
Submitted:
21 June 2024
Posted:
24 June 2024
You are already at the latest version
Abstract
Diffuse type of gastric cancer (GC) has an increasing prevalence worldwide, especially in Western countries, is usually diagnosed at advanced stages, and has no efficacious treatment options. Epidemiological studies have reported an increased mortality from GC after occupational exposure to well-studied pro-carcinogen that are metabolically activated by Cyp P450 enzymes through aryl hydrocarbon receptor (AhR). Substantial studies support the involvement of AhR in gastric carcinogenesis. However, little is known about the role of AhR in diffuse GC, as compared to intestinal GC. In a cohort of 29 gastric tumors, we described a significantly increased AhR protein and mRNA expression levels in GCs, independently of subtypes and clinical parameters. AhR and RhoA mRNA expression were correlated in diffuse GC. Further, our study characterized how AhR affects gene expression in diffuse GC. Using qRT-PCR, we compared the expression levels of AhR, Cyp1A1 and Cyp1B1 to the expression of genes in a panel previously described. In diffuse GC, Cyp1A1 expression correlated with genes involved in IGF signalling, EMT (VIM), migration (MMP2). In an in vitro assay using the poorly differentiated KATOIII epithelial cell line, two well-known ligands for AhR (TCDD and BaP) induced mRNA expression of CYP1A1, IL1b, as well as UGT1, NQO1 and AhRR to a lower extent. We also observed a strong increase in Cyp1B1 expression in diffuse GC, along with a lower TCDD-increased Cyp1B1 expression as compared to Cyp1A1 in KATOIII cells, and immunostaining in stromal cells. In intestinal GC, Cyp1B1 inversely correlated with several genes including IDO1 (generating endogenous kynurenin-e AhR ligand). Our data provide evidence for a major role of AhR in GC, as an environmental xenobiotics receptor, through different mechanisms and pathways in diffuse and intestinal GC. Our results support continued efforts to clarify the identities of exogenous AhR ligands in diffuse GC in order to define new therapeutic strategies.
Keywords:
gastric cancers
; aryl hydrocarbon receptor (AhR)
; diffuse-gastric subtype
; intestinal-gastric subtype
; benzo[a]pyrene (BaP)
; 2
; 3
; 7
; 8-tetrachlorodibenzeno-p-dioxin (TCDD or dioxin)
; xenobiotic metabolism
; tryptophan metabolism.
1. Introduction
Gastric cancer (GC) still remains the fourth leading cause of cancer mortality [1,2,3], with high heterogeneity in different subtypes according to the classification proposed by the World Health Organization [4]. Regardless of the country, the majority of gastric tumors are adenocarcinomas, which can be more histologically classified into diffuse-and intestinal- sub types according to the Lauren classification [5]. While intestinal- subtype GC is well differentiated and related to Helicobacter pylori infection, the diffuse- subtype is poorly differentiated, and can be seen in familial (germline mutation in the CDH1 gene) or sporadic settings, as an infiltrating and scattered type [6,7] with unknown origin. The prevalence of the diffuse type is increasing worldwide especially in Western countries [8]. The majority of patients with diffuse- GC are usually diagnosed at advanced stages, with positive axillary nodes (83%) and peritoneal carcinomatosis (18.6%) [6,9,10].
The hypothesis of a link between exposure to persistent organic pollutants (POPs including dioxin) and cancers, including GC, is supported by several epidemiological studies of accidentally exposed populations in Seveso [11]. It is also known that polycyclic polyhalogenated hydrocarbons (like dioxins and PCBs) and polycyclic aromatic hydrocarbons (PAHs) including benzo[a]pyrene from tobacco-smoke and biomass burning) may link to relatively high health risk including cancers. We have previously shown that environmental chemicals such as persistent organic pollutants (POPs) are involved in breast cancers [12], and may accumulate in the Omentum adipose tissue of patients with GC [13]. In addition to TCDD, the prototypical and most potent known environmental ligand in animals and humans, other widespread environmental POPs contaminants bind aryl hydrocarbon receptor (AhR, a basic helix-loop-helix transcription factor) with strong affinities [14,15] and may chronically activate AhR in cancer agression.
For decades, AhR has been studied for its role in environmental chemical toxicity and a mediator of unintended consequences of human pollution characterized as a regulator of xenobiotic metabolic enzymes such as cytochromes P450 Cyp1A1 and Cyp1B1 [16]. Numerous studies have now demonstrated that AhR is involved in important cellular and pathological processes such as tumor initiation and progression including control of proliferation and migration, angiogenesis and control of the immune system [12,15,17,18,19,20,21,22,23]. However, the expression of AhR and AhR-signaling pathways has not yet been investigated for Western patients with diffuse GCs.
Various animal experimental data have provided substantial support for an association between AhR expression/function and GCs (23-28). Transgenic mice that express a constitutively active dioxin/AhR mutant (CA-AhR), due to a deletion in the ligand binding domain of AhR, rapidly developed stomach cancers [24,25]. Several tissues of CA-AhR mice have shown life-long continuous low-level activity of AhR, and this model is useful to mimic the dioxin exposure of humans in the general population [26]. Mice exposed to the chemical carcinogen BaP [27] also developed gastric tumors. In humans, elevated AhR expression has been reported in GC [28,29].
In this study, we have analyzed the expression of AhR in a series of diffuse- and intestinal-subpopulations of gastric tumors, the expression of xenobiotic metabolic enzymes such as cytochromes P450 (Cyp1A1 and Cyp1B1) and the expression of a large panel of genes known to be regulated by AhR in several cancers. We have also analyzed the effect of exogenous environmental Ahr ligands on 2 gastric cell lines in vitro.
2. Materials and Methods
2.1. Patients and Tissue Samples
A total of 29 patients underwent partial gastrectomy for histopathologically-confirmed gastric adenocarcinoma primary tumor tissue in the Lariboisiere Hospital (Paris, France) from 2005 to 2014. All patients provided written informed consent prior to their inclusion in the study. Biopsies (provided before 2014) were taken for diagnostic and research purposes, and analysis was permitted by the Ethical Committee of Lariboisiere Hospital (Paris). Eligibility criteria included (1) gastric carcinoma identified by histopathological examination, (2) no other malignancy, (3) no pre-operative chemotherapy or radiotherapy, and (4) availability of complete clinical, histological and biological data. Normal (non-malignant) samples refer to samples harvested from the stomach, from sites distant from the tumor. Immediately after surgery, fresh gastric tumors and their matched normal mucosa were stored in liquid nitrogen until mRNA extraction; other tumor samples and their adjacent normal tissues were routinely fixed in 10% buffered formalin and embedded in paraffin for histological analysis. As previously described [10], the population was divided into two groups according to the histological status of GC: diffuse-subtype adenocarcinoma (a poorly differentiated, infiltrating and scattered type) or intestinal-subtype adenocarcinoma (a well differentiated and clustered subtype), according to the Lauren Classification (see Table 1).The malignancy of infiltrating carcinomas was scored according to the TNM staging system (Stage I to IV) as previously described [10]: first according to AJCC7 [30], revised from IGCA [31,32] and AJCC8 [33]. This TNM staging includes T scores for the primary tumor (T1-T4), N scores (lymph node metastasis) and M scores (metastasis).
2.2. Total RNA Preparation and qRT-PCR
The conditions for total RNA extraction, complementary cDNA synthesis and qRT-PCR conditions were as previouly described [12,34]. Primers for AhR, AhRR and other genes were selected using the Oligo 6.0 program (National Biosciences, Plymouth, MN) [12,29]. Each sample was normalized on the basis of its TBP content as previously described [10,12]. Results, expressed as N-fold differences in target gene expressions relative to the TBP gene (and termed “Ntarget”), were determined as Ntarget = 2ΔCtsample, where the ΔCt value of the sample was determined by subtracting the average Ct value of the specific target gene from the average Ct value of the TBP gene. Ntarget values of the samples were subsequently normalized so that the median of Ntarget values for normal gastric tissues (n=11) was 1. Target gene expression was normalized to their transcription level of house keeping genes TBP, P0 and PPIA. Preliminary analysis of gene expression did not indicate changes in median basal levels in normal samples in the same patients (with either diffuse- or intestinal- GC subtypes). For each gene expression, normalized RNA values of 3 (or more) were considered to represent gene overexpression in tumor samples, and values 0.33 (or less) represented gene underexpression.
2.3. Immunohistochemistry
Immunohistochemical labeling (IHC) was performed on paraffin sections (4 μm), as previously described [12,29]. Immunohistochemical staining for AhR and Cyp1B1 (Santa Cruz, dilution 1/50 and 1/200 respectively) were performed using the Ventana Autostainer (USA). Specificity was checked by control staining performed in the absence of primary antibody and with positive tissue [12] The antigen-antibody complex was visualized using DAB as chromogen. Immunostaigning was analysed blindly in duplicate by two specialists including a certified pathologist.
2.4. Cell Culture
The human gastric cell lines, KATO-III obtained from a poorly differentiated gastric adenocarcinoma, and AGS from moderately differentiated GC were acquired from ATCC (Manassas, VA, USA). Cells were maintained in Dulbecco’s modified Eagle medium supplemented with 10% heat-inactivated fetal bovine serum, 0.5% penicillin-streptomycin, and 2 nM of L-glutamine (Gibco, Saint Aubin, France). Cells were grown at 37˚C in a humidified 5% CO2/95% air atmosphere. Exponentially growing cells were trypsinized and seeded in flasks; the medium was replaced every 24h; when cells reached 70-80% confluence by microscopic examination, the medium was changed, and compounds (2, 3, 7, 8-tetrachlorodibenzeno-p-dioxin or TCDD (1-30 nM) and BaP (10 μM) (gift from P Balaguer, Montpellier, France) were added for 16-24h. Control experiments included the addition of CHH 223191 (10 μM), a full AhR antagonist (a gift from P Balaguer)
2.5. Statistical Analysis
As the mRNA levels of gene expression did not fit a Gaussian distribution, the relative expression of genes was characterized by the median and the range rather than their mean values and coefficient of variation [10,12,29]. For each gene, differences of expression between tumors versus normal tissues (fold change) were analyzed as previously described [10,12]. Differences in the number of samples that over- (>3-fold) or and under- (<3-fold) expressed were analyzed using the Chi2-square test [29]. The relationships between expressions of genes in GC were determined using non parametric Spearman’s rank correlation test. Relationships between expression levels and clinical parameters were analyzed using non parametric Kruskal-Wallis (or Mann-Whitney) and Chi-square tests, as indicated in each Table. Statistical analyses were performed using Prism 5.03 software (GraphPad, San Diego, CA, USA). Differences were considered significant at confidence levels greater than 95% (p<0.05).
3. Results
3.1. Patient Characteristics
The clinical characteristics of the patients are shown in Table 1. Patients with diffuse-subtype GC were younger as than patients with intestinal-subtype (57 [27–71] years and 75 [59–82)]) years respectively, p=0.0004). Both subtypes of carcinoma had large tumors (>50 mm) and tumor invasion (T3-T4) [10]. Within each subtype, half of the patients smoked. Patients with diffuse adenocarcinoma had more lymphatic invasion (p=0.0014) and metastasis (31% vs 6%) than patients with intestinal-subtype. Most diffuse GC was TNM stage III-IV, while patients with intestinal-subtype were stage I, II and III. Vascular and neural invasion did not differ among different GC subtypes.
3.2. High AhR Expression in Gastric Tumors Both at the mRNA and Protein Levels
The cohort of GC specimens were first used to assess AhR mRNA expression levels. As compared to normal gastric tissue samples, AhR expression was significantly increased in gastric tumors (x1.94, p=0.002), both diffuse-and intestinal- subtypes GC (x2.12, p=0.001 and x1.60, p=0.003, respectively) (Table 2). Moreover, AhR mRNA expression was independent of classical clinical parameters in GCs and their subtypes, i.e., gender, age, tumor grade, lymphatic invasion, metastasis status, TNM stage, vascular or neural invasion (Supplementary Table S1).
Most importantly at the protein expression levels, strong nuclear AhR staining was observed in GC (Figure 1B) in epithelial and stromal cells, including fibroblasts, endothelial and immune cells [29] as compared to the weak cytoplasmic and nuclear staining observed in epithelial cells in non-tumoral tissue ((Figure 1A).
Altogether, these results indicate that AhR (mRNA and protein) is significantly increased in GCss, both in diffuse- and intestinal- adenocarcinoma, as compared to control samples.
3.3. Expression of AhR-Target Genes Encoding Xenobiotic Metabolizing Enzymes in Gastric Cancers
As GC express high AhR expression levels (mRNA and protein), we underwent to analyse changes in the expression of classic target genes of AhR, such as CYP1A1 and CYP1B1, two genes involved in xenobiotic metabolism [16]. CYP1A1 was expressed at a low level in normal tissues (Table 2). As compared to the non-tumoral tissue, enhanced expression of Cyp1A1 was attained with an increase of more than 3-fold observed in 5/29 of GC cases (17%) and in 23% diffuse GC. CYP1B1 expression was significantly increased in diffuse GC (p=0.014, 92% lymphatic invasion, Table 2 and Table 3). At the protein level, Cyp1B1 was mainly observed in the stromal compartment in diffuse GC (Figure 1C). In all tumors, CYP1B1 expression was independent of clinical parameters (gender, age, smoking, tumor grade (T), except for an increase with lymphatic invasion (p<0.02) and TNM (p<0.05 respectively) (Table 3).
3.4. AhR Ligands such as Environmental Ligands Induced mRNA Expression of CYP1A, IL1β, UGT1A1 and AhRR in Gastric Epithelial Cell Lines
We further examined the effect of two AhR ligands: TCDD (2,3,7,8-tetrachlorodibenzo-p-dioxin or dioxin), the most potent known environmental AhR ligand, and BaP (benzo[a]pyrene, a prototypical polycyclic aromatic hydrocarbon found in combustion processes and cigarette smoke) on mRNA expression of several genes in two epithelial gastric cell lines (KATO III and AGS) using qRT-PCR. These cells are poorly (KATO III) and moderately (AGS) differentiated, with high AhR expression.
In KATO III cells, TCDD (30 nM) strongly increased expression of CYP1A1 as compared to control cells (x15, p<0.0001) (Figure 2d). The TCDD-induced CYPA1 expression was reversed by CHH 223191, a full AhR antagonist (p<0.001) (Figure 2d). BaP (10 μM), a well-studied pro-carcinogen, also increased CYP1A expression (x5, p<0.001) (Figure 2d). TCDD (30 nM) also significantly stimulated the expression of IL1B (x5.5, p<0.0001), UGT1A and AhRR (x2, p<0.0001) (Figure 2c, e,f). We did not detect significant effects of TCDD (or BaP) on AhR and ARNT expression in KATO III cells (Figure 2a, b). In contrast to CYP1A1, basal and TCDD-increased levels of CYP1B1 were low in epithelial KATO III cells (Figure 3A) or undetectable (qPCR threshold >CT50) in AGS cells (not shown). Studies in AGS cells also showed that CYP1A, but not AhR or AhRR, was significantly increased following TCDD treatment (Figure 3B a-c).
3.5. Correlations of AhR, Cyp1A1 and Cyp1B1, AhRR, Expression with a Panel of Genes Involved in AhR-Related Signaling Pathways
AhR is known to activate several signalling pathways governing proliferation, epithelial-mesenchymal transition (EMT), cell migration, inflammation, immunity and angiogenesis in cancers [12,15,17,18,19,20,21,22,23]. We then compared the mRNA expression levels of AhR, CYP1A1 and CYP1B1 with the expression of 36 genes involved in EMT, cell proliferation and migration, immunity and angiogenesis, that we previously described in our cohort of GCss [10,29]. AhR expression was correlated with RHOA expression in both GC subtypes (Table 4). In diffuse GC, CYP1A1 expression correlated with several genes such as growth factors IGF1 (p=0.001), IGFR2 (p=0.015), genes involved in EMT such as VIM (p=0.007), SNAI2 and ZEB2 (p=0.04), and migration (MMP2, p=0.01) (Table 4). CYP1B1 expression was only inversely correlated with ERRB2 (p=0.01). Moreover, significantly increased AhRR expression (x2.65, p<0.01) positively correlated mainly with IGF1 (p=0.0001), TWIST2 and ZEB2 (p<0.02), MMP2 (p<0.002) and NRP1 (p=0.02) in diffuse, but not intestinal GC (Table 4). In intestinal GC, low expression of CYP1A1 and CYPB1 inversely correlated with IDO1 in intestinal GC (p=0.014 and p=0.001, respectively, Table 4). Low CYP1B1 expression also correlated with genes that were not significantly increased including growth factors and receptors, EMT (VIM, SNAI2, TWIST2), VEGF and NRP1 (Table 4).
4. Discussion
Although AhR is known for its role in environmental chemical toxicity, as a mediator of unintended consequences of human pollution, and its involvement in tumor initiation and progression [24,25,26,28,35], the relationship between AhR expression, pollution linked AhR-dependent function and Western patients with GCs remains unexplored. Because diffuse GC is increasing in prevalence worldwide in Western countries, usually diagnosed at advanced stages, and has no efficacious treatment options, exploration of its cellular and molecular causes is crucial. We reported significant high expression of AhR in our Western cohort of GC, independently of their clinical subtypes [29]. A link between exposure to persistent organic pollutants (POPs) and diffuse GC is supported by the study of accidentally exposed populations in Seveso [11]. In the study presented here, we have analyzed the expression of AhR and several AhR-regulated genes in a series of gastric tumors including diffuse- and intestinal-GC. Furthermore, we have studied the impact of two AhR ligands well-known for their critical role in cancer development linked to pollution [26,27].
Expression of AhR and RHOA. Using RT-PCR, we found a correlation between the expression of AhR and RHOA in GCs independently of their subtypes (Table 4), as previously documented for other types of cancers [12,36,37]. A higher expression of RHOA has been found in diffuse GC with 85% overexpression (>3fold) as compared to normal samples, along with 50% in intestinal subtype) [10]. The functional and coordinated role of RhoA in the development of cancers involves several processes such as cell proliferation, migration, invasion and angiogenesis [36,37,38,39,40]. Increase RhoA activity is correlated with worse overall survival in diffuse patients [41]. Interestingly, RHOA transcription has recently been shown to be initiated by a ligand-AhR-Arnt complex. Somatic alterations in RHOA and CDH1 have been reported in aggressive diffuse GC and generally associated with familial disease [42,43]. However, our Western cohort of diffuse GC by the Lauren classification did not include familial GC.
Expression and distribution of Cyp1A1 and Cyp1B1. Cytochrome P450-1 enzymes are inducible forms of the cytochrome P450 family of xenobiotic metabolizing enzymes [44,45]. In our study, Cyp1A1 was detected at very low level in stomach, but overexpressed (>3) in 23% in diffuse GC as compared to non-tumoral tissue. Cyp1A1 was also highly induced by TCDD, a non-genotoxic AhR, in undifferentiated diffuse GC (Kato III) cells. Cyp1B1 was the most significantly expressed form in diffuse GC, as previously reported in a wide range of human cancers including breast, colon, lung and others [44]. Cyp1A1 and Cyp1B1 have a central role in tumor development and in the activation step of pro-carcinogen compounds such as benzopyrene (BaP) [16,45,46,47,48,49]. BaP (benzo[a]pyrene) is a prototypical polycyclic aromatic hydrocarbon (PAH) found in tobacco and combustion processes such as biomass burning [50,51].
Our in vitro experiments using two GC cell lines (KATO III and AGS) indicated that TCDD, and BaP to a lesser extent, strongly induced CYP1A1, UGT1A1 and NQO1 in KATO III epithelial cells, but low levels of CYP1B1, as compared to unexposed cells (Figs 2 and 3A). Cyp1A1 was also increased following TCDD treatment in AGS cells as compared to unexposed cells. Functional DRE enhancer element has been identified in vitro and in vivo for AhR target genes including CYP1A1, CYP1B1 and NQO1, which encode phase I and II xenobiotic metabolizing enzymes [52]. Our in vivo study also shows that CYP1B1expression is significantly increased (p=0.014) in diffuse GC, as compared to intestinal GC. At the protein level, Cyp1B1 was mainly observed in the stromal compartment in diffuse GC (Figure 1C). It is well known that stromal cells, such as fibroblasts and macrophages, express CYP1B1, but not CYP1A1, in response to TCDD or benzopyrene [53,54]. Taken together, our results suggest a cell-specific distribution of Cyp1B1 and Cyp1A1 in diffuse GC. Our results also suggest that activated AhR may contribute to the tumor-stroma interaction (through Cyp1A1 and Cyp1B1) in diffuse GC.
Funtional role of AhR, Cyp1A1 and Cyp1B1. Animal and clinical data provide evidence for the role of AhR in gastric tumorigenesis, implicating the receptor in regulation of tumor growth, EMT, migration, invasion and cancer aggression [24,25,28,35]. The connection between EMT, a process allowing cells to transition from epithelial to mesenchymal, and tumorigenesis has been established in human cancers involving several pathways such as activation of Wnt/beta-catenin signaling through Cyp1s [55,56], or hedgehog signaling [57,58]. It is also well known that Cyp1A1 and Cyp1B1 have important roles in both tumor development (cell invasion, migration, and disease progression), in addition to the metabolic activation of BaP related to carcinogenesis. Analysis of CYP1s and co-regulated genes using large scale analysis may also be helpful for functional studies [59]. Using RT-qPCR, we compared the expression of CYP1s with the expression levels of 36 genes coding for proteins that have been previously studied in the same cohort of patients [10,29]. These genes were selected on the basis of their roles in proliferation, the IGF pathway, the EMT signature, migration, angiogenesis or immunity. In diffuse GC, but not intestinal GC, CYP1A1 expression was strongly correlated with expression of genes involved in proliferation (IGF1, p=0.001), EMT signature such as VIM, p=0.007, SLUG and ZEB2 (p=0.04), and migration (MMP2, p=0.01); these genes were previously shown to be correlated with IGF1 [10]. Interestingly, the promoters of CYP1A1, VIM, SNAI2/SLUG contain a xenobiotic responsive element (DRE) sequence that when bound by AhR-ARNT heterodimers (canonical pathway) leads to their transcription. Activation of the AhR pathway by TCDD has been previously shown to enhance cancer cell invasion through metalloproteinases [60,61]. Environmental pollutants have been found to contribute to EMT and mesenchymal markers that provide invasion, migration and subsequent metastasis [61,62,63]. In contrast to diffuse GC, the low expression of CYP1A1 and CYP1B1 in intestinal GC inversely correlated with IDO1 expression (p<0.02). The IDO enzyme mediates the early steps of tryptophan metabolism leading to kynurenine, an endogenous AhR ligand produced in intestinal but not in diffuse GC [29]. Thus, environmental pollutants vs endogenous kynurenin may have differently effects on AhR-dependent gene expression in GCs.
Whether or not the increase of CYP1s expression that we observed in diffuse as compared to intestinal GC is due to exposure to a specific or to multiple POPs remains to be established [64]. We have previously reported the significant and widespread increase of a substantial set of POPs such as polychlorinated dioxins (PCDDs/PCDFs), polychlorobiphenyls (PCBs) and polybrominated flame retardants (such as PBDE 209, a carcinogenic intermediate of BaP) in human omental tissue (fat deposits) from French patients with diffuse-GC, as compared to control biopsies [13]. Coexposure of TCDD and PBDE209 was observed in 33% of the omentum from patients with diffuse GC. Interestingly, an increased incidence of hepatocellular carcinomas was observed in rodents upon exposure to PBDE-209 as well as an increased CYP1A1 mRNA expression levels in Caco-2 cells [65].
Expression of AhRR. Our results reveal that AhRR mRNA expression is strongly increased in the cohort of diffuse GC when compared to normal tissues of the same anatomical origin (x2.65, p=0.007) (Table 2). The effect of dioxin on Cyp1A1 and AhRR expression in the undifferentiated KATO III as compared to AGS gastric epithelial cells also support a role of xenobiotic compounds in vivo. Expression of AhRR correlated with Cyp1A1 (p=0.007), IGF1 (p<0.0001), and with genes involved in EMT (TWIST2, ZEB2) and migration (MMP2, p=0.002). Our observations in patients with metastatic diffuse GC further indicate a significant decrease of AhRR expression. Our results suggest that AhRR may represent an independent prognosis factor in diffuse GC, as we previously reported for breast cancer [12]. Poor prognosis was previously correlated with a decreased expression of AhRR in GCs from an Asian gastric cohort, but without discrimination between subtypes [66]. Moreover, loss of AhRR correlates with an aggressive tumorigenic phenotype in several tumors including colon, cervical, and ovarian carcinoma [67].
In conclusion, this pilot study explores two forms of GCs, diffuse- and intestinal- GC that lead to metastases in the peritoneal cavity. The induction of CYP1s through AhR activation may potentially serve as a biomarker for exposure to xenobiotics in diffuse GC. In vitro experiments indicate that TCDD strongly induce CYP1A1 in epithelial cells. Expression of Cyp1A1 strongly correlated with that of IGF, genes involved in EMT and migration. Increased expression of Cyp1B1 was observed in diffuse GC. Cyp1B1 activates a large number of pollutants which may result in activation of pro-cancer signaling pathways. AhR may contribute to the tumor-stroma interaction (through Cyp1A1 and Cyp1B1) in diffuse GC. Whether clinical factors such as smoking are prognostic factors remain to be investigated in GCs. We argue that reduction of exposure to subsets of environmental ligands could be important to prevent primary diffuse GC. A recent study revealed that exposure to environment pollutants such as POPs and BaP may reduce the efficacy of chemotherapy [68].
We acknowledge that our study has limitations. Because of the relatively low sample size in this report (n=29), certainly the results need to be confirmed using a larger cohort of gastric tumor samples with different clinical characteristics (including early and advanced stages). Nonetheless our pilot study shed light on the impact of AhR and related signaling pathays in Western patients with GCs. In addition, it will be interesting to extend the coverage to different geographical population settings. This will allow us to understand if the signaling pathways identified in GC subtypes are characteristic only in Western patients or can also be observed in patients from other geographical distributions. Further in vitro and in vivo studies with a larger cohort of gastric tumor samples will provide a better understanding of the complexity of the effect of different ligands on the regulation of the AhR pathway and may contribute to the development of novel clinically relevant agonists or antagonists.
As summarized in Figure 4, the present study provides new insights about the diversity of AhR functions in the development of cancer including GCs. It is likely that binding of various ligands is central to this carcinogenesis. The gastric epithelium is constantly exposed to exogenous AhR ligands such as dietary compounds and environmental toxins (PAH and other dioxin-like compounds), which enable to strong activation of AhR. Furthermore, the endogenous AhR kynurenine is produced through metabolism of tryptophan by IDO1 which is induced in stromal cells, or by TDO2 which can be up-regulated in tumor cells and in the tumor stroma. Our findings merit further studies with a larger cohort of gastric tumor samples with different clinical characteristics, including early and advanced tumor stages.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Relationship between AhR mRNA expression and clinical biological parameters in a series of 29 gastric cancers including diffuse- and intestinal subtypes.
Funding
This work was supported by INSERM (Institut National de la Santé et de la Recherche Medicale, Paris) and CNRS (Centre national de la Recherche Scientifique (MP-A).
Authors Contributions
Conceptualization: MP-A; Methodology: CP, SV, MP-A; Software: CP, M P-A, VB, Data curation: MP collected the clinical samples and data from patients. Writing original draft: MP-A; rewriting: VB. Funding acquisition: MP-A, MP; All authors have read and agreed to the published version of the manuscript.
Availability of data and materials
A total of 29 patients underwent partial gastrectomy for histopathologically-confirmed gastric adenocarcinoma primary tissue in the lariboisiere Hospital (Paris, France) from 2005-2014.
Ethics approval and consent to participate
The studies involving human participants were approved by the Ethical Commitee of Lariboisiere Hospital (Paris, France). The patients/participants provided their written informed consent to participate in scientific studies. Written informed consent was obtained from the participants before the collection of any samples, and the specimens were irreversibly de-identified.. All experiments involving the handling of human tissues were performed in accordance with Tenets of the Declaration of Helsinki. Biopsies from patients underwent partial gastrectomy for gastric adenocarcinoma tumor tissue from 2005-2014. The cohort of patients was previously described in Perrot-Applanat et al., Oncol Letters 2019, 18:674-686.
Patient consent for publication
The patients/participants provided their written informed consent to participate in this study and associated publications.
Acknowledgments
The authors greatly thank Matti Ullah and Mrs Iman Aldybiat (INSERM U1275, Lariboisiere Hospital), P Castagnet (Department of Pathology, Lariboisiere Hospital, Paris, France), Ivan Bieche (Department of Genetics, Pharmacogenomics Unit-Institut Curie, University of Paris Descartes-Paris5, Paris 75005). We also thank Peter Brooks for assistance in editing the English language of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study, in the collection, analyses, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef]
- Tan, P.; Yeoh, K.G. Genetics and Molecular Pathogenesis of Gastric Adenocarcinoma. Gastroenterology 2015, 149, 1153–1162. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Jass, J.R.; Sobin, L.H. Histological Classification of Gastric Tumours. In Histological typing of oesophageal and gastric tumors. World Health Organization. International Histological Classification of Tumours, Berlin, Heidelberg: Springer, 1990.
- Lauren, P. The Two Histological Main Types of Gastric Carcinoma: Diffuse and So-Called Intestinal-Type Carcinoma. An Attempt at a Histo-Clinical Classification. Acta Pathol. Microbiol. Scand. 1965, 64, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Piessen, G.; Messager, M.; Leteurtre, E.; Jean-Pierre, T.; Mariette, C. Signet ring cell histology is an independent predictor of poor prognosis in gastric adenocarcinoma regardless of tumoral clinical presentation. Ann. Surg. 2009, 250, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Piessen, G.; Messager, M.; Le Malicot, K.; Robb, W.B.; Di Fiore, F.; Guilbert, M.; Moreau, M.; Christophe, V.; Adenis, A.; Mariette, C. Phase II/III multicentre randomised controlled trial evaluating a strategy of primary surgery and adjuvant chemotherapy versus peri-operative chemotherapy for resectable gastric signet ring cell adenocarcinomas - PRODIGE 19 - FFCD1103 - ADCI002. BMC Cancer 2013, 13, 281. [Google Scholar] [CrossRef]
- Henson, D.E.; Dittus, C.; Younes, M.; Nguyen, H.; Albores-Saavedra, J. Differential trends in the intestinal and diffuse types of gastric carcinoma in the United States, 1973-2000: increase in the signet ring cell type. Arch Pathol Lab Med 2004, 128, 765–770. [Google Scholar] [CrossRef]
- Taghavi, S.; Jayarajan, S.N.; Davey, A.; Willis, A.I. Prognostic significance of signet ring gastric cancer. J. Clin. Oncol. 2012, 30, 3493–3498. [Google Scholar] [CrossRef]
- Perrot-Applanat, M.; Vacher, S.; Pimpie, C.; Chemlali, W.; Derieux, S.; Pocard, M.; Bieche, I. Differential gene expression in growth factors, epithelial mesenchymal transition and chemotaxis in the diffuse type compared with the intestinal type of gastric cancer. Oncol. Lett. 2019, 18, 674–686. [Google Scholar] [CrossRef]
- Pesatori, A.C.; Consonni, D.; Rubagotti, M.; Grillo, P.; Bertazzi, P.A. Cancer incidence in the population exposed to dioxin after the "Seveso accident": twenty years of follow-up. Environ. Health 2009, 8, 39. [Google Scholar] [CrossRef]
- Vacher, S.; Castagnet, P.; Chemlali, W.; Lallemand, F.; Meseure, D.; Pocard, M.; Bieche, I.; Perrot-Applanat, M. High AHR expression in breast tumors correlates with expression of genes from several signaling pathways namely inflammation and endogenous tryptophan metabolism. PLoS One 2018, 13, e0190619. [Google Scholar] [CrossRef] [PubMed]
- Perrot-Applanat, M.; Pimpie, C.; Cano-Sancho, G.; Antignac, J.P.; Pocard, M. Detection of Persistent Organic Pollutants in Omental Adipose Tissue from Patients with Diffuse-Gastric Cancer: A Pilot Study. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Denison, M.S.; Nagy, S.R. Activation of the aryl hydrocarbon receptor by structurally diverse exogenous and endogenous chemicals. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 309–334. [Google Scholar] [CrossRef] [PubMed]
- Denison, M.S.; Soshilov, A.A.; He, G.; DeGroot, D.E.; Zhao, B. Exactly the same but different: promiscuity and diversity in the molecular mechanisms of action of the aryl hydrocarbon (dioxin) receptor. Toxicol. Sci. 2011, 124, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Go, R.E.; Hwang, K.A.; Choi, K.C. Cytochrome P450 1 family and cancers. J. Steroid Biochem. Mol. Biol. 2015, 147, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Salguero, P.; Pineau, T.; Hilbert, D.M.; McPhail, T.; Lee, S.S.; Kimura, S.; Nebert, D.W.; Rudikoff, S.; Ward, J.M.; Gonzalez, F.J. Immune system impairment and hepatic fibrosis in mice lacking the dioxin-binding Ah receptor. Science 1995, 268, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Safe, S. Molecular biology of the Ah receptor and its role in carcinogenesis. Toxicol. Lett. 2001, 120, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Roman, A.C.; Carvajal-Gonzalez, J.M.; Rico-Leo, E.M.; Fernandez-Salguero, P.M. Dioxin receptor deficiency impairs angiogenesis by a mechanism involving VEGF-A depletion in the endothelium and transforming growth factor-beta overexpression in the stroma. J. Biol. Chem. 2009, 284, 25135–25148. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Cao, Z.; Wang, X. Role of aryl hydrocarbon receptor in cancer. Biochim. Biophys. Acta 2013, 1836, 197–210. [Google Scholar] [CrossRef]
- Quintana, F.J.; Sherr, D.H. Aryl hydrocarbon receptor control of adaptive immunity. Pharmacol. Rev. 2013, 65, 1148–1161. [Google Scholar] [CrossRef]
- Stockinger, B.; Di Meglio, P.; Gialitakis, M.; Duarte, J.H. The aryl hydrocarbon receptor: multitasking in the immune system. Annu. Rev. Immunol. 2014, 32, 403–432. [Google Scholar] [CrossRef] [PubMed]
- Litzenburger, U.M.; Opitz, C.A.; Sahm, F.; Rauschenbach, K.J.; Trump, S.; Winter, M.; Ott, M.; Ochs, K.; Lutz, C.; Liu, X.; et al. Constitutive IDO expression in human cancer is sustained by an autocrine signaling loop involving IL-6, STAT3 and the AHR. Oncotarget 2014, 5, 1038–1051. [Google Scholar] [CrossRef] [PubMed]
- Andersson, P.; McGuire, J.; Rubio, C.; Gradin, K.; Whitelaw, M.L.; Pettersson, S.; Hanberg, A.; Poellinger, L. A constitutively active dioxin/aryl hydrocarbon receptor induces stomach tumors. Proc. Natl. Acad. Sci. U S A 2002, 99, 9990–9995. [Google Scholar] [CrossRef]
- Andersson, P.; Rubio, C.; Poellinger, L.; Hanberg, A. Gastric hamartomatous tumours in a transgenic mouse model expressing an activated dioxin/Ah receptor. Anticancer. Res. 2005, 25, 903–911. [Google Scholar] [PubMed]
- Brunnberg, S.; Andersson, P.; Lindstam, M.; Paulson, I.; Poellinger, L.; Hanberg, A. The constitutively active Ah receptor (CA-Ahr) mouse as a potential model for dioxin exposure--effects in vital organs. Toxicology 2006, 224, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Labib, S.; Guo, C.H.; Williams, A.; Yauk, C.L.; White, P.A.; Halappanavar, S. Toxicogenomic outcomes predictive of forestomach carcinogenesis following exposure to benzo(a)pyrene: relevance to human cancer risk. Toxicol. Appl. Pharmacol. 2013, 273, 269–280. [Google Scholar] [CrossRef]
- Peng, T.L.; Chen, J.; Mao, W.; Song, X.; Chen, M.H. Aryl hydrocarbon receptor pathway activation enhances gastric cancer cell invasiveness likely through a c-Jun-dependent induction of matrix metalloproteinase-9. BMC Cell Biol. 2009, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- Perrot-Applanat, M.; Pimpie, C.; Vacher, S.; Bieche, I.; Pocard, M.; Baud, V. Differential Expression of Genes Involved in Metabolism and Immune Response in Diffuse and Intestinal Gastric Cancers, a Pilot Ptudy. Biomedicines 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Washington, K. 7th edition of the AJCC cancer staging manual: stomach. Ann. Surg. Oncol. 2010, 17, 3077–3079. [Google Scholar] [CrossRef]
- Sano, T.; Coit, D.G.; Kim, H.H.; Roviello, F.; Kassab, P.; Wittekind, C.; Yamamoto, Y.; Ohashi, Y. Proposal of a new stage grouping of gastric cancer for TNM classification: International Gastric Cancer Association staging project. Gastric Cancer 2017, 20, 217–225. [Google Scholar] [CrossRef]
- Shu, P.; Qin, J.; Shen, K.; Chen, W.; Liu, F.; Fang, Y.; Wang, X.; Wang, H.; Shen, Z.; Sun, Y.; et al. The IGCA staging system is more accurate than AJCC7 system in stratifying survival of patients with gastric cancer in stage III. BMC Cancer 2017, 17, 238. [Google Scholar] [CrossRef]
- Ji, X.; Bu, Z.D.; Yan, Y.; Li, Z.Y.; Wu, A.W.; Zhang, L.H.; Zhang, J.; Wu, X.J.; Zong, X.L.; Li, S.X.; et al. The 8th edition of the American Joint Committee on Cancer tumor-node-metastasis staging system for gastric cancer is superior to the 7th edition: results from a Chinese mono-institutional study of 1663 patients. Gastric Cancer, 1007. [Google Scholar] [CrossRef]
- Bieche, I.; Onody, P.; Laurendeau, I.; Olivi, M.; Vidaud, D.; Lidereau, R.; Vidaud, M. Real-time reverse transcription-PCR assay for future management of ERBB2-based clinical applications. Clin. Chem. 1999, 45, 1148–1156. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.F.; Chen, J.; Mao, W.; Wang, Y.H.; Chen, M.H. Downregulation of aryl hydrocarbon receptor expression decreases gastric cancer cell growth and invasion. Oncol. Rep. 2013, 30, 364–370. [Google Scholar] [CrossRef]
- Nam, S.; Lee, Y.; Kim, J.H. RHOA protein expression correlates with clinical features in gastric cancer: a systematic review and meta-analysis. BMC Cancer 2022, 22, 798. [Google Scholar] [CrossRef] [PubMed]
- Kakiuchi, M.; Nishizawa, T.; Ueda, H.; Gotoh, K.; Tanaka, A.; Hayashi, A.; Yamamoto, S.; Tatsuno, K.; Katoh, H.; Watanabe, Y.; et al. Recurrent gain-of-function mutations of RHOA in diffuse-type gastric carcinoma. Nat. Genet. 2014, 46, 583–587. [Google Scholar] [CrossRef]
- Ridley, A.J. RhoA, RhoB and RhoC have different roles in cancer cell migration. J. Microsc. 2013, 251, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Guo, Y.; Xu, B. Expressions of Ras Homolog Gene Family, Member A (RhoA) and Cyclooxygenase-2 (COX-2) Proteins in Early Gastric Cancer and Their Role in the Development of Gastric Cancer. Med. Sci. Monit. 2017, 23, 2979–2984. [Google Scholar] [CrossRef] [PubMed]
- Lawson, C.D.; Ridley, A.J. Rho GTPase signaling complexes in cell migration and invasion. J. Cell Biol. 2018, 217, 447–457. [Google Scholar] [CrossRef]
- Yoon, J.H.; Choi, W.S.; Kim, O.; Choi, B.J.; Nam, S.W.; Lee, J.Y.; Park, W.S. Gastrokine 1 inhibits gastric cancer cell migration and invasion by downregulating RhoA expression. Gastric Cancer 2017, 20, 274–285. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research, N. Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014, 513, 202–209. [Google Scholar] [CrossRef]
- Wang, R.; Song, S.; Harada, K.; Ghazanfari Amlashi, F.; Badgwell, B.; Pizzi, M.P.; Xu, Y.; Zhao, W.; Dong, X.; Jin, J.; et al. Multiplex profiling of peritoneal metastases from gastric adenocarcinoma identified novel targets and molecular subtypes that predict treatment response. Gut 2020, 69, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Murray, G.I.; Taylor, M.C.; McFadyen, M.C.; McKay, J.A.; Greenlee, W.F.; Burke, M.D.; Melvin, W.T. Tumor-specific expression of cytochrome P450 CYP1B1. Cancer Res. 1997, 57, 3026–3031. [Google Scholar] [PubMed]
- Murray, G.I.; Taylor, M.C.; Burke, M.D.; Melvin, W.T. Enhanced expression of cytochrome P450 in stomach cancer. Br. J. Cancer 1998, 77, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Hayes, C.L.; Yamazaki, H.; Amin, S.; Hecht, S.S.; Guengerich, F.P.; Sutter, T.R. Activation of chemically diverse procarcinogens by human cytochrome P-450 1B1. Cancer Res. 1996, 56, 2979–2984. [Google Scholar] [PubMed]
- Buters, J.T.; Mahadevan, B.; Quintanilla-Martinez, L.; Gonzalez, F.J.; Greim, H.; Baird, W.M.; Luch, A. Cytochrome P450 1B1 determines susceptibility to dibenzo[a,l]pyrene-induced tumor formation. Chem. Res. Toxicol. 2002, 15, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Shiizaki, K.; Kawanishi, M.; Yagi, T. Modulation of benzo[a]pyrene-DNA adduct formation by CYP1 inducer and inhibitor. Genes. Environ. 2017, 39, 14. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Snyder, M.; Kenison, J.E.; Yang, K.; Lara, B.; Lydell, E.; Bennani, K.; Novikov, O.; Federico, A.; Monti, S.; et al. How the AHR Became Important in Cancer: The Role of Chronically Active AHR in Cancer Aggression. Int. J. Mol. Sci. 2020, 22. [Google Scholar] [CrossRef] [PubMed]
- Nebert, D.W.; Shi, Z.; Galvez-Peralta, M.; Uno, S.; Dragin, N. Oral benzo[a]pyrene: understanding pharmacokinetics, detoxication, and consequences--Cyp1 knockout mouse lines as a paradigm. Mol. Pharmacol. 2013, 84, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, X.; Wang, Y.; Bai, P.; Hayakawa, K.; Zhang, L.; Tang, N. Characteristics and Influencing Factors of Polycyclic Aromatic Hydrocarbons Emitted from Open Burning and Stove Burning of Biomass: A Brief Review. Int. J. Environ. Res. Public. Health 2022, 19. [Google Scholar] [CrossRef]
- Dere, E.; Lo, R.; Celius, T.; Matthews, J.; Zacharewski, T.R. Integration of genome-wide computation DRE search, AhR ChIP-chip and gene expression analyses of TCDD-elicited responses in the mouse liver. BMC Genomics 2011, 12, 365. [Google Scholar] [CrossRef]
- Eltom, S.E.; Larsen, M.C.; Jefcoate, C.R. Expression of CYP1B1 but not CYP1A1 by primary cultured human mammary stromal fibroblasts constitutively and in response to dioxin exposure: role of the Ah receptor. Carcinogenesis 1998, 19, 1437–1444. [Google Scholar] [CrossRef]
- Sparfel, L.; Pinel-Marie, M.L.; Boize, M.; Koscielny, S.; Desmots, S.; Pery, A.; Fardel, O. Transcriptional signature of human macrophages exposed to the environmental contaminant benzo(a)pyrene. Toxicol. Sci. 2010, 114, 247–259. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Baek, H.S.; Ye, D.J.; Shin, S.; Kim, D.; Chun, Y.J. CYP1B1 Enhances Cell Proliferation and Metastasis through Induction of EMT and Activation of Wnt/beta-Catenin Signaling via Sp1 Upregulation. PLoS ONE 2016, 11, e0151598. [Google Scholar] [CrossRef]
- Mohamed, H.T.; Gadalla, R.; El-Husseiny, N.; Hassan, H.; Wang, Z.; Ibrahim, S.A.; El-Shinawi, M.; Sherr, D.H.; Mohamed, M.M. Inflammatory breast cancer: Activation of the aryl hydrocarbon receptor and its target CYP1B1 correlates closely with Wnt5a/b-beta-catenin signalling, the stem cell phenotype and disease progression. J. Adv. Res. 2019, 16, 75–86. [Google Scholar] [CrossRef]
- Ohta, H.; Aoyagi, K.; Fukaya, M.; Danjoh, I.; Ohta, A.; Isohata, N.; Saeki, N.; Taniguchi, H.; Sakamoto, H.; Shimoda, T.; et al. Cross talk between hedgehog and epithelial-mesenchymal transition pathways in gastric pit cells and in diffuse-type gastric cancers. Br. J. Cancer 2009, 100, 389–398. [Google Scholar] [CrossRef]
- Katoh, M. Genomic testing, tumor microenvironment and targeted therapy of Hedgehog-related human cancers. Clin Sci (Lond) 2019, 133, 953–970. [Google Scholar] [CrossRef]
- Weitzman, J.B. Co-regulation of mouse genes predicts function. J. Biol. 2004, 3, 19. [Google Scholar] [CrossRef]
- Ishida, M.; Mikami, S.; Kikuchi, E.; Kosaka, T.; Miyajima, A.; Nakagawa, K.; Mukai, M.; Okada, Y.; Oya, M. Activation of the aryl hydrocarbon receptor pathway enhances cancer cell invasion by upregulating the MMP expression and is associated with poor prognosis in upper urinary tract urothelial cancer. Carcinogenesis 2010, 31, 287–295. [Google Scholar] [CrossRef]
- Barouki, R.; Coumoul, X. Cell migration and metastasis markers as targets of environmental pollutants and the Aryl hydrocarbon receptor. Cell Adh Migr. 2010, 4, 72–76. [Google Scholar] [CrossRef]
- Otsuki, S.; Inokuchi, M.; Enjoji, M.; Ishikawa, T.; Takagi, Y.; Kato, K.; Yamada, H.; Kojima, K.; Sugihara, K. Vimentin expression is associated with decreased survival in gastric cancer. Oncol. Rep. 2011, 25, 1235–1242. [Google Scholar] [CrossRef]
- Ochieng, J.; Nangami, G.N.; Ogunkua, O.; Miousse, I.R.; Koturbash, I.; Odero-Marah, V.; McCawley, L.J.; Nangia-Makker, P.; Ahmed, N.; Luqmani, Y.; et al. The impact of low-dose carcinogens and environmental disruptors on tissue invasion and metastasis. Carcinogenesis 2015, 36 Suppl. 1, S128–159. [Google Scholar] [CrossRef]
- Lagunas-Rangel, F.A.; Linnea-Niemi, J.V.; Kudlak, B.; Williams, M.J.; Jonsson, J.; Schioth, H.B. Role of the Synergistic Interactions of Environmental Pollutants in the Development of Cancer. Geohealth 2022, 6, e2021GH000552. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yuan, J.; Che, S.; Zhang, L.; Ruan, Z.; Sun, X. Decabromodiphenyl ether induces ROS-mediated intestinal toxicity through the Keap1-Nrf2 pathway. J. Biochem. Mol. Toxicol. 2022, 36, e22995. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Wang, D.D.; Zhao, B.W.; Wang, W.; Yuan, S.Q.; Huang, C.Y.; Chen, Y.M.; Zheng, Y.; Keshari, R.P.; Xia, J.C.; et al. Poor prognosis of gastric adenocarcinoma with decreased expression of AHRR. PLoS One 2012, 7, e43555. [Google Scholar] [CrossRef] [PubMed]
- Zudaire, E.; Cuesta, N.; Murty, V.; Woodson, K.; Adams, L.; Gonzalez, N.; Martinez, A.; Narayan, G.; Kirsch, I.; Franklin, W.; et al. The aryl hydrocarbon receptor repressor is a putative tumor suppressor gene in multiple human cancers. J. Clin. Investig. 2008, 118, 640–650. [Google Scholar] [CrossRef]
- Lagunas-Rangel, F.A.; Liu, W.; Schioth, H.B. Can Exposure to Environmental Pollutants Be Associated with Less Effective Chemotherapy in Cancer Patients? Int. J. Environ. Res. Public. Health 2022, 19. [Google Scholar] [CrossRef]
Figure 1.
Immunohistochemical staining AhR and Cyp1B1 in peritumoral and diffuse GCs. AhR in peritumoral gastric tissue (A); weak cytoplasmic and/or nuclear staining were observed in glandular tissue and stroma. In tumoral tissue (B and D), strong AhR immunostaining is observed in most cells, both epithelial and stromal compartments. Cyp1B1 (C) and AhR (D) immunostaining are shown on the same tumor (diffuse GC). Cyp1B1 was mainly observed in the stromal compartment in diffuse GC (C). Original magnification x20. Bar scale, 500 μm.
Figure 1.
Immunohistochemical staining AhR and Cyp1B1 in peritumoral and diffuse GCs. AhR in peritumoral gastric tissue (A); weak cytoplasmic and/or nuclear staining were observed in glandular tissue and stroma. In tumoral tissue (B and D), strong AhR immunostaining is observed in most cells, both epithelial and stromal compartments. Cyp1B1 (C) and AhR (D) immunostaining are shown on the same tumor (diffuse GC). Cyp1B1 was mainly observed in the stromal compartment in diffuse GC (C). Original magnification x20. Bar scale, 500 μm.
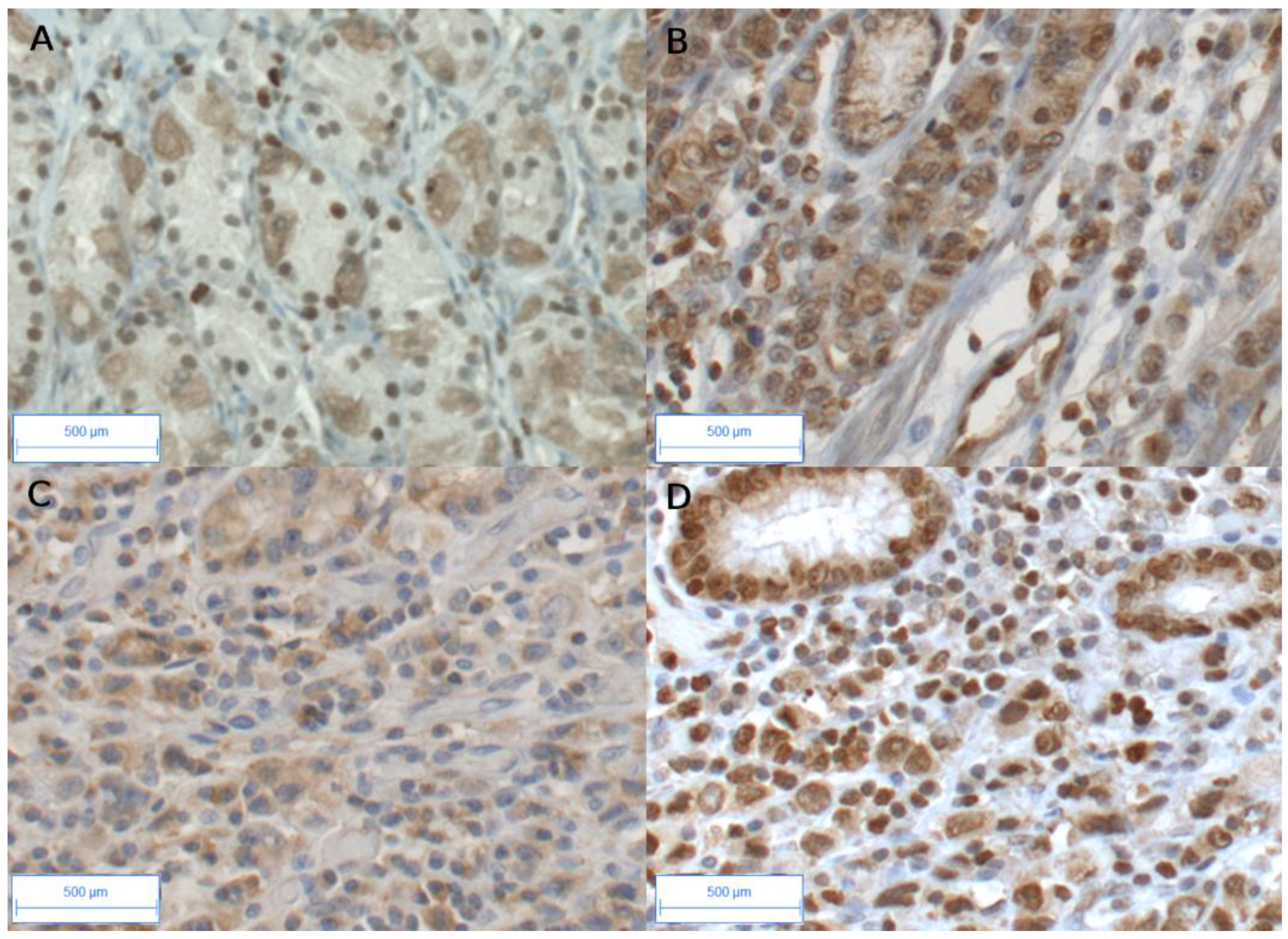
Figure 2.
mRNA expression levels of AhR, ARNT, Cyp1A1, IL1B and UGT1A1 in KATO III gastric cells treated with two different ligands. KATO III cells were cultivated in the absence (Ctrl) or presence of TCDD (2,3,7,8-tetrachlorodibenzo-p-dioxin i.e., dioxin, 30 nM) or BaP (benzo[a]pyrene) (10µM) for 16h. Cells were incubated with (grey column) or without (black column) CHH22319. Expression of genes AhR (a), ARNT (b), AhRR (c), CYP1A1 (d), IL1β (e) and UGT1A1 (f) were determined by qRT-PCR. All experiments were performed in triplicate. Results were expressed as means +/- S.E.M and normalized so that the mean of the control cells was 1. Three levels of statistical significance are distinguished: **p-value<0.04; ***p-value<0.001, ****p-value<0.0001.
Figure 2.
mRNA expression levels of AhR, ARNT, Cyp1A1, IL1B and UGT1A1 in KATO III gastric cells treated with two different ligands. KATO III cells were cultivated in the absence (Ctrl) or presence of TCDD (2,3,7,8-tetrachlorodibenzo-p-dioxin i.e., dioxin, 30 nM) or BaP (benzo[a]pyrene) (10µM) for 16h. Cells were incubated with (grey column) or without (black column) CHH22319. Expression of genes AhR (a), ARNT (b), AhRR (c), CYP1A1 (d), IL1β (e) and UGT1A1 (f) were determined by qRT-PCR. All experiments were performed in triplicate. Results were expressed as means +/- S.E.M and normalized so that the mean of the control cells was 1. Three levels of statistical significance are distinguished: **p-value<0.04; ***p-value<0.001, ****p-value<0.0001.
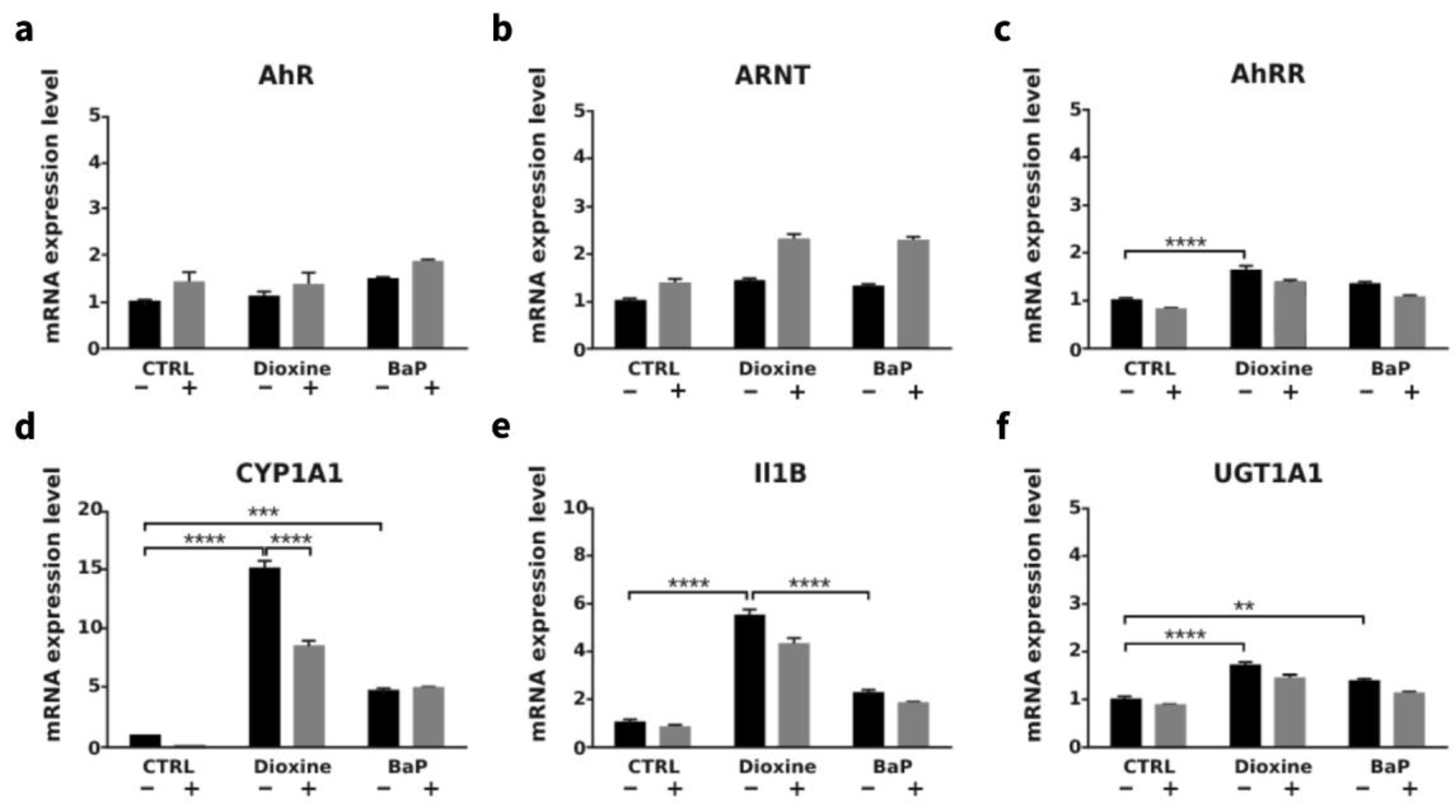
Figure 3.
mRNA expression levels of AhR, Cyp1A1, Cyp1B1 and AhRR in gastric epithelial cancer cells treated with TCDD. Figure 3A: Kato III cells were cultivated in the presence or absence of dioxin. Expression levels of Cyp1A1 and Cyp1B1 were determined by qRT-PCR in the same experiment. Results were expressed as arbitrary values (means +/- S.E.M) vs CT35 for Cyp1A1 and CYP1B1; basal level of CYP1B1 was undetectable in AGS cells (CT>50). Figure 3B: AGS cells were cultivated in the absence (Ctrl in black) or presence of TCDD (2,3,7,8-tetrachlorodibenzo-p-dioxin) (0.01-10 nM, in grey). Expression levels of AhR, CYP1A1, AhRR were determined by qRT-PCR in the same experiment All experiments were performed in triplicate. Results were expressed as means +/- S.E.M and normalized so that the mean of the control cells was 1 (Fig. B a-c). ***p-value<0.001.
Figure 3.
mRNA expression levels of AhR, Cyp1A1, Cyp1B1 and AhRR in gastric epithelial cancer cells treated with TCDD. Figure 3A: Kato III cells were cultivated in the presence or absence of dioxin. Expression levels of Cyp1A1 and Cyp1B1 were determined by qRT-PCR in the same experiment. Results were expressed as arbitrary values (means +/- S.E.M) vs CT35 for Cyp1A1 and CYP1B1; basal level of CYP1B1 was undetectable in AGS cells (CT>50). Figure 3B: AGS cells were cultivated in the absence (Ctrl in black) or presence of TCDD (2,3,7,8-tetrachlorodibenzo-p-dioxin) (0.01-10 nM, in grey). Expression levels of AhR, CYP1A1, AhRR were determined by qRT-PCR in the same experiment All experiments were performed in triplicate. Results were expressed as means +/- S.E.M and normalized so that the mean of the control cells was 1 (Fig. B a-c). ***p-value<0.001.
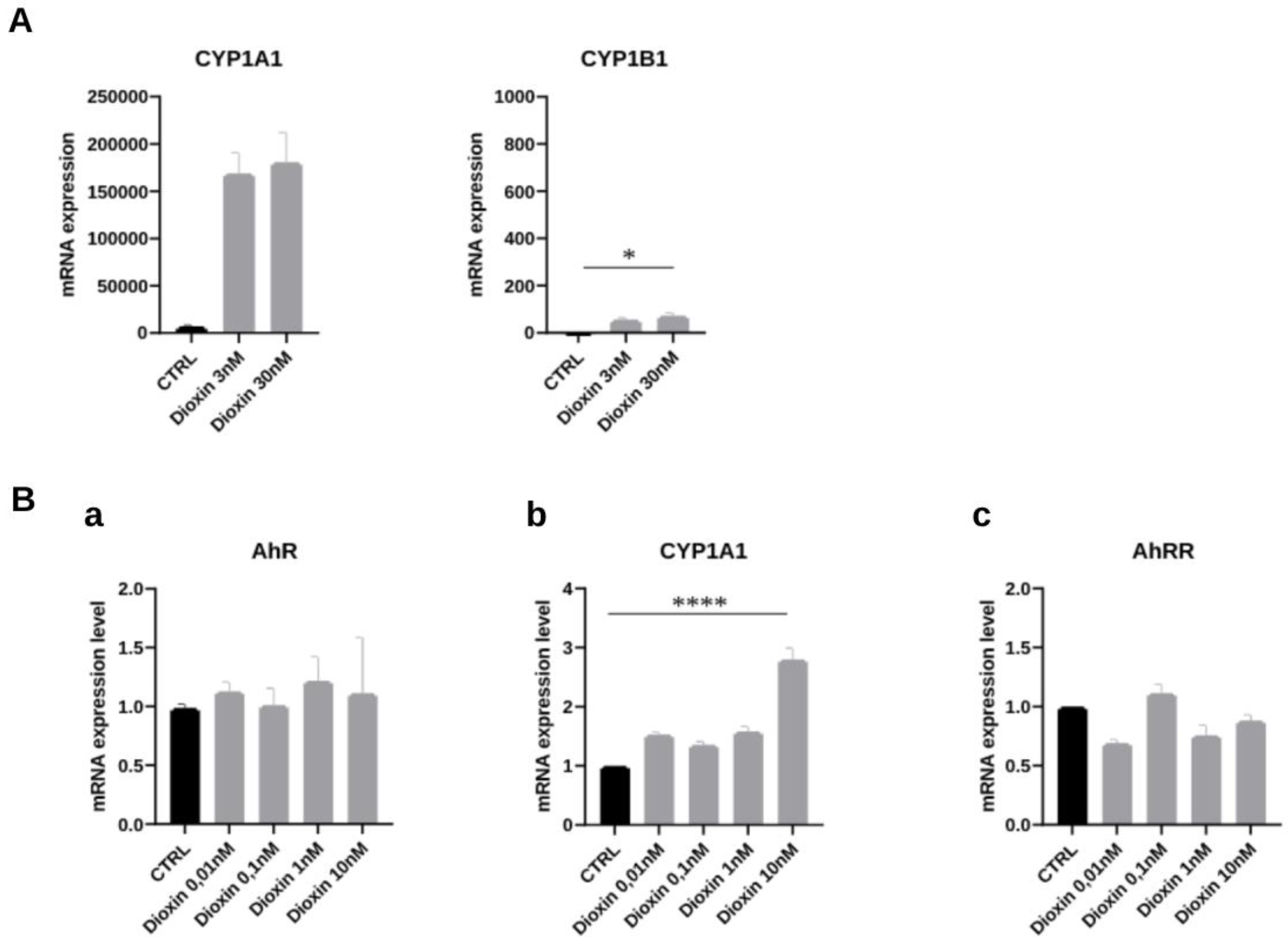
Figure 4.
AhR role in cancer biology; environmental compounds at the crossroads of toxicity and several signaling pathways.
Figure 4.
AhR role in cancer biology; environmental compounds at the crossroads of toxicity and several signaling pathways.
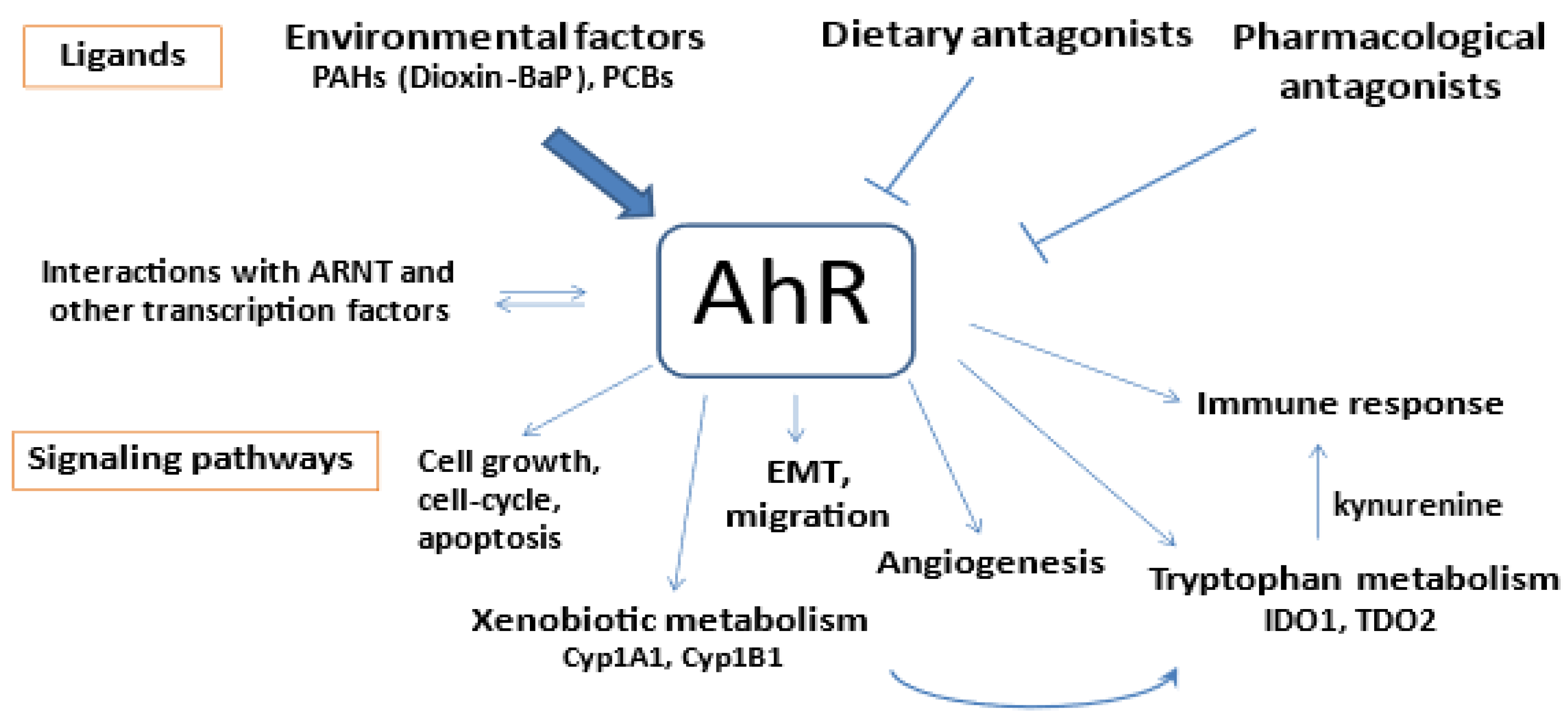

Table 1.
Clinicopathological characteristics of gastric carcinoma patients accordingly with histopathological subtype.
Table 1.
Clinicopathological characteristics of gastric carcinoma patients accordingly with histopathological subtype.
| Total GC (n=29) | Diffuse/poorly cohesive GC* (n=13) (45%) | Intestinal-subtype GC** (n=16) (55%) |
P-value a | |
|---|---|---|---|---|
| Gender, n (%) Male Female |
13/29 16/29 |
6/13 (46%) 7/13 (54%) |
7/16 (43%) 9/16 (56%) |
0.90 (NS) |
| Age (years, median) | 63 +/-17 | 57 (27-71) | 75 (59-82) | 0.0004 b |
| Smoking Negative Positive |
12/22 10/22 |
4/12 3/10 |
8/12 7/10 |
0.77 (NS) |
| Tumor size (mm), n (%) <50 >=50 |
10/27 17/27 |
4/11 (36 %) 7/11 (64 %) |
6/16 (37 %) 10/16 (63 %) |
0.10 (NS) b 0.95 (NS) |
| Depth tumor invasion (T) T1-T2 T3-T4 |
6/29 23/29 |
2/13 (15%) 11/13 (85%) |
4/16 (33 %) 12/16 (67 %) |
0.66 (NS) |
| Lymphatic invasion (N) Negative Positive |
11/28 17/28 |
1/13 (7%) 12/13 (92%) |
10/15 (67 %) 5/15 (33%) |
0.0014 |
| Metastasis (M), n (%) Negative Positive |
24/29 5/29 |
9/13 (69%) 4/13 (31%) |
15/16 (94%) 1/16 (6%) |
0.14 (NS) |
| TNM status I-II III-IV |
16/29 13/29 |
5/13 (38.5%) 8/13 (61.5%) |
11/16 (69%) 5/16 (31%) |
0.10 (NS) |
| Vascular invasion, n + (%) Negative Positive |
9/29 20/29 |
3/13 (23%) 10/13 (77%) |
6/16 (38%) 10/16 (62%) |
0.67 (NS) |
| Neural invasion, n (%) Negative Positive |
23/29 6/29 |
2/13 (15%) 11/13 (68%) |
4/16 (25%) 12/16 (75%) |
0.66 (NS) |
*poorly cohesive adenocarcinoma/diffuse-type carcinoma; ** intestinal-type adenocarcinoma. a, Chi-square test, Yates’ continuity corrected chi-square test or Fisher’s exact test if appropriate; b, Mann Whitney. NS= not statistically different.
Table 2.
Statistical analysis of mRNA expression levels of AhR, CYP1B1, CYP1A1 and AhRR in gastric cancers (all GC, diffuse- and intestinal-subtypes).
Table 2.
Statistical analysis of mRNA expression levels of AhR, CYP1B1, CYP1A1 and AhRR in gastric cancers (all GC, diffuse- and intestinal-subtypes).
| Genes | nontumoral gastric tissues (n=11) | All tumors (n=29) | p-valuea | Diffuse-GC vs PT (n=13) |
p-valuea | Intestinal-GC vs PT (n=16) |
p-valuea | p-value (GC sub-types) |
|---|---|---|---|---|---|---|---|---|
| AhR and target genes | ||||||||
| AhR | 1 (0.37-1.64) | 1.94 (0.55-3.53) | 0.002 | 2.12 (0.55-3.35) | 0.001 | 1.60 (0.65-3.53) | 0.003 | 0.13 (NS) |
| CYP1B1 | 1 (0.52-2.90) | 1.45 (0.13-4.90) | 0.91 (NS) | 1.62 (0.43-4.90) | 0.014 | 1.22 (0.13-4.0) | 0.82 (NS) | 0.19 (NS) |
| CYP1A1* | 0 (0-5.6) | 1.37 (0-86) | 0.26 (NS) | 0.43 (0-30) | 0.73 (NS) | NS | ||
| AhRR | 1 (0.23-1.66) | 1.25 (0.19-3.93) | >0.999 (NS) | 2.65 (0.74-3.96) | 0.007 | 0.89 (0.19-3.85) | 0.88 (NS) | 0.017 |
Median (range) of gene mRNA expression levels; p value (Mann Whitney). Significant p value in bold. Comparative basal levels of genes in normal gastric tissue (x1) are as follows: AhR (71), Cyp1A1 (0), Cyp1B1 (372), AhRR (21). NS= not statistically different.
Table 3.
Relationship between CYP1B1, CYP1A1 and AhRR expression with clinical parameters in gastric adenocarcinomas (including sub-populations).
Table 3.
Relationship between CYP1B1, CYP1A1 and AhRR expression with clinical parameters in gastric adenocarcinomas (including sub-populations).
| All tumors, n=29 | Diffuse sub-type GC, n=13 | Intestinal sub-type GC, n=16 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| AhRR | Cyp1B1 | Cyp1A1 | n= | AhRR | Cyp1B1 | Cyp1A1 | n= | AhRR | Cyp1B1 | Cyp1A1 | |
|
Gender Male (n=13) Female (n=16) |
p= 0.75 1.63 (0.29-3.96) 1.18 (0.29-3.85) |
p= 0.47 1.62 (0.13-4.03) 1.34 (0.32-4.9) |
p= 0.063 1.37 (0-30.1) 0 (0-86) |
6 7 |
p= 0.42 2.2 (1.23-3.96) 2.65 (0.74-3.53) |
p= 0.92 1.76 (1.15-2.32) 1.47 (0.43-4.9) |
p= 0.83 1.38 (0-4.01) 0 (0-86) |
7 9 |
p= 0.88 0.93 (0.29-2.38) 0.85 (0.19-3.85) |
p= 0.67 1.45 (0.13-4) 1.16 (0.32-2.5) |
p= 0.058 0.76 (0-30.1) 0 (0-1.39) |
|
Age <60 years (n=9) >60 years(n=20) |
p= 0.27 1.66 (0.74-3.96) 1.17 (0.19-3.85) |
p= 0.18 1.47 (1.23-4.9) 1.21 (0.13-4) |
p= 0.97 0.67 (0-30.1) 0.65 (0-86) |
8 5 |
p= 0.27 1.64 (0.74-3.96) 3.17 (1.15-3.66) |
p= 0.49 1.68 (1.23-4.9) 1.62 (0.43-2.44) |
p= 0.16 0.33 (0-4.01) 1.51 (0-86) |
1 15 |
ND 2.38 0.85 (0.19-3.85) |
ND 1.45 1.16 (0.13-4) |
ND 30.1 0.25 (0-3.31) |
|
Smoking Negative (n=12) Positive (n=10) |
p= 0.54 1.21 (0.42-1.67) 1.65 (0.29-3.85) |
p= 0.34 1.63 (0.36-4) 0.98 (0.13-5) |
p= 0.42 0.33 (0-1.98) 0.43 (0-86) |
4 3 |
p= 0.46 1.43 (0.74-1.66) 3.17 (0.74-3.53) |
p= 0.23 1.78 (1.31-2.32) 2.44 (1.85-4.9) |
p= 0.40 0.33 (0-1.37) 62.55 (0-86) |
8 7 |
p= 0.84 1.02 (0.42-1.67) 0.93 (0.29-3.85) |
p= 0.054 1.63 (0.36-4) 0.49 (0.13-3) |
p= 0.86 0.38 (0-1.98) 0.25 (0-30.1) |
|
Tumor invasion (T) T1-T2 (n=6) T3-T4 (n=23) |
p= 0.11 0.43 (0.19-3.5) 1.31 (0.42-3.87) |
p= 0.18 0.94 (0.13-2.44) 1.47 (0.32-4.9) |
p= 0.63 0.47 (0-86) 0.67 (0-30.1) |
2 11 |
ND 3.35 (3.17-3.53) 1.66 (0.74-3.96) |
ND 2.14 (1.85-2.44) 1.47 (0.43-4.9) |
ND 74.3 (62.5-86) 0.67 (0-4.01) |
4 12 |
p= 0.002 0.31 (0.19-0.53) 1.22 (0.42-3.85) |
p= 0.055 0.46 (0.13-1.2) 1.52 (0.32-4) |
p= 0.36 0.12 (0-0.7) 0.68 (0-30.1) |
|
Lymphatic invasion negative (n=11) positive (n=17) |
p= 0.032 0.74 (0.19-3.85) 1.67 (0.42-3.96) |
p= 0.014 0.5 (0.3-2.5) 1.6 (0.4-4.9) |
p= 0.06 0 (0-1.39) 1 (0-86) |
1 12 |
ND 1.63 2.7 (0.74-3.96) |
ND 2.32 1.54 (0.43-4.9) |
ND 1.37 1.03 (0-86) |
10 5 |
p= 0.25 0.73 (0.19-3.85) 1.31 (0.42-2.38) |
p= 0.019 0.52 (0.13-2.5) 1.59 (1.27-4) |
p= 0.04 0 (0-1.39) 1 (0-30.1) |
|
Metastasis (M) negative (n=24) positive (n=5) |
p= 0.59 1.28 (0.19-4.0) 1.15 (0.74-2.38) |
p= 0.55 1.39 (0.13-4) 1.45 (0.43-4.9) |
p= 0.59 0.68 (0-86) 0 (0-30.1) |
9 4 |
p= 0.006 3.17 (1.23-3.96) 0.94 (0.74-1.63) |
p= 0.82 1.62 (1.15-2.44) 1.86 (0.43-4.9) |
p= 0.06 1.51 (0-86) 0 (0-1.37) |
15 1 |
ND 0.85 (0.19-3.85) 2.38 |
ND 1.16 (0.13-4) 1.45 |
ND 0.25 (0-3.31) 30.1 |
|
TNM I-II (n=16) III-IV (n=13) |
p= 0.35 1.06 (0.19-3.96) 1.63 (0.42-3.66) |
p= 0.045 1.19 (0.13-3) 1.62 (0.43-4.9) |
p= 0.10 0.12 (0-4.01) 1.37 (0-86) |
5 8 |
p= 0.50 2.65 (1.23-3.96) 2.19 (0.74-3.66) |
p= 0.68 1.47 (1.23-2.17) 1.73 (0.43-4.9) |
p= 0.83 0.67 (0-4.01) 1.38 (0-86) |
11 5 |
p= 0.22 0.74 (0.19-3.85) 1.31 (0.42-2.38) |
p= 0.038 0.52 (0.13-3) 1.59 (1.27-4) |
p= 0.097 0 (0-3.31) 1 (0-30.1) |
|
Vascular invasion, negative (n=9) positive (n=20) |
p=0.09 0.74 (0.2-4) 1.65 (0.3-3.8) |
p= 0.39 1.27 (0.36-2.32) 1.54 (0.13-4.9) |
p= 0.45 0.61 (0-4.01) 0.71 (0-86) |
3 10 |
p= 0.84 1.63 (0.74-3.96) 2.70 (0.74-3.66) |
p= 0.57 1.9 (1.4-2.32) 1.54 (0.43-4.9) |
p>0.9999 1.37 (0-4.01) 1.03 (0-86) |
6 10 |
p= 0.17 0.62 (0.19-1.31) 1.09 (0.29-3.85) |
p= 0.53 0.94 (0.36-1.6) 1.56 (0.13-4) |
p= 0.41 0.30 (0-1) 0.5 (0-30.1) |
|
Neural invasion negative (n=6) positive (n=23) |
p= 0.38 0.79 (0.19-3.53) 1.31 (0.29-3.96) |
p= 0.41 1.76 (0.72-2.51) 1.4 (0.13-4.9) |
p= 0.38 1.04 (0-86) 0.61 (0-30.1) |
2 11 |
ND 3.35 (3.17-3.53) 1.66 (0.74-3.96) |
ND 2.14 (1.85-2.44) 1.47 (0.43-4.9) |
ND 74.3 (62.5-85.9) 0.67 (0-4.01) |
4 12 |
p= 0.12 0.63 (0.19-0.85) 1.22 (0.29-3.85) |
p= 0.52 1.41 (0.72-2.5) 0.89 (0.13-4) |
p= 0.76 0.35 (0-1.39) 0.43 (0-30.1) |
Median (range) of gene mRNA expression levels; p value (Mann Whitney). Significant p value in bold; tendancy in italic. ND= not determined.
Table 4.
Correlations of selected genes analysed in the study in relation to AhR and “AhR-related signalling pathways” in subtypes GC.
Table 4.
Correlations of selected genes analysed in the study in relation to AhR and “AhR-related signalling pathways” in subtypes GC.
| Diffuse GC | Intestinal GC | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genes | AHR | AHR | AHRR | AHRR | CYP1B1 | CYP1B1 | CYP1A1 | CYP1A1 | AHR | AHR | AHRR | AHRR | CYP1B1 | CYP1B1 | CYP1A1 | CYP1A1 |
| r | P value | r | P value | r | P value | r | P value | r | P value | r | P value | r | P value | r | P value | |
| AHR | x | -0.031 | 0.92 | -0.066 | 0.83 | 0.181 | 0.553 | x | 0.679 | 0.004 | 0.018 | 0.95 | 0.112 | 0.68 | ||
| AHRR | -0.031 | 0.92 | x | -0.107 | 0.73 | 0.711 | 0.007 | 0.679 | 0.004 | x | 0.253 | 0.34 | 0.241 | 0.368 | ||
| CYP1B1 | -0.066 | 0.83 | -0.107 | 0.73 | x | 0.342 | 0.253 | 0.018 | 0.95 | 0.253 | 0.34 | x | 0.655 | 0.006 | ||
| Growth factors and receptors (n=10) | ||||||||||||||||
| IGF1 | 0.184 | 0.55 | 0.865 | 0.0001 | 0.155 | 0.71 | 0.846 | 0.001 | -0.113 | 0.68 | 0.168 | 0.53 | 0.765 | 0.001 | 0.371 | 0.16 |
| IGF1R | 0.596 | 0.032 | 0.267 | 0.38 | -0.159 | 0.6 | 0.345 | 0.24 | -0.053 | 0.84 | 0.077 | 0.78 | 0.667 | 0.005 | 0.521 | 0.04 |
| FGFR1 | -0.234 | 0.441 | 0.542 | 0.055 | 0.269 | 0.37 | 0.444 | 0.13 | -0.021 | 0.94 | 0.132 | 0.62 | 0.794 | <0.0001 | 0.455 | 0.08 |
| FGF7 | 0 | 1 | 0.576 | 0.04 | -0.055 | 0.86 | 0.424 | 0.15 | -0.127 | 0.64 | 0.047 | 0.86 | 0.721 | 0.002 | 0.401 | 0.12 |
| IGF2 | 0.041 | 0.89 | 0.119 | 0.7 | -0.033 | 0.91 | -0.043 | 0.69 | -0.093 | 0.73 | 0.144 | 0.59 | 0.774 | <0.0001 | 0.337 | 0.2 |
| IGFR2 | 0.562 | 0.046 | 0.375 | 0.21 | -0.06 | 0.84 | 0.655 | 0.015 | 0.169 | 0.53 | 0.018 | 0.95 | 0.524 | 0.037 | 0.364 | 0.15 |
| IRS1 | -0.259 | 0.39 | 0.457 | 0.12 | -0.121 | 0.69 | 0.384 | 0.19 | 0.056 | 0.84 | 0.471 | 0.07 | 0.915 | <0.0001 | 0.531 | 0.05 |
| IRS2 | 0.259 | 0.39 | 0.501 | 0.08 | 0.005 | 0.99 | 0.623 | 0.02 | 0.113 | 0.68 | 0.203 | 0.45 | 0.582 | 0.018 | 0.05 | 0.83 |
| ERBB2 | 0.341 | 0.25 | 0.529 | 0.06 | -0.676 | 0.01 | 0.291 | 0.34 | 0.533 | 0.03 | 0.288 | 0.28 | -0.141 | 0.602 | 0.097 | 0.72 |
| EMT and migration (n=10) | ||||||||||||||||
| VIM | 0.135 | 0.66 | 0.518 | 0.07 | 0.280 | 0.35 | 0.709 | 0.007 | 0.186 | 0.49 | 0.132 | 0.62 | 0.812 | <0.0001 | 0.379 | 0.15 |
| CDH1 | 0.501 | 0.08 | 0.57 | 0.09 | -0.264 | 0.38 | 0.592 | 0.03 | 0.284 | 0.286 | 0.085 | 0.75 | 0.051 | 0.85 | 0.214 | 0.43 |
| SNAI1 | 0.239 | 0.43 | 0.102 | 0.74 | 0.066 | 0.83 | 0.131 | 0.67 | 0.087 | 0.75 | 0.068 | 0.8 | 0.577 | 0.02 | 0.241 | 0.37 |
| TGFB1 | 0.297 | 0.32 | 0.182 | 0.55 | -0.511 | 0.83 | -0.165 | 0.59 | 0.21 | 0.39 | -0.091 | 0.74 | 0.453 | 0.08 | 0.118 | 0.66 |
| RUNX3 | -0.771 | 0.8 | 0.202 | 0.51 | -0.313 | 0.30 | -0.065 | 0.83 | 0.195 | 0.47 | 0.081 | 0.76 | -0.199 | 0.46 | -0.071 | 0.8 |
| SNAI2 | 0.317 | 0.29 | 0.441 | 0.13 | 0.044 | 0.89 | 0.605 | 0.04 | 0.282 | 0.29 | 0.171 | 0.53 | 0.711 | 0.002 | 0.131 | 0.63 |
| TWIST2 | -0.005 | 0.99 | 0.667 | 0.013 | 0.115 | 0.71 | 0.504 | 0.08 | 0.121 | 0.66 | 0.109 | 0.69 | 0.827 | <0.0001 | 0.307 | 0.25 |
| ZEB2 | 0.033 | 0.91 | 0.661 | 0.014 | -0.038 | 0.90 | 0.602 | 0.04 | 0.133 | 0.62 | 0.242 | 0.37 | 0.477 | 0.001 | 0.462 | 0.07 |
| RHOA | 0.600 | 0.034 | 0.176 | 0.56 | -0.203 | 0.50 | 0.484 | 0.09 | 0.693 | 0.003 | 0.324 | 0.22 | -0.056 | 0.84 | 0.171 | 0.53 |
| RHOB | -0.215 | 0.48 | 0.295 | 0.23 | 0.429 | 0.14 | 0.444 | 0.13 | -0.094 | 0.73 | 0.041 | 0.88 | 0.559 | 0.02 | 0.201 | 0.45 |
| Cell proliferation and migration (n=3) | ||||||||||||||||
| Ki67 | 0.463 | 0.11 | 0.328 | 0.27 | -0.176 | 0.56 | 0.537 | 0.26 | 0.277 | 0.3 | -0.135 | 0.61 | -0.665 | 0.006 | -0.381 | 0.1 |
| MMP2 | -0.099 | 0.74 | 0.774 | 0.002 | 0.164 | 0.59 | 0.701 | 0.01 | 0.121 | 0.65 | 0.118 | 0.66 | 0.800 | <0.0001 | 0.335 | 0.2 |
| MMP9 | 0.193 | 0.53 | 0.005 | 0.99 | -0.511 | 0.07 | -0.271 | 0.32 | 0.139 | 0.61 | 0.041 | 0.88 | 0.185 | 0.49 | 0.07 | 0.8 |
| Immunity (n=5) | ||||||||||||||||
| IDO1 | 0.528 | 0.07 | -0.228 | 0.44 | -0.440 | 0.13 | -0.245 | 0.42 | 0.272 | 0.3 | -0.025 | 0.94 | -0.727 | 0.001 | -0.601 | 0.014 |
| TDO2 | 0.267 | 0.38 | 0.330 | 0.27 | -0.280 | 0.35 | 0.048 | 0.87 | 0.046 | 0.87 | 0.103 | 0.7 | 0.218 | 0.41 | 0.163 | 0.54 |
| PD1 | 0.534 | 0.06 | 0.033 | 0.92 | 0.115 | 0.71 | 0.209 | 0.49 | -0.155 | 0.56 | -0.132 | 0.62 | 0.056 | 0.84 | -0.261 | 0.25 |
| CD274 | 0.446 | 0.13 | -0.437 | 0.13 | -0.060 | 0.85 | -0.435 | 0.11 | 0.199 | 0.46 | 0.156 | 0.56 | -0.251 | 0.34 | -0.332 | 0.15 |
| PDL2 | 0.332 | 0.26 | 0.080 | 0.79 | 0.091 | 0.77 | 0.266 | 0.38 | -0.015 | 0.95 | -0.172 | 0.517 | 0.455 | 0.08 | -0.032 | 0.76 |
| Angiogenesis (n=6) | ||||||||||||||||
| FLT1 | 0.559 | 0.047 | 0.303 | 0.31 | 0.214 | 0.48 | 0.319 | 0.29 | -0.407 | 0.12 | -0.394 | 0.13 | 0.032 | 0.91 | -0.176 | 0.51 |
| VEGF165 | 0.402 | 0.17 | 0.358 | 0.23 | 0.011 | 0.97 | 0.364 | 0.22 | -0.531 | 0.03 | -0.641 | 0.01 | -0.321 | 0.23 | -0.013 | 0.68 |
| VEGF189 | 0.306 | 0.31 | 0.088 | 0.77 | -0.137 | 0.65 | 0.114 | 0.71 | -0.181 | 0.51 | -0.238 | 0.37 | -0.156 | 0.56 | -0.046 | 0.86 |
| KDR | -0.187 | 0.54 | -0.151 | 0.62 | -0.368 | 0.22 | -0.387 | 0.19 | -0.081 | 0.77 | -0.079 | 0.77 | 0.482 | 0.06 | 0.061 | 0.82 |
| VEGFC | 0.179 | 0.56 | 0.328 | 0.27 | 0 | 1 | 0.199 | 0.51 | 0.01 | 0.88 | 0.132 | 0.63 | 0.788 | 0.0001 | 0.353 | 0.18 |
| NRP1 | 0.185 | 0.55 | 0.614 | 0.02 | 0.033 | 0.91 | 0.411 | 0.16 | 0.131 | 0.62 | 0.168 | 0.53 | 0.838 | <0.0001 | 0.324 | 0.22 |
r, Spearman’s rank test (relationship between two quantitative parameters). Values in bold type are statistically significant at confidence level greater than 99% (p value<0.01) and r>0.6. Cyp1B1 and AhRR were not significantly increased in intestinal GC and statistical correlations should be considered with caution (see Table 2).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



