
Submitted:
26 June 2024
Posted:
28 June 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
A severe consequence of SARS-CoV-2 infection that manifests as systemic inflammation and multi-organ involvement is called Multisystem Inflammatory Syndrome in Children (MIS-C). This review examines the possible relationship between gut barrier integrity, the microbiome, dysregulation of interleukin-6 (IL-6) signaling, and MIS-C. Clinical and biochemical features of MIS-C are similar to those of other hyperinflammatory syndromes, suggesting a dysregulated immune response. One possible explanation for the systemic inflammation seen in MIS-C patients is SARS-CoV-2-induced dysregulation of the IL-6 signaling pathway. In addition, new data suggest a reciprocal link between gut barrier integrity and IL-6. SARS-CoV-2 exhibits bacteriophage-like behavior, highlighting the role of bacteria as a reservoir for the virus and emphasizing the importance of understanding the bacteriophagic mechanism of the virus in fecal-oral transmission. Increased translocation of viral products and bacterial toxins may result from disrupting the intestinal barrier and cause systemic inflammation. On the other hand, systemic inflammation can weaken the integrity of the intestinal barrier, which feeds back into the loop of immunological dysregulation. In the context of MIS-C, understanding the interaction between SARS-CoV-2 infection, IL-6, and gut barrier integrity may shed light on the etiology of the disease and guide treatment options.
Keywords:
MIS-C
; IL-6
; gut barrier dysfunction
; zonulin levels
; microbiome
; SARS-CoV-2 bacteriophage behavior
; Long COVID-19
1. Introduction
COVID-19 is not as severe in children as it is in adults in most situations. Children are the target of around 18% of coronavirus infections overall and 1% of COVID-19 cases. The 5-15 age group has a higher incidence of COVID-19. Children in the United States have a seropositivity rate of approximately 75%, which is greater than that of adults. Fortunately, the majority of pediatric patients (>90%) with SARS-CoV-2 infection are asymptomatic or only show moderate signs such as weakness, dry cough, and low fever. In 2021, only 2% of all infected children in the United States required intensive care unit (ICU) care. 27% had moderate symptoms similar to influenza, 5% had substantial pneumonic symptoms, and 66% of all infected children exhibited no symptoms at all [1]. SARS-CoV-2 caused about 48 out of every 100,000 children under the age of 18 to be hospitalized. The prevalence is highest in children aged 0–4 years (66.8 per 100,000), next in children aged 12–17 years (59.9 per 100,000), and lowest in children aged 5–11 years (25 per 100,000 cases). As of February 2021, the number of child deaths from COVID-19 had dropped to 0.17 per 100,000 persons in seven countries—the US, the UK, Italy, Germany, Spain, and France. Among children under 18, COVID-19 was responsible for only 0.48% of the total predicted deaths from all causes in an average year [2]. Similar findings have been reported in other works [3-9].
Several investigations have shown that with the appearance of new less pathogenic variants, the percentage of children and adolescents with COVID-19 who needed to be admitted to the ICU fell sharply from the beginning of the pandemic across all age categories, even though community infection rates eventually increased [10,11]. Wang et al. [11] found that in children under five years old, Omicron infection was associated with a decreased probability of hospitalization, intensive care support, and hospitalizations than Delta infection in the United States. 1.7% of children and adolescents with COVID-19 were admitted to ICU by the Omicron wave in England. All children and adolescents who died within 28 days of being hospitalized for COVID-19 had medical comorbidities, as did the majority of those who needed to be admitted to the ICU [12,13].
Numerous theories have been presented to explain why COVID-19 is less severe in infants. Thymic involution is one plausible cause. A group of researchers has demonstrated that the frequency of COVID-19 hospital admissions in several nations regularly doubles every 16 years of age. They concluded that, in contrast to T-cell production, the probability of hospitalization for COVID-19 increased exponentially with age [14]. Moreover, men are more likely than women to be hospitalized with COVID-19 [15-17]. Men generate fewer T cells and have a higher rate of thymic involution than women. Consequently, thymic involution may be partially responsible for the hospitalization rate associated with COVID-19. However, the researchers found that a further 49-75% of the under-20s are protected from serious disease by other factors. Thymic involution is therefore one, but not the only, explanation for the increasing hospitalization rate with age [14].
To evaluate the immunological responses of children and adults, asymptomatic individuals, those with moderate disease, and individuals who experienced severe COVID-19, Banoun [18] conducted a review study. The majority of adults have antibodies (and T cells) that cross-react with SARS-CoV-2 and other human coronaviruses such as common colds. In adults, however, this response is not related to the severity of the disease; rather, there appears to be a qualitative rather than a quantitative difference (i.e., children have IgM and anti-S, whereas adults have IgG, IgA, and anti-N). Adults who are mildly affected share some of the child's cross-reactivity. The immaturity of the T cells involved is where the differences between children and adults lie at the cellular level [18]. Salivary and mucosal IgA levels are positively associated with the absence of SARS-CoV-2 infection and negatively correlated with age, and are distinct and more potent than serum IgA [19]. In children without symptoms, the innate cellular response in the upper and lower airways is more efficient; the interferon response occurs earlier and involves immune cells instead of epithelial cells, which are linked to hyper-inflammation [20,21]. Given that SARS-CoV-2 has the capacity to inhibit interferon 1 responses, this initial reaction is essential [21].
Regulatory T cells (Tregs), which play an important role in keeping the inflammatory response under control, are found in the respiratory tissues of children. Studies have shown that the lungs and other mucosal and non-lymphoid tissues in children have the highest levels of Tregs [22]. Adult lung tissue has a much lower frequency of Tregs among all CD4+ T cells, at about 5%, compared to pediatric lung tissue, which has a frequency of 15%. The increased concentration of Tregs in a child's respiratory tissues is probably caused by the respiratory system's persistent exposure to inhaled pathogens and antigens, which requires a careful balance between tissue maintenance and immunological responses [22].
When comparing mildly affected children and adults to those severely affected with COVID-19, there is a qualitative difference in the response of neutrophils and macrophages/monocytes, which are also accountable for hyper-inflammation: in severe cases, there is an increase in the number of classical monocytes and dysfunctional neutrophils [20,23].
Banoun [18] also reiterated the importance of the gut microbiome (GM) on the severity of COVID-19, as GM and respiratory infections are strongly linked and may influence the host's response to pneumonia. The synthesis of short-chain fatty acids (SCFAs), the control of systemic inflammation, the establishment of oral immunological tolerance via Tregs, and the management of extra-intestinal T cell populations are some of the potential pathways [24]. Therefore, this paper hypothesizes that gut dysbiosis may also influence the pathogenesis of the multisystem inflammatory response syndrome in children (MIS-C) and clinical outcomes in affected children, and presents evidence to support this proposal.
2. Long COVID-19 and (MIS-C) are Different
Long COVID-19 and MIS-C are two distinct disorders caused by the SARS-CoV-2 virus [25]. Long COVID-19, sometimes called Post-Acute Sequelae of SARS-CoV-2 (PASC), is a syndrome in which people diagnosed with COVID-19 have a variety of symptoms that affect their daily activities, including physical, mental, emotional, and behavioral problems that persist long after the initial infection [26]. These symptoms, which include cardiovascular symptoms (heart palpitations, for example), breathing difficulties, and extreme fatigue, can last for months or even years [26]. On the other hand, MIS-C is an uncommon illness that may occur in children infected with SARS-CoV-2. It involves inflammation in various organs, including the brain, skin, eyes, heart, lungs, kidneys, and the gastrointestinal tract. Although it can be life-threatening, the majority of children recover with medical attention [25,27,28].
3. The GM Supports Barrier Protection Functionality
The pathophysiology of numerous inflammatory and immunological disorders is intimately linked to the integrity of the intestinal barrier [29-31], a dynamic structure that interacts with and responds to a range of stimuli. It is composed of surface mucus, the epithelial layer, and immunological defenses [30]. The mucosal and epithelial components of the physical barrier are closely associated with several cellular junctions, such as adherens junctions (AJ), tight junctions (TJs), and desmosomes [32]. The intestinal epithelium serves its main function as a barrier limiting the interaction between luminal elements (e.g., gut bacteria), the core immune system, and other parts of the organism [33]. Additionally, the normal GM constitutes the majority of the biological barrier, which also controls the intestinal micro-ecological equilibrium [34]. Increased epithelial permeability and possibly microbial dysbiosis lead to a disruption of the intestinal barrier, resulting in a leaky gut. This can allow antigens, bacteria, and toxins from the lumen to enter the circulation and cause a variety of systemic side effects, such as oxidative stress, increased inflammation, and reduced insulin sensitivity (i.e. insulin resistance) [29,35,36].
The GM plays a critical role in maintaining the integrity of the intestinal barrier through several mechanisms, including the production of metabolites that directly regulate the barrier's permeability and the modulation of the immune system's response to the microbiome [37,38]. SCFAs, a type of microbial metabolite, have been shown to improve intestinal barrier function by stimulating the synthesis of TJ proteins, which are critical for preserving the integrity of the epithelial layer. In addition, SCFAs contain anti-inflammatory properties that can help reduce the permeability of the barrier and prevent the entry of hazardous chemicals and pathogens [37].
4. The Link between Gut Microbiome Dysbiosis and MIS-C
Constant fever, vomiting, diarrhea, skin irritation, abdominal pain, and, in severe cases, hypotension and shock are the hallmarks of MIS-C [25-27] One of the most common MIS-C symptoms is gastrointestinal distress, raising the possibility that the GM may act as both a local and systemic inflammatory modulator [39,40].
The human gastrointestinal tract, particularly the colon, is home to trillions of microorganisms, that contain hundreds of times more genes in them than there are in the human genome [41,42]. In this sense, the human body can be thought of as a supra (or meta)-organism. The composition and activity of the GM are co-evolved with the host. It is influenced throughout life, from birth to old age, by a dynamic and complex interaction between the host genome and lifestyle variables, particularly nutrition, which is becoming recognized as the primary modulator of microbial activity [43,44].
This microbial population produces vitamins and ferments macromolecules like proteins, lipids, and carbohydrates. These metabolic effects are collectively known as co-metabolism [45]. Furthermore, the GM is primarily responsible for maintaining host homeostasis through the "training" of host immunity and structural/protective actions against commensal pathogens, specifically safeguarding the intestinal barrier [45]. Eubiosis is maintained by the GM when it is healthy. In pathological circumstances, the GM experiences an unbalanced state of dysbiosis wherein there is either a decrease in beneficial commensals, a proliferation of opportunistic pathogens, or both [46].
The intestinal barrier typically blocks substances and microbes from moving from the lumen to the circulation. However, intestinal dysbiosis, or dysregulation of the gut flora, may result in a disorder called "leaky gut syndrome", characterized by increased permeability that may trigger the innate immune system and promote low-grade inflammation. In recent times, GM dysbiosis has been linked to extra-intestinal as well as intestinal diseases. These include chronic diseases that are particularly prevalent in the elderly, such as diabetes and its severe vascular consequences [47-49]. In adults, it is increasingly recognized that the gut acts as a reservoir for SARS-CoV-2 [50], and that dysbiosis and GI barrier disruption induce inflammatory activation in severe COVID-19 [51,52].
Numerous investigations have revealed that GM is also affected in adult patients with long-term COVID. One study found that GM dysbiosis continued for up to 30 days after disease resolution, which may be related to persistent symptoms of PASC [53]. It is known that angiotensin-converting enzyme 2 (ACE2) regulates the synthesis of neutral amino acid transporters in the gut [54], which regulate the composition of the GM and, ultimately, the immune responses in the body [55,56]. In COVID-19 patients who already had age-related disorders, ACE2 imbalance has been associated with poor outcomes (including increased disease severity and mortality rate) through its impact on intestinal dysbiosis [57].
In a 6-month follow-up, Liu et al. [58] confirmed that long-term COVID patients had different GM species than controls and a consistently decreased diversity. Remarkably, the dysbiosis pattern was different among those who initially had COVID-19 but did not have long-term COVID-19. In the long-COVID-19 subgroup, tiredness, respiratory, and neuropsychiatric problems were strongly correlated with rising fecal relative abundance of opportunistic microorganisms [58]. Additional research revealed that long COVID-19 is associated with dysbiosis of the GM in patients who have recovered a year after being discharged, suggesting that the GM may be crucial in long COVID-19 [59].
Although the aforementioned findings explicitly demonstrated that alterations in the microbiome have a deleterious effect on the extended clinical course of COVID-19 in adults, there has been a lack of research on this crucial topic in MIS-C. The composition, diversity, and abundance of the gut microbiome were found to be different in MIS-C cases compared to COVID-19 cases and healthy controls. At the phylum level, the MIS-C group had an abundance of Bacteroidetes, whereas the healthy children had Firmicutes. When MIS-C was compared to the SARS-CoV-2 group and the healthy control group, the relative abundance of Bacteroidetes increased, and the Firmicutes: Bacteroides ratio significantly dropped [60]. Romani et al. [61] investigated the GM of children with COVID-19, including four cases of MIS-C. Veillonella, Ruminococcus, Clostridium, Dialister, and Streptococcus were more prevalent in the GM of MIS-C patients, while Bifidobacterium, Blautia, Granulicatella, and Prevotella were less prevalent.
Pro-inflammatory taxa were more prevalent and anti-inflammatory taxa were less prevalent in children with MIS-C. Children with MIS-C and COVID-19 [60] had lower concentrations of Faecalibacterium prausnitzii, which has been demonstrated to use butyrate to reduce intestinal mucosal inflammation and maintain gut physiology [62,63]. Therefore, it has been proposed that "alterations of the intestinal microbiome may contribute to the pathophysiology of MIS-C predisposing factors" [60].
4.1. SARS-CoV-2 Infection Impairs Gut Barrier Integrity by Inducing Zonulin Release
For successful maintenance of intestinal homeostasis, mucosal integrity and management of intestinal permeability are essential [64]. From a structural perspective, the gut barrier can be defined as a working structure composed of mechanical components like mucus and the layer of epithelial cells. Intestinal bacteria are secluded from the underlying lamina propria by the dynamic physical and biochemical barrier known as the intestinal mucosa, which also inhibits the infiltration of pathogenic and antigenic molecules [65]. Despite the presence of chemical and physical barriers in the intestines, some pathogens can cross through it and interact with intestinal epithelial cells (IECs). Several sensing pattern recognition receptors (PRRs) are expressed by IECs, which are essential for maintaining the intestinal mucosal immune response's homeostasis [66].
In response to bacterial signals, PRRs can initiate a physiological inflammatory response that promotes progenitor cell proliferation, epithelial cell survival, and pathogen elimination—all of which enhance intestinal integrity [67]. Crucially, intercellular junctional complexes consisting of desmosomes, TJs, AJs, and gap junctions (GJ) connect IECs. Together, these complexes establish intercellular communication, control paracellular permeability, and allow passive entry of water, ions, and nutrients. Maintaining intracellular adhesion helps to create a semi-permeable protective barrier that prevents pathogenic and commensal microbes and antigens from penetrating the lamina propria [68,69].
As was already mentioned, children with MIS-C frequently experience GI symptoms, which can lead to a severe hyper-inflammatory reaction and cardiac problems. A SARS-CoV-2 infection is known to cause MIS-C several weeks later, and it is known that the viral load in respiratory secretions decreases for 7–10 days following infection [70-72]. It seems doubtful that the initial respiratory tract infection in children with MIS-C is related to the disease, as the majority of them have negative nasopharyngeal viral swabs [70]. Furthermore, several lines of evidence demonstrated that SARS-CoV-2 can establish robust infection and replication in human IECs, which may contribute to GI dysfunction and potential fecal-oral transmission in some patients [73-75]. Patients with SARS-CoV-2 infection had elevated zonulin levels, which were linked to worse outcomes [40,76-79]. Furthermore, high serum zonulin levels disrupt the brain-blood barrier (BBB), allowing viruses to enter the brain and induce serious neurological symptoms [40,80]. Numerous autoimmune and hyper-inflammatory illnesses, including celiac disease [81], inflammatory bowel disease [82], and Kawasaki disease [83], have been linked to elevated circulating zonulin levels and increased intestinal permeability [82,84].
In 2021, Yonker et al. [40] examined bio specimens from 100 children: 19 had acute COVID-19, 26 had MIS-C, and 55 were controls. Reverse transcription PCR (RT-PCR) was used to detect SARS-CoV-2 in stool samples, and zonulin and other indicators of mucosal barrier integrity were tested in plasma. Zonulin is a member of a structurally and functionally similar family of proteins that modulate intercellular TJss to reversibly regulate intestinal permeability [85-87]. Large antigens, such as viral antigens generated from SARS-CoV-2 present in the GI tract, should not be able to pass through the gut lumen and into the bloodstream when the intestinal mucosal barrier is intact and functioning [50]. According to this seminal research, the prolonged presence of undigested SARS-CoV-2 viruses causes the gastrointestinal tract to release more zonulin, which increases intestinal permeability [40].
Elevated zonulin levels suggest that intestinal epithelial TJss are failing, possibly allowing SARS-CoV-2 antigens to penetrate the blood. Several events, including gut dysbiosis [88], trigger the release of zonulin in a myeloid differentiation primary response 88 (MyD88)-dependent manner, which enables zonulin to bind to its target, the protease-activated receptor 2 (PAR2), and subsequently transactivate the epidermal growth factor receptor (EGFR) [89]. This initiates a chain of events that results in the phosphorylation of TJ proteins, such as myosin 1c and zonula occludens 1 (ZO1), which in turn leads to the disintegration of TJs and an increase in the permeability of paracellular membranes to macromolecules [90]. This, in turn, accelerates the trafficking of viral antigens into the bloodstream (Figure 1), resulting in hyper-inflammation [40].
The development of zonulin-dependent loss of gut integrity in infants affected with MIS-C, but not COVID-19, suggests that a chronic form of SARS-CoV-2-induced dysbiosis in the gut leads to a progressive deterioration of mucosal barrier integrity. Viremia was not found in MIS-C, although antigenemia and viremia have been demonstrated to correlate with severe acute COVID-19 in adults [72,91].
Yonker et al. [40] found SARS-CoV-2 cleaved soluble component of spike ( S1), and nucleocapsid antigens in the blood of children suffering from MIS-C, even though the children had been exposed to or had been infected with the virus weeks before. Compared to MIS-C patients and healthy controls (p < 0.0001), and children with acute COVID-19 (p < 0.001), SARS-CoV-2 spike protein levels were significantly higher. They also found that patients with MIS-C had significantly higher levels of SARS-CoV-2 S1 protein than healthy controls (p = 0.004) and children with acute COVID-19 (p = 0.02). Compared with healthy controls, the researchers found no discernible increase in blood levels of SARS-CoV-2 antigen in children with acute COVID-19 [40].
4.2. Bacteriophage-Like Behavior of SARS-CoV-2 and MISC
In addition to SARS-CoV-2-mediated damage to the integrity of the intestinal barrier induced by zonulin release, other important research performed by Brogna and colleagues revealed that this virus is capable of infecting intestinal bacteria, acting like a bacteriophage [92-96]. According to a previous study by that group, the blood, feces, and urine of COVID-19 patients contained toxin-like peptides that were almost identical to the poisonous elements of animal venoms, including conotoxin, phospholipases A2, phosphodiesterases, zinc metal proteinases, and bradykinins [97]. Later research revealed that SARS-CoV-2 might replicate autonomously in bacterial cultures derived from patient feces for up to 30 days and longer [98]. Viral-like structures ranging in size from 25 to 100 nm were observed by electron microscopy interacting with and within bacterial cell walls (Figure 2). The presence of SARS-CoV-2 nucleocapsid protein both inside and outside of the bacteria was verified by immunolabeling; specifically two anti-inflammatory gut bacterial species normally present in a healthy human GM (Faecalibacterium prausnitzii and Dorea formicigenerans) [94]. These species were previously found to be considerably reduced in severe COVID-19 cases [53,99] and in children with MIS-C [60]. Importantly, both bacteria have been shown to use butyrate, thereby preserving gut physiology and reducing gut mucosal inflammation [62,63].
In addition, de novo synthesis of SARS-CoV-2 spike protein was detected in the bacterial cultures using 15N-labelled nitrogen as a source, accompanied by an increase in viral RNA load [94]. The discovery that SARS-CoV-2 can infect bacteria cells that are a component of the normal human GM and synthesize both nucleic acid and viral peptides has major consequences for human health, especially concerning the regulation and control of the virus spread. For example, the ability of SARS-CoV-2 to infect and replicate in human gut bacteria may lead to a scenario in which infected bacteria continue to harbor the virus for prolonged periods, possibly even after the systemic infection of the human host has resolved [94]. Epidemiological studies have shown that this may result in prolonged viral shedding through the intestinal tract, potentially increasing the likelihood of subsequent transmission. Furthermore, prolonged survival due to continuous replication of the virus in the intestinal lumen could allow recurrent revival of systemic SARS-CoV-2 infection in the person, leading to COVID-19 relapse without the need for external reinfection [94].
The combined deleterious effects of SARS-CoV-2 infection reflected as increased zonulin release [40] and the decrease in commensal bacterial populations due to the bacteriophage-like effect of SARS-CoV-2 [92-96] could further compromise the integrity of the intestinal barrier, allowing the passage of the spike protein and toxins released by infected bacteria.
5. The Link between IL-6 Levels, Gut Barrier Integrity, and MIS-C
During the COVID-19 pandemic, several meta-analyses demonstrated that high levels of IL-6 have been linked to some adverse health effects, including acute respiratory distress syndrome, death, and ICU hospitalization. Serum IL-6 levels were almost three times greater in patients with such complex types of COVID-19 than in patients with less severe disease [100-104]. However, extensive research allows us to understand that, depending on the specific circumstances of the immune response, IL-6 is a cytokine with multiple functions, having both pro- and anti-inflammatory effects [105]. Initially discovered as B-cell stimulatory factor 2 (BSF-2), IL-6 stimulates activated B cells to produce immunoglobulin (Ig) [106]. At low, physiological concentrations, it regulates many important immune functions. Innate and adaptive immune systems are largely developed and activated in large part by IL-6. This cytokine induces monocytes to differentiate into macrophages rather than dendritic cells (DCs) in the innate immune system [107].
Moreover, the increase of anti-apoptotic proteins that support T cell survival is associated with IL-6 signaling [108-110]. Furthermore, IL-6 inhibits transforming growth factor beta (TGF-β)-mediated maturation of Tregs and promotes the development of untrained CD4+ T cells into effector T cell subgroups, such as pathogen-specific effector Th17 cells [111,112]. Therefore, an increase in IL-6 levels causes immune dysregulation, as in COVID-19´s cytokine storm, and also in other autoimmune disorders and cancer [111,113-118]. IL-6 is also essential for epithelial proliferation and wound repair [119] and for immune-epithelial-bacteria communication in both beneficial as well as harmful processes. For example, Pediococcus acidilactici K15, a lactic acid bacterial strain, promotes the secretion of IL-6 in BDCA1+ DCs (mDC1) via its double-stranded RNA and enhances the synthesis of protective IgA antibodies [120].
Conversely, high levels of IL-6 can cause inflammation, and many probiotics work to lower these levels to treat intestinal disease [121]. In the context of ulcerative colitis (UC), a study found that at physiological levels, IL-6 controls epithelial barrier function by modulating the expression of TJs-related proteins. In contrast, IL-6 levels in the plasma of UC patients were elevated and increased as the disease worsened. Overproduction of IL-6 has also been shown to damage the intestinal epithelial cell barrier and control barrier function by increasing zonulin release. Conversely, when an anti-IL-6 antibody was added, the amount of zonulin was lower than in the control group [122].
It was discovered that long-term SARS-CoV-2 infection in the GI tract triggered zonulin release in children with MIS-C, with subsequent trafficking of SARS-CoV-2 antigens into the circulation, resulting in hyper-inflammation and signs of a cytokine storm, including significantly higher levels of TNF-α, IL-6, IL-10, and IL-1β [40]. Scientists demonstrated that SARS-CoV-2 RNA is still present in the GI tract for weeks after initial infection and viral antigenemia is correlated with zonulin-induced increased permeability (leaky gut) of the mucosal barrier [40].
Thus, it is likely that the high zonulin levels found in children with MIS-C [40] are due to excessive concentrations of IL-6 released by SARS-CoV-2 infected cells at the GI. In addition, several works have demonstrated that in MIS-C, impairment of the innate and adaptive immune responses is responsible for the prolonged presence of the virus in the GI tract [123-128].
6. Preventive and Therapeutic Strategies for MIS-C
Yonker et al. [40] used larazotide, a zonulin antagonist, to treat children with MIS-C and evaluated the impact on antigenemia and the children's clinical outcomes. After receiving larazotide treatment for MIS-C, the patient's plasma SARS-CoV-2 spike antigen levels and inflammatory markers decreased concurrently, leading to a greater clinical improvement than what was currently possible with known treatments. In adults with COVID-19 and long COVID, Brogna et al. [129] discovered that patients who started early antibiotic treatment showed a statistically significant decrease in recovery time, and such treatment was critical for maintaining high blood oxygen saturation levels. Delayed antibiotic initiation within the first 3 days increased the risk of pneumonia in both vaccinated and unvaccinated patients.
Furthermore, it is noteworthy that a considerable proportion of patients who were administered antibiotics within the initial three days and throughout the full seven days of the acute phase did not experience long COVID. One of the main contributing factors to the development of the disease appears to be the bacteriophage behavior of SARS-CoV-2 during the acute and post-COVID-19 phases. Early antibiotic treatment appears to be essential for halting the progression of disease, controlling toxin release from infected bacteria, and preventing viral replication in the GM [129].
Another potential approach could involve employing anti-IL-6 antibodies such as tocilizumab or sarilumab. However, it has been shown that IL-6 suppression in COVID-19 patients affects the neutralizing capacity of anti-SARS CoV-2 antibodies [130]. Considering that the neutralizing activity of anti-SARS-CoV-2 antibodies determines protection against symptomatic infection [131], the study conducted by Della-Torre calls for a rigorous reevaluation of the risk of reinfection and severe illness in patients receiving anti-IL-6 antibodies [130].
Certain Gram-negative bacterial strains, such as Escherichia coli, Prevotella, Pseudomonas, and Salmonella spp., have been found to induce intestinal zonulin release, while other strains, primarily Gram-positive ones, like Bifidobacterium and Lactobacillus spp., have been found to decrease zonulin levels. These findings are consistent with previous research conducted on cell lines and animal models (reviewed in [132]).Probiotics are live microorganisms that, when administered in the right quantities and for the right length of time, benefit the health of the host [133]. Through their surface molecules and metabolites, probiotics and intestinal symbionts can alter the host's intestinal barrier function [134]. Several studies have shown that probiotics reduce both intestinal permeability and epithelial barrier dysfunction in gastrointestinal disorders, thereby demonstrating the role of GM in improving intestinal barrier function and protection (Figure 3) against pathogens [135-137].
At present, there is no doubt about the link between GM and the regulation of zonulin release, as some probiotic strains have been shown to improve gut barrier function by affecting the expression of zonulin and TJ proteins [132,138-140]. Probiotics dramatically enhanced gut barrier functioning, according to a meta-analysis of data from a total of 26 randomized controlled trials (n = 1891). Specifically, the trans-epithelial resistance (TER) was significantly enhanced, while serum zonulin, endotoxin, and lipopolysaccharide levels were importantly reduced. Moreover, probiotic groups outperformed control groups in lowering inflammatory markers like IL-6, tumor necrosis factor-α (TNF-α), and C reactive protein. Additionally, probiotics can regulate the composition of the GM by increasing the enrichment of Lactobacillus and Bifidobacterium [137].
Oral probiotic therapy has also been utilized in pediatric and adult populations to reduce the incidence and severity of respiratory infections. In this regard, it is interesting to mention that in 95% of MIS-C patients, hypovitaminosis D was found [141]. The severity of MIS-C and 25-hydroxyvitamin D [(25-(OH) D] levels were found to be significantly correlated; children with more severe MIS-C also had lower 25(OH) D levels. Echocardiograms showed worse left ventricular systolic and diastolic performance in MIS-C patients with decreased vitamin D levels [141]. The effects of vitamin D on the immune system and inflammatory response have been the subject of extensive investigation in recent years. Vitamin D affects immunological response as well as the proliferation and differentiation of immune cells through binding to vitamin D receptors [142-145]. Vitamin D additionally controls the proliferation and antibody synthesis in B lymphocytes [146]. It has been established that immunological illnesses and disorders are more likely to occur in people with low serum levels of 25(OH) D [147], and some scientists proposed that MIS-C may be the cause of hypovitaminosis D [148].
There is mounting evidence that vitamin D levels are related to the composition of the GM [149]. A systematic analysis identified 25 research papers that investigated the relationship between vitamin D and GM; it was found that supplementing with vitamin D caused a notable increase in the abundance of Firmicutes and Bifidobacterium, and Firmicutes were found to be correlated with serum Vitamin D levels [150]. Vitamin D insufficiency affects over 80% of people in certain geographic regions and has been connected to inflammation and gut dysbiosis [151-153]. In a study of 80 vitamin D-deficient women, its administration (after 8 weeks) markedly increased GM diversity. More precisely, the abundance of the probiotic taxa Bifidobacterium and Akkermansi increased in parallel with the Bacteroidetes to Firmicutes ratio [149].
By strengthening the gut-lung axis and controlling the host inflammatory response, probiotics can elicit antiviral effects [154]. In a pediatric experiment (n = 31 prebiotics, 31 probiotics, and 32 placebos), preterm infants were given a prebiotic combination of galacto-oligosaccharide and polydextrose or the probiotic Lactobacillus rhamnosus GG mixed with breast milk throughout the first 60 days of life. There were fewer cases of virus-associated respiratory tract infections in the probiotic and prebiotic groups (p = 0.022 and p < 0.001, respectively) [155]. Probiotics may lessen symptoms of upper respiratory tract infections and stabilize GM diversity, according to a study that supplied Lab4P probiotics (comprising lactobacilli and bifidobacteria) daily to 220 overweight and obese adults [154,156]. This may be especially important for COVID-19 infections, where obesity is linked to worse outcomes [157,158].
Research into the treatment of COVID-19 in children using oral microbial interventions is scarce, despite its potential [39,159]. Probiotics have only been tested in a small number of adult patients with COVID-19, but the results were encouraging, showing reductions in viral load, hospitalization duration, death, and diarrhea frequency [160-164]. Adults with moderate-to-severe COVID-19 were given a supplementary oral dose of the Bifidobacterium animalis sp. Lactis strain (n = 20 probiotic, 24 non-probiotic). The probiotic group experienced a five-day reduction in hospital stay (p < 0.001) and a corresponding decrease in IL-6 levels (p < 0.001) [162].
A single-center, quadruple-blinded, randomized trial was carried out by other researchers on adult outpatients with symptomatic Covid-19. For 30 days, subjects were randomly assigned to either a probiotic formula (including Lactiplantibacillus plantarum KABP022, KABP023, and KAPB033, as well as Pediococcus acidilactici KABP021) or a placebo [161]. Of the 147 patients, 78 (53.1%) in the probiotic group experienced complete remission, while 41 (28.1%) in the placebo group did. There were no hospitalizations or deaths during the research, and the nasopharyngeal viral load, lung infiltrates, and duration of both digestive and non-digestive symptoms were all reduced when compared to placebo. There were no significant differences in fecal microbiome composition between probiotic and placebo groups, although probiotic treatment boosted specific IgM and IgG antibodies against SARS-CoV2 when compared to placebo. Therefore, rather than altering the diversity of the colonic microbiome, it is assumed that probiotics predominantly enhance the host´s immune system [161]. They promote the synthesis of immunoglobulins, specifically IgA and type I interferons, and boost the induction of interleukins and the activation of macrophages, natural killer cells, and T-helper cells [165,166].
Probiotic supplementation may therefore help to restore gut health by enhancing immune responses, reducing inflammation, and reinforcing the epithelial barrier, thus possibly decreasing susceptibility to severe SARS-CoV-2 infection. It is suggested that probiotics should be used as a prophylactic rather than a treatment for severe cases of MIS-C, as once a cytokine storm is activated it is very difficult to control.
7. Conclusions
Research has shown that viral infections of the respiratory tract, such as H1N1 influenza A virus [167] and respiratory syncytial virus [168], can affect GM in children. Although the complex relationship between invasive viruses and host physiology is not yet fully understood, increasing data suggests that the microbiome may influence the course of viral diseases [169]. The GM diversity has been suggested as a predictive biomarker of COVID-19. This diversity is characterized by a lower ratio of Firmicutes to Bacteroidetes, a higher abundance of Proteobacteria, and a lower abundance of beneficial butyrate-producing bacteria [170], and it has been proposed that the microbiome of children may have a high level of protective resilience due to characteristics such as the comparatively high abundance of Bifidobacterium [39]. Research suggests that changes in the composition of the gut microbiome may play a role in the production of inflammatory cytokines induced by SARS-CoV-2, which could trigger the cytokine storm [171].
Lower levels of Bifidobacterium have been linked to the severity of SARS-CoV-2 infection [53,171-175]. Prebiotics are frequently used to enhance the effects of probiotics [176], which have been successfully tested and hypothesized to improve SARS-CoV-2 symptoms [160,161,177-179].
Through their metabolic byproducts and end products, Bifidobacterium spp. symbiotically feed other gut microorganisms, which in turn improve butyrate synthesis and decrease inflammation [180]. The anti-inflammatory effects of Bifidobacterium may be one of the methods by which it increases "natural immunity”," hence defending against the consequences of SARS-CoV-2 infection [181]. It has been demonstrated that Bifidobacterium binds TNF-α [182,183]. According to Cervantes and Hong [184], this binding could absorb TNF-α from the gut, which will then decrease it in the bloodstream and eventually adsorb it from the lungs and other affected tissues (the "gut-lung axis"). Moreover, Bifidobacterium species modulate the helper T cell response and raise levels of the anti-inflammatory cytokine interleukin IL-10. Furthermore, Bifidobacterium stimulates the Th1 immune response while suppressing the Th2 immunological response [185].
Regarding the pathogenic role of SARS-CoV-2, Brogna et al. [92,94,96] discovered that this virus, by exhibiting a bacteriophage-like behavior, infects and destroys two important anti-inflammatory bacteria (Faecalibacterium prausnitzii and Dorea formicigenerans) that are normally present in a healthy gut. Interestingly, both species are butyrate-producing bacteria. Butyrate is an important short-chain fatty acid that contributes to gut health by serving as an energy source for the body, stimulating immunological responses, having anti-inflammatory effects, and supporting colon cell function [62,186,187]. Previously, the prevalence of these species was found to be much lower in sever COVID-19 cases [53,99], in post-acute COVID-19 syndrome (PACS) [58,188], and in children with MIS-C [60]. Such a reduction in the number of these bacteria could be due to the destruction caused by the lytic phase of SARS-CoV-2 [92]. A relevant study demonstrated that fecal transplant from PACS patients into germ-free animals resulted in lung inflammation and worse outcomes following pulmonary infection with multidrug-resistant Klebsiella pneumoniae. Transplanted mice also performed poorly in cognitive tests. These findings showed that altered GM may significantly contribute to PACS [189].
In addition to the destruction of beneficial bacteria by SARS-COV-2 [92,94,96], this virus also induces an excessive IL-6-mediated zonulin release that increases intestinal permeability (leaky gut), so the spike protein and toxins released by the bacteria pass into the bloodstream causing MIS-C [40]. Finally, there is evidence that if damage to the mucosal epithelium is prevented and treated early in the disease, MIS-C may not develop [40]. Given that gut dysbiosis may predispose children to MIS-C, it is important to strengthen their microbiome by consuming probiotics, particularly butyrate-producing Bifidobacterium, and diets high in plant fiber (prebiotics).
Author Contributions
Conceptualization, E.M.R., A.R.-C.; formal analysis, A.Z., E.M.R., M.R., D.C., A.H.J., V.N.U., M.F., C.B., M.P., A.R.-C.; investigation, A.Z., E.M.R., M.R., D.C., A.H.J., V.N.U., M.F., C.B., M.P., A.R.-C.; data curation, A.Z., E.M.R., M.R., D.C., A.H.J., V.N.U., M.F., C.B., M.P., A.R.-C.; writing—original draft preparation, A.Z., E.M.R., M.R., D.C., A.H.J., V.N.U., M.F., C.B., M.P., A.R.-C.; writing—review and editing, A.Z., E.M.R., M.R., D.C., A.H.J., V.N.U., M.F., C.B., M.P., A.R.-C.; visualization, A.Z., A.R.-C.; supervision, E.M.R., A.R.-C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Forrest, C.B.; Burrows, E.K.; Mejias, A.; Razzaghi, H.; Christakis, D.; Jhaveri, R.; Lee, G.M.; Pajor, N.M.; Rao, S.; Thacker, D. Severity of acute COVID-19 in children< 18 years old March 2020 to December 2021. Pediatrics 2022, 149, e2021055765. [Google Scholar]
- Bhopal, S.S.; Bagaria, J.; Olabi, B.; Bhopal, R. Children and young people remain at low risk of COVID-19 mortality. The Lancet Child & Adolescent Health 2021, 5, e12–e13. [Google Scholar]
- Brodin, P. SARS-CoV-2 infections in children: Understanding diverse outcomes. Immunity 2022, 55, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Curtis, N. Why does the severity of COVID-19 differ with age?: understanding the mechanisms underlying the age gradient in outcome following SARS-CoV-2 infection. The Pediatric Infectious Disease Journal 2022, 41, e36–e45. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. Why is COVID-19 less severe in children? A review of the proposed mechanisms underlying the age-related difference in severity of SARS-CoV-2 infections. Archives of disease in childhood 2021, 106, 429–439. [Google Scholar] [CrossRef]
- Bundle, N.; Dave, N.; Pharris, A.; Spiteri, G.; Deogan, C.; Suk, J.E. COVID-19 trends and severity among symptomatic children aged 0–17 years in 10 European Union countries, 3 August 2020 to 3 October 2021. Eurosurveillance 2021, 26, 2101098. [Google Scholar] [CrossRef] [PubMed]
- Thors, V.; Bjornsdottir, K.L.; Love, T.; Haraldsson, A. SARS-CoV-2 infections in icelandic children: close follow-up of all confirmed cases in a nationwide study. The Pediatric infectious disease journal 2022, 41, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Gu, J.; Chen, Q.; Deng, N.; Li, J.; Huang, L.; Zhou, X. Clinical and epidemiological characteristics of pediatric SARS-CoV-2 infections in China: A multicenter case series. PLoS medicine 2020, 17, e1003130. [Google Scholar] [CrossRef]
- Say, D.; Crawford, N.; McNab, S.; Wurzel, D.; Steer, A.; Tosif, S. Post-acute COVID-19 outcomes in children with mild and asymptomatic disease. The Lancet Child & Adolescent Health 2021, 5, e22–e23. [Google Scholar]
- Cloete, J.; Kruger, A.; Masha, M.; du Plessis, N.M.; Mawela, D.; Tshukudu, M.; Manyane, T.; Komane, L.; Venter, M.; Jassat, W. Paediatric hospitalisations due to COVID-19 during the first SARS-CoV-2 omicron (B. 1.1. 529) variant wave in South Africa: a multicentre observational study. The Lancet Child & Adolescent Health 2022, 6, 294–302. [Google Scholar]
- Wang, L.; Berger, N.A.; Kaelber, D.C.; Davis, P.B.; Volkow, N.D.; Xu, R. Incidence rates and clinical outcomes of SARS-CoV-2 infection with the omicron and delta variants in children younger than 5 years in the US. JAMA pediatrics 2022, 176, 811–813. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.L.; Harwood, R.; Kenny, S.; Cruz, J.; Clark, M.; Davis, P.J.; Draper, E.S.; Hargreaves, D.; Ladhani, S.N.; Gent, N. Pediatric hospitalizations and ICU admissions due to COVID-19 and pediatric inflammatory multisystem syndrome temporally associated with SARS-CoV-2 in England. JAMA pediatrics 2023, 177, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Ungar, S.P.; Solomon, S.; Stachel, A.; Shust, G.F.; Clouser, K.N.; Bhavsar, S.M.; Lighter, J. Hospital and ICU admission risk associated with comorbidities among children with COVID-19 ancestral strains. Clinical Pediatrics 2023, 62, 1048–1058. [Google Scholar] [CrossRef] [PubMed]
- Palmer, S.; Cunniffe, N.; Donnelly, R. COVID-19 hospitalization rates rise exponentially with age, inversely proportional to thymic T-cell production. Journal of the Royal Society Interface 2021, 18, 20200982. [Google Scholar] [CrossRef]
- Gomez, J.M.D.; Du-Fay-de-Lavallaz, J.M.; Fugar, S.; Sarau, A.; Simmons, J.A.; Clark, B.; Sanghani, R.M.; Aggarwal, N.T.; Williams, K.A.; Doukky, R. Sex differences in COVID-19 hospitalization and mortality. Journal of Women's health 2021, 30, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Biolè, C.; Bianco, M.; Núñez-Gil, I.J.; Cerrato, E.; Spirito, A.; Roubin, S.R.; Viana-Llamas, M.C.; Gonzalez, A.; Castro-Mejía, A.F.; Eid, C.M. Gender Differences in the Presentation and Outcomes of Hospitalized Patients With COVID-19. Journal of Hospital Medicine 2021, 16, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Chinn, J.; De Ferrante, M.; Kirby, K.A.; Hohmann, S.F.; Amin, A. Male gender is a predictor of higher mortality in hospitalized adults with COVID-19. PloS one 2021, 16, e0254066. [Google Scholar] [CrossRef] [PubMed]
- Banoun, H. Why are children and many adults not affected by COVID-19? Role of the host immune response. Infectious Diseases Research 2022, 3, 18. [Google Scholar] [CrossRef]
- Tsukinoki, K.; Yamamoto, T.; Saito, J.; Sakaguchi, W.; Iguchi, K.; Inoue, Y.; Ishii, S.; Sato, C.; Yokoyama, M.; Shiraishi, Y. Prevalence of saliva immunoglobulin A antibodies reactive with severe acute respiratory syndrome coronavirus 2 among Japanese people unexposed to the virus. Microbiology and Immunology 2022, 66, 403–410. [Google Scholar] [CrossRef]
- Yoshida, M.; Worlock, K.B.; Huang, N.; Lindeboom, R.G.; Butler, C.R.; Kumasaka, N.; Dominguez Conde, C.; Mamanova, L.; Bolt, L.; Richardson, L. Local and systemic responses to SARS-CoV-2 infection in children and adults. Nature 2022, 602, 321–327. [Google Scholar] [CrossRef]
- Vono, M.; Huttner, A.; Lemeille, S.; Martinez-Murillo, P.; Meyer, B.; Baggio, S.; Sharma, S.; Thiriard, A.; Marchant, A.; Godeke, G.-J. Robust innate responses to SARS-CoV-2 in children resolve faster than in adults without compromising adaptive immunity. Cell reports 2021, 37. [Google Scholar] [CrossRef] [PubMed]
- Traxinger, B.R.; Richert-Spuhler, L.E.; Lund, J.M. Mucosal tissue regulatory T cells are integral in balancing immunity and tolerance at portals of antigen entry. Mucosal Immunology 2022, 15, 398–407. [Google Scholar] [CrossRef]
- McKenna, E.; Wubben, R.; Isaza-Correa, J.M.; Melo, A.M.; Mhaonaigh, A.U.; Conlon, N.; O’Donnell, J.S.; Ní Cheallaigh, C.; Hurley, T.; Stevenson, N.J. Neutrophils in COVID-19: not innocent bystanders. Frontiers in Immunology 2022, 13, 864387. [Google Scholar] [CrossRef] [PubMed]
- Chunxi, L.; Haiyue, L.; Yanxia, L.; Jianbing, P.; Jin, S. The gut microbiota and respiratory diseases: new evidence. Journal of immunology research 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Fraser, R.; Orta-Resendiz, A.; Dockrell, D.; Müller-Trutwin, M.; Mazein, A. Severe COVID-19 versus multisystem inflammatory syndrome: comparing two critical outcomes of SARS-CoV-2 infection. European Respiratory Review 2023, 32. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.; Siles, N.; Kelley, M.; Wylie, D.; Melamed, E.; Brode, W.M. Clinical characteristics of Long COVID patients presenting to a dedicated academic post-COVID-19 clinic in Central Texas. Scientific Reports 2023, 13, 21971. [Google Scholar] [CrossRef] [PubMed]
- Constantin, T.; Pék, T.; Horváth, Z.; Garan, D.; Szabó, A.J. Multisystem inflammatory syndrome in children (MIS-C): Implications for long COVID. Inflammopharmacology 2023, 31, 2221–2236. [Google Scholar] [CrossRef] [PubMed]
- Rowley, A.H. Understanding SARS-CoV-2-related multisystem inflammatory syndrome in children. Nature Reviews Immunology 2020, 20, 453–454. [Google Scholar] [CrossRef]
- Wells, J.M.; Brummer, R.J.; Derrien, M.; MacDonald, T.T.; Troost, F.; Cani, P.D.; Theodorou, V.; Dekker, J.; Méheust, A.; De Vos, W.M. Homeostasis of the gut barrier and potential biomarkers. American Journal of Physiology-Gastrointestinal and Liver Physiology 2017, 312, G171–G193. [Google Scholar] [CrossRef]
- Camilleri, M. Leaky gut: mechanisms, measurement and clinical implications in humans. Gut 2019, 68, 1516–1526. [Google Scholar] [CrossRef]
- Binienda, A.; Twardowska, A.; Makaro, A.; Salaga, M. Dietary carbohydrates and lipids in the pathogenesis of leaky gut syndrome: An overview. International journal of molecular sciences 2020, 21, 8368. [Google Scholar] [CrossRef]
- Hiippala, K.; Jouhten, H.; Ronkainen, A.; Hartikainen, A.; Kainulainen, V.; Jalanka, J.; Satokari, R. The potential of gut commensals in reinforcing intestinal barrier function and alleviating inflammation. Nutrients 2018, 10, 988. [Google Scholar] [CrossRef] [PubMed]
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: a therapeutic target? Nature reviews Gastroenterology & hepatology 2017, 14, 9–21. [Google Scholar]
- Li, X.-Y.; He, C.; Zhu, Y.; Lu, N.-H. Role of gut microbiota on intestinal barrier function in acute pancreatitis. World journal of gastroenterology 2020, 26, 2187. [Google Scholar] [CrossRef]
- Mu, Q.; Kirby, J.; Reilly, C.M.; Luo, X.M. Leaky gut as a danger signal for autoimmune diseases. Frontiers in immunology 2017, 8, 269575. [Google Scholar] [CrossRef] [PubMed]
- Régnier, M.; Van Hul, M.; Knauf, C.; Cani, P.D. Gut microbiome, endocrine control of gut barrier function and metabolic diseases. Journal of Endocrinology 2021, 248, R67–R82. [Google Scholar] [CrossRef]
- Ghosh, S.; Whitley, C.S.; Haribabu, B.; Jala, V.R. Regulation of intestinal barrier function by microbial metabolites. Cellular and molecular gastroenterology and hepatology 2021, 11, 1463–1482. [Google Scholar] [CrossRef] [PubMed]
- Di Vincenzo, F.; Del Gaudio, A.; Petito, V.; Lopetuso, L.R.; Scaldaferri, F. Gut microbiota, intestinal permeability, and systemic inflammation: A narrative review. Internal and emergency medicine 2024, 19, 275–293. [Google Scholar] [CrossRef]
- Bacorn, M.; Romero-Soto, H.N.; Levy, S.; Chen, Q.; Hourigan, S.K. The Gut Microbiome of Children during the COVID-19 Pandemic. Microorganisms 2022, 10, 2460. [Google Scholar] [CrossRef]
- Yonker, L.M.; Gilboa, T.; Ogata, A.F.; Senussi, Y.; Lazarovits, R.; Boribong, B.P.; Bartsch, Y.C.; Loiselle, M.; Rivas, M.N.; Porritt, R.A. Multisystem inflammatory syndrome in children is driven by zonulin-dependent loss of gut mucosal barrier. The Journal of Clinical Investigation 2021, 131. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.-X.; Chen, X.-Y.; Wang, J.-Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J. Microbiota in health and diseases. Signal transduction and targeted therapy 2022, 7, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochemical journal 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Barone, M.; D'Amico, F.; Brigidi, P.; Turroni, S. Gut microbiome–micronutrient interaction: The key to controlling the bioavailability of minerals and vitamins? Biofactors 2022, 48, 307–314. [Google Scholar] [CrossRef]
- Corbin, K.D.; Carnero, E.A.; Dirks, B.; Igudesman, D.; Yi, F.; Marcus, A.; Davis, T.L.; Pratley, R.E.; Rittmann, B.E.; Krajmalnik-Brown, R. Host-diet-gut microbiome interactions influence human energy balance: A randomized clinical trial. Nature communications 2023, 14, 3161. [Google Scholar] [CrossRef]
- Krishnan, S.; Alden, N.; Lee, K. Pathways and functions of gut microbiota metabolism impacting host physiology. Current opinion in biotechnology 2015, 36, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.; Kandalgaonkar, M.R.; Golonka, R.M.; Yeoh, B.S.; Vijay-Kumar, M.; Saha, P. Crosstalk between gut microbiota and host immunity: impact on inflammation and immunotherapy. Biomedicines 2023, 11, 294. [Google Scholar] [CrossRef] [PubMed]
- Ascher, S.; Reinhardt, C. The gut microbiota: an emerging risk factor for cardiovascular and cerebrovascular disease. European Journal of Immunology 2018, 48, 564–575. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Clément, K. The gut microbiome, diet, and links to cardiometabolic and chronic disorders. Nature Reviews Nephrology 2016, 12, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.W.; Kitai, T.; Hazen, S.L. Gut microbiota in cardiovascular health and disease. Circulation research 2017, 120, 1183–1196. [Google Scholar] [CrossRef]
- Gaebler, C.; Wang, Z.; Lorenzi, J.C.; Muecksch, F.; Finkin, S.; Tokuyama, M.; Cho, A.; Jankovic, M.; Schaefer-Babajew, D.; Oliveira, T.Y. Evolution of antibody immunity to SARS-CoV-2. Nature 2021, 591, 639–644. [Google Scholar] [CrossRef]
- Giron, L.B.; Dweep, H.; Yin, X.; Wang, H.; Damra, M.; Goldman, A.R.; Gorman, N.; Palmer, C.S.; Tang, H.-Y.; Shaikh, M.W. Plasma markers of disrupted gut permeability in severe COVID-19 patients. Frontiers in immunology 2021, 12, 686240. [Google Scholar] [CrossRef]
- Trottein, F.; Sokol, H. Potential causes and consequences of gastrointestinal disorders during a SARS-CoV-2 infection. Cell reports 2020, 32. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, Y.K.; Zuo, T.; Lui, G.C.-Y.; Zhang, F.; Liu, Q.; Li, A.Y.; Chung, A.C.; Cheung, C.P.; Tso, E.Y.; Fung, K.S. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19. Gut 2021, 70, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Camargo, S.M.; Singer, D.; Makrides, V.; Huggel, K.; Pos, K.M.; Wagner, C.A.; Kuba, K.; Danilczyk, U.; Skovby, F.; Kleta, R. Tissue-specific amino acid transporter partners ACE2 and collectrin differentially interact with hartnup mutations. Gastroenterology 2009, 136, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Perlot, T.; Rehman, A.; Trichereau, J.; Ishiguro, H.; Paolino, M.; Sigl, V.; Hanada, T.; Hanada, R.; Lipinski, S. ACE2 links amino acid malnutrition to microbial ecology and intestinal inflammation. Nature 2012, 487, 477–481. [Google Scholar] [CrossRef]
- Perlot, T.; Penninger, J.M. ACE2–From the renin–angiotensin system to gut microbiota and malnutrition. Microbes and infection 2013, 15, 866–873. [Google Scholar] [CrossRef]
- Viana, S.D.; Nunes, S.; Reis, F. ACE2 imbalance as a key player for the poor outcomes in COVID-19 patients with age-related comorbidities–role of gut microbiota dysbiosis. Ageing research reviews 2020, 62, 101123. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Mak, J.W.Y.; Su, Q.; Yeoh, Y.K.; Lui, G.C.-Y.; Ng, S.S.S.; Zhang, F.; Li, A.Y.; Lu, W.; Hui, D.S.-C. Gut microbiota dynamics in a prospective cohort of patients with post-acute COVID-19 syndrome. Gut 2022, 71, 544–552. [Google Scholar] [PubMed]
- Zhang, D.; Zhou, Y.; Ma, Y.; Chen, P.; Tang, J.; Yang, B.; Li, H.; Liang, M.; Xue, Y.; Liu, Y. Gut microbiota dysbiosis correlates with long COVID-19 at one-year after discharge. Journal of Korean Medical Science 2023, 38. [Google Scholar] [CrossRef]
- Suskun, C.; Kilic, O.; Yilmaz Ciftdogan, D.; Guven, S.; Karbuz, A.; Ozkaya Parlakay, A.; Kara, Y.; Kacmaz, E.; Sahin, A.; Boga, A. Intestinal microbiota composition of children with infection with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and multisystem inflammatory syndrome (MIS-C). European journal of pediatrics 2022, 181, 3175–3191. [Google Scholar] [CrossRef]
- Romani, L.; Del Chierico, F.; Macari, G.; Pane, S.; Ristori, M.V.; Guarrasi, V.; Gardini, S.; Pascucci, G.R.; Cotugno, N.; Perno, C.F. The relationship between pediatric gut microbiota and SARS-CoV-2 infection. Frontiers in cellular and infection microbiology 2022, 12, 908492. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhang, M.; Wang, Y.; Dorfman, R.G.; Liu, H.; Yu, T.; Chen, X.; Tang, D.; Xu, L.; Yin, Y. Faecalibacterium prausnitzii produces butyrate to maintain Th17/Treg balance and to ameliorate colorectal colitis by inhibiting histone deacetylase 1. Inflammatory bowel diseases 2018, 24, 1926–1940. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, B.; Luckey, D.; Taneja, V. Autoimmunity-associated gut commensals modulate gut permeability and immunity in humanized mice. Military medicine 2019, 184, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Konig, J.; Wells, J.; Cani, P.; Garcia-Rodenas, C.; MacDonald, T.; Mercenier, A.; Whyte, J.; Troost, F.; Brummer, R. Human intestinal barrier function in health and disease. Clin Transl Gastroenterol. 2016; 7 (10): e196.
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue barriers 2017, 5, e1373208. [Google Scholar] [CrossRef]
- Santaolalla, R.; Abreu, M.T. Innate immunity in the small intestine. Current opinion in gastroenterology 2012, 28, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.; Trinchieri, G. Innate immune mechanisms of colitis and colitis-associated colorectal cancer. Nature Reviews Immunology 2011, 11, 9–20. [Google Scholar] [CrossRef]
- Marchiando, A.M.; Graham, W.V.; Turner, J.R. Epithelial barriers in homeostasis and disease. Annual Review of Pathology: Mechanisms of Disease 2010, 5, 119–144. [Google Scholar] [CrossRef]
- Laukoetter, M.G.; Bruewer, M.; Nusrat, A. Regulation of the intestinal epithelial barrier by the apical junctional complex. Current opinion in gastroenterology 2006, 22, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Yonker, L.; Neilan, A.; Bartsch, Y. Pediatric SARS-CoV-2: clinical presentation, infectivity, and immune responses [manuscript published online ahead of print 20 August 2020]. J Pediatr 2020, 10. [Google Scholar]
- Wang, Y.; Zhang, L.; Sang, L.; Ye, F.; Ruan, S.; Zhong, B.; Song, T.; Alshukairi, A.N.; Chen, R.; Zhang, Z. Kinetics of viral load and antibody response in relation to COVID-19 severity. The Journal of clinical investigation 2020, 130, 5235–5244. [Google Scholar] [CrossRef]
- Fajnzylber, J.; Regan, J.; Coxen, K.; Corry, H.; Wong, C.; Rosenthal, A.; Worrall, D.; Giguel, F.; Piechocka-Trocha, A.; Atyeo, C. SARS-CoV-2 viral load is associated with increased disease severity and mortality. Nature communications 2020, 11, 5493. [Google Scholar] [CrossRef] [PubMed]
- Minami, S.; Matsumoto, N.; Omori, H.; Nakamura, Y.; Tamiya, S.; Nouda, R.; Nurdin, J.A.; Yamasaki, M.; Kotaki, T.; Kanai, Y. Effective SARS-CoV-2 replication of monolayers of intestinal epithelial cells differentiated from human induced pluripotent stem cells. Scientific Reports 2023, 13, 11610. [Google Scholar] [CrossRef] [PubMed]
- Zang, R.; Castro, M.F.G.; McCune, B.T.; Zeng, Q.; Rothlauf, P.W.; Sonnek, N.M.; Liu, Z.; Brulois, K.F.; Wang, X.; Greenberg, H.B. TMPRSS2 and TMPRSS4 promote SARS-CoV-2 infection of human small intestinal enterocytes. Science immunology 2020, 5, eabc3582. [Google Scholar] [CrossRef]
- Stanifer, M.L.; Kee, C.; Cortese, M.; Zumaran, C.M.; Triana, S.; Mukenhirn, M.; Kraeusslich, H.-G.; Alexandrov, T.; Bartenschlager, R.; Boulant, S. Critical role of type III interferon in controlling SARS-CoV-2 infection in human intestinal epithelial cells. Cell reports 2020, 32. [Google Scholar] [CrossRef]
- Kılıç, A.O.; Akın, F.; Yazar, A.; Metin Akcan, Ö.; Topcu, C.; Aydın, O. Zonulin and claudin-5 levels in multisystem inflammatory syndrome and SARS-CoV-2 infection in children. Journal of Paediatrics and Child Health 2022, 58, 1561–1565. [Google Scholar] [CrossRef] [PubMed]
- Okuyucu, M.; Kehribar, D.Y.; Çapraz, M.; Çapraz, A.; Arslan, M.; Çelik, Z.B.; Usta, B.; Birinci, A.; Ozgen, M.; Özgen, M. The relationship between COVID-19 disease severity and zonulin levels. Cureus 2022, 14. [Google Scholar] [CrossRef]
- Hensley-McBain, T.; Manuzak, J.A. Zonulin as a biomarker and potential therapeutic target in multisystem inflammatory syndrome in children. The Journal of clinical investigation 2021, 131. [Google Scholar] [CrossRef]
- Palomino-Kobayashi, L.A.; Ymaña, B.; Ruiz, J.; Mayanga-Herrera, A.; Ugarte-Gil, M.F.; Pons, M.J. Zonulin, a marker of gut permeability, is associated with mortality in a cohort of hospitalised peruvian COVID-19 patients. Frontiers in cellular and infection microbiology 2022, 12, 1000291. [Google Scholar]
- Llorens, S.; Nava, E.; Muñoz-López, M.; Sánchez-Larsen, Á.; Segura, T. Neurological symptoms of COVID-19: the zonulin hypothesis. Frontiers in Immunology 2021, 12, 665300. [Google Scholar] [CrossRef]
- Drago, S.; El Asmar, R.; Di Pierro, M.; Grazia Clemente, M.; Sapone, A.T.A.; Thakar, M.; Iacono, G.; Carroccio, A.; D'Agate, C.; Not, T. Gliadin, zonulin and gut permeability: Effects on celiac and non-celiac intestinal mucosa and intestinal cell lines. Scandinavian journal of gastroenterology 2006, 41, 408–419. [Google Scholar] [CrossRef]
- Wyatt, J.; Vogelsang, H.; Hübl, W.; Waldhoer, T.; Lochs, H. Intestinal permeability and the prediction of relapse in Crohn's disease. The Lancet 1993, 341, 1437–1439. [Google Scholar] [CrossRef] [PubMed]
- Rivas, M.N.; Wakita, D.; Franklin, M.K.; Carvalho, T.T.; Abolhesn, A.; Gomez, A.C.; Fishbein, M.C.; Chen, S.; Lehman, T.J.; Sato, K. Intestinal permeability and IgA provoke immune vasculitis linked to cardiovascular inflammation. Immunity 2019, 51, 508–521. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A. All disease begins in the (leaky) gut: role of zonulin-mediated gut permeability in the pathogenesis of some chronic inflammatory diseases. F1000Research 2020, 9. [Google Scholar] [CrossRef]
- Fasano, A.; Not, T.; Wang, W.; Uzzau, S.; Berti, I.; Tommasini, A.; Goldblum, S.E. Zonulin, a newly discovered modulator of intestinal permeability, and its expression in coeliac disease. The Lancet 2000, 355, 1518–1519. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A. Regulation of intercellular tight junctions by zonula occludens toxin and its eukaryotic analogue zonulin. Annals of the New York Academy of Sciences 2000, 915, 214–222. [Google Scholar] [CrossRef]
- Wang, W.; Uzzau, S.; Goldblum, S.E.; Fasano, A. Human zonulin, a potential modulator of intestinal tight junctions. Journal of cell science 2000, 113, 4435–4440. [Google Scholar] [CrossRef] [PubMed]
- El Asmar, R.; Panigrahi, P.; Bamford, P.; Berti, I.; Not, T.; Coppa, G.V.; Catassi, C.; Fasano, A. Host-dependent zonulin secretion causes the impairment of the small intestine barrier function after bacterial exposure. Gastroenterology 2002, 123, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.E.; Sapone, A.; Fasano, A.; Vogel, S.N. Gliadin stimulation of murine macrophage inflammatory gene expression and intestinal permeability are MyD88-dependent: role of the innate immune response in Celiac disease. The Journal of Immunology 2006, 176, 2512–2521. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A. Intestinal permeability and its regulation by zonulin: diagnostic and therapeutic implications. Clinical Gastroenterology and Hepatology 2012, 10, 1096–1100. [Google Scholar] [CrossRef]
- Ogata, A.F.; Maley, A.M.; Wu, C.; Gilboa, T.; Norman, M.; Lazarovits, R.; Mao, C.-P.; Newton, G.; Chang, M.; Nguyen, K. Ultra-sensitive serial profiling of SARS-CoV-2 antigens and antibodies in plasma to understand disease progression in COVID-19 patients with severe disease. Clinical chemistry 2020, 66, 1562–1572. [Google Scholar] [CrossRef]
- Brogna, C.; Brogna, B.; Bisaccia, D.R.; Lauritano, F.; Marino, G.; Montano, L.; Cristoni, S.; Prisco, M.; Piscopo, M. Could SARS-CoV-2 have bacteriophage behavior or induce the activity of other bacteriophages? Vaccines 2022, 10, 708. [Google Scholar] [CrossRef] [PubMed]
- Brogna, C.; Viduto, V.; Fabrowski, M.; Cristoni, S.; Marino, G.; Montano, L.; Piscopo, M. The importance of the gut microbiome in the pathogenesis and transmission of SARS-CoV-2: Someone on Earth:“.. we moved at the speed of Science!”-Science from the center of the Universe:“Hey man, I’m still waiting for you in the 50s!”. Gut Microbes 2023, 15, 2244718. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, M.; Querci, M.; Brogna, C.; Ponti, J.; Cristoni, S.; Markov, P.V.; Valsesia, A.; Leoni, G.; Benedetti, A.; Wiss, T. Evidence of SARS-CoV-2 bacteriophage potential in human gut microbiota. F1000Research 2022, 11, 292. [Google Scholar] [CrossRef]
- Brogna, C.; Costanzo, V.; Brogna, B.; Bisaccia, D.R.; Brogna, G.; Giuliano, M.; Montano, L.; Viduto, V.; Cristoni, S.; Fabrowski, M. Analysis of bacteriophage behavior of a human RNA virus, SARS-CoV-2, through the integrated approach of immunofluorescence microscopy, proteomics and D-amino acid quantification. International Journal of Molecular Sciences 2023, 24, 3929. [Google Scholar] [CrossRef]
- Brogna, C.; Cristoni, S.; Petrillo, M.; Bisaccia, D.R.; Lauritano, F.; Montano, L.; Prisco, M.; Piscopo, M. The first report on detecting SARS-CoV-2 inside bacteria of the human gut microbiome: A case series on asymptomatic family members and a child with COVID-19. F1000Research 2022, 11, 135. [Google Scholar] [CrossRef]
- Brogna, C.; Cristoni, S.; Petrillo, M.; Querci, M.; Piazza, O.; Van den Eede, G. Toxin-like peptides in plasma, urine and faecal samples from COVID-19 patients. F1000Research 2021, 10. [Google Scholar] [CrossRef]
- Petrillo, M.; Brogna, C.; Cristoni, S.; Querci, M.; Piazza, O.; Van den Eede, G. Increase of SARS-CoV-2 RNA load in faecal samples prompts for rethinking of SARS-CoV-2 biology and COVID-19 epidemiology. F1000Research 2021, 10. [Google Scholar] [CrossRef]
- Zuo, T.; Zhang, F.; Lui, G.C.; Yeoh, Y.K.; Li, A.Y.; Zhan, H.; Wan, Y.; Chung, A.C.; Cheung, C.P.; Chen, N. Alterations in gut microbiota of patients with COVID-19 during time of hospitalization. Gastroenterology 2020, 159, 944–955. [Google Scholar] [CrossRef] [PubMed]
- Coomes, E.A.; Haghbayan, H. Interleukin-6 in COVID-19: a systematic review and meta-analysis. Reviews in medical virology 2020, 30, 1–9. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, J.; Yang, Y.; Ma, H.; Li, Z.; Zhang, J.; Cheng, J.; Zhang, X.; Zhao, Y.; Xia, Z. The role of interleukin-6 in monitoring severe case of coronavirus disease 2019. EMBO molecular medicine 2020, 12, e12421. [Google Scholar] [CrossRef]
- Zhu, J.; Pang, J.; Ji, P.; Zhong, Z.; Li, H.; Li, B.; Zhang, J. Elevated interleukin-6 is associated with severity of COVID-19: a meta-analysis. Journal of medical virology 2021, 93, 35. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.-X.; Agbana, Y.L.; Sun, Z.-S.; Fei, S.-W.; Zhao, H.-Q.; Zhou, X.-N.; Chen, J.-H.; Kassegne, K. Increased interleukin-6 is associated with long COVID-19: a systematic review and meta-analysis. Infectious Diseases of Poverty 2023, 12, 43. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.; Fatima, R.; Assaly, R. Elevated interleukin-6 and severe COVID-19: a meta-analysis. Journal of medical virology 2020, 92, 2283. [Google Scholar] [CrossRef]
- Aliyu, M.; Zohora, F.T.; Anka, A.U.; Ali, K.; Maleknia, S.; Saffarioun, M.; Azizi, G. Interleukin-6 cytokine: An overview of the immune regulation, immune dysregulation, and therapeutic approach. International Immunopharmacology 2022, 111, 109130. [Google Scholar] [CrossRef]
- Kishimoto, T. The biology of interleukin-6. Blood 1989, 74, 1–10. [Google Scholar] [CrossRef]
- Chomarat, P.; Banchereau, J.; Davoust, J.; Karolina Palucka, A. IL-6 switches the differentiation of monocytes from dendritic cells to macrophages. Nature immunology 2000, 1, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Narimatsu, M.; Maeda, H.; Itoh, S.; Atsumi, T.; Ohtani, T.; Nishida, K.; Itoh, M.; Kamimura, D.; Park, S.-J.; Mizuno, K. Tissue-specific autoregulation of the stat3 gene and its role in interleukin-6-induced survival signals in T cells. Molecular and cellular biology 2001. [Google Scholar] [CrossRef] [PubMed]
- Teague, T.K.; Schaefer, B.C.; Hildeman, D.; Bender, J.; Mitchell, T.; Kappler, J.W.; Marrack, P. Activation-induced inhibition of interleukin 6–mediated T cell survival and signal transducer and activator of transcription 1 signaling. The Journal of experimental medicine 2000, 191, 915–926. [Google Scholar] [CrossRef]
- Curnow, S.J.; Scheel-Toellner, D.; Jenkinson, W.; Raza, K.; Durrani, O.M.; Faint, J.M.; Rauz, S.; Wloka, K.; Pilling, D.; Rose-John, S. Inhibition of T cell apoptosis in the aqueous humor of patients with uveitis by IL-6/soluble IL-6 receptor trans-signaling. The Journal of Immunology 2004, 173, 5290–5297. [Google Scholar] [CrossRef]
- Kimura, A.; Kishimoto, T. IL-6: regulator of Treg/Th17 balance. European journal of immunology 2010, 40, 1830–1835. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. Therapeutic targeting of the interleukin-6 receptor. Annual review of pharmacology and toxicology 2012, 52, 199–219. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F.; Finotto, S. IL-6 signaling in autoimmunity, chronic inflammation and inflammation-associated cancer. Cytokine & growth factor reviews 2011, 22, 83–89. [Google Scholar]
- Aricha, R.; Mizrachi, K.; Fuchs, S.; Souroujon, M.C. Blocking of IL-6 suppresses experimental autoimmune myasthenia gravis. Journal of autoimmunity 2011, 36, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. International immunology 2021, 33, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.E.; Maerz, M.D.; Buckner, J.H. IL-6: a cytokine at the crossroads of autoimmunity. Current opinion in immunology 2018, 55, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Iwakura, T.; Matsui, K.; Kawaguchi, H.; Obana, M.; Hayama, A.; Maeda, M.; Izumi, Y.; Komuro, I.; Ohsugi, Y. IL-6-mediated Th17 differentiation through RORγt is essential for the initiation of experimental autoimmune myocarditis. Cardiovascular research 2011, 91, 640–648. [Google Scholar] [CrossRef]
- Narazaki, M.; Tanaka, T.; Kishimoto, T. The role and therapeutic targeting of IL-6 in rheumatoid arthritis. Expert review of clinical immunology 2017, 13, 535–551. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, K.A.; Manieri, N.A.; Liu, T.-C.; Stappenbeck, T.S. IL-6 stimulates intestinal epithelial proliferation and repair after injury. PloS one 2014, 9, e114195. [Google Scholar] [CrossRef]
- Kawashima, T.; Ikari, N.; Kouchi, T.; Kowatari, Y.; Kubota, Y.; Shimojo, N.; Tsuji, N.M. The molecular mechanism for activating IgA production by Pediococcus acidilactici K15 and the clinical impact in a randomized trial. Scientific reports 2018, 8, 5065. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, B.; Wang, T.; Gao, L.; Yang, Z.-j.; Wang, F.-f.; Shang, H.-w.; Hua, R.; Xu, J.-d. Biological characteristics of IL-6 and related intestinal diseases. International journal of biological sciences 2021, 17, 204. [Google Scholar] [CrossRef]
- Li, Y.; Jia, Y.; Cui, T.; Zhang, J. IL-6/STAT3 signaling pathway regulates the proliferation and damage of intestinal epithelial cells in patients with ulcerative colitis via H3K27ac. Experimental and Therapeutic Medicine 2021, 22, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Isaza-Correa, J.; Ryan, L.; Kelly, L.; Allen, J.; Melo, A.; Jones, J.; Huggard, D.; Ryan, E.; Ó Maoldomhnaigh, C.; Geoghehan, S. Innate immune dysregulation in multisystem inflammatory syndrome in children (MIS-C). Scientific Reports 2023, 13, 16463. [Google Scholar] [CrossRef] [PubMed]
- Ghazizadeh Esslami, G.; Mamishi, S.; Pourakbari, B.; Mahmoudi, S. Systematic review and meta-analysis on the serological, immunological, and cardiac parameters of the multisystem inflammatory syndrome (MIS-C) associated with SARS-CoV-2 infection. Journal of Medical Virology 2023, 95, e28927. [Google Scholar] [CrossRef] [PubMed]
- Rybkina, K.; Bell, J.N.; Bradley, M.C.; Wohlbold, T.; Scafuro, M.; Meng, W.; Korenberg, R.C.; Davis-Porada, J.; Anderson, B.R.; Weller, R.J. SARS-CoV-2 infection and recovery in children: distinct T cell responses in MIS-C compared to COVID-19. Journal of Experimental Medicine 2023, 220, e20221518. [Google Scholar] [CrossRef] [PubMed]
- Gruber, C.N.; Patel, R.S.; Trachtman, R.; Lepow, L.; Amanat, F.; Krammer, F.; Wilson, K.M.; Onel, K.; Geanon, D.; Tuballes, K. Mapping Systemic Inflammation and Antibody Responses in Multisystem Inflammatory Syndrome in Children (MIS-C). Cell 2023, 186, 3325. [Google Scholar] [CrossRef] [PubMed]
- Zerra, P.E.; Stowell, J.; Verkerke, H.; McCoy, J.; Jones, J.; Graciaa, S.; Lu, A.; Hussaini, L.; Anderson, E.J.; Rostad, C.A. Factor H autoantibodies contribute to complement dysregulation in multisystem inflammatory syndrome in children (MIS-C). American journal of hematology 2023, 98, E98–E101. [Google Scholar] [CrossRef]
- Lee, M.J.; Leong, M.W.; Rustagi, A.; Beck, A.; Zeng, L.; Holmes, S.; Qi, L.S.; Blish, C.A. SARS-CoV-2 escapes direct NK cell killing through Nsp1-mediated downregulation of ligands for NKG2D. Cell reports 2022, 41. [Google Scholar] [CrossRef] [PubMed]
- Brogna, C.; Montano, L.; Zanolin, M.E.; Bisaccia, D.R.; Ciammetti, G.; Viduto, V.; Fabrowski, M.; Baig, A.M.; Gerlach, J.; Gennaro, I. A retrospective cohort study on early antibiotic use in vaccinated and unvaccinated COVID-19 patients. Journal of Medical Virology 2024, 96, e29507. [Google Scholar] [CrossRef]
- Della-Torre, E.; Criscuolo, E.; Lanzillotta, M.; Locatelli, M.; Clementi, N.; Mancini, N.; Dagna, L.; Group, C.-B.S. IL-1 and IL-6 inhibition affects the neutralising activity of anti-SARS-CoV-2 antibodies in patients with COVID-19. The Lancet. Rheumatology 2021, 3, e829. [Google Scholar] [CrossRef]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nature medicine 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Veres-Székely, A.; Szász, C.; Pap, D.; Szebeni, B.; Bokrossy, P.; Vannay, Á. Zonulin as a potential therapeutic target in microbiota-gut-brain axis disorders: encouraging results and emerging questions. International Journal of Molecular Sciences 2023, 24, 7548. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature reviews Gastroenterology & hepatology.
- Liu, Q.; Yu, Z.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Surface components and metabolites of probiotics for regulation of intestinal epithelial barrier. Microbial Cell Factories 2020, 19, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Andrew, H.; Kirschner, B.S.; Guandalini, S. Is Lactobacillus GG helpful in children with Crohn’s disease? Results of a preliminary, open-label study. Journal of pediatric gastroenterology and nutrition 2000, 31, 453–457. [Google Scholar]
- Ewaschuk, J.B.; Diaz, H.; Meddings, L.; Diederichs, B.; Dmytrash, A.; Backer, J.; Looijer-van Langen, M.; Madsen, K.L. Secreted bioactive factors from Bifidobacterium infantis enhance epithelial cell barrier function. American journal of physiology-gastrointestinal and liver physiology 2008, 295, G1025–G1034. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, Z.; Tang, P.; Wu, Y.; Zhang, A.; Li, D.; Wang, C.-Z.; Wan, J.-Y.; Yao, H.; Yuan, C.-S. Probiotics fortify intestinal barrier function: a systematic review and meta-analysis of randomized trials. Frontiers in Immunology 2023, 14, 1143548. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, X.; Jiang, Y.; Zhao, W.; Guo, T.; Cao, Y.; Teng, J.; Hao, X.; Zhao, J.; Yang, Z. Antioxidant status and gut microbiota change in an aging mouse model as influenced by exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibetan kefir. Journal of dairy science 2017, 100, 6025–6041. [Google Scholar] [CrossRef] [PubMed]
- Petrof, E.O.; Kojima, K.; Ropeleski, M.J.; Musch, M.W.; Tao, Y.; De Simone, C.; Chang, E.B. Probiotics inhibit nuclear factor-κB and induce heat shock proteins in colonic epithelial cells through proteasome inhibition. Gastroenterology 2004, 127, 1474–1487. [Google Scholar] [CrossRef] [PubMed]
- Martorell, P.; Alvarez, B.; Llopis, S.; Navarro, V.; Ortiz, P.; Gonzalez, N.; Balaguer, F.; Rojas, A.; Chenoll, E.; Ramon, D. Heat-treated Bifidobacterium longum CECT-7347: a whole-cell postbiotic with antioxidant, anti-inflammatory, and gut-barrier protection properties. Antioxidants 2021, 10, 536. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, D.; Benzon, B.; Srsen, S.; Polic, B.; Vukovic Novogradec, A.; Milic, P.; Markic, J. The Impact of Vitamin D Levels on Clinical Manifestations of Multisystem Inflammatory Syndrome in Children: A Cross-Sectional Study. Life 2023, 13, 674. [Google Scholar] [CrossRef]
- Bikle, D. Nonclassic actions of vitamin D. The Journal of Clinical Endocrinology & Metabolism 2009, 94, 26–34. [Google Scholar]
- Cozzolino, M.; Vidal, M.; Arcidiacono, M.V.; Tebas, P.; Yarasheski, K.E.; Dusso, A.S. HIV-protease inhibitors impair vitamin D bioactivation to 1, 25-dihydroxyvitamin D. Aids 2003, 17, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Cantorna, M.T.; Zhu, Y.; Froicu, M.; Wittke, A. Vitamin D status, 1, 25-dihydroxyvitamin D3, and the immune system. The American journal of clinical nutrition 2004, 80, 1717S–1720S. [Google Scholar] [CrossRef]
- Cippitelli, M.; Santoni, A. Vitamin D3: a transcriptional modulator of the interferon-γ gene. European journal of immunology 1998, 28, 3017–3030. [Google Scholar] [CrossRef]
- Chen, S.; Sims, G.P.; Chen, X.X.; Gu, Y.Y.; Chen, S.; Lipsky, P.E. Modulatory effects of 1, 25-dihydroxyvitamin D3 on human B cell differentiation. The Journal of Immunology 2007, 179, 1634–1647. [Google Scholar] [CrossRef] [PubMed]
- Charoenngam, N.; Holick, M.F. Immunologic effects of vitamin D on human health and disease. Nutrients 2020, 12, 2097. [Google Scholar] [CrossRef]
- Smolders, J.; van den Ouweland, J.; Geven, C.; Pickkers, P.; Kox, M. Vitamin D deficiency in COVID-19: Mixing up cause and consequence. Metabolism-Clinical and Experimental 2021, 115. [Google Scholar] [CrossRef]
- Singh, P.; Rawat, A.; Alwakeel, M.; Sharif, E.; Al Khodor, S. The potential role of vitamin D supplementation as a gut microbiota modifier in healthy individuals. Scientific reports 2020, 10, 21641. [Google Scholar] [CrossRef]
- Bellerba, F.; Muzio, V.; Gnagnarella, P.; Facciotti, F.; Chiocca, S.; Bossi, P.; Cortinovis, D.; Chiaradonna, F.; Serrano, D.; Raimondi, S. The association between vitamin D and gut microbiota: a systematic review of human studies. Nutrients 2021, 13, 3378. [Google Scholar] [CrossRef]
- Forrest, K.Y.; Stuhldreher, W.L. Prevalence and correlates of vitamin D deficiency in US adults. Nutrition research 2011, 31, 48–54. [Google Scholar] [CrossRef]
- Cashman, K.D.; Dowling, K.G.; Škrabáková, Z.; Gonzalez-Gross, M.; Valtueña, J.; De Henauw, S.; Moreno, L.; Damsgaard, C.T.; Michaelsen, K.F.; Mølgaard, C. Vitamin D deficiency in Europe: pandemic? The American journal of clinical nutrition 2016, 103, 1033–1044. [Google Scholar] [CrossRef]
- Sharif, E.A.; Rizk, N. The prevalence of vitamin D deficiency among female college students at Qatar University. 2011.
- Mullish, B.H.; Marchesi, J.R.; McDonald, J.A.; Pass, D.A.; Masetti, G.; Michael, D.R.; Plummer, S.; Jack, A.A.; Davies, T.S.; Hughes, T.R. Probiotics reduce self-reported symptoms of upper respiratory tract infection in overweight and obese adults: should we be considering probiotics during viral pandemics? Gut Microbes 2021, 13, 1900997. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, G.; Borrazzo, C.; Pinacchio, C.; Santinelli, L.; Innocenti, G.P.; Cavallari, E.N.; Celani, L.; Marazzato, M.; Alessandri, F.; Ruberto, F. Oral bacteriotherapy in patients with COVID-19: a retrospective cohort study. Frontiers in Nutrition 2021, 7, 613928. [Google Scholar] [CrossRef] [PubMed]
- Michael, D.; Davies, T.; Jack, A.; Masetti, G.; Marchesi, J.; Wang, D.; Mullish, B.; Plummer, S. Daily supplementation with the Lab4P probiotic consortium induces significant weight loss in overweight adults. Scientific reports 2021, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- García-Mena, J.; Corona-Cervantes, K.; Cuervo-Zanatta, D.; Benitez-Guerrero, T.; Vélez-Ixta, J.M.; Zavala-Torres, N.G.; Villalobos-Flores, L.E.; Hernández-Quiroz, F.; Perez-Cruz, C.; Murugesan, S. Gut microbiota in a population highly affected by obesity and type 2 diabetes and susceptibility to COVID-19. World Journal of Gastroenterology 2021, 27, 7065. [Google Scholar] [CrossRef] [PubMed]
- Ivashkin, V.; Fomin, V.; Moiseev, S.; Brovko, M.; Maslennikov, R.; Ulyanin, A.; Sholomova, V.; Vasilyeva, M.; Trush, E.; Shifrin, O. Efficacy of a Probiotic Consisting of Lacticaseibacillus rhamnosus PDV 1705, Bifidobacterium bifidum PDV 0903, Bifidobacterium longum subsp. infantis PDV 1911, and Bifidobacterium longum subsp. longum PDV 2301 in the Treatment of Hospitalized Patients with COVID-19: a Randomized Controlled Trial. Probiotics and antimicrobial proteins 2021, 1–9. [Google Scholar]
- Kurian, S.J.; Unnikrishnan, M.K.; Miraj, S.S.; Bagchi, D.; Banerjee, M.; Reddy, B.S.; Rodrigues, G.S.; Manu, M.K.; Saravu, K.; Mukhopadhyay, C. Probiotics in prevention and treatment of COVID-19: current perspective and future prospects. Archives of medical research 2021, 52, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, H.S.; Bilen, Ö. Oral booster probiotic bifidobacteria in SARS-COV-2 patients. International journal of immunopathology and pharmacology 2021, 35, 20587384211059677. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Castrellón, P.; Gandara-Martí, T.; Abreu Y Abreu, A.T.; Nieto-Rufino, C.D.; López-Orduña, E.; Jiménez-Escobar, I.; Jiménez-Gutiérrez, C.; López-Velazquez, G.; Espadaler-Mazo, J. Probiotic improves symptomatic and viral clearance in Covid19 outpatients: a randomized, quadruple-blinded, placebo-controlled trial. Gut Microbes 2022, 14, 2018899. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Z.; Mak, J.W.; Chow, K.M.; Lui, G.; Li, T.C.; Wong, C.K.; Chan, P.K.; Ching, J.Y.; Fujiwara, Y. Gut microbiota-derived synbiotic formula (SIM01) as a novel adjuvant therapy for COVID-19: An open-label pilot study. Journal of Gastroenterology and Hepatology 2022, 37, 823–831. [Google Scholar] [CrossRef]
- Saviano, A.; Potenza, A.; Siciliano, V.; Petruzziello, C.; Tarli, C.; Migneco, A.; Nasella, F.; Franceschi, F.; Ojetti, V. COVID-19 pneumonia and gut inflammation: the role of a mix of three probiotic strains in reducing inflammatory markers and need for oxygen support. Journal of Clinical Medicine 2022, 11, 3758. [Google Scholar] [CrossRef]
- Zhao, Y.; Dong, B.R.; Hao, Q. Probiotics for preventing acute upper respiratory tract infections. Cochrane database of systematic reviews 2022. [Google Scholar]
- Arena, M.P.; Capozzi, V.; Russo, P.; Drider, D.; Spano, G.; Fiocco, D. Immunobiosis and probiosis: antimicrobial activity of lactic acid bacteria with a focus on their antiviral and antifungal properties. Applied microbiology and biotechnology 2018, 102, 9949–9958. [Google Scholar] [CrossRef]
- Kang, J.Y.; Lee, D.K.; Ha, N.J.; Shin, H.S. Antiviral effects of Lactobacillus ruminis SPM0211 and Bifidobacterium longum SPM1205 and SPM1206 on rotavirus-infected Caco-2 cells and a neonatal mouse model. Journal of Microbiology 2015, 53, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Hanada, S.; Pirzadeh, M.; Carver, K.Y.; Deng, J.C. Respiratory viral infection-induced microbiome alterations and secondary bacterial pneumonia. Frontiers in immunology 2018, 9, 418009. [Google Scholar] [CrossRef]
- Harding, J.N.; Siefker, D.; Vu, L.; You, D.; DeVincenzo, J.; Pierre, J.; Cormier, S.A. Altered gut microbiota in infants is associated with respiratory syncytial virus disease severity. BMC microbiology 2020, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Harper, A.; Vijayakumar, V.; Ouwehand, A.C.; Ter Haar, J.; Obis, D.; Espadaler, J.; Binda, S.; Desiraju, S.; Day, R. Viral infections, the microbiome, and probiotics. Frontiers in Cellular and Infection Microbiology 2021, 10, 596166. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Rosário, A.; Marques, C.; Pinheiro, H.; Araújo, J.R.; Ribeiro, P.; Rocha, R.; Mota, I.; Pestana, D.; Ribeiro, R.; Pereira, A. Gut microbiota diversity and C-reactive protein are predictors of disease severity in COVID-19 patients. Frontiers in Microbiology 2021, 12, 705020. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Zhang, G.; Wang, X.; Guo, M.; Zeng, W.; Xu, Z.; Cao, D.; Pan, A.; Wang, Y.; Zhang, K. Analysis of the intestinal microbiota in COVID-19 patients and its correlation with the inflammatory factor IL-18. Medicine in Microecology 2020, 5, 100023. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Cai, H.; Shen, Y. Management of corona virus disease-19 (COVID-19): the Zhejiang experience. J Zhejiang Univ (Med Sci) 49 (1): 0. 2020.
- Reinold, J.; Farahpour, F.; Fehring, C.; Dolff, S.; Konik, M.; Korth, J.; van Baal, L.; Hoffmann, D.; Buer, J.; Witzke, O. A pro-inflammatory gut microbiome characterizes SARS-CoV-2 infected patients and a reduction in the connectivity of an anti-inflammatory bacterial network associates with severe COVID-19. Frontiers in Cellular and Infection Microbiology 2021, 11, 747816. [Google Scholar] [CrossRef]
- Zuo, T.; Wu, X.; Wen, W.; Lan, P. Gut microbiome alterations in COVID-19. Genomics, Proteomics and Bioinformatics 2021, 19, 679–688. [Google Scholar] [CrossRef]
- Hazan, S.; Stollman, N.; Bozkurt, H.S.; Dave, S.; Papoutsis, A.J.; Daniels, J.; Barrows, B.D.; Quigley, E.M.; Borody, T.J. Lost microbes of COVID-19: Bifidobacterium, Faecalibacterium depletion and decreased microbiome diversity associated with SARS-CoV-2 infection severity. BMJ open Gastroenterology 2022, 9, e000871. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, H.S.; Quigley, E.M. The probiotic Bifidobacterium in the management of Coronavirus: A theoretical basis. International Journal of Immunopathology and Pharmacology 2020, 34, 2058738420961304. [Google Scholar] [CrossRef] [PubMed]
- Khaled, J.M. Probiotics, prebiotics, and COVID-19 infection: A review article. Saudi journal of biological sciences 2021, 28, 865–869. [Google Scholar] [CrossRef]
- Khodavirdipour, A.; Asadimanesh, M.; Masoumi, S.A. Impact of SARS-CoV-2 genetic blueprints on the oral manifestation of COVID-19: a case report. Global Medical Genetics 2021, 8, 183–185. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Moens, F.; Selak, M.; Rivière, A.; Leroy, F. Summer Meeting 2013: growth and physiology of bifidobacteria. Journal of applied microbiology 2014, 116, 477–491. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.J.; Shin, H.S. Antimicrobial and immunomodulatory effects of bifidobacterium strains: A review. Journal of Microbiology and Biotechnology 2020, 30, 1793. [Google Scholar] [CrossRef]
- Hughes, K.R.; Harnisch, L.C.; Alcon-Giner, C.; Mitra, S.; Wright, C.J.; Ketskemety, J.; van Sinderen, D.; Watson, A.J.; Hall, L.J. Bifidobacterium breve reduces apoptotic epithelial cell shedding in an exopolysaccharide and MyD88-dependent manner. Open biology 2017, 7, 160155. [Google Scholar] [CrossRef]
- Dyakov, I.N.; Mavletova, D.A.; Chernyshova, I.N.; Snegireva, N.A.; Gavrilova, M.V.; Bushkova, K.K.; Dyachkova, M.S.; Alekseeva, M.G.; Danilenko, V.N. FN3 protein fragment containing two type III fibronectin domains from B. longum GT15 binds to human tumor necrosis factor alpha in vitro. Anaerobe 2020, 65, 102247. [Google Scholar] [CrossRef]
- Cervantes, J.; Hong, B.-y. The gut–lung axis in tuberculosis. Pathogens and Disease 2017, 75, ftx097. [Google Scholar] [CrossRef]
- Ruiz, L.; Delgado, S.; Ruas-Madiedo, P.; Sánchez, B.; Margolles, A. Bifidobacteria and their molecular communication with the immune system. Frontiers in microbiology 2017, 8, 302400. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, M.; Martín, R.; Torres-Maravilla, E.; Chadi, S.; González-Dávila, P.; Sokol, H.; Langella, P.; Chain, F.; Bermúdez-Humarán, L.G. Butyrate mediates anti-inflammatory effects of Faecalibacterium prausnitzii in intestinal epithelial cells through Dact3. Gut Microbes 2020, 12, 1826748. [Google Scholar] [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The controversial role of human gut lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, J.; Zhang, D.; Ma, W.-L.; Wang, X. Linking the gut microbiota to persistent symptoms in survivors of COVID-19 after discharge. Journal of Microbiology 2021, 59, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Mendes de Almeida, V.; Engel, D.F.; Ricci, M.F.; Cruz, C.S.; Lopes, Í.S.; Alves, D.A.; d’Auriol, M.; Magalhães, J.; Machado, E.C.; Rocha, V.M. Gut microbiota from patients with COVID-19 cause alterations in mice that resemble post-COVID symptoms. Gut Microbes 2023, 15, 2249146. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Zonulin is a molecular gatekeeper that regulates TJs between intestinal epithelial cells. Normally, these TJs serve as a selective barrier, preventing chemicals and pathogens from entering the bloodstream from the intestines. SARS-CoV-2 infection in the GI tract causes localized inflammation of the mucosa, which in turn triggers zonulin release. This, in turn, enhances gut permeability, allowing SARS-CoV-2 antigens, such as the spike protein's superantigen-like motif, to pass through mucosal barriers and enter the bloodstream. Parts of the figure were drawn using pictures from Servier Medical Art. Servier Medical Art by Servier is licensed under a Creative Commons Attribution 3.0 Unported License.
Figure 1.
Zonulin is a molecular gatekeeper that regulates TJs between intestinal epithelial cells. Normally, these TJs serve as a selective barrier, preventing chemicals and pathogens from entering the bloodstream from the intestines. SARS-CoV-2 infection in the GI tract causes localized inflammation of the mucosa, which in turn triggers zonulin release. This, in turn, enhances gut permeability, allowing SARS-CoV-2 antigens, such as the spike protein's superantigen-like motif, to pass through mucosal barriers and enter the bloodstream. Parts of the figure were drawn using pictures from Servier Medical Art. Servier Medical Art by Servier is licensed under a Creative Commons Attribution 3.0 Unported License.
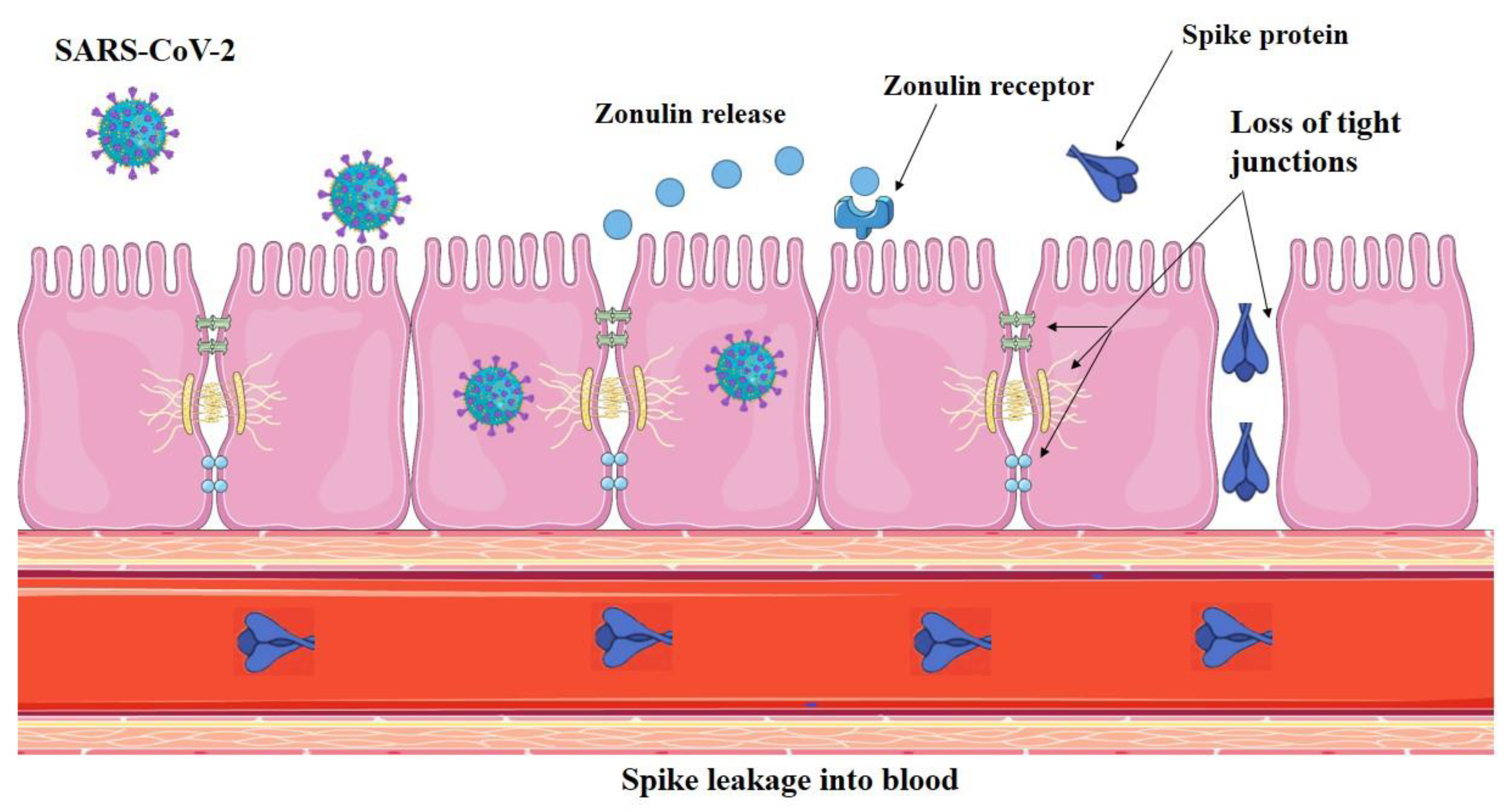
Figure 2.
SARS-CoV-2 can infect intestinal bacteria, thus showing a bacteriophage-like behavior. Panels a and b: Images from a transmission electron microscope revealed the presence of SARS-CoV-2 inside two bacteria, marked by black arrows. Source: [93]. This figure is open access and is distributed under the Creative Commons Attribution Non-Commercial (CC BY-NC 4.0) license.
Figure 2.
SARS-CoV-2 can infect intestinal bacteria, thus showing a bacteriophage-like behavior. Panels a and b: Images from a transmission electron microscope revealed the presence of SARS-CoV-2 inside two bacteria, marked by black arrows. Source: [93]. This figure is open access and is distributed under the Creative Commons Attribution Non-Commercial (CC BY-NC 4.0) license.
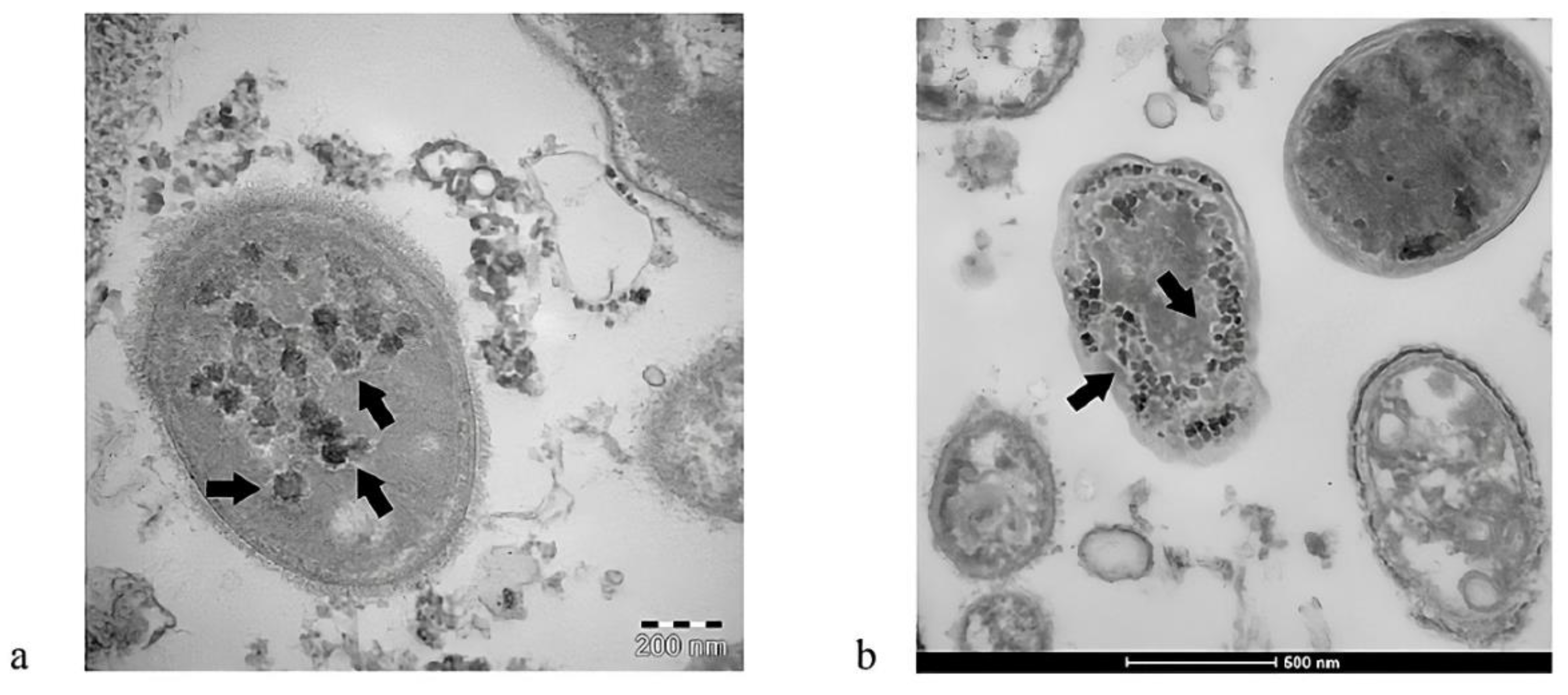
Figure 3.
A graphical representation shows the influence of probiotics on intestinal barrier function in immunological and inflammatory conditions. Source: [137]. This figure is open access and is distributed under the Creative Commons Attribution Non-Commercial (CC BY-NC 4.0) license.
Figure 3.
A graphical representation shows the influence of probiotics on intestinal barrier function in immunological and inflammatory conditions. Source: [137]. This figure is open access and is distributed under the Creative Commons Attribution Non-Commercial (CC BY-NC 4.0) license.
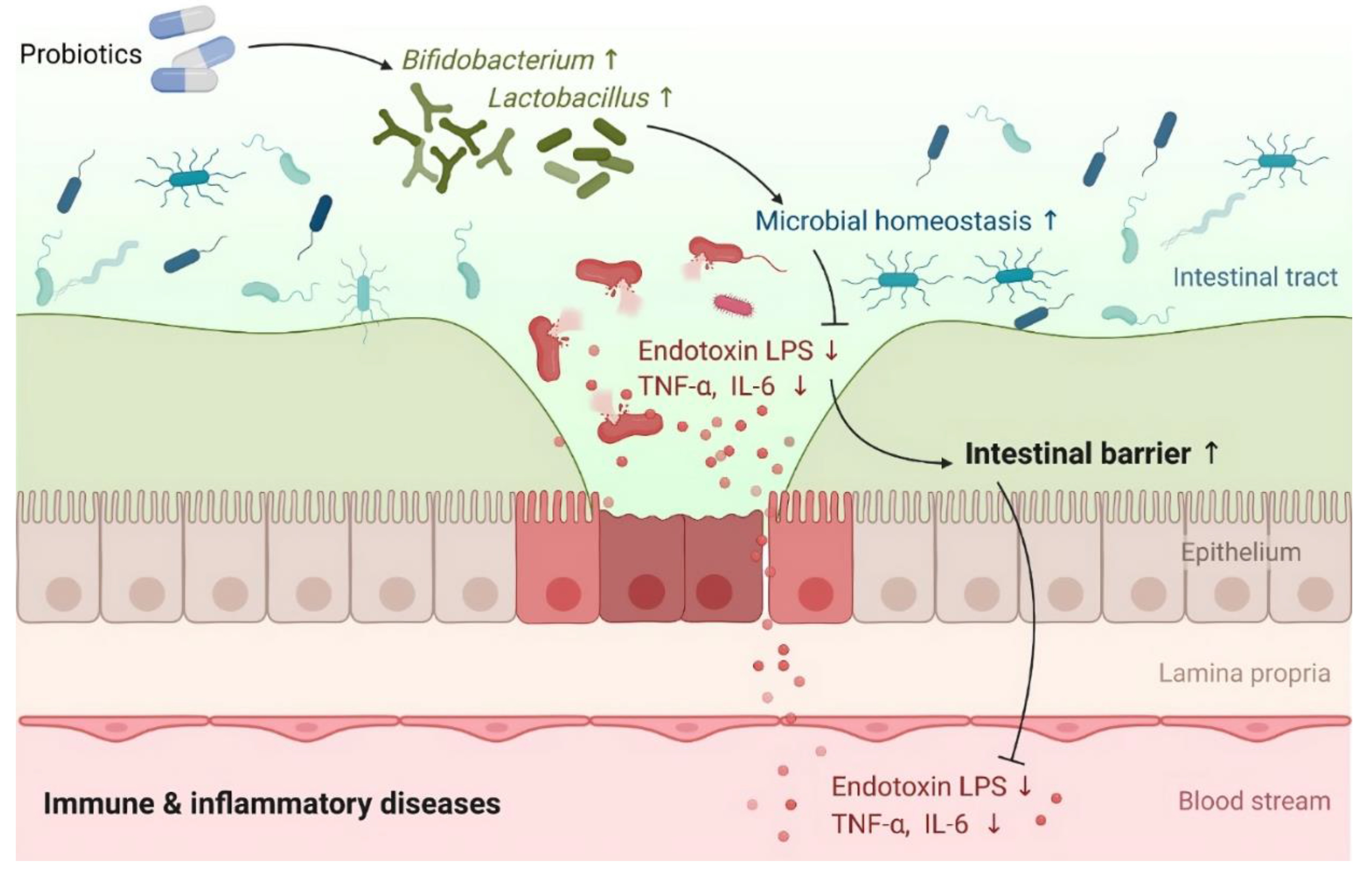
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



