Submitted:
21 June 2024
Posted:
24 June 2024
You are already at the latest version
Abstract
The study is based on information as the property of matter and its evolution from chemical to digital information. Consequences of these forms of information include molecular, modular, cel-lular, non-human, and human individual interactions, as well as human-machine interaction, and the formation of corresponding modules through self-organizing selective processes. The function of modular structures is decision-making and is analogous to the behavior of an intelligent agent. The evolutionary modular model is proposed based on the interaction of evolutionary factors (matter, energy, information, self-organization, natural selection, and heritability) among themselves and with the arrow of time. In conclusion, we realize that the essence of the evolution of living organ-isms is modular evolution. Modules through which the survival and reproduction of living organ-isms is ensured, are formed by two basic molecular processes, which are molecular recognition or information and modular self-organization.
Keywords:
evolution
; information
; module
; self-organization
; molecular recognition
; intelligent agent
; evolutionary factors
; modular evolution
1. Introduction
In the scientific opinion, it is accepted that concepts of information based on the technique of signal transmission or entropy, interactions, and self-organizing selective processes during which modules are formed in the living world as intelligent agents or Maxwell's demons cannot be explained.
In two previous studies [1,2], the idea that information is a property of matter [3], is material [4,5,6], is the ability to organize and interact with matter [7], information nourishes evolution [8], has genetic and non-genetic nature [9], interactions are a consequence of information [10] has been supported.
Perhaps, even Ch. Adami's definition [11] that information is the ability to make predictions better than expected by chance can be translated as the ability to explain why two or more elements of a community are more often found together than expected by chance. Also, the concept of information from the structural and symbolic division [12] implies its material origin. In these studies, it has been highlighted through a mental experiment that modules behave like demons of information as a property of matter from which they are composed.
Furthermore, it has been demonstrated that an evolutionary tree can be constructed based on the formats of information from which various types of interactions have emerged. From all these interactions, the components of living organisms are formed, meaning those modules that ensure their survival and reproduction [13].
In this context, a fundamental problem arises for the study at hand. It is accepted that the life of every living organism is a permanent chain of decisions and actions resulting from these decisions [14]. This means that the function of each module is a decision-making process linked to an action, and the action itself is another decision-making process. In each link of this chain, a function appears. The probability of performing this function, converted into bits, is called semantic information [1,2,15]. Hence, semantic information arises from the interactions between modules and the environment. As a result, the behavior of each module can and should be the behavior of an intelligent agent.
Has it not been accepted, for four decades now, that to live means to know [16]?
Finally, it is proposed that the factors of evolution are those that participate in or affect the formation of modules. Such factors include matter, energy, information, self-organization, natural selection, and heritability. Based on the interaction among these evolutionary factors and the arrow of time, the modular evolution model is constructed from the discussion of which several interesting problems arise.
2. The Structure of the Module Arises from Information
It is argued that meaning is usage [17]. I believe a more accurate perspective is that meaning is a function: something is used when its function is recognized because the function occurs before the usage. Equally accurate is the notion that the structure arises from information, and not the structure arises from usage [17]. We discuss these two aphorisms about the concept of the module in living organisms. When conceptualizing a module as a system, we refer to its definition [18]: a system is a community of elements that interact with one another. This definition lays the groundwork for solving the problem of what elements of a system interact with each other. For example, what does an enzyme do to recognize a substrate, or what does a substrate do to recognize an enzyme? Certainly, there is recognition, and for this recognition, chemists publish a specialized journal titled "Molecular Recognition”. Thus, we must accept that the structure of the module is born primarily from molecular recognition, meaning from information. Rightfully so, molecular recognition and self-organization are considered two main characteristics of molecules [19]. It is a well-known idea that a module can and should be analyzed, on one hand, as a structure and, on the other hand, as a function [20].
From the structural perspective, a module is a community of molecules, cellular elements or cells, individuals of the same species, or individuals of different species, whose interactions perform a function or form an effector that executes a function [2]. Let us analyze a molecular community, such as a group of ATP or GTP molecules. Both molecules are considered universal energy coins. During evolution, they specialized in two directions. In cells that use more energy, ATP molecules have an advantage because their formation has a lower cost, whereas in cells specialized for signaling, GTP molecules are mainly used, even though the latter has a higher formation cost due to the G-C binding having three hydrogen bonds compared to the two in A-T and A-U bindings.
In a previous study [15], it has been shown that genes are also modules, specifically genetic modules. What is the structure of genetic or epigenetic modules? I do not think the structure of genetic modules can be identified solely by a fragment of nucleic acids. The same can be said for complex neural modules often identified with the term "memes" by R. Dawkins [21]. If genes and memes are modules with effector chemical modules such as the groups of ATP and DTP molecules or as the function of an allosteric enzyme, they are considered modules without effectors [2].
What we know today about the structure of modules is that their formation is a result of self-organizing processes [12,20,22,23,24]. By self-organization, we imply the interactions of elements at the lowest level of a system, from which emerges a global behavior at the highest level of this system [25]. From this perspective, the question arises of how the function of the module manifests in a way that acts as an intelligent agent and why modules are real intelligent agents.
3. Module as an Intelligent Agent or Decision-Maker
Today, scientific thought supports the idea that modules in the living world are formed by self-organizing processes or more precisely by self-organizing selective processes. From this, we can write that modules are formed through self-organization, where the term "module" (M) implies the term "self-organization" (SO). About a decade ago, in a study [26], it was argued that self-organization, which, as mentioned above, arises from self-organizing processes, is a process similar to human decision-making. According to these authors, the similarity in words is the result of using the same mathematical formulation belonging to probabilistic methods:
- -
- Self-organization is the process of evaluating the probabilities of states in a system in search of a more thermodynamically stable state.
- -
- Decision-making is the process of evaluating alternatives (states) of decision-making in search of the most stable preferential alternative.
From these claims, we conclude that self-organization can be considered decision-making (SO is DM), and decision-making can be considered self-organization (DM is SO). If the axiom of transitivity is to be used, we can assume that when the formation of modules (D) implies self-organization (SO) and self-organization implies decision-making (DM), then the formation of modules (D) can be considered decision-making (DM). It should be emphasized that decision-making is much more complex: several agents are involved in a decision. Thus, it remains to be accepted that each module is formed and behaves as an intelligent agent. As an intelligent agent, the module is defined in the functional perspective:
A module, much like an intelligent agent (A) perceives and stores information about the object (O) it needs to act upon, which we metaphorically call a lock. Thus, based on this information, the agent forms the effector (E), meaning the entirety that opens/unlocks the lock and in this way, the necessary function is performed. It should be emphasized that the formation of each module is not only a self-organizing process but also a selective one, and as such, the module does not always behave as a decision-maker or an intelligent agent.
In the first stage, intramodular selection occurs, meaning within the structure of the module, and in the second stage, intermodular selection occurs when the module interacts with other modules or with the environment. From this, we conclude that the classical variation-selection dyad [27] can be replaced by the dyad of intra and intermodular selection, which is in support of the idea of the marriage of self-organization with natural selection [28]. Before explaining the behavior of modules as intelligent agents, it is necessary to conceptualize these modules as real, not imaginary demons, from whose behavior semantic information is created.
4. Modules Are Real Demons: Definition of Information
As mentioned in the introduction to this study, the factor that makes what E. Schrodinger [29] calls organized matter possible is information as property of this matter. Accepting the idea that information is a property of matter undermines the concept that information in a system comes from the outside [30,31]. Let's look at Figure 1 where in a container divided into two parts, there are two molecules, one red and the other green. In placing the molecules in a container with two partitions, an observer distinguishes four microstates (Figure 1).
Let's assume that the two molecules are the enzyme and its substrate. As a result, they bind together in the region of their steric or chemical information (Figure 1b), and the observer discerns only two microstates. We also assume that due to a physical barrier, the enzyme-substrate aggregate does not pass into the neighboring compartment of the container. After a while, the observer notices several microstates because a container separation can be formed by one or more aggregates. Above all, it is expected to find a group of molecules formed in a corner of each compartment without the intervention of the imaginary demon.
The factor that produces organized matter must be sought within the matter itself and not outside it or the container. The two halves of a container are different because the information is a change that makes a difference [32], or because something becomes a source of information when it is present in two or more states [33]. From here, we formulate the definition of interaction information: The information of interaction is the property of the constituent elements of material and non-material systems that enables their interaction both within and between systems. This interaction forms the structure of modules that function as intelligent agents. Below, we will attempt to argue this definition even further.
5. Modular Thinking and Modular Biology
Modules, just like coins, have two sides. On one side is the structure, and on the other is the function. Above, the definition of the module has been given in terms of both structure and function and now we need to understand and explain the origin and nature of the module simultaneously as both structure and function. Thinking about the module as both structure and function is like thinking popularly [34], thinking in terms of trees [35], selection thinking and is called modular thinking.
In this article, we will try to explain modular thinking through the main types of modules formed in every living organism, including humans, language, and human society.
In Figure 2, marked with the letter A are indicated the agents that perceive and store information about a stimulus that the module must react to or about a task that the module must fulfill, which are marked with the letter O, and metaphorically called the lock. Based on this information, effectors or corresponding keys, marked with the letter E, are formed to perform the appropriate function.
In contrast to the classical concept of information, the value of the semantic information of a module is not discrete, meaning either 0 or 1, but fluctuates according to the probability of the function's execution with the value pi.
If the probability of the execution of the function, pi, is less than or equal to 0.5, the semantic information (Is) is calculated using the formula: Is=log2 pi (1)
If pi is greater than 0.5, then this formula is used: Is=1+(1−log2 pi)
We cannot overlook the fact that the non-discrete nature, non-deterministic, and dependent on semantic information, resembles quantum information [36]. Just like quantum thinking is like modular thinking. However, this is an observation that requires special attention.
In Figure 2a, a genetic module is presented in which the agent (A) carries information about a DNA segment, meaning it is a gene, for example, a gene that breaks down lactose. In contrast, lactase, as the gene's product, is an effector (E), the key that opens the lock, which breaks down lactose (O), and the necessary function is performed.
The analogy often drawn between genes and memes is in their common nature as intelligent agents (Figure 2a and 2b). Let us recall the concept of the mental model formulated about eight decades ago [37] that the mental model is an internal representation of the external reality. From this concept, it has been concluded that DNA is also an internal representation of external reality [38].
In a sentence, the subject is called the agent (Figure 2b). Humans produce and understand language because, before acting or undergoing an action, they recognize the object and react with effectors (verbs) solely based on the information gathered from this object [1].
The concept of modulome is equally interesting as it replaces not only the agent as information but also the genome. By modulome, we understand the information of a cell, individual, species, or human community in which DNA is a part of the general information. It is not difficult to understand that the effectors of an individual, species, or human community are respectively the modules, individuals, and institutions (Figure 2), while modulome is a community of intelligent agents.
The common nature of modules as intelligent agents also has unique aspects that need to be discussed. Thus, the naming of different forms of information is based on semantic information. For instance, the structure of the genetic module is formed by the chemical information of interaction. According to this concept, processes like duplication, transcription, translation, etc., do not contain genetic or epigenetic information. It is considered genetic information only when the effector is formed as the product of a gene. Similarly, it is considered cellular information when an effector is formed through cellular interactions. Naturally, chemical, genetic, and epigenetic information are included in cellular information because, as mentioned below, both information and the respective modules have a hierarchical nature.
6. The Tree of Life Is Based on the Formats of Interaction Information
The construction of the Tree of Life is based on the principle of common descent, and from here, it follows that the increase in the complexity of living organisms is the result of a series of major evolutionary transformations. According to the well-known evolutionists J. Maynard – Smith and E. Szathmary [13], these transitions involve changes in how information is stored and transmitted.
The Tree of Life is based on the types of information, the types of interactions that cause these forms, and the modules they form (Figure 3).
These modules in the Hutchinsonian niche space perform their respective tasks to ensure survival and reproduction in different periods of the evolution of life. It is emphasized that life processes performed through modules form a continuous chain of decision-making and actions resulting from this decision-making: decision-making / decision decision-making / decision...[14].
In this figure, it is shown that the types of information that appear in the living world, as well as the matter itself, have their roots in the primordial nucleosynthesis of H and He. This line is divided into three types of interactions: A1 (modular interactions), A2 (individual non-human interactions), and A3 (chemical-genetic interactions). Modular interactions occur from all types of information interaction, and all these interactions create a unique form of information called semantic information. Simultaneously, with the emergence of the first living organisms, some types of information appear, such as species information (A2.1), endosymbiotic (A2.2), coevolutionary (A2.3), and holobiontic (A2.4), which enable interactions between individuals of the same species and individuals of different species (A2) by forming corresponding modules.
Chemical-genetic molecular interactions are the result of chemical, genetic, and epigenetic information, which form respective modules with the same name (A3). Around two billion years ago, cell interactions (A4) began, which are the result of somatic cell information (multicellularity, A4.1), sexual cell information (sexual reproduction, A4.2), and neural cell information (birth of the nervous system, A4.3). Simple neuro-endocrine and complex neural modules were formed from neural cell information (A4.3), and complex modules, such as cognitive modules, and individual human interactions (A5), were born. The birth of human interactions, communication, and language learning marked the emergence of the Cognitive Revolution, about 70,000 years ago [39].
The development of language and learning through language has been influenced by information from mirror neurons (A5.1), linguistic information (A5.2), and social, cultural, and linguistic information (A5.3). These factors have led to the formation of gesture modules, word, and sentence modules, as well as complex social-cultural and linguistic modules.
Scientific opinion must answer the question of whether humanity is currently living in the Second Cognitive Revolution, and consequently, it is involved in its sixth interaction, the human-machine interaction. I am convinced that humanity is on the verge of this interaction caused by digital information, and perhaps quantum information will also be added to this information not too far in the future. On the other hand, some other issues should be discussed, among which four of them are mentioned:
Firstly, the table diagram of Figure 3 cannot be considered a "tree," and we consider this fact as positive. Perhaps in the future, this type of tree could be divided into columns and rows similar to those of the periodic table of elements, and the systematics of the future might be based on modules and modular networks. Secondly, Figure 3 shows that species have appeared simultaneously with the emergence of life (A2.1) and can be considered modules. On the other hand, it is shown that throughout evolution, but especially in the first living organisms, collective evolution has always been present [40], as evidenced by the formation of modules among individuals of different species (endosymbiotic, coevolutionary, and holobiontic modules) and the fact that horizontal gene transfer is a normal phenomenon in evolution.
Thirdly, we propose that the emergence of cultural inheritance as a second system of inheritance is linked to the appearance of mirror neurons because, just as a DNA sequence forms another sequence (molecular replication), the same module is formed in both the one making a gesture and in the one observing this gesture (individual replication) [41]. Thus, the formation of gesture modules has been a prerequisite for the formation of word modules, sentence modules, and social, cultural, and linguistic modules.
Finally, the concept of information as a property of matter excludes such thoughts in the field of biosemiotics that coding rules are not dictated by laws of the physical and chemical world [42] that plants can use signs (icons, indices, or symbols) to communicate between them [43] that structure arises from usage and not from information or that meaning is usage and not function [17] etc.
7. The Model of Modular Evolution
7.1. Module Description
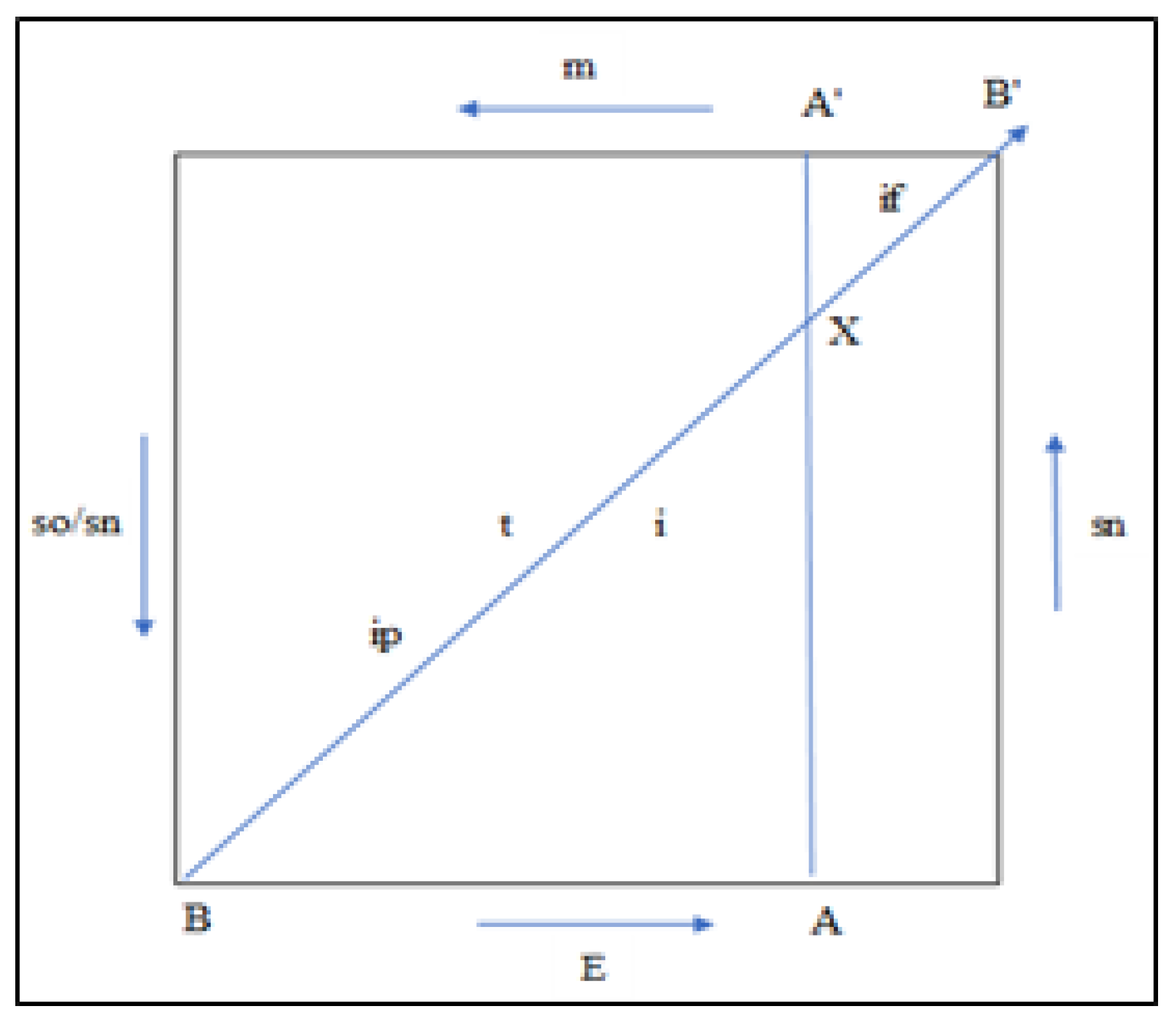
The model of modular evolution is constructed based on the connections between evolutionary factors and the arrow of time among them (Figure 4).
In Figure 4 the value of evolutionary factors (E, m, so, sn, i, and h2), ranging from 0 to 1, indicates the direction of the arrow in the four quadrants of the square and on its diagonal, simultaneously representing time and the sum of the values of semantic information of all modules within a taxon.
The position of each taxon is a point on the arrow of time or the diagonal representing semantic information. The essence of the model lies in the connections between the factors and with the arrow of time. For example, the value of energy is the mass converted into energy, and energy and mass are represented in the model as complementary magnitudes.
Similarly, it is implied that energy and information increase along the arrow of time. The interpretation of the relationships is more complex between self-organization (so), natural selection (sn), and heritability (h2). Heritability is a statistical concept that indicates how many variations in traits or variations in the expression of semantic information are attributed to variations in the forms of information exchange, and how many of them are "noise" dictated by the nonlinear dynamics in the process of module formation.
Self-organization (so) is expressed as the percentage of formed modules, while natural selection (sn) is expressed as the percentage of eliminated modules. Next, we will attempt to argue how the value of heritability (h2) and natural selection (sn) decreases over time.
7.2. Information Is Not Matter But One of Its Properties
In Figure 4 with the letter X on the diagonal of the square representing both time and the value of information, the hypothetical taxon's position is indicated.
Assuming life originated four billion years ago, taxon X in our model belongs approximately to that group of living organisms that appeared around a billion years ago, including the first multicellular organisms, sexually reproducing organisms, and those equipped with a simple nervous system. If we draw a perpendicular line from point X to the side/edge in Figure 4, we obtain the right-angled triangle OXe, where OX is the hypotenuse, Xe is the cathetus with a value of "so," and the other cathetus eO has a value equal to the mass converted into energy.
From here we can write: i2 = e2 + so2 (1)
Formula 1 shows that the squared information of taxon X is equal to the squared sum of energy and the squared number of modules formed. The fact that information is a larger quantity than energy (Formula 1) does not support the idea of the equivalence of matter, energy, and information [44,45]. Also, the lack of this equivalence does not support the notion that information has "mass," that it is a "pure form of matter," a physical unit [44,45], a special state of matter [46], even though it is true that information is present in every particle [45], and this permanent presence in the matter is information as its property. Naturally, just like matter and energy, information has evolved to the extent that its highest form is now immaterial information.
7.3. Evolutionary Factors: e = m∙i
P. Lherminier [31] has put forth the interesting idea that "each species is a unique solution to the equation that balances energy/matter expenditures with the gain of information (i)." In Figure 3, the position of each species is any point on the diagonal and indicates that the value of semantic information (i) increases simultaneously with the energy used as mass converted into energy (e) but decreases the mass of unconverted matter (m). This means that information (i) and mass converted into energy (e), which is the expenditure or use of energy, increase in parallel, but matter as unconverted mass (m) decreases.
On the other hand, with the increase in self-organizing selection processes (so) or the increase in the number of modules, the mass of unconverted matter decreases.
Comments of this nature are evident from the graphical representation of the modular evolution model. What needs to be emphasized in this model is the fact that the factors or forces that enable evolution must be sought in the energy-matter and information relationships. This is also illustrated by the equation e = m∙i.
Of course, this fact does not dismiss the idea that natural selection, mutations, migrations, and genetic drift are factors in evolution. However, it should be emphasized that these factors change the genetic composition of the population, leading to microevolutionary processes, but this is not a sure proof that these changes are automatically responsible for macroevolutionary processes such as the formation of species.
The proposed model is an attempt to show that the formation of species is linked to the transformations of matter/energy under the pressure of semantic information arising from the decision-making functions of modules as intelligent agents.
From here, it follows that the so-called functional information law [47] applies only to living systems. E. Zuckerkandl and L. Pauling [48] were the first to note that genetic distances between taxons can be converted into absolute geological time. In the evolution model, absolute geological time is represented by the arrow of time that runs parallel to the increase in information.
Let's assume that the population of a hypothetical species, for its parameter values, has the position x on the arrow of time (Figure 4). This position corresponds to the concept that each species is a field of recombination of a group of individuals where only the connections between the groups create a stable relationship between energy/matter expenses and information gain [31]. By drawing a perpendicular line from point x to the four sides of the square, similar triangles xAB and XA'B' are obtained.
Based on the similarity of the triangles xAB and xA'B', we can write:
AB/A'B' = xB/xB'= xA/ xA'
or e/m = ip/if(t0/t1) = s/he)
We acknowledge that the ip/if ratio, which represents the amount of information before the population of type x is formed (ip) compared to the amount of information after the formation of population x until today (if), remains constant. This constant characterizes populations of type x and has an i value. Such a constant exists for every other type of living world.
ip/if = i
As a result, we can write:
e/m = i and e = m∙i
The similarity of the equation e = m∙i with the famous formula E=mc2 is evident but is this similarity essential or just an intellectual game (!?) The problem is to be discussed. However, let's continue with our idea. We can replace the ratio ip/if with the ratio t0/t1 because the diagonal also represents time in years. Thus, t0 represents the time elapsed from the birth of population x, and t1 represents the time elapsed from the birth of population x to the present. Let's assume that life originated approximately four billion years ago, and the first fossil of a living organism belongs to an age of 3.8 billion years, meaning t0 = 200,000,000 and t1 = 3,800,000,000.
From this data, we can calculate the value of coefficient i because:
ip/if = t0/t1 = i
i = 200 000 000/ 3 800 000 000 = 0,05
We calculate the coefficient i for humans, where t1 = 200,000 years (the time when modern humans appeared) and t0 = 4,000,000,000 - 200,000 = 3,999,800,000 is the time before the appearance of humans.
Therefore: t0/t1 = 3,999,800,000 / 200,000 = i = 19,999 or 20,000.
The replacement of the coefficient i specifically for the first living fossils (i = 0.05) and for humans (i = 19,999) in the formula e = m∙i shows that for the same mass (m), humans possess an energy quantity many times larger. In other words, throughout evolution, more and more work are accomplished with a smaller mass because evolution is the process of energizing the mass of living systems through the increase in the number of modules or modularization of mass, as well as the increase in semantic, functional, or modular information.
On the other hand, almost a century ago, L. Lapicque [49] found that for the excitation of invertebrate muscles like Aplysia, 703 ergs are needed, while for vertebrates like Rana esculenta, 518 ergs are required. From this, A.T. Pol [50] concluded that during evolution, the quality of information increases as a force of interaction and reorganization of matter, and energy use is minimized. This means that more evolved organisms use less energy for the same work, but their use is more efficient.
Regardless of the incorrect concept of predatory information, according to which living systems "seize" information from the outside and not that information is a property of the matter they are composed of, P. Lherminier [31] has given an interesting idea when he writes that "each species is a unique solution to the equation that balances material/energy expenditures with information gain." The formula e = m∙i is the equation that shows how matter/energy expenditures are balanced with information gain in every world living species. As a result, the author's thought supports the modular evolution model and vice versa. However, we emphasize once again that individuals within a species should not be labeled as predators and victims, as P. Lherminier [31] writes, because, above all, interaction occurs between them based on the information they possess.
Another interesting connection worth discussing is the relationship of inheritance or heritability, as described earlier as an evolution factor. The heritability coefficient (h) reduction is explained by the increased number of neural modules and their role in higher organisms. Furthermore, the presence and role of these modules also explain the low level of selective pressure (sn).
The equation e = m∙i reflects the connections between evolution factors over time from the emergence of life, during which interactions based primarily on information have occurred and continue. In contrast, the equation E = m.c2 explains the energy/matter equivalence since the time of the Big Bang when interactions were mainly based on force.
The fact that during the formation of a helium atom from four hydrogen atoms, 0.774% of the mass of the hydrogen atoms is lost suggests that the electromagnetic origin of mass might not cause this loss. Like biological and chemical evolution, energy growth is balanced by a mass reduction through the interaction of information, which acts as a complementary component of this process (Figure 4). From this, it can be accepted that the peaks of chemical and biological evolution are heavy elements like uranium and higher animals, with humans at the forefront, possessing a very high energy potential.
The inventory of nature and everything humans create is formed over time due to the interaction of matter, energy, and information. This is why these universal parameters are considered the basic factors of chemical evolution, the origin of life, biological evolution, and socio-cultural and linguistic evolution. From the interaction between these three parameters, three other factors of evolution emerged simultaneously: self-organization, natural selection, and heredity. The modular evolution model helps us explain simultaneously the interconnections over time of all evolutionary factors.
It's worth noting that the Book of Life is not just the DNA molecule but also all the different types of information interactions with the corresponding modules mentioned above. Thus, the difference between chimpanzees and modern humans should not be sought in their DNA sequences but in the fact that humans are an energetic paradox [51]. This is why we consider evolution a process of modularization and energization of living systems through the interaction of information. In simpler terms, evolution is the acquisition and growth of information from which, over time, more numerous and complex modules are formed. Such evolution is modular evolution.
The idea that embryonic development is not solely governed by the genetic community is correct, but it does not mean that their absence is compensated for by spatial-temporal conditions [52]. Below, it will be shown that modularity and self-organization in living organisms are consequences of information. In a recent study [53], the issue of purpose in living organisms is discussed. According to these authors, directed purpose depends on persistence, meaning the continuous presence over time of a function or an adaptation, as well as plasticity, which, in our opinion, also involves persistence.
Their formula for calculating persistence (P) is:
P= GN _R 1- R where:
G represents the number of successful moves; N represents the total number of moves, and R represents a successful move that could also occur by chance. The value derived from this formula is almost the same as that used to measure the amount of semantic (Is) modular or functional information: Is= log2 pi ) p1, and Is= 1+ (1-log2 pi) This fact supports the idea that persistence, dependent on the directed purpose of living organisms, is the value of semantic information. It can also be said that the birth of semantic information as the interaction of a module with the environment or other modules is the birth of a directed purpose.
From this, it follows that the birth of a purpose from a modular function occurs in every module of living organisms because all types of modules are valid for survival and reproduction. Hence, we should dismiss the notion that biologists sin when accepting function and meaningful information in every module.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
None.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bajrami, Z.; Rrokaj, S. Modular Evolution: A Hypothesis for Language Acquisition. International Journal of Language and Linguistic 2023, 10, 16–26. [Google Scholar]
- Bajrami, Z. Informacioni dhe evolucioni modular; Akademia e Shkencave të Shqipërisë: Tiranë, 2024; ISBN 9789928809193. [Google Scholar]
- Spirkin, A.G. Dialectical Materialism; Progress Publishers: Moscow, 1983. [Google Scholar]
- Landauer, R. Information Is Physical. Physics Today 1991, 44, 23–29. [Google Scholar] [CrossRef]
- Landauer, R. Information Is a Physical Entity. Physica A: Statistical Mechanics and its Applications 1999, 263, 63–67. [Google Scholar] [CrossRef]
- Mingers, J. Information and Meaning: Foundations for an Intersubjective Account. Information Systems Journal 1995, 5, 285–306. [Google Scholar] [CrossRef]
- Stonier, T. Information as a Basic Property of the Universe. Biosystems 1996, 38, 135–140. [Google Scholar] [CrossRef]
- Lehn, J.-M. Toward Complex Matter: Supramolecular Chemistry and Self-Organization. Proc Natl Acad Sci U S A 2002, 99, 4763–4768. [Google Scholar] [CrossRef]
- Cabej, N.R. On the Origin and Nature of Nongenetic Information in Eumetazoans. Annals of the New York Academy of Sciences 2023, 1525, 104–127. [Google Scholar] [CrossRef]
- Gershenson, C.; Fernández, N. Complexity and Information: Measuring Emergence, Self-Organization, and Homeostasis at Multiple Scales. Complexity 2012, 18, 29–44. [Google Scholar] [CrossRef]
- Adami, C. The Evolution of Biological Information: How Evolution Creates Complexity, from Viruses to Brains; Princeton University Press; ISBN 978-0-691-24115-9.
- Haken, H.; Portugali, J. Information and Self-Organization. Entropy 2017, 19, 18. [Google Scholar] [CrossRef]
- Szathmáry, E.; Smith, J.M. The Major Evolutionary Transitions. Nature 1995, 374, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Kovalenko, T.; Vincent, S.; Yukalov, V.I.; Sornette, D. Calibration of Quantum Decision Theory: Aversion to Large Losses and Predictability of Probabilistic Choices. J. Phys. Complex. 2023, 4, 015009. [Google Scholar] [CrossRef]
- Bajrami, Z. The Modular Concept of Gene. Journal of Natural Sciences Research 2013, 3, 125–130. [Google Scholar]
- Maturana, H.R.; Varela, F.J. The Tree of Knowledge: The Biological Roots of Human Understanding; Revised edition.; Shambhala: Boston, 1992; ISBN 978-0-87773-642-4. [Google Scholar]
- Tomasello, M. Constructing a Language: A Usage-Based Theory of Language Acquisition; First Edition.; Harvard University Press: Cambridge, Mass, 2005; ISBN 978-0-674-01764-1. [Google Scholar]
- Von Bertalanffy, L. General System Theory: Foundations, Development, Applications (Revised Edition) by Ludwig Von Bertalanffy: New Paperback (1969) | GoldenWavesOfBooks. Available online: https://www.abebooks.com/9780807604533/General-System-Theory-Foundations-Development-0807604534/plp (accessed on 29 May 2024).
- Sugita, M.; Onishi, I.; Irisa, M.; Yoshida, N.; Hirata, F. Molecular Recognition and Self-Organization in Life Phenomena Studied by a Statistical Mechanics of Molecular Liquids, the RISM/3D-RISM Theory. Molecules 2021, 26, 271. [Google Scholar] [CrossRef] [PubMed]
- Modularity in Development and Evolution; Schlosser, G. , Wagner, G.P., Eds.; University of Chicago Press: Chicago, 2004; ISBN 978-0-226-73853-6. [Google Scholar]
- Dawkins, R. The Selfish Gene: 40th Anniversary Edition; Oxford University Press: New York, NY, 2016; ISBN 978-0-19-878860-7. [Google Scholar]
- Wagner, G.P. Homologues, Natural Kinds and the Evolution of Modularity. Amer Zool 1996, 36, 36–43. [Google Scholar] [CrossRef]
- Wagner, G.P.; Lynch, V.J. Evolutionary Novelties. Curr Biol 2010, 20, R48–52. [Google Scholar] [CrossRef]
- Haken, H. Information and Self-Organization: A Macroscopic Approach to Complex Systems; Softcover reprint of hardcover 3rd ed. 2006 edition; Springer: Berlin, 2010; ISBN 978-3-642-06957-4. [Google Scholar]
- Self-Organization in Biological Systems; Camazine, S., Ed.; Princeton studies in complexity; 2. print., and 1. paperback print.; Princeton Univ. Press: Princeton, NJ, 2003; ISBN 978-0-691-11624-2. ISBN 978-0-691-11624-2.
- Yukalov, V.I.; Sornette, D. Self-Organization in Complex Systems as Decision Making. Advs. Complex Syst. 2014, 17, 1450016. [Google Scholar] [CrossRef]
- Forterre, P.I.P. Darwin’s Goldmine Is Still Open: Variation and Selection Run the World. Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, S.A. The Origins of Order: Self-Organization and Selection in Evolution; 1st edition.; Oxford University Press: New York, 1993; ISBN 978-0-19-507951-7. [Google Scholar]
- Schrödinger, E. What Is Life? & Mind and Matter: The Physical Aspect of the Living Cell; Cambridge University Press, 1974.
- Alemi, M. Life, Energy and Information. In The Amazing Journey of Reason; SpringerBriefs in Computer Science; Springer International Publishing: Cham, 2020; pp. 1–10. ISBN 978-3-030-25961-7. [Google Scholar]
- Lherminier, P. La prédation informative : vers un nouveau concept d’espèce. Comptes Rendus. Biologies 2018, 341, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Bateson, G. Steps to an Ecology of Mind; University of Chicago Press ed.; University of Chicago Press: Chicago, 2000; ISBN 978-0-226-03906-0.
- Shanon, C. A Mathematical Theory of Communication. 27, 379–423.
- Ernst Mayr Evolution and Anthropology: A Centennial Appraisal.; Washington, 1959.
- O’hara, R.J. Population Thinking and Tree Thinking in Systematics. Zoologica Scripta 1997, 26, 323–329. [Google Scholar] [CrossRef]
- Collins, N. January 18 2023.
- N, E.; Craik, K.J.W. The Nature of Explanation. Journal of Philosophy 1943, 40, 667. [Google Scholar] [CrossRef]
- Adami, C. The Use of Information Theory in Evolutionary Biology. 2012, 1256, 49–65. [CrossRef]
- Harari, Y.N. Sapiens: A Brief History of Humankind; Illustrated edition.; Harper: New York, NY, 2015; ISBN 978-0-06-231609-7. [Google Scholar]
- Vetsigian, K.; Woese, C.; Goldenfeld, N. Collective Evolution and the Genetic Code. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 10696–10701. [Google Scholar] [CrossRef] [PubMed]
- Rizzolatti, G.; Fabbri-Destro, M.; Cattaneo, L. Mirror Neurons and Their Clinical Relevance. Nat Clin Pract Neurol 2009, 5, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, M. From Biosemiotics to Code Biology. Biol Theory 2014, 9, 239–249. [Google Scholar] [CrossRef]
- Witzany, G. Plant Communication from Biosemiotic Perspective: Differences in Abiotic and Biotic Signal Perception Determine Content Arrangement of Response Behavior. Context Determines Meaning of Meta-, Inter- and Intraorganismic Plant Signaling. Plant Signaling & Behavior 2006, 1, 169–178. [Google Scholar] [CrossRef]
- Vopson, M.M. The Mass-Energy-Information Equivalence Principle. AIP Advances 2019, 9, 095206. [Google Scholar] [CrossRef]
- Vopson, M.M. Experimental Protocol for Testing the Mass–Energy–Information Equivalence Principle. AIP Advances 2022, 12, 035311. [Google Scholar] [CrossRef]
- Thompson, A. A Proposed Experimental Test for the Mass-Energy-Information Equivalence Principle. Scilight 2022, 2022, 091111. [Google Scholar] [CrossRef]
- Wong, M.L.; Cleland, C.E.; Arend, D.; Bartlett, S.; Cleaves, H.J.; Demarest, H.; Prabhu, A.; Lunine, J.I.; Hazen, R.M. On the Roles of Function and Selection in Evolving Systems. Proc. Natl. Acad. Sci. U.S.A. 2023, 120, e2310223120. [Google Scholar] [CrossRef]
- Zuckerkandl, E.; Pauling, L. Molecules as Documents of Evolutionary History. J Theor Biol 1965, 8, 357–366. [Google Scholar] [CrossRef]
- Lapicque, L. L’excitabilité En Fonction Du Temps : La Chronaxie, Sa Signification et Sa Mesure Par Louis Lapicque. 1926; Generic, 2022.
- Pol, A. Life: Energy-Information Relationship within Materials Systems: General Outline of the Concept T. Computers Math. Applic. 1990, 20, 269–285. [Google Scholar] [CrossRef]
- Pontzer, H.; Brown, M.H.; Raichlen, D.A.; Dunsworth, H.; Hare, B.; Walker, K.; Luke, A.; Dugas, L.R.; Durazo-Arvizu, R.; Schoeller, D.; et al. Metabolic Acceleration and the Evolution of Human Brain Size and Life History. Nature 2016, 533, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Layer, P.G. Brains Emerging: On Modularity and Self-Organisation of Neural Development In Vivo and In Vitro. In Emergence and Modularity in Life Sciences; Wegner, L.H., Lüttge, U., Eds.; Springer International Publishing: Cham, 2019; pp. 145–169. ISBN 978-3-030-06128-9. [Google Scholar]
- Lee, J.; McShea, D. Operationalizing Goal Directedness; An Empirical Route to Advancing a Philosophical Discussion. 2020, 12.
Figure 1.
Four microstates of two molecules (red and green) in a container divided into two parts (a) and two states (b) (According to [30]).
Figure 1.
Four microstates of two molecules (red and green) in a container divided into two parts (a) and two states (b) (According to [30]).
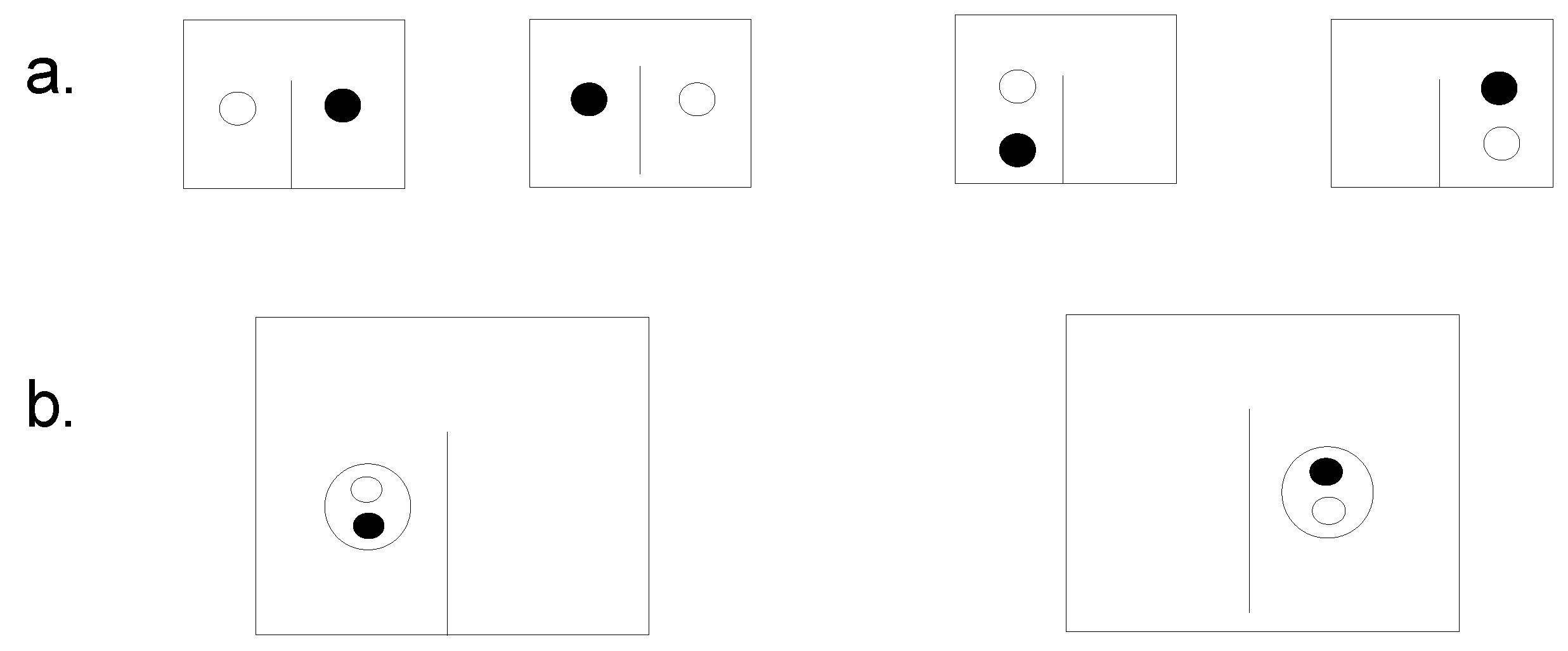
Figure 2.
Modules as functional units.
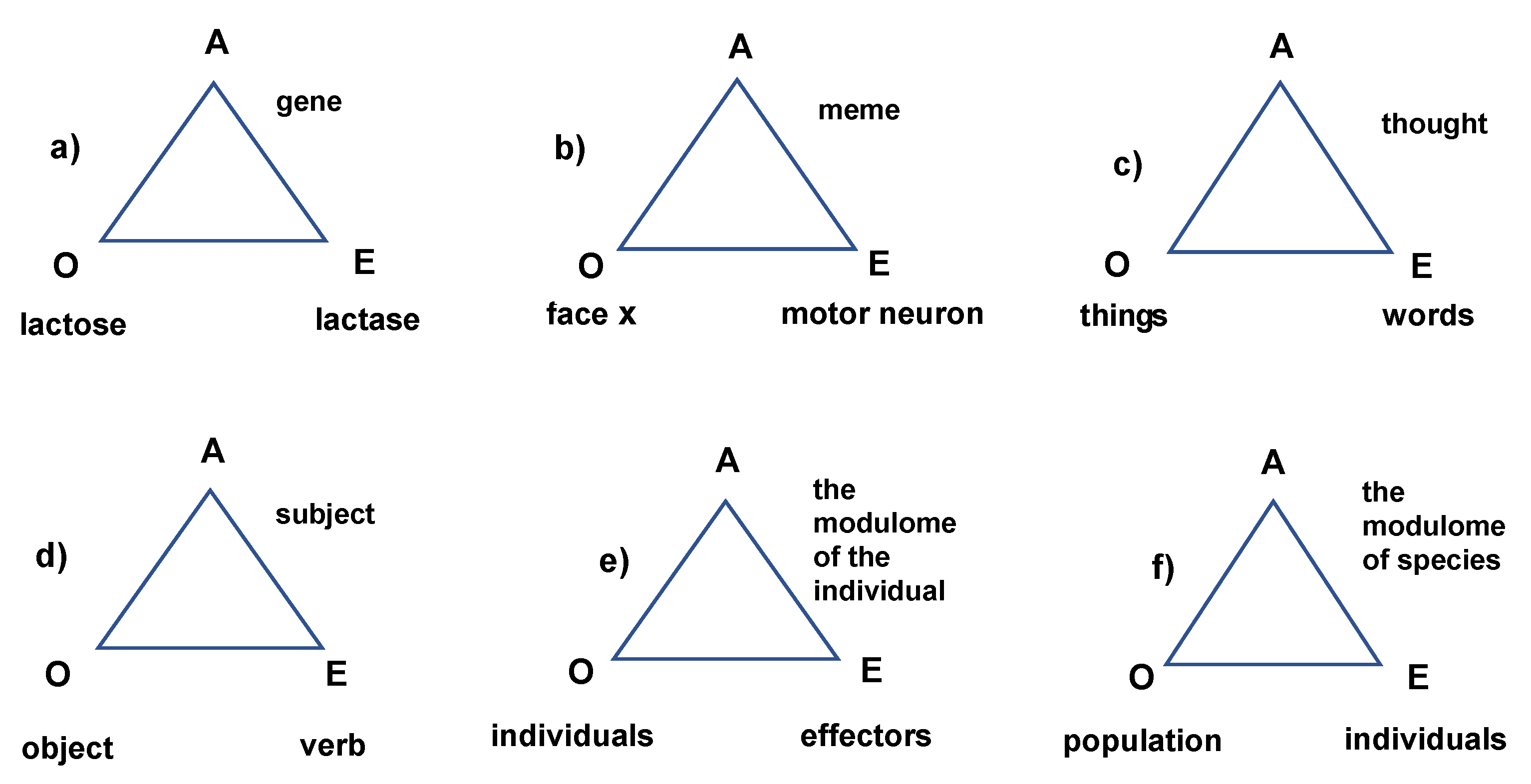
Figure 3.
Tree of Life, the construction of which is based on the types of interactions, information formats, and corresponding modules.
Figure 3.
Tree of Life, the construction of which is based on the types of interactions, information formats, and corresponding modules.
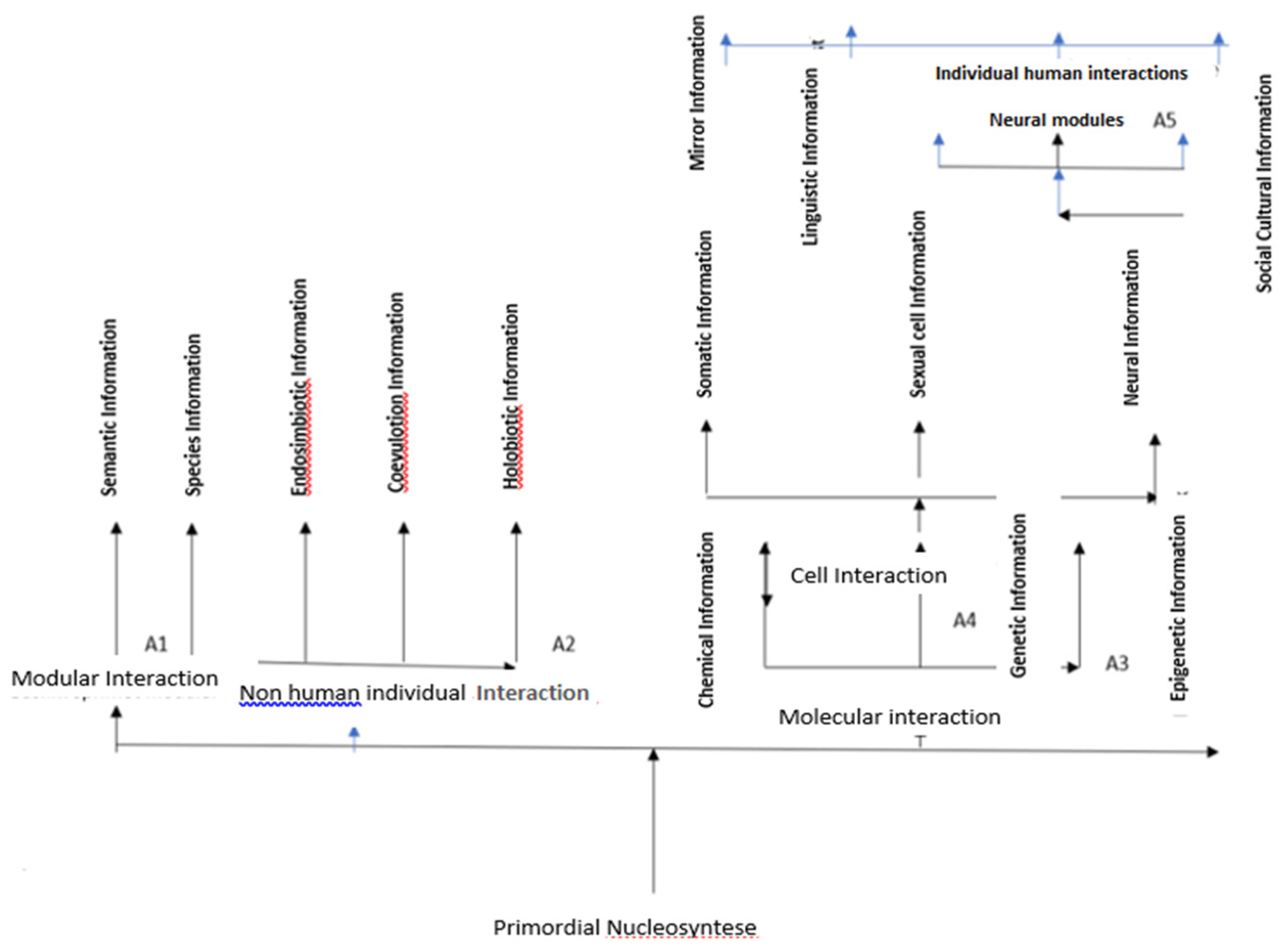
Figure 4.
The evolutionary modular model: energy (E), matter (m), information (i), natural selection (ns), self-organization (so), and heritability (h2).
Figure 4.
The evolutionary modular model: energy (E), matter (m), information (i), natural selection (ns), self-organization (so), and heritability (h2).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



