
Submitted:
20 June 2024
Posted:
20 June 2024
You are already at the latest version
Abstract
Salinity stress emerges as an important menace to global food security, particularly affecting susceptible crops such as common bean plants (Phaseolus vulgaris L.). The disruption of ion balances caused by salt stress withing plants cells reduces water and nutrient absorption, and hampers growth and productivity. Sacchorhiza polyschides, a brown macroalga rich in essential elements, and macrominerals, has drawn attention as a natural biostimulant, being a potential remedy to bolster crop resilience against salt stress. This study delves into the effect of water extracts of Sacchorhiza polyschides (SPE) across varying concentrations (1%, 2%, and 5%) in common bean plants under salt stress (68.4 mM NaCl) conditions. Two different methods of application were used: by spraying the aerial part of the plant or by irrigation together with the watering solution. The effects of both systems of application were studied on the aerial parts of the plants. SPE exhibited significant enhancements in morphological parameters, chlorophyll content, carbon-nitrogen activities, and antioxidant enzymatic system activities, alongside increasing osmolyte accumulation, compared to salt-stressed plants without biostimulation. Moreover, SPE reduced reactive oxygen species (ROS) levels caused by salt stress, mitigating lipid peroxidation. The concentrations of 2% and 5% SPE applied by spraying showed the most significant results in most of the parameters analyzed. By shedding light on the underlying science behind this biostimulant effect, this research underscores the potential of SPE to enhance crop resilience and augment food production in salt-challenged environments.
Keywords:
Salt stress
; Saccorhiza polyschides extract
; Foliar application
; Irrigation
; Phaseolus vulgaris L.
; Enzyme activities
; Osmolytes
; Reactive oxygen species.
1. Introduction
Salinity stress is an environmental constraint that produces substantial limitations on crop productivity and quality worldwide [1], especially in those crops which are sensitive to this stress such as common bean (Phaseolus vulgaris L.) [2]. Common bean is a vital source of proteins and essential nutrients for millions of people worldwide. However, plant growth and productivity can be significantly affected by environmental stressors, with salt stress being a major concern [3]. Excessive salt concentration produces damage in plants by disrupting water uptake, producing ion toxicity and interfering with nutrient uptake [4]. For instance, excessive salts in the soil can replace essential nutrients like potassium (K+) and calcium (Ca2+), affecting vital cellular processes [5]. Ion toxicity results from the buildup of toxic amounts of Na+ in plant tissues and cells, which negatively impacts the plant's ability to grow and develop. Na+ is not necessary for plants, in contrast to animals (except in some C4 plants) [6,7] and Its overabundance will be extremely harmful to plants, causing cytosolic K+ efflux, which will disrupt cellular homeostasis, oxidative stress, interfere with K+ and Ca2+ functions, nutrient shortage, stunted growth, and even cell death [8,9].
During the emergence and development of saline stress inside the plant, all major processes such as photosynthesis, protein synthesis, energy metabolism are affected. The first response is the reduction in the speed of extension of the leaf surface, followed by stopping the extension with the intensification of stress [10]. The oxidative stress caused by salinity increases the formation of reactive oxygen species (ROS), such as superoxide anion (O2−) and hydrogen peroxide (H2O2), which can perturbate the structure of cells, leading to DNA mutation, protein denaturation and lipid peroxidation [11]. Maintaining vital processes in these conditions of high salinity therefore requires a resistance of the plant to dehydration, by an adaptation of its osmotic potential to restore water relations and ensure a suitable water supply, also by means of an effective control of intracellular ion fluxes [12]. To prevent ROS-induced damage and lessen its negative effects, plant cells are outfitted with highly developed antioxidant defense systems, both enzymatic and non-enzymatic. Within each organelle of cells, there are one or more antioxidants that work to detoxify a specific ROS [13], therefore, plants exposed to salinity and water stressors have adopted the important strategy of enhanced synthesis and accumulation of low molecular weight compounds. To survive under stressful circumstances, plants often store free amino acids, non-structural carbohydrates, and quaternary ammonium compounds [14].
In recent years, natural biostimulant exploration has emerged as a promising avenue for improving crop resilience against salt stress [15]. Saccorhiza polyschides (SP) is a large and bulky brown macroalgae species belonging to Phyllariaceae family, whose size varies from 1.5 to more than 5 m in length with up to 3.5 m width. Usually found on the lowest shore part, it is widely distributed on west European, Mediterranean and north-west African coasts. Samples of SP were taken from natural algae fields located in Sidi Rahal marine coast, in the south of Casablanca, Morocco. This alga possesses a unique bioactive composition with significant amounts of relevant macrominerals, and trace elements essential for plant growth [16]. Apart from this, this edible seaweed has the potential of ensuring the production of large amounts of biomass for feed and food [17]. The appeal of such a natural solution lies in its potential to mitigate environmental and ecological impacts of traditional agricultural interventions [18]. As a potential biostimulant source, SP could provide a natural and sustainable solution to mitigate adverse effects of salt stress on crops like common beans, and other salt-sensitive plants, enhancing their resilience in saline environments. Mamede et al. [19] reported that the application of the S. polyschides extract (SPE) increased soil fertility and hence, turnip green plants enhanced their root system, photosynthetic activity and the essential nutrient uptake.
The current investigation focuses on the exploration of the role of SPE at different concentrations (1%, 2%, and 5%) in the mitigation of salt stress (68.4 mM NaCl) in common bean plants. The biostimulant effect is analysed testing physiological and biochemical processes on the plant such as growth parameters, indole acetic acid, sugars and amino acids, chlorophyll content, antioxidant enzyme activities and carbon-nitrogen enzyme activities. The objective was not only to scientifically understand the biostimulant phenomenon, but also to provide practical insights of applications in agriculture to alleviate salt-induced stress in common beans crops.
2. Materials and Methods
2.1. Graphical experimental design
In this study, we investigated the use of brown seaweed extract derived from Saccorhiza polyschides (SPE) to improve the ability of common bean (Phaseolus vulgaris L.) plants, a major global legume, to withstand salt stress. We evaluated the impact of the foliar application and irrigation with SPE in water at different concentrations (0, 1, 2, 5% (v/v)) on common bean plants under salt stress. The experimental design and the main results of this study are illustrated in Figure 1.
2.2. Preparation of Seaweed Extract
Saccorhiza polyschides (SP) specimens were carefully selected and identified and picked with complete thalli. Samples were thoroughly rinsed with tap water to eliminate salt and any potentially unwanted substances or dirt. SP full thalli were subsequently dried and finely ground. Crude extract stock solutions were obtained by mixing 100 g of dried algae powder with 1 L of distilled water. For two hours, these solutions were heated, and then filtered to eliminate any solid debris. Resulting filtrate served as 100% crude extract stock, from which experimental biostimulant solutions were later prepared. Working biostimulant solutions were made by diluting the crude extract with additional distilled water to obtain experimental solutions containing 1%, 2%, and 5% of Saccorhiza polyschides extracts (SPE).
2.2.1. Chemical composition of Saccorhiza polyschides Extract
The chemical composition of SPE was analyzed. The measurements of indole acetic acid and free amino acids were made using the protocols for plants given below. Secondary metabolites, such as flavonoids and polyphenols, were also determined according to Bouchmaa et al. [19].
2.3. Plant Material and Growth Conditions
Seeds of a variety of common bean (Phaseolus vulgaris L.var. nanus (L.)) were selected based on size and color. After two to three minutes of disinfection with a 5% sodium hypochlorite solution, they were rinsed with distilled water. Sterilized seeds were sown, and, after germination, five plants were selected and cultivated per pot. They were kept at 22°C with a 15 h lighting photoperiod and an irradiance of 250 μmole m-2 s-1. A total of 32 pots were used for the present experiment (4 repetitions for each condition were made) (Figure 1).
Plants were sprayed every 5 days with different SPE concentrations (1%, 2%, and 5%) and were watered every 4 days with freshwater (NSS), with NaCl solution (68.4 mM for SS), or with NaCl solution mixed with the SPE in the irrigation method. After 2 months, all plants were harvested and morphologically characterized, measuring growth parameters, mainly length, weight, and leaf diameter. Chemical and enzymatic analyses were carried out using freshly harvested plant leaves (Figure 1).
2.4. Estimation of Malondialdehyde Content
Malondialdehyde (MDA) content was measured using the method described by Bouchmaa et al. [20] to ascertain the extent of lipid peroxidation with a few minor modifications. To create a cell homogenate, trichloroacetic acid (20%) and tetrabutylammonium hydroxide (TBA) (0.67%) were added to plant cell homogenate and mixed with a vortex. Following an hour of boiling, ice was used to cool the mixture, and 1 mL of n-butanol was added before centrifuging for 12 minutes at 12,000 g. Afterwards, the supernatant was collected and analyzed with a spectrophotometer at 532 and 600 nm.
2.5. Estimation of O2- Content
The superoxide (O2-) content in common bean plant leaves was measured according to Kubiś [21].
2.6. Estimation of H2O2 Content
The content of peroxide (H2O2) was ascertained in accordance with Mrid et al. [22]. After pulverizing 0.2 g of fresh leaves with 1 mL of 0.1% TCA, the mixture was centrifuged at 12,000 g for 15 minutes. The obtained supernatant was combined with 2 mL of 1 M iodide potassium (KI) and 1 mL of 10 mM phosphate buffer pH 7. Following a 1-hour incubation period in a dark environment, the optical density (OD) was measured at 390 nm. A standard curve was used to calculate the concentrations of H2O2.
2.7. Chlorophyll Assay
Leaves chlorophyll content was measured using the methodology described by Arnon [23]. During the process, 800 µL of 80% acetone was mixed with 200 µl of soluble plant homogenate pellets in water. Three independent replicates of the amount of chlorophyll were measured following a 72-hour incubation period at 4°C. The levels of chlorophyll-a and chlorophyll-b were evaluated in the following ways:
Chlorophyll-a (mg/ml) = (0.0127*OD663) - (0.0269* OD645)
Chlorophyll-b (mg/ml) = (0.0229*OD645) - (0.00468* OD663)
2.8. Plant Extract Preparation for Measurement of Soluble Sugar, Amino Acid, and Indole Acetic Acid Contents
After drying the leaves of common bean at 37 °C and using 80% ethanol, 200 mg of the dried material were extracted at 4°C. The amount of soluble sugar, amino acids, and indole acetic acid in the supernatant was measured after centrifugation at 5,000 g for 15 minutes at 4°C.
2.8.1. Estimation of Indole Acetic Acid Content
The amount of indole acetic acid (IAA) generated in common bean leaves was determined according to Ennoury et al. [24], using Salkowski's reagent method and commercial IAA as standard. Thus, 500 μL of this reagent (2% 1.5 M ferric chloride (FeCl3) in 36% perchloric acid (HClO4)) was combined with 1.5 mL of common bean plant extract, and the mixture was incubated for 30 minutes at 25°C. Subsequently, absorbance was determined at 530 nm using a blank containing 500 μL of Salkowski's reagent and 1.5 mL of distilled water.
2.8.2. Estimation of Amino Acid content
Amino acid content was evaluated by applying the methodology described in Sun et al. [25]. Thus, 1.5 mL of 2% ninhydrin solution (solubilized in 0.2 M citrate buffer, pH 5 and ethylene glycol) was mixed with an aliquot of 500 µL plant extract. The mixture was boiled for 15 minutes after being stirred with a Vortex. After allowing this mixture to cool at room temperature, the absorbance was measured at 546 nm. A calibration curve made with various glycine concentrations was used to calculate the amount of amino acids.
2.8.3. Estimation of Soluble Sugar Content
Soluble sugar concentration was determined using the method reported by Yemm and Willis [26]. Thus, 2 mL of anthrone reagent (1 g/L of anthrone dissolved in 95% sulfuric acid H2SO4) and 1 mL of common bean leaf extract were mixed. After being vortexed and heated to 100°C in a water bath for 10 minutes, the mixture was left to cool on ice. The absorbance of the samples was measured at 630 nm, and a calibration curve with various concentrations of anhydrous glucose was used to calculate the sugar content of the samples.
2.9. Extraction and Assay of Antioxidant Activities
For the enzyme extraction process, fresh common bean leaves (0.2 g) were combined with a buffer that contained 10 mM magnesium chloride, 100 mM HEPES-KOH, 20 µM FAD, 1 mM phenylmethylsulfonyl fluoride, and 14 mM β-mercaptoethanol. At 4°C and 20,000 x g, the homogenate was centrifuged for 20 minutes. The supernatant was used to determine the subsequent enzyme activities. Thereby, Beauchamp and Fridovich [27] methodology was used to measure the superoxide dismutase (SOD) activity (EC 1.15.1.1).
Glutathione reductase (GR) activity was determined using NADPH oxidation at 340 nm (extinction coefficient 6.2 mM- 1 cm-1) in accordance with Mrid et al. [23]. In this regard, 0.2 mM GSSG and 0.2 mM NADPH were combined with 0.1 M phosphate buffer, pH 7.8. Glutathione peroxidase (GPx) activity was assessed at 360 nm after NADPH oxidation. The Bouchmaa et al. [20] protocol was used to measure the enzyme activity.
Glutathione S-Transferase (GST) activity was measured using the protocol outlined by Latique et al. [28]. The reaction mixture was supplemented with 0.1 M sodium buffer pH 6.5, 2 mM 1-chloro-2,4-dinitrobenzene, 5 mM glutathione, and 0.1% triton x100. Then, a molar extinction coefficient of 9.6 mM−1 cm−1 was used to measure the activity of GST at 340 nm.
2.10. Extraction and Assay of Nitrogen and Carbon Activities
Using a 100 mM Tris-HCl buffer (pH 8) mixture containing 10 mM MgCl2, 1.4 mM glycerol, 1.4 mM β-mercaptoethanol, 1 mM EGTA, 1 mM phenylmethylsulfonyl fluoride (PMSF), 16.5 µM chymostatin, 1 mM EDTA, and 9.4 µM leupeptin, common bean leaves were ground in a cooled mortar. The homogenate was centrifuged for 15 minutes at 4°C at 12,000 x g. Solid ammonium sulphate was added to the supernatant after 30 minutes, until it reached 60 % saturation. The pellet was resuspended in 300 µL of extraction buffer and utilized for enzyme activities after the saturated supernatant underwent another centrifugation.
The method described by El Omari et al. [29] was used to measure the activity of phosphoenolpyruvate carboxylase (PEPC). Glutamine synthetase (GS) and Aspartate aminotransferase (AAT) activities were measured using methods outlined by Kchikich et al. [30], and Glutamate dehydrogenase (GDH) activity was measured spectrophotometrically for 30 minutes at 340 nm, as described by Ben Mrid et al. [31].
Protein Determination
The total soluble protein content of the enzyme extracts was determined using the Bradford method [32], with the protein standard for the calibration curve being bovine serum albumin (BSA).
2.11. Statistical Analysis
For statistical analysis, all datasets were treated by ANOVA using general linear model in SAS, incorporating fixed salt stress level and SPE treatment. Comparison was achieved using Tukey´s test. Data analysis was performed using SAS 9.4 software. A p -value below 0.05 was considered significant.
To create a biplot of the studied parameters, principal component analysis (PCA) was carried out under IBM SPSS Statistics version 25.0, using KMO and Bartlett’s test as well as Oblimin with Kaiser Normalization in Rotation Method.
3. Results
3.1. Saccorhiza polyschides Extract Composition
SPE composition was determined (Table 1). Flavonoid and polyphenol secondary metabolytes presented amounts of 4.494 mg. g-1 DW and 1.327 mg. g-1 DW, respectively. Furthermore, these results showed also amounts of 0.264 mg. g-1 DW and 0.353 mg. g-1 DW for IAA and the free amino acids respectively.
3.2. Saccorhiza polyschides Extract enhanced the Growth Parameters of Common Bean Plants under Salt Stress
Results of salt stress (68.4 mM NaCl) effect on growth parameters of common bean plants, mainly Shoot length (cm), plant fresh weight (g), and leaf diameter (cm) are shown in Figure 2. These results clearly indicated that salt stress (SS) has a detrimental effect on the growth of common bean plants. As a matter of fact, plant length and weight, and leaf diameter were significantly reduced by salt stress compared to control (NSS).
Results shown in Figure 2 indicated also that SPE applied by spraying method had a strong biostimulant effect. Overall, a significant improvement was observed in all plant growth parameters analyzed in SPE treatments combined with salt stress conditions (SPE +SS), compared to those irrigated only with SS. The best results in all parameters (plant length, weight and leaf diameter) were observed using 5% of SPE under SS, followed by 2% SPE + SS. In irrigation method, SPE application did not influence the reduction of SS symptoms, except for leaf diameter when plants were irrigated with 2% or 5% SPE.
According to these results, it clearly seems that, under SS conditions, 5% SPE treatment with spraying method is more efficient than irrigation method in terms of the performances of growth parameters. However, compared to control results (NSS), all treatments showed equal or less effect except in the two cases of plant length parameter using 5% and 2% SPE in spraying method.
3.3. Saccorhiza polyschides Extract Enhanced Chlorophyll and Osmolytes in Common Bean Plants under Salt Stress
3.3.1. Chlorophyll content increase in common bean leaves after biostimulation
As shown in Figure 3, chlorophyll content in common bean plants was negatively impacted by salt stress (SS) when compared to control (NSS), showing a reduction of 40%.
When compared to salt stress (SS control), chlorophyll content in stressed plants significantly increased after SPE application, both with irrigation and spraying conditions, and at all biostimulant concentrations. All treatments showed relevant improvements compared to SS control, but SPE being sprayed produced better results that when it was irrigated. The best result was observed using 5% SPE in both application methods compared to SS, but only the application of SPE 5% by spraying increased the chlorophyll content by 11% with respect to the content of the NSS plants.
3.3.2. Indole Acetic Acid and Osmolytes content (Soluble sugars and Amino acids) increase in common bean leaves after biostimulation
Within the present investigation, salt stress induced a significant reduction in IAA and osmolytes content in common bean plants but with variable magnitudes depending on osmolyte type (Figure 4).
In this regard, and compared to NSS control, IAA, soluble sugar and AA contents have been decreased under salt stress conditions. SPE application helped re-establish and increase hormone content in common bean plants under salt-stressed conditions. This positive effect was important, particularly at 5% SPE being sprayed. So, in terms of IAA, the best result was recorded using 5% SPE being sprayed which generated an important increase compared to SS control (Figure 4 a). However, the content of AIA leaves under all SPE treatments was always lower than those found in NSS plants.
SPE treatments significantly increased the soluble sugar content of common bean plants under SS control (Figure 4b), being this increase proportional to biostimulant concentrations. The application of SPE by spraying showed better results than by irrigation. As a matter of fact, the highest value recorded with 5% of SPE treatment in spraying method showed a great increase of soluble sugars compared to SS plants, being the content higher than those found in NSS plants.
In the case of amino acid (AA) content in common bean plants under salinity conditions (Figure 4 c), all SPE treatments were higher than SS control with almost same improvement levels in both irrigation and spraying methods. Best results were recorded with 5% SPE in both methods providing enhanced AA content in stressed common bean plants in comparison to the control (SS). However, compared to NSS control, all SPE treatments results were lower except 5% SPE which were almost equal in both irrigation and spraying methods.
3.4. Saccorhiza polyschides Extract Limits Oxidative Stress Markers in Common Bean Plants under Salt Stress
The presence of NaCl in soil produced oxidative stress, which in the end produced lipid peroxidation in the cells and could be measured as MDA content. In fact, the MDA content was 2.5 times higher in SS plants than in NSS plants (Figure 5 a). In addition, O2- and H2O2 contents, which are the oxidant agents that produce lipid peroxidation, were also significantly increased under SS conditions (Figure 5 b and c). However, the application of SPE, by both methods, reduced the lipid peroxidation levels in all concentrations assayed (Figure 5.a) although the best treatment was SPE 5%. In line with this, the SPE application reduced the levels of O2- (Figure 5 b) and H2O2 (Figure 5 c) compared to SS plants, being the O2- and H2O2 concentrations almost lower than in NSS when SPE 5% was sprayed on the leaves.
3.5. Saccorhiza polyschides Extract Boosts Antioxidant Enzymatic Activities in Common Bean Plants under Salt Stress
Salt stress increased all antioxidant activities assayed (SOD, GPx, GST and GR) in common bean plants (Figure 6), being 5% the best concentrations for all activities except for GPx in which SPE 1% was the best application. The irrigation methods also increased the activities assayed but to a minor extent and normally, at the highest concentration (SPE 5%).
3.6. Saccorhiza polyschides Extract Boosts the Carbon-Nitrogen Enzymatic System of Common Bean Plants under Salt Stress
The effects of salt stress in common bean plants, in terms of carbon-nitrogen enzymatic system, compared to NSS conditions showed variable results (Figure 7). In control plants, SS led to a marked increase in GDH and AAT activities compared to NSS (Figure 7b and d). However, PEPC and GS activities decreased under SS conditions compared to NSS (Figure 7a and c).
Moreover, the enzymatic system in stressed common bean plants treated with SPE using the three concentrations was significantly increased, especially when it was sprayed using 2% and 5%, for PEPC, GDH and GS compared to SS (Figure 7a, b and c).
SPE treatment for plants under SS helped restoring AAT activity, especially when this extract was sprayed compared to SS (Figure 7d), but were slightly higher in all three SPE treatments (1%, 2% and 5%) with almost the same level compared to SS (Figure 7d). However, although the SPE applied by irrigation also increased AAT activity compared to NSS, the levels never reached those measured in SS plants.
3.7. Principal Component Analysis (PCA)
In our research, we performed two principal component analyses (PCA) to investigate the relationships between the treatments applied and the morpho-physiological (Figure 8) and biochemical traits (Figure 9) observed in common bean plants. As depicted in Figure 8A, the first PCA component (PC1) explained approximately 52.38% of the total variation, and was related to the treatment, being the SS plants located at the negative part of this component. The second component (PC2) accounted only for 27.64 % of the variance observed in the dataset.
The scatter plot of treatments (Figure 8B) revealed that under salt stress, the effects of SPE at the concentration of 2% and 5% (SSPE2% and SSPE5%) were positively associated with the measured common bean growth parameters. However, these associations were almost completely lost when irrigation was used at 1% SPE (ISPE1%). SSPE application at 2% and 5% was negatively related to SS.
The PCA shown in Figure 9 revealed the relationship between the treatments, the enzymatic activities and oxidative stress markers analyzed in common bean plants under the different treatments. In this case, the PC1 and PC2 explained 51.44 % and 26.50 %, of the total variation respectively, and collectively accounted for 77.94 %. The pattern matrix with rotation method showed that all enzymes were correlated with the positive part of the PC1, whereas the oxidative stress markers such as lipid peroxidation (MDA), H2O2 and O2- content were correlated with the negative part of PC1. However, the PC2 seems to be related to the treatment, being NSS and SPE-biostimulated (SPC) plants in the positive part, and SS plants in the negative part (Figure 9A). The IAA content was a parameter not related to these treatments.
The scatter plot of treatments (Figure 9B) illustrated that SPE at 2 and 5 % concentrations by means of spraying (SSPE) had a positive correlation with NSS and a negative correlation with SS plants. However, the rest of applications were between NSS and SS plants.
4. Discussion
The main objective of this work is to evaluate the putative biostimulant effect of Saccorhiza polyschides extracts (SPE) on common bean plants under salt stress condition (68.4 mM NaCl), testing two types of SPE application: by spraying the aerial part of the plant or by irrigation. Salt stress has one of the most inhibitory impacts on several glycophytic crops such as Solanum, Capsicum, or Triticum plants, causing reduced growth, development, and productivity [33,34] The application of algae biostimulants has been described as a useful tool for improving crop protection against several stresses [35,36], and brown seaweeds have been largely employed mostly due to their year-round availability and large size. Most of the experiments in this sense have been developed using Ascophyllum nodosum extracts, which used as fertilizers enhance the growth, the quality and resilience under salt stress conditions in Lactuca sativa, Arabidopsis thaliana, and Solanum lycoperssicum among others [37,38,39]. However, as far as we know, no experiments were made using SPE to improve the resilience and productivity of common bean plants under salt stress conditions.
The biostimulant action of algae extracts was attributed to high amounts of macro- and micronutrients, hormones, phenolics, amino acids, carotenoids, vitamins and polyamines [40,41]. SPE contained significant levels of free amino acids, and auxins (Table 1), which could provide insight into why its application positively impacts on the growth of plants and their ability to withstand salt stress conditions. Similarly, a study done by Hussein et al. [43] on the effects of seaweed liquid extracts (SLEs) prepared from three Egyptian seaweeds (Ulva fasciata, Cystoseira compressa, and Laurencia obtusa) on seed priming, growth, and salinity stress mitigation on Vigna sinensis and Zea mays revealed that the physiologic stimulant effect of the micronutrient content of these SLEs and the presence of plant growth regulating substances could be used to understand the stimulating effects of SLEs on the growth of these plants.
The increment of salt in the substrate modifies the electrical conductivity of the soil, limiting the uptake of water by most of the glycophytic plants [44] such as common bean. The reduction in water content impairs the transpiration and stomal conductance, affecting the expansion of old leaves and the emergence of new ones [45]. This effect reduces plant growth, as we observed in common bean plants under salt stress, which reduced their shoot length, fresh weight and leaf diameter (Figure 2). Salt stress also affected photosynthetic parameters negatively, producing pigment degradation (Figure 3) and reducing the carbon fixation capacity of the plant, which lead to accumulation of less soluble sugars and amino acids (Figure 4). However, the application of algae extracts increased growth parameters, chlorophyll content and photosynthesis in Cucurbita pepo [46] and Triticcum aestivum [47] under salt stress. In accordance with this, 5% SPE application by spraying significantly increased growth parameters (Figure 2) and chlorophyll content (Figure 3) in common bean plants under salt stress compared to plants without biostimulation. The irrigation method only had significant effect at 2% and 5% SPE on leaf diameter and chlorophyll content (Figure 2 and Figure 3), suggesting an effect of SPE on cytokinin synthesis and/or mobilization from the root to the leaves [48].
The accumulation of osmolytes into the cells is a well-known strategy of halophyte plants to facilitate water entrance and protect cells and subcellular structures upon salt stress [45]. However, common bean plants reduced soluble sugar and amino acid content under salt stress (Figure 4). Similarly, HM et al. [49] reported a progressive decrease in amino acid content in maize and broad bean plants with increasing salt stress, as well as a decrease in soluble sugar content. Although, the SPE application increased their content to the levels of those found in non-stressed plants (Figure 4). This result has also been noted in tomato plants treated with A. nodosum extract [38], and the exogenous application of other algal extracts significantly increased osmolytes accumulation in plants exposed to various types of stress [50]. In this regard, a variety of organic compounds found in algal extracts have been described, including amino acids (such as proline, glycine, glutamine) and soluble sugars (such as glucose, sucrose, mannitol) that regulate osmotic balance and mitigate stress in plants.
When subject to salt stress, plants suffer an accumulation of reactive oxidizing species (ROS), such as superoxide anion (O2-) radicals and hydrogen peroxide (H2O2) [51], which could cause adverse effects such as increased lipid peroxidation as evidenced by elevated malondialdehyde (MDA) levels [51].
Recorded results of the current investigation revealed a noteworthy rise in lipid peroxidation damage, and a high O2- and H2O2 content in common bean plants under salt stress conditions (Figure 5). However, the application of 2% or 5% SPE significantly reduced the ROS content in salt-stressed plants to that level found in control plants or even less, resulting in a drastic reduction in the lipid peroxidation levels. This could be attributed to SPE boosting non-enzymatic antioxidant activity pathways as well as enzymatic ones [36]. The content of natural antioxidants in SPE (Table 1), including polyphenols, plays an important role in reducing lipid peroxidation. Compounds recognized as excellent anti-oxidants are found in algae comprising certain pigments such as carotenoids, astaxanthin, fucoxanthin, and polyphenols such as phenolic acids, flavonoids, and tannins [52].
Apart from antioxidant compounds, plants use enzymatic systems in order to deal with oxidative stress. Superoxide dismutase (SOD), glutathione peroxidase (GPx), glutathione-S-transferase (GST) and glutathione reductase (GR) create an intricate network to reduce, absorb, and scavenge ROS detoxification, allowing cellular survival [53]. It is well-known that SOD is the first line of defense against damage caused by ROS because it catalyzes the dismutation of O2− into oxygen and H2O2 [54]. A noteworthy rise in SOD activity was observed in the present research in reaction to salt stress. Nevertheless, it was overcome by plants treated with sprayed SPE mainly at 5%, which, in addition to neutralizing the harmful salt stress effect, further increased SOD activity. As expected, in SPE-treated plants under stress, the increase in SOD activity was accompanied by a decrease in O2- content (Figure 5 and Figure 6). Hence, the H2O2 generated by SOD could be used for other types of procedures like cell wall reinforcement [55] as well as a signaling molecule that activates plant defense systems and get detoxified by other antioxidant enzymes such as peroxidases and catalases. These results suggest that SPE could help common bean plants to defend themselves against salinity-induced oxidative stress.
In the present study, common bean leaves exposed to salt stress presented an increase in GPx and GR activities, especially when plants were sprayed with SPE rather than irrigated (Figure 4). GPx catalyzes the reduction of the generated H2O2 by SOD to water and oxygen, using GSH directly as a reducing agent [53]. GSSG reduction through glutathione reductase (GR) facilitates the regeneration of GSH. By employing NADPH as a reducing agent, the GR enzyme maintains a balanced ratio of GSSG/GSH, which is essential for preserving the oxidative balance in plant cells [56].
GST activity also plays an essential role for ROS detoxification [57]. Plants under salinity have higher GSH content which is linked to higher GST and GPx activities (Figure 6; [57]). Additionally, SPE treatments have been shown to reduce the oxidative stress response in common bean plants, especially at 5% of SPE (Figure 4c), suggesting it provides tolerance to salt toxicity. In sum, SPE reduced the production of ROS and minimized oxidative damage caused by salinity by modulating the antioxidant defense system, allowing common beans to develop normally even in salt stressed environments. In line with these results, Latique et al. [58] examined liquid seaweed extract effects on mitigating salt stress on wheat plants, resulting in increased growth, antioxidant enzymes, and boosting the overall health of crops subject to salt stress.
The present investigation highlighted how salt stress influences carbon and nitrogen metabolism in common bean plant leaves with and without SPE treatment. Salt stress caused a striking drop in PEPC activity (Figure 7), which is known as a key enzyme in carbon metabolism. Considering that there is plenty of evidence showing adverse salt toxicity effects on PEPC performance [59], this enzyme plays diverse functions in plant cells, by exchanging phosphoenolpyruvate (PEP) into oxaloacetate and inorganic phosphate (Pi) in the presence of HCO3 [60]. This enzyme has the potential to support the replenishment of intermediates necessary for nitrogen assimilation and biosynthesis in the tricarboxylic acid cycle (TCA). This is necessary not only for amino acids synthesis, but also for carbohydrates and nucleic acid production [59,28]. Furthermore, PEPC activity showed a significant increase in SPE treatment, particularly by spraying application (Figure 7a), suggesting a potential alleviating effect on demand for carbon skeletons required for nitrogen assimilation in common bean plants under salt stress conditions.
GDH activity rose while GS activity was significantly suppressed under salt stress (Figure 7b and c). As an essential enzyme for the metabolism of nitrogen, GS produces glutamine by condensing glutamate and ammonia [61]. Moreover, GDH primarily functions as a bidirectional enzyme reaction, catalyzing the assimilation of ammonium to glutamate and the deamination of glutamate to 2-oxoglutarate and ammonium [31]. The increasing GDH activity and GS/GOGAT cycle decreasing capacity under salt stress have been previously reported by various studies [61,62,63]. The results presented here may indicate that ammonium assimilation under salinity stress might largely depend on GDH pathway. As reported, SPE increased GDH and GS activities, much more by spraying than by irrigation, highlighting the ability of SPE to stimulate ammonium assimilation in common bean plants. AAT activity plays a crucial role in the synthesis of amino acids from glutamate, making it a necessary enzyme for nitrogen metabolism in plants [65]. Salt stress markedly increased AAT activity, compared to control plants (Figure 7d); however, when stressed plants were treated with SPE by irrigation, AAT activity was reduced compared to SPE-sprayed plants, which showed a significant increase. AAT is an aminotransferase, oxaloacetate and glutamate are produced when AAT facilitates the reversible transfer the amino group of aspartate to α-ketoglutarate [65]. AATs have been shown to be crucial in several physiological functions in plants, involving the carbon skeleton recycling that occurs when roots absorb ammonia. The current observations of elevated AAT activity in SPE-treated plants is consistent with the rise in the amino acid pool. The ammonia that tends to build up in salty environments is detoxified in large part by the keto acids and certain amino acids [66]. Considering all these results SPE treatment could be assessed as an excellent potential to induce resistance in common bean plants against salt stress toxicity due to its potential for modulating carbon and nitrogen metabolism, antioxidant system, morphological parameters and content in osmolytes.
Overall, the PCA results reveal that the treatments applied have a significant effect on the morpho-physiological biochemical (Figure 8) and enzymatical traits (Figure 9) in common bean plants. The positive association between the SSPE2% and SSPE5% treatments and the measured parameters under salt stress suggests that these treatments are effective in mitigating the negative effects of salt stress. In contrast, the relationship between SSPE1% and the measured parameters under salt stress indicates that this treatment may not be as effective as the SSPE2% and SSPE5% in these conditions.
Therefore, our findings highlight the need for a precise selection of SPE concentrations and an effective application method for agricultural use.
5. Conclusions
This study highlights the effectiveness of Saccorhiza polyschides extract in alleviating salt stress in common bean plants. Salt stress impaired growth and physiological functions, reducing chlorophyll content, photosynthetic activity, and essential metabolic processes. SPE treatment significantly countered these effects, enhancing osmolytes accumulation and plant growth, chlorophyll content as well as antioxidant defenses. Key findings include increased activity of nitrogen metabolism enzymes and antioxidant enzymes, and decreased ROS and lipid peroxidation levels with SPE application.
The SPE application by spraying in leaves, especially at 2% and 5%, seems to be a potent biostimulant able to enhance common bean resilience to salt stress, making SPE a sustainable solution for improving crop productivity in saline environments.
Author Contributions
Conceptualization, Nada. Nhhala; methodology, Nada. Nhhala, Anass. Kchikich and Amr Kchikich; software, Nada. Nhhala, Amr. Kchikich; validation, Mohamed. Nhiri, Penélope García-Angulo; formal analysis, Mohamed. Nhiri, Penélope García-Angulo, and Nada. Nhhala; research, Mohamed. Nhiri, Penélope García-Angulo, Salma Latique and Nada. Nhhala; writing—Nada Nhhala; writing—review and editing, Penélope García-Angulo, Salma Latique, Mohamed. Nhiri, Nada Nhhala, and Anass Kchikich; supervision, Mohamed. Nhiri, Penélope García-Angulo. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data used to support the finding of this study are included in the article.
Acknowledgments
We thank Rafael Calvo for the assistance with the English manuscript correction.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Sofy, M.R.; Elhawat, N.; Tarek Alshaal Glycine Betaine Counters Salinity Stress by Maintaining High K+/Na+ Ratio and Antioxidant Defense via Limiting Na+ Uptake in Common Bean (Phaseolus Vulgaris L.). Ecotoxicol. Environ. Saf. 2020, 200. [CrossRef]
- Wekesa, C.; Asudi, G.O.; Okoth, P.; Reichelt, M.; Muoma, J.O.; Furch, A.C.U.; Oelmüller, R. Rhizobia Contribute to Salinity Tolerance in Common Beans (Phaseolus Vulgaris L.). Cells 2022, 11. [CrossRef]
- Ehtaiwesh, A.; 2022, U. The Effect of Salinity on Nutrient Availability and Uptake in Crop Plants. jas.sabu.edu.lyAF EhtaiweshScientific J. Appl. Sci. Sabratha Univ. 2022•jas.sabu.edu.ly.
- Bouzroud, S.; Henkrar, F.; Fahr, M.; Smouni, A. Salt Stress Responses and Alleviation Strategies in Legumes: A Review of the Current Knowledge. 3 Biotech 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Kronzucker, H.J.; Coskun, D.; Schulze, L.M.; Wong, J.R.; Britto, D.T. Sodium as Nutrient and Toxicant. Plant Soil 2013, 369, 1–23. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Martínez, V.; Benito, B.; Rubio, F. Comparison between Arabidopsis and Rice for Main Pathways of K+ and Na+ Uptake by Roots. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- PLETT, D.C.; Plant, I.M.-; environment, cell &; 2010, Na+ Transport in Glycophytic Plants: What We Know and Would like to Know. Wiley Online Libr. CRAIG PLETT, IS MøllerPlant, cell Environ. 2010•Wiley Online Libr. 2009, 33, 612–626. [CrossRef]
- Cabot, C.; Sibole, J. V.; Barceló, J.; Poschenrieder, C. Lessons from Crop Plants Struggling with Salinity. Plant Sci. 2014, 226, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt Tolerance and Salinity Effects on Plants: A Review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Höhn, A.; Jung, T.; Grune, T. Pathophysiological Importance of Aggregated Damaged Proteins. Free Radic. Biol. Med. 2014, 71, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Yang, Y. How Plants Tolerate Salt Stress. Curr. Issues Mol. Biol. 2023, 45, 5914–5934. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.S.A.; Ashraf, M.; Ali, Q. Soil Salinity as a Selection Pressure Is a Key Determinant for the Evolution of Salt Tolerance in Blue Panicgrass (Panicum Antidotale Retz.). Flora Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 37–45. [Google Scholar] [CrossRef]
- Jatav, K.S.; Agarwal, R.M.; Tomar, N.S.; Tyagi, S.R. Nitrogen Metabolism, Growth and Yield Responses of Wheat (Triticum Aestivum L.) to Restricted Water Supply and Varying Potassium Treatments. J Indian Bot Soc 2014, 93, 177–189. [Google Scholar]
- Francesca, S.; Arena, C.; Mele, B.H.; Schettini, C.; Ambrosino, P.; Barone, A.; Rigano, M.M. The Use of a Plant-Based Biostimulant Improves Plant Performances and Fruit Quality in Tomato Plants Grown at Elevated Temperatures. mdpi.comS Fr. C Arena, B Hay Mele, C Schettini, P Ambrosino, A Barone, MM RiganoAgronomy, 2020•mdpi.com 2020, 10. [CrossRef]
- Soares, C.; Švarc-Gajić, J.; Oliva-Teles, M.T.; Pinto, E.; Nastić, N.; Savić, S.; Almeida, A.; Delerue-Matos, C. Mineral Composition of Subcritical Water Extracts of Saccorhiza Polyschides, a Brown Seaweed Used as Fertilizer in the North of Portugal. J. Mar. Sci. Eng. 2020, 8, 244. [Google Scholar] [CrossRef]
- Cardoso, C.; Almeida, J.; Coelho, I.; Delgado, I.; Gomes, R.; Quintã, R.; Bandarra, N.M.; Afonso, C. Farming a Wild Seaweed and Changes to Its Composition, Bioactivity, and Bioaccessibility: The Saccorhiza Polyschides Case Study. Aquaculture 2023, 566, 739217. [Google Scholar] [CrossRef]
- Senousy, H.H.; Hamoud, Y.A.; Abu-Elsaoud, A.M.; Mahmoud Al zoubi, O.; Abdelbaky, N.F.; Zia-ur-Rehman, M.; Usman, M.; Soliman, M.H. Algal Bio-Stimulants Enhance Salt Tolerance in Common Bean: Dissecting Morphological, Physiological, and Genetic Mechanisms for Stress Adaptation. Plants 2023, 12, 3714. [Google Scholar] [CrossRef] [PubMed]
- Mamede, M.; Cotas, J.; Pereira, L.; Bahcevandziev, K. Seaweed Polysaccharides as Potential Biostimulants in Turnip Greens Production. Hortic. 2024, 10, 130. [Google Scholar] [CrossRef]
- Bouchmaa, N.; Mrid, R. Ben; Kabach, I.; Zouaoui, Z.; Karrouchi, K.; Chtibi, H.; Zyad, A.; Cacciola, F.; Nhiri, M. Beta Vulgaris Subsp. Maritima: A Valuable Food with High Added Health Benefits. Appl. Sci. 2022, 12, 1866. [Google Scholar] [CrossRef]
- Bouchmaa, N.; Ben Mrid, R.; Boukharsa, Y.; Nhiri, M.; Ait Mouse, H.; Taoufik, J.; Ansar, M.; Zyad, A. Cytotoxicity of New Pyridazin-3(2H)-One Derivatives Orchestrating Oxidative Stress in Human Triple-Negative Breast Cancer (MDA-MB-468). Arch. Pharm. (Weinheim). 2018, 351, 1800128. [Google Scholar] [CrossRef] [PubMed]
- Kubiś, J. Exogenous Spermidine Differentially Alters Activities of Some Scavenging System Enzymes, H2O2 and Superoxide Radical Levels in Water-Stressed Cucumber Leaves. J. Plant Physiol. 2008, 165, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Mrid, R. Ben; Bouchmaa, N.; Bouargalne, Y.; Ramdan, B.; Karrouchi, K.; Kabach, I.; Karbane, M. El; Idir, A.; Zyad, A.; Nhiri, M. Phytochemical Characterization, Antioxidant and In Vitro Cytotoxic Activity Evaluation of Juniperus Oxycedrus Subsp. Oxycedrus Needles and Berries. Molecules 2019, 24, 502. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Ennoury, A.; BenMrid, R.; Nhhala, N.; Roussi, Z.; Latique, S.; Zouaoui, Z.; Nhiri, M. River’s Ulva Intestinalis Extract Protects Common Bean Plants (Phaseolus Vulgaris L.) against Salt Stress. South African J. Bot. 2022, 150, 334–341. [Google Scholar] [CrossRef]
- Sun, S.W.; Lin, Y.C.; Weng, Y.M.; Chen, M.J. Efficiency Improvements on Ninhydrin Method for Amino Acid Quantification. J. Food Compos. Anal. 2006, 19, 112–117. [Google Scholar] [CrossRef]
- YEMM, E.W.; WILLIS, A.J. The Estimation of Carbohydrates in Plant Extracts by Anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, C.; Fridovich, I. Superoxide Dismutase: Improved Assays and an Assay Applicable to Acrylamide Gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Latique, S.; Mrid, R. Ben; Kabach, I.; Kchikich, A.; Sammama, H.; Yasri, A.; Nhiri, M.; El Kaoua, M.; Douira, A.; Selmaoui, K. Foliar Application of Ulva Rigida Water Extracts Improves Salinity Tolerance in Wheat (Triticum Durum L.). Agronomy 2021, 11, 265. [Google Scholar] [CrossRef]
- El Omari, R.; Ben Mrid, R.; Amakran, A.; Nhiri, M. Effect of Fungicide (Maneb) on Antioxydant System and Carbon Assimilation in Leaves of Sorghum Plants. Russ. J. Plant Physiol. 2018, 65, 237–243. [Google Scholar] [CrossRef]
- Kchikich, A.; El Omari, R.; Kabach, I.; Yasri, A.; Nhiri, M.; Ben Mrid, R. Effect of Arbuscular Mycorrhizal Fungus and the γ-Aminobutyric Acid Treatment in Nitrate Assimilation under Nitrogen Deficiency in Sorghum Plant. Russ. J. Plant Physiol. 2021, 68, 901–908. [Google Scholar] [CrossRef]
- Ben MRid, R.; El Omari, R.; BOuaRgalne, Y.; El Mourabit, N.; Nhiri, M. Activities of Carbon and Nitrogen Metabolism Enzymes during Germinating Sorghum Seeds and Early Seedlings Growth. Cereal Res. Commun. 2017, 45, 587–597. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing Sustainability by Improving Plant Salt Tolerance through Macro-and Micro-Algal Biostimulants. Biology (Basel). 2020, 9, 1–21. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Potential Biochemical Indicators of Salinity Tolerance in Plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing Sustainability by Improving Plant Salt Tolerance through Macro-and Micro-Algal Biostimulants. Biology (Basel). 2020, 9, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Stasio, E. Di; James, M.; Oosten, V.; Silletti, S.; Raimondi, G.; Dell’aversana, E.; Carillo, P.; Maggio, A. Ascophyllum Nodosum-Based Algal Extracts Act as Enhancers of Growth, Fruit Quality, and Adaptation to Stress in Salinized Tomato Plants. SpringerE Di Stasio, MJ Van Oosten, S Silletti, G Raimondi, E Dell’Aversana, P Carillo, A MaggioJournal Appl. Phycol. 2018•Springer 2018, 30, 2675–2686. [CrossRef]
- Jithesh, M.N.; Pushp, ·; Shukla, S.; Kant, · P; Joshi, J.; Critchley, A.T.; Prithiviraj, · B Physiological and Transcriptomics Analyses Reveal That Ascophyllum Nodosum Extracts Induce Salinity Tolerance in Arabidopsis by Regulating the Expression Of. SpringerMN Jithesh, PS Shukla, P Kant, J Joshi, Critchley, B PrithivirajJournal plant growth Regul. 2019•Springer 2019, 38, 463–478. [CrossRef]
- Carmody, N.; Goñi, O.; Łangowski, Ł.; O’Connell, S. Ascophyllum Nodosum Extract Biostimulant Processing and Its Impact on Enhancing Heat Stress Tolerance During Tomato Fruit Set. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Stirk, W.A.; Rengasamy, K.R.R.; Kulkarni, M.G.; van Staden, J. Plant Biostimulants from Seaweed: An Overview. Chem. Biol. Plant Biostimulants 2020, 33–55. [Google Scholar] [CrossRef]
- Hussein, M.H.; Eltanahy, E.; Al Bakry, A.F.; Elsafty, N.; Elshamy, M.M. Seaweed Extracts as Prospective Plant Growth Bio-Stimulant and Salinity Stress Alleviator for Vigna Sinensis and Zea Mays. J. Appl. Phycol. 2021, 33, 1273–1291. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Jagendorf, A.; science, J.Z.-C.; 2005, Understanding and Improving Salt Tolerance in Plants. Wiley Online Libr. Chinnusamy, A Jagendorf, JK ZhuCrop Sci. 2005•Wiley Online Libr. 2005, 45, 437–448. [CrossRef]
- Negrão, S.; Schmöckel, S.; botany, M.T.-A. of; 2017, Evaluating Physiological Responses of Plants to Salinity Stress. Acad. Negrão, SM Schmöckel, M TesterAnnals Bot. 2017•academic.oup.com. [CrossRef]
- Rouphael, Y.; De Micco, V.; Arena, C.; Raimondi, G.; Colla, G.; De Pascale, S. Effect of Ecklonia Maxima Seaweed Extract on Yield, Mineral Composition, Gas Exchange, and Leaf Anatomy of Zucchini Squash Grown under Saline Conditions. SpringerY Rouphael, V Micco, C Arena, G Raimondi, G Colla, S PascaleJournal Appl. Phycol. 2017•Springer 2017, 29, 459–470. [CrossRef]
- Ibrahim, W.M.; Ali, R.M.; Hemida, K.A.; Sayed, M.A. Role of Ulva Lactuca Extract in Alleviation of Salinity Stress on Wheat Seedlings. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef]
- Wally, O.S.D.; Critchley, A.T.; Hiltz, D.; Craigie, J.S.; Han, X.; Zaharia, L.I.; Abrams, S.R.; Prithiviraj, B. Regulation of Phytohormone Biosynthesis and Accumulation in Arabidopsis Following Treatment with Commercial Extract from the Marine Macroalga Ascophyllum Nodosum. J. Plant Growth Regul. 2013, 32, 324–339. [Google Scholar] [CrossRef]
- HM, A.; Shaddad, M.; & N.B.-J. of S.P.; 2010, The Role of Amino Acids in Improvement in Salt Tolerance of Crop Plants. cyberleninka.ruAES HM, MAK Shaddad, N BarakatJournal Stress Physiol. Biochem. 2010•cyberleninka.ru 2010, 6, 25–37.
- Battacharyya, D.D.; Babgohari, M. Seaweed Extracts as Biostimulants in Horticulture. 2015.
- Singh Kesawat, M.; Satheesh, N.; Singh Kherawat, B.; Kumar, A.; Kim, H.-U.; Chung, S.-M.; Kumar, M.; Vigyan Kendra, K.; Keshwanand, S. Regulation of Reactive Oxygen Species during Salt Stress in Plants and Their Crosstalk with Other Signaling Molecules—Current Perspectives and Future Directions. mdpi.com MS Kesawat, N Satheesh, BS Kherawat, A Kumar, HU Kim, SM Chung, M Kumar Plants, 2023•mdpi.com 2023. [CrossRef]
- Nabil-Adam, A.; Shreadah, M.A.; Abd El-Moneam, N.M.; El-Assar, S.A. Marine Algae of the Genus Gracilaria as Multi Products Source for Different Biotechnological and Medical Applications. Recent Pat. Biotechnol. 2020, 14, 203–228. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent Developments in Enzymatic Antioxidant Defence Mechanism in Plants with Special Reference to Abiotic Stress. Biology (Basel). 2021, 10.
- Ighodaro, O.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alexandria J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- De la Rubia, A.G.; Centeno, M.L.; Moreno-González, V.; De Castro, M.; García-Angulo, P. Perception and First Defense Responses Against Pseudomonas syringae Pv. phaseolicola in Phaseolus vulgaris: Identification of Wall-Associated Kinase Receptors. Phytopathology 2021, 111, 2332–2342. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Loscalzo, J. Metabolic Responses to Reductive Stress. Antioxidants Redox Signal. 2020, 32, 1330–1347. [Google Scholar] [CrossRef] [PubMed]
- Gapińska, M.; Skłodowska, M.; Gabara, B. Effect of Short- and Long-Term Salinity on the Activities of Antioxidative Enzymes and Lipid Peroxidation in Tomato Roots. Acta Physiol. Plant. 2008, 30, 11–18. [Google Scholar] [CrossRef]
- Latique, S.; Mohamed Aymen, E.; Halima, C.; Chérif, H.; Mimoun, E.K. Alleviation of Salt Stress in Durum Wheat (Triticum Durum L.) Seedlings Through the Application of Liquid Seaweed Extracts of Fucus Spiralis. Commun. Soil Sci. Plant Anal. 2017, 48, 2582–2593. [Google Scholar] [CrossRef]
- Sharma, S.; Joshi, J.; Kataria, S.; Verma, S.K.; Chatterjee, S.; Jain, M.; Pathak, K.; Rastogi, A.; Brestic, M. Regulation of the Calvin Cycle under Abiotic Stresses: An Overview. In Plant Life under Changing Environment: Responses and Management; 2020; pp. 681–717 ISBN 9780128182048.
- O’Leary, B.; Park, J.; Plaxton, W.C. The Remarkable Diversity of Plant PEPC (Phosphoenolpyruvate Carboxylase): Recent Insights into the Physiological Functions and Post-Translational Controls of Non-Photosynthetic PEPCs. Biochem. J. 2011, 436, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Betti, M.; García-Calderón, M.; Pérez-Delgado, C.M.; Credali, A.; Estivill, G.; Galván, F.; Vega, J.M.; Márquez, A.J. Glutamine Synthetase in Legumes: Recent Advances in Enzyme Structure and Functional Genomics. Int. J. Mol. Sci. 2012, 13, 7994–8024. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.G.; Shah, K.; Dubey, R.S. Salinity Induced Behavioural Changes in Malate Dehydrogenase and Glutamate Dehydrogenase Activities in Rice Seedlings of Differing Salt Tolerance. Plant Sci. 2000, 156, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Farhangi-Abriz, S.; Torabian, S. Biochar Improved Nodulation and Nitrogen Metabolism of Soybean under Salt Stress. Symbiosis 2018, 74, 215–223. [Google Scholar] [CrossRef]
- Zhang, X.; He, P.; Guo, R.; Huang, K.; Huang, X. Effects of Salt Stress on Root Morphology, Carbon and Nitrogen Metabolism, and Yield of Tartary Buckwheat. Sci. Rep. 2023, 13. [Google Scholar] [CrossRef]
- Zhou, Y.; Cai, H.; Xiao, J.; Li, X.; Zhang, Q.; Lian, X. Over-Expression of Aspartate Aminotransferase Genes in Rice Resulted in Altered Nitrogen Metabolism and Increased Amino Acid Content in Seeds. Theor. Appl. Genet. 2009, 118, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Ramanjulu, S.; Veeranjaneyulu, K.; Sudhakar, C. Short-Term Shifts in Nitrogen Metabolism in Mulberry Morus Alba under Salt Shock. Phytochemistry 1994, 37, 991–995. [Google Scholar] [CrossRef]
Figure 1.
Overview of experimental design used to test the effect of different SPE applications (1%, 2% and 5%) in common bean plants after 2 months growing under non salinity stress (NSS; water) and salinity stress (SS; 68.4 mM of NaCl) culture conditions. Two different SPE applications were used: by spraying or mixed in the irrigation water. A summary of main affected parameters under these different conditions is also shown in the lower part (Illustration made using Biorender).
Figure 1.
Overview of experimental design used to test the effect of different SPE applications (1%, 2% and 5%) in common bean plants after 2 months growing under non salinity stress (NSS; water) and salinity stress (SS; 68.4 mM of NaCl) culture conditions. Two different SPE applications were used: by spraying or mixed in the irrigation water. A summary of main affected parameters under these different conditions is also shown in the lower part (Illustration made using Biorender).
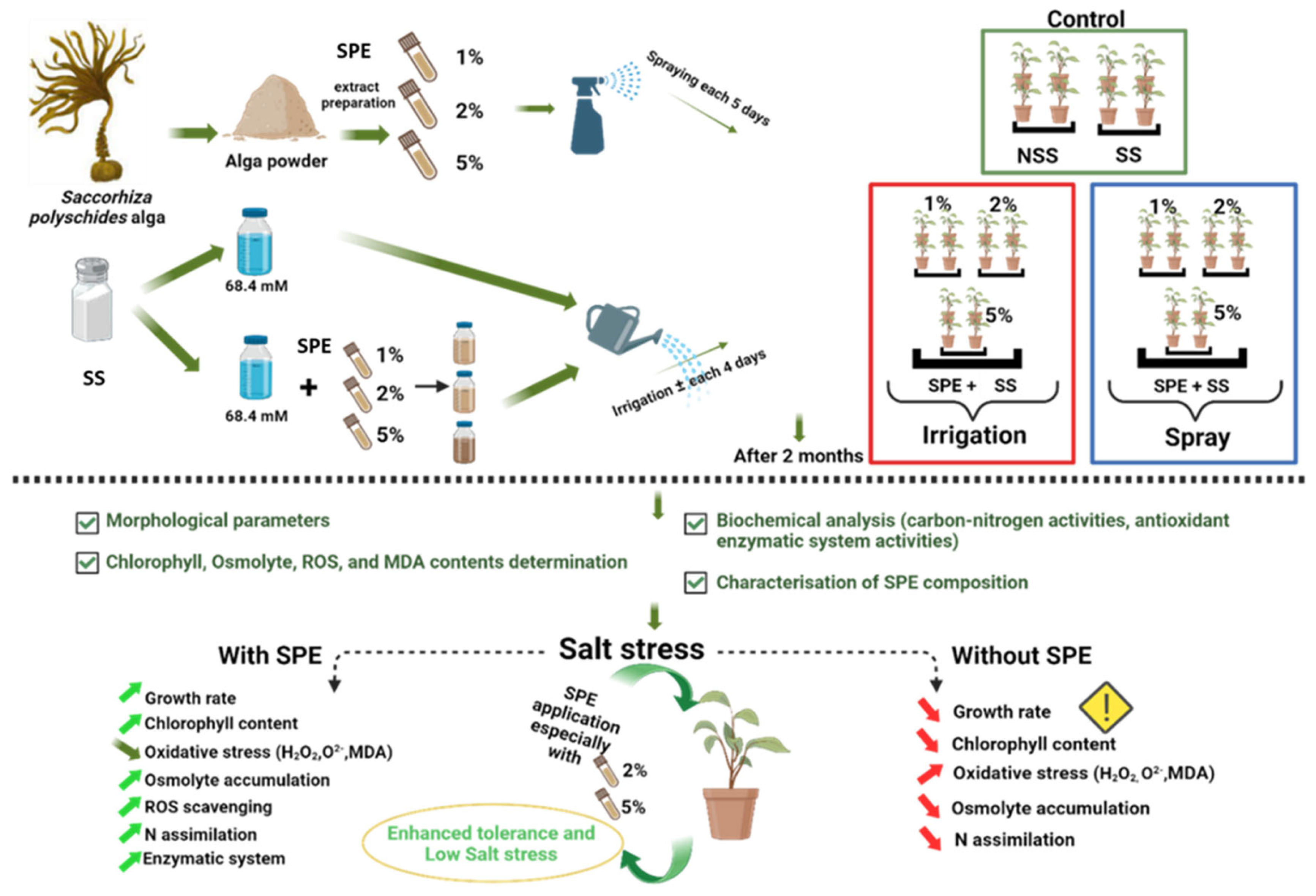
Figure 2.
Biostimulant effect of 1%, 2% and 5% SPE applied by irrigation method (SPE Irrigation + SS), or spraying method (SPE Spraying + SS) on common bean plants growth parameters: shoot length (a), fresh weight (b) and leaf diameter (c), under salt stress (68.4 mM of NaCl). Results of control plants neither with biostimulation nor salt stress (NSS), and under salt stress without biostimulation (SS) were also included for comparison. The value is the average ± standard deviation (SD) of three or four independent observations. Different letters indicate significant differences by Tukey’s test (p < 0.05).
Figure 2.
Biostimulant effect of 1%, 2% and 5% SPE applied by irrigation method (SPE Irrigation + SS), or spraying method (SPE Spraying + SS) on common bean plants growth parameters: shoot length (a), fresh weight (b) and leaf diameter (c), under salt stress (68.4 mM of NaCl). Results of control plants neither with biostimulation nor salt stress (NSS), and under salt stress without biostimulation (SS) were also included for comparison. The value is the average ± standard deviation (SD) of three or four independent observations. Different letters indicate significant differences by Tukey’s test (p < 0.05).
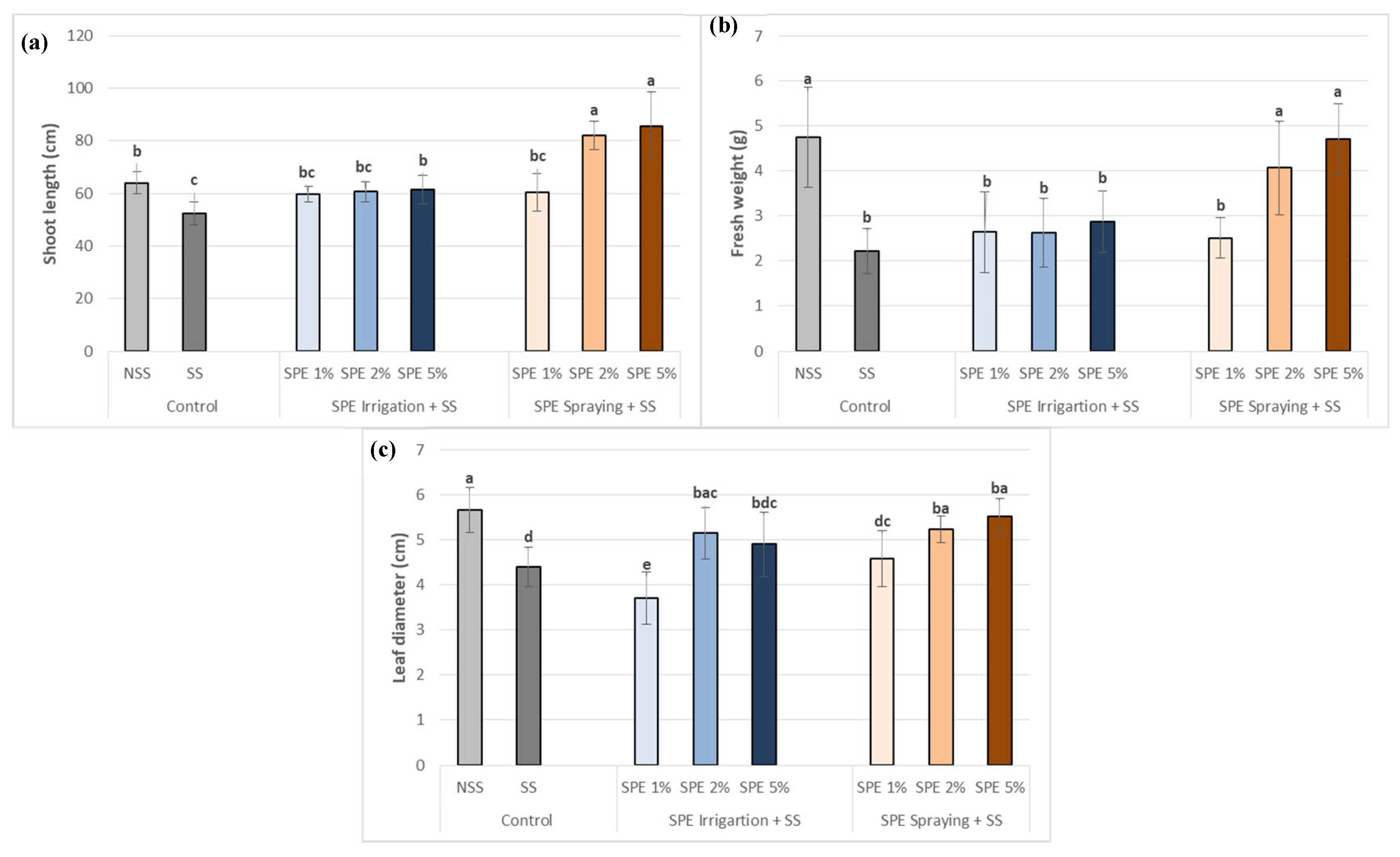
Figure 3.
Biostimulant effect of 1%, 2% and 5% SPE applied by irrigation method (SPE Irrigation + SS), or spraying method (SPE Spraying + SS) on total chlorophyll content of common bean plants under salt stress (68.4 mM of NaCl). Results of control plants neither with biostimulation nor salt stress (NSS), and under salt stress without biostimulation (SS) were also included for comparison. The value is the average ± standard deviation (SD) of three or four independent observations. Different letters indicate significant differences by Tukey’s test (p < 0.05).
Figure 3.
Biostimulant effect of 1%, 2% and 5% SPE applied by irrigation method (SPE Irrigation + SS), or spraying method (SPE Spraying + SS) on total chlorophyll content of common bean plants under salt stress (68.4 mM of NaCl). Results of control plants neither with biostimulation nor salt stress (NSS), and under salt stress without biostimulation (SS) were also included for comparison. The value is the average ± standard deviation (SD) of three or four independent observations. Different letters indicate significant differences by Tukey’s test (p < 0.05).
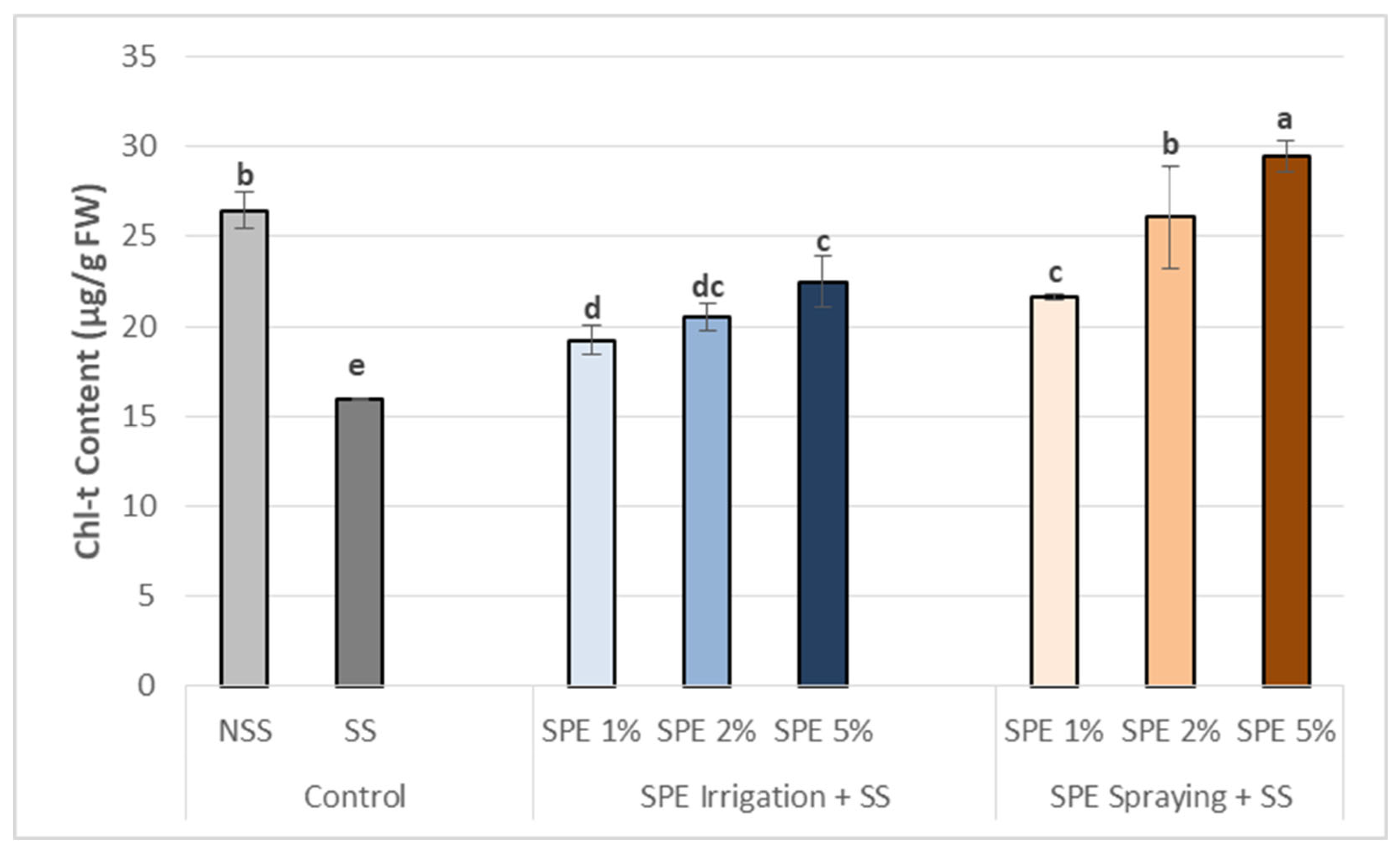
Figure 4.
Biostimulant effect of 1%, 2% and 5% SPE applied by irrigation method (SPE Irrigation + SS), or spraying method (SPE Spraying + SS) on common bean content of IAA (a), soluble sugars (b) and amino acid (AA) (c), under salt stress (68.4 mM of NaCl). Results of control plants neither with biostimulation nor salt stress (NSS), and under salt stress without biostimulation (SS) were also included for comparison. The value is the average ± standard deviation (SD) of three or four independent observations. Different letters indicate significant differences by Tukey’s test (p < 0.05).
Figure 4.
Biostimulant effect of 1%, 2% and 5% SPE applied by irrigation method (SPE Irrigation + SS), or spraying method (SPE Spraying + SS) on common bean content of IAA (a), soluble sugars (b) and amino acid (AA) (c), under salt stress (68.4 mM of NaCl). Results of control plants neither with biostimulation nor salt stress (NSS), and under salt stress without biostimulation (SS) were also included for comparison. The value is the average ± standard deviation (SD) of three or four independent observations. Different letters indicate significant differences by Tukey’s test (p < 0.05).
Figure 5.
Biostimulant effect of 1%, 2% and 5% SPE applied by irrigation method (SPE Irrigation + SS), or spraying method (SPE Spraying + SS) on common bean lipid peroxidation (MDA) (a), superoxide O2- (b) and hydrogen peroxide H2O2 (c), under salt stress (68.4 mM of NaCl). Results of control plants neither with biostimulation nor salt stress (NSS), and under salt stress without biostimulation (SS) were also included for comparison. The value is the average ± standard deviation (SD) of three or four independent observations. Different letters indicate significant differences by Tukey’s test (p < 0.05).
Figure 5.
Biostimulant effect of 1%, 2% and 5% SPE applied by irrigation method (SPE Irrigation + SS), or spraying method (SPE Spraying + SS) on common bean lipid peroxidation (MDA) (a), superoxide O2- (b) and hydrogen peroxide H2O2 (c), under salt stress (68.4 mM of NaCl). Results of control plants neither with biostimulation nor salt stress (NSS), and under salt stress without biostimulation (SS) were also included for comparison. The value is the average ± standard deviation (SD) of three or four independent observations. Different letters indicate significant differences by Tukey’s test (p < 0.05).
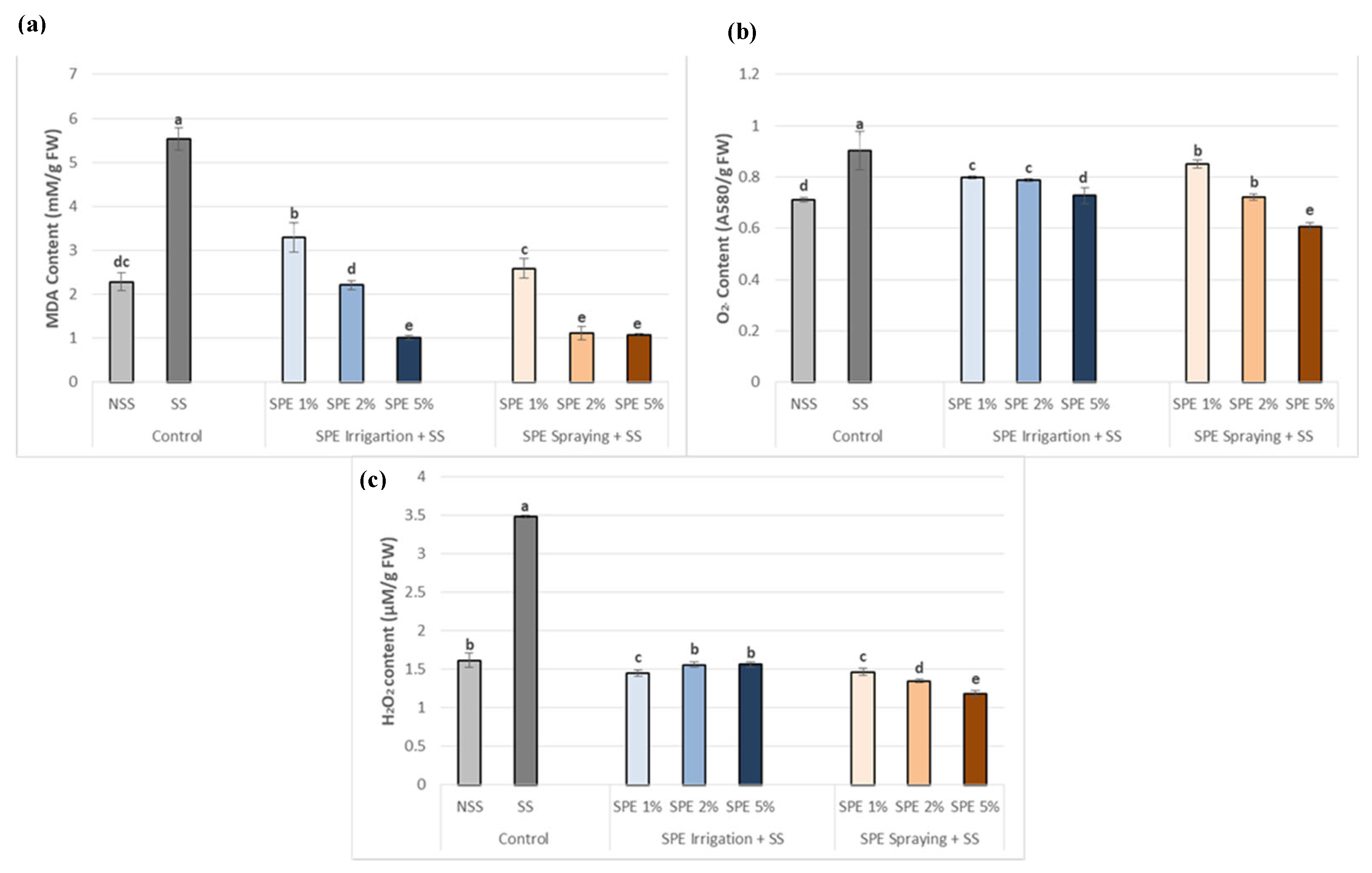
Figure 6.
Biostimulant effect of 1%, 2% and 5% SPE applied by irrigation method (SPE Irrigation + SS), or spraying method (SPE Spraying + SS) on common bean antioxidant enzyme activities; superoxide dismutase (SOD) (a), glutathione peroxidase (GPx) (b), glutathione-S-transferase (GST) (c), and glutathione reductase (GR) (d), under salt stress (68.4 mM of NaCl). Results of control plants neither with biostimulation nor salt stress (NSS), and under salt stress without biostimulation (SS) were also included for comparison. The value is the average ± standard deviation (SD) of three or four independent observations. Different letters indicate significant differences by Tukey’s test (p < 0.05).
Figure 6.
Biostimulant effect of 1%, 2% and 5% SPE applied by irrigation method (SPE Irrigation + SS), or spraying method (SPE Spraying + SS) on common bean antioxidant enzyme activities; superoxide dismutase (SOD) (a), glutathione peroxidase (GPx) (b), glutathione-S-transferase (GST) (c), and glutathione reductase (GR) (d), under salt stress (68.4 mM of NaCl). Results of control plants neither with biostimulation nor salt stress (NSS), and under salt stress without biostimulation (SS) were also included for comparison. The value is the average ± standard deviation (SD) of three or four independent observations. Different letters indicate significant differences by Tukey’s test (p < 0.05).
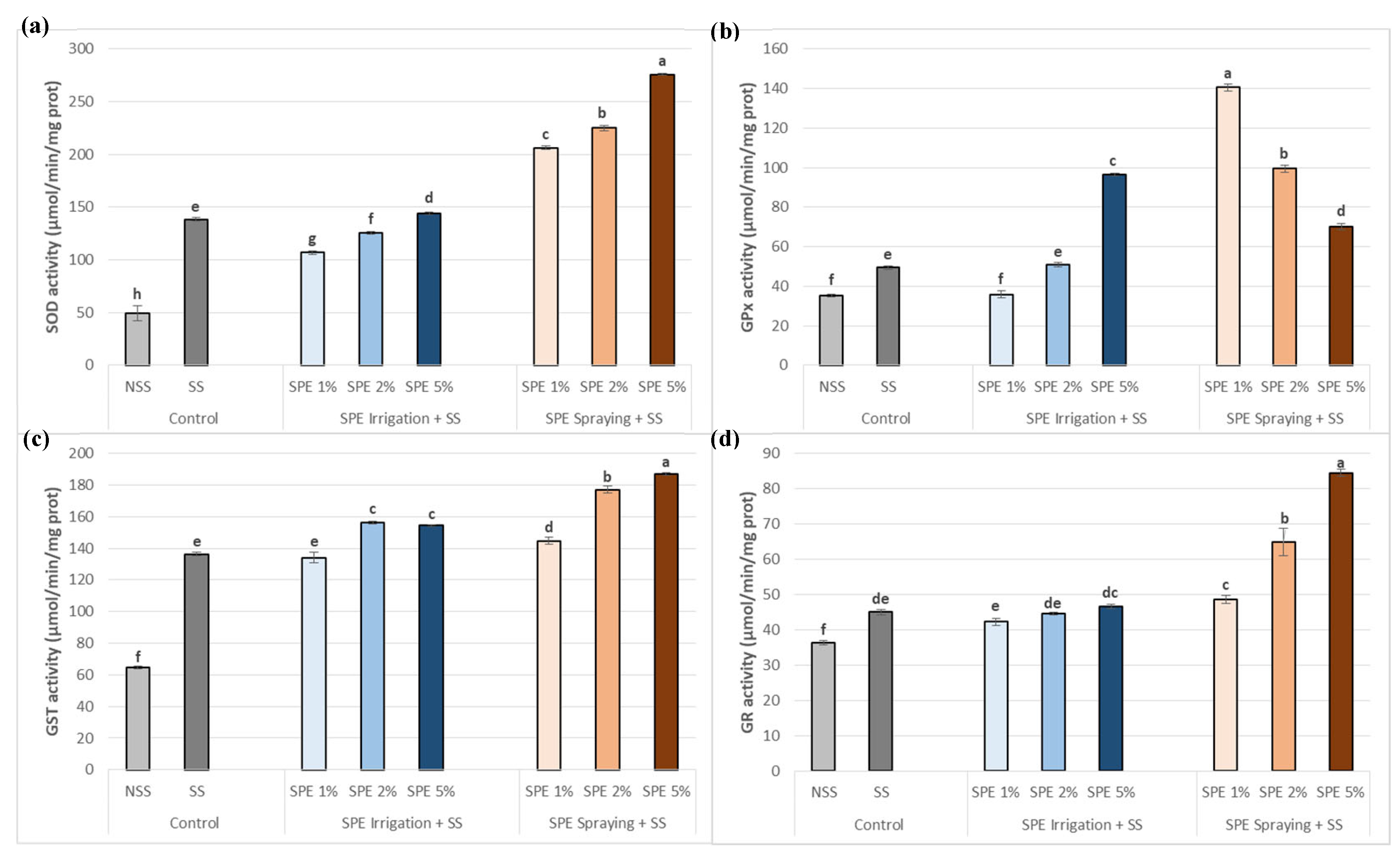
Figure 7.
Biostimulant effect of 1%, 2% and 5% SPE applied by irrigation method (SPE Irrigation + SS), or spraying method (SPE Spraying + SS) on common bean carbon-nitrogen enzyme activities: phosphoenolpyruvate carboxylase (PEPC) (a); glutamate dehydrogenase (GDH) (b), Glutamine synthase (GS) (c) and Aspartate Aminotransferase (AAT) (d), under salt stress (68.4 mM of NaCl). Results of control plants neither with biostimulation nor salt stress (NSS), and under salt stress without biostimulation (SS) were also included for comparison. The value is the average ± standard deviation (SD) of three or four independent observations. Different letters indicate significant differences by Tukey’s test (p < 0.05).
Figure 7.
Biostimulant effect of 1%, 2% and 5% SPE applied by irrigation method (SPE Irrigation + SS), or spraying method (SPE Spraying + SS) on common bean carbon-nitrogen enzyme activities: phosphoenolpyruvate carboxylase (PEPC) (a); glutamate dehydrogenase (GDH) (b), Glutamine synthase (GS) (c) and Aspartate Aminotransferase (AAT) (d), under salt stress (68.4 mM of NaCl). Results of control plants neither with biostimulation nor salt stress (NSS), and under salt stress without biostimulation (SS) were also included for comparison. The value is the average ± standard deviation (SD) of three or four independent observations. Different letters indicate significant differences by Tukey’s test (p < 0.05).
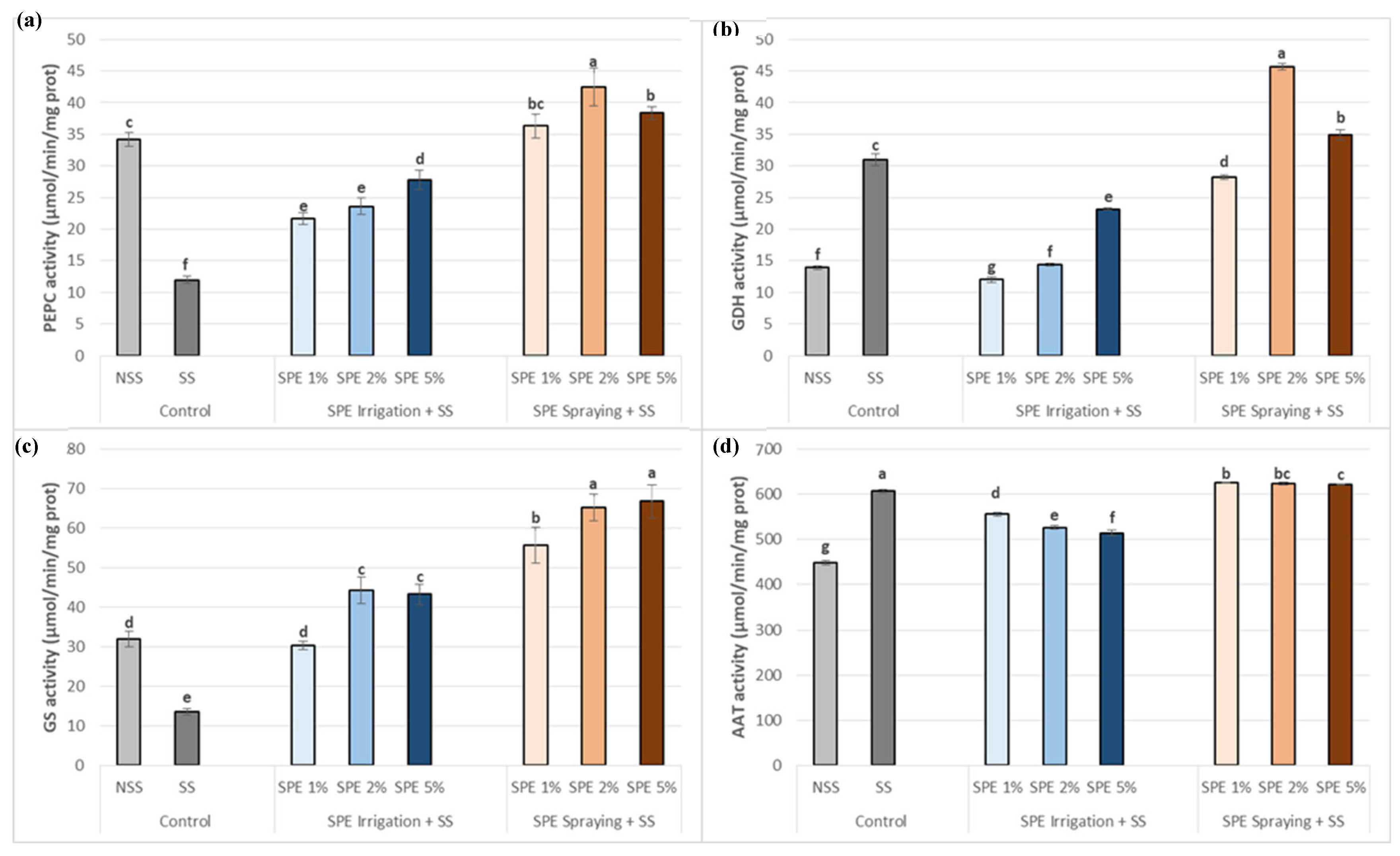
Figure 8.
Principal component analysis (PCA) of morpho-physiological and biochemical traits observed in common bean plants (A) and scatter plot of treatment (B). The variables included were: shoot length (SL), fresh weight (FW), leaf diameter (LD), total chlorophyll content. The total treatments (TT) included were: control (NSS: irrigation only with water), NaCl application (SS: irrigation only with 68.4 mM of NaCl), Saccorhiza polyschides extract (SPE) at different concentration (SPC): SPE 1%, SPE2% or SPE5% and 68.4 mM of NaCl applied through two application methods (AM): irrigation (I) and spraying (S).
Figure 8.
Principal component analysis (PCA) of morpho-physiological and biochemical traits observed in common bean plants (A) and scatter plot of treatment (B). The variables included were: shoot length (SL), fresh weight (FW), leaf diameter (LD), total chlorophyll content. The total treatments (TT) included were: control (NSS: irrigation only with water), NaCl application (SS: irrigation only with 68.4 mM of NaCl), Saccorhiza polyschides extract (SPE) at different concentration (SPC): SPE 1%, SPE2% or SPE5% and 68.4 mM of NaCl applied through two application methods (AM): irrigation (I) and spraying (S).
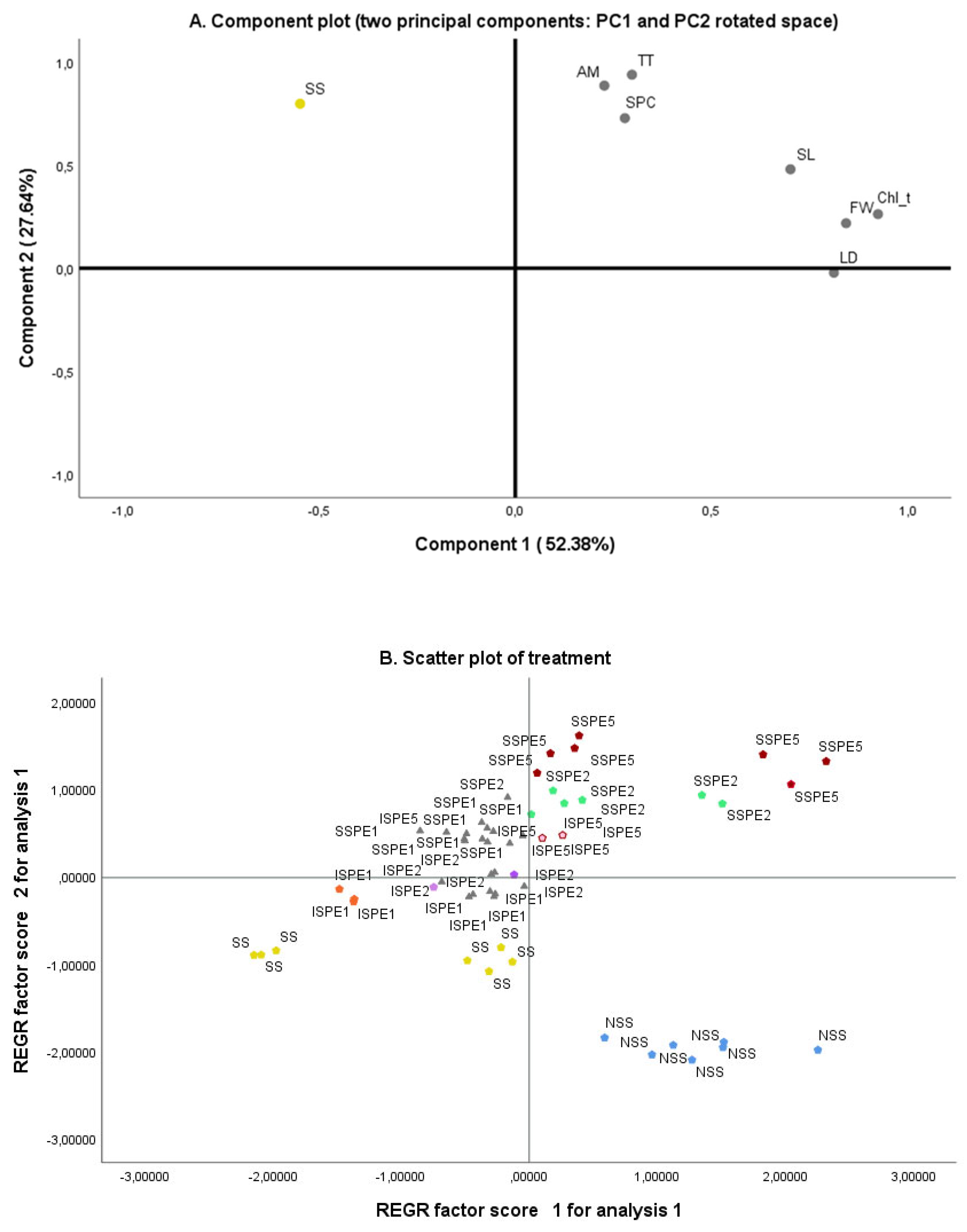
Figure 9.
Principal component analysis (PCA) of enzyme activities and some biochemical traits observed in common bean plants (A) and scatter plot of treatment (B). The enzymatic activities included were: superoxide dismutase (SOD), glutathione-s-transferase (GST), glutathione-peroxidase (GPx), phosphoenolpyruvate carboxylase (PEPC), aspartate aminotransferase (AAT), glutamate dehydrogenase (GDH), glutamine synthetase (GS). The content of indolacetic acid (IAA), amino acids, soluble sugars, malonaldehyde acid (MDA), H2O2 and O2- was also included. The treatments (TT) included were: control (NSS: irrigation only with water), NaCl (SS: irrigation only with 68.4 mM of NaCl), Saccorhiza polyschides extract (SPE) at different concentration (SPC): SPE 1%, SPE2% or SPE5% and 68.4 mM of NaCl applied through two methods (AM): irrigation (I) and spraying (S).
Figure 9.
Principal component analysis (PCA) of enzyme activities and some biochemical traits observed in common bean plants (A) and scatter plot of treatment (B). The enzymatic activities included were: superoxide dismutase (SOD), glutathione-s-transferase (GST), glutathione-peroxidase (GPx), phosphoenolpyruvate carboxylase (PEPC), aspartate aminotransferase (AAT), glutamate dehydrogenase (GDH), glutamine synthetase (GS). The content of indolacetic acid (IAA), amino acids, soluble sugars, malonaldehyde acid (MDA), H2O2 and O2- was also included. The treatments (TT) included were: control (NSS: irrigation only with water), NaCl (SS: irrigation only with 68.4 mM of NaCl), Saccorhiza polyschides extract (SPE) at different concentration (SPC): SPE 1%, SPE2% or SPE5% and 68.4 mM of NaCl applied through two methods (AM): irrigation (I) and spraying (S).
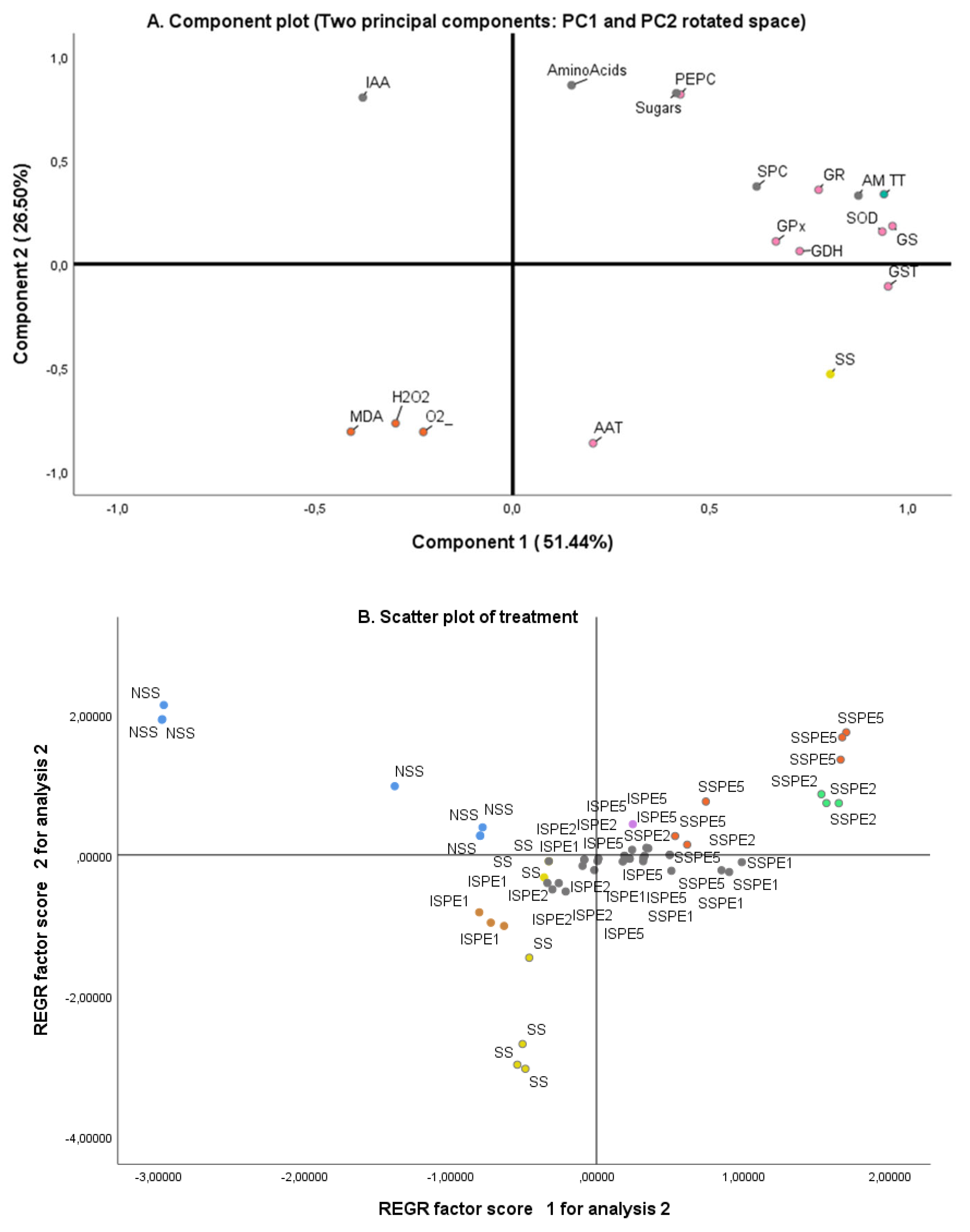

Table 1.
Polyphenol, flavonoid, indole acetic acid (IAA) and amino acid (AA) content of Saccorhiza polyschides water extract (SPE) at 100%. Values represent the mean of four replicates ± standard deviation (SD).
Table 1.
Polyphenol, flavonoid, indole acetic acid (IAA) and amino acid (AA) content of Saccorhiza polyschides water extract (SPE) at 100%. Values represent the mean of four replicates ± standard deviation (SD).
| Saccorhiza polyschides | Polyphenols | Flavonoids | IAA | AA | |
| (mg/g DW) | |||||
| 5.494 ± 0.026 | 1.327 ± 0.012 | 0.264 ± 0.004 | 0.353 ± 0.009 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



