
Submitted:
21 June 2024
Posted:
24 June 2024
You are already at the latest version
Abstract
Detection methods have been developed to prevent transmission of zoonotic or xenozoonotic porcine viruses after transplantation of pig organs or cells to the recipient (xenotransplantation). Eleven xenotransplantation-relevant viruses including porcine cytomegalovirus, a porcine roseolovirus (PCMV/PRV), porcine lymphotropic herpesviruses -1, -2, -3 (PLHV-1, 2, 3), porcine parvovirus (PPV), porcine circovirus 2, 3, 4 (PCV2, 3, 4), hepatitis E virus genotype 3 (HEV3), porcine endogenous retrovirus-C (PERV-C) and recombinant PERV-A/C have been selected. In the past, several pig breeds, minipigs and genetically modified pigs generated for xenotransplantation had been analyzed using these methods. Here, spleen, liver and blood samples from 10 German slaughterhouse pigs were screened using both, PCR-based and immunological assays. Five viruses: PCMV/PRV, PLHV-1, PLHV-3 and PERV-C were found in all animals, and PCV3 in one animal. Some animals were latently infected with PCMV/PRV as only virus-specific antibodies were detected. Others were also PCR positive in spleen and/or liver, indicative of an ongoing infection. These results provide important information on the viruses that infect German slaughterhouse pigs and together with results of previous studies revealed that the methods and test strategies efficiently work under field conditions.
Keywords:
porcine viruses
; xenotransplantation
; virus safety
; herpes viruses x porcine endogenous retroviruses x latency
1. Introduction
Xenotransplantation is under development to alleviate the shortage of human organs for transplantation [1]. Pigs are the species of choice and multiple genetically modified pigs have been generated to prevent hyperacute and acute rejection of pig organs [2]. In preclinical studies with non-human primates using organs from such pigs, using novel immunosuppressive drugs, considerable survival times of the xenotransplant were achieved in the last several years. Based on these studies, a heart from a pig with 10 genetic modifications (10 GE pig) was transplanted into a patient at the University of Maryland in Baltimore (UMB) in 2022. The patient survived 2 months [3]. In the year before, surgeons from the New York University (NYU) Langone Transplant Institute have xenotransplanted αGalT-KO pig thymo-kidneys into a brain-dead human recipient, which lasted for 54 hours [4,5]. In a study at the University of Alabama in Birmingham (UAB), decedent recipients were nephrectomised and bilaterally transplanted with kidneys from 10 GE pigs. Patients received a conventional immunosuppression and the first case was monitored for 74 hours during which urine production was poor and creatinine clearance did not improve [6]. Furthermore, at NYU 10 GE pig hearts were transplanted into brain-dead patients [7]. In China, a 50-year-old clinically dead man was the first person to receive a liver from a pig [8]. Also in China, at the Xijing Hospital of the Air Force Medical University in Xian, the kidney of a multi-gene-edited pig was transplanted into a brain-dead human. The kidney was functioning well for more than 13 [9]. At the UMB a second heart was transplanted into a patient, who died after 6 weeks [10]. Recently, pig kidneys with 69 genetic modifications were transplanted at the Massachusetts General Hospital in Boston, the patient was in excellent health and had been discharged from hospital [11]. He died after 7 weeks from a heart attack, the kidney worked still well at that time. At the NYU Langone Transplant Institute, a left ventricular assist device and a pig kidney with only one genetic modification, a knock out of the gene responsible for the production of an alpha-gal residues, was transplanted in a living person [12].
Xenotransplantation could result in the transmission of potentially pathogenic microorganisms to the recipient. The transmission of the porcine cytomegalovirus, a porcine roseolovirus (PCMV/PRV), to the pig heart recipient in Baltimore [3], and the virus contributed obviously to the death of the patient, demonstrates how important it is to prevent virus transmission [13]. In the past it was shown in several studies that transmission of PCMV/PRV with pig heart and kidney transplants into non-human primates (NHP) resulted in a significant reduction of the survival time of the xenotransplant [14,15,16,17]. It is important to be aware that microorganisms including viruses such as human immunodeficiency virus, human cytomegalovirus and rabies virus have also been transmitted in allotransplantations [18].
The data mentioned above underlines the importance of the safety research in this field. Consequently, in the last few years numerous PCR-based and immunological methods have been developed to detect porcine viruses [19,20,21,22]. At present it is still unclear which viruses may pose a risk during xenotransplantation, i.e., which viruses are xenotransplantation-relevant [23,24]. On one hand there are known zoonotic viruses (zoonosis means inducing disease in the recipient) such as the hepatitis E virus, genotype 3 (HEV3). On the other hand we now know viruses which do not harm healthy people, but harm patients in the context of xenotransplantation such as PCMV/PRV. These viruses should be called xenozoonotic [25]. Unfortunately, tests for these pig viruses are not available at most veterinary diagnostic institutes, but only in very few specialized laboratories [26].
To test the developed methods under field conditions, we have used them in the past for screening not only pigs generated for xenotransplantation and the corresponding non-human primate recipients [16,27], but also for a comprehensive screening of pig breeds such as the indigenous Greek black pigs [28], the Göttingen minipigs [29,30,31,32], the Aachen minipigs [33], the Mini LEWE minipigs [34], Göttingen minipigs with dippity pig syndrome [35], and Greek pigs with erythema multiforme [36]. Here, we screen German slaughterhouse pigs using PCR-based and immunological methods able to detect pig viruses and demonstrate again the robust functionality of these methods. Furthermore, we compare the data to previously tested pig breeds
2. Materials and Methods
2.1. Animals and Tissues
Tissue samples from spleen, liver and clotted blood were obtained from pigs presenting a breeding between Duroc-boars Danish landrace x Danish- Large White (Edelschwein) hybrid sows from a slaughterhouse near Berlin, their age was 6 months. Sex and weight are shown in Table 1. The samples were frozen at -80oC until analyzed.
2.2. Isolation of DNA and RNA
DNA was isolated from the tissues and purified PBMCs according to the manufacturer’s instructions using the DNeasy Blood & Tissue kit (Qiagen, Hilden, Germany). DNA concentrations were determined using a NanoDrop ND-1000 (Thermo Fisher Scientific Inc., Worcester, MA, USA) or Qubit device (Invitrogen, Thermo Fisher Scientific Inc., Worcester, MA, USA).
2.3. Real-Time PCR for the Detection of DNA Viruses
Real-time PCRs using specific primers and probes were performed with a defined sensitivity to detect PCMV/PRV (sensitivity 10 copies), PLHV-1 (1 copy), PLHV-2 (1 copy), PLHV-3 (1 copy), PCV2 (1 copy), PCV3 (10 copies), PCV4 (100 copies) and PPV1 (10 copies) as described previously (Table 2) [28]. All protocols were performed using the SensiFAST Probe No-ROX kit (Meridian Bioscience, Cincinnati, OH, USA) in a reaction volume of 16 µl plus 4 µl (100 ng) of DNA template. All real-time PCRs were carried out as duplex PCRs testing simultaneously the viral gene of interest and porcine glyceraldehyde-3-phosphate-dehydrogenase (pGAPDH) as internal control for each sample. Furthermore, virus specific gene blocks were used to verify the functionality of the PCR [34]. Real-time PCR reactions were carried out with a qTOWER3 G qPCR cycler (Analytik Jena, Jena, Germany) and the real-time PCR conditions as previously described [28].
2.4. Real-Time Reverse Transcriptase PCR for the Detection of HEV3
Real-time reverse transcriptase-PCR (real-time RT-PCR) as described by Jothikumar et al. [37] was carried out to detect hepatitis E virus genotype 3 (HEV3). All real-time RT-PCR reactions were performed in a reaction volume of 16 µl using SensiFAST Probe No-ROX One-Step Kit (Meridian Bioscience, Cincinnati, OH, USA) plus 4 µl (100 ng) template RNA. The reaction was performed with a qTOWER3 G PCR cycler (Analytik Jena, Jena, Germany). The temperature-time profile applied consists of a reverse transcriptase step of 30 minutes at 50°C, followed by an activation step of 15 minutes at 95°C and 45 cycles comprising a step of 10 seconds at 95°C, followed by a step of 20 seconds at 55°C and 15 seconds at 72°C [38].
2.5. Conventional and Real-Time PCR for the Detection of PERVs
A conventional PCR was performed to determine the presence of PERV-C. PERV-C was detected using a set of primers with an amplicon length of 288 bp (described as PCR4 in [45]). It was carried out with AmpliTaq DNA Polymerase (Applied Biosystems, Waltham, MA, USA) and was set up with a Biometra TRIO cycler (Analytik Jena, Jena, Germany). The following temperature-time profile was used: 95°C for 10 minutes (activation step), followed by 45 cycles at 95°C for 15 seconds (denaturation), 55°C for 30 seconds (annealing) and 72°C for 30 seconds (extension) and a final extension at 72°C for 5 minutes.
A conventional PCR to determine the presence of human-tropic PERV-A/C, was set up using specific primer pairs (Table 2) [46]. The PERV-A/C long primer mix detects an amplicon of 1266 bp length. It was carried out with AmpliTaq DNA Polymerase (Applied Biosystems, Waltham, MA, USA) and was set up with a Biometra TRIO cycler (Analytik Jena, Jena, Germany). The following temperature-time profile was used: 95°C for 10 minutes (activation step), followed by 45 cycles at 95°C for 15 seconds (denaturation), 55°C for 30 seconds (annealing) and 72°C for 90 seconds (extension) and a final single cycle at 72°C for 5 minutes.
2.6. Western Blot Analysis
3. Results
3.1. Results of the PCR-Based Screening
DNA was isolated from the spleen and liver samples and real-time PCRs were performed to screen for the DNA viruses (Table 3). In addition, a conventional PCR and a real-time reverse transcriptase PCR were performed to test for PERV-A/C and HEV3, respectively (Table 3). PCMV/PRV was found in the spleen of 6 and in the liver of one of 10 animals tested. PLHV-1 was found in all 10 animals, both in the spleen and the liver. The same result was obtained for PLHV-3. PCV3 was found in the spleen and the liver of one animal. PERV-C was found integrated in the genome of all animals using a real-time PCR method and in 9 of 10 animals using conventional PCRs designated PCR1 and PCR4 [45]. The real-time testing for PERV-C was positive in all 10 animals. The animals were negative for the other viruses (PLHV-2, PPV-1, PCV2, PCV4, PERV-A/C and HEV) in both organs (Table 3).
3.1. Results of the Western Blot-Based Screening
Plasma samples from the slaughterhouse pigs were screened for antibodies against PCMV/PRV by Western blotting using the recombinant R2 domain of the gB protein of PCMV/PRV as antigen. All animals were antibody positive, highighting that all animals were PCMV/PRV infected, either latently or an active infection (Table 1, Figure 1).
4. Discussion
To apply our methods developed to detect xenotransplantation-relevant porcine viruses, we screened 10 German slaughterhouse pigs. Surprisingly, the animals contained four DNA viruses, PCMV/PRV, PLHV-1, PLHV-3 and PCV3, although the animals were healthy and intended for consumption. There are hundreds of publications investigating the prevalence of pig viruses in animals world-wide. In most cases they were screened for HEV [48,49,50], influenza virus [51,52], porcine reproductive and respiratory syndrome (PRRSV) [53] and Aujeszky’s disease virus [52]. For example, in the case of PRRSV, 74.3% of 1039 serum samples were seropositive and 1.9% of the 1027 meat samples were positive by PCR in Canadian abattoirs. The virus was infectious when meat was ingested by uninfected pigs [53].
When we recently screened indigenous Greek black pigs using the same methods as described here, we found PCMV/PRV, PLHV-1, PLHV-2, PLHV-3, PCV2, and PCV3 [28] (Table 4). These animals were among all pigs we tested, the breed containing the highest number of pig viruses (Table 4). Since the animals were healthy and their meat was indented for consumption, it is possible that the animals have a natural resilience to virus infections, e.g., due to antiviral restriction factors [28]. Whether the German slaughterhouse pigs also have such restriction factors remains unclear.
All German slaughterhouse pigs had antibodies against PCMV/PRV, indicating that all were infected, despite the fact that virus DNA was not found in liver and spleen of all animals by real-time PCR. This confirms previous results that PCMV/PRV, like all herpesviruses, establishes latency, during which it can no longer be detected using PCR methods [47]. This was the case for four animals when testing the DNA from spleen and nine animals when testing the DNA from liver. This is the only virus where differences between the detection in the liver and the detection in the spleen was observed, all other viruses were found in both organs concurrently.
Obviously at this stage of infection more cells containing PCMV/PRV DNA remained in the spleen compared to the liver. Whether these cells are still expressing PCMV/PRV genes or even produce virus particles or whether the DNA is derived from circulating latently infected cells, remains unknown. As a rule, PCMV/PRV can be detected by PCR using nasal swabs only when an animal is freshly infected and suffers from rhinitis. PCMV/PRV can only be detected in blood and organs of young animals by PCR only until approximately 19 weeks after infection [47]. In older animals, only the detection of virus-specific antibodies provides reliable data. For this either a Western blot analysis using recombinant proteins [19,47] or an ELISA using synthetic peptides derived from the same region of the glycoprotein B were applied [54]. Both methods gave comparable results [55]. Antibodies against PCMV/PRV can also be detected in the blood of young animals. However, these antibodies are mostly from the infected mother sow, transmitted to the piglet by colostrum [47]. The transmission of PCMV/PRV to the first patient receiving a pig heart in Baltimore showed that screening for PCMV/PRV is not trivial. In this case unfortunately only nasal swab samples were taken from the donor animal and analyzed by PCR [3]. However, nasal swabs will only be positive when the animal is freshly infected and do not allow detection of latently infected animals [47,56].
Although PCMV/PRV is widely distributed in pigs worldwide and nearly all German slaughterhouse pigs are PCMV-positive [19, this manuscript], the impact of PCMV/PRV on pig breeding seems to be low. PCMV/PRV infection is mostly acquired early in life and infection results in seroconversion and life-long latent infection [57]. Fatalities were mainly observed in piglets less than 3 weeks old and virus-infected sows are prone to abortion [57]. There are no antiviral drugs and no vaccines against PCMV/PRV. However, the virus can be easily removed from a pig population by early weaning [48]. Since PCMV/PRV is a roseolovirus, closely related to human herpesviruses 6 and 7 (HHV6, 7) and only distantly related to the human cytomegalovirus (HCMV), all drugs effective against HCMV do not act against PCMV/PRV [59,60]. Various drugs against HCMV that were used to treat the first patient receiving a pig heart in Baltimore did not reduce the PCMV/PRV load in the patient’s blood [61].
When we screened for porcine lymphotropic herpesviruses, PLHV-1 and PLHV-3 were found in all animals. PLHV-1/3 are gammaherpesviruses widely distributed in pigs (up to 80%% in single farms), but have not been associated with any pig disease (for review see [62]). At present there are no antiviral drugs or vaccines available. In contrast to PCMV/PRV, Caesarean delivery was not or only partially successful to eliminate these viruses [63,64]. PLHV was detected in all of the eight genetically modified pigs used for orthotopic heart transplantation, however, the virus was not transmitted to the baboon recipients [16].
When we screened for circoviruses, we found only one pig which was infected with PCV3. The virus was found in both, the liver and the spleen of animal 4. PCV1 is not pathogenic for pigs and was therefore not included into the screening. However, since we do not know whether it may be pathogenic for humans, it should be included in future testing of pigs intended for xenotransplantation. PCV2 induces an entire complex of porcine circovirus diseases (PCVD), there exist several vaccines against PCV2 which are able to prevent diseases, but in most cases, they do not prevent the transmission of the virus (for review see [65]). PCV3 was discovered in 2016, it is common in domestic pigs and wild boars world-wide (for review see [66]). Since PCV3 was also found in healthy pigs, there is clear evidence that PCV3 is pathogenic since virus clones were able to induce porcine dermatitis and nephropathy syndrome (PDNS) in specified pathogen-free animals [67] PCV3 was also found in Greek pigs with erythema multiform [36] and in Göttingen minipigs with dippity pig syndrome [35]. In contrast to our findings in a German slaughterhouse, the prevalence of PCV2, PCV3, and PCV4 in slaughterhouses in one province in China was 56.8, 80, and 9.4%, respectively [68]. PCV3 was also found in pigs generated for xenotransplantation, which appeared clinically healthy and the virus was transmitted in a few cases to baboons after transplantation of hearts from these pigs [69].
When we screened for HEV, all animals were negative. This is remarkable because in other slaughterhouses higher amounts of positive animals were found. For example, HEV IgG was detected in sera from 167 pigs among 250 tested animals (67.6 %) and HEV RNA was detected 25 (11.0 %) liver samples in Dutch slaughterhouses [50]. HEV RNA was detected in 6.3% and HEV IgG in 40% of 5,033 serum samples from market-weight pigs at 25 slaughterhouses in 10 US states [49]. HEV is of great importance for xenotransplantation because it represents a proven zoonotic virus, it may induce diseases in humans, mainly a moderate hepatitis and neurological syndromes (for review see [70,71]). HEV was found in Göttingen minipigs which are produced under specified pathogen-free conditions using real-time PCR and antibody detection by Western blot assays and a transplacental mother to piglet transmission was demonstrated [29].
Since all pigs carry PERV-A and PERV-B proviruses in their genome, we only tested for PERV-C and recombinant PERV-A/C. PERV-C was found in all German slaughterhouse pigs as well in all other pig breeds screened for, with exception of the Göttingen minipigs at the Göttingen university (Table 4). It is interesting that in the German slaughterhouse pigs PERV-C was detected in all pigs using the real-time PCR, but PERV-C was not detected in animal 3 using conventional PCRs (Table 3). The PCRs used were designated PCR1 and PCR4 by Kaulitz et al. [45]. PCR4 was shown highly reliable in detection of PERV-C, comparable in reliability to PCR1 developed by Takeuchi et al. [72]. The reason for the discrepancy between PCR1 and PCR4 on one hand and the real-time PCR on the other hand is under investigation (Jhelum et al., in preparation). Recombinants of PERV-A with PERV-C (PERV-A/C), which are characterized by higher virus titers, and the ability to infect human cells, were not found in all investigated animals. This confirm previous findings that PERV-A/C recombinants are rare and found mainly in minipigs (for review see [73]).
The methods used here and in previous investigations (Table 4) can not only be used for screening animals generated for xenotransplantation. They should also be used to screen all pigs used for biomedical research, because the presence of these viruses may interfere with the result of numerous biomedical experiments. Most importantly, these methods can also be used to eliminate these viruses from pig herds produced for consumption. This would improve the health of the animals, minimize losses in pig breeding and lead to higher profits in meat production. Xenotransplantation using pig cells, tissues and organs may be save and prolong the lives of patients, but it may also be associated with the transmission of porcine microorganisms to the recipient, eventually resulting in emerging infectious diseases. For this reason, the health of both the donor animals and the human recipients represents a special and sensitive case of the One Health concept [74].
5. Conclusions
PCR-based and immunological methods were used to screen German slaughterhouse pigs for eleven xenotransplantation-relevant viruses including PERV. All pigs with exception of one contained PERV-C in their genome. In addition, four DNA viruses were found. Together with previous screening of numerous other pig breeds this indicates that these detection methods work well under field conditions. These diagnostic tests will be used to screen multiple genetically modified pigs designated for clinical trials of xenotransplantation. Furthermore, they can also be used to screen pigs used in biomedical research to prevent that the viruses influence the resulting data. In pork production they may be used to reduce losses from these virus infections.
Author Contributions
Conceptualization, J.D.; methodology, H.J.; validation, H.J., J.D. and B.K.; investigation, H.J.; writing—original draft preparation, J.D.; writing—review and editing, H.J., J.D. and B.K.; supervision, J.D. and B.K.; funding acquisition, J.D. and B.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Deutsche Forschungsgemeinschaft, TRR127, and the APC was funded by Freie Universität Berlin.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable
Data Availability Statement
Data is contained within the article.
Acknowledgments
We would like to thank Dr. Kostja Renko, German Federal Institute for Risk Assessment, Unit Strategies for Toxicological Assessments, Department Experimental Toxicology and ZEBET, Berlin, and Dr. Gordon Mählis, Veterinäramt Potsdam-Mittelmark for help when acquiring the pig samples, Julia Raddatz and Axel Teigeler, Institute of Virology, for excellent technical support. Above all we thank Ludwig Krabben, Institute of Virology, for critical reading the manuscript and permanent advice.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Cooper, D.K.C.; Gaston, R.; Eckhoff, D.; Ladowski, J.; Yamamoto, T.; Wang, L.; Iwase, H.; Hara, H.; Tector, M.; Tector, A.J. Xenotransplantation-the current status and prospects. Br Med Bull. 2018, 125, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.; Yacoub, M.H.; Ayares, D.; Yamada, K.; Eisenson, D.; Griffith, B.P.; Mohiuddin, M.M.; Eyestone, W.; Venter, J.C.; Smolenski, R.T.; Rothblatt, M. Physiological basis for xenotransplantation from genetically modified pigs to humans. Physiol Rev. 2024, 104, 1409–1459. [Google Scholar] [CrossRef]
- Mohiuddin, M.M.; Singh, A.K.; Scobie, L.; Goerlich, C.E.; Grazioli, A.; Saharia, K.; Crossan, C.; Burke, A.; Drachenberg, C.; Oguz, C.; Zhang, T.; Lewis, B.; Hershfeld, A.; Sentz, F.; Tatarov, I.; Mudd, S.; Braileanu, G.; Rice, K.; Paolini, J.F.; Bondensgaard, K.; Vaught, T.; Kuravi, K.; Sorrells, L.; Dandro, A.; Ayares, D.; Lau, C.; Griffith, B.P. Graft dysfunction in compassionate use of genetically engineered pig-to-human cardiac xenotransplantation: A case report. Lancet 2023, 402, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Loupy, A.; Goutaudier, V.; Giarraputo, A.; Mezine, F.; Morgand, E.; Robin, B.; Khalil, K.; Mehta, S.; Keating, B.; Dandro, A.; Certain, A.; Tharaux, P.L.; Narula, N.; Tissier, R.; Giraud, S.; Hauet, T.; Pass, H.I.; Sannier, A.; Wu, M.; Griesemer, A.; Ayares, D.; Tatapudi, V.; Stern, J.; Lefaucheur, C.; Bruneval, P.; Mangiola, M.; Montgomery, R.A. Immune response after pig-to-human kidney xenotransplantation: A multimodal phenotyping study. Lancet 2023, 402, 1158–1169. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, R.A.; Stern, J.M.; Lonze, B.E.; Tatapudi, V.S.; Mangiola, M.; Wu, M.; Weldon, E.; Lawson, N.; Deterville, C.; Dieter, R.A.; Sullivan, B.; Boulton, G.; Parent, B.; Piper, G.; Sommer, P.; Cawthon, S.; Duggan, E.; Ayares, D.; Dandro, A.; Fazio-Kroll, A.; Kokkinaki, M.; Burdorf, L.; Lorber, M.; Boeke, J.D.; Pass, H.; Keating, B.; Griesemer, A.; Ali, N.M.; Mehta, S.A.; Stewart, Z.A. Results of Two Cases of Pig-to-Human Kidney Xenotransplantation. N Engl J Med. 2022, 386, 1889–1898. [Google Scholar] [CrossRef] [PubMed]
- Locke, J.E.; Kumar, V.; Anderson, D.; Porrett, P.M. Normal graft function after pig-to-human kidney xenotransplant. JAMA Surg 2023, 158, 1106–1108. [Google Scholar] [CrossRef] [PubMed]
- Moazami, N.; Stern, J.M.; Khalil, K.; Kim, J.I.; Narula, N.; Mangiola, M.; Weldon, E.P.; Kagermazova, L.; James, L.; Lawson, N.; Piper, G.L.; Sommer, P.M.; Reyentovich, A.; Bamira, D.; Saraon, T.; Kadosh, B.S.; DiVita, M.; Goldberg, R.I.; Hussain, S.T.; Chan, J.; Ngai, J.; Jan, T.; Ali, N.M.; Tatapudi, V.S.; Segev, D.L.; Bisen, S.; Jaffe, I.S.; Piegari, B.; Kowalski, H.; Kokkinaki, M.; Monahan, J.; Sorrells, L.; Burdorf, L.; Boeke, J.D.; Pass, H.; Goparaju, C.; Keating, B.; Ayares, D.; Lorber, M.; Griesemer, A.; Mehta, S.A.; Smith, D.E.; Montgomery, R.A. Pig-to-human heart xenotransplantation in two recently deceased human recipients. Nat Med 2023, 29, 1989–1997. [Google Scholar] [CrossRef]
- First pig liver transplanted into a person lasts for 10 days. Nature 2024, 627, 710–711. Available online: https://www.nature.com/articles/d41586-024-00853-8 (accessed on 13 June 2024). [CrossRef] [PubMed]
- Kidney transplantation in China. Available online: https://www.globaltimes.cn/page/202404/1310234.shtml (accessed on 13 June 2024).
- Transplantation of the second heart in Baltimore. Available online: https://www.nytimes.com/2023/10/31/health/pig-heart-transplant-faucette.html?smid=nytcore-ios-share&referringSource=articleShare (accessed on 13 June 2024).
- Kidney in living individual in Boston STAT. Available online: https://www.statnews.com/2024/03/21/first-xenotransplantation-crispr-gene-edited-pig-kidney-egenesis/?utm_campaign=breaking_news&utm_medium=email&_hsmi=299239261&utm_content=299239261&utm_source=hs_email (accessed on 13 June 2024).
- Available online: https://nyulangone.org/news/first-ever-combined-heart-pump-gene-edited-pig-kidney-transplant-gives-new-hope-patient-terminal-illness (accessed on 8 May 2024).
- Fishman, J.A. Risks of Infectious Disease in Xenotransplantation. N Engl J Med. 2022, 387, 2258–2267. [Google Scholar] [CrossRef]
- Yamada, K.; Tasaki, M.; Sekijima, M.; Wilkinson, R.A.; Villani, V.; Moran, S.G.; Cormack, T.A.; Hanekamp, I.M.; Hawley, R.J.; Arn, J.S.; Fishman, J.A.; Shimizu, A.; Sachs, D.H. Porcine cytomegalovirus infection is associated with early rejection of kidney grafts in a pig to baboon xenotransplantation model. Transplantation. 2014, 98, 411–418.3. [Google Scholar] [CrossRef]
- Sekijima, M.; Waki, S.; Sahara, H.; Tasaki, M.; Wilkinson, R.A.; Villani, V.; Shimatsu, Y.; Nakano, K.; Matsunari, H.; Nagashima, H.; Fishman, J.A.; Shimizu, A.; Yamada, K. Results of life-supporting galactosyltransferase knockout kidneys in cynomolgus monkeys using two different sources of galactosyltransferase knockout Swine. Transplantation. 2014, 98, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Denner, J.; Längin, M.; Reichart, B.; Krüger, L.; Fiebig, U.; Mokelke, M.; Radan, J.; Mayr, T.; Milusev, A.; Luther, F.; Sorvillo, N.; Rieben, R.; Brenner, P.; Walz, C.; Wolf, E.; Roshani, B.; Stahl-Hennig, C.; Abicht, J.M. Impact of porcine cytomegalovirus on long-term orthotopic cardiac xenotransplant survival. Sci Rep. 2020, 10, 17531. [Google Scholar] [CrossRef]
- Denner, J. Xenotransplantation and porcine cytomegalovirus. Xenotransplantation. 2015, 22, 329–335. [Google Scholar] [CrossRef]
- Nellore, A.; Fishman, J.A. Donor-derived infections and infectious risk in xenotransplantation and allotransplantation. Xenotransplantation. 2018, 25, e12423. [Google Scholar] [CrossRef] [PubMed]
- Plotzki, E.; Keller, M.; Ivanusic, D.; Denner, J. A new Western blot assay for the detection of porcine cytomegalovirus (PCMV). J Immunol Methods. 2016, 437, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Noordergraaf, J.; Schucker, A.; Martin, M.; Schuurman, H.J.; Ordway, B.; Cooley, K.; Sheffler, M.; Theis, K.; Armstrong, C.; Klein, L.; Hansen, D.; Olson, M.; Schlechter, L.; Spizzo, T. Pathogen eliminationand prevention within a regulated, Designated Pathogen Free, closed pig herd for long-term breeding and production of xeno-transplantation materials. Xenotransplantation 2018, 25, e12428. [Google Scholar] [CrossRef]
- Denner, J. Sensitive detection systems for infectious agents in xenotransplantation. Xenotransplantation. 2020, 18, e12594. [Google Scholar] [CrossRef] [PubMed]
- Otabi, H.; Miura, H.; Uryu, H.; Kobayashi-Harada, R.; Abe, K.; Nakano, K.; Umeyama, K.; Hasegawa, K.; Tsukahara, T.; Nagashima, H.; Inoue, R. Development of a panel for detection of pathogens in xenotransplantation donor pigs. Xenotransplantation. 2023, 30, e12825. [Google Scholar] [CrossRef]
- Groenendaal, H.; Costard, S.; Ballard, R.; Bienhoff, S.; Challen, D.C.; Dominguez, B.J.; Kern, D.R.; Miller, D.; Noordergraaf, J.; Rudenko, L.; Schuurman, H.J.; Spizzo, T.; Sturos, M.; Zollers, B.; Fishman, J.A. Expert opinion on the identification, risk assessment, and mitigation of microorganisms and parasites relevant to xenotransplantation products from pigs. Xenotransplantation 2023, e12815. [Google Scholar] [CrossRef]
- Krüger, L.; Kristiansen, Y.; Reuber, E.; Möller, L.; Laue, M.; Reimer, C.; Denner, J. A Comprehensive Strategy for Screening for Xenotransplantation-Relevant Viruses in a Second Isolated Population of Gottingen Minipigs. Viruses 2019, 12, 38. [Google Scholar] [CrossRef]
- Denner, J. Zoonosis and xenozoonosis in xenotransplantation: A proposal for a new classification. Zoonoses Public Health. 2023, 70, 578–579. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. Limited availability of methods for the detection of xenotransplantation-relevant viruses in veterinary laboratories. Xenotransplantation 2024, 31, e12851. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.A.; Ludwig, S.; Ludwig, B.; Rotem, A.; Barkai, U.; Bornstein, S.R.; Denner, J. Islet cell transplantation from Gottingen minipigs to cynomolgus monkeys: Analysis of virus safety. Xenotransplantation. 2016, 23, 320–327. [Google Scholar] [CrossRef]
- Jhelum, H.; Papatsiros, V.; Papakonstantinou, G.; Krabben, L.; Kaufer, B.; Denner, J. Screening for Viruses in Indigenous Greek Black Pigs. Microorganisms 2024, 12, 315. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.A.; Morozov, A.V.; Rotem, A.; Barkai, U.; Bornstein, S.; Denner, J. Extended Microbiological Characterization of Göttingen Minipigs in the Context of Xenotransplantation: Detection and Vertical Transmission of Hepatitis E Virus. PLoS ONE 2015, 10, e0139893. [Google Scholar] [CrossRef]
- Morozov, V.A.; Plotzki, E.; Rotem, A.; Barkai, U.; Denner, J. Extended microbiological characterization of Göttingen minipigs: Porcine cytomegalovirus and other viruses. Xenotransplantation. 2016, 23, 490–496. [Google Scholar] [CrossRef]
- Heinze, J.; Plotzki, E.; Denner, J. Virus Safety of Xenotransplantation: Prevalence of Porcine Circovirus 2 (PCV2) in Pigs. Ann Virol Res 2016, 2, 1023. [Google Scholar] [CrossRef]
- Krüger, L.; Kristiansen, Y.; Reuber, E.; Möller, L.; Laue, M.; Reimer, C.; Denner, J. A Comprehensive Strategy for Screening for Xenotransplantation-Relevant Viruses in a Second Isolated Population of Gottingen Minipigs. Viruses. 2019, 12, 38. [Google Scholar] [CrossRef]
- Halecker, S.; Metzger, J.; Strube, C.; Krabben, L.; Kaufer, B.; Denner, J. Virological and Parasitological Characterization of Mini-LEWE Minipigs Using Improved Screening Methods and an Overview of Data on Various Minipig Breeds. Microorganisms. 2021, 9, 2617. [Google Scholar] [CrossRef]
- Plotzki, E.; Heinrichs, G.; Kubícková, B.; Ulrich, R.G.; Denner, J. Microbiological characterization of a newly established pig breed, Aachen Minipigs. Xenotransplantation 2016, 23, 159–167. [Google Scholar] [CrossRef]
- Jhelum, H.; Grand, N.; Jacobsen, K.R.; Halecker, S.; Salerno, M.; Prate, R.; Krüger, L.; Kristiansen, Y.; Krabben, L.; Möller, L.; Laue, M.; Kaufer, B.; Kaaber, K.; Denner, J. First virological and pathological study of Gottingen Minipigs with Dippity Pig Syndrome (DPS). PLoS ONE 2023, 18, e0281521. [Google Scholar] [CrossRef] [PubMed]
- Halecker, S.; Papatsiros, V.; Psalla, D.; Krabben, L.; Kaufer, B.; Denner, J. Virological characterization of pigs with erythema multiforme. Microorganisms 2022, 10, 652. [Google Scholar] [CrossRef] [PubMed]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods. 2006, 131, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Mueller, N.J.; Barth, R.N.; Yamamoto, S.; Kitamura, H.; Patience, C.; Yamada, K.; Cooper, D.K.; Sachs, D.H.; Kaur, A.; Fishman, J.A. Activation of cytomegalovirus in pig-to-primate organ xenotransplantation. J. Virol. 2002, 76, 4734–4740. [Google Scholar] [CrossRef] [PubMed]
- Chmielewicz, B.; Goltz, M.; Franz, T.; Bauer, C.; Brema, S.; Ellerbrok, H.; Beckmann, S.; Rziha, H.-J.; Lahrmann, K.-H.; Romero, C.; et al. A novel porcine gammaherpesvirus. Virology 2003, 308, 317–329. [Google Scholar] [CrossRef]
- McMahon, K.J.; Minihan, D.; Campion, E.M.; Loughran, S.T.; Allan, G.; McNeilly, F.; Walls, D. Infection of pigs in Ireland with lymphotropic gamma-herpesviruses and relationship to postweaning multisystemic wasting syndrome. Vet. Microbiol. 2006, 116, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Xiao, Y.; Li, X.; Li, S.; Xie, N.; Yan, X.; Li, X.; Zhu, J. Development and application of a quadruplex real-time PCR assay for differential detection of porcine circoviruses (PCV1 to PCV4) in Jiangsu province of China from 2016 to 2020. Transbound. Emerg. Dis. 2021, 68, 1615–1624. [Google Scholar] [CrossRef]
- Palinski, R.; Piñeyro, P.; Shang, P.; Yuan, F.; Guo, R.; Fang, Y.; Byers, E.; Hause, B.M. A Novel Porcine Circovirus Distantly Related to Known Circoviruses Is Associated with Porcine Dermatitis and Nephropathy Syndrome and Reproductive Failure. J. Virol. 2017, 91, e01879. [Google Scholar] [CrossRef]
- Opriessnig, T.; Shen, H.G.; Pal, N.; Ramamoorthy, S.; Huang, Y.W.; Lager, K.M.; Beach, N.M.; Halbur, P.G.; Meng, X. A Live-Attenuated Chimeric Porcine Circovirus Type 2 (PCV2) Vaccine Is Transmitted to Contact Pigs but Is Not Upregulated by Concurrent Infection with Porcine Parvovirus (PPV) and Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) and Is Efficacious in a PCV2b-PRRSV-PPV Challenge Model. Clin. Vaccine Immunol. 2011, 18, 1261–1268. [Google Scholar]
- Duvigneau, J.; Hartl, R.; Groiss, S.; Gemeiner, M. Quantitative simultaneous multiplex real-time PCR for the detection of porcine cytokines. J. Immunol. Methods. 2005, 306, 16–27. [Google Scholar] [CrossRef]
- Kaulitz, D.; Mihica, D.; Adlhoch, C.; Semaan, M.; Denner, J. Improved pig donor screening including newly identified variants of porcine endogenous retrovirus-C (PERV-C). Arch. Virol. 2013, 158, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.C.; Quinn, G.; Suling, K.M.; Oldmixon, B.A.; Van Tine, B.A.; Cina, R.; Arn, S.; Huang, C.A.; Scobie, L.; Onions, D.E.; et al. Identification of Exogenous Forms of Human-Tropic Porcine Endogenous Retrovirus in Miniature Swine. J. Virol. 2004, 78, 2494–2501. [Google Scholar] [CrossRef] [PubMed]
- Halecker, S.; Hansen, S.; Krabben, L.; Ebner, F.; Kaufer, B.; Denner, J. How, where and when to screen for porcine cytomegalovirus (PCMV) in donor pigs for xenotransplantation. Sci. Rep. 2022, 12, 21545. [Google Scholar] [CrossRef]
- Chelli, E.; Suffredini, E.; De Santis, P.; De Medici, D.; Di Bella, S.; D’Amato, S.; Gucciardi, F.; Guercio, A.; Ostanello, F.; Perrone, V.; Purpari, G.; Scavia, G.S.; Schembri, P.; Varcasia, B.M.; Di Bartolo, I. Hepatitis E Virus Occurrence in Pigs Slaughtered in Italy. Animals 2021, 11, 277. [Google Scholar] [CrossRef] [PubMed]
- Sooryanarain, H.; Heffron, C.L.; Hill, D.E.; Fredericks, J.; Rosenthal, B.M.; Werre, S.R.; Opriessnig, T.; Meng, X.J. Hepatitis E Virus in Pigs from Slaughterhouses, United States, 2017-2019. Emerg Infect Dis. 2020, 26, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Boxman, I.L.A.; Verhoef, L.; Dop, P.Y.; Vennema, H.; Dirks, R.A.M.; Opsteegh, M. Int J Food Microbiol. 2022, 379, 109830. [CrossRef]
- Jallow, M.M.; Barry, M.A.; Fall, A.; Ndiaye, N.K.; Kiori, D.; Sy, S.; Goudiaby, D.; Niang, M.N.; Fall, G.; Fall, M.; Dia, N. Influenza A Virus in Pigs in Senegal and Risk Assessment of Avian Influenza Virus (AIV) Emergence and Transmission to Human. Microorganisms 2023, 11, 1961. [Google Scholar] [CrossRef] [PubMed]
- Elbers, A.R.; Tielen, M.J.; Cromwijk, W.A.; Hunneman, W.A. Sero-epidemiological screening of pig sera collected at the slaughterhouse to detect herds infected with Aujeszky’s disease virus, porcine influenza virus and Actinobacillus (Haemophilus) pleuropneumoniae in the framework of an integrated quality control (IQC) system. Vet Q. 1990, 12, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Magar, R.; Larochelle, R. Evaluation of the presence of porcine reproductive and respiratory syndrome virus in pig meat and experimental transmission following oral exposure. Can J Vet Res. 2004, 68, 259–266. [Google Scholar]
- Fischer, N.; Gulich, B.; Keßler, B.; Längin, M.; Fishman, J.A.; Wolf, E.; Boller, K.; Tönjes, R.R.; Godehardt, A.W. PCR and peptide based PCMV detection in pig - development and application of a combined testing procedure differentiating newly from latent infected pigs. Xenotransplantation. 2023, 30, e12803. [Google Scholar] [CrossRef]
- Denner, J.; Jhelum, H.; Hansen, S.; Kaufer, B.B. Comparison of methods for the detection of porcine cytomegalovirus/roseolovirus in relation to biosafety monitoring of xenotransplantation products. Xenotransplantation. 2023, 13, e12835. [Google Scholar] [CrossRef]
- Plowright, W.; Edington, N.; Watt, R.G. The behaviour of porcine cytomegalovirus in commercial pig herds. J Hyg (Lond). 1976, 76, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Edington, N. Porcine cytomegalovirus. In Diseases of Swine (eds Straw, B.E. et al.) (Iowa State University Press, 1999).
- Egerer, S.; Fiebig, U.; Kessler, B.; Zakhartchenko, V.; Kurome, M.; Reichart, B.; Kupatt, C.; Klymiuk, N.; Wolf, E.; Denner, J.; Bähr, A. Early weaning completely eliminates porcine cytomegalovirus from a newly established pig donor facility for xenotransplantation. Xenotransplantation. 2018, 25, e12449. [Google Scholar] [CrossRef] [PubMed]
- Mueller, N.J.; Sulling, K.; Gollackner, B.; Yamamoto, S.; Knosalla, C.; Wilkinson, R.A.; Kaur, A.; Sachs, D.H.; Yamada, K.; Cooper, D.K.; Patience, C.; Fishman, J.A. Reduced efficacy of ganciclovir against porcine and baboon cytomegalovirus in pig-to-baboon xenotransplantation. Am J Transplant. 2003, 3, 1057–1064. [Google Scholar] [CrossRef]
- Fryer, J.F.; Griffiths, P.D.; Emery, V.C.; Clark, D.A. Susceptibility of porcine cytomegalovirus to antiviral drugs. J Antimicrob Chemother. 2004, 53, 975–980. [Google Scholar] [CrossRef]
- Denner, J. First transplantation of a pig heart from a multiple gene-modified donor, porcine cytomegalovirus/roseolovirus, and antiviral drugs. Xenotransplantation. 2023, e12800. [Google Scholar] [CrossRef]
- Denner, J. Porcine Lymphotropic Herpesviruses (PLHVs) and Xenotranplantation. Viruses. 2021, 13, 1072. [Google Scholar] [CrossRef]
- Mueller, N.J.; Kuwaki, K.; Knosalla, C.; Dor, F.J.; Gollackner, B.; Wilkinson, R.A.; Arn, S.; Sachs, D.H.; Cooper, D.K.; Fishman, J.A. Early weaning of piglets fails to exclude porcine lymphotropic herpesvirus. Xenotransplantation 2005, 12, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Tucker, A.; McNeill, F.; Meehan, B.; Galbraith, D.; McArdle, P.; Allan, G.; Patience, C. Methods for the exclusion of circoviruses and gammaherpesviruses from pigs. Xenotransplantation 2003, 10, 343–348. [Google Scholar] [CrossRef]
- Denner, J.; Mankertz, A. Porcine Circoviruses and Xenotransplantation. Viruses. 2017, 9, 83. [Google Scholar] [CrossRef]
- Opriessnig, T.; Xiao, C.T.; Mueller, N.J.; Denner, J. Emergence of novel circoviruses in humans and pigs and their possible importance for xenotransplantation and blood transfusions. Xenotransplantation. 2024, 31, e12842. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Wu, J.; Jiang, M.; Xie, Y.; Bu, W.; Liu, C.; Zhang, G.; Luo, M. A Novel Technique for Constructing Infectious Cloning of Type 3 Porcine Circovirus. Front Microbiol. 2020, 11, 1067. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Li, Y.; Zhang, X.; He, J.; Ma, H. Prevalence of Porcine Circoviruses in Slaughterhouses in Central Shanxi Province, China. Front Vet Sci. 2022, 9, 820914. [Google Scholar] [CrossRef] [PubMed]
- Krüger, L.; Längin, M.; Reichart, B.; Fiebig, U.; Kristiansen, Y.; Prinz, C.; Kessler, B.; Egerer, S.; Wolf, E.; Abicht, J.M.; Denner, J. Transmission of Porcine Circovirus 3 (PCV3) by xenotransplantation of pig hearts into baboons. Viruses. 2019, 11, 650. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Meng, X.J. Hepatitis E virus: Host tropism and zoonotic infection. Curr Opin Microbiol. 2021, 59, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. Xenotransplantation and Hepatitis E virus. Xenotransplantation. 2015, 22, 167–173. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Patience, C.; Magre, S.; Weiss, R.A.; Banerjee, P.T.; Le Tissier, P.; Stoye, J.P. Host range and interference studies of three classes of pig endogenous retrovirus. J Virol. 1998, 72, 9986–9991. [Google Scholar] [CrossRef] [PubMed]
- Denner, J.; Schuurman, H.J. High Prevalence of Recombinant Porcine Endogenous Retroviruses (PERV-A/Cs) in Minipigs: A Review on Origin and Presence. Viruses. 2021, 13, 1869. [Google Scholar] [CrossRef]
- Denner, J. Xenotransplantation - A special case of One Health. One Health. 2017, 3, 17–22. [Google Scholar] [CrossRef]
Figure 1.
Western blot analysis of German slautherhouse pigs. A, schematic presentation of the glycoprotein gB of PCMV/PRV and localisation of the sequence corresponding to the recombinant protein R2 used as antigen (nucleotides according to Acc. No:AF268039) [19], B, result of the Western blot of 10 animals using R2, PC, positive control, serum from a PCMV/PRV-positive pig, NC, serum from a PCMV/PRV-negative pig.
Figure 1.
Western blot analysis of German slautherhouse pigs. A, schematic presentation of the glycoprotein gB of PCMV/PRV and localisation of the sequence corresponding to the recombinant protein R2 used as antigen (nucleotides according to Acc. No:AF268039) [19], B, result of the Western blot of 10 animals using R2, PC, positive control, serum from a PCMV/PRV-positive pig, NC, serum from a PCMV/PRV-negative pig.
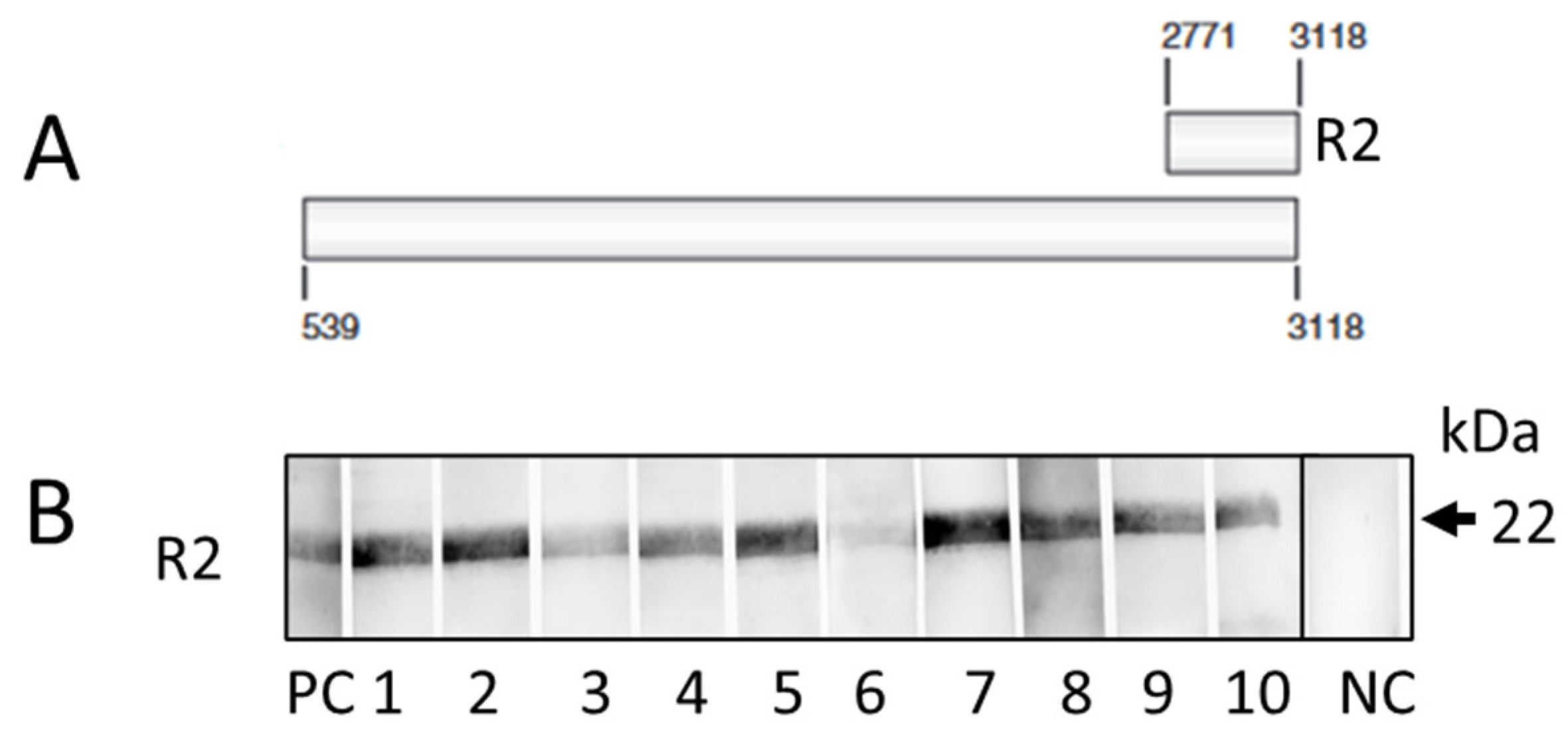

Table 1.
Sex and weight of the tested slaughterhouse pigs and results of the Western blot assay for antibodies against PCMV/PRV.
Table 1.
Sex and weight of the tested slaughterhouse pigs and results of the Western blot assay for antibodies against PCMV/PRV.
| Animal | Sex |
Weight (kg)* |
PCMV/PRV |
| Western blot | |||
| 1 | male | 110.9 | + |
| 2 | male | 108.1 | + |
| 3 | male | 97.6 | + |
| 4 | female | 97.6 | + |
| 5 | female | 99.1 | + |
| 6 | male | 93.8 | + |
| 7 | female | 108.2 | + |
| 8 | male | 109.3 | + |
| 9 | female | 95.6 | + |
| 10 | female | 94.7 | + |
| Total | 5 and 5 | 10/10 |
*after bleeding out; +, positive PCR result; -, negative PCR result,.
Table 2.
Oligonucleotides for the primers and probes used in this study.
| Virus | Primer/Probe | Sequence 5′-3′ | Reference |
| HEV3 | JVHEV3-Fwd | GGT GGT TTC TGG GGT GAC | Jothikumar et al. 2006 [37] |
| JVHEV3-Rev | AGG GGT TGG TTG GAT GAA | ||
| JVHEV3-Probe | 6FAM-TGA TTC TCA GCC CTT CGC-BHQ | ||
| PCMV/PRV | PCMV-Fwd | ACT TCG TCG CAG CTC ATC TGA | Mueller et al. 2002 [38] |
| PCMV-Rev | GTT CTG GGA TTC CGA GGT TG | ||
| PCMV-Probe | 6FAM-CAG GGC GGC GGT CGA GCT C-BHQ | ||
| PLHV-1 | PLHV-1 (1125)-Fwd | CTC ACC TCC AAA TAC AGC GA | Chmielewicz et al. 2003 [39] |
| PLHV-1 (1125)-Rev | GCT TGA ATC GTG TGT TCC ATA G | ||
| PLHV-1 (1125)-Probe | 6FAM-CTG GTC TAC TGA ATC GCC GCT AAC AG-TAMR | ||
| PLHV-2 | PLHV-2 (1155)-Fwd | GTC ACC TGC AAA TAC ACA GG | Chmielewicz et al. 2003 [39] |
| PLHV-2 (1155)-Rev | GGC TTG AAT CGT ATG TTC CAT AT | ||
| PLHV-2 (1155)-Probe | 6FAM-CTG GTC TAC TGA AGC GCT GCC AAT AG-TAMRA | ||
| PLHV-3 | PLHV-3 (210s)-Fwd | AAC AGC GCC AGA AAA AAA GG | McMahon et al. 2006 [40] |
| PLHV-3 (210as)-Rev | GGA AAG GTA GAA GGT GAA CCA TAA AA | ||
| PLHV-3 (210)-Probe | 6-FAM CCA AAG AGG AAA ATC-MGB | ||
| PCV2 | PCV2 (F2020)-Fwd | CTG AGT CTT TTT TAT CAC TTC GTA ATG GT | Chen et al. 2021 [41] |
| PCV2 (F2020)-Rev | ACT GCG TTC GAA AAC AGT ATA TAC GA | ||
| PCV2 (F2020)-Probe | 6FAM-TTA AGT GGG GGG TCT TTA AGA TTA AAT TCT CTG AAT TGT-BHQ2 | ||
| PCV3 | PCV3-Fwd | AGT GCT CCC CAT TGA ACG | Palinski et al. 2017 [42] |
| PCV3-Rev | ACA CAG CCG TTA CTT CAC | ||
| PCV3-Probe | 6FAM-ACC CCA TGG CTC AAC ACA TAT GAC C-BHQ1 | ||
| PCV4 | PCV4 (F2020)-Fwd | ATT ATT AAA CAG ACT TTA TTT GTG TCA TCA CTT | Chen et al. 2021 [41] |
| PCV4 (F2020)-Rev | ACA GGG ATA ATG CGT AGT GAT CAC T | ||
| PCV4 (F2020)-Probe | 6FAM-ATA CTA CAC TTG ATC TTA GCC AAA AGG CTC GTT GA-BHQ1 | ||
| PPV1 | PPV1-Fwd | CAG AAT CAG CAA CCT CAC CA | Opriessnig et al. 2011 [43] |
| PPV1-Rev | GCT GCT GGT GTG TAT GGA AG | ||
| PPV1-Probe | 6FAM-TGC AAG CTT/ZEN/AAT GGT CGC ACT AGA CA-BHQ1 | ||
| pGAPDH | pGAPDH-Fwd | ACA TGG CCT CCA AGG AGT AAG A | Duvigneau et al. 2005 [44] |
| pGAPDH-Rev | GAT CGA GTT GGG GCT GTG ACT | ||
| pGAPDH-Probe | HEX-CCA CCA ACC CCA GCA AGA G-BHQ1 | ||
| PERV-C, PCR1 | PERV-envC-Fwd | GAT TAG AAC TGG AAG CCC CAA GTG CTC T | Kaulitz et al., 2013 [45] |
| PERV-envC-Rev | TCT GAT CCA GAA GTT ATG TTA GAG GAT GGT | ||
| PERV-C, PCR4 | envC.2 for | GATTAGAACTGGAAGCCCCAAGTGCTCT | |
| envC.2 rev | TCTGATCCAGAAGTTATGTTAGAGGATGGT | ||
| PERV-A/C | PERV-A env VRBF-Fwd | CCT ACC AGT TAT AAT CAA TTT AAT TAT GGC | Wood et al. 2004 [46] |
| PERV-C env TMR-Rev | CTC AAA CCA CCC TTG AGT AGT TTC C | ||
Fwd = forward primer, Rev = reverse primer.
Table 3.
Screening for pig viruses in spleen and liver of German slaughterhouse pigs (mean ct values).
Table 3.
Screening for pig viruses in spleen and liver of German slaughterhouse pigs (mean ct values).
| SPLEEN | |||||||||||||
| Animal |
PCMV /PRV |
PLHV-1 | PLHV-2 | PLHV-3 | PPV-1 | PCV2 | PCV3 | PCV4 | PERV-C | PERV-C | PERV-A/C | HEV | |
| Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | PCR1 | PCR4 | PCR | Real-time RT-PCR | |
| 1 | 29.61 | 31.97 | n.d. | 27.41 | n.d. | n.d. | n.d. | n.d. | 24.57 | + | + | - | n.d. |
| 2 | 31.01 | 31.15 | n.d. | 28.24 | n.d. | n.d. | n.d. | n.d. | 21.67 | + | + | - | n.d. |
| 3 | 30.37 | 32.26 | n.d. | 27.77 | n.d. | n.d. | n.d. | n.d. | 26.43 | - | - | - | n.d. |
| 4 | n.d. | 31.21 | n.d. | 29.07 | n.d. | n.d. | 20.09 | n.d. | 25.11 | + | + | - | n.d. |
| 5 | 32.64 | 29.01 | n.d. | 35.62 | n.d. | n.d. | n.d. | n.d. | 25.78 | + | + | - | n.d. |
| 6 | n.d. | 31.75 | n.d. | 35.23 | n.d. | n.d. | n.d. | n.d. | 26.36 | + | + | - | n.d. |
| 7 | 34.10 | 28.56 | n.d. | 31.67 | n.d. | n.d. | n.d. | n.d. | 25.78 | + | + | - | n.d. |
| 8 | n.d. | 30.07 | n.d. | 28.96 | n.d. | n.d. | n.d. | n.d. | 23.49 | + | + | - | n.d. |
| 9 | n.d. | 33.95 | n.d. | 32.73 | n.d. | n.d. | n.d. | n.d. | 29.40 | + | + | - | n.d. |
| 10 | 36.49 | 32.57 | n.d. | 33.81 | n.d. | n.d. | n.d. | n.d. | 27.12 | + | + | - | n.d. |
| Total | 6/10 | 10/10 | 0/10 | 10/10 | 0/10 | 0/10 | 1/10 | 0/10 | 10/10 | 9/10 | 9/10 | 0/10 | 0/10 |
| LIVER | |||||||||||||
| Animal |
PCMV /PRV |
PLHV-1 | PLHV-2 | PLHV-3 | PPV-1 | PCV2 | PCV3 | PCV4 | PERVC | PERV-C | PERV-A/C | HEV | |
| Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | PCR1 | PCR4 | PCR | Real-time RT-PCR | |
| 1 | 36.19 | 31.35 | n.d. | 31.75 | n.d. | n.d. | n.d. | n.d. | 28.66 | + | + | - | n.d. |
| 2 | n.d. | 31.68 | n.d. | 29.91 | n.d. | n.d. | n.d. | n.d. | 22.00 | + | + | - | n.d. |
| 3 | n.d. | 30.79 | n.d. | 33.00 | n.d. | n.d. | n.d. | n.d. | 31.05 | - | - | - | n.d. |
| 4 | n.d. | 30.12 | n.d. | 32.21 | n.d. | n.d. | 25.00 | n.d. | 26.46 | + | + | - | n.d. |
| 5 | n.d. | 27.64 | n.d. | 35.47 | n.d. | n.d. | n.d. | n.d. | 28.90 | + | + | - | n.d. |
| 6 | n.d. | 29.89 | n.d. | 34.92 | n.d. | n.d. | n.d. | n.d. | 26.19 | + | + | - | n.d. |
| 7 | n.d. | 27.46 | n.d. | 34.21 | n.d. | n.d. | n.d. | n.d. | 27.42 | + | + | - | n.d. |
| 8 | n.d. | 29.48 | n.d. | 31.81 | n.d. | n.d. | n.d. | n.d. | 23.05 | + | + | - | n.d. |
| 9 | n.d. | 34.84 | n.d. | 37.02 | n.d. | n.d. | n.d. | n.d. | 31.68 | + | + | - | n.d. |
| 10 | n.d. | 30.68 | n.d. | 35.50 | n.d. | n.d. | n.d. | n.d. | 33.57 | + | + | - | n.d. |
| Total | 1/10 | 10/10 | 010 | 10/10 | 0/10 | 0/10 | 1/10 | 0/10 | 10/10 | 9/10 | 9/10 | o/10 | 0/10 |
n.d., not detected; ct values are shown.
Table 4.
Summary of virus testing of different pig breeds.
| Pig breed |
Virus Detection method |
PCMV/PRV | PLHV-1 | PLHV-2 | PLHV-3 | PPV-1 | PCV1 | PCV2 | PCV3 | PCV4 | PERV-C | PERV-A/C | HEV3 | Reference | ||
|
Facility/ Institution |
Real-time PCR | Western blot |
Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR | Real-time PCR, PCR | PCR | Real-time RT-PCR | Western blot | ||
| Göttingen minipigs | Ellegaard Göttingen Minipigs A/S, Denmark | 12/39 (30%) | 8/67 (12%) | 0/14 (0%) | n.t. | n.t. | n.t. | n.t. | 3/21 (14%) | 0/10 (0%) | n.t. | 28/28 (100%) | 3/13 (23%) |
9/40 (22.5%) | 2/22 (9%) | Morozov et al. [29,30], Plotzki et al. [19], Heinze et al. [31], |
| Göttingen minipigs | University Göttingen, Göttingen, Germany | 0/10 (0%) | n.t. | 2/11 (18%) | 2/11 (18%) | 2/11 (18%) | n.t. | n.t. | 2/10 (20%) | 0/10 (0%) | n.t. | 0/10 (0%) | 0/10 (0%) |
0/10 (0%) | n.t. | Krüger et al. [24] |
| Göttingen minipigs with dippity pig syndrome | Ellegaard Göttingen Minipigs A/S, Denmark, Marshall BioResources, North Rose, New York |
3/7 (42%) |
n.t, | 0/7 (0%) |
0/7 (0%) |
n.t. | n.t. | 3/7 (42%) |
0/7 (0%) |
2/7 (29%) |
0/7 (0%) |
7/7 (100%) |
0/1 (0%) |
0/1 (9%) |
n.t. | Jhelum et al. [35] |
| Aachen minipigs | Aachen Minipig, Heinsberg, Germany |
5/18 (28%) |
n.t. | 0/18 (0%) |
5/18 (28%) |
3/18 (16%) |
n.t. | n.t. | 6/10 (60%) |
n.t. | n.t. | 13/13 (100%) | 2/8 (25%) |
12/18 (67%) |
4/18 (22%) |
Plotzki et al. [33] |
| Mini LEWE | University of Veterinary Medicine Hannover, Germany | 0/10 (0%) |
n.t. | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) | 10/10 (110%) |
0/10 (0%) |
0/10 (0%) | n.t. | Halecker et al. [34] |
| Indigenous Greek black pigs | Four farms in Greece | 16/21 (76%) |
11/11 (100%) |
12/21 (57%) |
15/21 (71%) |
21/21 (100%) |
0/21 (0%) |
n.t. | 21/21 (100%) |
6/21 (29%) |
0/21 (0%) |
11/21 (52%) |
0/21 (0%) |
0/21 (0%) |
n.t. | Jhelum et al. [28] |
| Greek pigs with erythema multiforme | Farm in Greece | 0/5 (0%) |
n.t. | 5/5 (100%) |
1/5 (20%) |
4/5 (80%) |
n.t. | 0/5 (0%) |
1/5 (20%) |
1/5 (20%) |
0/5 (0%) |
5/5 (100%) |
0/5 (0%) |
n.t. | n.t. | Halecker et al. [36] |
| German slaughterhouse pigs | Slaughterhouse near Berlin, Germany |
6/10 (60%) |
10/10 (100%) |
10/10 (100%) |
0/10 (0%) |
10/10 (100%) |
0/10 (0%) |
n.t. | 0/10 (0%) |
1/10 (10%) |
0/10 (0%) |
10/10 (100%) |
0/10 (0%) |
0/10 (0%) |
n.t. | This manuscript |
n.t., not tested; light brown, virus present in some or all animals tested; light green, virus absent in all animals tested.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



