
Submitted:
20 June 2024
Posted:
21 June 2024
You are already at the latest version
Abstract
Despite advances in our knowledge of systemic lupus erythematosus (SLE) has increased over time, there are still many challenges in deciphering the precise mechanisms involved in the development and progression of the disease. Recent evidence has raised questions about the effectiveness of the programmed cell death protein 1 (PD-1) in suppressing autoreactive CD4+ T cells during the autoimmune response. Research has ventured into investigate the potential impact of PD-1 on various CD4+ T cell subpopulations, including T follicular helper (Tfh) cells, circulating Tfh (cTfh) cells, and T peripheral helper (Tph) cells, all of which exhibit substantial PD-1 expression and have been closely related with several autoimmune disorders including SLE. This review aims to highlight the complex role of PD-1 in autoimmunity and emphasizes the imperative for further research to elucidate its functions during autoreactive T-cell response. Additionally, we address the potential of PD-1 or its ligands as a possible therapeutic target in SLE.
Keywords:
PD-1
; PD-L1/PD-L2
; Systemic lupus erythematosus
; T follicular helper (Tfh) cells
; T peripheral helper (Tph) cells.
1. Introduction
Autoimmune diseases are reported to affect 5–8% of the world population [1]. They have garnered significant attention in research due to the multitude of immunopathological pathways characterized by aberrant innate and adaptive responses and the absence of effective therapeutic interventions.
Systemic lupus erythematosus (SLE) is considered the prototype of autoimmune diseases due to the complexity of the molecular and immunological pathways involved in its development and progression [2]. Although the production of autoantibodies targeting nuclear antigens is characteristic of the disease, this process involves a wide range of mechanisms. One of the most important is the differentiation and maturation of T cell-dependent B cells. During this process, CD4+ T cells that have evaded central and peripheral tolerance mechanisms collaborate with B cells, providing them with the necessary signals for their complete differentiation into antibody-producing plasma cells or long-lived memory cells [3].
During this interaction, T cells also require co-stimulatory signals for their optimal activation, differentiation, and function. However, ultimately, they also require inhibitory signals to maintain immune balance, limit auto-reactivity, and prevent tissue injury. These signals, known as immune checkpoints, are added to co-stimulatory signals to inhibit T cell receptor (TCR) pathway signaling, thus counteracting cellular overactivation and contributing to immune response control. Nevertheless, it has been reported that dysregulation in the function or expression of these checkpoints leads to a failure in controlling T cell exhaustion, favoring autoimmunity [4].
One of the most important is the programmed cell death protein 1 (PD-1) receptor, which has been reported to prevent autoimmunity since its deficiency or the blockade of the PD-1 signaling pathway exacerbates disease progression in several autoimmune mouse models [5,6,7,8,9], as well as observations in which mutations in the PDCD1 gene are associated with susceptibility to several human autoimmune disorders [10,11,12,13]. PD-1 controls central and peripheral self-tolerance and prevents autoimmunity by suppressing autoreactive T cells. However, it has been reported that effector PD-1+/highCD4+ T cells are increased in patients with several autoimmune diseases associated with disease activity (reviewed in [14]), like SLE. This suggests that those cells are not functionally restricted by their high expression of PD-1.
This review provides a comprehensive overview of our current understanding of PD-1 and its association with the mechanisms involved in autoimmune responses with a special focus on SLE. We aim to provide potential explanations for why PD-1 may not effectively carry out its suppressive functions, even when overexpressed on CD4+ T cells, and raise important questions about PD-1's function for future research in this area.
2. PD-1 Expression and Function. What Do We know?
Our understanding of the PD-1 molecule began with identifying its cDNA in 1991 [15], followed by the discovery that PD-1 transcript expression occurred in T cells just before their activation and that its function was associated with inducing cell death. Interestingly, it also was observed that mice lacking PD-1 expression exhibited abnormal activation of both B and T cells [16]. It was further demonstrated that these PD-1-deficient mice developed some diseases such as cardiomyopathy, arthritis, and nephritis, highlighting its importance not only as a negative regulator of the immune response but also in developing autoimmune-related diseases [17,18,19].
PD-1 (CD279) is a signal-transducing type I protein composed of 260 amino acids, that is expressed on the cell surface of activated T cells, B cells, thymocytes, natural killer (NK) cells, natural killer T (NKT) cells, macrophages, and dendritic cells (DC) [20,21,22]. Structurally, PD-1 is composed of an extracellular immunoglobulin variable region (called (IgV)-like domain), followed by a constant region (called (IgC)-like domain) a transmembrane domain, and an intracellular domain. The (IgV)-like domain of PD-1 contains the binding site for its ligands, while the intracellular domain contains two tyrosine residues: an immunoreceptor tyrosine-based inhibitory motif (ITIM) and an immunoreceptor tyrosine-based switch motif (ITSM) [23,24]. PD-1 has two major ligands known as PD-L1 (CD274) [25] and PD-L2 (CD273) [26], that share the extracellular structure with PD-1 (Figure 1). PD-1 belongs to the CD28/CTLA-4 superfamily with 21–33% sequence identity with other family members like CTLA-4, CD28, and ICOS [27].
The PD-1 pathway plays a pivotal role in maintaining immune balance, serving as a negative regulator of T cell receptor (TCR) signaling and suppressing T cell activation triggered by auto-antigens, thus PD-1 prevents excessive activation following an adaptive immune response [28]. It has also been shown that PD-1 negatively regulates B cell responses by inhibiting B cell receptor (BCR) signaling [29].
During chronic infections, PD-1 signaling plays a central role in perpetuating T cell exhaustion, a state in which T cells progressively lose their effector functions after their activation [20,30,31,32]. Furthermore, it has been well-documented that tumor cells exploit the PD-1 signaling pathway to evade immune responses, indeed the use of antibodies that blockade the activation of this pathway has been widely used in cancer immuno-therapies [33]. However, despite the extensive knowledge regarding PD-1's involvement in cancer responses, its role in autoimmunity, particularly in the pathogenesis of diseases like SLE, is still largely unexplored.
3. PD-1/PD-L1/PD-L2 in Central and Peripheral Tolerance
The most widely accepted role for PD-1 in autoimmunity is during the mechanisms of autoreactive T cell deletion during central and peripheral cell tolerance [34]. The initial observations regarding the significance of PD-1 in thymic selection mechanisms were conducted by Hiroyuki Nishimura et al. in 2000. They investigated the impact of PD-1 deficiency on thymocyte differentiation, finding that PD-1 negatively regulates beta selection, while modulating positive selection, thus, PD-1 significantly altered the mature T cell repertoire [35]. These findings were subsequently corroborated by others that also indicated a role for PD-1 during thymic selection [36,37]. More recently, it was reported that mice lacking the expression of both PD-1 and the autoimmune regulator (AIRE) genes developed fatal adulthood autoimmunity. This phenomenon was not observed in mice lacking other combinations of tolerance mediators, suggesting that the cooperation between PD-1 and AIRE plays a pivotal role in mediating central tolerance and autoimmune development [38].
Peripheral tolerance of T cells in the context of PD-1 is mediated by the expression of PD-L1 and PD-L2 on antigen-presenting cells (APCs), such as dendritic cells, which through this expression determine T cell inhibition [39]. However, a decrease in the expression of PD-1 or its ligands, which activate the inhibitory pathway, could be closely linked with the development of autoimmune diseases including SLE (as discussed below). This is due to the lack of inhibition of autoreactive T cells.
In line with this, recent research has focused on studying two subpopulations of T helper (Th) cells whose frequencies appear to be increased and associated with the pathogenesis of some autoimmune conditions. T follicular helper (Tfh) cells are usually identified by a CD4+CXCR5+PD-1+ phenotype. These cells represent a specialized subpopulation primarily located within germinal centers (GC) of secondary lymphoid organs, which play a crucial role in mediating the differentiation of B cells into autoantibody-producing cells, a function that is primarily achieved through the expression of IL-21 [40]. IL-21 is a key regulator of various processes within the immune system and autoimmunity. It is the main cytokine required to enable mechanisms of germinal center formation, affinity maturation, and generation of plasma cells and memory B cells [41]. Furthermore, IL-21 is also required for the complete generation of Tfh cells [42,43]. A small proportion of Tfh cells called circulating Tfh (cTfh) cells can be found in the peripheral blood, and they have been extensively associated with autoimmune diseases such as systemic lupus erythematosus (SLE) [44,45,46,47,48], (RA) [49,50,51], Sjögren's syndrome (pSS) [52,53,54,55,56] and multiple sclerosis (MS) [57,58]. Although Tfh cells had been the main CD4+ T cell subpopulation involved in maintaining the generation of autoantibodies, recent research has focused on a novel cell subpopulation that appears to be more closely associated with the pathogenesis of autoimmune diseases.
T peripheral helper (Tph) cells are characterized by a CD4+CXCR5-PD-1+/high phenotype. The absence of CXCR5 leads them to exit germinal centers and position themselves within inflamed tissues by expressing other chemokine receptors such as CCR2 and CCR5, where Tph cells induce B cell differentiation through the production of IL-21 [14]. Increased numbers of Tph cells have also been observed in SLE [59,60,61,62], RA [58,59,60,61], and pSS [54,67,68].
A shared characteristic between Tfh, cTfh, and Tph cells is the high expression of PD-1 on their cell surface, which is in some cases associated with the disease activity. Because the frequencies of cTfh and Tph cells are increased in these autoimmune diseases, it is reasonably suggested that PD-1 may not be exerting its suppressive functions on these cellular subpopulations, this potentially indicated a break in the peripheral tolerance mediated by an aberrant function of PD-1.
Another important subpopulation of T cells responsible for mediating peripheral tolerance and tissue damage are regulatory T cells (Treg), which are identified by presenting a CD4+CD25+Foxp3+ phenotype. In addition, they express activation markers and immunomodulatory molecules such as CTLA-4, PD-1, and PD-L1 [69]. Treg cells are divided into two groups: those that express Foxp3 and differentiate in the thymus known as natural Tregs (nTreg), while those that differentiate from naïve CD4+ T cells in the periphery or through in vitro stimulation known as peripheral (pTreg) or induced (iTreg) Treg cells [70]. Increasing evidence has shown that the PD-1 pathway is associated with the differentiation of pTreg cells. Francisco, Loise M, et al, demonstrated that PD-L1-deficient antigen-presenting cells failed to induce the polarization of naïve CD4+ T cells into pTreg cells. When CD4+ T cells were primed with PD-L1 in vitro, iTregs were induced, demonstrating that PD-L1 maintained Foxp3 expression and the suppressive function of iTreg cells [71]. This could contribute to our understanding of how the deficiency in PD-L1 expression by APC is associated with the development of autoimmune diseases. Similar observations have been made for Th1 cells, which, in a PD-L1 manner, polarized to Treg cells [72].
Recently it was demonstrated higher expression of PD-1 but lower expression of PD-L1 on CD4+CD25+FOXP3+ Treg cells after stimulation in cell culture, which was negatively associated with the SLEDAI index in SLE patients [73]. Furthermore, it has been shown that the stimulation of PD-1 with specific agonists can inhibit autoreactive T cells and restore Treg cell homeostasis [74], highlighting the importance of the PD-1 pathway Treg cells and peripheral tolerance (Figure 2).
4. Regulation of PD-1 Expression and Its Relationship with the Autoimmune Response in SLE
To understand the role of PD-1 in the autoimmune response in SLE and other autoimmune disorders it is important to consider how the expression of PD-1, its ligands, and the downstream signaling pathways are regulated.
4.1. Stimulation of PD-1 Expression
PDCD1 located in chromosome 2 is the gene encoding PD-1 in humans, whereas Pdcd1 located in chromosome 1 is their counterpart in mice. PD-1 transcription initiates with the TCR activation and cytokine stimulation that activate various transcription factors including NFATc1 (activated by TCR signaling) [75], STAT3, STAT4, and STAT5 (activated by IL-6, IL-12, IL-2, IL-21) [76,77], the ISGF3 complex (activated by IFNα) [78], and FOXO1, AP-1, NF-κB (activated by other various cytokines) [79,80]. These transcription factors directly bind to specific enhancers and promoters located in cis-regulatory elements comprising two conserved regions: Conserved Region B (CR-B) and Conserved Region C (CR-C), of the PD-1 gene in both humans and mice [81] (Figure 3A).
This could offer an initial explanation for the overexpression of PD-1 on the surface of CD4+ T cells, like cTfh and Tph cells, especially in autoimmune diseases such as SLE, where there is constant and repetitive stimulation of T cells by self-antigens and proinflammatory cytokines like IL-6, IL-21, and IFNα [82]. After translation, PD-1 is stabilized by fucosylation in the endoplasmic reticulum and Golgi apparatus [83]. The presence of PD-1 has been identified in vesicles proximal to the Golgi and trans-Golgi network, hinting at the potential role of these vesicles as reservoirs for PD-1 awaiting activation through TCR [84].
4.2. Repression of PD-1 Expression
Other transcription factor-binding sites near CR-B and CR-C serve for repression of the PD-1 expression. These include an RBP-Jκ-binding site upstream of CR-C [85], a Blimp-1-binding site downstream of CR-C [86], and a T-bet-binding site at -0.5 kb of Pdcd1 [87].
Regarding this, it has demonstrated that increased expression of the transcription factor Blimp-1 was correlated with the overexpression of PD-1 in CD8+ T cells during chronic viral infection, in the same work was found that upon the conditional deletion of Blimp-1, there was also a reduction in the expression of this inhibitory receptor [88]. However, a couple of years later, it was demonstrated that PD-1 can be repressed by Blimp-1 in CD8+ T cells by the suppression of the expression of NFATc1, which acts as a transcriptional activator of Pdcd1 in acute viral infection [86]. Despite these contradictory initial results in response to the infection state, it is important to highlight that there is an association between Blimp-1 and PD-1 expression in CD8+ T cells. Despite Blimp-1 could play a different role in CD8+ and CD4+ T cells, there has not been explored the possible association between this transcription factor and the PD-1 regulation in CD4+ T cells during normal and autoimmune responses, even though Blimp-1 has been linked to exhausted protein markers such as PD-1 [89,90]. Tph cells are characterized by expressing the Blimp-1 transcription factor which serves as a mediator of its differentiation. As Tph cells overexpress PD-1 on their surface, could be interesting to study the mechanisms involved in the regulation of the PDCD1 gene by Blimp-1.
5. PD-1 Ligands Emerged as a New and Important Focus of Study in the Field of SLE
PD-L1 is coded by the Cd274 gene situated on chromosome 9 in humans and chromosome 19 in mice. Similarly, PD-L2 is encoded by the Pdcd1lg2 gene, which is found on chromosome 9 in humans and chromosome 19 in mice [91]. PD-L1 and PD-L2 belong to B7 superfamily members. Both are type I transmembrane glycoproteins composed of (IgV)- and (IgC)-like domains. Despite PD-L1 and PD-L2 sharing 30–40% of amino acid homology in humans, it has been reported that PD-L1 is the main ligan though PD-1 activates its downstream pathway [92].
PD-L1 and PD-L2 have distinct expression patterns by various components of the immune system as well as other tissue cells [93]. Both are expressed in activated T cells, B cells, dendritic cells, macrophages, and bone marrow-derived mast cells [94]. Additionally, PD-L1 is also expressed by NK cells, and non-immunological cells like endothelial, epithelial, and keratinocytes [95,96].
The PD-1/PD-L1/2 ligation is not restrictive, as PD-L1 can also bind to CD80, while PD-L2 can also bind to the repulsive guidance molecule b (RGMb) [97,98,99]. The binding between PD-L1 and CD80 occurs in cis (on the same cell surface) [100,101]. As the activation of the pathway needs the binding of PD-1 with any of PD-L1 or PD-L2, the availability of the ligands is necessary for the successful inhibitory function of PD-1 in T cells. Therefore, PD-L1/CD80 cis-binding on antigen-presenting cells (APCs) can reduce the number of PD-L1 available for trans-binding interaction (cell to cell) with PD-1 expressed on T cells. This not only leads to diminished PD-1 inhibitory signaling [102] but also reduced CTLA4-CD80 immune regulatory axes [103]. However, the PD-L1/CD80 interaction does not obstruct the binding of CD80 to CD28, consequently, they can form a trimeric complex that engages CD28 and thereby conveys a normal co-stimulatory signal [102] (Figure 3B).
Exploring the availability of PD-L1/L2 to PD-1 in SLE could yield valuable insights, potentially positioning the interaction as a promising therapeutic target in autoimmune responses, as it has been demonstrated in a recent study that the interaction between astrocytic PD-L1 with microglial PD-1 is necessary for the attenuation of autoimmune central nervous system inflammation in acute and progressive stages of a mouse model of multiple sclerosis [104]. Also, T cells from the synovial fluid of active rheumatoid arthritis patients exhibited decreased PD-1-mediated inhibition of their proliferation, especially at suboptimal concentrations of PD-L1. The treatment of those cells with PDL-1.Fc antibodies demonstrate an improvement in the reduction of their T cell response and ameliorate the severity of collagen-induced arthritis (CIA) [105]. In line with this, it has been demonstrated that the disruption of PD-L1/CD80 cis-binding by CD80 antibodies can alleviate autoimmunity due to the accessibility of PD-1/PD-L1 interaction [106].
The inhibitory PD-1/PD-L1/2 pathway can also be restricted by the soluble form of PD-1 (sPD-1). sPD-1 is generated from an alternative splicing event where the exon3, which contains the coding sequence for the transmembrane domain of PD-1 is deleted from the mRNA transcript. The expression of this transcript is upregulated following T-cell activation, leading to the release of sPD-1 into the plasma [107]. It has been reported that sPD-1 can interfere with PD-L1 or PD-L2, preventing the binding of the ligands to the transmembrane form of PD-1, and consequently blocking the negative signal [108] (Figure 3C). In this regard, it has been found elevated levels of sPD-1 in patients with autoimmune diseases such as SLE [109,110,111], RA [112,113,114], pSS [115,116], and the antineutrophil cytoplasmic antibody (ANCA)-associated vasculitides (AAV) [117], that could be promoting the lack of cellular inhibition by blocked the inhibitory effect of membrane-bound PD-1 on T-cell activation.
PD-L1 and PD-L2 can also be found in a soluble form (sPD-L1 and sPD-L2, respectively), which are generated through proteolytic cleavage of membrane-bound protein by metalloproteases like ADAM or metalloproteinases (MMPs) [118], or by alternative splicing that induces a protein product that lacks the transmembrane domain and is secreted in a soluble form [119,120]. In the context of cancer, it has been reported that sPD-L1 influences the activation of the inhibitory function of PD-1 expressed in T cells by binding to PD-1 and leading to the activation of the downstream pathway [121]. However, the role of sPD-L2 in the activation of the inhibitory pathway remains unclear. Understanding the mechanisms of how sPD-L1 and sPD-L2 influence the pathogenesis of SLE and other autoimmune disorders could be challenging at this moment due to varying reports. While some authors indicated elevated levels of these ligands compared to healthy controls, others reported decreased levels, and in some autoimmune disorders, there is a lack of sufficient information. In Table 1 we summarize some of the findings of PD-1 and its ligands as well as the soluble forms in SLE and other autoimmune diseases.
6. Regulation of PD-L1 and PD-L2 Expression and Its Association with the Autoimmune Response
It is well known that the major inducers for PD-L1 and PD-L2 expression are interferon (IFN) cytokines including type I (IFNα, IFNβ) and type II (IFNγ) interferons. IFN-pathway through a JAK/STAT signaling promotes the activation and expression of the interferon regulatory transcription factor 1 (IRF1) that induces the amplification of the PD-L1/2 gene by direct binding to the PD-L1 promoter [142]. After PD-L1 (and probably PD-L2) translation, the protein can be modified by post-translational modifications such as glycosylation, acetylation, tyrosine phosphorylation, and mono-ubiquitination upon epidermal growth factor (EGF) stimulation [143]. The knowledge about post-translational modifications in PD-L1 has primarily stemmed from studies focused on cancer. However, there are currently no works evaluating the effect of these modifications in the context of autoimmunity. Therefore, it would be important to understand the regulatory mechanisms of both PD-L1 and PD-L2, as they could serve as potential therapeutic targets.
PD-L1 protein can be stabilized by glycosylation in N35, N192, N200, and N219 residues increasing the half-life of the protein and regulating the PD-1/PD-L1 interaction [144]. PD-L1 has an extra glycosylation site at N64 (91). PD-L1 palmitoylation in the C272 site of cysteine residues [145,146], and phosphorylation in Try112 [147] have also been implicated in stabilizing the protein and cell surface distribution in some cancer cells. In contrast, some modifications like ubiquitination mediated by the CDK4 cyclin and phosphorylation in S195 of PD-L1 induce its degradation [147].
Recent findings have also demonstrated the association of two proteins members of the chemokine-like factor (CKLF)-like MARVEL (CMTM) family with the stability of PD-L1 expression in cancer and normal myeloid cells of both humans and mice [148,149]. CMTM6 is a type 3 transmembrane protein that binds to PD-L1, maintaining its expression on the cell surface without modifying the constitutive IFN-induced mRNA expression. CMTM6 is also found in recycling endosomes with PD-L1, preventing its ubiquitination and degradation by lysosomes. Consequently, the depletion of CMTM6 leads to a reduction in PD-L1 expression on the cell surface [148,149]. CMTM4 is another member of this family that shares 55% sequence similarity to CMTM6 but has received less attention in research, despite also demonstrating similar functions to CMTM6 in the regulation of PD-L1 [149].
CMTM6 and CMTM4 are just being explored in autoimmune diseases. Using a genetic association network model, Davis et al. conducted a resequencing analysis of exomes from SLE patients and healthy individuals, identifying CMTM4 as a new strong candidate for study in SLE as it emerged as an implicated susceptibility gene [150], this discovery suggests that CMTM4 may play a role in the pathogenesis of SLE, potentially contributing to our understanding of the disease's genetic basis and offering new avenues for research into its mechanisms and therapeutic interventions.
Another study performed in pSS disease that analyzed the serum levels of CMTM6, found increased concentrations of the molecule in pSS patients compared with non-pSS and healthy controls and found up-regulated expression of CMTM6 in labial glands of those pSS patients. In this study, CMTM6 also positively correlates with sPD-1 and sPD-L1 [116]. Recently, it was reported that lower PD-L1 expression in monocytes of patients with AVV was associated with increased lysosomal degradation of PD-L1 because of reduced expression of CMTM6 [151]. Further studies of CMTM6 and CMTM4 are needed to understand their implication in PD-L1 or PD-L2 in the autoimmune context (Figure 4).
7. Is It Possible that PD-1/PD-L1/2 Has also Another Important Role beyond Inhibition?
The binding of PD-1 with its ligands initiates a cascade of signals that results in the inhibition of T-cell signaling. When PD-1 binds to PD-L1/2, the ITIM, and ITSM domains are phosphorylated. This phosphorylation leads to the activation of the phosphatase Src homology region domain-containing phosphatase 2 (SHP2) and its requirement to the ITSM domain. SHP2 then dephosphorylates the kinases ZAP70, PI3K, and Ras, resulting in the restriction of the downstream TCR signaling pathway [21,22]. PD-1 can also inhibit the CD28 co-stimulatory signaling by limiting the PI3K/AKT, resulting in a strong inhibition of T cell activation [152]. In B cells, the PD-1-PD-L1/2 interaction also results in the phosphorylation of the tyrosine residue in ITIM/ITSM domains that recruits SHP1/SHP2. SHP1/SHP2 further dephosphorylates B-cell receptor (BCR) proximal signaling molecules that attenuate the activation of the downstream pathway, leading to the loss of B-cell functions [153]. However, despite the well-defined signaling pathway downstream of PD-1, the question remains open: is it possible that PD-1 may exert other functions beyond the inhibition of the cellular response?
In this context, a phosphoproteomic analysis revealed that not all the pathways downstream of PD-1 are inhibitory. Instead, PD-1 ligation resulted in increased phosphorylation at several tyrosine sites, some of which were associated with the activation of cellular functions [154]. This notion was further supported by studies in which it was observed that following PD-1 ligation, the transcriptional signature of proliferating CD4+ T cells was enrichment in genes associated with an activated state. Furthermore, they observed that the genes induced after PD-1 ligation significantly differed between PD-L1 and PD-L2 stimulation [155,156]. These observations are consistent with the idea that PD-1 may be involved in regulating specific cellular functions beyond its inhibitory role.
Given that PD-1 is a marker for GC-resident Tfh cells [157], and that cTfh, as well as Tph cells, express high levels of PD-1 on their cell surface as discussed above, it could be interesting to assess whether or not is it a role played by this protein in the differentiation or function of these cells, that have been mainly implicated with the pathogenesis of SLE and other autoimmune disorders.
The idea that PD-1 is important in germinal center responses potentially leading to antibody production has been increasing over the years. In 2010, Good-Jaboson et al. [158] demonstrated that PD-1 regulated the survival of germinal center B cells and the development of long-lived plasma cells. They observed that PD-1 was upregulated in Tfh cells, while PD-L1 and PD-L2 were upregulated in germinal center B cells. In their study, they also found that mice deficient in PD-1, PD-L2, or both PD-L1/L2 exhibited reduced numbers of long-lived plasma cells [158]. However, how the does the deficiency of PD-1 or its ligands is related to impaired humoral responses remains a question. They proposed that the loss of PD-1 signaling resulted in an increased number of Tfh cells but reduced synthesis of IL-4 and IL-21 mRNA by these cells. This, in turn, leads to a deficient cooperative response, as is well known that IL-21 is required for optimal differentiation and maturation of long-lived plasma cells and antibody production [159]. This notion was supported using Pdcd1–/– mice and demonstrating that the deficiency of PD-1 led to an increased expression of BCL6 in Tfh cells, and increased numbers of these cells within the germinal center. However, the expression of interferon regulatory factor 4 (IRF4), which is essential to produce IL-21, was diminished in these cells. Consequently, Tfh cells from Pdcd1–/– mice produced less IL-21 than those from wild-type (WT) mice [160]. More recently, a study conducted in bone marrow chimeras, where both PD-L1-sufficient and -deficient B cells were evaluated, also demonstrated that interactions between PD-1 on Tfh cells and PD-L1 on B cells optimized the competitiveness and affinity maturation of the B cells by controlling Tfh cells positioning and function [161]. They propose that PD-1 has a bystander signaling mode for the recruitment of Tfh cells into the follicle. This function is carried out in synergy with ICOS, another member of the B7-family of co-stimulatory molecules that is important for Tfh cell development [162]. Furthermore, it is discussed that both ICOS and PD-1 are required for optimal IL-21 production by Tfh cells, which could be a possible explanation for why these cells express high levels of PD-1 on their surface. However, it also has been demonstrated that IL-21 is essential to produce PD-1 [163], which suggests a positive feedback loop.
Most of the information available about the functions performed by PD-1 and its ligands has been obtained from studies involving CD8+ T cells in cancer, where CD8+ T cells expressing PD-1 have been considered as an exhausted T cell subpopulation [164]. However, it appears that the functions of PD-1 may vary depending on the context of the disease and the T cell subpopulation expressing it. In this sense, Tfh, cTfh, and Tph cells are distinguished by their high expression of IL-21 [50,54,165], the cytokine through which they perform their primary helper functions, along with the expression of other cytokines during the humoral response. Therefore, it is suggested to evaluate whether these cells can be considered as an exhausted T cell subpopulation and new studies are necessary to elucidate the role of PD-1 beyond inhibition in CD4+ T cells in the context of autoimmune response.
8. PD-1 as a Possible Therapeutic Target in SLE
The proposal to develop biological agents targeting co-inhibitory pathways to treat patients with autoimmune diseases is not new [166]. Based on the discussion above, it is clear that a better understanding of the PD-1/PD-L1/L2 axes in SLE and other autoimmune diseases could lead to the design of new therapeutic strategies that can exclusively target the PD-1 pathway or be combined with other treatments. From our perspective, two therapeutic approaches could be followed: facilitating PD-1's accessibility to its ligands and administering PD-1 agonists.
8.1. Accessibility between PD-1 and Its Ligands
Daisuke Sugiura et al. generated anti-CD80 antibodies that dissociate the cis-PD-L1-CD80 complex. This dissociation allows PD-L1 interaction with PD-1 leading to the activation and potentiation of the inhibition of T cells, resulting in the attenuation of autoimmune symptoms [106].
8.2. PD-1 Agonists
Previously, a study was conducted using a murine model to evaluate the impact of systemically activating PD-1 through the administration of PD-L1-Ig in SLE-prone mice. The results demonstrated that PD-L1-Ig activated the PD-1 pathway, leading to the suppression of Th17 cell generation, a decrease in cytokine levels, including IFN-γ, IL-17, and IL-10, as well as reduced production of anti-dsDNA autoantibodies. These effects collectively contributed to the mitigation of kidney disease and an extension of the survival of these SLE-prone mice [167]. Using computational design, a small molecule named PD-MP1 was created to specifically bind to both human and murine PD-1 at the PD-L1 interface. The trimerization of PD-MP1 acts as a PD-1 agonist leading to significantly inhibiting the activation of murine T cells, this suggests that PD-MP1 holds potential as a PD-1 agonist for the treatment of autoimmune diseases and inflammatory conditions [168]. Also, it has developed PD-1 agonist molecules called ImmTAAI. These molecules can bind to target cells and imitate PD-L1 function, leading to highly effective activation of the PD-1 receptor on interacting T cells, and therefore achieve immune suppression [169]. The development of nanoparticles coated with dexamethasone has shown therapeutic efficacy in improving lupus nephritis (LN) in MRL/lpr mice. They specifically target CD4+ T, activating the PD-1 and TIGIT (T cell immunoglobulin and ITIM domain) signaling, leading to the inhibition of effector T cells, and enhancing the immunosuppressive activities of Treg [170]. More recently, natural autoantibodies that downregulate specific autoimmune disorders have been discovered. These autoantibodies are called regulatory rheumatoid factor (regRF). regRF specifically binds to PD-1 on activated T cells, and it has been reported that serum containing regRF reduces the number of PD-1+CD4+ lymphocytes. Therefore, RegRF utilizes the PD-1 pathway to regulate activated CD4+ T lymphocytes [171].
Although the potential of activating the PD-1 co-inhibitory pathway to alleviate autoimmunity is still being explored, it appears to be a promising therapeutic approach. In fact, there are PD-1 agonists ongoing in phase I and II in clinical trials, and researchers are collecting information to propose novel strategies such as combining low doses of IL-2 with PD-1 agonists [172]. Certainly, this has the potential to revolutionize conventional strategies and bring us closer to the era of precision medicine.
9. Conclusions
PD-1 is a molecule whose function appears to largely depend on the environment and context in which it is found. Most of the studies about PD-1 and PD-L1/2 expression, regulation, and function, have been made mainly on CD8+ T cells in the pathogenesis of cancer. However, this review emphasized the importance of directing new assays to elucidate the mechanisms operating PD-1’s function on CD4+ T cells during the autoimmune response, as it has been suggested that PD-1 could play a role in the differentiation and function of various T cell subpopulations such as Tfh, cTfh, and Tph cells, that are associated with some autoimmune disorders, which suggest a more complex regulation of this molecule.
We highlighted some of the most recent findings about PD-1 and PD-L1/L2 axis in systemic lupus erythematosus to demonstrate both their inhibitory and non-inhibitory functions and their association with disease activity. We suggest that a deeper understanding of the interaction between B and T cells in a PD-1-dependent manner could have potential value in identifying new therapeutic targets for the disease.
Author Contributions
Conceptualization, literature search, and writing—original draft preparation, Nefertari Sagrero-Fabela; literature search, and figure editing, Ramón Chávez-Mireles; review and editing, Diana Celeste Salazar-Camarena and Claudia Azucena-Pañafox Sánchez; supervision, Claudia Azucena-Pañafox Sánchez. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
All figures in this review were created with “BioRender.com” licensed by DCSC.
Data Availability Statement
Data is contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fugger L, Jensen LT, Rossjohn J. Challenges, Progress, and Prospects of Developing Therapies to Treat Autoimmune Diseases. Cell [Internet]. 2020 Apr 2 [cited 2023 Aug 8];181(1):63–80. Available from: http://www.cell.com/article/S0092867420302695/fulltext.
- Tsokos GC, Lo MS, Reis PC, Sullivan KE. New insights into the immunopathogenesis of systemic lupus erythematosus. Nature Reviews Rheumatology 2016 12:12 [Internet]. 2016 Nov 22 [cited 2024 May 13];12(12):716–30. Available from: https://www.nature.com/articles/nrrheum.2016.186.
- Pisetsky DS, Lipsky PE. New insights into the role of antinuclear antibodies in systemic lupus erythematosus. Nature Reviews Rheumatology 2020 16:10 [Internet]. 2020 Sep 3 [cited 2024 Jan 8];16(10):565–79. Available from: https://www.nature.com/articles/s41584-020-0480-7.
- Burke KP, Patterson DG, Liang D, Sharpe AH. Immune checkpoint receptors in autoimmunity. Curr Opin Immunol. 2023 Feb 1;80:102283.
- Reynoso ED, Elpek KG, Francisco L, Bronson R, Bellemare-Pelletier A, Sharpe AH, et al. Intestinal Tolerance Is Converted to Autoimmune Enteritis upon PD-1 Ligand Blockade. The Journal of Immunology [Internet]. 2009 Feb 15 [cited 2023 Sep 21];182(4):2102–12. Available from:. [CrossRef]
- Lucas JA, Menke J, Rabacal WA, Schoen FJ, Sharpe AH, Kelley VR. Programmed Death Ligand 1 Regulates a Critical Checkpoint for Autoimmune Myocarditis and Pneumonitis in MRL Mice. The Journal of Immunology [Internet]. 2008 Aug 15 [cited 2023 Sep 21];181(4):2513–21. Available from:. [CrossRef]
- Salama AD, Chitnis T, Imitola J, Akiba H, Tushima F, Azuma M, et al. Critical Role of the Programmed Death-1 (PD-1) Pathway in Regulation of Experimental Autoimmune Encephalomyelitis. Journal of Experimental Medicine [Internet]. 2003 Jul 7 [cited 2023 Sep 21];198(1):71–8. Available from: http://www.jem.org/cgi/doi/10.1084/jem.20022119.
- Ansari MJI, Salama AD, Chitnis T, Smith RN, Yagita H, Akiba H, et al. The Programmed Death-1 (PD-1) Pathway Regulates Autoimmune Diabetes in Nonobese Diabetic (NOD) Mice. Journal of Experimental Medicine [Internet]. 2003 Jul 7 [cited 2023 Sep 21];198(1):63–9. Available from: http://www.jem.org/cgi/doi/10.1084/jem.20022125.
- Nishimura H, Nose M, Hiai H, Minato N, Honjo T. Development of Lupus-like Autoimmune Diseases by Disruption of the PD-1 Gene Encoding an ITIM Motif-Carrying Immunoreceptor. Immunity. 1999 Aug 1;11(2):141–51.
- Kroner A, Mehling M, Hemmer B, Rieckmann P, Toyka K V., Mäurer M, et al. A PD-1 polymorphism is associated with disease progression in multiple sclerosis. Ann Neurol [Internet]. 2005 Jul 1 [cited 2023 Sep 21];58(1):50–7. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/ana.20514.
- Lee YH, Woo JH, Choi SJ, Ji JD, Song GG. Association of programmed cell death 1 polymorphisms and systemic lupus erythematosus: a meta-analysis. [Internet]. 2009 Jan 1 [cited 2023 Sep 21];18(1):9–15. Available from: https://journals.sagepub.com/doi/10.1177/0961203308093923?url_ver=Z39.88-2003&rfr_id=ori%3Arid%3Acrossref.org&rfr_dat=cr_pub++0pubmed.
- Gao J, Gai N, Wang L, Liu K, Liu XH, Wei LT, et al. Meta-analysis of programmed cell death 1 polymorphisms with systemic lupus erythematosus risk. Oncotarget [Internet]. 2017 Mar 18 [cited 2023 Sep 22];8(22):36885–97. Available from: https://www.oncotarget.com/article/16378/text/.
- Lee YH, Bae SC, Kim JH, Song GG. Meta-analysis of genetic polymorphisms in programmed cell death 1. Associations with rheumatoid arthritis, ankylosing spondylitis, and type 1 diabetes susceptibility. Z Rheumatol [Internet]. 2015 Apr 1 [cited 2023 Sep 22];74(3):230–9. Available from: https://pubmed.ncbi.nlm.nih.gov/24942602/.
- Huang Y, Ba X, Han L, Wang H, Lin W, Chen Z, et al. T peripheral helper cells in autoimmune diseases: What do we know? Front Immunol. 2023;14.
- Ishida Y, Agata Y, Shibahara K, Honjo T. Induced expression of PD-1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death. EMBO Journal. 1992;11(11):3887–95.
- Nishimura H, Minato N, Nakano T, Honjo T. Immunological studies on PD-1-deficient mice: implication of PD-1 as a negative regulator for B cell responses. Vol. 10, International Immunology. 1998.
- Okazaki T, Tanaka Y, Nishio R, Mitsuiye T, Mizoguchi A, Wang J, et al. Autoantibodies against cardiac troponin I are responsible for dilated cardiomyopathy in PD-1-deficient mice. Nat Med [Internet]. 2003 Dec [cited 2023 Jul 30];9(12):1477–83. Available from: https://pubmed.ncbi.nlm.nih.gov/14595408/.
- Nishimura H, Nose M, Hiai H, Minato N, Honjo T. Development of Lupus-like Autoimmune Diseases by Disruption of the PD-1 Gene Encoding an ITIM Motif-Carrying Immunoreceptor. Immunity. 1999;11:141–51.
- Nishimura H, Okazaki T, Tanaka Y, Nakatani K, Hara M, Matsumori A, et al. Autoimmune dilated cardiomyopathy in PD-1 receptor-deficient mice. Science [Internet]. 2001 Jan 12 [cited 2023 Jul 30];291(5502):319–22. Available from: https://pubmed.ncbi.nlm.nih.gov/11209085/.
- Jubel JM, Barbati ZR, Burger C, Wirtz DC, Schildberg FA. The Role of PD-1 in Acute and Chronic Infection. Front Immunol. 2020 Mar 24;11:524474.
- Boussiotis VA. Molecular and Biochemical Aspects of the PD-1 Checkpoint Pathway. New England Journal of Medicine [Internet]. 2016 Nov 3 [cited 2023 Sep 21];375(18):1767–78. Available from: https://www.nejm.org/doi/10.1056/NEJMra1514296.
- Schildberg FA, Klein SR, Freeman GJ, Sharpe AH. Coinhibitory Pathways in the B7-CD28 Ligand-Receptor Family. Immunity [Internet]. 2016 May 17 [cited 2023 Sep 21];44(5):955–72. Available from: http://www.cell.com/article/S107476131630156X/fulltext.
- Chemnitz JM, Parry R V., Nichols KE, June CH, Riley JL. SHP-1 and SHP-2 associate with immunoreceptor tyrosine-based switch motif of programmed death 1 upon primary human T cell stimulation, but only receptor ligation prevents T cell activation. J Immunol [Internet]. 2004 Jul 15 [cited 2023 Aug 3];173(2):945–54. Available from: https://pubmed.ncbi.nlm.nih.gov/15240681/.
- Okazaki T, Maeda A, Nishimura H, Kurosaki T, Honjo T. PD-1 immunoreceptor inhibits B cell receptor-mediated signaling by recruiting src homology 2-domain-containing tyrosine phosphatase 2 to phosphotyrosine. Proc Natl Acad Sci U S A [Internet]. 2001 Nov 20 [cited 2023 Aug 3];98(24):13866–71. Available from: https://pubmed.ncbi.nlm.nih.gov/11698646/.
- Freeman GJ, Long AJ, Iwai Y, Bourque K, Chernova T, Nishimura H, et al. Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J Exp Med [Internet]. 2000 Oct 2 [cited 2023 Aug 2];192(7):1027–34. Available from: https://pubmed.ncbi.nlm.nih.gov/11015443/.
- Latchman Y, Wood CR, Chernova T, Chaudhary D, Borde M, Chernova I, et al. PD-L2 is a second ligand for PD-1 and inhibits T cell activation. Nat Immunol [Internet]. 2001 Mar [cited 2023 Aug 2];2(3):261–8. Available from: https://pubmed.ncbi.nlm.nih.gov/11224527/.
- Dermani FK, Samadi P, Rahmani G, Kohlan AK, Najafi R. PD-1/PD-L1 immune checkpoint: Potential target for cancer therapy. J Cell Physiol [Internet]. 2019 Feb 1 [cited 2023 Aug 11];234(2):1313–25. Available from: https://pubmed.ncbi.nlm.nih.gov/30191996/.
- Ahn E, Araki K, Hashimoto M, Li W, Riley JL, Cheung J, et al. Role of PD-1 during effector CD8 T cell differentiation. Proc Natl Acad Sci U S A [Internet]. 2018 May 1 [cited 2023 Aug 11];115(18):4749–54. Available from: https://pubmed.ncbi.nlm.nih.gov/29654146/.
- Fritz JM, Lenardo MJ. Development of immune checkpoint therapy for cancer. J Exp Med [Internet]. 2019 Jun 1 [cited 2023 Aug 11];216(6):1244–54. Available from: https://pubmed.ncbi.nlm.nih.gov/31068379/.
- Catakovic K, Klieser E, Neureiter D, Geisberger R. T cell exhaustion: from pathophysiological basics to tumor immunotherapy. Cell Communication and Signaling 2017 15:1 [Internet]. 2017 Jan 5 [cited 2023 Sep 21];15(1):1–16. Available from: https://biosignaling.biomedcentral.com/articles/10.1186/s12964-016-0160-z.
- Wherry EJ, Kurachi M. Molecular and cellular insights into T cell exhaustion. Nature Reviews Immunology 2015 15:8 [Internet]. 2015 Jul 24 [cited 2023 Sep 21];15(8):486–99. Available from: https://www.nature.com/articles/nri3862.
- Pauken KE, Wherry EJ. Overcoming T cell exhaustion in infection and cancer. Trends Immunol [Internet]. 2015 Apr 1 [cited 2023 Sep 21];36(4):265–76. Available from: http://www.cell.com/article/S1471490615000393/fulltext.
- Wakabayashi G, Lee YC, Luh F, Kuo CN, Chang WC, Yen Y. Development and clinical applications of cancer immunotherapy against PD-1 signaling pathway. J Biomed Sci [Internet]. 2019 Dec 5 [cited 2023 Sep 21];26(1):1–13. Available from: https://jbiomedsci.biomedcentral.com/articles/10.1186/s12929-019-0588-8.
- Fife BT, Pauken KE. The role of the PD-1 pathway in autoimmunity and peripheral tolerance. Ann N Y Acad Sci [Internet]. 2011 Jan 1 [cited 2023 Sep 22];1217(1):45–59. Available from: https://onlinelibrary.wiley.com/doi/full/10.1111/j.1749-6632.2010.05919.x.
- Nishimura H, Honjo T, Minato N. Facilitation of β Selection and Modification of Positive Selection in the Thymus of Pd-1–Deficient Mice. Journal of Experimental Medicine [Internet]. 2000 Mar 6 [cited 2023 Sep 22];191(5):891–8. Available from: http://www.jem.org/cgi/current/full/191/5/891.
- Keir ME, Latchman YE, Freeman GJ, Sharpe AH. Programmed Death-1 (PD-1):PD-Ligand 1 Interactions Inhibit TCR-Mediated Positive Selection of Thymocytes. J Immunol [Internet]. 2005 Dec 12 [cited 2023 Sep 22];175(11):7372. Available from: /pmc/articles/PMC2779139/.
- Blank C, Brown I, Marks R, Nishimura H, Honjo T, Gajewski TF. Absence of Programmed Death Receptor 1 Alters Thymic Development and Enhances Generation of CD4/CD8 Double-Negative TCR-Transgenic T Cells. The Journal of Immunology [Internet]. 2003 Nov 1 [cited 2023 Sep 22];171(9):4574–81. Available from:. [CrossRef]
- Policheni AN, Teh CE, Robbins A, Tuzlak S, Strasser A, Gray DHD. PD-1 cooperates with AIRE-mediated tolerance to prevent lethal autoimmune disease. Proc Natl Acad Sci U S A [Internet]. 2022 Apr 12 [cited 2023 Sep 22];119(15):e2120149119. Available from: https://www.pnas.org/doi/abs/10.1073/pnas.2120149119.
- Probst HC, McCoy K, Okazaki T, Honjo T, Van Den Broek M. Resting dendritic cells induce peripheral CD8+ T cell tolerance in vivo through PD-1 and CTLA-4. Probst, H C; McCoy, K; Okazaki, T; Honjo, T; van den Broek, Maries (2005) Resting dendritic cells induce peripheral CD8+ T cell tolerance in vivo through PD-1 and CTLA-4 Nature Immunology, 6:280-286 [Internet]. 2005 [cited 2023 Oct 18];6(3):280–6. Available from: https://www.zora.uzh.ch/id/eprint/136849/.
- Wei X, Niu X. T follicular helper cells in autoimmune diseases. Vol. 134, Journal of Autoimmunity. Academic Press; 2023.
- Mesas-Fernández A, Bodner E, Hilke FJ, Meier K, Ghoreschi K, Solimani F. Interleukin-21 in autoimmune and inflammatory skin diseases. Vol. 53, European Journal of Immunology. John Wiley and Sons Inc; 2023.
- Vogelzang A, McGuire HM, Yu D, Sprent J, Mackay CR, King C. A Fundamental Role for Interleukin-21 in the Generation of T Follicular Helper Cells. Immunity [Internet]. 2008 Jul 18 [cited 2024 May 13];29(1):127–37. Available from: http://www.cell.com/article/S1074761308002744/fulltext.
- Nurieva RI, Chung Y, Hwang D, Yang XO, Kang HS, Ma L, et al. Generation of T Follicular Helper Cells Is Mediated by Interleukin-21 but Independent of T Helper 1, 2, or 17 Cell Lineages. Immunity [Internet]. 2008 Jul 18 [cited 2024 May 13];29(1):138–49. Available from: http://www.cell.com/article/S1074761308002732/fulltext.
- Zhou H, Hu B, Huang N, Mo X, Li W, Zhang B, et al. Aberrant T cell subsets and cytokines expression profile in systemic lupus erythematosus. Clin Rheumatol [Internet]. 2018 Sep 1 [cited 2023 Dec 2];37(9):2405–13. Available from: https://link.springer.com/article/10.1007/s10067-018-4124-0.
- Han L, Yang X, Yu Y, Wan W, Lv L, Zou H. Associations of circulating CXCR3–PD-1+CD4+T cells with disease activity of systemic lupus erythematosus. Mod Rheumatol [Internet]. 2019 May 4 [cited 2023 Dec 2];29(3):461–9. Available from:. [CrossRef]
- Zhang X, Lindwall E, Gauthier C, Lyman J, Spencer N, Alarakhia A, et al. Circulating CXCR5+CD4+helper T cells in systemic lupus erythematosus patients share phenotypic properties with germinal center follicular helper T cells and promote antibody production. [Internet]. 2015 Feb 5 [cited 2023 Dec 2];24(9):909–17. Available from: https://journals.sagepub.com/doi/10.1177/0961203314567750?url_ver=Z39.88-2003&rfr_id=ori%3Arid%3Acrossref.org&rfr_dat=cr_pub++0pubmed.
- Choi JY, Ho JHE, Pasoto SG, Bunin V, Kim ST, Carrasco S, et al. Circulating follicular helper-like T cells in systemic lupus erythematosus: Association with disease activity. Arthritis and Rheumatology. 2015 Apr 1;67(4):988–99.
- Wang L, Zhao P, Ma L, Shan Y, Jiang Z, Wang J, et al. Increased Interleukin 21 and Follicular Helper T-like Cells and Reduced Interleukin 10+ B cells in Patients with New-onset Systemic Lupus Erythematosus. J Rheumatol [Internet]. 2014 Sep 1 [cited 2023 Dec 2];41(9):1781–92. Available from: https://www.jrheum.org/content/41/9/1781.
- Cao G, Chi S, Wang X, Sun J, Zhang Y. CD4+CXCR5+PD-1+ T Follicular Helper Cells Play a Pivotal Role in the Development of Rheumatoid Arthritis. Medical Science Monitor. 2019 Apr 25;25:3032–40.
- Liu R, Wu Q, Su D, Che N, Chen H, Geng L, et al. A regulatory effect of IL-21 on T follicular helper-like cell and B cell in rheumatoid arthritis. Arthritis Res Ther [Internet]. 2012 Nov 23 [cited 2023 Dec 3];14(6):1–12. Available from: https://arthritis-research.biomedcentral.com/articles/10.1186/ar4100.
- Wang X, Yang C, Xu F, Qi L, Wang J, Yang P. Imbalance of circulating Tfr/Tfh ratio in patients with rheumatoid arthritis. Clin Exp Med [Internet]. 2019 Feb 1 [cited 2023 Dec 3];19(1):55–64. Available from: https://link.springer.com/article/10.1007/s10238-018-0530-5.
- Fonseca VR, Romão VC, Agua-Doce A, Santos M, López-Presa D, Ferreira AC, et al. The Ratio of Blood T Follicular Regulatory Cells to T Follicular Helper Cells Marks Ectopic Lymphoid Structure Formation While Activated Follicular Helper T Cells Indicate Disease Activity in Primary Sjögren’s Syndrome. Arthritis and Rheumatology. 2018 May 1;70(5):774–84.
- Kim JW, Lee J, Hong SM, Lee J, Cho M La, Park SH. Circulating CCR7loPD-1hi Follicular Helper T Cells Indicate Disease Activity and Glandular Inflammation in Patients with Primary Sjögren’s Syndrome. Immune Netw [Internet]. 2019 Aug 27 [cited 2023 Dec 4];19(4). Available from: . [CrossRef]
- Pontarini E, Murray-Brown WJ, Croia C, Lucchesi D, Conway J, Rivellese F, et al. Unique expansion of IL-21+ Tfh and Tph cells under control of ICOS identifies Sjögren’s syndrome with ectopic germinal centres and MALT lymphoma. Ann Rheum Dis [Internet]. 2020 Dec 1 [cited 2023 Dec 4];79(12):1588–99. Available from: https://ard.bmj.com/content/79/12/1588.
- Szabó K, Papp G, Szántó A, Tarr T, Zeher M. A comprehensive investigation on the distribution of circulating follicular T helper cells and B cell subsets in primary Sjögren’s syndrome and systemic lupus erythematosus. Clin Exp Immunol [Internet]. 2015 Dec 22 [cited 2023 Dec 4];183(1):76–89. Available from:. [CrossRef]
- Szabó K, Jámbor I, Szántó A, Horváth IF, Tarr T, Nakken B, et al. The Imbalance of Circulating Follicular T Helper Cell Subsets in Primary Sjögren’s Syndrome Associates With Serological Alterations and Abnormal B-Cell Distribution. Front Immunol. 2021 Mar 19;12:639975.
- Haque R, Kim Y, Park K, Jang H, Kim SY, Lee H, et al. Altered distributions in circulating follicular helper and follicular regulatory T cells accountable for imbalanced cytokine production in multiple sclerosis. Clin Exp Immunol [Internet]. 2021 Jun 17 [cited 2023 Dec 4];205(1):75–88. Available from:. [CrossRef]
- Guo J, Zhao C, Wu F, Tao L, Zhang C, Zhao D, et al. T follicular helper-like cells are involved in the pathogenesis of experimental autoimmune encephalomyelitis. Front Immunol. 2018 May 7;9(MAY):348539.
- Ribeiro F, Romão VC, Rosa S, Jesus K, Água-Doce A, Barreira SC, et al. Different antibody-associated autoimmune diseases have distinct patterns of T follicular cell dysregulation. Scientific Reports 2022 12:1 [Internet]. 2022 Oct 21 [cited 2023 Dec 2];12(1):1–8. Available from: https://www.nature.com/articles/s41598-022-21576-8.
- Lin J, Yu Y, Ma J, Ren C, Chen W. PD-1+CXCR5−CD4+T cells are correlated with the severity of systemic lupus erythematosus. Rheumatology [Internet]. 2019 Dec 1 [cited 2023 Dec 2];58(12):2188–92. Available from:. [CrossRef]
- Makiyama A, Chiba A, Noto D, Murayama G, Yamaji K, Tamura N, et al. Expanded circulating peripheral helper T cells in systemic lupus erythematosus: association with disease activity and B cell differentiation. Rheumatology [Internet]. 2019 Oct 1 [cited 2023 Dec 2];58(10):1861–9. Available from:. [CrossRef]
- Sagrero-Fabela N, Ortíz-Lazareno PC, Salazar-Camarena DC, Cruz A, Cerpa-Cruz S, Muñoz-Valle JF, et al. BAFFR expression in circulating T follicular helper (CD4+CXCR5+PD-1+) and T peripheral helper (CD4+CXCR5−PD-1+) cells in systemic lupus erythematosus. [Internet]. 2023 Jul 17 [cited 2023 Sep 25]; Available from: https://journals.sagepub.com/doi/10.1177/09612033231189804?url_ver=Z39.88-2003&rfr_id=ori%3Arid%3Acrossref.org&rfr_dat=cr_pub++0pubmed.
- Luo Q, Fu P, Guo Y, Fu B, Guo Y, Huang Q, et al. Increased TIGIT+PD-1+CXCR5-CD4+T cells are associated with disease activity in rheumatoid arthritis. Exp Ther Med [Internet]. 2022 Aug 31 [cited 2023 Dec 3];24(4). Available from: /pmc/articles/PMC9468811/.
- Zhao L, Li Z, Zeng X, Xia C, Xu L, Xu Q, et al. Circulating CD4+ FoxP3– CXCR5– CXCR3+ PD-1hi cells are elevated in active rheumatoid arthritis and reflect the severity of the disease. Int J Rheum Dis [Internet]. 2021 Aug 1 [cited 2023 Dec 3];24(8):1032–9. Available from: https://onlinelibrary.wiley.com/doi/full/10.1111/1756-185X.14170.
- Rao DA, Gurish MF, Marshall JL, Slowikowski K, Fonseka CY, Liu Y, et al. Pathologically expanded peripheral T helper cell subset drives B cells in rheumatoid arthritis. Nature 2017 542:7639 [Internet]. 2017 Feb 2 [cited 2023 Dec 3];542(7639):110–4. Available from: https://www.nature.com/articles/nature20810.
- Sakuragi T, Yamada H, Haraguchi A, Kai K, Fukushi J ichi, Ikemura S, et al. Autoreactivity of Peripheral Helper T Cells in the Joints of Rheumatoid Arthritis. The Journal of Immunology [Internet]. 2021 May 1 [cited 2023 Dec 3];206(9):2045–51. Available from:. [CrossRef]
- Chen W, Yang F, Lin J. Tph Cells Expanded in Primary Sjögren’s Syndrome. Front Med (Lausanne). 2022 Jun 9;9:900349.
- Dupré A, Pascaud J, Rivière E, Paoletti A, Ly B, Mingueneau M, et al. Association between T follicular helper cells and T peripheral helper cells with B-cell biomarkers and disease activity in primary Sjögren syndrome. RMD Open [Internet]. 2021 Mar 1 [cited 2023 Dec 4];7(1):e001442. Available from: https://rmdopen.bmj.com/content/7/1/e001442.
- Rosenblum MD, Way SS, Abbas AK. Regulatory T cell memory. Nature Reviews Immunology 2015 16:2 [Internet]. 2015 Dec 21 [cited 2023 Oct 18];16(2):90–101. Available from: https://www.nature.com/articles/nri.2015.1.
- Dominguez-Villar M, Hafler DA. Regulatory T cells in autoimmune disease. Vol. 19, Nature Immunology. Nature Publishing Group; 2018. p. 665–73.
- Francisco LM, Salinas VH, Brown KE, Vanguri VK, Freeman GJ, Kuchroo VK, et al. PD-L1 regulates the development, maintenance, and function of induced regulatory T cells. Journal of Experimental Medicine [Internet]. 2009 Dec 21 [cited 2023 Oct 18];206(13):3015–29. Available from: www.jem.org/cgi/doi/10.1084/jem.20090847.
- Amarnath S, Mangus CW, Wang JCM, Wei F, He A, Kapoor V, et al. The PDL1-PD1 axis converts human T H1 cells into regulatory T cells. Sci Transl Med [Internet]. 2011 Nov 30 [cited 2023 Oct 18];3(111). Available from: https://www.science.org/doi/10.1126/scitranslmed.3003130.
- Zhao L, Zhou X, Zhou X, Wang H, Gu L, Ke Y, et al. Low expressions of PD-L1 and CTLA-4 by induced CD4+CD25+ Foxp3+ Tregs in patients with SLE and their correlation with the disease activity. Cytokine. 2020 Sep 1;133:155119.
- Ferreira RC, Castro Dopico X, Oliveira JJ, Rainbow DB, Yang JH, Trzupek D, et al. Chronic Immune Activation in Systemic Lupus Erythematosus and the Autoimmune PTPN22 Trp620 Risk Allele Drive the Expansion of FOXP3+ Regulatory T Cells and PD-1 Expression. Front Immunol. 2019 Nov 8;10.
- Oestreich KJ, Yoon H, Ahmed R, Boss JM. NFATc1 regulates PD-1 expression upon T cell activation. J Immunol [Internet]. 2008 Oct 1 [cited 2023 Aug 2];181(7):4832–9. Available from: https://pubmed.ncbi.nlm.nih.gov/18802087/.
- Austin JW, Lu P, Majumder P, Ahmed R, Boss JM. STAT3, STAT4, NFATc1, and CTCF regulate PD-1 through multiple novel regulatory regions in murine T cells. J Immunol [Internet]. 2014 May 15 [cited 2023 Aug 3];192(10):4876–86. Available from: https://pubmed.ncbi.nlm.nih.gov/24711622/.
- Wang G, Tajima M, Honjo T, Ohta A. STAT5 interferes with PD-1 transcriptional activation and affects CD8+ T-cell sensitivity to PD-1-dependent immunoregulation. Int Immunol [Internet]. 2021 Nov 1 [cited 2023 Aug 3];33(11):563–72. Available from: https://pubmed.ncbi.nlm.nih.gov/34453440/.
- Terawaki S, Chikuma S, Shibayama S, Hayashi T, Yoshida T, Okazaki T, et al. IFN-α directly promotes programmed cell death-1 transcription and limits the duration of T cell-mediated immunity. J Immunol [Internet]. 2011 Mar 1 [cited 2023 Aug 3];186(5):2772–9. Available from: https://pubmed.ncbi.nlm.nih.gov/21263073/.
- Xiao G, Deng A, Liu H, Ge G, Liu X. Activator protein 1 suppresses antitumor T-cell function via the induction of programmed death 1. Proc Natl Acad Sci U S A [Internet]. 2012 Sep 18 [cited 2023 Aug 3];109(38):15419–24. Available from: https://pubmed.ncbi.nlm.nih.gov/22949674/.
- Staron MM, Gray SM, Marshall HD, Parish IA, Chen JH, Perry CJ, et al. The transcription factor FoxO1 sustains expression of the inhibitory receptor PD-1 and survival of antiviral CD8(+) T cells during chronic infection. Immunity [Internet]. 2014 Nov 20 [cited 2023 Aug 3];41(5):802–14. Available from: https://pubmed.ncbi.nlm.nih.gov/25464856/.
- Chi Z, Lu Y, Yang Y, Li B, Lu P. Transcriptional and epigenetic regulation of PD-1 expression. Cell Mol Life Sci [Internet]. 2021 Apr 1 [cited 2023 Aug 2];78(7):3239–46. Available from: https://pubmed.ncbi.nlm.nih.gov/33738533/.
- Tsokos GC, Lo MS, Reis PC, Sullivan KE. New insights into the immunopathogenesis of systemic lupus erythematosus. Vol. 12, Nature Reviews Rheumatology. Nature Publishing Group; 2016. p. 716–30.
- Okada M, Chikuma S, Kondo T, Hibino S, Machiyama H, Yokosuka T, et al. Blockage of Core Fucosylation Reduces Cell-Surface Expression of PD-1 and Promotes Anti-tumor Immune Responses of T Cells. Cell Rep [Internet]. 2017 Aug 1 [cited 2023 Dec 11];20(5):1017–28. Available from: http://www.cell.com/article/S2211124717309932/fulltext.
- Pentcheva-Hoang T, Chen L, Pardoll DM, Allison JP. Programmed death-1 concentration at the immunological synapse is determined by ligand affinity and availability. Proc Natl Acad Sci U S A [Internet]. 2007 Nov 6 [cited 2023 Dec 11];104(45):17765–70. Available from: https://www.pnas.org/doi/abs/10.1073/pnas.0708767104.
- Mathieu M, Cotta-Grand N, Daudelin JF, Thébault P, Labrecque N. Notch signaling regulates PD-1 expression during CD8+ T-cell activation. Immunol Cell Biol [Internet]. 2013 Jan 1 [cited 2023 Sep 21];91(1):82–8. Available from: https://onlinelibrary.wiley.com/doi/full/10.1038/icb.2012.53.
- Lu P, Youngblood BA, Austin JJ, Mohammed AUR, Butler R, Ahmed R, et al. Blimp-1 represses CD8 T cell expression of PD-1 using a feed-forward transcriptional circuit during acute viral infection. J Exp Med [Internet]. 2014 Mar [cited 2023 Aug 3];211(3):515–27. Available from: https://pubmed.ncbi.nlm.nih.gov/24590765/.
- Kao C, Oestreich KJ, Paley MA, Crawford A, Angelosanto JM, Ali MAA, et al. Transcription factor T-bet represses expression of the inhibitory receptor PD-1 and sustains virus-specific CD8+ T cell responses during chronic infection. Nat Immunol [Internet]. 2011 May [cited 2023 Aug 3];12(7):663–71. Available from: https://pubmed.ncbi.nlm.nih.gov/21623380/.
- Shin H, Blackburn SD, Intlekofer AM, Kao C, Angelosanto JM, Reiner SL, et al. A role for the transcriptional repressor Blimp-1 in CD8(+) T cell exhaustion during chronic viral infection. Immunity [Internet]. 2009 Aug 21 [cited 2023 Aug 3];31(2):309–20. Available from: https://pubmed.ncbi.nlm.nih.gov/19664943/.
- Shankar EM, Che KF, Messmer D, Lifson JD, Larsson M. Expression of a broad array of negative costimulatory molecules and Blimp-1 in T cells following priming by HIV-1 pulsed dendritic cells. Molecular Medicine [Internet]. 2011 Mar 17 [cited 2023 Sep 30];17(3–4):229–40. Available from: https://molmed.biomedcentral.com/articles/10.2119/molmed.2010.00175.
- Che KF, Shankar EM, Muthu S, Zandi S, Sigvardsson M, Hinkula J, et al. p38 mitogen-activated protein kinase/signal transducer and activator of transcription-3 pathway signaling regulates expression of inhibitory molecules in T Cells activated by HIV-1-exposed dendritic cells. Molecular Medicine [Internet]. 2012 Aug 3 [cited 2023 Sep 30];18(8):1169–82. Available from: https://molmed.biomedcentral.com/articles/10.2119/molmed.2012.00103.
- Philips EA, Garcia-España A, Tocheva AS, Ahearn IM, Adam KR, Pan R, et al. The structural features that distinguish PD-L2 from PD-L1 emerged in placental mammals. Journal of Biological Chemistry [Internet]. 2020 Apr 3 [cited 2023 Dec 11];295(14):4372–80. Available from: http://www.jbc.org/article/S0021925817487047/fulltext.
- Bardhan K, Anagnostou T, Boussiotis VA. The PD1:PD-L1/2 Pathway from Discovery to Clinical Implementation. Front Immunol [Internet]. 2016 [cited 2023 Aug 11];7(DEC). Available from: https://pubmed.ncbi.nlm.nih.gov/28018338/.
- Ishida M, Iwai Y, Tanaka Y, Okazaki T, Freeman GJ, Minato N, et al. Differential expression of PD-L1 and PD-L2, ligands for an inhibitory receptor PD-1, in the cells of lymphohematopoietic tissues. Immunol Lett [Internet]. 2002 Oct 21 [cited 2023 Aug 2];84(1):57–62. Available from: https://pubmed.ncbi.nlm.nih.gov/12161284/.
- Ibañez-Vega J, Vilchez C, Jimenez K, Guevara C, Burgos PI, Naves R. Cellular and molecular regulation of the programmed death-1/programmed death ligand system and its role in multiple sclerosis and other autoimmune diseases. J Autoimmun [Internet]. 2021 Sep 1 [cited 2023 Aug 11];123. Available from: https://pubmed.ncbi.nlm.nih.gov/34311143/.
- Sun C, Mezzadra R, Schumacher TN. Regulation and Function of the PD-L1 Checkpoint. Immunity [Internet]. 2018 Mar 20 [cited 2023 Aug 11];48(3):434–52. Available from: https://pubmed.ncbi.nlm.nih.gov/29562194/.
- Yi M, Niu M, Xu L, Luo S, Wu K. Regulation of PD-L1 expression in the tumor microenvironment. J Hematol Oncol [Internet]. 2021 Dec 1 [cited 2023 Aug 11];14(1). Available from: https://pubmed.ncbi.nlm.nih.gov/33413496/.
- Nie X, Chen W, Zhu Y, Huang B, Yu W, Wu Z, et al. B7-DC (PD-L2) costimulation of CD4+ T-helper 1 response via RGMb. Cell Mol Immunol [Internet]. 2018 Oct 1 [cited 2023 Aug 11];15(10):888–97. Available from: https://pubmed.ncbi.nlm.nih.gov/28479601/.
- Muraro E, Romanò R, Fanetti G, Vaccher E, Turturici I, Lupato V, et al. Tissue and circulating PD-L2: moving from health and immune-mediated diseases to head and neck oncology. Crit Rev Oncol Hematol [Internet]. 2022 Jul 1 [cited 2023 Aug 11];175. Available from: https://pubmed.ncbi.nlm.nih.gov/35569724/.
- Mayoux M, Roller A, Pulko V, Sammicheli S, Chen S, Sum E, et al. Dendritic cells dictate responses to PD-L1 blockade cancer immunotherapy. Sci Transl Med [Internet]. 2020 Mar 11 [cited 2023 Aug 11];12(534). Available from: https://pubmed.ncbi.nlm.nih.gov/32161104/.
- Anton Van Der Merwe P, Bodian DL, Daenke S, Linsley P, Davis SJ. CD80 (B7-1) Binds Both CD28 and CTLA-4 with a Low Affinity and Very Fast Kinetics. J Exp Med [Internet]. 1997 [cited 2023 Sep 8];185(3):393–403. Available from: http://rupress.org/jem/article-pdf/185/3/393/1680533/5372.pdf.
- Butte MJ, Peña-Cruz V, Kim MJ, Freeman GJ, Sharpe AH. Interaction of human PD-L1 and B7-1. Mol Immunol. 2008 Aug 1;45(13):3567–72.
- Sugiura D, Maruhashi T, Okazaki IM, Shimizu K, Maeda TK, Takemoto T, et al. Restriction of PD-1 function by cis-PD-L1/CD80 interactions is required for optimal T cell responses. Science (1979) [Internet]. 2019 [cited 2023 Aug 8];364(6440):558–66. Available from: https://www.science.org.
- Zhao Y, Lee CK, Lin CH, Gassen RB, Xu X, Huang Z, et al. Article PD-L1:CD80 Cis-Heterodimer Triggers the Co-stimulatory Receptor CD28 While Repressing the Inhibitory PD-1 and CTLA-4 Pathways. [cited 2023 Aug 8]; Available from: . [CrossRef]
- Linnerbauer M, Beyer T, Nirschl L, Farrenkopf D, Lößlein L, Vandrey O, et al. PD-L1 positive astrocytes attenuate inflammatory functions of PD-1 positive microglia in models of autoimmune neuroinflammation. Nature Communications 2023 14:1 [Internet]. 2023 Sep 9 [cited 2023 Sep 22];14(1):1–17. Available from: https://www.nature.com/articles/s41467-023-40982-8.
- Raptopoulou AP, Bertsias G, Makrygiannakis D, Verginis P, Kritikos I, Tzardi M, et al. The programmed death 1/programmed death ligand 1 inhibitory pathway is up-regulated in rheumatoid synovium and regulates peripheral T cell responses in human and murine arthritis. Arthritis Rheum. 2010 Jul;62(7):1870–80.
- Sugiura D, Okazaki I mi, Maeda TK, Maruhashi T, Shimizu K, Arakaki R, et al. PD-1 agonism by anti-CD80 inhibits T cell activation and alleviates autoimmunity. Nat Immunol. 2022 Mar 1;23(3):399–410.
- Nielsen C, Ohm-Laursen L, Barington T, Husby S, Lillevang ST. Alternative splice variants of the human PD-1 gene. Cell Immunol. 2005 Jun 1;235(2):109–16.
- Gu D, Ao X, Yang Y, Chen Z, Xu X. Soluble immune checkpoints in cancer: production, function and biological significance. Journal for ImmunoTherapy of Cancer 2018 6:1 [Internet]. 2018 Nov 27 [cited 2023 Sep 8];6(1):1–14. Available from: https://jitc.biomedcentral.com/articles/10.1186/s40425-018-0449-0.
- Király Z, Nagy E, Bokor L, Kovács A, Marschalkó M, Hidvégi B. The Possible Clinical Significance of a Decreased Serum Level of Soluble PD-L1 in Discoid Lupus Erythematosus, but Not in Subacute Cutaneous Lupus Erythematosus—A Pilot Study. Journal of Clinical Medicine 2023, Vol 12, Page 5648 [Internet]. 2023 Aug 30 [cited 2023 Oct 3];12(17):5648. Available from: https://www.mdpi.com/2077-0383/12/17/5648/htm.
- Hirahara S, Katsumata Y, Kawasumi H, Kawaguchi Y, Harigai M. Serum levels of soluble programmed cell death protein 1 and soluble programmed cell death protein ligand 2 are increased in systemic lupus erythematosus and associated with the disease activity. [Internet]. 2020 Apr 7 [cited 2023 Oct 3];29(7):686–96. Available from: https://journals.sagepub.com/doi/10.1177/0961203320916517?url_ver=Z39.88-2003&rfr_id=ori%3Arid%3Acrossref.org&rfr_dat=cr_pub++0pubmed.
- Du Y, Nie L, Xu L, Wu X, Zhang S, Xue J. Serum levels of soluble programmed death-1 (sPD-1) and soluble programmed death ligand 1(sPD-L1) in systemic lupus erythematosus: Association with activity and severity. Scand J Immunol [Internet]. 2020 Jul 1 [cited 2023 Oct 3];92(1). Available from: https://pubmed.ncbi.nlm.nih.gov/32243638/.
- Xu L, Jiang L, Nie L, Zhang S, Liu L, Du Y, et al. Soluble programmed death molecule 1 (sPD-1) as a predictor of interstitial lung disease in rheumatoid arthritis. BMC Immunol [Internet]. 2021 Dec 1 [cited 2023 Oct 3];22(1):1–10. Available from: https://bmcimmunol.biomedcentral.com/articles/10.1186/s12865-021-00460-6.
- Greisen SR, Rasmussen TK, Stengaard-Pedersen K, Hetland ML, Horslev-Petersen K, Hvid M, et al. Increased soluble programmed death-1 (sPD-1) is associated with disease activity and radiographic progression in early rheumatoid arthritis. Scand J Rheumatol [Internet]. 2014 [cited 2023 Oct 4];43(2):101–8. Available from: https://www.tandfonline.com/doi/abs/10.3109/03009742.2013.823517.
- Liu C, Jiang J, Gao L, Wang X, Hu X, Wu M, et al. Soluble PD-1 aggravates progression of collagen-induced arthritis through Th1 and Th17 pathways. Arthritis Res Ther [Internet]. 2015 Nov 25 [cited 2023 Oct 4];17(1):1–13. Available from: https://arthritis-research.biomedcentral.com/articles/10.1186/s13075-015-0859-z.
- Chen Y, Wang Y, Xu L, Zhu W, Xu C, Xu M, et al. Influence of total glucosides of paeony on PD-1/PD-L1 expression in primary Sjögren’s syndrome. Int J Rheum Dis [Internet]. 2019 Feb 1 [cited 2023 Oct 4];22(2):200–6. Available from: https://onlinelibrary.wiley.com/doi/full/10.1111/1756-185X.13391.
- Qian S, Xu J, Zhao S, Yang P, Yang C. CMTM6: increased circulating level and up-regulated expression in labial salivary glands in patients with primary Sjogren’s syndrome. Clin Exp Immunol [Internet]. 2022 Jan 28 [cited 2023 Oct 4];207(1):65–71. Available from:. [CrossRef]
- Yoon T, Ahn SS, Jung SM, Song JJ, Park YB, Lee SW. Serum soluble programmed cell death protein 1 could predict the current activity and severity of antineutrophil cytoplasmic antibody-associated vasculitis: a monocentric prospective study. Clin Exp Rheumatol [Internet]. 2019 Mar 1 [cited 2023 Oct 4];37(2):116–21. Available from: https://www.clinexprheumatol.org/abstract.asp?a=13748.
- Bailly C, Thuru X, Quesnel B. Soluble programmed death ligand-1 (Spd-l1): A pool of circulating proteins implicated in health and diseases. Vol. 13, Cancers. MDPI; 2021.
- He XH, Xu LH, Liu Y. Identification of a novel splice variant of human PD-L1 mRNA encoding an isoform-lacking Igv-like domain. Acta Pharmacologica Sinica 2005 26:4 [Internet]. 2005 Apr [cited 2023 Oct 3];26(4):462–8. Available from: https://www.nature.com/articles/aps200570.
- He XH, Liu Y, Xu LH, Zeng YY. Cloning and Identification of Two Novel Splice Variants of Human PD-L2. Acta Biochim Biophys Sin (Shanghai) [Internet]. 2004 Apr 1 [cited 2023 Oct 3];36(4):284–9. Available from:. [CrossRef]
- Niu M, Liu Y, Yi M, Jiao D, Wu K. Biological Characteristics and Clinical Significance of Soluble PD-1/PD-L1 and Exosomal PD-L1 in Cancer. Front Immunol. 2022 Mar 21;13:827921.
- Bertsias GK, Nakou M, Choulaki C, Raptopoulou A, Papadimitraki E, Goulielmos G, et al. Genetic, immunologic, and immunohistochemical analysis of the programmed death 1/programmed death ligand 1 pathway in human systemic lupus erythematosus. Arthritis Rheum. 2009 Jan;60(1):207–18.
- Liu MF, Weng CT, Weng MY. Variable increased expression of program death-1 and program death-1 ligands on peripheral mononuclear cells is not impaired in patients with systemic lupus erythematosus. J Biomed Biotechnol. 2009;2009.
- Luo Q, Huang Z, Ye J, Deng Y, Fang L, Li X, et al. PD-L1-expressing neutrophils as a novel indicator to assess disease activity and severity of systemic lupus erythematosus. Arthritis Res Ther [Internet]. 2016 Feb 11 [cited 2023 Dec 2];18(1):1–11. Available from: https://arthritis-research.biomedcentral.com/articles/10.1186/s13075-016-0942-0.
- Jia XY, Zhu Q qing, Wang YY, Lu Y, Li ZJ, Li BQ, et al. The role and clinical significance of programmed cell death- ligand 1 expressed on CD19+B-cells and subsets in systemic lupus erythematosus. Clinical Immunology. 2019 Jan 1;198:89–99.
- Zhu Q, Li Y, Zhang L, Wang M, Chen Z, Shi J, et al. Patients with systemic lupus erythematosus show increased proportions of CD19+CD20− B cells and secretion of related autoantibodies. Clin Rheumatol [Internet]. 2021 Jan 1 [cited 2023 Dec 2];40(1):151–65. Available from: https://link.springer.com/article/10.1007/s10067-020-05220-2.
- Rincon-Arevalo H, Wiedemann A, Stefanski AL, Lettau M, Szelinski F, Fuchs S, et al. Deep Phenotyping of CD11c+ B Cells in Systemic Autoimmunity and Controls. Front Immunol. 2021 Mar 12;12:635615.
- Shi H, Ye J, Teng J, Yin Y, Hu Q, Wu X, et al. Elevated serum autoantibodies against co-inhibitory PD-1 facilitate T cell proliferation and correlate with disease activity in new-onset systemic lupus erythematosus patients. Arthritis Res Ther [Internet]. 2017 Mar 9 [cited 2023 Dec 2];19(1):1–10. Available from: https://arthritis-research.biomedcentral.com/articles/10.1186/s13075-017-1258-4.
- Tong M, Fang X, Yang J, Wu P, Guo Y, Sun J. Abnormal membrane-bound and soluble programmed death ligand 2 (PD-L2) expression in systemic lupus erythematosus is associated with disease activity. Immunol Lett. 2020 Nov 1;227:96–101.
- Xiong J, Yang J, Sun Y, Chen Y, Guo Y, Liu C, et al. Dysregulated PD-L2 is correlated with disease activity and inflammation in rheumatoid arthritis. Immunogenetics [Internet]. 2023 Oct 1 [cited 2023 Dec 4];75(5):425–31. Available from: https://link.springer.com/article/10.1007/s00251-023-01307-7.
- Guo Y, Walsh AM, Canavan M, Wechalekar MD, Cole S, Yin X, et al. Immune checkpoint inhibitor PD-1 pathway is down-regulated in synovium at various stages of rheumatoid arthritis disease progression. PLoS One [Internet]. 2018 Feb 1 [cited 2023 Dec 4];13(2):e0192704. Available from: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0192704.
- Matsuda K, Miyoshi H, Hiraoka K, Hamada T, Yoshida S, Ishibashi Y, et al. Clinicopathological value of programmed cell death 1 (PD-1) and programmed cell death ligand 1 (PD-L1) expression in synovium of patients with rheumatoid arthritis. Clin Exp Med [Internet]. 2018 Nov 1 [cited 2023 Dec 4];18(4):487–94. Available from: https://link.springer.com/article/10.1007/s10238-018-0515-4.
- Moret FM, van der Wurff-Jacobs KMG, Bijlsma JWJ, Lafeber FPJG, van Roon JAG. Synovial T cell hyporesponsiveness to myeloid dendritic cells is reversed by preventing PD-1/PD-L1 interactions. Arthritis Res Ther [Internet]. 2014 Nov 30 [cited 2023 Dec 4];16(6):1–8. Available from: https://arthritis-research.biomedcentral.com/articles/10.1186/s13075-014-0497-x.
- Wan B, Nie H, Liu A, Feng G, He D, Xu R, et al. Aberrant Regulation of Synovial T Cell Activation by Soluble Costimulatory Molecules in Rheumatoid Arthritis. The Journal of Immunology [Internet]. 2006 Dec 15 [cited 2023 Dec 4];177(12):8844–50. Available from:. [CrossRef]
- Greisen SR, Rasmussen TK, Stengaard-Pedersen K, Hetland ML, Horslev-Petersen K, Hvid M, et al. Increased soluble programmed death-1 (sPD-1) is associated with disease activity and radiographic progression in early rheumatoid arthritis. Scand J Rheumatol [Internet]. 2014 [cited 2023 Dec 4];43(2):101–8. Available from: https://www.tandfonline.com/doi/abs/10.3109/03009742.2013.823517.
- Bommarito D, Hall C, Taams LS, Corrigall VM. Inflammatory cytokines compromise programmed cell death-1 (PD-1)-mediated T cell suppression in inflammatory arthritis through up-regulation of soluble PD-1. Clin Exp Immunol [Internet]. 2017 May 9 [cited 2023 Dec 4];188(3):455–66. Available from:. [CrossRef]
- Kobayashi M, Kawano S, Hatachi S, Kurimoto C, Okazaki T, Iwai Y, et al. Enhanced expression of programmed death-1 (PD-1)/PD-L1 in salivary glands of patients with Sjögren’s syndrome. J Rheumatol. 2005;32(11).
- Loureiro-Amig J, Franco-Jarav C, Perurena-Priet J, Palacio C, Martínez-Vall F, Soláns-Laqu R. Serum CXCL13, BAFF, IL-21 and IL-22 levels are related to disease activity and lymphocyte profile in primary Sjögren’s syndrome. Clin Exp Rheumatol. 2021;39(6):S131–9.
- Nishikawa A, Suzuki K, Kassai Y, Gotou Y, Takiguchi M, Miyazaki T, et al. Identification of definitive serum biomarkers associated with disease activity in primary Sjögren’s syndrome. Arthritis Res Ther [Internet]. 2016 May 14 [cited 2023 Dec 5];18(1):1–10. Available from: https://arthritis-research.biomedcentral.com/articles/10.1186/s13075-016-1006-1.
- Linnerbauer M, Beyer T, Nirschl L, Farrenkopf D, Lößlein L, Vandrey O, et al. PD-L1 positive astrocytes attenuate inflammatory functions of PD-1 positive microglia in models of autoimmune neuroinflammation. Nature Communications 2023 14:1 [Internet]. 2023 Sep 9 [cited 2023 Dec 7];14(1):1–17. Available from: https://www.nature.com/articles/s41467-023-40982-8.
- Trabattoni D, Saresella M, Pacei M, Marventano I, Mendozzi L, Rovaris M, et al. Costimulatory Pathways in Multiple Sclerosis: Distinctive Expression of PD-1 and PD-L1 in Patients with Different Patterns of Disease. The Journal of Immunology [Internet]. 2009 Oct 15 [cited 2023 Dec 7];183(8):4984–93. Available from:. [CrossRef]
- Garcia-Diaz A, Shin DS, Moreno BH, Saco J, Escuin-Ordinas H, Rodriguez GA, et al. Interferon Receptor Signaling Pathways Regulating PD-L1 and PD-L2 Expression. Cell Rep [Internet]. 2017 May 9 [cited 2023 Sep 29];19(6):1189–201. Available from: http://www.cell.com/article/S2211124717305259/fulltext.
- Horita H, Law A, Hong S, Middleton K. Identifying Regulatory Posttranslational Modifications of PD-L1: A Focus on Monoubiquitinaton. Neoplasia (United States). 2017 Apr 1;19(4):346–53.
- Li CW, Lim SO, Xia W, Lee HH, Chan LC, Kuo CW, et al. Glycosylation and stabilization of programmed death ligand-1 suppresses T-cell activity. Nature Communications 2016 7:1 [Internet]. 2016 Aug 30 [cited 2023 Dec 10];7(1):1–11. Available from: https://www.nature.com/articles/ncomms12632.
- Yao H, Lan J, Li C, Shi H, Brosseau JP, Wang H, et al. Inhibiting PD-L1 palmitoylation enhances T-cell immune responses against tumours. Nature Biomedical Engineering 2019 3:4 [Internet]. 2019 Mar 25 [cited 2023 Dec 10];3(4):306–17. Available from: https://www.nature.com/articles/s41551-019-0375-6.
- Linder ME, Deschenes RJ. Palmitoylation: policing protein stability and traffic. Nature Reviews Molecular Cell Biology 2007 8:1 [Internet]. 2007 Jan [cited 2023 Dec 10];8(1):74–84. Available from: https://www.nature.com/articles/nrm2084.
- Yu X, Li W, Young KH, Li Y. Posttranslational Modifications in PD-L1 Turnover and Function: From Cradle to Grave. Biomedicines 2021, Vol 9, Page 1702 [Internet]. 2021 Nov 16 [cited 2023 Dec 10];9(11):1702. Available from: https://www.mdpi.com/2227-9059/9/11/1702/htm.
- Burr ML, Sparbier CE, Chan YC, Williamson JC, Woods K, Beavis PA, et al. CMTM6 maintains the expression of PD-L1 and regulates anti-tumour immunity. Nature 2017 549:7670 [Internet]. 2017 Aug 16 [cited 2023 Sep 29];549(7670):101–5. Available from: https://www.nature.com/articles/nature23643.
- Mezzadra R, Sun C, Jae LT, Gomez-Eerland R, De Vries E, Wu W, et al. Identification of CMTM6 and CMTM4 as PD-L1 protein regulators. Nature 2017 549:7670 [Internet]. 2017 Aug 16 [cited 2023 Sep 29];549(7670):106–10. Available from: https://www.nature.com/articles/nature23669.
- Davis NA, Lareau CA, White BC, Pandey A, Wiley G, Montgomery CG, et al. Encore: Genetic Association Interaction Network Centrality Pipeline and Application to SLE Exome Data. Genet Epidemiol [Internet]. 2013 Sep 1 [cited 2023 Sep 29];37(6):614–21. Available from: https://onlinelibrary.wiley.com/doi/full/10.1002/gepi.21739.
- Zeisbrich M, Chevalier N, Sehnert B, Rizzi M, Venhoff N, Thiel J, et al. CMTM6-Deficient Monocytes in ANCA-Associated Vasculitis Fail to Present the Immune Checkpoint PD-L1. Front Immunol. 2021 May 24;12:673912.
- Hui E, Cheung J, Zhu J, Su X, Taylor MJ, Wallweber HA, et al. T cell costimulatory receptor CD28 is a primary target for PD-1-mediated inhibition. Science [Internet]. 2017 Mar 31 [cited 2023 Aug 11];355(6332):1428–33. Available from: https://pubmed.ncbi.nlm.nih.gov/28280247/.
- Zamani MR, Aslani S, Salmaninejad A, Javan MR, Rezaei N. PD-1/PD-L and autoimmunity: A growing relationship. Cell Immunol [Internet]. 2016 Dec 1 [cited 2023 Aug 11];310:27–41. Available from: https://pubmed.ncbi.nlm.nih.gov/27660198/.
- Tocheva AS, Peled M, Strazza M, Adam KR, Lerrer S, Nayak S, et al. Quantitative phosphoproteomic analysis reveals involvement of PD-1 in multiple T cell functions. Journal of Biological Chemistry. 2020 Dec 25;295(52):18036–50.
- Strazza M, Bukhari S, Tocheva AS, Mor A. PD-1-induced proliferating T cells exhibit a distinct transcriptional signature. Immunology [Internet]. 2021 Nov 1 [cited 2023 Oct 10];164(3):555–68. Available from: https://onlinelibrary.wiley.com/doi/full/10.1111/imm.13388.
- Lerrer S, Tocheva AS, Bukhari S, Adam K, Mor A. PD-1-stimulated T cell subsets are transcriptionally and functionally distinct. iScience. 2021 Sep 24;24(9):103020.
- Haynes NM, Allen CDC, Lesley R, Ansel KM, Killeen N, Cyster JG. Role of CXCR5 and CCR7 in Follicular Th Cell Positioning and Appearance of a Programmed Cell Death Gene-1High Germinal Center-Associated Subpopulation. The Journal of Immunology [Internet]. 2007 Oct 15 [cited 2023 Oct 7];179(8):5099–108. Available from:. [CrossRef]
- Good-Jacobson KL, Szumilas CG, Chen L, Sharpe AH, Tomayko MM, Shlomchik MJ. PD-1 regulates germinal center B cell survival and the formation and affinity of long-lived plasma cells. Nat Immunol. 2010 Jun;11(6):535–42.
- Ding BB, Bi E, Chen H, Yu JJ, Ye BH. IL-21 and CD40L Synergistically Promote Plasma Cell Differentiation through Upregulation of Blimp-1 in Human B Cells. The Journal of Immunology [Internet]. 2013 Feb 15 [cited 2023 Oct 7];190(4):1827–36. Available from:. [CrossRef]
- Kawamoto S, Tran TH, Maruya M, Suzuki K, Doi Y, Tsutsui Y, et al. The inhibitory receptor PD-1 regulates IgA selection and bacterial composition in the gut. Science (1979). 2012 Apr 27;336(6080):485–9.
- Shi J, Hou S, Fang Q, Liu X, Liu X, Correspondence HQ, et al. PD-1 Controls Follicular T Helper Cell Positioning and Function Article PD-1 Controls Follicular T Helper Cell Positioning and Function. Immunity [Internet]. 2018 [cited 2023 Sep 8];49:264–74. Available from: . [CrossRef]
- Choi YS, Kageyama R, Eto D, Escobar TC, Johnston RJ, Monticelli L, et al. ICOS Receptor Instructs T Follicular Helper Cell versus Effector Cell Differentiation via Induction of the Transcriptional Repressor Bcl6. Immunity. 2011 Jun 24;34(6):932–46.
- Kinter AL, Godbout EJ, McNally JP, Sereti I, Roby GA, O’Shea MA, et al. The Common γ-Chain Cytokines IL-2, IL-7, IL-15, and IL-21 Induce the Expression of Programmed Death-1 and Its Ligands. The Journal of Immunology [Internet]. 2008 Nov 15 [cited 2023 Oct 7];181(10):6738–46. Available from:. [CrossRef]
- Haymaker C, Wu R, Bernatchez C, Radvanyi L. PD-1 and BTLA and CD8+ T-cell “exhaustion” in cancer: “Exercising” an alternative viewpoint. Oncoimmunology [Internet]. 2012 Aug 8 [cited 2024 May 14];1(5):735. Available from: /pmc/articles/PMC3429577/.
- Dolff S, Abdulahad WH, Westra J, Doornbos-van der Meer B, Limburg PC, Kallenberg CGM, et al. Increase in IL-21 producing T-cells in patients with systemic lupus erythematosus. Arthritis Res Ther [Internet]. 2011 Sep 29 [cited 2024 Jan 9];13(5):1–10. Available from: https://arthritis-research.biomedcentral.com/articles/10.1186/ar3474.
- Paluch C, Santos AM, Anzilotti C, Cornall RJ, Davis SJ. Immune checkpoints as therapeutic targets in autoimmunity. Vol. 9, Frontiers in Immunology. Frontiers Media S.A.; 2018.
- Liao W, Zheng H, Wu S, Zhang Y, Wang W, Zhang Z, et al. The Systemic Activation of Programmed Death 1-PD-L1 Axis Protects Systemic Lupus Erythematosus Model from Nephritis. Am J Nephrol. 2017 Nov 1;46(5):371–9.
- Bryan CM, Rocklin GJ, Bick MJ, Ford A, Majri-Morrison S, Kroll A V, et al. Computational design of a synthetic PD-1 agonist. Proc Natl Acad Sci U S A [Internet]. 2021; Available from: . [CrossRef]
- Curnock AP, Bossi G, Kumaran J, Bawden LJ, Figueiredo R, Tawar R, et al. Cell-targeted PD-1 agonists that mimic PD-L1 are potent T cell inhibitors. JCI Insight. 2021 Oct 22;6(20).
- Guo Q, Chen C, Wu Z, Zhang W, Wang L, Yu J, et al. Engineered PD-1/TIGIT dual-activating cell-membrane nanoparticles with dexamethasone act synergistically to shape the effector T cell/Treg balance and alleviate systemic lupus erythematosus. Biomaterials. 2022 Jun 1;285.
- Khramova T, Beduleva L, Sidorov A, Terentiev A, Menshikov I. Regulatory Rheumatoid Factor is Specific to PD-1 and Uses PD-1 Pathway to Control CD4 T Lymphocytes. Immunol Invest. 2023;
- Wang B, Chen C, Liu X, Zhou S, Xu T, Wu M. The effect of combining PD-1 agonist and low-dose Interleukin-2 on treating systemic lupus erythematosus. Vol. 14, Frontiers in Immunology. Frontiers Media S.A.; 2023.
Figure 1.
Structure of PD-1 and its ligands PD-L1 and PD-L2. A) The structure of PD-1 is composed of an (IgV)-like domain and (IgC)-like domain in the extracellular region, followed by a transmembrane domain, and the intracellular region is composed of an ITIM and ITSM domains. B) The structure of its ligands PD-1 and PD-L2 shared the extracellular structure with PD-1. PD-1: Programmed death 1; PD-L1: Programmed cell death-ligand 1; PD-L2: Programmed cell death-ligand 2; ITIM: Immunoreceptor Tyrosine-based Inhibitory Motif; ITSM: Immunoreceptor Tyrosine-based Switch Motif.
Figure 1.
Structure of PD-1 and its ligands PD-L1 and PD-L2. A) The structure of PD-1 is composed of an (IgV)-like domain and (IgC)-like domain in the extracellular region, followed by a transmembrane domain, and the intracellular region is composed of an ITIM and ITSM domains. B) The structure of its ligands PD-1 and PD-L2 shared the extracellular structure with PD-1. PD-1: Programmed death 1; PD-L1: Programmed cell death-ligand 1; PD-L2: Programmed cell death-ligand 2; ITIM: Immunoreceptor Tyrosine-based Inhibitory Motif; ITSM: Immunoreceptor Tyrosine-based Switch Motif.
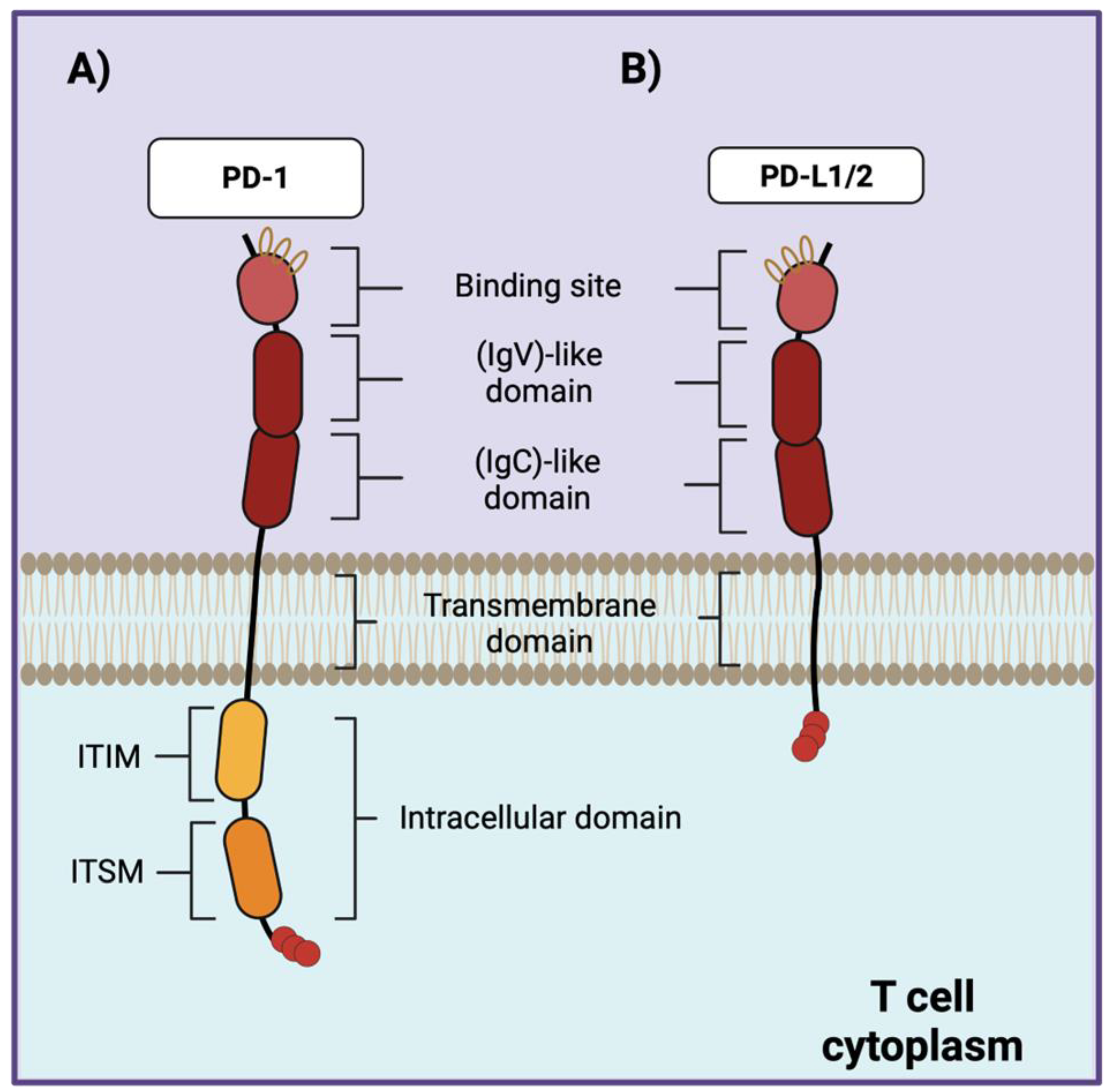
Figure 2.
PD-1 expression on the surface of different CD4+ T cells in autoimmunity. PD-1 expression has been studied in different subpopulations of CD4+ T cells like Tfh, cTfh, Tph, Treg, and Tfr cells. Tfh: T follicular helper cells, cTfh: circulating T follicular helper cells; Tph: T peripheral helper cells; Treg: regulatory T cells; Tfr: follicular regulatory T cells.
Figure 2.
PD-1 expression on the surface of different CD4+ T cells in autoimmunity. PD-1 expression has been studied in different subpopulations of CD4+ T cells like Tfh, cTfh, Tph, Treg, and Tfr cells. Tfh: T follicular helper cells, cTfh: circulating T follicular helper cells; Tph: T peripheral helper cells; Treg: regulatory T cells; Tfr: follicular regulatory T cells.
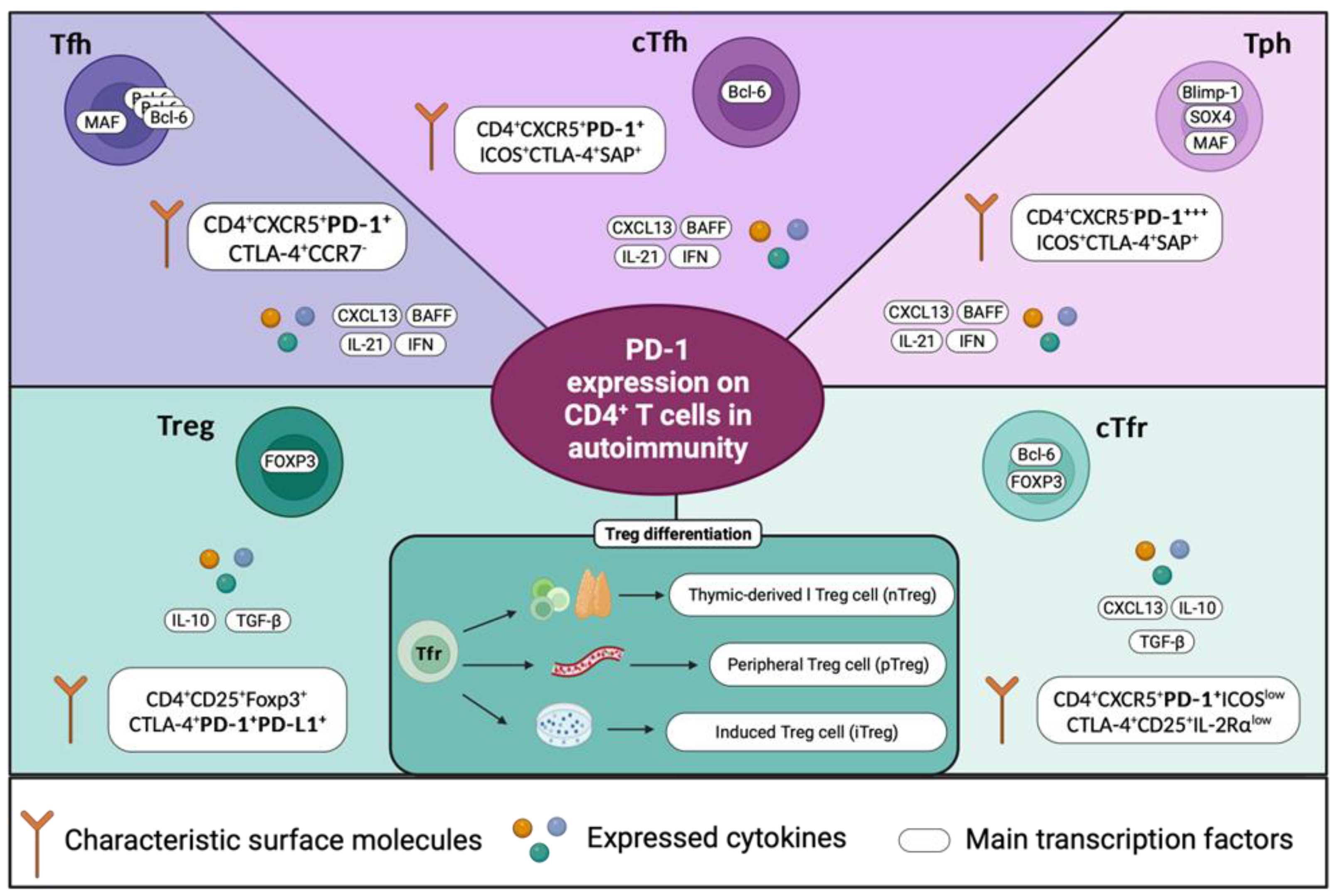
Figure 3.
Expression of PD-1 and its ligands PD-L1 and PD-L2. A) The expression of PD-1 is stimulated by various transcription factors generated after TCR activation and cytokine stimulation. B) PD-L1 can also bind to CD80, and PD-L2 can also bind to RGMb molecule. C) PD-1 and its ligands can also be found in a soluble form generated by alternative splicing or proteolytic cleavage. PD-1: Programmed death 1; PD-L1: Programmed cell death-ligand 1; PD-L2: Programmed cell death-ligand 2; TCR: T-cell receptor; RGMb: Repulsive Guidance Molecule b.
Figure 3.
Expression of PD-1 and its ligands PD-L1 and PD-L2. A) The expression of PD-1 is stimulated by various transcription factors generated after TCR activation and cytokine stimulation. B) PD-L1 can also bind to CD80, and PD-L2 can also bind to RGMb molecule. C) PD-1 and its ligands can also be found in a soluble form generated by alternative splicing or proteolytic cleavage. PD-1: Programmed death 1; PD-L1: Programmed cell death-ligand 1; PD-L2: Programmed cell death-ligand 2; TCR: T-cell receptor; RGMb: Repulsive Guidance Molecule b.
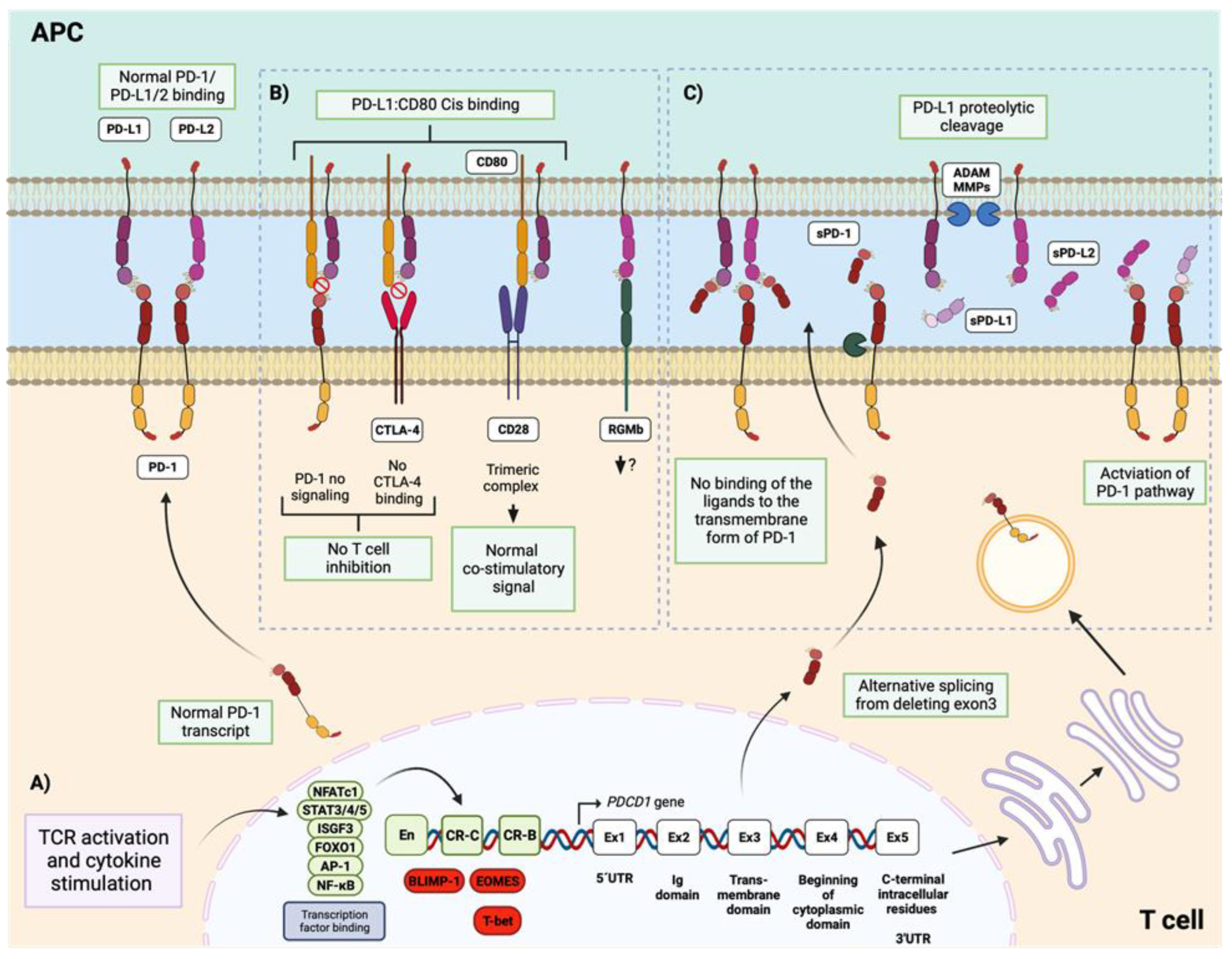
Figure 4.
Regulation of PD-L1 expression. CMTM6/4 can bind to PD-L1 on the plasma membrane, maintaining its expression on the cell surface as well as in early endosomes. This interaction favors recycling, thereby preventing ubiquitination and degradation by the proteasome. In contrast, CDK4 promotes the ubiquitination of PD-L1. HDAC2 deacetylates PD-L1, facilitating its translocation to the nucleus where PD-L1 can activate genes involved in the regulation of its own expression. Post-translational modifications, such as glycosylation (N35, N192, N200, N219), phosphorylation (Try112), and palmitoylation (C272), contribute to the stabilization, increased expression, extended half-life, and maintenance of cellular distribution of PD-L1. Conversely, ubiquitination (CSN5, c-Cbl, and CDK4) and phosphorylation (S195) promote lysosomal and proteasomal degradation. HDAC2: histone deacetylase; CDK4: Cyclin D/cyclin-dependent kinase 4; PD-1: Programmed death 1; PD-L1: Programmed cell death-ligand 1; CMTM6: CKLF-like MARVEL transmembrane domain-containing protein 6; IFN-y/a/B: interferon-gamma/alpha/betha; IRF1: Interferon-gamma receptor; GAS: Gamma interferon Activated Sites; STAT: Signal Transducer and Activator of Transcription; JAK: janus kinase mRNA: messenger RNA; ER: endoplasmic reticulum.
Figure 4.
Regulation of PD-L1 expression. CMTM6/4 can bind to PD-L1 on the plasma membrane, maintaining its expression on the cell surface as well as in early endosomes. This interaction favors recycling, thereby preventing ubiquitination and degradation by the proteasome. In contrast, CDK4 promotes the ubiquitination of PD-L1. HDAC2 deacetylates PD-L1, facilitating its translocation to the nucleus where PD-L1 can activate genes involved in the regulation of its own expression. Post-translational modifications, such as glycosylation (N35, N192, N200, N219), phosphorylation (Try112), and palmitoylation (C272), contribute to the stabilization, increased expression, extended half-life, and maintenance of cellular distribution of PD-L1. Conversely, ubiquitination (CSN5, c-Cbl, and CDK4) and phosphorylation (S195) promote lysosomal and proteasomal degradation. HDAC2: histone deacetylase; CDK4: Cyclin D/cyclin-dependent kinase 4; PD-1: Programmed death 1; PD-L1: Programmed cell death-ligand 1; CMTM6: CKLF-like MARVEL transmembrane domain-containing protein 6; IFN-y/a/B: interferon-gamma/alpha/betha; IRF1: Interferon-gamma receptor; GAS: Gamma interferon Activated Sites; STAT: Signal Transducer and Activator of Transcription; JAK: janus kinase mRNA: messenger RNA; ER: endoplasmic reticulum.
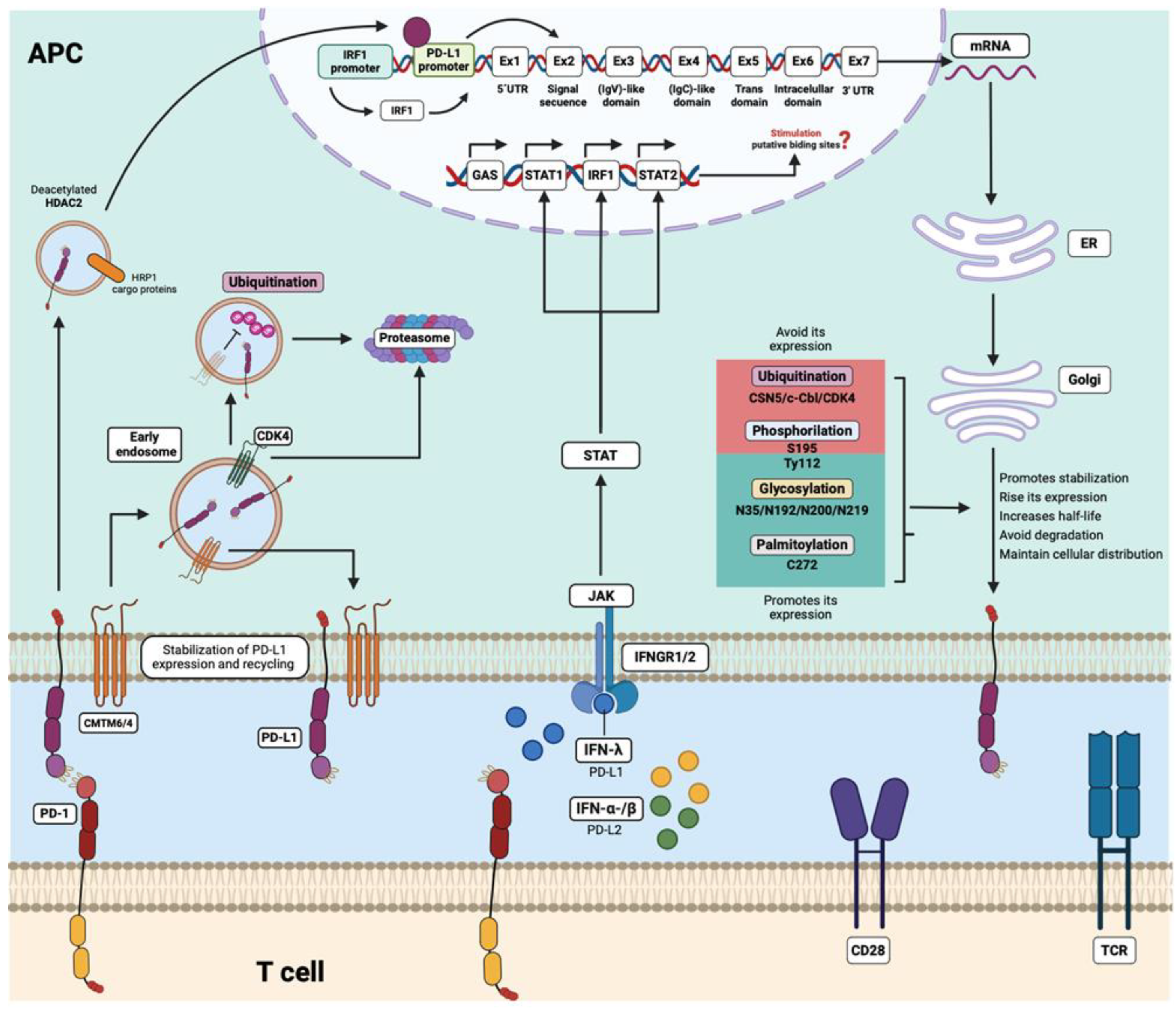

Table 1.
PD-1, PD-L1, and PD-L2 tissue, cell, and serum expression in systemic lupus erythematosus and other autoimmune diseases.
Table 1.
PD-1, PD-L1, and PD-L2 tissue, cell, and serum expression in systemic lupus erythematosus and other autoimmune diseases.
| Disease | Evaluation | Finding | Reference |
|---|---|---|---|
| SLE | Tissue expression | PD-1 and PD-L1 were identified in the kidney of lupus nephritis patients. | Bertsias, George K et al. 2009 [122] |
| Cell expression | Increased percentages of PD-1-expressing CD3+ T cells and PD-1-expressing CD19+ B cells in SLE. High expression of PD-L1-expressing CD19+B cells and PD-L2-expressing CD14+ monocytes. |
Liu, Ming-Fei et al. 2009 [123] |
|
| Elevated frequency of PD-L1-expressing neutrophils in SLE. This percentage decreased after receiving a 15-day regular treatment with corticosteroids and immunosuppressive drugs. | Luo, Qing et al. 2016 [124] | ||
| PD-L1 was significantly higher in CD19+ cells of SLE patients with active disease and LN. The expression of PD-L1 was increased in double-negative B cells (DN) and plasma cells (PC). The percentage of CD19+PD-L1+ cells correlated with the disease activity index and the T follicular helper cells (Tfh) frequency. |
Jia, Xiao-Yun et al. 2019 [125] |
||
| Higher frequency of PD-1, PD-L1, PD-L2, and CD86 in CD19+CD20- B than in CD19+CD20+ B cells. | Zhu, Qingqing et al. 2021 [126] |
||
| Higher expression of PD-1 and PD-L1 in CD11c+ B cells. | Rincon-Arevalo, Hector et al. 2021 [127] | ||
| Soluble levels | Increased serum levels of anti-PD-1 IgG in new-onset SLE patients. | Shi, Hui et al. 2017 [128] | |
| Higher serum levels of sPD-1 and sPD-L1. | Du, Yan et al. 2020 [111] | ||
| Higher serum levels of sPD-1 and sPD-L2 in SLE patients with high disease activity. sPD-L1 was not elevated in SLE patients. |
Hirahara, Shinya et al. 2020 [110] | ||
| Higher expression of membrane-bound PD-L2 on monocytes. Lower serum levels of sPD-L2. |
Tong, Min et al. 2020 [129] | ||
| RA | Tissue expression | PD-L2 was highly expressed on macrophages in the synovial tissue. | Xiong, Jian et al. 2023 [130] |
| Low PD-L1 expression in RA synovial tissue | Guo, Yanxia et al. 2018 [131] | ||
| PD-1 expression on synovium infiltrating lymphocytes. PD-L1 expression on synovial lining cells. |
Matsuda, Kotaro et al. 2018 [132] | ||
| Cell expression | Increased percentages of PD-L2-expressing CD14+ monocytes. | Xiong, Jian et al. 2023 [130] | |
| Increased PD-L1 expression on synovial fluid mDCs compared with peripheral blood mDCs | Moret, Frederique M et al. 2014 [133] | ||
| Soluble levels | Lower sPD-L2 in the serum of RA patients. | Xiong, Jian et al. 2023 [130] | |
| sPD-1 levels are increased in ACPA+ but not ACPA- early RA | Guo, Yanxia et al. 2018 [131] | ||
| Elevated sPD-1 and sPD-L1 levels in serum and synovial fluid of RA | Wan, Bing et al. 2006 [134] | ||
| Higher sPD-1 in early and chronic RA | Greisen, S R et al. 2014 [135] | ||
| Elevated sPD-1 in RA serum | Bommarito, D et al. 2017 [ 136] | ||
| pSS | Tissue expression |
PD-1 expression on infiltrating lymphocytes in the salivary gland from 52% of SS patients. PD-L1 expression on ductal and acinar epithelial cells from 68% of SS patients. |
Kobayashi, Masaya et al. 2005 [137] |
| PD-1 and PD-L1 expression in labial glands were higher than in non-pSS controls | Qian, Sirui et al. 2022 [116] | ||
| Cell expression | Increased expression of PD-L1 in CD11c+ B cells of pSS patients | Rincon-Arevalo, Hector et al. 2021 [127] | |
| Serum levels | Lower sPD-L2 serum levels in pSS patients | Loureiro-Amigo, Jose et al. 2021 [138] | |
| Elevated sPD-L2 serum levels in pSS patients | Nishikawa, Ayumi et al. 2016 [139] | ||
| Elevated sPD-1 and sPD-L1 serum levels in pSS patients | Qian, Sirui et al. 2022 [116] | ||
| MS | Tissue expression | Astrocytes in white matter lesions from MS patients upregulate PD-L1 in response to aryl hydrocarbon receptor and interferon signaling | Linnerbauer, Mathias et al. 2023 [140] |
| Cell expression | Frequency of PD-L1-expressing CD19+ B cells, and PD-L1+/IL-10+CD14+ monocytes are higher in stable multiple sclerosis patients compared to acute MS (AMS) patients | Trabattoni, Daria et al. 2009 [141] | |
| Serum levels | No information |
Systemic Lupus Erythematosus (SLE), Rheumatoid Arthritis (RA), Primary Sjögren's Syndrome (pSS), and Primary Sjögren's Syndrome (MS).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



