
Submitted:
21 June 2024
Posted:
21 June 2024
You are already at the latest version
Abstract
Climate change has altered the frequency and severity of extreme weather, which can affect vegetation condition and habitat quality for wildlife. Declines in vegetation productivity during droughts and heatwaves can negatively impact animals that depend on vegetation for water and nutrition. The ability to detect such effects on habitat suitability can reveal refugia, which can be prioritised for protection to improve threatened species conservation. We used the normalised difference vegetation index (NDVI) to look at relationships between vegetation productivity and the presence of koalas (Phascolarctos cinereus) in potential habitat throughout much of their range. Using a large, long-term koala presence dataset, we tested the hypothesis that locations where koalas had been observed would exhibit higher NDVI values than a random, representative sample from the same vegetation group (i.e. woodlands, open forests or tall open forests). We also identified the minimum NDVI threshold at which koalas occurred across time for each vegetation group and compared these to the minimum NDVI values observed across potential koala habitat before and during the millennium drought; one of the worst recorded in Australia. Additionally, we investigated whether vegetation above the minimum NDVI thresholds was significantly closer to perennial water than unsuitable vegetation. We found that koalas tend to occur at locations exhibiting higher NDVI than average for all vegetation groups. We also found that 49% of all vegetation groups maintained a minimum NDVI above the koalas’ NDVI threshold before drought, equating to 190,227 km2, which declined to 166,746 km2 during drought (a 12% reduction). We also found that unsuitable vegetation tended to occur farther from perennial water than suitable vegetation for some vegetation groups. Areas that remained above the NDVI thresholds during the drought should be considered potential refugia for populations during an event of similar magnitude and could indicate future core habitat extent.
Keywords:
NDVI
; koala
; herbivore
; remote sensing
; forest
; Landsat
; drought
; habitat
1. Introduction
Climate change is impacting ecosystems around the world. Global mean land surface temperatures were 1.34 to 1.83°C higher in 2011-2020 than in 1850-1900 [1]. A common component of climate change is increased extreme climate events, such as heatwaves and droughts [2]. Such events can drive rapid change in vegetation and wildlife populations [3,4]. Declining productivity is a common response of vegetation to climate extremes, including drought, and this may negatively impact higher trophic levels [5,6]. For example, declining forage productivity can reduce habitat suitability for browsing herbivores via reduced nutrient, energy, and water availability, and result in warmer microclimates from a loss of vegetation cover [7,8]. Analysing this process spatially can indicate the distribution of potentially suitable habitat in future environments (i.e. refugia).
Remotely-sensed measures of ecosystem function, such as the Normalised Difference Vegetation Index (NDVI), can reveal changes to vegetation suitability for animals during extreme conditions provided data are available across time and at multiple spatial scales [9,10,11]. The NDVI is a measure of vegetation productivity and green biomass, it is the difference between red and near-infrared wavelengths in radiation reflected from vegetation (chlorophyll absorbance & mesophyll reflectance, respectively). NDVI is highly correlated with several vegetation characteristics including fraction of absorbed photosynthetically-active radiation, foliar nitrogen and chlorophyll, net primary production, and evapotranspiration [12,13]. Multiple studies have used NDVI to characterise the magnitude of drought impacts on vegetation condition and productivity [14,15]. The index also has been used in numerous studies of plant-animal interactions, including as a measure of forage phenology in assessing the trophic effects of climate on body mass of red deer (Cervus elaphus, [16]), and to test optimal foraging and species-energy theory by analysing vegetation productivity in relation to species diversity, abundance and landscape use for a review see [12].
Species distributions have shifted under anthropogenic climate change partly because of changes in habitat suitability, including for specialist herbivores [17,18,19]. In this study, we used the NDVI to identify productivity thresholds for the occurrence of a specialist arboreal folivore, the koala (Phascolarctos cinereus), throughout most of its distribution across eastern Australia. Using a long-term, continental-scale koala observation dataset, vegetation mapping, and time-series NDVI from Landsat sensors, we tested the hypothesis (H1) that locations where koalas had been observed would exhibit higher NDVI values than a random, representative sample from the same vegetation group (i.e. woodlands, open forests and tall open forests). We then obtained the minimum NDVI values at which koalas were observed for each vegetation group. We used these values to represent a NDVI threshold for koala occurrence and investigated how severe drought affected the distribution of potential koala habitat based on the thresholds and changes with drought to landscape NDVI, within vegetation groups. Potential habitat in this context means vegetation groups in which koalas are commonly found.
We tested the hypothesis that drought would reduce the area of potential habitat that is above the NDVI threshold for koalas (H2). We also investigated whether this suitable vegetation was significantly closer to perennial water than unsuitable vegetation (H3), before and during the drought. In this study, we use the term suitable or unsuitable vegetation to describe vegetation that is above or below koala NDVI thresholds in vegetation groups commonly associated with koala habitat. Whether koalas are present in a particular area can depend on several other factors that were beyond the scope of this study. By quantifying the responses both of our target species (the koala) and of the quality of its potential habitat via fluctuations in NDVI, we present a simple, powerful analysis capturing the impact of climate stress on vegetation and vegetation-dependant fauna which can be considered alongside other factors that influence habitat suitability.
2. Materials and Methods
Study Area
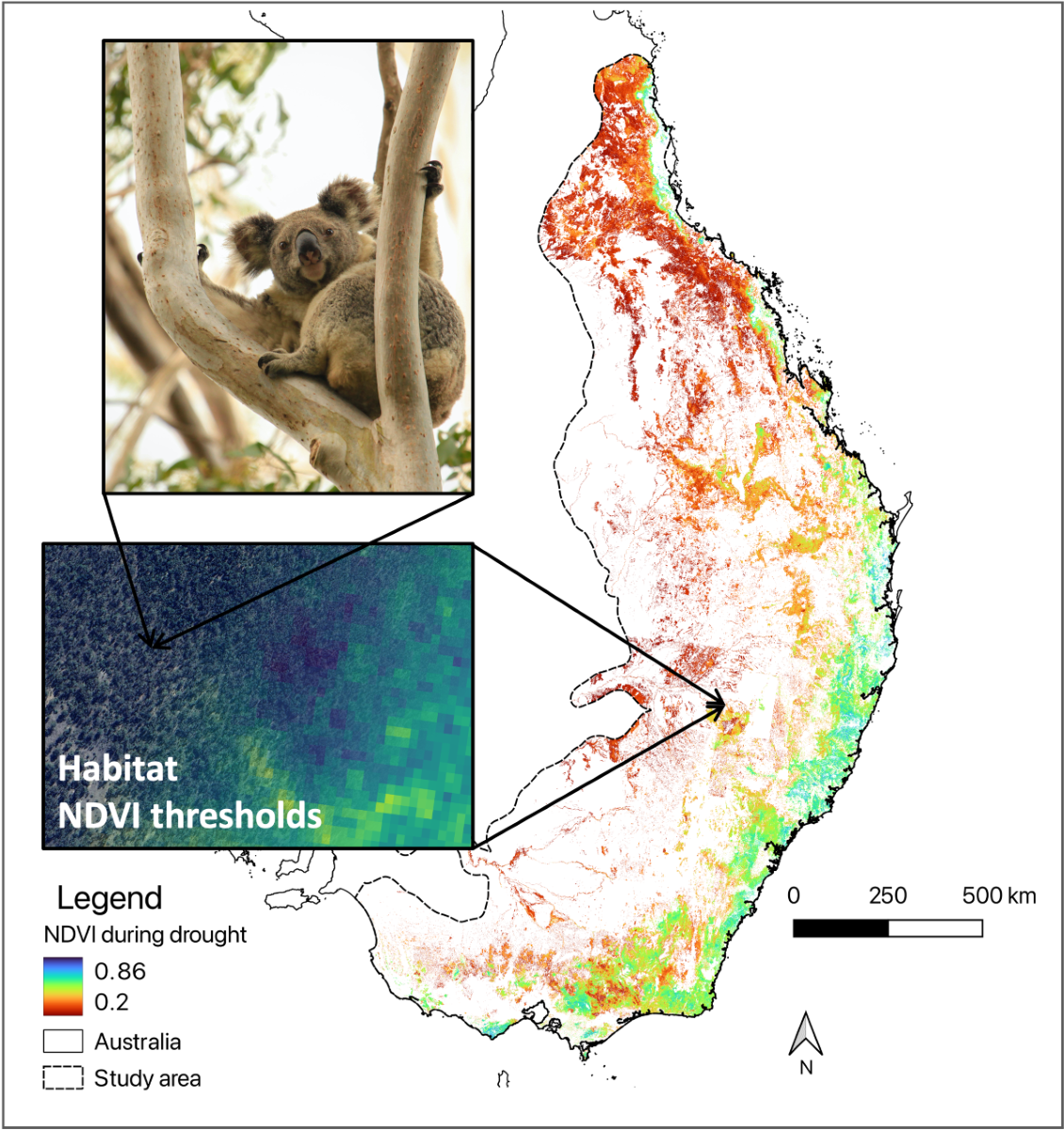
The study area encompasses most of the koala’s distribution, primarily across the Australian states of Queensland, New South Wales, Victoria, and the Australian Capital Territory (Figure 1). Tree cover in the area is predominately temperate evergreen broadleaf forests and woodlands, dominated by trees from the genus Eucalyptus, Angophora and Corymbia. Maximum mean annual temperatures (MAT) across the range have increased by 0.4-0.6°C per decade since 1970 [20], and the median MAT is 18.8°C, increasing to above 20°C in the north-west [21].
Study Species
The koala (Phascolarctos cinereus) is a specialist folivore that browses primarily on the foliage of eucalypt trees, from which they also obtain most of their water. Koalas have a wide distribution across the eastern states of Australia where they are associated with forests and woodlands dominated by evergreen trees in the genus Eucalyptus. Koalas have relatively high site fidelity, with sometimes overlapping home ranges that can vary widely in size, from less than 2 ha to over 100 ha in more open woodland environments [22]. Once common, large population declines, particularly in Queensland and New South Wales, have resulted in koalas in those states being recently listed as endangered [17,23,24,25]. Direct and indirect effects of heatwaves and droughts associated with climate change have contributed to koala mortality and population declines [10,26,27,28], along with habitat clearing and degradation [17,29,30]. Population decreases in semi-arid bioregions tend to align with drought periods [17,24,31].
NDVI of Koala Observations Compared to Landscapes
We obtained koala observations from the Species Profile and Threats Database (SPRAT) for the period 1995-2009 to identify koala observations for which NDVI and vegetation group could be determined [32] (Table 1). The observations extended throughout the koala range in eastern Australian states and territories (NSW, Queensland and ACT) where koalas are listed in legislation as endangered. For the state of Victoria, where the koala is not listed, we obtained observations from the Victorian Biodiversity Atlas [33]. The koala records had been compiled from many sources, including structured scientific and community surveys, and opportunistic sightings. We filtered for locational uncertainty of 100m or less and removed all unlikely (e.g. aquatic) or duplicate observations. This included removing records that were not associated with a precise date, to allow us to obtain NDVI values associated with each koala observation.

Table 1.
Data sources of koala records and landscape layers used in this study.
| Source | Variable | Study Use | Native Resolution | Source Website |
|---|---|---|---|---|
| Landsat satellite data cube hosted by Geoscience Aus. & NCI | Normalised Difference Vegetation Index [NDVI] (from corrected reflectance rasters) | Landscape vegetation condition and koala site vegetation condition | 25 m x 25 m | http://pid.geoscience.gov.au/dataset/ga/144643 |
| Species Profile and Threats Database (SPRAT) | Koala presence | Identify associated vegetation condition | Vector points | https://www.environment.gov.au/sprat |
| National Vegetation Information System (NVIS v6.0) | Vegetation group (based on NVIS major vegetation groups [MVG]) raster | Assign NDVI and koala presence observations into vegetation groups | 100 m x100 m | https://www.environment.gov.au/fed |
| Surface Hydrology Lines (Regional) | Perennial water courses and bodies | Calculate distance of observations and potential habitat to water | Vector lines and polygons | http://pid.geoscience.gov.au/dataset/ga/83107 |
Koala observation site NDVI
We extracted NDVI measurements at the filtered koala observation sites to derive suitability thresholds for koala associated vegetation groups. We used reflectance data from multispectral Landsat satellites 5 and 7 from 1995-2009 (25 m ground resolution, Table 1), which have equivalent spectral bands (i.e. red: 0.6-0.7 µm & 0.63-0.69 µm, and near-infrared [NIR]: 0.7-0.8 µm & 0.77-0.90 µm, respectively). Before 1999, Landsat 5 operated and the image timeseries had a 16-day frequency; for the remainder, both satellites 5 and 7 were operating and two images were acquired, and used, in each 16-day interval. NDVI was calculated following [34]:
We constrained the data to 0.2 ≤ NDVI ≤ 0.86, as values below this window are typically not considered vegetation and those above may be affected by saturation of NDVI for the vegetation groups and spatial resolution of our data [35,36]. These data were extracted from Landsat scenes stored in a timeseries datacube hosted by Digital Earth Australia (DEA), which is processed with a nadir-corrected bidirectional reflectance distribution function [37]. We masked poor-quality pixels including cloud using the ‘Fmask’ function built-in to the DEA platform [38]. We then collected the Landsat NDVI value nearest in time to each koala observation, within a maximum of 14-days either side of the observation. We used an arithmetic mean NDVI value from a 3x3 window of pixels around the location to get a measure more representative of the habitat and reduce the effect of NDVI outliers.
Vegetation group assignment
We assigned the retained koala observations to vegetation groups based on the Major Vegetation Groups dataset (MVG, Table 1) and analysed each vegetation group separately, because differences in NDVI across these groups driven by differences in vegetation composition and biomass limits comparisons between them [12]. To do this we used Python software packages xarray [39], and geopandas [40]. The MVG data are based on structural and floristic information including dominant genus, growth form, height, and cover [41]. It has been assembled from data collected at various on-ground dates; here we used the most recent combined version (version 6.0, [41]). We excluded records from MVGs including less than 500 koala observations, to maintain large sample sizes within groups and because less koala habitat occurs in these groups.
Comparing koala observation NDVI with mean NDVI for each vegetation group
To test whether the NDVI of koala sites differed from the mean NDVI for that vegetation group, we used generalised linear mixed modelling (GLMM). As a response variable, we used presence of koalas and drew an equally sized random subset of background points from the NDVI data source for respective vegetation groups in the landscape (woodland, open forest, and tall open forest, n=1,500 each). We paired these background landscape NDVI values to the times of koala observations, which were from anytime in 1995-2009, to control for temporal variation in NDVI. We modelled a binary response, either observation or background-landscape, with NDVI as a predictor in separate binomial GLMMs for each vegetation group. To account for spatial autocorrelation of NDVI, we included an exponential covariance term based on the spatial coordinates of observations. We then tested the model residuals for spatial autocorrelation. To fit these GLMMs, we used the software packages glmmTMB [42] and DHARMa [43] in the R environment [44].
NDVI Thresholds for Suitable Vegetation
We determined minimum thresholds of NDVI associated with koala observations by analysing the empirical cumulative distribution functions for all observations (1995-2009), using the statsmodel Python package [45]. The NDVI value, for each vegetation group, with an estimated probability equal to 5%.
is the NDVI below which koala occurrence is not likely and was selected as the threshold above which vegetation productivity is sufficient to support koalas.
Landscape NDVI in drought and non-drought periods
From 2001 to 2009, Australia experienced one of its worst droughts on record, dubbed the millennium drought [46] which had many harmful ecological impacts [47]. The rainfall anomaly was 12-13% below the mean of the previous century. We obtained landscape NDVI data for this period also from the DEA [37]. We used a subset of the Landsat NDVI imagery across the koala range from 2005-2009, which was the latter part of the millennium drought, when cumulative effects would have been greatest. We reduced this timeseries to the 5th percentile value for each pixel to produce an image of minimal vegetation productivity without capturing low outliers (e.g. unmasked cloud). We repeated this process for 1995-1999 to make comparisons with drought.
Vegetation group assignment
To limit the analyses to vegetation groups most frequently associated with koala observations in the landscape, we combined the NVIS-derived groups as described previously with the minimum NDVI images. Rasters of each vegetation group and the NDVI images were aligned spatially, to account for the different resolutions and projections. Taking a Boolean mask of each vegetation group we multiplied it by the NDVI, resulting in a landscape NDVI layer with resolution of the Landsat input, divided into vegetation groups. This process was repeated for the two time periods, using a raster calculator [48].
Applying koala NDVI thresholds across the landscape
We applied the suitability thresholds for each vegetation group to the drought and non-drought NDVI images using a raster calculator. The result was a set of NDVI rasters of suitable vegetation, for pre- and drought periods which excluded potential habitat vegetation deemed unsuitable to support koalas because the NDVI value of that vegetation was below the relevant vegetation-group threshold. We then computed difference layers by subtracting pre-drought from drought to reveal how much potential habitat shifted from suitable pre-drought to unsuitable in drought.
Proximity of Suitable Vegetation to Water
To understand the spatial proximity of vegetation above the NDVI threshold for koalas in relation to distance to perennial surface water, we combined the suitability maps with surface hydrology data [49]. These data describe perennial watercourses and water bodies for the continent (Table 1). We converted the vectors to a raster at the spatial resolution of the NDVI data, then created a proximity raster of distance of each pixel to the target (water) pixels, using the Rasterize function (GDAL, [50]). With the same software, we multiplied the proximity raster by the mask of vegetation suitability rasters for each of two time periods and the two suitability groups, producing rasters of distance (km) to suitable and unsuitable vegetation pixels. To test whether the distance to perennial water of suitable pixels differed from the average distance to water for each vegetation group, we used generalised linear mixed modelling (GLMM). We used a subset of pixels from each vegetation group, which we randomly selected from across the landscape, made up of half suitable and half unsuitable (total n=4,500). The modelling approach was similar to that of koala site ~ NDVI (H1), except the binomial response was pixels above or below NDVI thresholds and the predictor, distance to water, was logarithmically transformed because of its Euclidean structure. We used this same model structure to test if koala observations were closer to water than random sites, with the response variable being binary: koala site or random location. Here, we used the same coordinates as the subset of observations used in the methods for H1.
3. Results
3.1. NDVI of Koala Observations Compared to Landscapes
3.2. NDVI Thresholds for Suitable Vegetation
Figure 4 shows the minimum (5th percentile) identified in the cumulative distributions of NDVI for koala observation pixels for each vegetation group (i.e., vegetation thresholds; woodlands = 0.393, open forests = 0.436, tall open forests = 0.549). Woodland followed by open forest, had thresholds lower than the tall open forests, reflecting the different productivity levels of each vegetation group.
Across the entire study area, the effect of drought was observed most strongly in the minimum NDVI rather than the mean NDVI; changes to the mean were small and not significant in all vegetation groups. The landscape NDVI minimum across vegetation groups declined significantly between pre- and late-drought periods (all p < 0.01), due to declines in all vegetation groups across periods. Figure 5 shows distributions in both periods for respective vegetation groups.
The total area of all three vegetation groups was 429,997 km2. During the pre-drought period, 49% of this area remained above the koala NDVI threshold for all vegetation groups (Table 2; Figure 6). This means approximately half of potential koala habitat was not sufficiently productive to continuously support koalas (i.e. suitable vegetation). During the drought, we observed a 12% reduction in suitable vegetation area, across vegetation groups. This affirmed our second hypothesis, that NDVI decline in drought reduces the area of potential habitat for koalas (H2). Figure 7 shows an example of this reduction within a landscape, where of 606 ha of suitable vegetation prior to drought, 16.8% declined below thresholds in drought (102 ha).
3.3. Proximity of Suitable Vegetation to Water
The mean distance to water from koala observations was 5 km for woodland observations (SD=7.9 km), 1.8 km for open forest (SD=2.4 km), and 1.3 km for tall open forest (SD=1.6 km). The maximum distances at which koalas were observed from perennial water were 51.4 km, 32.2 km, and 21.1 km in the three vegetation groups respectively. Koala observations were significantly closer to water in open forests than random points taken from the same landscape type (p < 0.01), but this pattern was not found in woodlands or tall open forests (p = 0.93, and p = 0.3, respectively).
We found that suitable and unsuitable vegetation could occur 40 km or more from water, in all vegetation groups. However, there was a significant effect of distance to water on suitability (i.e. vegetation above or below NDVI minimum threshold) in tall open forest (p < 0.001) and open forests (p = 0.04) but not in woodlands (p = 0.8).
4. Discussion
Climate change is challenging our abilities to understand rapidly changing current and future habitat suitability and species distributions, creating a need for methods to monitor such changes and identify potential refugia, thus allowing better conservation for wildlife and their habitat [51]. For the first time, we identified thresholds of the minimum NDVI associated with the presence of a specialist folivore, the koala, allowing us to assess the extent of potential habitat before and during a major drought. This study extends previous work focusing on local abundance of folivores with NDVI (e.g., [52]) by considering the relationship between NDVI and ecological niche at broader spatial scales. Recognising a basic relationship with available energy, we did not aim to build a model of the whole NDVI-animal relationship, as has been done elsewhere (e.g. NDVI-abundance, [53]), instead we identified bounds of that relationship and extrapolated these limits in space.
NDVI of Koala Observations Compared to Landscapes
In all three vegetation groups, we found that koalas were more likely to be observed in areas of the landscape that had higher NDVI values than the average for that vegetation group. These findings are consistent with earlier, regional koala occupancy modelling, based on acoustic detection, that showed landscape NDVI had a significant, positive effect even within a relatively mesic region, the North Coast and Northern Tablelands of NSW [54]. Similarly, [52] found that locations where another specialist eucalypt folivore, the greater glider (Petauroides volans), was observed had significantly higher NDVI on average than areas where they were not observed. That study also showed a positive relationship between abundance and NDVI.
Several studies have suggested that NDVI may correlate to available energy from forage [52,55,56] and preferentially foraging in patches of higher energy would allow animals to increase energy intake while reducing expenditure, although the ability to metabolise the forage in these patches is also important [57,58]. NDVI is linked to forage quality and available energy in several ways. NDVI is a measure of photosynthetic capacity, the system comprising pigments and proteins, of which the latter is a limiting nutrient for koalas and many other folivores [59,60]. Furthermore, NDVI is highly correlated with net primary productivity and canopy biomass that makeup forage energy [12,13,61]. Declines in productivity and biomass will lead to lower NDVI values and can also reflect less available energy for folivores [62]. Additionally, lower canopy biomass means less shade and evapotranspiration, resulting in hotter micro-climates that can reduce habitat suitability [63].
Since our data were sourced from koala observations that included opportunistic sightings, rather than a random sample across the entire range, there may be bias in observations towards areas with more observers, which may also be more productive in terms of NDVI [64]. We expect this effect to be less pronounced in the vegetation groups that are more common along the coast (e.g., tall open forest and open forest) where human populations are most concentrated and therefore tend to coexist with koalas. This potential bias could not be entirely avoided, but the inclusion of data from scientific surveys and research sites in more remote areas, as well as the consistent pattern across all forest groups and with earlier studies of folivore occurrence and NDVI, provide additional support to our findings.
NDVI Thresholds for Suitable Vegetation
The minimum NDVI thresholds for koala observations differed across vegetation groups, which is in keeping with the differing distributions of NDVI in those groups. However, in all vegetation groups, the minimum NDVI values associated with koala observations were always higher than the minimum value possible for that landscape group. Accordingly, we also found that a sizable area of all three vegetation groups fell below the NDVI minimum, even prior to drought. The area of potential habitat above the NDVI thresholds associated with koala occurrence declined by 12% during the millennium drought, supporting the notion that during drought, koala habitat contracts in spatial extent. The largest portion of potential habitat that fell below thresholds was in woodlands, but in all vegetation groups there was contraction.
The western, inland extent of the koala’s range is typically comprised of woodlands, with intersecting riparian areas where koalas may occur in higher densities relative to other areas [65]. Water is thought to be a limiting factor for both tree and koala abundance in such regions, and previous studies have identified relationships between foliar NDVI and environmentally available water in these western woodlands, particularly in riparian areas predominated by the locally preferred koala browse species, Eucalyptus camaldulensis [17,86]. Notably, these riparian forests, embedded in the broader woodland landscape, were often classified as tall open forests, due to their structural profile that differed from the surrounding woodland landscape. This likely contributed to the overall proportion of tall open forests that fell below the NDVI threshold during the drought period, and the significant relationship between distance to perennial water and tall open forest above the NDVI threshold. Although these riparian forests tend to be phreatophytic, maintaining hydration longer during dry conditions, other studies have found that the NDVI values associated with E. camaldulensis dominated riparian forests can decline substantially during prolonged drought conditions [86]. Other types of tall open forests in our study typically occur closer to the coast and experience higher rainfall, so water is less likely to be a limiting factor on vegetation productivity and NDVI.
In times of drought, koalas may be critically stressed in landscapes that fall below the vegetation productivity threshold associated with their occurrence. Reduced forage water supply can lead to heat-induced water stress in koalas if free water is not available [51]. Dehydration of koalas has been documented within population crashes during drought [31]. Koalas in refugial areas that remain above the threshold of vegetation productivity during times of drought or extreme heat are more likely to survive [31]. The periodic loss of habitat suitability due to drought impacts is particularly concerning for species like the koala and other arboreal folivores that have less capacity for migration [65] and due to forage specialisation, cannot adapt by expanding their dietary breadth [66]. Such contractions in suitable vegetation may also impact populations by way of habitat fragmentation [67].
Using the relationship between NDVI minimum thresholds and change in vegetation conditions from pre-drought to drought periods, we have revealed the extent of potential koala habitat contraction during the millenium drought. In a future of increasingly severe droughts [68], both the extent of habitat contraction and the duration of time in which habitat drops below koala NDVI thresholds could increase. Our analysis considered the extant vegetation, by using reflectance data and forest-type mapping, unlike many distribution or population models [51,69,70]. [51] estimated the extent and distribution of suitability based on a biophysical, spatial model which does recognise the importance of changes to koala habitat quality. Like the results of that study, we found contractions in potential koala habitat during drought occurred mainly in the inland parts of the koala’s distribution (Figure 6). Such contraction of populations to refugia during drought has been documented [9,31]; however, the contraction of potential habitat in drought has not, to our knowledge, been revealed before this study.
Our study overcomes some of the limitations of predicting species’ habitat distribution based on forecasts of climate averages. Such approaches do not incorporate climate extremes well, nor the trophic interactions caused by those extremes [71]. Whereas NDVI integrates the many and varied effects of realised ecological drought across landscape vegetation. The drought studied here did contain spatial variability, including substantial effects in the south of Queensland [47]. Our inclusion of the less affected northern, tropical regions may have reduced the observed drought effect for vegetation groups also occurring there.
Notably, a sizable portion of the vegetation in the Victorian central and eastern highlands that fell below NDVI thresholds during drought was directly affected by the Black Summer Fires of 2009 (Figure 6, see also [72]. The NDVI of burnt forest is typically lower than the same area before fire, particularly if canopy scorch occurs [73]. The millennium drought conditions contributed to the intensity of the 2009 fires [74,75] and in the short-term, fire can reduce koala populations and habitat quality due to forage loss [76]. Some vegetation may recover productivity quickly via epicormic regrowth, providing potential forage for koalas [77], although this recovery was slowed in some areas by ongoing drought [78]. Other possible drivers of NDVI minima in some cases, could include land clearing [30] and tree dieback [79].
Proximity of Suitable Vegetation to Water
On average, the distance between suitable vegetation and perennial water was 7km, which is greater than the width of even the largest koala home ranges. However, we did find that unsuitable vegetation tended to occur significantly farther from water bodies than suitable vegetation for some vegetation groups. During drought, the distance to water was slightly less for suitable vegetation than before drought, but still 6.4 km on average. Our analysis of the suitability maps does not suggest that access to free-standing water is a habitat requirement for most koalas; however, it’s possible that koalas in the western part of their range may rely on it more often [17].
[65] found that presence of free-standing water influenced the home range size of koalas in western Queensland, and koalas in arid sites spent more time in riparian parts of their habitat compared to those in relatively mesic sites (~80% compared to ~40%). Although koalas are thought to obtain most of their water from leaves, they will also drink free water with increasing frequency as temperatures increase [80]. Sources of free water may be available in addition to those used in this study, including during some stages of drought. These sources include ephemeral creeks, waterholes, farm dams and bores, although koalas survived prior to the construction of the latter two sources. In addition, it is likely that trees in some areas also access water from underground sources that were not considered in this study but could contribute to NDVI and vegetation suitability. More research is needed to understand how water, both in the koala’s environment and in their diet of leaves, contributes to habitat suitability and the distribution and resilience of koala populations.
Further Applications, in Other Systems and in Management
Having confirmed the existence of vegetation-specific thresholds for koalas and mapped potential koala habitat during drought, it would be valuable to expand the study temporally. The full Landsat time-series could be utilised to estimate length of periods of suitability. This may reveal areas that are resistant to drought and only shift during the most extreme conditions (i.e., highly skewed temporal distributions). Perhaps more valuably, the Landsat data might also provide insight into the link between koala presence, recovery from drought and its interval, including interactions with temperature which may increase the speed of drought onset [81].
Analyses of habitat productivity thresholds, like that here, need not be limited to specialist folivores, such as the koala. Other possible systems of study include herbivores that are limited by forage quality in environments that experience extreme periods, particularly those systems with vegetation sensitive to climate change. However, stressors other than climate extremes could also be studied using our approach, provided the stressor affects vegetation NDVI (e.g., seasonal insect outbreaks). Examples of other possible study systems include savanna grass specialists of El-Niño affected South Africa [82], or environments in China where vegetation loss in drought is substantial [83]. Such systems could feature generalist herbivores because the NDVI is responsive to productivity in most plants [34,84]; however, analyses may be more complicated if diets are diverse.
5. Conclusions
This section is not mandatory but can be added to the manuscript if the discussion is unusually long or complex.
Our results consider one measure of habitat quality based on vegetation productivity, but the results may be useful to environmental managers precisely because of that simplicity. This is because NDVI integrates many environmental effects on vegetation and indicates extant vegetation. Models that include more parameters and complex influences can be harder to comprehend for management purposes [85]. The maps produced in this study provide an outline of areas of potential koala habitat that could be conserved through future climatic extremes.
Author Contributions
Conceptualization, I.K., K.Y., C.M. and B.M.; methodology, C.M. and I.K.; software, C.M. and I.K.; formal analysis, I.K., C.M., M.E. and K.Y.; investigation, I.K., C.M., M.E., K.Y. and B.M.; resources, C.M., K.Y., B.M.; data curation, C.M., I.K.; writing—original draft preparation, I.K.; writing—review and editing, I.K., C.M., J.E., K.Y. and B.M.; visualization, I.K., J.E.; supervision, K.Y., B.M.; project administration, I.K., K.Y., B.M.; funding acquisition, K.Y., B.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NSW Department of Climate Change, Energy, the Environment and Water (New South Wales, Australia), project number A13782.
Data Availability Statement
The code, input and extracted data can be found at the following webpage: https://hie-pub.westernsydney.edu.au/projects/ndvi_koala_analysis/
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhongming, Z, Linong, L, Xiaona, Y, Wangqiang, Z, Wei, L, et al. 2021, AR6 Climate Change 2021: The Physical Science Basis.
- Calvin, K, Dasgupta, D, Krinner, G, Mukherji, A, Thorne, PW, Trisos, C, Romero, J, et al. 2023, IPCC, 2023: Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, H. Lee and J. Romero (eds.)]. IPCC, Geneva, Switzerland., P Arias et al. (eds.), <https://www.ipcc.ch/report/ar6/syr/>.
- Wagner, B, Baker, PJ, Stewart, SB, Lumsden, LF, Nelson, JL, Cripps, JK, Durkin, LK, et al. 2020, Climate change drives habitat contraction of a nocturnal arboreal marsupial at its physiological limits, Ecosphere, vol. 11, no. 10.
- Walther, G-R, Post, E, Convey, P, Menzel, A, Parmesan, C, Beebee, TJC, Fromentin, J-M, et al. 2002, Ecological responses to recent climate change, Nature, vol. 416, no. 6879, pp. 389-395.
- Dublin, HT & Ogutu, JO 2015, Population regulation of African buffalo in the Mara-Serengeti ecosystem, Wildlife Research, vol. 42, no. 5, p. 382.
- Gedir, J V., Cain, JW, Harris, G, & Turnbull, TT 2015, Effects of climate change on long-term population growth of pronghorn in an arid environment, Ecosphere, vol. 6, no. 10, pp. 1-20.
- Birnie-Gauvin, K, Peiman, KS, Raubenheimer, D, & Cooke, SJ 2017, Nutritional physiology and ecology of wildlife in a changing world, Conservation Physiology, vol. 5, no. 1.
- Fuller, A, Mitchell, D, Maloney, SK, & Hetem, RS 2016, Towards a mechanistic understanding of the responses of large terrestrial mammals to heat and aridity associated with climate change, Climate Change Responses, vol. 3, no. 1, p. 10.
- Smith, AG, McAlpine, CA, Rhodes, JR, Lunney, D, Seabrook, L, & Baxter, G 2013, Out on a limb: Habitat use of a specialist folivore, the koala, at the edge of its range in a modified semi-arid landscape, Landscape Ecology, vol. 28, no. 3, pp. 415-426.
- Crowther, MS, Lunney, D, Lemon, J, Stalenberg, E, Wheeler, R, Madani, G, Ross, KA, et al. 2014, Climate-mediated habitat selection in an arboreal folivore, Ecography, vol. 37, no. 4, pp. 336-343.
- Davies, N, Gramotnev, G, Seabrook, L, McAlpine, C, Baxter, G, Lunney, D, & Bradley, A 2014, Climate-driven changes in diet composition and physiological stress in an arboreal folivore at the semi-arid edge of its distribution, Biological Conservation, vol. 172, pp. 80-88.
- Pettorelli, N, Ryan, S, Mueller, T, Bunnefeld, N, Jedrzejewska, B, Lima, M, & Kausrud, K 2011, The Normalized Difference Vegetation Index (NDVI): unforeseen successes in animal ecology, Climate Research, vol. 46, no. 1, pp. 15-27.
- Gamon, JA, Field, CB, Goulden, ML, Griffin, KL, Hartley, AE, Joel, G, Penuelas, J, et al. 1995, Relationships Between NDVI, Canopy Structure, and Photosynthesis in Three Californian Vegetation Types, Ecological Applications, vol. 5, no. 1, pp. 28-41.
- Karnieli, A, Agam, N, Pinker, RT, Anderson, M, Imhoff, ML, Gutman, GG, Panov, N, et al. 2010, Use of NDVI and Land Surface Temperature for Drought Assessment: Merits and Limitations, Journal of Climate, vol. 23, no. 3, pp. 618-633.
- Verdon-Kidd, DC, Scanlon, BR, Ren, T, & Fernando, DN 2017, A comparative study of historical droughts over Texas, USA and Murray-Darling Basin, Australia: Factors influencing initialization and cessation, Global and Planetary Change, vol. 149, pp. 123-138.
- Mysterud, A, Yoccoz, NG, Langvatn, R, Pettorelli, N, & Stenseth, NC 2008, Hierarchical path analysis of deer responses to direct and indirect effects of climate in northern forest, Philosophical Transactions of the Royal Society B: Biological Sciences, vol. 1501, no. 363, pp. 2359-2368. [CrossRef]
- Seabrook, L, McAlpine, C, Baxter, G, Rhodes, J, Bradley, A, & Lunney, D 2011, Drought-driven change in wildlife distribution and numbers: A case study of koalas in south west Queensland, Wildlife Research, vol. 38, no. 6, pp. 509-524.
- Lovari, S, Franceschi, S, Chiatante, G, Fattorini, L, Fattorini, N, & Ferretti, F 2020, Climatic changes and the fate of mountain herbivores, Climatic Change, vol. 162, no. 4, pp. 2319-2337.
- Parida, M, Hoffmann, AA, & Hill, MP 2015, Climate change expected to drive habitat loss for two key herbivore species in an alpine environment, Journal of Biogeography, vol. 42, no. 7, pp. 1210-1221.
- Trewin, B 2016, The Australian Climate Observations Reference Network--Surface Air Temperature ACORN, J. Appl. Met. Climatol, vol. 48, pp. 2160-2168.
- BOM 2020a, Mean monthly and mean annual maximum, minimum and mean temperature data (base climatological data sets).
- Youngentob, KN, Marsh, KJ, & Skewes, J 2021, A review of koala habitat assessment criteria and methods, Canberra, <https://www.awe.gov.au/environment/epbc/publications>.
- Predavec, M, Lunney, D, Shannon, I, Lemon, J, Sonawane, I, & Crowther, M 2018, Using repeat citizen science surveys of koalas to assess their population trend in the north-west of New South Wales: Scale matters. Australian Mammalogy, vol. 40, no. 1, 47-57. [CrossRef]
- Lunney, D, Predavec, M, Sonawane, I, Kavanagh, R, Barrott-Brown, G, Phillips, S, Callaghan, J, et al. 2017, The remaining koalas (Phascolarctos cinereus) of the Pilliga forests, north-west New South Wales: refugial persistence or a population on the road to extinction?, Pacific Conservation Biology, vol. 23, no. 3, pp. 277-294.
- DAWE 2022, Conservation Advice for Phascolarctos cinereus (Koala) combined populations of Queensland, New South Wales and the Australian Capital Territory, Canberra.
- Beale, PK, Marsh, KJ, Foley, WJ, & Moore, BD 2018, A hot lunch for herbivores: physiological effects of elevated temperatures on mammalian feeding ecology, Biological Reviews, vol. 93, no. 1, pp. 674-692.
- Adam, D, Johnston, SD, Beard, L, Nicolson, V, Gaughan, JB, Lisle, AT, FitzGibbon, S, et al. 2020, Body temperature of free-ranging koalas (Phascolarctos cinereus) in south-east Queensland, International Journal of Biometeorology, vol. 64, no. 8, pp. 1305-1318.
- Youngentob, KN, Lindenmayer, DB, Marsh, KJ, Krockenberger, AK, & Foley, WJ 2021, Food intake: an overlooked driver of climate change casualties?, Trends in Ecology & Evolution, vol. 36, no. 8, pp. 676-678.
- Reside, AE, Beher, J, Cosgrove, AJ, Evans, MC, Seabrook, L, Silcock, JL, Wenger, AS, et al. 2017, Ecological consequences of land clearing and policy reform in Queensland, Pacific Conservation Biology, vol. 23, no. 3, pp. 219-230.
- Taylor, MFJ 2020, Destruction of Koala habitat increased after listing as vulnerable in 2012, WWF-Australia.
- Gordon, G, Brown, AS, & Pulsford, T 1988, A koala (Phascolarctos cinereus Goldfuss) population crash during drought and heatwave conditions in south-western Queensland, Australian Journal of Ecology, vol. 13, no. 4, pp. 451-461.
- DOEE, Department of Environment and Energy 2015, Species Profile and Threats Database (SPRAT). https://www.environment.gov.au/cgi-bin/sprat/public/sprat.pl.
- Department of Environment, Land, W and P 2021, Victorian Biodiversity Atlas.
- Tucker, CJ 1979, Red and photographic infrared linear combinations for monitoring vegetation, Remote Sensing of Environment, vol. 8, no. 2, pp. 127-150.
- Asrar, G, Fuchs, M, Kanemasu, ET, & Hatfield, JL 1984, Estimating Absorbed Photosynthetic Radiation and Leaf Area Index from Spectral Reflectance in Wheat1, Agronomy Journal, vol. 76, pp. 300-306.
- Sebastiani, A, Salvati, R, & Manes, F 2023, Comparing leaf area index estimates in a Mediterranean forest using field measurements, Landsat 8, and Sentinel-2 data, Ecological Processes, vol. 12, no. 1.
- Krause, C, Dunn, B, Bishop-Taylor, R, Adams, C, Burton, C, Alger, M, Chua, S, et al. 2021, Digital Earth Australia notebooks and tools repository.
- Zhu, Z, Wang, S, & Woodcock, CE 2015, Improvement and expansion of the Fmask algorithm: Cloud, cloud shadow, and snow detection for Landsats 4-7, 8, and Sentinel 2 images, Remote Sensing of Environment, vol. 159, pp. 269-277.
- Hoyer, S & Hamman, JJ 2017, xarray: N-D labeled Arrays and Datasets in Python, Journal of Open Research Software, vol. 5, pp. 1-6.
- Jordahl, K, Bossche, JV den, Wasserman, J, McBride, J, Gerard, J, Fleischmann, M, Tratner, J, et al. 2019, Geopandas/geopandas: V0. 6.1.
- DAWE 2020, National Vegetation Information System V6.0.
- Brooks, ME, Kristensen, K, Van Benthem, KJ, Magnusson, A, Berg, CW, Nielsen, A, Skaug, HJ, et al. 2017, glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling, The R journal, vol. 9, no. 2, pp. 378-400.
- Hartig, F 2022, DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models.
- R Core Team 2022, R: A Language and Environment for Statistical Computing.
- Seabold, S & Perktold, J 2010,Statsmodels: Econometric and Statistical Modeling with Python, in, Proceedings of the 9th Python in Science Conference, pp.92-96.
- Van Dijk, AIJM, Beck, HE, Crosbie, RS, De Jeu, RAM, Liu, YY, Podger, GM, Timbal, B, et al. 2013, The Millennium Drought in southeast Australia (2001-2009): Natural and human causes and implications for water resources, ecosystems, economy, and society, Water Resources Research, vol. 49, no. 2, pp. 1040-1057.
- Jiao, T, Williams, CA, Rogan, J, De Kauwe, MG, & Medlyn, BE 2020, Drought Impacts on Australian Vegetation During the Millennium Drought Measured With Multisource Spaceborne Remote Sensing, Journal of Geophysical Research: Biogeosciences, vol. 125, no. 2, pp. 1-19.
- QGIS.org 2021, QGIS Geographic Information System.
- Crossman, S & Li, O 2015, Surface Hydrology Lines (Regional). http://pid.geoscience.gov.au/dataset/ga/83107.
- Rouault, E, Warmerdam, F, Schwehr, K, Kiselev, A, Butler, H, Łoskot, M, Szekeres, T, et al. 2022, GDAL.
- Briscoe, NJ, Kearney, MR, Taylor, CA, & Wintle, BA 2016, Unpacking the mechanisms captured by a correlative species distribution model to improve predictions of climate refugia, Global Change Biology, vol. 22, no. 7, pp. 2425-2439.
- Youngentob, KN, Yoon, H-J, Stein, J, Lindenmayer, DB, & Held, AA 2015, Where the wild things are: using remotely sensed forest productivity to assess arboreal marsupial species richness and abundance, Diversity and Distributions, vol. 21, no. 8, pp. 977-990.
- Pettorelli, N, Bro-Jorgensen, J, Durant, SM, Blackburn, T, & Carbone, C 2009, Energy availability and density estimates in African ungulates, American Naturalist, vol. 173, no. 5, pp. 698-704.
- Law, B, Brassil, T, Gonsalves, L, Roe, P, Truskinger, A, & McConville, A 2018, Passive acoustics and sound recognition provide new insights on status and resilience of an iconic endangered marsupial (koala Phascolarctos cinereus) to timber harvesting, PLoS ONE, vol. 13, no. 10, pp. 1-20.
- Law, B, Caccamo, G, Roe, P, Truskinger, A, Brassil, T, Gonsalves, L, McConville, A, et al. 2017, Development and field validation of a regional, management-scale habitat model: A koala Phascolarctos cinereus case study, Ecology and Evolution, vol. 7, no. 18, pp. 7475-7489.
- McArt, SH, Spalinger, DE, Collins, WB, Schoen, ER, Stevenson, T, & Bucho, M 2009, Summer dietary nitrogen availability as a potential bottom-up constraint on moose in south-central Alaska, Ecology, vol. 90, no. 5, pp. 1400-1411.
- Graham, P 2019,Optimal foraging theory: an introduction, in, Encyclopedia of animal behaviour, Elsevier Academic Press, pp.111-117.
- Stephens, DW & Krebs, JR 1986, Foraging Theory, Princeton University Press.
- Evans, JR 1989, Photosynthesis and nitrogen relationships in leaves of C3 plants, Oecologia, , no. 78, pp. 9-19.
- Moore, BD, & Foley, WJ 2005, Tree use by koalas in a chemically complex landscape, Nature, vol. 7041, no. 435, pp. 488. https://www.nature.com/articles/nature03551.pdf.
- Kozlowski, TT & Pallardy, SG 2002, Acclimation and adaptive responses of woody plants to environmental stresses, Bot. Rev, vol. 68, no. 2, pp. 270-334.
- Wright, IJ, Reich, PB, Westoby, M, Ackerly, DD, Baruch, Z, Bongers, F, Cavender-Bares, J, et al. 2004, The worldwide leaf economics spectrum, <www.nature.com/nature>.
- Ellis, W, Melzer, A, Clifton, I, & Carrick, F 2010, Climate change and the koala Phascolarctos cinereus: water and energy, Australian Zoologist, vol. 35, no. 2, pp. 369-377.
- Donohue, RJ, Roderick, ML, & McVicar, TR 2008, Deriving consistent long-term vegetation information from AVHRR reflectance data using a cover-triangle-based framework, Remote Sensing of Environment, vol. 112, no. 6, pp. 2938-2949.
- Davies, N, Gramotnev, G, Seabrook, L, Bradley, A, Baxter, G, Rhodes, J, Lunney, D, et al. 2013, Movement patterns of an arboreal marsupial at the edge of its range: A case study of the koala, Movement Ecology, vol. 1, no. 1, pp. 1-15.
- DeGabriel, JL, Moore, BD, Felton, AM, Ganzhorn, JU, Stolter, C, Wallis, IR, Johnson, CN, et al. 2014, Translating nutritional ecology from the laboratory to the field: milestones in linking plant chemistry to population regulation in mammalian browsers, Oikos, vol. 123, no. 3, pp. 298-308.
- Crowther, MS, Rus, AI, Mella, VSA, Krockenberger, MB, Lindsay, J, Moore, BD, & McArthur, C 2022, Patch quality and habitat fragmentation shape the foraging patterns of a specialist folivore, Behavioral Ecology.
- CSIRO & BoM 2023, State of The Climate 2022, viewed 14 May 2024, <https://www.csiro.au/en/research/environmental-impacts/climate-change/state-of-the-climate>.
- Santika, T, Mcalpine, CA, Lunney, D, Wilson, KA, & Rhodes, JR 2014, Modelling species distributional shifts across broad spatial extents by linking dynamic occupancy models with public-based surveys, Diversity and Distributions, vol. 20, no. 7, pp. 786-796.
- Adams-Hosking, C, Grantham, HS, Rhodes, JR, McAlpine, C, & Moss, PT 2011, Modelling climate-change-induced shifts in the distribution of the koala, Wildlife Research, vol. 38, no. 2, p. 122.
- Fitzpatrick, MC & Hargrove, WW 2009, The projection of species distribution models and the problem of non-analog climate, Biodiversity and Conservation, vol. 18, no. 8, pp. 2255-2261.
- Nolan, RH, Lane, PNJ, Benyon, RG, Bradstock, RA, & Mitchell, PJ 2015, Trends in evapotranspiration and streamflow following wildfire in resprouting eucalypt forests, Journal of Hydrology, vol. 524, no. January, pp. 614-624.
- Caccamo, G, Bradstock, R, Collins, L, Penman, T, & Watson, P 2015, Using MODIS data to analyse post-fire vegetation recovery in Australian eucalypt forests, Journal of Spatial Science, vol. 60, no. 2, pp. 341-352.
- Choat, B, Brodribb, TJ, Brodersen, CR, Duursma, RA, López, R, & Medlyn, BE 2018, Triggers of tree mortality under drought, Nature, vol. 558, no. 7711, pp. 531-539.
- Yebra, M, Quan, X, Riaño, D, Rozas Larraondo, P, van Dijk, AIJM, & Cary, GJ 2018, A fuel moisture content and flammability monitoring methodology for continental Australia based on optical remote sensing, Remote Sensing of Environment, vol. 212, no. November 2017, pp. 260-272.
- Johnson, DC & Shapcott, A 2024, Koala forest habitat recovery varies with fire severity, Forest Ecology and Management, vol. 556, no. May 2023, p. 121704.
- Lane, MR, Youngentob, KN, Clark, RG, & Marsh, KJ 2024, The nutritional quality of post-fire eucalypt regrowth and its consumption by koalas in the New South Wales Southern Tablelands, Australian Journal of Zoology, vol. 71, no. 3.
- Rifai, SW, De Kauwe, MG, Gallagher, R V., Cernusak, LA, Meir, P, & Pitman, AJ 2024, Burn Severity and Post-Fire Weather Are Key to Predicting Time-To-Recover From Australian Forest Fires, Earths Future, vol. 12, no. 4.
- Losso, A, Challis, A, Gauthey, A, Nolan, RH, Hislop, S, Roff, A, Boer, MM, et al. 2022, Canopy dieback and recovery in Australian native forests following extreme drought, Scientific Reports, vol. 12, no. 1, pp. 1-14.
- Mella, VSA, McArthur, C, Krockenberger, MB, Frend, R, & Crowther, MS 2019, Needing a drink: Rainfall and temperature drive the use of free water by a threatened arboreal folivore, PLoS ONE, vol. 14, no. 5, pp. 1-16.
- Kirono, DGC, Round, V, Heady, C, Chiew, FHS, & Osbrough, S 2020, Drought projections for Australia: Updated results and analysis of model simulations, Weather and Climate Extremes, vol. 30.
- Donaldson, JE, Parr, CL, Mangena, EH, & Archibald, S 2020, Droughts Decouple African Savanna Grazers from Their Preferred Forage with Consequences for Grassland Productivity, Ecosystems, vol. 23, no. 3, pp. 689-701.
- Guo, W, Huang, S, Huang, Q, Leng, G, Mu, Z, Han, Z, Wei, X, et al. 2023, Drought trigger thresholds for different levels of vegetation loss in China and their dynamics, Agricultural and Forest Meteorology, vol. 331, no. October 2022, p. 109349.
- Pettorelli, N 2013, The Normalized Difference Vegetation Index, Oxford University Press.
- Mitchell, DL, Soto-Berelov, M, Langford, WT, & Jones, SD 2021, Factors confounding koala habitat mapping at multiple decision-making scales, Ecological Management and Restoration, vol. 22, no. 2, pp. 171-182.
- Doody, TM, Benger, SN, Pritchard, JL, & Overton, IC 2014, Ecological response of Eucalyptus camaldulensis (river red gum) to extended drought and flooding along the River Murray, South Australia (1997-2011) and implications for environmental flow management, Marine and Freshwater Research, vol. 65, no. 12, pp. 1082-1093.
- BOM 2020b, High resolution mean monthly and mean annual rainfall data (base climatological data sets).
- GEBCO Bathymetric Compilation Group 2021, The GEBCO_2021 Grid - a continuous terrain model of the global oceans and land.
Figure 1.
Study area covering most of the koala range in eastern Australia (i.e. coloured area). Colours showing mean annual precipitation [87] classes and shading representing elevation [88], increasing from light at sea level to dark at highest mountains.
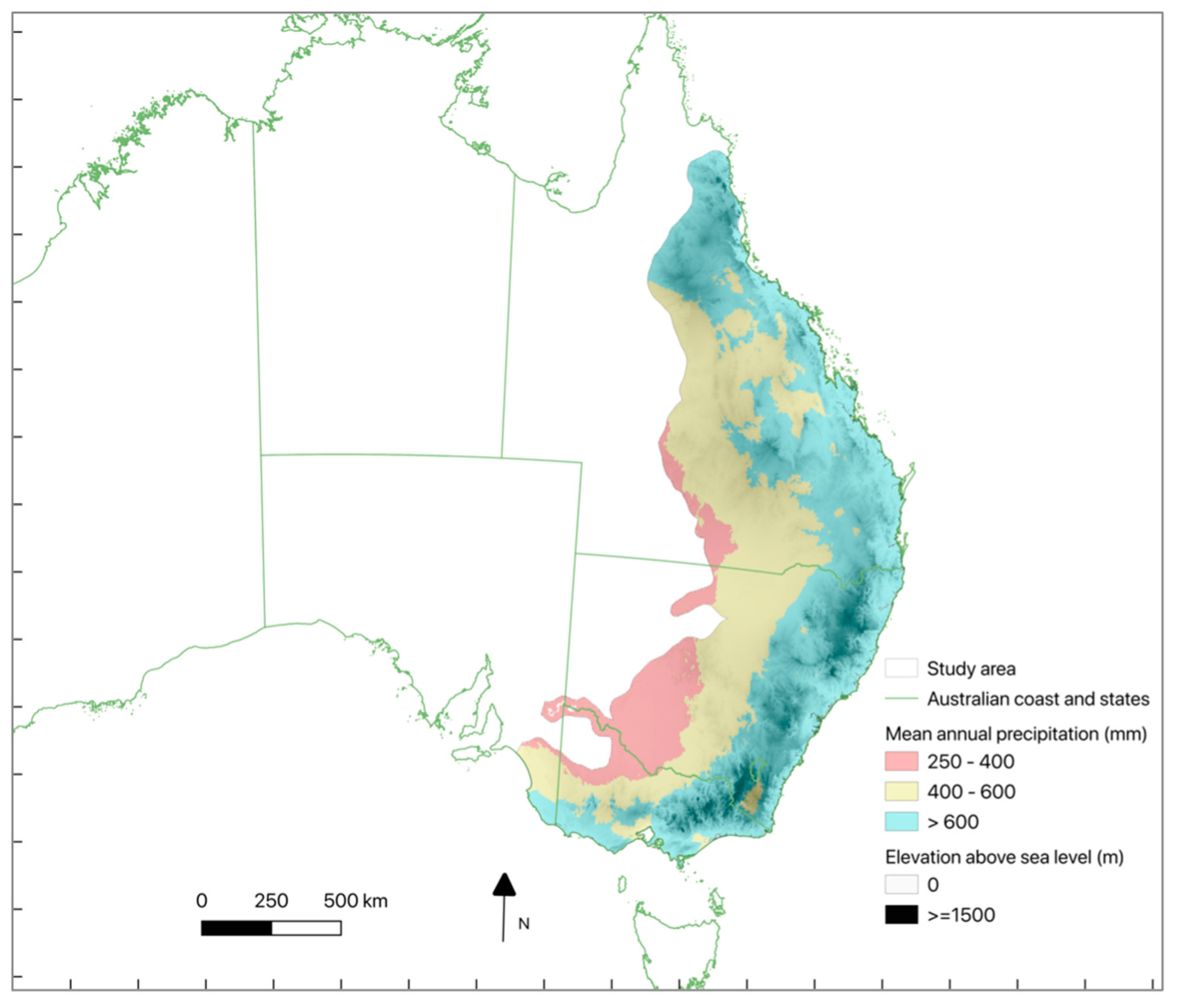
Figure 2.
Violin plot of NDVI by vegetation group (colour) and data type (letters & shades): koala site (K, dark) and random landscape location (L, light). n = 4,500. Landscape NDVI is random in space, within the respective vegetation group, paired in time to koala site NDVI which is the NDVI value at time and place of observations from anytime in the study period (1995-2009). Each violin is scaled to have the same area. Shown inside the violin is a simplified boxplot with a white dot at the median.
Figure 2.
Violin plot of NDVI by vegetation group (colour) and data type (letters & shades): koala site (K, dark) and random landscape location (L, light). n = 4,500. Landscape NDVI is random in space, within the respective vegetation group, paired in time to koala site NDVI which is the NDVI value at time and place of observations from anytime in the study period (1995-2009). Each violin is scaled to have the same area. Shown inside the violin is a simplified boxplot with a white dot at the median.
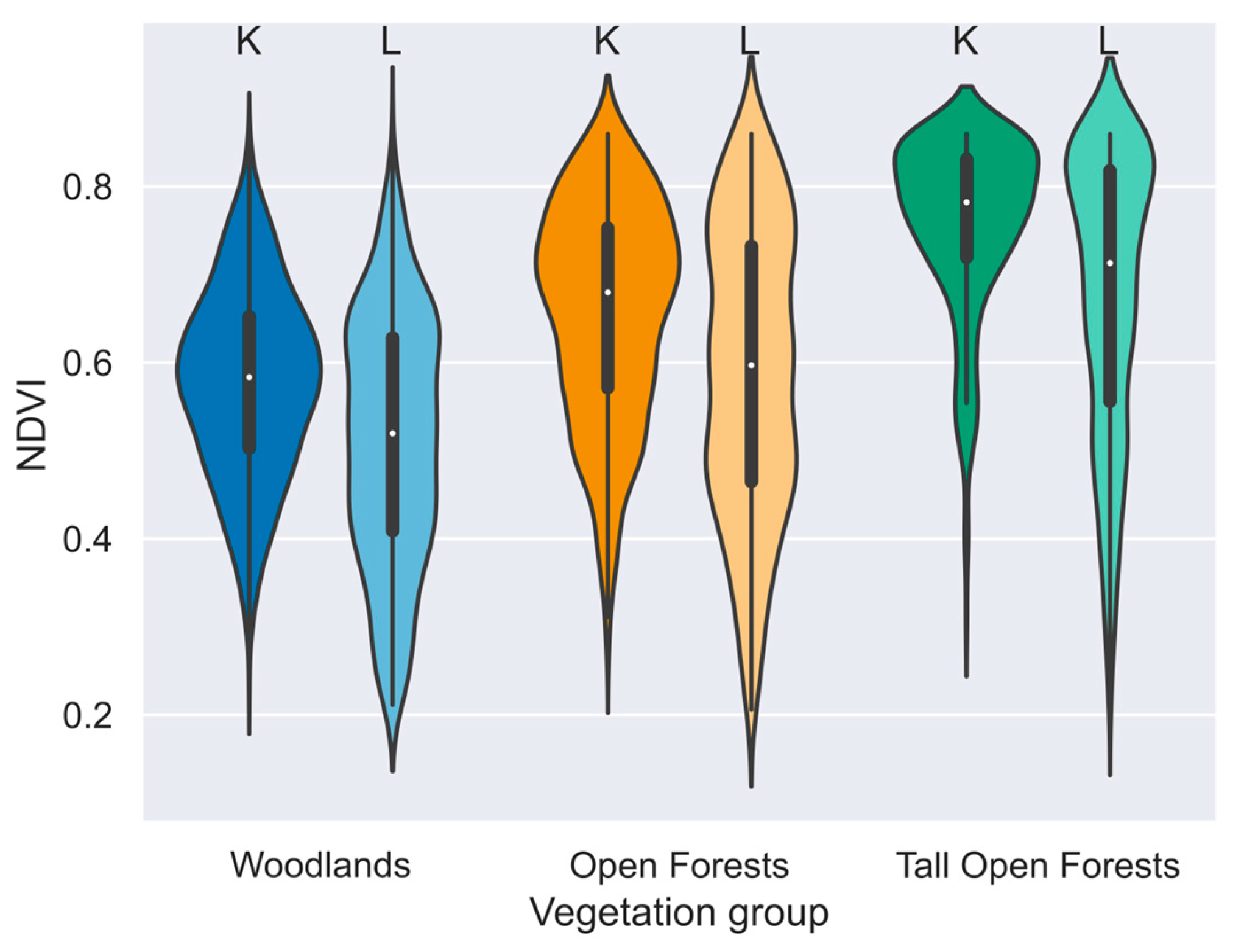
Figure 3.
Combined effect size plot of three binomial generalised linear mixed models, one per vegetation group, to determine whether NDVI is a significant explanatory variable for the differences between where koalas have been observed and random pixels across the koala range within the same vegetation groups. Effect sizes are logarithm odds ratios. Error bars represent 95% confidence intervals with those not crossing zero indicating a significant effect. P-values; Woodlands = 3.58e-05, Open Forests = 8.41e-12, Tall Open Forests = 2.71e-12. n = 4500 (i.e. n = 1500 per vegetation group/model). Equation: GLMM = Class ~ NDVI + SpatialCovariance (where class is koala site or random location and NDVI is standardised).
Figure 3.
Combined effect size plot of three binomial generalised linear mixed models, one per vegetation group, to determine whether NDVI is a significant explanatory variable for the differences between where koalas have been observed and random pixels across the koala range within the same vegetation groups. Effect sizes are logarithm odds ratios. Error bars represent 95% confidence intervals with those not crossing zero indicating a significant effect. P-values; Woodlands = 3.58e-05, Open Forests = 8.41e-12, Tall Open Forests = 2.71e-12. n = 4500 (i.e. n = 1500 per vegetation group/model). Equation: GLMM = Class ~ NDVI + SpatialCovariance (where class is koala site or random location and NDVI is standardised).
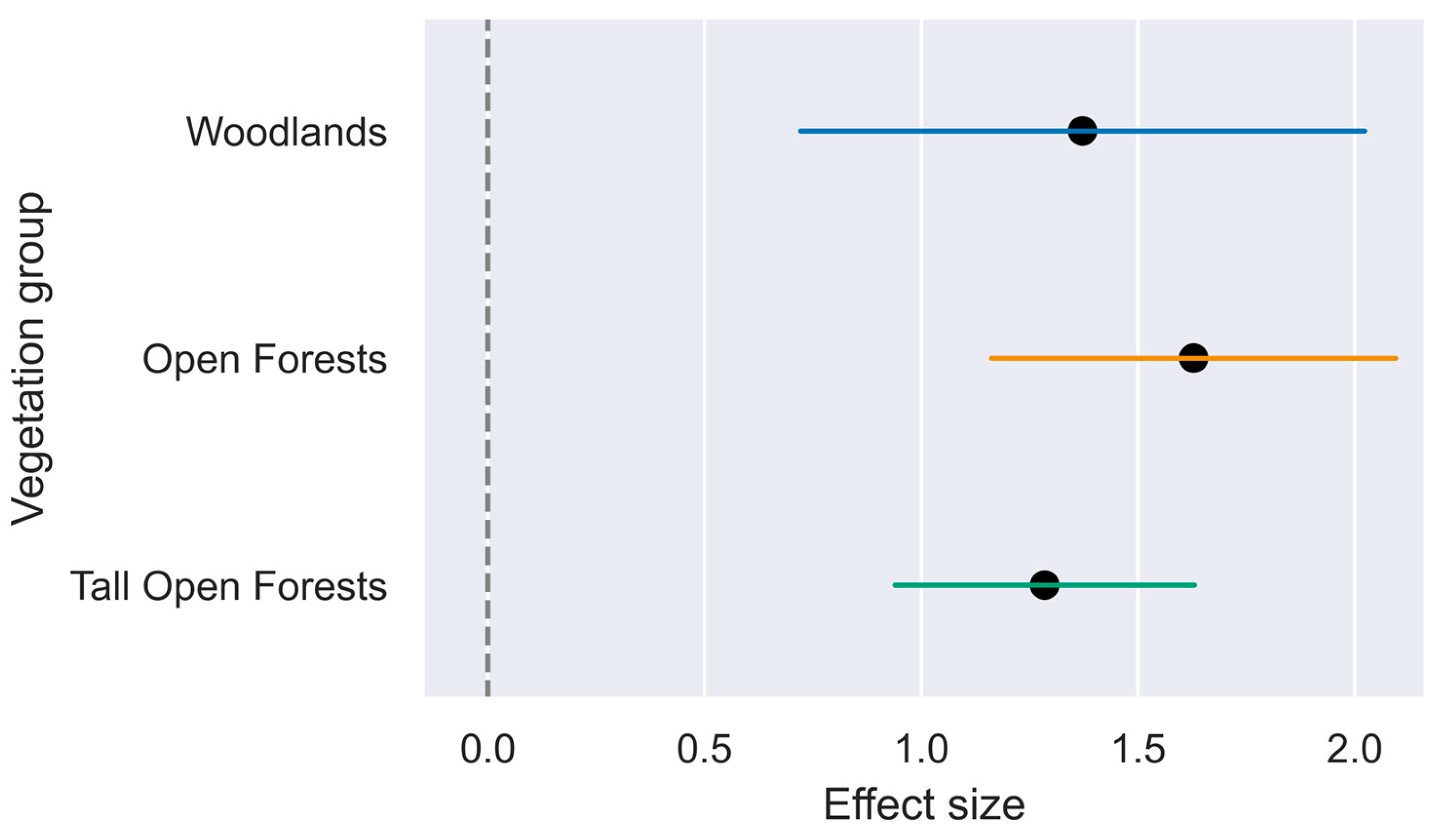
Figure 4.
Empirical cumulative distributions of NDVI in koala sites at time of observation with the 5th percentile thresholds overlayed as dashed lines (i.e. Proportion = 0.05). The colours represent vegetation groups. The cut-off in maximum NDVI=0.86 is visible, this threshold was used as an upper bound of the data to avoid potential saturation of NDVI, which is known to occur above this level for this data type.
Figure 4.
Empirical cumulative distributions of NDVI in koala sites at time of observation with the 5th percentile thresholds overlayed as dashed lines (i.e. Proportion = 0.05). The colours represent vegetation groups. The cut-off in maximum NDVI=0.86 is visible, this threshold was used as an upper bound of the data to avoid potential saturation of NDVI, which is known to occur above this level for this data type.
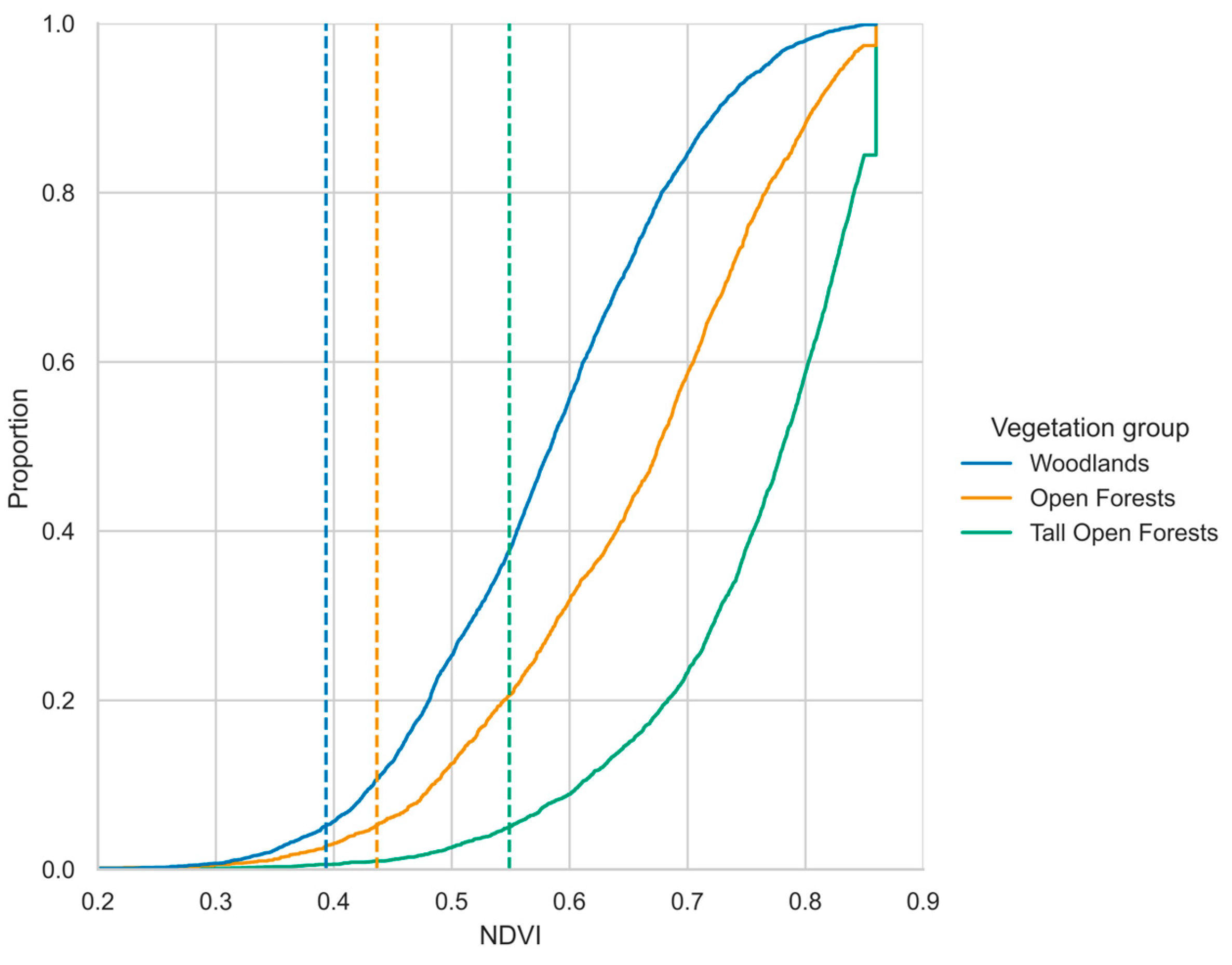
Figure 5.
Difference between pre-drought (1995-1999) and drought (2005-2009) periods in NDVI minimum (5th percentile) of vegetation groups in the landscape. NDVI minimum is the 5th percentile across time for each pixel. Boxes are interquartile range with a median dividing line.
Figure 5.
Difference between pre-drought (1995-1999) and drought (2005-2009) periods in NDVI minimum (5th percentile) of vegetation groups in the landscape. NDVI minimum is the 5th percentile across time for each pixel. Boxes are interquartile range with a median dividing line.
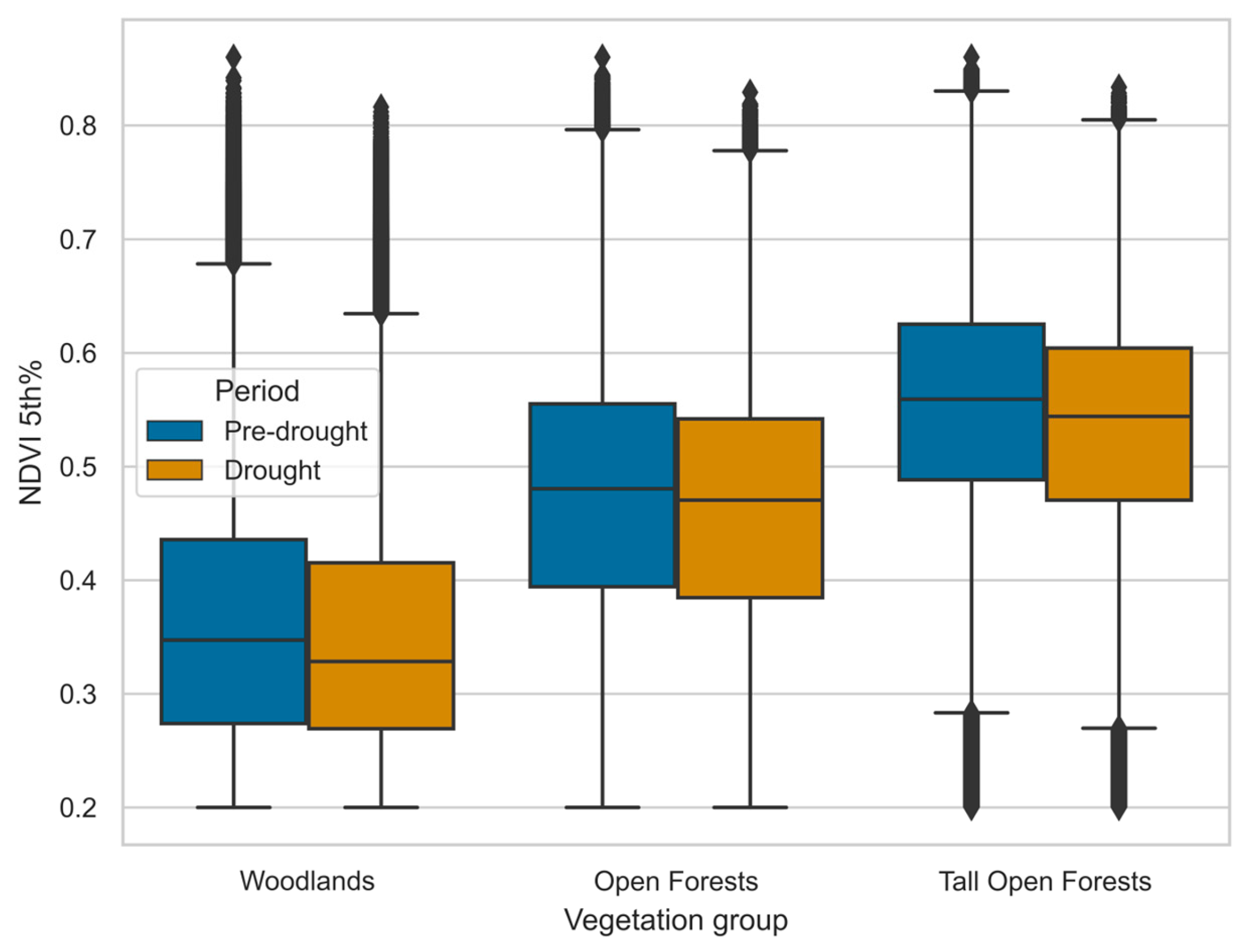
Figure 6.
Maps of potential koala habitat that was a) above the thresholds in 1995-1999 (pre-drought), b) potential habitat that had been above thresholds in 1995-1999 that was now below thresholds in 2005-2009 (drought).
Figure 6.
Maps of potential koala habitat that was a) above the thresholds in 1995-1999 (pre-drought), b) potential habitat that had been above thresholds in 1995-1999 that was now below thresholds in 2005-2009 (drought).
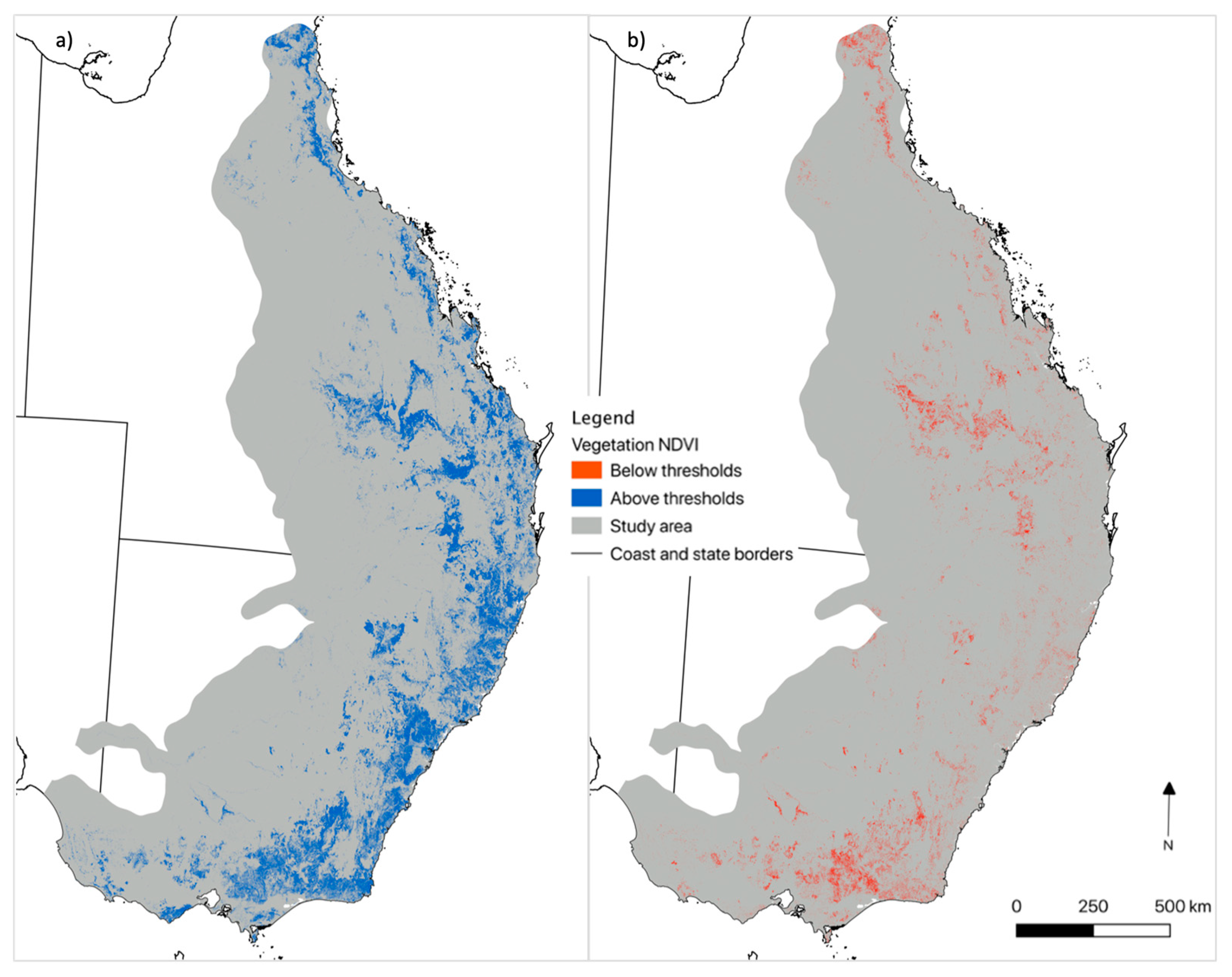
Figure 7.
Map of potential koala habitat in the Liverpool Range, NSW that was above (blue) the thresholds in 1995-1999 (pre-drought), and vegetation that had been above in 1995-1999 that was now below (red) in 2005-2009 (drought). Central coordinate: 150.23443, -31.93875 (coordinate system = EPSG:4326). Basemap is the NSW Imagery dataset, accessed via https://datasets.seed.nsw.gov.au/dataset/5601b661-3352-47ad-a8e5-09278927226f.
Figure 7.
Map of potential koala habitat in the Liverpool Range, NSW that was above (blue) the thresholds in 1995-1999 (pre-drought), and vegetation that had been above in 1995-1999 that was now below (red) in 2005-2009 (drought). Central coordinate: 150.23443, -31.93875 (coordinate system = EPSG:4326). Basemap is the NSW Imagery dataset, accessed via https://datasets.seed.nsw.gov.au/dataset/5601b661-3352-47ad-a8e5-09278927226f.
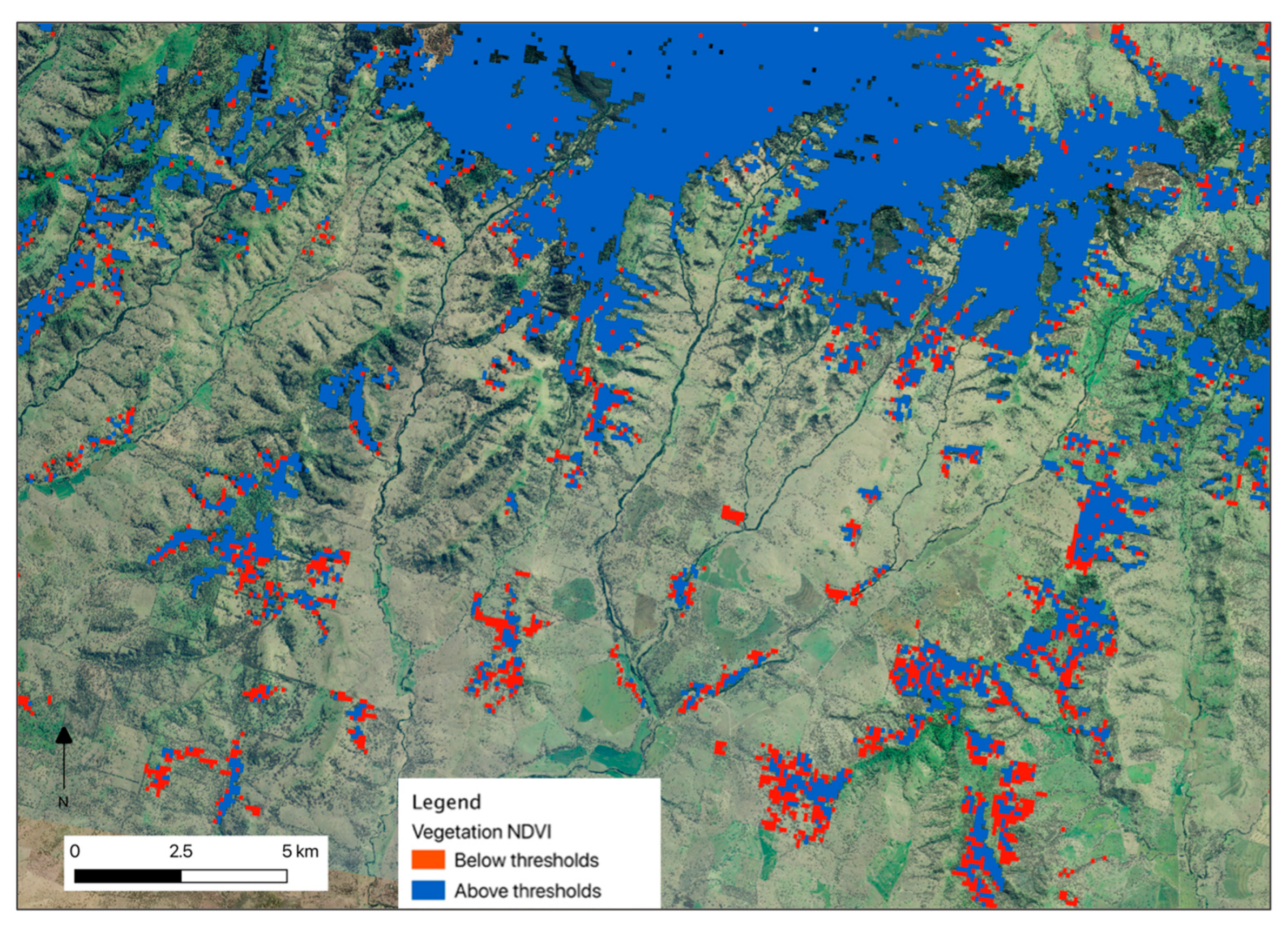
Table 2.
Vegetation areas above-thresholds, per group, in the landscape minimum NDVI data for 1995-1999 and 2005-2009 time periods. Thresholds based on koala observation sites, identified by the lowest likely NDVI (5th percentile) of each vegetation group.
Table 2.
Vegetation areas above-thresholds, per group, in the landscape minimum NDVI data for 1995-1999 and 2005-2009 time periods. Thresholds based on koala observation sites, identified by the lowest likely NDVI (5th percentile) of each vegetation group.
| Area of Vegetation above the NDVI Thresholds (km2) | Difference between Periods (%) | ||||
|---|---|---|---|---|---|
| Vegetation Group | Pre-Drought 1995-1999 |
Drought 2005-2009 |
|||
| Woodlands | 100,305 | 84,370 | -15.8% | ||
| Open Forests | 54,904 | 51,180 | -6.8% | ||
| Tall Open Forests |
35,017 | 31,195 | -10.9% | ||
| All groups | 190,227 | 166,746 | -12.3% | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



