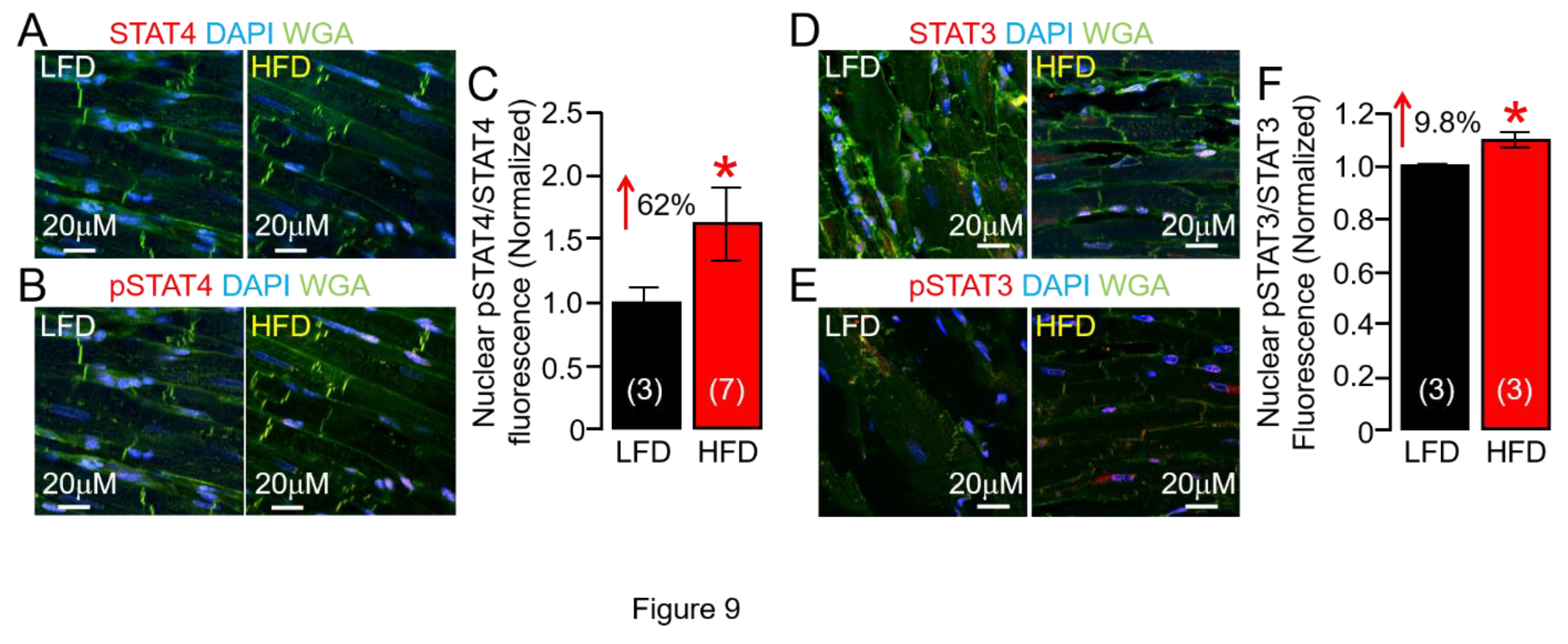
Figure 10.
Lipotoxicity induces lipid droplet remodeling and increased expression of IL-6R in guinea pig ventricular tissue slices. Confocal images of permeabilized adult guinea pig ventricular cardiomyocytes showing lipid droplet (magenta) accumulation in untreated (A) and PA-BSA (B, a potent inducer of lipotoxicity). Compared to BSA-treated control conditions, PA-BSA-treated ventricular myocytes significantly increased lipid droplet count (C) and size (D) and promoted IL-6R expression (E-G). Data columns are mean ± S.E.M, n = 16-37 images. *Statistical significance at P < 0.05.
Figure 10.
Lipotoxicity induces lipid droplet remodeling and increased expression of IL-6R in guinea pig ventricular tissue slices. Confocal images of permeabilized adult guinea pig ventricular cardiomyocytes showing lipid droplet (magenta) accumulation in untreated (A) and PA-BSA (B, a potent inducer of lipotoxicity). Compared to BSA-treated control conditions, PA-BSA-treated ventricular myocytes significantly increased lipid droplet count (C) and size (D) and promoted IL-6R expression (E-G). Data columns are mean ± S.E.M, n = 16-37 images. *Statistical significance at P < 0.05.
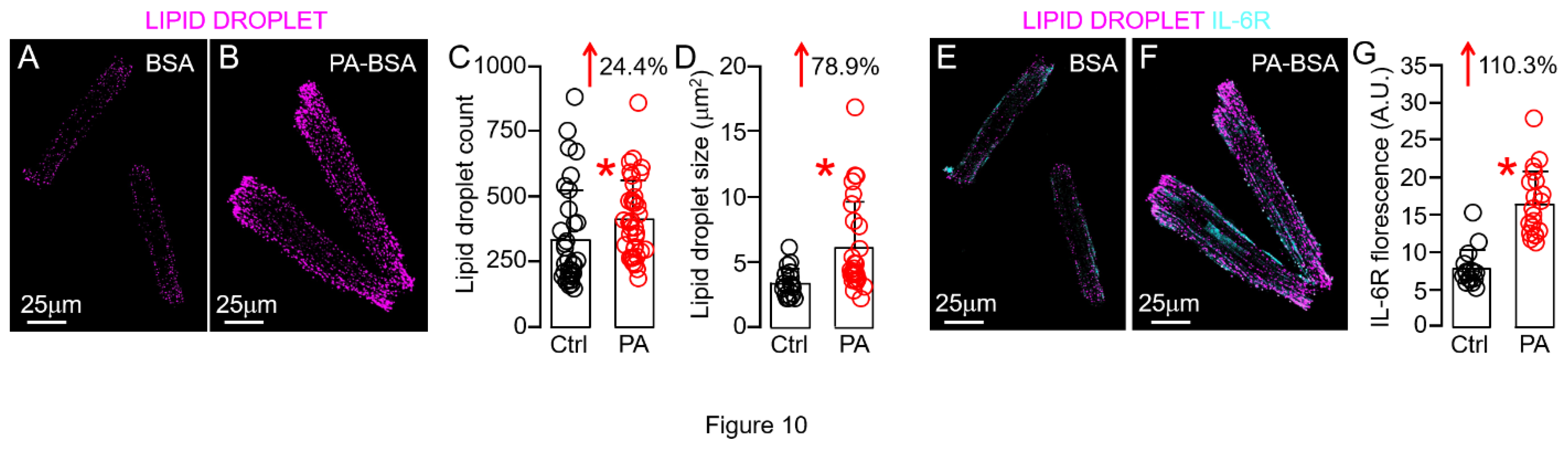

Table 1.
List and sequence of primers used in this manuscript for qPCR amplification.
| Gene | Sequence (5’- > 3´) | Gene ID |
|---|---|---|
| IL-6R | sense GGGTCGGGCTTCAAGATGTTA antisense AACGGTGCCTGTATTCTGGG |
100730490 |
| JAK2 | sense CTTAGATTACGCCGCCCAGC antisense TGTGCCGGTATGACCCTCTA |
100722908 |
| KCNQ1 | sense GCTGTTCTCTGAGGGTCTTCCA antisense CCATCCACCCTGAACTCTTTCT |
100379230 |
| KCNE1 | sense TCCCAGGAAAACTGTCAGCTC antisense CGGTTCTGAGGAAGCGGATT |
100135562 |
Table 2.
QTc interval measured in guinea pigs challenged with different dietary interventions.
| Conditions | QTc (ms) (Basal) |
QTc (ms) (Post-intervention) |
P Value |
Δ QTc (ms) |
n |
|---|---|---|---|---|---|
| Low fat diet (LFD) | 279.8±15.2 | 269.4±18.1 | 0.665 | -10.5±4.51 | 8 |
| High fat diet (HFD) | 260.4±15.1 | 323.9±15.2* | 0.0018 | 63.32±10.9 | 15 |
| Oleic acid diet (OAD) | 294.7±6.14 | 307.8±5.58 | 0.137 | 13.1±8.09 | 8 |
Data are means ± S.E.M. * p < 0.05 compared to corresponding basal controls, two-tailed paired Student’s t test.
Table 3.
Effect of overactive pro-inflammatory cytokine and IKs inhibition on QTc interval measured in guinea pigs.
Table 3.
Effect of overactive pro-inflammatory cytokine and IKs inhibition on QTc interval measured in guinea pigs.
| Conditions | QTc (ms) (Basal) |
QTc (ms) (Post-intervention) |
P Value |
Δ QTc (ms) |
n |
|---|---|---|---|---|---|
| Vehicle | 297.3±2.54 | 302.7±3.94 | 0.298 | 5.42±1.45 | 4 |
| IL-6-sIL-6R | 289.3±0.72 | 310.9±4.22* | 0.003 | 21.66±6.12 | 5 |
| hyperIL-6 (hIL-6) | 299.7±2.61 | 333.9±8.63* | 0.004 | 34.8±8.15 | 8 |
| hIL-6+Olamkicept | 292.9±5.47 | 293.8±4.24 | 0.899 | 0.93±8.30 | 3 |
| Coumermycin | 299.1±5.40 | 337.5±10.7* | 0.01 | 38.4±10.9 | 7 |
| IL-6-sIL-6R+IL-18 | 292.3±10.7 | 319.2±10.7* | 0.0004 | 26.95±3.98 | 6 |
| Chromanol293B | 308.6±4.24 | 327.9±5.68* | 0.018 | 19.23±3.35 | 8 |
| Chromanol293B+ISO- 10 min | 308.6±4.24 | 342.5±7.69* | 0.004 | 33.0±10.4 | 6 |
| Chromanol293B+ISO+Cytomix- 10 min | 308.6±4.24 | 363.6±6.48* | 0.0036 | 54.2±11.02 | 3 |
| Chromanol293B+ISO+Cytomix- 30 min | 308.6±4.24 | 377.8±15.3* | 0.03 | 68.4±17.8 | 3 |
Data are means ± S.E.M. * p < 0.05 compared to corresponding basal controls, two-tailed paired Student’s t test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



