
Submitted:
21 June 2024
Posted:
24 June 2024
You are already at the latest version
Abstract
Chemical pollution poses a significant threat to human health, with detrimental effects on various physiological systems, including the respiratory, cardiovascular, mental, and perinatal domains. While the impact of pollution on these systems has been extensively studied, the intricate relationship between chemical pollution and immunity remains a critical area of investigation. The focus of this study is to elucidate the relationship between chemical pollution and human immunity. To accomplish this task, this study presents a comprehensive review that encompasses in vitro, ex vivo, and in vivo studies, shedding light on the ways in which chemical pollution can modulate human immunity. Our aim is to unveil the complex mechanisms by which environmental contaminants compromise the delicate balance of the body's defence systems going beyond the well-established associations with defence system delving into the less-explored link between chemical exposure and various immune disorders, adding urgency to our understanding of the underlying mechanisms and their implications for public health.
Keywords:
Environmental pollutant
; innate and adaptive immunity
; immunomodulation
; developmental toxicology
; pregnancy
1. Introduction
The exponential growth of the world economy and its accompanying exploitation of resources and industrial production have caused an unprecedented transfer of contaminants to the environment, with multiple impacts on health. Because these human-induced impacts on earth systems are so extensive, a new definition of this geological era has been proposed, the Anthropocene. Although the last few decades have been characterised by marked technological innovation that aims at the concept of sustainability as a way to improve the quality of the environment, anthropogenic activities continue to have a profound effect on human health, mainly due to poor remediation actions [1]. Humans are regularly exposed to many environmental chemicals that may have potentially toxic effects on health. These chemicals can enter the human body through various routes: ingestion of chemicals via the consumption of contaminated food or water, inhalation of airborne pollutants through the respiratory system, as well as dermal absorption through the skin.
The most recent WHO environmental burden of disease estimates that, every year, 13% of deaths in the 28 European Member States are attributable to environmental stressors [2]. In terms of the absolute number of deaths attributable to the environment, the European Environment Agency concluded that 90% of deaths attributable to the environment result from non-communicable diseases (NCDs), including cancers, cardiovascular disease, diabetes and chronic lung illnesses (EEA Report No 21/2019). The interplay between the immune system and non-communicable diseases is complex and multifaceted. Chronic inflammation, immune dysregulation, and lifestyle factors can significantly impact the prevention and management of NCDs. Understanding this relationship can provide insights into the prevention and management of NCDs.
The exposome concept was introduced in the field of epidemiology by Wild in 2005 to encompass “the totality of human environmental exposures from conception onwards, complementing the genome” [3]. Therefore, the exposome concept provides a description of lifelong (from the prenatal period) exposure history. It is now known that the exposome poses new challenges in assessing the relationship between exposure to environmental factors (pollution, climate change, lifestyle, and diet) and health. Indeed, there is increasing evidence that genetic variants explain a limited fraction of the variability in the risk of chronic diseases, as shown by studies in monozygotic twins, leaving a potentially large role for environmental exposures and interaction between environmental and genetic factors [4]. The molecular processes underlying human health and disease are highly complex. Often, genetic and environmental factors contribute to a given disease or phenotype in a non-additive manner, yielding a gene–environment interaction [5]. Epigenetic mechanisms, including DNA methylation, histone modification and non-coding RNA, can modulate gene expression levels without changing the underlying DNA sequence. Moreover, many epigenetic modifications are dynamic, reflecting cumulative environmental exposures throughout the lifespan and correlating with ageing related diseases and outcomes [6].
In 2012, Wild has enhanced his approach, describing three overlapping domains within the exposome that refer to different factors, such as: i) the general external environment (including factors such as the urban–rural environment, climate factors, social capital and education); ii) the specific external environment (including diet, physical activity, tobacco, infections, occupation, etc.); iii) the internal environment including internal biological factors, such as metabolic factors, gut microbioflora, inflammation, oxidative stress and ageing [7]. This, together with the results of studies on animal models, would suggest a pathological mechanism of “multiple hits'' [8]. According to this model, the rearrangement of gene expression through epigenetic mechanisms can involve different organ systems and is caused by a combination of genetic predisposition and prenatal injury (the "first-hit"), which may not be enough to change the adult phenotype on its own. However, tissue imbalances resulting from the perinatal insults and/or adverse stressors/exposures during postnatal life may serve as a "second-hit", which might reveal or accentuate the underlying abnormalities leading to disease states.
All chemicals found in the environment are referred to as pollutants or xenobiotics. Many of such chemicals can persist in the environment over long periods of time and, for this reason, are called persistent organic pollutants (POPs). POPs are characterised by ubiquity and persistence in the environment, and it is known that the derived products can interact with the environment and undergo biotransformation and bioaccumulation processes [9]. Among the anthropogenic environmental pollutants, in addition to certain natural compounds like heavy metals, pesticides , drugs, and synthetic chemical compounds generated during production processes (hydrocarbons, dioxins, polychlorinated diphenyls, polybromodiphenyl ethers, etc.) can contribute to the pathogenesis of several diseases in living organisms.
The immune system is an ancient defence system formed by all metazoans while struggling with various internal and external factors, whose perturbation may lead to increased susceptibility to pathogens and diseases. Inflammation is a common response to a variety of stressors, including xenobiotics. Under normal conditions, inflammation is a healthy and adaptive process that both combats infection and aids in repairing damage to tissues. However, xenobiotics can elicit prolonged, severe, and/or inappropriate inflammatory responses that play a causal role in the progression of biological events, linking a molecular initiating event to an adverse outcome pathway (AOP) [10]. Ordering relevant and causally linked events in the response cascade is critical for applying this information to Environmental Risk Assessment, which is why the AOP framework was created [11].
In this view, the immune system has a recognized central role in many processes involving chronic diseases, and the frontiers between communicable and non-communicable diseases also require additional exploration [12]. The recognition of altered immune system function in many chronic disease states has proven to be pivotal in non-communicable diseases that are characterised by a chronic low level of inflammation, including autoimmune [13] and neurodegenerative conditions [14], cardiovascular diseases [15], cancer [16], diabetes mellitus and Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD) [17,18]. Therefore, compared with other toxicity endpoints (i.e., genotoxicity, endocrine toxicity or developmental toxicity), there are still many challenges for the introduction of immunotoxicity endpoint evaluation [19].
So far, there is still insufficient knowledge about the effects of pollutants on the different cell populations of the immune system, and the knowledge of related toxicological mechanisms remains elusive. Many findings have suggested that a subset of environmental pollutants can bind the receptors involved in metabolism, oxidative stress, immune response and inflammation, such as the aryl hydrocarbon receptors (AhR) [20]. The complexity of AhR signalling arises from multiple factors, including the diverse ligands that activate the receptor, the expression level of AhR itself, and its interaction with the AhR nuclear translocator [21]. Indeed, AhR engages in crosstalk with the AhR repressor or other transcription factors and signalling pathways and, in this way, it can also mediate non-genomic effects [22].
In recent decades, AhR has been increasingly recognized as an important modulator of disease because of AhR’s role in the regulation of the redox system and of immune and inflammatory responses [23]. AhR has significant effects on the control of adaptive immunity, modulating T cell differentiation and function directly and indirectly through its effects on antigen-presenting cells. For instance, 2,3,7,8-tetraclorodibenzo-p-dioxin (TCCD) is a potent xenobiotic ligand of AhR and, from this, it was found to inhibit immune responses [24], showing an effect linked to the induction of CD4+ T cells with a regulatory phenotype [25]. Indeed, the ability to activate AhR has been demonstrated for other contaminants, such as polycyclic aromatic hydrocarbons (PAHs) [26] and polybrominated diphenyl ethers (PBDEs) [27]. The production of reactive oxygen species (ROS) and organic metabolites due to the dysregulation of this receptor can cause serious consequences on biological macromolecules such as proteins, lipids and DNA [28]. Indeed, oxidative stress, inflammation and apoptosis can compromise immune response, with the possible development of infections, tumours and lung diseases [29].
In this review, we analyse how environmental pollutants can modulate immune response, reviewing the biological effects of the most common environmental pollutants on different immune cells in in vitro, ex vivo and in vivo studies. Indeed, we will devote special attention to pregnancy, describing the effects of pollutants on immune response during different windows of prenatal life (Figure 1).
2. Pollutants and Immune System Modulation: In Vitro, Ex Vivo and In Vivo Models
A large set of environmental pollutants have been investigated for their impact on immune response. In this paper, we will review the main biological effects of a list of the most widespread chemicals that have been detected in the environment and whose transfer to humans has been documented. In these perspectives, we will focus on the effects of specific classes of toxicants on the immune regulation of human primary and/or immortalized cell lines and in animal models.
2.1. Metals
Heavy metals, such as arsenic, lead, mercury and cadmium, are a group of highly toxic chemical pollutants that have garnered significant attention due to their detrimental effects on the environment and human health. These elements, which occur naturally in the Earth's environment, can find their way into the environment through various industrial processes, mining activities, and the burning of fossil fuels (anthropogenic activities). One of the most concerning aspects of heavy metals as chemical pollutants is their persistence in the environment. These substances do not degrade over time, accumulating in soil, water, and air and consequently, they can enter the food chain, leading to bioaccumulation, where the concentration of these toxic metals increases as one moves up the food web [30,31,32]. Below, we describe the effects of some of the most common heavy metals analysing their effects in vitro, ex vivo and in vivo models.
2.1.1. Arsenic
Several national and international organizations have classified Arsenic (As) as the most toxic and carcinogenic inorganic environmental pollutant. It exists in nature in both organic and inorganic form. The inorganic form is more toxic and accumulates in exposed organisms; it is found in the environment in the form of arsenite (AsIII) or arsenate (AsV); The main route of arsenic exposure is the consumption of contaminated drinking water. Many studies have found a positive relationship between arsenic (As) exposure and the onset of human pathologies including cancer (lung and bladder) and liver, respiratory, cardiovascular and immune diseases [33,34]. In terms of immune system dysregulation, it has been demonstrated that As reduces either the expression level of cytokines (IFN-γ, IL-4 and IL-10) [35,36,37] and induces apoptotic mechanisms in different cell types, such as B and T lymphocytes, macrophages and neutrophils [38,39]. It has been shown that As can drive M2 polarization of macrophages, triggering the CD206 marker, TGF-β1 and Arg1 genes that have been shown to be involved in fibrinogenic process [40]. Arsenic-induced M2 polarization is also favoured via miR-21 regulation of phosphatase and tensin homologue deletion on chromosome ten (PTEN), implicated in chronic hepatic fibrosis and in the development of arsenicosis [41].
Microarray analysis of peripheral blood mononuclear cells (PBMCs) purified from subjects exposed to increasing As concentrations has also shown that this pollutant modulates T cell receptor expression, the cell cycle and apoptotic processes. Furthermore, As increases the expression of inflammatory molecules such as cytokines or growth factors [42] and induces the hypomethylation of leukocyte DNA, which is associated with an increased risk of inflammatory skin lesions [43]. Arsenic has also a negative effect on innate and humoral immunity, altering macrophage and B lymphocyte functions and stimulating oxidative stress signalling, DNA damage and cytotoxicity in T cells and in human polymorphonuclear neutrophils [44,45,46].
2.1.2. Lead
Lead (Pb) is available in nature in relatively low amounts. Anthropogenic activities, such as manufacturing, mining, and fossil fuel burning, have contributed to increase the release of Pb in the environment. Lead induces toxic effects on various tissues of the human body, including the immune system, causing increased allergies, infectious and autoimmune diseases [47,48]. For instance, in vitro studies have shown that Pb treatments induce the dysregulation of proinflammatory cytokine production [49,50,51] and impair THP-1 monocyte/macrophage cell viability at low concentrations. Zhao and coworkers showed that Pb exposure induces the quiescence of haematopoietic stem cells through Wnt3a/β-catenin signalling and decreases the expression of CD70 on bone-marrow-resident macrophages [52]. These observations were confirmed by ex vivo studies on a population of chronically exposed workers in which higher levels of IFN-γ, IL-2, IL-12 and IL-17 were observed [53,54]. Lead can cross the placenta [55] and can have adverse effects on birth outcomes, possibly by accumulating in the placenta and causing reduced nutrient transfer and oxidative stress and abnormal functioning [56].
2.1.3. Mercury
Mercury (Hg) is a highly toxic metal, and exposure to its organic and inorganic chemical compounds causes adverse effects on humans, inducing genetic damage, neurological, kidney, cardiac and immunological diseases [57]. The predominant toxic Hg forms include: elemental Hg (Hg0), ionic Hg and organic (o)Hg, such as methylmercury (MeHg), which is classified as the most toxic among them.
In the innate immune system, Hg can influence the activity of different cell subtypes, such as neutrophils, monocytes/macrophages, NK cells and dendritic cells. It has been shown that in vitro exposure of neutrophils at low concentrations (≤5 μM) of HgCl2 causes an increase in the production of superoxide ion [58], inhibiting the apoptosis of neutrophils, while at higher concentrations, Hg displays cytotoxic effects. At the same concentrations of pollutant, macrophages show functional impairment of both phagocytic and migratory activity [59,60], showing a reduction in NO production and a greater synthesis of inflammatory cytokines, such as IL-6 and TNF-α. Moreover, after Hg exposure in co-culture assays of intestinal epithelial cells and THP-1 macrophages, mimicking the gastrointestinal tract, the metal induced a pro-inflammatory response with an increase in IL-8 and IL-1β cytokine production [61].
Mercury derivatives also show genotoxic effects in vitro and in vivo, in terms of both chromosome aberrations and loss of cell proliferative capacity, compromising the metabolic, immune and nervous systems [62]. Dietary MeHg intake (3.9 μg/gram) by mice and rats resulted in a 42-44% suppression of the tumoricidal activity of blood and splenic NK cells, as well as the proliferation of T and B cells of the adaptive immune system [63,64]. These events have been associated with an increase in intracellular oxidative stress [65,66] which causes the activation of transcription factors and the expression of genes encoding pro-inflammatory cytokines.
2.1.4. Cadmium
Cadmium (Cd) is a heavy metal naturally present in the environment (in the soil, water and air) and is mainly used in the steel industry, in plastics and as a component of batteries. It is released into the environment through industrial and domestic activities, such as the combustion of fossils fuels (coal, diesel, gasoline, etc.), the incineration of industrial waste (in particular batteries and plastics containing Cd), the production of metal alloys and the manufacture of fertilizer phosphates. Food, drinking water and the inhalation of cigarette smoke, both in active and passive form, are sources of this heavy metal. Target organs for Cd-induced toxicity include the liver, kidneys, lungs, testes, prostate, heart, skeletal system, nervous and immune system.
Following environmental and/or industrial exposure, it has been shown that Cd induces inflammation and oxidative damage in neutrophils and macrophages, showing an increase in the production of ROS [67,68]. These results have been further confirmed by experimental studies carried out on rodent macrophages and macrophage cell lines [69]. Cd-induced M1 polarization has been observed in in vitro and in vivo rodent macrophages via the JAK2/STAT3 pathway, contributing to the atherosclerotic process [70]. Furthermore, it has been shown that intrauterine exposure to Cd can provoke a dramatic decrease in the levels of IFN-γ and IL-2 cytokines in offspring and affects the activity of adaptive immunity cell populations in C57Bl/6 mice. In female offspring, the CD4+ and CD8+ T cell populations increase after exposure to Cd, while the percentage of NK cells and granulocytes decreases. In addition, a decrease in splenic Treg cells has been shown in both male and female offspring [71,72]. Another study in wild boars highlighted that Cd downregulates the expression of different pro-inflammatory cytokines (TNF-α, IL-12p40, several TLRs, CD14, MD2, BD2, MyD88, p65 and NOS2) in macrophages, negatively affecting their immune role [70].
In a study on workers occupationally exposed to Cd, epigenetic alterations in the expression of miRNAs associated with inflammation, carcinogenesis and their possible correlation with immune profile were evaluated. MiRNA profiling showed that miR-221 and miR-155 were modulated in exposed workers with reference to a higher percentage of the Th17 population [73] (See Table 1 for summary).
2.2. Persistent Organic Pollutants (POPs)
Among the most dangerous organic pollutants for health, persistent polyhalogenated organic compounds or POPs (so-called because they withstand biochemical and photolysis processes by remaining in the environment for a long time) [74] are very stable chemicals whose lipophilic nature determines their bioaccumulation in animal adipose tissues, thus allowing their biomagnification along the food chain. Due to their chemical-physical characteristics, POPs can cross the placenta, causing exposure starting from intrauterine life, but also can accumulate in breast milk, which is particularly rich in lipids, determining their intake in post-natal life [75]. The most common POPs are polycyclic aromatic hydrocarbons (PHAs), organochlorine pesticides (OCPs), polychlorinated biphenyls (PCBs), perfluorinated compounds (PCFs), dioxins and brominated flame retardants (BFRs) that will be discussed in dedicated sections.
2.2.1. Dioxins
Dioxins are highly toxic POPs, produced from natural processes like forest fires as well as the burning of trash by anthropic activities. Due to their highly lipophilic nature, they undergo biomagnification along the food chain to eventually accumulate in human tissues. Of all the analysed congeners, 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) showed the highest toxicity for living organisms [76]. Several in vitro, ex vivo and in vivo studies have been conducted to determine the adverse effects of dioxins on the various components of the immune system, with immune-suppressive effects. Regarding innate response, a compromise of the defence system against pathogenic organisms and cancer cells was observed; in particular, TCDD impairs the correct functioning of macrophages, NK cells, neutrophils and dendritic cells [77,78,79]. In adaptive response, dioxin compromises the production of antibodies by B lymphocytes and the cytotoxic activity of T lymphocytes and also attenuates the IgE mediated hypersensitivity response [80,81]. It has been observed that, in THP-1-derived macrophages, TCDD treatment induces alterations in adherence, adhesion molecule expression, morphology, multiple cytokine/chemokine production and the expression of total mRNA [82]. Few studies have focused on TCDD’s multigenerational and transgenerational effects on human reproductive health, despite the high amount of evidence in animal models of such effects on male and female reproductive health [83].
2.2.2. Polychlorinated Biphenyls (PCBs)
Polychlorinated biphenyls are environmental pollutants of industrial origin. Although their production has been interrupted many years ago , they persist in the environment due to their resistance to chemical, physical and biological degradation processes. Like other organic pollutants, they accumulate in soil and can enter the food chain reaching humans. Numerous studies have shown that PCBs can alter the proper functioning of the immune system at several levels. For example, in macrophage cell lines, treatment with PCB 126, a dioxin-like polybromophenyl, has a proinflammatory immunostimulating activity [84]. In vitro, the congeners non-dioxin-like polychlorinated biphenyls (NDL-PCBs), NDL-PCB 153 and NDL-PCB 180 display an immunosuppressive effect by reducing the expression of the proinflammatory cytokines TNF-α and IL-6 and the expression of reactive species [85]. PCBs are also capable of altering the physiological antiviral cellular response [86]. In an in vivo mouse model, PCB promoted an alteration of collagen in the female bladder compared to the male one, indicating an inflammatory event that results in the impairment of immune system functions [87]. Moreover, studies from PCB-exposed populations have revealed DNA methylation differences in CpG sites with effects in oestradiol CpG site sizes and affecting the immune system [88].
2.2.3. Brominated Flame Retardants (BFRs)
Brominated Flame Retardants (BFRs) are chemicals used in plastics synthesis, textile products and electronic and electrical devices; they are added to industrial products to make them less flammable. It has been shown that the flame retardants contained in various products are released into the environment through combustion, percolation, and degradation [89]. Due to their increasing use, BFRs have become globally widespread pollutants over the years. They have been found both at the level of different environmental matrices (air, aquatic sediments, soil, dust, etc.), and in animal organs and tissues, such as blood, mother's milk and adipose tissue [90,91].
There are four main classes of BFRs: 1) polybrominated diphenyl ethers (PBDEs), present in plastics, textiles, electronic devices and circuits; 2) hexabromocyclododecanes (HBCDDs) used for thermal insulation in buildings; 3) tetrabromobisphenol A (TBBPA) and other phenols used in thermoplastics and televisions; 4) polybrominated biphenyls (PBBs) used in consumer devices and textiles [92]. The immunotoxic effects of the most studied flame retardants will be discussed below.
2.2.3.1. Polybrominated Diphenyl Ethers (PBDEs)
Polybrominated diphenyl ethers are chemicals consisting of two aromatic rings joined by an ester bond with bromine atoms. The commercial use of PBDEs dates back to 1976 and the three most used blends were penta-BDE, octa-BDE and deca-BDE. Since 2004, the production of penta and octa-BDE has been banned, both in the United States and in the European Union, due to their negative effects on the environment and health. The deca-BDEs have been banned in the United States since 2010. On the basis of the number and position of the bromine atoms, we can distinguish 209 congeners with different chemical-physical characteristics that affect their capacity for bioaccumulation and toxicity in the liver, kidney, gut and thyroid in terms of oxidative and mitochondrial damage and apoptosis [90,93]. Human population studies have shown that PBDEs accumulate in adipose tissue, hair, human serum, breast milk and placenta. Among the most toxic and widespread congeners in the environment, BDE-47 (a tetra-BDE) and BDE-209 (a deca-BDE) are the most relevant in terms of health risks. Experimental data on animal models have shown that BDE-47 can have neurotoxic, cardiotoxic, hepatotoxic and teratogenic effects on zebrafish larvae and adults and in fish [94,95,96].
Several studies have investigated the immunotoxic effects of PBDEs. In in vitro studies, BDE-47 significantly reduced the expression and secretion of the proinflammatory response M1 macrophage marker genes IL-6, TNF-α and IL-1β and the expression of metalloelastase 12, a gene essential in the motility of the macrophage, recruitment of neutrophils and release of cytokines and chemokines in inflammatory processes [97]. Indeed, the same authors showed that BDE-47 can have diverse action mechanisms on human macrophage cell lines affecting small extracellular vesicles biogenesis [98] and the content of their miRNA cargo. Furthermore, they demonstrated that BDE-47 treatment can regulate the sEVs’ cargo with purposeful consequences toward downstream events and target bystander macrophages. Exacerbating the macrophage LPS-induced pro-inflammatory response [99] as well as altering A549 epithelial lung cells which modulate mRNA expression of tight junctions, adhesion molecules, cytokines and EMT (epithelial-mesenchymal transition) markers [100]. Finally, it has been shown that BDE-47 can also induce cardiovascular toxicity, activating PPARγ in THP-1 monocytes, inducing the foam cell formation typical of a proatherogenic process [101]. In an animal model, it was observed that they stimulate an increase in the production of ROS in macrophages, leading to the remodelling of the cellular physiology of phagocytes and, therefore, to the destruction of the functional activity of the macrophage [102,103,104]. Recently, Balb/c mice treated with different doses of BDE-47 by gavage showed that BDE-47 significantly reduced antibody response and induced histopathological effects on the liver, spleen, small intestine and thyroid, in the absence of general toxicity, suggesting that exposure to BDE-47 may perturb innate and adaptive immune responses [105].
2.2.3.2. Hexabromocyclododecane (HBCDD) and Tetrabromobisphenol A (TBBPA)
Hexabromocyclododecane (HBCDD) is a non-aromatic brominated cyclic alkane; it is mainly used in expanded propylene insulation foam [106,107], but also in tapestry fabrics, curtains and wall coverings [108]. HBCDD is added to plastic material but, since it is not chemically linked to it, it can transfer into the environment and accumulate in soil, sediments, and dust particles. It is involved in the trophic chain to humans through the ingestion of dust [109,110], bioaccumulating in blood, adipose tissue and breast milk [111,112,113]. Immunotoxicity studies on rats have highlighted that HBCDD reduces splenocytes and increases IgG and neutrophils, contributing to immune system imbalance [114]. Tetrabromobisphenol A (TBBPA) is a flame retardant used in thermoplastics production. At the cellular level, Barańska and collaborators found that TBBPA has a genotoxic mechanism because it causes oxidative damage in purines and pyrimidines in PBMCs [114,115,116]. Furthermore, a study conducted on human dendritic cells isolated from healthy subjects by Canbaz and collaborators showed that exposure to this flame retardant can induce an inflammatory phenotype typical of Th2 response [117]. Furthermore, for the same substance, neurotoxic, nephrotoxic, hepatotoxic and immunotoxic effects have been reported which can induce ROS expression, promoting the production of inflammatory cytokines and compromising mitochondrial function [118]. A correlation has also been demonstrated between human exposure to both classes of pollutants and thyroid and neurological disorders, reproductive health, immunological, oncological, and cardiovascular diseases [119,120].
2.2.4. Bisphenol A
Bisphenol A (BPA) is a chemical that has been synthesized since the 1960s and is widely used in industrialized countries. BPA is used in the production of polycarbonate plastics, in food-grade containers and in epoxy resins that make up the internal protective coating found in most food and beverage cans, and it is also used for dental devices and in thermal receipt paper. Under the pressure of EFSA, between 2010 and 2015, many studies have been conducted which are aimed at understanding the possible effects of BPA on immune system cells and tissues. Balistreri and collaborators showed that BPA alters the function of neutrophils in a dose-dependent manner, causing an increase in ROS production in these cells through a calcium-dependent process. Bisphenol A reduces both the chemotactic ability of neutrophils towards other cell types of the immune system, and the ability of neutrophils themselves to eliminate pathogens; these data therefore suggest that BPA has the ability to compromise some fundamental functions of innate immune response [121]. Bisphenol A also promotes the polarization of M1 macrophages in terms of cell number, gene expression of CD11c and iNOS markers, and the production of pro-inflammatory cytokines via the upregulation of transcription factor IRF5 expression involved in the polarization process [122]. The upregulation of pro-inflammatory cytokines was also induced in the murine macrophage cell line RAW264.7 by the BPA analogue BPF (4,4'-Methylenediphenol) and, together with the involvement of the JAK2/STAT3 pathway, they contributed to M1 polarization [123,124].
BPA also showed immunotoxic effects on adaptive immune response. An in vivo study conducted on mice showed that animals treated with a single dose (250 µg/kg) of BPA in the neonatal period and subsequently subjected to the induction of breast cancer had no significant changes in the population of B lymphocytes nor damage to the spleen or peripheral lymph nodes, but instead showed a significant difference in IgM reactivity capable of recognizing tumour antigens decreased in guinea pigs [125].
2.2.5. Perfluorooctanesulfonic Acid and Perfluoroctanoic Acid
Perfluorooctanesulfonic acid (PFOS) and perfluoroctanoic acid (PFOA) are perfluoroalkyl organic substances (PFAS). They are chemical compounds containing long carbon chains that are used to increase resistance to high temperatures in numerous products, such as fabrics, carpets, clothing, food grade paper, non-stick cookware and firefighting foams. PFOA and PFOS remain in the environment for a long time and, due to their chemical stability, resist degradation processes, accumulating in the soil, air and water. PFAS are able to provoke the release of the pro-inflammatory cytokine IL-1β and caspase-1 in THP-1 macrophages; they also activate the innate immune response through the AIM2 receptor of inflammasome, inducing inflammation in the lung, liver and kidneys of mice [126]. Population studies have shown that PFAS can accumulate in human organs and tissues; for example, PFOS has been found in the liver, kidneys, lungs, hair, breast milk and urine, but it accumulates predominantly in the blood [127,128]. In vitro and in vivo data have shown that PFOSs and PFOAs have toxic effects on multiple cell types and organs of the immune system. An in vitro study showed that treatment of human T cells with increasing concentrations of PFOS induced a reduction in the expression of IL-2, a cytokine essential for the correct functioning of leukocytes and whose reduction was observed in various autoimmune diseases [129]. An in vivo study conducted in mice showed that PFOS treatment inhibits T cell proliferation. The analysis of the biochemical pathways involved in immune cell signalling revealed the inhibition of genes involved in cell cycle regulation and response to oxidative stress [104]. In vivo studies in mice performed by Torres and collaborators showed that PFOS exposure reduced immune cell populations in some organs and also led to an increase in the number of cells in others, suggesting a possible de novo localization in cells and a decrease in the activity of some organs like the spleen and liver [130]. Taylor and coworkers showed that PFOA exposure can reduce B-cell subtypes and, consequently, IgM antibody primary response in female mice[131]. See (Table 2 for a summary).
The Figure 2 displays the similarities and the differences among the main classes of environmental pollutants in the modulation of immune response.
3. Developmental Immunotoxicology
The term developmental immunotoxicology (DIT) has been used to identify a new approach to understanding the effects of immune development perturbation on adverse outcomes [132]. DIT addresses the impact of xenobiotics on immune responses during the early stages of immune system development/maturation compared to adults. Immuno toxicology studies have defined some critical windows of vulnerability during development [133] where xenobiotics can exert their effects , but sometimes stressors may be applied during rather broad periods which often cover the entire immune developmental period, such as in highly polluted areas where environmental stressors can be relevant for a large period of prenatal and post-natal life. Furthermore, considering the accumulation of evidence supporting the hypothesis of the Developmental Origin of Health and Disease (DOHaD) [134,135], disturbances of the immune system during foetal development are of particular interest for possible long-term consequences. According to epidemiological studies, adverse effects of in utero exposures on the immune system have been associated with higher rates of chronic immune disorders in adults, including autoimmunity, immune deficiency, inflammation and allergic reactions [136].
Although these associations are of considerable importance, examining only outcomes directly linked to immune system elements fails to capture the complete range of health risks associated with DIT. In fact, from the early stages of life, immune cells are found in all tissues and organs where they serve as homeostatic regulators of the physiological function of the tissue and as sentinels to defend it. In the liver, for example, the functional state of hepatocytes (the majority of cell populations) can be significantly altered by a small number of resident immune cells (i.e., Kupffer cells). This occurs similarly between microglia and astrocytes in the central nervous system and many other organs [137]. For these reasons, the involvement of DIT events underlying behavioural disorders and metabolic dysfunction has been suggested [138].
During pregnancy, major adaptations occur in the maternal immune system to protect the mother and her future baby from pathogens; therefore, the perturbation of this balance may have detrimental immune responses against the allogeneic foetus. Pro- and anti-inflammatory stimuli alternate during gestation, and their regulation is the basis of the success of numerous key steps of the pregnancy. For example, the implantation of the embryo on the uterine wall is facilitated by a period of active inflammation, essential for the remodelling of the maternal uterus [139]. On the contrary, the phases of active foetal growth and development are accompanied by a relative inflammatory quiescence [140]. Failure to induce these systemic changes predisposes women to adverse pregnancy outcomes, such as recurrent miscarriage [141], preeclampsia [142] and preterm birth [143]. Additionally, depending on the duration and severity, this inappropriate activation of the maternal immune response may have transgenerational consequences. For instance, the activation of the mother's immune/inflammatory response has been linked to negative neurobehavioral outcomes in offspring [144,145,146,147]. These findings are noteworthy from an immunological standpoint, as the physiological development of the foetal brain involves a variety of immunological factors at the maternal-foetal interface [148]. For example, in the case of extremely preterm infants, placental methylation of genes related to inflammation has been observed to be associated with a decreased risk of cognitive impairment and decreased neonatal systemic inflammation [149].
Evidence suggests that prenatal exposure to xenobiotics may play a role in altering the epigenetic regulation of immune-related genes that influence the development of regulatory T cell (Treg) networks or the ratio of Th1 and Th2 cells. Because Th1/Th2 immunity is closely linked to disease, epigenetic changes caused by an adverse intrauterine environment could explain the occurrence of disease susceptibility later in life [150]. In parallel with the direct modulation of epigenetic regulation, numerous toxic substances can impact the endocrine system. Given its close collaboration with the immune system in guiding development from gestation to early childhood, endocrine disruptor chemicals may also trigger DIT [151].
In this section, we will overview the effects of the most relevant xenobiotics during pregnancy with specific reference to the immune dysregulation.
Arsenic toxicity has been linked to many possible pathways, including oxidative stress, epigenetic modification, interference with DNA repair mechanisms and alterations of the immune system [152,153]. Exposure to As during pregnancy can result in DIT interacting directly and indirectly with the foetal environment. In both cases, the interference is strongly related to the placental function that is permeable to As while also being particularly susceptible to its detrimental consequences, including oxidative stress, inflammation and immune system perturbation [33]. In two separate cohort studies from Bangladesh and United States, prenatal As exposure was associated with lower percentages of CD4+ T cells and, in particular, activated memory CD4+ T helper cells in cord blood [154,155]. Although a direct comparison of exposure levels between the two populations is not possible, the effects of As exposure appear to be consistent. In the study on the Bangladeshi population, As exposure is determined from the levels found in drinking water samples, whereas in the U.S. study, exposure is assessed from maternal toenail samples. Evidence of the similarity in findings showed that both cohorts demonstrated relatively higher susceptibility to infectious diseases at different stages of childhood, which was associated with prenatal arsenic (As) exposure [156,157]. This suggests that prenatal As exposure causes overall immunosuppression in offspring. Among the mechanisms of action that could explain the negative effects of As exposure on immune system cells, the most widely accepted appears to be the induction of oxidative stress. Arsenic exposure has been shown to trigger oxidative stress in peripheral blood, in both adults and children [158,159]. Studies conducted on populations exposed to high levels of As have demonstrated that prenatal exposure to arsenic increases the expression of numerous markers of inflammatory and oxidative stress in umbilical cord blood and placenta [33,160].
The long-term effects on the immune system of arsenic exposure during pregnancy may be mediated by epigenetic mechanisms. Specifically, exposure to arsenic has been shown to induce alterations in DNA methylation patterns, which can influence gene expression and immune cell function [161,162]. Studies on mice have suggested that one possible mechanism linking prenatal As exposure to health effects later in life may be epigenome alteration. Cytosine methylation (e.g., CpG methylation) has been suggested as a valuable candidate for As-related health effects [163].
The toxic effects related to Hg exposure, in particular to methylmercury (MetHg), mainly affect the nervous system [164]. However, as previously mentioned, in vitro experimental evidence showed possible associations between Hg exposure and immunotoxicity [61,65,67]. Mercury can be transferred from mother to foetus through the placenta [164], and in utero exposure to Hg has been associated with a higher risk of respiratory infection in infants during the first year of life [165]. Mercury levels during pregnancy and postpartum have been linked to the frequency of natural Treg and NKT cells as shown in the New Hampshire Birth Cohort Study. Higher natural Treg counts have also been associated with an increased risk of Hg exposure as measured by seafood consumption. Elevated Tregs may represent a counterbalancing mechanism in response to the increased autoimmune pressure associated with low-level Hg exposure [166]. In a study focused on a highly exposed population from the Faroe Islands, Oulhote and colleagues reported a correlation between prenatal methylmercury (MetHg) exposure and a decreased total white blood cell count, specifically affecting monocytes, basophils, CD3+ T and CD4+ T lymphocytes, and CD19+ B lymphocytes at the age of 5 [167]. It could be hypothesized that such an alteration in the balance of white blood cells could affect inflammatory cytokine levels. In this view, a multicentric cohort study conducted in five European countries (France, Greece, Norway, Spain and the United Kingdom) showed that in utero exposure to Hg was correlated with poorer childhood metabolic status and higher levels of TNF-α, IL-6 and IL-1 at 6 and 12 years. In this report, in fact, the authors suggested that changes in key inflammatory cytokines and the metabolic profile are strictly interconnected [168]. In an Amazonian Brazil population exposed to high levels of mercury, a positive association between mercury levels and serum concentration of antinuclear antibodies was found, suggesting a possible role of mercury in autoimmune disease [169]. In a previous study conducted in the same population, Pinheiro and colleagues reported elevated glutathione levels and decreased catalase activity in response to Hg exposure [170]. Mercury's capacity to modulate the synthesis and activity of enzymes within the endogenous antioxidant system may underlie its effects on immune cells. By disrupting the normal function of these enzymes, mercury can impair the antioxidant defences of immune cells, resulting in altered immune responses and increased vulnerability to infections and diseases.
Studies on the effects of Pb during pregnancy suggest that exposure to the metal can lead to adverse outcomes, such as an increase of enzymes involved in oxidative stress, tissue damage and the dysregulation of inflammatory pathways [171], and a polarization towards a Th2-type response with elevated serum IgE levels [51,172]. Indeed, lead exposure can affect thymic function, altering the development and maturation of immune cells [173] . This can cause an imbalance in immune functionality, shifting towards a more pro-inflammatory response and increasing the susceptibility to allergies and autoimmune diseases. In utero Pb exposure has been identified as a risk factor for childhood asthma later in life [47]. Indeed, Pb-related immunomodulating effects have also been highlighted by cross-sectional studies that revealed associations between blood Pb levels and biomarkers linked to allergy and infectious diseases in children [174,175].
Cadmium exposure induces oxidative stress by generating ROS, through which it exerts its toxicity. Cd exposure can occur through the consumption of contaminated food as well as through the inhalation of cigarette smoke. Smoking pregnant women have considerably greater amounts of Cd in their blood, placenta and umbilical cord blood. The concentration of Cd was associated with a higher expression of miRNA in cord plasma [176]. Cd exposure was associated with respiratory symptoms in adolescents. One proposed mechanism of action is increased vulnerability to acute respiratory infections in the early years of life [177,178]. The accumulation of Cd in the placenta has been proven in both in vitro and in epidemiological studies [179,180]. Cadmium concentrations have been analysed in cord blood, maternal blood and placental tissue, with demonstrated Cd-induced oxidative stress that adversely affects birth outcomes [181]. Sanders and co-workers found a significant association between in utero Cd exposure and DNA methylation patterns in the leukocytes of newborns and their mothers. It is worth noting that the methylation pattern was Cd specific; in fact, when compared to those produced using cotinine rather than Cd levels, the methylation patterns were non-overlapping [182]. Cadmium levels in the urine of pregnant women were associated with a lower absolute number of CD3+ and CD4+ lymphocytes and lower levels of IL-4 and IL-6 in girls, while an inverse association with the absolute count of CD3+ and CD8+ was only seen in males, according to a recent cohort study done in Wuhan, China [183]. Similarly, a sex-related effect was noted in a study conducted by Nygaard and colleagues. They reported that the concentration of Cd in maternal nails, sampled immediately after the delivery, was associated with a decreased number of T helper memory cells in cord blood [155]. Moreover, studies in rats have demonstrated that the sex-specific effects of cadmium on immune system cells might result from its interaction with sexual hormones, particularly 17β-estradiol [184].
Exposure to PFAS and PFOA has been associated with reduced immunogenicity of vaccines in offspring in two independent cohort studies from Norway and the Faroe Islands [185,186,187]. In the cohort of Faroese children, prenatal exposure to MetHg, PCBs and PFASs was also associated with higher levels of autoantibodies [188]. However, the authors concluded that more comprehensive cohort studies are needed to determine whether the presence and concentrations of these autoantibodies can predict the occurrence and severity of autoimmune diseases, or whether their presence should be considered merely a consequence of tissue damage.
PFAS exposure during pregnancy has also been linked to increased risks of asthma and respiratory syncytial virus infection during childhood [189]. On the same line, a study conducted among the National Health and Nutrition Examination Survey (NHANES) survey reported an increased risk of asthma in adolescent exposed to high concentration of PFOA [190,191]. Moreover, Wang and colleagues found a positive relationship between maternal levels of PFOA and PFOS and IgE levels in the cord blood. Interestingly, the authors do not report any association between exposure levels and asthma symptoms when models are corrected for other possible confounders [192]. However, it is relevant to report that several other studies demonstrated an inverse association between PFAS exposure and the risk of Eczema and Rhinitis [193].
Epidemiological studies in birth cohorts have shown that newborns exposed to high doses of PCBs in maternal blood, or children exposed to higher levels of PCBs in early childhood, have reduced thymus size, increased onset of respiratory disease and reduced immune response following vaccination [194,195]. PCB exposure during pregnancy has been linked to abnormal cellular immunity development at 6 and 16 months after birth: subjects most exposed to PCBs had significantly higher expression of CD3+ T-lymphocytes, B-lymphocytes, and activated B-lymphocytes, while NK cells were less expressed. In the same study, altered serum immunoglobulins were found in subjects exposed to higher levels of PCBs in utero compared to those less exposed [196].TCDD exposure during pregnancy has been observed to impact the miRNA profile of the foetal thymus, affecting the regulation of a wide number of genes that may impair immune system development [197]. TCDD also has a harmful effect on reproductive health, inducing epigenetic modifications in both human germlines, such as DNA methylation, suggesting that, if TCDD exposure happens during the initial germ cell development, the alteration can be transmitted to subsequent generations [198]. Few studies have focused on TCDD’s multigenerational and transgenerational effects on human reproductive health, despite the quantity of evidence of such effects on male and female reproductive health in animal models. These studies show that paternal ancestral TCDD exposure substantially contributed to pregnancy outcome and foetal health, although pregnancy outcome was considered tightly related to the woman's health [198].
Cases of occupational occurrence of allergic contact dermatitis in workers exposed to plastic based on BPA, and in some cases to bis-phenol F (BPF), have been reported, and for this reason such compounds are classified as highly allergenic [199]. Several human birth cohort studies have reported an association between prenatal BPA exposure and allergy symptoms such as asthma and wheezing in children of different ages [200,201]. Although many articles mention allergenic effects as an additional way to demonstrate the immunotoxicity of BPA, there are currently few studies investigating the sensitization effects of BPA analogues (i.e., BPF and bisphenol S [BPS]). Therefore, Mendyet and coworkers analysed the NHANES data and found a positive correlation between urinary BPF levels and current asthma and hay fever, while BPS was associated with a higher likelihood of asthma in men [202].
In the United States, blood PBDE levels range from 30-100 ng/g of lipids in adults, but the alarming health concern was mainly based on children who showed blood PBDE levels 3 to 9-fold higher than adults. PBDEs disrupt endocrine, immune, reproductive and nervous systems. Studies performed on the Boston Birth Cohort provided evidence that in utero exposure to PBDEs may epigenetically reprogramme the offspring's immunological response through promoter methylation of a proinflammatory gene [203]. These data agree with the in vitro results using macrophage-like cell lines, where cytokine production and miRNA expression were modulated by BDE-47 treatment [97,98,99].
It is becoming clear that DIT and prenatal programming may have a crucial role in raising the risk of infectious and noncommunicable disease incidence; thus, greater efforts are needed to understand the events leading to DIT in order to effectively counteract them.
An initial line of intervention could be to evaluate populations exposed to various concentration ranges by standardizing exposure variables and identifying priority outcomes for multicenter studies. Most of the studies mentioned in this paragraph examine the association between exposure and immune system outcomes in highly exposed populations. While these studies provide valuable insights into the effects of high levels of exposure, they do not fully reflect potential health risks. The immune system responses and health outcomes observed at high exposure levels might differ significantly from those at lower levels, potentially leading to an underestimation or overestimation of risks. For example, focusing on highly exposed populations might cause studies to miss subtler effects that occur at lower, more common exposure levels. Conversely, highly exposed populations also represent an important focus for studying co-exposures.
For instance, in a cohort of pregnant women living in a highly contaminated area, Longo and coworkers studied the effects of simultaneous exposure to a multi-pollutant mixture using a WQS regression model, demonstrating the association between the expression levels of immune relevant miRNAs and levels of a suite of inorganic and organic elements in the sera of pregnant women. The study emphasized the concurrent assessment of essential elements, recognizing their indispensable role in maintaining physiological balance. This approach allowed the interplay between exposure to environmental pollutants and the availability of essential elements to be unravelled, shedding light on potential synergistic or antagonistic effects, with particular reference to epigenetic alterations inherent in the maintenance of the redox state and cellular homeostasis during pregnancy [204].
4. Conclusion
The intricate relationship between chemical pollution and immunity underscores the profound impact of environmental factors on human health. In an era of an increasingly polluted world where chemicals pervade our air, water, food and soil, safeguarding the resilience of the human immune system is of paramount importance for the well-being of global populations. This situation and the associated risks are greatly amplified in areas where contaminants from industrial (i.e. heavy metals, persistent organic pollutants (POPs), etc.) and from hazardous waste pollutants (microplastics, endocrine disrupting chemicals, etc.) are released in the environment leading to a complex mixture of substances determined to be harmful to human health. An estimation of the population living near landfills in the EU is about 30 million people, about 6% of the total European population, and it is estimated that there are 2.5 million contaminated sites in Europe, with potentially significant adverse health effects [205]. In this regard, the European Parliament has issued the REACH Regulation [Regulation on the Registration, Evaluation, Authorisation and Restriction of Chemical Substances (EC) No. 1907/2006], to define the risks that may be posed by chemicals. Indeed, despite some impressive associations demonstrated by epidemiological studies on highly exposed populations, the role of pollutants on pathophysiological mechanisms responsible for the onset of diseases is often unknown. The REACH legislation provides toxicological studies in validated cell and animal model systems to assess the dose-response to chemicals, the effects on molecular mechanisms, tissue damages, accumulation and biomagnification under controlled dose and time exposures. The main outcome of these studies is to evaluate, for each pollutant, the No Observed Adverse Effect Level (NOAEL) and the Lowest Observed Adverse Effect Level (LOAEL), normally used to establish exposure threshold values in risk assessment guidelines. It is also noteworthy that the most recent studies in toxicology are focused on exposure to low-dose pollutant mixtures to better simulate the environmental conditions in real life. The early results suggest that low dose mixtures could induce dysmetabolism and pollutants could have a synergistic effect on systemic inflammatory response, highlighting the need for policy makers to reassess the threshold values in risk assessment and improve the strategies to protect human health [206,207,208] intervention strategy to alleviate the burden of immune system pathologies involves targeted environmental management and public health initiatives. In 2018, Bourguignon and coworkers proposed an approach aimed at reducing the exposure to complex mixtures during pregnancy by implementing global initiatives to mitigate the possible consequences of combinations of adverse lifestyle factors. The theory, akin to the “Hygiene Theory” developed to decrease the burden of disease from communicable diseases, entails reducing environmental exposures irrespective of identifying a causal link between exposure and illness. Indeed, considering that the effects of interactions among different pollutants on fetal and neonatal health are often unpredictable, and the long-term consequences are challenging to replicate experimentally, a strategy based on a priori preventive approach could reduce the disease burden earlier compared to identifying causal links between exposure and illness [209]. In this view, Dietert and colleagues proposed another intervention strategy aimed at reducing the burden of immune system pathologies. According to the authors, a wide range of adult-onset chronic conditions could be prevented by an early diagnosis and intervention strategy on “entryway diseases” during first years of life [137]. In this view, the identification of biomarkers of effect, emerges as a potent tool for early disease detection for the understanding of the mechanisms underlying pollutant-induced pathologies. Indeed, advancements in scientific knowledge on the evidence-based link between environmental pollution and altered immune response suggest the importance to i) improve understanding of exposure risks; ii) develop policy tools, guidelines, and recommendations for policymakers and authorities; iii) enhance awareness and training for health professionals and citizens. Accurate and continuous information on the issue will promote general knowledge and an improvement of habits in the environmental health domain [210]. By implementing targeted interventions, we can pave the way towards a healthier and more resilient population, with a reduced burden of immune system pathologies.
Author Contributions
Conceptualisation, V.L. and F.C.; writing—original draft preparation, G.D., A.L., S.R. and N.A., writing—review and editing, F.M.C., M.L.C., P.C., V.L. and N.A.; supervision, P.C.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the “Progetto Obiettivo Piano Sanitario Nazionale 2019 – Linea 4 – Piano Nazionale Prevenzione della Regione Siciliana (Italy)”.
Acknowledgments
The authors express their gratitude to Mrs. Antonina Azzolina and Mrs. Doriana Lo Piparo for their assistance with writing.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviation
| (o)Hg | Organic Mercury |
| A549 | Human lung adenocarcinoma cells |
| AhR | Aryl hydrocarbon receptor |
| AIM2 | Absent in Melanoma 2 receptor |
| AOP | Adverse Outcome Pathway |
| Arg1 | Arginase 1 |
| ARNT | Aryl hydrocarbon receptor nuclear translocator |
| As | Arsenic |
| BD2 | Beta-defensin 2 |
| BDE-209 | Decabromodiphenyl ether |
| BDE-47 | Tetrabromodiphenyl ether |
| BFRs | Brominated flame retardants |
| BPA | Bisphenol A |
| BPF | Bisphenol F |
| BPS | Bisphenol S |
| Cd | Cadmium |
| CD11c | Cluster of Differentiation 11c |
| CD14 | Cluster of Differentiation 14 |
| CD19 | Cluster of Differentiation 19 |
| CD206 | Cluster of Differentiation 206 |
| CD4+ | Cluster of Differentiation 4 positive (T-helper cells) |
| CD70 | Cluster of Differentiation 70 |
| CD8+ | Cluster of Differentiation 8 positive (Cytotoxic T cells) |
| CpG | Cytosine-phosphate-guanine (DNA sequence) |
| deca-BDE | Decabromodiphenyl ether |
| DIT | Developmental Immunotoxicity |
| DNA | Deoxyribonucleic acid |
| DOHaD | Developmental Origins of Health and Disease |
| EEA | European Environment Agency |
| EFSA | European Food Safety Authority |
| EMT | Epithelial-mesenchymal transition |
| HBCDD | Hexabromocyclododecane |
| HBCDDs | Hexabromocyclododecanes |
| Hg | Mercury |
| Hg0 | Elemental Mercury |
| HgCl2 | Mercury (II) chloride |
| IFN-γ | Interferon-gamma |
| IgE | Immunoglobulin E |
| IgG | Immunoglobulin G |
| IgM | Immunoglobulin M |
| IL-10 | Interleukin-10 |
| IL-12 | Interleukin-12 |
| IL-12p40 | Interleukin-12 subunit p40 |
| IL-17 | Interleukin-17 |
| IL-1β | Interleukin-1 beta |
| IL-2 | Interleukin-2 |
| IL-4 | Interleukin-4 |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin-8 |
| iNOS | Inducible nitric oxide synthase |
| IRF5 | Interferon regulatory factor 5 |
| JAK2 | Janus kinase 2 |
| STAT3 | Signal transducer and activator of transcription 3 |
| LPS | Lipopolysaccharide |
| M1 | Classically activated macrophages |
| M2 | Alternatively activated macrophages |
| MASLD | Metabolic Dysfunction-Associated Steatotic Liver Disease |
| MD2 | Myeloid differentiation factor 2 |
| MeHg | Methylmercury |
| MyD88 | Myeloid differentiation primary response 88 |
| NDL-PCB | Non-dioxin-like polychlorinated biphenyls |
| NHANES | National Health and Nutrition Examination Survey |
| NK cells | Natural killer cells |
| NO | Nitric oxide |
| NOS2 | Nitric oxide synthase 2 |
| OCPs | Organochlorine pesticides |
| p65 | RelA (p65) transcription factor |
| Pb | Lead |
| PBBs | Polybrominated biphenyls |
| PBDEs | Polybrominated diphenyl ethers |
| PBMCs | Peripheral blood mononuclear cells |
| PCBs | Polychlorinated Biphenyls (PCBs) |
| PCFs | Perfluorinated chemicals |
| PFAS | Per- and polyfluoroalkyl substances |
| PFOA | Perfluorooctanoic acid |
| PFOS | Perfluorooctanesulfonic acid |
| PHAs | polycyclic aromatic hydrocarbons |
| POPs | Persistent organic pollutants |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| PTEN | Phosphatase and tensin homolog |
| RAW264.7 | Murine macrophage cell line |
| RNA | Ribonucleic acid |
| ROS | Reactive oxygen species |
| sEVs | Small extracellular vesicles |
| TBBPA | Tetrabromobisphenol A |
| TCDD | 2,3,7,8-Tetrachlorodibenzo-p-dioxin |
| TGF-β1 | Transforming growth factor beta 1 |
| Th17 | T-helper 17 cells |
| Th2 | T-helper 2 cells |
| THP-1 | Human monocytic cell line |
| TLRs | Toll-like receptors |
| TNF-α | Tumor necrosis factor alpha |
| Treg | Regulatory T cells |
| WHO | World Health Organization |
| Wnt3a | Wingless-related integration site 3A |
| WQS | Weighted Quantile Sum regression |
References
- Kuppusamy, S.; Venkateswarlu, K.; Megharaj, M.; Mayilswami, S.; Lee, Y.B. Risk-based remediation of polluted sites: A critical perspective. Chemosphere 2017, 186, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, S.R.; Shrivastava, P.S.; Ramasamy, J. Preventing diseases through promotion of a healthier environment: World Health Organization. Ann Trop Med Public 2016, 9, 364–U115. [Google Scholar] [CrossRef]
- Wild, C.P. Complementing the genome with an "exposome": the outstanding challenge of environmental exposure measurement in molecular epidemiology. Cancer epidemiology, biomarkers & prevention : a publication of the American Association for Cancer Research, cosponsored by the American Society of Preventive Oncology 2005, 14, 1847–50. [Google Scholar]
- Rappaport, S.M. Genetic Factors Are Not the Major Causes of Chronic Diseases. PloS one 2016, 11, e0154387. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, S.M. Redefining environmental exposure for disease etiology. NPJ Syst Biol Appl 2018, 4, 30. [Google Scholar] [CrossRef] [PubMed]
- Sen, P.; Shah, P.P.; Nativio, R.; Berger, S.L. Epigenetic Mechanisms of Longevity and Aging. Cell 2016, 166, 822–839. [Google Scholar] [CrossRef] [PubMed]
- Wild, C.P. The exposome: from concept to utility. Int J Epidemiol 2012, 41, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, V.; Cardoso, R.C.; Puttabyatappa, M. Developmental Programming, a Pathway to Disease. Endocrinology 2016, 157, 1328–40. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Q.; Loganath, A.; Chong, Y.S.; Tan, J.; Obbard, J.P. Persistent organic pollutants and adverse health effects in humans. Journal of toxicology and environmental health 2006, 69. [Google Scholar]
- Ankley, G.T.; Bennett, R.S.; Erickson, R.J.; Hoff, D.J.; Hornung, M.W.; Johnson, R.D.; Mount, D.R.; Nichols, J.W.; Russom, C.L.; Schmieder, P.K.; Serrrano, J.A.; Tietge, J.E.; Villeneuve, D.L. Adverse outcome pathways: a conceptual framework to support ecotoxicology research and risk assessment. Environ Toxicol Chem 2010, 29, 730–41. [Google Scholar] [CrossRef]
- Bajard, L.; Adamovsky, O.; Audouze, K.; Baken, K.; Barouki, R.; Beltman, J.B.; Beronius, A.; Bonefeld-Jorgensen, E.C.; Cano-Sancho, G.; de Baat, M.L.; Di Tillio, F.; Fernandez, M.F.; FitzGerald, R.E.; Gundacker, C.; Hernandez, A.F.; Hilscherova, K.; Karakitsios, S.; Kuchovska, E.; Long, M.; Luijten, M.; Majid, S.; Marx-Stoelting, P.; Mustieles, V.; Negi, C.K.; Sarigiannis, D.; Scholz, S.; Sovadinova, I.; Stierum, R.; Tanabe, S.; Tollefsen, K.E.; van den Brand, A.D.; Vogs, C.; Wielsoe, M.; Wittwehr, C.; Blaha, L. Application of AOPs to assist regulatory assessment of chemical risks - Case studies, needs and recommendations. Environmental research 2023, 217, 114650. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; Miller, A.H.; Mantovani, A.; Weyand, C.M.; Barzilai, N.; Goronzy, J.J.; Rando, T.A.; Effros, R.B.; Lucia, A.; Kleinstreuer, N.; Slavich, G.M. Chronic inflammation in the etiology of disease across the life span. Nature medicine 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Rao, X.; Sigdel, K.R. Regulation of Inflammation in Autoimmune Disease. J Immunol Res 2019, 2019, 7403796. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Kummer, M.P.; Latz, E. Innate immune activation in neurodegenerative disease. Nat Rev Immunol 2014, 14, 463–77. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, L.; Fabbri, E. Inflammageing: chronic inflammation in ageing, cardiovascular disease, and frailty. Nat Rev Cardiol 2018, 15, 505–522. [Google Scholar] [CrossRef]
- Taniguchi, K.; Karin, M. NF-kappaB, inflammation, immunity and cancer: coming of age. Nat Rev Immunol 2018, 18, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Henao-Mejia, J.; Flavell, R.A. Innate immune receptors: key regulators of metabolic disease progression. Cell Metab 2013, 17, 873–882. [Google Scholar] [CrossRef]
- Kazankov, K.; Jorgensen, S.M. D.; Thomsen, K.L.; Moller, H.J.; Vilstrup, H.; George, J.; Schuppan, D.; Gronbaek, H. The role of macrophages in nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Nat Rev Gastroenterol Hepatol 2019, 16, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Mangino, M.; Roederer, M.; Beddall, M.H.; Nestle, F.O.; Spector, T.D. Innate and adaptive immune traits are differentially affected by genetic and environmental factors. Nat Commun 2017, 8, 13850. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Rui, K.; Wang, X.; Peng, N.; Zhou, W.; Shi, X.; Lu, L.; Hu, D.; Tian, J. The aryl hydrocarbon receptor in immune regulation and autoimmune pathogenesis. J Autoimmun 2023, 138, 103049. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Vazquez, C.; Quintana, F.J. Regulation of the Immune Response by the Aryl Hydrocarbon Receptor. Immunity 2018, 48, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Grosskopf, H.; Walter, K.; Karkossa, I.; von Bergen, M.; Schubert, K. Non-Genomic AhR-Signaling Modulates the Immune Response in Endotoxin-Activated Macrophages After Activation by the Environmental Stressor BaP. Frontiers in immunology 2021, 12, 620270. [Google Scholar] [CrossRef] [PubMed]
- Opitz, C.A.; Holfelder, P.; Prentzell, M.T.; Trump, S. The complex biology of aryl hydrocarbon receptor activation in cancer and beyond. Biochem Pharmacol 2023, 216, 115798. [Google Scholar] [CrossRef] [PubMed]
- Kerkvliet, N.I.; Baecher-Steppan, L.; Smith, B.B.; Youngberg, J.A.; Henderson, M.C.; Buhler, D.R. Role of the Ah locus in suppression of cytotoxic T lymphocyte activity by halogenated aromatic hydrocarbons (PCBs and TCDD): structure-activity relationships and effects in C57Bl/6 mice congenic at the Ah locus. Fundam Appl Toxicol 1990, 14, 532–41. [Google Scholar] [CrossRef] [PubMed]
- Marshall, N.B.; Vorachek, W.R.; Steppan, L.B.; Mourich, D.V.; Kerkvliet, N.I. Functional characterization and gene expression analysis of CD4+ CD25+ regulatory T cells generated in mice treated with 2,3,7,8-tetrachlorodibenzo-p-dioxin. Journal of immunology 2008, 181, 2382–91. [Google Scholar] [CrossRef] [PubMed]
- O'Driscoll, C.A.; Gallo, M.E.; Fechner, J.H.; Schauer, J.J.; Mezrich, J.D. Real-world PM extracts differentially enhance Th17 differentiation and activate the aryl hydrocarbon receptor (AHR). Toxicology 2019, 414, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Jung, A.M.; Beitel, S.C.; Gutenkunst, S.L.; Billheimer, D.; Jahnke, S.A.; Littau, S.R.; White, M.; Hoppe-Jones, C.; Cherrington, N.; Burgess, J.L. Excretion of polybrominated diphenyl ethers and AhR activation in breastmilk among firefighters. Toxicol Sci 2023, 192, 223–32. [Google Scholar] [CrossRef] [PubMed]
- Checa, J.; Aran, J.M. Reactive Oxygen Species: Drivers of Physiological and Pathological Processes. J Inflamm Res 2020, 13, 1057–1073. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, F.S.; Lanzetti, M.; Nesi, R.T.; Nagato, A.C.; Silva, C.P. E.; Kennedy-Feitosa, E.; Melo, A.C.; Cattani-Cavalieri, I.; Porto, L.C.; Valenca, S.S. Oxidative Stress and Inflammation in Acute and Chronic Lung Injuries. Antioxidants (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Ahmed, A.S. S.; Sultana, S.; Habib, A.; Ullah, H.; Musa, N.; Hossain, M.B.; Rahman, M.M.; Sarker, M.S. I. Bioaccumulation of heavy metals in some commercially important fishes from a tropical river estuary suggests higher potential health risk in children than adults. PloS one 2019, 14, e0219336. [Google Scholar] [CrossRef]
- Onakpa, M.M.; Njan, A.A.; Kalu, O.C. A Review of Heavy Metal Contamination of Food Crops in Nigeria. Ann Glob Health 2018, 84, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Orosun, M.M.; Nwabachili, S.; Alshehri, R.F.; Omeje, M.; Alshdoukhi, I.F.; Okoro, H.K.; Ogunkunle, C.O.; Louis, H.; Abdulhamid, F.A.; Osahon, S.E.; Mohammed, A.U.; Ehinlafa, E.O.; Yunus, S.O.; Ife-Adediran, O. Potentially toxic metals in irrigation water, soil, and vegetables and their health risks using Monte Carlo models. Scientific reports 2023, 13, 21220. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Mahabbat-e Khoda, S.; Rekha, R.S.; Gardner, R.M.; Ameer, S.S.; Moore, S.; Ekstrom, E.C.; Vahter, M.; Raqib, R. Arsenic-associated oxidative stress, inflammation, and immune disruption in human placenta and cord blood. Environmental health perspectives 2011, 119, 258–64. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, N.; Banerjee, S.; Sen, R.; Bandyopadhyay, A.; Sarma, N.; Majumder, P.; Das, J.K.; Chatterjee, M.; Kabir, S.N.; Giri, A.K. Chronic arsenic exposure impairs macrophage functions in the exposed individuals. J Clin Immunol 2009, 29, 582–94. [Google Scholar] [CrossRef]
- Cho, Y.; Ahn, K.H.; Back, M.J.; Choi, J.M.; Ji, J.E.; Won, J.H.; Fu, Z.; Jang, J.M.; Kim, D.K. Age-related effects of sodium arsenite on splenocyte proliferation and Th1/Th2 cytokine production. Arch Pharm Res 2012, 35, 375–82. [Google Scholar] [CrossRef] [PubMed]
- Conde, P.; Acosta-Saavedra, L.C.; Goytia-Acevedo, R.C.; Calderon-Aranda, E.S. Sodium arsenite-induced inhibition of cell proliferation is related to inhibition of IL-2 mRNA expression in mouse activated T cells. Arch Toxicol 2007, 81, 251–9. [Google Scholar] [CrossRef] [PubMed]
- Morzadec, C.; Bouezzedine, F.; Macoch, M.; Fardel, O.; Vernhet, L. Inorganic arsenic impairs proliferation and cytokine expression in human primary T lymphocytes. Toxicology 2012, 300, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Binet, F.; Antoine, F.; Girard, D. Interaction between arsenic trioxide and human primary cells: emphasis on human cells of myeloid origin. Inflamm Allergy Drug Targets 2009, 8, 21–7. [Google Scholar] [CrossRef] [PubMed]
- Binet, F.; Cavalli, H.; Moisan, E.; Girard, D. Arsenic trioxide (AT) is a novel human neutrophil pro-apoptotic agent: effects of catalase on AT-induced apoptosis, degradation of cytoskeletal proteins and de novo protein synthesis. Br J Haematol 2006, 132, 349–58. [Google Scholar] [CrossRef]
- Xiao, T.; Zou, Z.; Xue, J.; Syed, B.M.; Sun, J.; Dai, X.; Shi, M.; Li, J.; Wei, S.; Tang, H.; Zhang, A.; Liu, Q. LncRNA H19-mediated M2 polarization of macrophages promotes myofibroblast differentiation in pulmonary fibrosis induced by arsenic exposure. Environmental pollution 2021, 268, 115810. [Google Scholar] [CrossRef]
- Xue, J.; Xiao, T.; Wei, S.; Sun, J.; Zou, Z.; Shi, M.; Sun, Q.; Dai, X.; Wu, L.; Li, J.; Xia, H.; Tang, H.; Zhang, A.; Liu, Q. miR-21-regulated M2 polarization of macrophage is involved in arsenicosis-induced hepatic fibrosis through the activation of hepatic stellate cells. J Cell Physiol 2021, 236, 6025–6041. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.M.; Chiou, H.Y.; Ho, I.C.; Chen, C.J.; Lee, T.C. Gene expression of inflammatory molecules in circulating lymphocytes from arsenic-exposed human subjects. Environmental health perspectives 2003, 111, 1429–38. [Google Scholar] [CrossRef] [PubMed]
- Pilsner, J.R.; Liu, X.; Ahsan, H.; Ilievski, V.; Slavkovich, V.; Levy, D.; Factor-Litvak, P.; Graziano, J.H.; Gamble, M.V. Folate deficiency, hyperhomocysteinemia, low urinary creatinine, and hypomethylation of leukocyte DNA are risk factors for arsenic-induced skin lesions. Environmental health perspectives 2009, 117, 254–60. [Google Scholar] [CrossRef] [PubMed]
- Dangleben, N.L.; Skibola, C.F.; Smith, M.T. Arsenic immunotoxicity: a review. Environmental health : a global access science source 2013, 12, 73. [Google Scholar] [CrossRef] [PubMed]
- Dashner-Titus, E.J.; Schilz, J.R.; Simmons, K.A.; Duncan, T.R.; Alvarez, S.C.; Hudson, L.G. Differential response of human T-lymphocytes to arsenic and uranium. Toxicol Lett 2020, 333, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.L.; Khan, M.I.; Wang, J.; Ali, R.; Ali, S.W.; Zahra, Q.U.; Kazmi, A.; Lolai, A.; Huang, Y.L.; Hussain, A.; Bilal, M.; Li, F.; Qiu, B. Role of receptor tyrosine kinases mediated signal transduction pathways in tumor growth and angiogenesis-New insight and futuristic vision. Int J Biol Macromol 2021, 180, 739–752. [Google Scholar] [CrossRef] [PubMed]
- Dietert, R.R.; Lee, J.E.; Hussain, I.; Piepenbrink, M. Developmental immunotoxicology of lead. Toxicol Appl Pharmacol 2004, 198, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, C.L.; Wu, K.H.; Wan, K.S. Effects of environmental lead exposure on T-helper cell-specific cytokines in children. J Immunotoxicol 2011, 8, 284–7. [Google Scholar] [CrossRef] [PubMed]
- Heo, Y.; Parsons, P.J.; Lawrence, D.A. Lead differentially modifies cytokine production in vitro and in vivo. Toxicol Appl Pharmacol 1996, 138, 149–57. [Google Scholar] [CrossRef] [PubMed]
- Krocova, Z.; Macela, A.; Kroca, M.; Hernychova, L. The immunomodulatory effect(s) of lead and cadmium on the cells of immune system in vitro. Toxicol In Vitro 2000, 14, 33–40. [Google Scholar] [CrossRef]
- Mishra, K.P.; Singh, V.K.; Rani, R.; Yadav, V.S.; Chandran, V.; Srivastava, S.P.; Seth, P.K. Effect of lead exposure on the immune response of some occupationally exposed individuals. Toxicology 2003, 188, 251–9. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wu, J.; Xu, H.; Li, Q.; Zhang, Y.; Zhai, Y.; Tang, M.; Liu, Y.; Liu, T.; Ye, Y.; He, M.; He, R.; Xu, Y.; Zhou, Z.; Kan, H.; Zhang, Y. Lead exposure suppresses the Wnt3a/beta-catenin signaling to increase the quiescence of hematopoietic stem cells via reducing the expression of CD70 on bone marrow-resident macrophages. Toxicol Sci 2023, 195, 123–142. [Google Scholar] [CrossRef] [PubMed]
- Dobrakowski, M.; Boron, M.; Czuba, Z.P.; Kasperczyk, A.; Machon-Grecka, A.; Kasperczyk, S. Cytokines related to three major types of cell-mediated immunity in short- and long-term exposures to lead compounds. J Immunotoxicol 2016, 13, 770–774. [Google Scholar] [CrossRef] [PubMed]
- Metryka, E.; Kupnicka, P.; Kapczuk, P.; Aszakiewicz, B.; Piotrowska, K.; Tkacz, M.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Lead (Pb) Accumulation in Human THP-1 Monocytes/Macrophages In Vitro and the Influence on Cell Apoptosis. Biol Trace Elem Res 2021, 199, 955–967. [Google Scholar] [CrossRef] [PubMed]
- Osman, K.; Akesson, A.; Berglund, M.; Bremme, K.; Schutz, A.; Ask, K.; Vahter, M. Toxic and essential elements in placentas of Swedish women. Clin Biochem 2000, 33, 131–8. [Google Scholar] [CrossRef] [PubMed]
- Llanos, M.N.; Ronco, A.M. Fetal growth restriction is related to placental levels of cadmium, lead and arsenic but not with antioxidant activities. Reproductive toxicology 2009, 27, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Zahir, F.; Rizwi, S.J.; Haq, S.K.; Khan, R.H. Low dose mercury toxicity and human health. Environ Toxicol Pharmacol 2005, 20, 351–60. [Google Scholar] [CrossRef] [PubMed]
- Jansson, G.; Harms-Ringdahl, M. Stimulating effects of mercuric- and silver ions on the superoxide anion production in human polymorphonuclear leukocytes. Free Radic Res Commun 1993, 18, 87–98. [Google Scholar] [CrossRef]
- Christensen, M.M.; Ellermann-Eriksen, S.; Rungby, J.; Mogensen, S.C. Comparison of the interaction of methyl mercury and mercuric chloride with murine macrophages. Arch Toxicol 1993, 67, 205–11. [Google Scholar] [CrossRef]
- InSug, O.; Datar, S.; Koch, C.J.; Shapiro, I.M.; Shenker, B.J. Mercuric compounds inhibit human monocyte function by inducing apoptosis: evidence for formation of reactive oxygen species, development of mitochondrial membrane permeability transition and loss of reductive reserve. Toxicology 1997, 124, 211–24. [Google Scholar] [CrossRef]
- Rodriguez-Viso, P.; Domene, A.; Velez, D.; Devesa, V.; Monedero, V.; Zuniga, M. Mercury toxic effects on the intestinal mucosa assayed on a bicameral in vitro model: Possible role of inflammatory response and oxidative stress. Food and chemical toxicology : an international journal published for the British Industrial Biological Research Association 2022, 166, 113224. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Alarcon, J.; Milic, M.; Bustamante-Montes, L.P.; Isaac-Olive, K.; Valencia-Quintana, R.; Ramirez-Duran, N. Genotoxicity of Mercury and Its Derivatives Demonstrated In Vitro and In Vivo in Human Populations Studies. Systematic Review. Toxics 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Ilback, N.G. Effects of methyl mercury exposure on spleen and blood natural killer (NK) cell activity in the mouse. Toxicology 1991, 67, 117–24. [Google Scholar] [CrossRef] [PubMed]
- Ilback, N.G.; Sundberg, J.; Oskarsson, A. Methyl mercury exposure via placenta and milk impairs natural killer (NK) cell function in newborn rats. Toxicol Lett 1991, 58, 149–58. [Google Scholar] [CrossRef] [PubMed]
- Migdal, C.; Foggia, L.; Tailhardat, M.; Courtellemont, P.; Haftek, M.; Serres, M. Sensitization effect of thimerosal is mediated in vitro via reactive oxygen species and calcium signaling. Toxicology 2010, 274, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Migdal, C.; Tailhardat, M.; Courtellemont, P.; Haftek, M.; Serres, M. Responsiveness of human monocyte-derived dendritic cells to thimerosal and mercury derivatives. Toxicol Appl Pharmacol 2010, 246, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Shukla, G.S.; Chandra, S.V. Effects of cadmium on superoxide dismutase and lipid peroxidation in liver and kidney of growing rats: in vivo and in vitro studies. Pharmacol Toxicol 1987, 60, 355–8. [Google Scholar] [CrossRef] [PubMed]
- Mates, J.M.; Segura, J.A.; Alonso, F.J.; Marquez, J. Roles of dioxins and heavy metals in cancer and neurological diseases using ROS-mediated mechanisms. Free Radic Biol Med 2010, 49, 1328–41. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mendoza, D.; Han, B.; van den Berg, H.; van den Brink, N.W. Cell-specific immune-modulation of cadmium on murine macrophages and mast cell lines in vitro. J Appl Toxicol 2019, 39, 992–1001. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Mo, L.; Huang, H.; Mo, L.; Zhu, W.; Li, W.; Yang, G.; Chen, L.; Wu, Y.; Song, J.; Yang, X. Cadmium contributes to atherosclerosis by affecting macrophage polarization. Food and chemical toxicology : an international journal published for the British Industrial Biological Research Association 2023, 173, 113603. [Google Scholar] [CrossRef]
- Hanson, M.L.; Holaskova, I.; Elliott, M.; Brundage, K.M.; Schafer, R.; Barnett, J.B. Prenatal cadmium exposure alters postnatal immune cell development and function. Toxicol Appl Pharmacol 2012, 261, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Holaskova, I.; Elliott, M.; Hanson, M.L.; Schafer, R.; Barnett, J.B. Prenatal cadmium exposure produces persistent changes to thymus and spleen cell phenotypic repertoire as well as the acquired immune response. Toxicol Appl Pharmacol 2012, 265, 181–9. [Google Scholar] [CrossRef] [PubMed]
- Goyal, T.; Mitra, P.; Singh, P.; Ghosh, R.; Sharma, S.; Sharma, P. Association of microRNA expression with changes in immune markers in workers with cadmium exposure. Chemosphere 2021, 274, 129615. [Google Scholar] [CrossRef] [PubMed]
- El-Shahawi, M.S.; Hamza, A.; Bashammakh, A.S.; Al-Saggaf, W.T. An overview on the accumulation, distribution, transformations, toxicity and analytical methods for the monitoring of persistent organic pollutants. Talanta 2010, 80, 1587–97. [Google Scholar] [CrossRef] [PubMed]
- Haddad, S.; Ayotte, P.; Verner, M.A. Derivation of exposure factors for infant lactational exposure to persistent organic pollutants (POPs). Regul Toxicol Pharmacol 2015, 71, 135–40. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y.; Tao, F.M.; Zeng, E.Y. Theoretical study on the chemical properties of polybrominated diphenyl ethers. Chemosphere 2008, 70, 901–7. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, B.P.; Warren, T.K.; Luong, H. Fewer T lymphocytes and decreased pulmonary influenza virus burden in mice exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). J Toxicol Environ Health A 2000, 61, 39–53. [Google Scholar] [PubMed]
- Marshall, N.B.; Kerkvliet, N.I. Dioxin and immune regulation: emerging role of aryl hydrocarbon receptor in the generation of regulatory T cells. Ann N Y Acad Sci 2010, 1183, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Warren, T.K.; Mitchell, K.A.; Lawrence, B.P. Exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) suppresses the humoral and cell-mediated immune responses to influenza A virus without affecting cytolytic activity in the lung. Toxicol Sci 2000, 56, 114–23. [Google Scholar] [CrossRef]
- Kerkvliet, N.I. Recent advances in understanding the mechanisms of TCDD immunotoxicity. Int Immunopharmacol 2002, 2, 277–91. [Google Scholar] [CrossRef]
- Torti, M.F.; Giovannoni, F.; Quintana, F.J.; Garcia, C.C. The Aryl Hydrocarbon Receptor as a Modulator of Anti-viral Immunity. Frontiers in immunology 2021, 12, 624293. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, N.; Han, Y.; Rao, K.; Ji, X.; Ma, M. 2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD)-induced suppression of immunity in THP-1-derived macrophages and the possible mechanisms. Environmental pollution 2021, 287, 117302. [Google Scholar] [CrossRef] [PubMed]
- Bruner-Tran, K.L.; Gnecco, J.; Ding, T.; Glore, D.R.; Pensabene, V.; Osteen, K.G. Exposure to the environmental endocrine disruptor TCDD and human reproductive dysfunction: Translating lessons from murine models. Reproductive toxicology 2017, 68, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Petriello, M.C.; Zhu, B.; Hennig, B. PCB 126 induces monocyte/macrophage polarization and inflammation through AhR and NF-kappaB pathways. Toxicol Appl Pharmacol 2019, 367, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Santoro, A.; Ferrante, M.C.; Di Guida, F.; Pirozzi, C.; Lama, A.; Simeoli, R.; Clausi, M.T.; Monnolo, A.; Mollica, M.P.; Mattace Raso, G.; Meli, R. Polychlorinated Biphenyls (PCB 101, 153, and 180) Impair Murine Macrophage Responsiveness to Lipopolysaccharide: Involvement of NF-kappaB Pathway. Toxicol Sci 2015, 147, 255–69. [Google Scholar] [CrossRef] [PubMed]
- Waugh, C.A.; Arukwe, A.; Jaspers, V.L. B. Deregulation of microRNA-155 and its transcription factor NF-kB by polychlorinated biphenyls during viral infections. APMIS 2018, 126, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, C.L.; Spiegelhoff, A.; Wang, K.; Lavery, T.; Nunez, A.; Manuel, R.; Hillers-Ziemer, L.; Arendt, L.M.; Stietz, K.P. K. The Bladder Is a Novel Target of Developmental Polychlorinated Biphenyl Exposure Linked to Increased Inflammatory Cells in the Bladder of Young Mice. Toxics 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Curtis, S.W.; Cobb, D.O.; Kilaru, V.; Terrell, M.L.; Marder, M.E.; Barr, D.B.; Marsit, C.J.; Marcus, M.; Conneely, K.N.; Smith, A.K. Genome-wide DNA methylation differences and polychlorinated biphenyl (PCB) exposure in a US population. Epigenetics 2021, 16, 338–352. [Google Scholar] [CrossRef] [PubMed]
- Sjodin, A.; Patterson, D.G. Jr.; Bergman, A. A review on human exposure to brominated flame retardants--particularly polybrominated diphenyl ethers. Environment international 2003, 29, 829–39. [Google Scholar] [CrossRef] [PubMed]
- McGrath, T.J.; Ball, A.S.; Clarke, B.O. Critical review of soil contamination by polybrominated diphenyl ethers (PBDEs) and novel brominated flame retardants (NBFRs); concentrations, sources and congener profiles. Environmental pollution 2017, 230, 741–757. [Google Scholar] [CrossRef]
- Shaw, S.D.; Blum, A.; Weber, R.; Kannan, K.; Rich, D.; Lucas, D.; Koshland, C.P.; Dobraca, D.; Hanson, S.; Birnbaum, L.S. Halogenated flame retardants: do the fire safety benefits justify the risks? Rev Environ Health 2010, 25, 261–305. [Google Scholar] [CrossRef] [PubMed]
- Chain, E.P. o. C. i. t. F.; Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; Del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Ron Hoogenboom, L.; Leblanc, J.-C.; Nebbia, C.S.; Nielsen, E.; Ntzani, E.; Petersen, A.; Sand, S.; Schwerdtle, T.; Wallace, H.; Benford, D.; Furst, P.; Hart, A.; Rose, M.; Schroeder, H.; Vrijheid, M.; Ioannidou, S.; Nikolic, M.; Bordajandi, L.R.; Vleminckx, C. Update of the risk assessment of polybrominated diphenyl ethers (PBDEs) in food. EFSA journal European Food Safety Authority 2024, 22, e8497. [Google Scholar]
- Xue, J.; Xiao, Q.; Zhang, M.; Li, D.; Wang, X. Toxic Effects and Mechanisms of Polybrominated Diphenyl Ethers. International journal of molecular sciences 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Alzualde, A.; Behl, M.; Sipes, N.S.; Hsieh, J.H.; Alday, A.; Tice, R.R.; Paules, R.S.; Muriana, A.; Quevedo, C. Toxicity profiling of flame retardants in zebrafish embryos using a battery of assays for developmental toxicity, neurotoxicity, cardiotoxicity and hepatotoxicity toward human relevance. Neurotoxicology and teratology 2018, 70, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Lema, S.C.; Schultz, I.R.; Scholz, N.L.; Incardona, J.P.; Swanson, P. Neural defects and cardiac arrhythmia in fish larvae following embryonic exposure to 2,2',4,4'-tetrabromodiphenyl ether (PBDE 47). Aquat Toxicol 2007, 82, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Parsons, A.; Lange, A.; Hutchinson, T.H.; Miyagawa, S.; Iguchi, T.; Kudoh, T.; Tyler, C.R. Molecular mechanisms and tissue targets of brominated flame retardants, BDE-47 and TBBPA, in embryo-larval life stages of zebrafish (Danio rerio). Aquat Toxicol 2019, 209, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.; Longo, A.; Di Sano, C.; Cigna, D.; Cibella, F.; Di Felice, G.; Colombo, P. In vitro exposure to 2,2',4,4'-tetrabromodiphenyl ether (PBDE-47) impairs innate inflammatory response. Chemosphere 2019, 219, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.; Longo, A.; Adamo, G.; Fiannaca, A.; Picciotto, S.; La Paglia, L.; Romancino, D.; La Rosa, M.; Urso, A.; Cibella, F.; Bongiovanni, A.; Colombo, P. 2,2'4,4'-Tetrabromodiphenyl Ether (PBDE-47) Modulates the Intracellular miRNA Profile, sEV Biogenesis and Their miRNA Cargo Exacerbating the LPS-Induced Pro-Inflammatory Response in THP-1 Macrophages. Frontiers in immunology 2021, 12, 664534. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.; Aloi, N.; Lo Presti, E.; Fiannaca, A.; Longo, A.; Adamo, G.; Urso, A.; Meraviglia, S.; Bongiovanni, A.; Cibella, F.; Colombo, P. Impact of the flame retardant 2,2'4,4'-tetrabromodiphenyl ether (PBDE-47) in THP-1 macrophage-like cell function via small extracellular vesicles. Frontiers in immunology 2022, 13, 1069207. [Google Scholar] [CrossRef]
- Albano, G.D.; Longo, V.; Montalbano, A.M.; Aloi, N.; Barone, R.; Cibella, F.; Profita, M.; Colombo, P. Extracellular vesicles from PBDE-47 treated M(LPS) THP-1 macrophages modulate the expression of markers of epithelial integrity, EMT, inflammation and muco-secretion in ALI culture of airway epithelium. Life Sci 2023, 322, 121616. [Google Scholar] [CrossRef]
- Ren, Q.; Xie, X.; Zhao, C.; Wen, Q.; Pan, R.; Du, Y. 2,2',4,4'-Tetrabromodiphenyl Ether (PBDE 47) Selectively Stimulates Proatherogenic PPARgamma Signatures in Human THP-1 Macrophages to Contribute to Foam Cell Formation. Chem Res Toxicol 2022, 35, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Babior, B.M. Phagocytes and oxidative stress. Am J Med 2000, 109, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Yang, F.; Hui, Y.; Xu, Y.; Lu, Y.; Liu, J. Cytotoxicity and apoptosis induction on RTG-2 cells of 2,2',4,4'-tetrabromodiphenyl ether (BDE-47) and decabrominated diphenyl ether (BDE-209). Toxicol In Vitro 2010, 24, 1190–6. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.Y.; Wan, B.; Guo, L.H.; Yang, Y.; Ren, X.M.; Zhang, H. In vivo immunotoxicity of perfluorooctane sulfonate in BALB/c mice: Identification of T-cell receptor and calcium-mediated signaling pathway disruption through gene expression profiling of the spleen. Chem Biol Interact 2015, 240, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Barletta, B.; Corinti, S.; Maranghi, F.; Tait, S.; Tassinari, R.; Martinelli, A.; Longo, A.; Longo, V.; Colombo, P.; Di Felice, G.; Butteroni, C. The environmental pollutant BDE-47 modulates immune responses in invitro and in vivo murine models. Chemosphere 2024, 349, 140739. [Google Scholar] [CrossRef] [PubMed]
- Birnbaum, L.S.; Staskal, D.F. Brominated flame retardants: cause for concern? Environmental health perspectives 2004, 112, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Schecter, A.; Szabo, D.T.; Miller, J.; Gent, T.L.; Malik-Bass, N.; Petersen, M.; Paepke, O.; Colacino, J.A.; Hynan, L.S.; Harris, T.R.; Malla, S.; Birnbaum, L.S. Hexabromocyclododecane (HBCD) stereoisomers in U.S. food from Dallas, Texas. Environmental health perspectives 2012, 120, 1260–4. [Google Scholar] [CrossRef] [PubMed]
- Kajiwara, N.; Sueoka, M.; Ohiwa, T.; Takigami, H. Determination of flame-retardant hexabromocyclododecane diastereomers in textiles. Chemosphere 2009, 74, 1485–9. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.A.; Harrad, S.; Covaci, A. Hexabromocyclododecanes and tetrabromobisphenol-A in indoor air and dust in Birmingham, U.K: implications for human exposure. Environmental science & technology 2008, 42, 6855–61. [Google Scholar]
- Remberger, M.; Sternbeck, J.; Palm, A.; Kaj, L.; Stromberg, K.; Brorstrom-Lunden, E. The environmental occurrence of hexabromocyclododecane in Sweden. Chemosphere 2004, 54, 9–21. [Google Scholar] [CrossRef]
- Kakimoto, K.; Akutsu, K.; Konishi, Y.; Tanaka, Y. Time trend of hexabromocyclododecane in the breast milk of Japanese women. Chemosphere 2008, 71, 1110–4. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, H.K.; Kvalem, H.E.; Thomsen, C.; Froshaug, M.; Haugen, M.; Becher, G.; Alexander, J.; Meltzer, H.M. Dietary exposure to brominated flame retardants correlates with male blood levels in a selected group of Norwegians with a wide range of seafood consumption. Mol Nutr Food Res 2008, 52, 217–27. [Google Scholar] [CrossRef] [PubMed]
- Pulkrabova, J.; Hradkova, P.; Hajslova, J.; Poustka, J.; Napravnikova, M.; Polacek, V. Brominated flame retardants and other organochlorine pollutants in human adipose tissue samples from the Czech Republic. Environment international 2009, 35, 63–8. [Google Scholar] [CrossRef] [PubMed]
- Chain, E.P. C. i. t. F.; Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; Del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.R.; Leblanc, J.-C.; Nebbia, C.S.; Nielsen, E.; Ntzani, E.; Petersen, A.; Sand, S.; Schwerdtle, T.; Wallace, H.; Benford, D.; Furst, P.; Rose, M.; Ioannidou, S.; Nikolic, M.; Bordajandi, L.R.; Vleminckx, C. Update of the risk assessment of hexabromocyclododecanes (HBCDDs) in food. EFSA journal European Food Safety Authority 2021, 19, e06421. [Google Scholar]
- Baranska, A.; Bukowska, B.; Michalowicz, J. Determination of Apoptotic Mechanism of Action of Tetrabromobisphenol A and Tetrabromobisphenol S in Human Peripheral Blood Mononuclear Cells: A Comparative Study. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Baranska, A.; Wozniak, A.; Mokra, K.; Michalowicz, J. Genotoxic Mechanism of Action of TBBPA, TBBPS and Selected Bromophenols in Human Peripheral Blood Mononuclear Cells. Frontiers in immunology 2022, 13, 869741. [Google Scholar] [CrossRef] [PubMed]
- Canbaz, D.; Lebre, M.C.; Logiantara, A.; van Ree, R.; van Rijt, L.S. Indoor pollutant hexabromocyclododecane enhances house dust mite-induced activation of human monocyte-derived dendritic cells. J Immunotoxicol 2016, 13, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Yu, Z.; Chen, H.; Han, Y.; Xiang, M.; Chen, X.; Ma, R.; Wang, Z. Tetrabromobisphenol A: Disposition, kinetics and toxicity in animals and humans. Environmental pollution 2019, 253, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Feiteiro, J.; Mariana, M.; Cairrao, E. Health toxicity effects of brominated flame retardants: From environmental to human exposure. Environmental pollution 2021, 285, 117475. [Google Scholar] [CrossRef]
- Miao, B.; Yakubu, S.; Zhu, Q.; Issaka, E.; Zhang, Y.; Adams, M. A Review on Tetrabromobisphenol A: Human Biomonitoring, Toxicity, Detection and Treatment in the Environment. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Balistrieri, A.; Hobohm, L.; Srivastava, T.; Meier, A.; Corriden, R. Alterations in human neutrophil function caused by bisphenol A. Am J Physiol Cell Physiol 2018, 315, C636–C642. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Li, M.; Wu, C.; Zhou, C.; Zhang, J.; Zhu, Q.; Shen, T. Bisphenol A promotes macrophage proinflammatory subtype polarization via upregulation of IRF5 expression in vitro. Toxicol In Vitro 2019, 60, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Kodila, A.; Franko, N.; Sollner Dolenc, M. A review on immunomodulatory effects of BPA analogues. Arch Toxicol 2023, 97, 1831–1846. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lin, Z.; Ye, L.; Chen, X.; Zhang, W.; Zhang, Z.; Luo, F.; Liu, Y.; Shi, M. Estrogen receptor-regulated SOCS3 modulation via JAK2/STAT3 pathway is involved in BPF-induced M1 polarization of macrophages. Toxicology 2020, 433-434, 152404. [Google Scholar] [CrossRef]
- Hernandez Avila, R.; Palacios-Arreola, M.I.; Nava-Castro, K.E.; Morales-Montor, J.; Ostoa-Saloma, P. Neonatal Bisphenol A Exposure Affects the IgM Humoral Immune Response to 4T1 Breast Carcinoma Cells in Mice. International journal of environmental research and public health 2019, 16. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Q.; Liu, T.; Yang, S.; Sun, L.; Zhao, Z.Y.; Li, L.Y.; She, Y.C.; Zheng, Y.Y.; Ye, X.Y.; Bao, Q.; Dong, G.H.; Li, C.W.; Cui, J. Perfluoroalkyl substance pollutants activate the innate immune system through the AIM2 inflammasome. Nat Commun 2021, 12, 2915. [Google Scholar] [CrossRef] [PubMed]
- Jian, J.M.; Chen, D.; Han, F.J.; Guo, Y.; Zeng, L.; Lu, X.; Wang, F. A short review on human exposure to and tissue distribution of per- and polyfluoroalkyl substances (PFASs). The Science of the total environment 2018, 636, 1058–1069. [Google Scholar] [CrossRef] [PubMed]
- Perez, F.; Nadal, M.; Navarro-Ortega, A.; Fabrega, F.; Domingo, J.L.; Barcelo, D.; Farre, M. Accumulation of perfluoroalkyl substances in human tissues. Environment international 2013, 59, 354–62. [Google Scholar] [CrossRef] [PubMed]
- Midgett, K.; Peden-Adams, M.M.; Gilkeson, G.S.; Kamen, D.L. In vitro evaluation of the effects of perfluorooctanesulfonic acid (PFOS) and perfluorooctanoic acid (PFOA) on IL-2 production in human T-cells. J Appl Toxicol 2015, 35, 459–65. [Google Scholar] [CrossRef]
- Torres, L.; Redko, A.; Limper, C.; Imbiakha, B.; Chang, S.; August, A. Effect of Perfluorooctanesulfonic acid (PFOS) on immune cell development and function in mice. Immunol Lett 2021, 233, 31–41. [Google Scholar] [CrossRef]
- Taylor, K.D.; Woodlief, T.L.; Ahmed, A.; Hu, Q.; Duncker, P.C.; DeWitt, J.C. Quantifying the impact of PFOA exposure on B-cell development and antibody production. Toxicol Sci 2023, 194, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Dietert, R.R. Developmental Immunotoxicity, Perinatal Programming, and Noncommunicable Diseases: Focus on Human Studies. Adv Med 2014, 2014, 867805. [Google Scholar] [CrossRef] [PubMed]
- Holsapple, M.P.; West, L.J.; Landreth, K.S. Species comparison of anatomical and functional immune system development. Birth Defects Res B Dev Reprod Toxicol 2003, 68, 321–34. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J. The fetal and infant origins of adult disease. BMJ 1990, 301, 1111. [Google Scholar] [CrossRef] [PubMed]
- Fleming, T.P.; Watkins, A.J.; Velazquez, M.A.; Mathers, J.C.; Prentice, A.M.; Stephenson, J.; Barker, M.; Saffery, R.; Yajnik, C.S.; Eckert, J.J.; Hanson, M.A.; Forrester, T.; Gluckman, P.D.; Godfrey, K.M. Origins of lifetime health around the time of conception: causes and consequences. Lancet 2018, 391, 1842–1852. [Google Scholar] [CrossRef] [PubMed]
- Hardy, A.; Benford, D.; Halldorsson, T.; Jeger, M.J.; Knutsen, H.K.; More, S.; Naegeli, H.; Noteborn, H.; Ockleford, C.; Ricci, A.; Rychen, G.; Schlatter, J.R.; Silano, V.; Solecki, R.; Turck, D.; Bresson, J.L.; Dusemund, B.; Gundert-Remy, U.; Kersting, M.; Lambré, C.; Penninks, A.; Tritscher, A.; Waalkens-Berendsen, I.; Woutersen, R.; Arcella, D.; Marques, D.C.; Dorne, J.L.; Kass, G.E. N.; Mortensen, A.; Comm, E.S. Guidance on the risk assessment of substances present in food intended for infants below 16 weeks of age. Efsa Journal 2017, 15. [Google Scholar]
- Dietert, R.R.; DeWitt, J.C.; Germolec, D.R.; Zelikoff, J.T. Breaking patterns of environmentally influenced disease for health risk reduction: immune perspectives. Environmental health perspectives 2010, 118, 1091–9. [Google Scholar] [CrossRef] [PubMed]
- Popescu, M.; Feldman, T.B.; Chitnis, T. Interplay Between Endocrine Disruptors and Immunity: Implications for Diseases of Autoreactive Etiology. Front Pharmacol 2021, 12, 626107. [Google Scholar] [CrossRef] [PubMed]
- Griffith, O.W.; Chavan, A.R.; Protopapas, S.; Maziarz, J.; Romero, R.; Wagner, G.P. Embryo implantation evolved from an ancestral inflammatory attachment reaction. Proc Natl Acad Sci U S A 2017, 114, E6566–E6575. [Google Scholar] [CrossRef] [PubMed]
- Jarmund, A.H.; Giskeodegard, G.F.; Ryssdal, M.; Steinkjer, B.; Stokkeland, L.M. T.; Madssen, T.S.; Stafne, S.N.; Stridsklev, S.; Moholdt, T.; Heimstad, R.; Vanky, E.; Iversen, A.C. Cytokine Patterns in Maternal Serum From First Trimester to Term and Beyond. Frontiers in immunology 2021, 12, 752660. [Google Scholar] [CrossRef]
- Kaislasuo, J.; Simpson, S.; Petersen, J.F.; Peng, G.; Aldo, P.; Lokkegaard, E.; Paidas, M.; Pal, L.; Guller, S.; Mor, G. IL-10 to TNFalpha ratios throughout early first trimester can discriminate healthy pregnancies from pregnancy losses. Am J Reprod Immunol 2020, 83, e13195. [Google Scholar] [CrossRef] [PubMed]
- Harmon, A.C.; Cornelius, D.C.; Amaral, L.M.; Faulkner, J.L.; Cunningham, M.W. Jr.; Wallace, K.; LaMarca, B. The role of inflammation in the pathology of preeclampsia. Clin Sci (Lond) 2016, 130, 409–19. [Google Scholar] [CrossRef] [PubMed]
- Prairie, E.; Cote, F.; Tsakpinoglou, M.; Mina, M.; Quiniou, C.; Leimert, K.; Olson, D.; Chemtob, S. The determinant role of IL-6 in the establishment of inflammation leading to spontaneous preterm birth. Cytokine Growth Factor Rev 2021, 59, 118–130. [Google Scholar] [CrossRef]
- Atladottir, H.O.; Thorsen, P.; Ostergaard, L.; Schendel, D.E.; Lemcke, S.; Abdallah, M.; Parner, E.T. Maternal infection requiring hospitalization during pregnancy and autism spectrum disorders. J Autism Dev Disord 2010, 40, 1423–30. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.J.; Hallenbeck, J.M. Effects of intrauterine inflammation on developing rat brain. J Neurosci Res 2002, 70, 570–9. [Google Scholar] [CrossRef]
- Ponzio, N.M.; Servatius, R.; Beck, K.; Marzouk, A.; Kreider, T. Cytokine levels during pregnancy influence immunological profiles and neurobehavioral patterns of the offspring. Ann N Y Acad Sci 2007, 1107, 118–28. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Li, J.; Garbett, K.; Mirnics, K.; Patterson, P.H. Maternal immune activation alters fetal brain development through interleukin-6. J Neurosci 2007, 27, 10695–702. [Google Scholar] [CrossRef] [PubMed]
- Lu-Culligan, A.; Iwasaki, A. The Role of Immune Factors in Shaping Fetal Neurodevelopment. Annu Rev Cell Dev Biol 2020, 36, 441–468. [Google Scholar] [CrossRef]
- Tomlinson, M.S.; Bommarito, P.A.; Martin, E.M.; Smeester, L.; Fichorova, R.N.; Onderdonk, A.B.; Kuban, K.C. K.; O'Shea, T.M.; Fry, R.C. Microorganisms in the human placenta are associated with altered CpG methylation of immune and inflammation-related genes. PloS one 2017, 12, e0188664. [Google Scholar] [CrossRef]
- Schaub, B.; Liu, J.; Hoppler, S.; Schleich, I.; Huehn, J.; Olek, S.; Wieczorek, G.; Illi, S.; von Mutius, E. Maternal farm exposure modulates neonatal immune mechanisms through regulatory T cells. J Allergy Clin Immunol 2009, 123, 774–82. [Google Scholar] [CrossRef] [PubMed]
- Schjenken, J.E.; Green, E.S.; Overduin, T.S.; Mah, C.Y.; Russell, D.L.; Robertson, S.A. Endocrine Disruptor Compounds-A Cause of Impaired Immune Tolerance Driving Inflammatory Disorders of Pregnancy? Front Endocrinol (Lausanne) 2021, 12, 607539. [Google Scholar] [CrossRef] [PubMed]
- Attreed, S.E.; Navas-Acien, A.; Heaney, C.D. Arsenic and Immune Response to Infection During Pregnancy and Early Life. Curr Environ Health Rep 2017, 4, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Wang, X.; Burchiel, S.W. Toxicity of environmentally-relevant concentrations of arsenic on developing T lymphocyte. Environ Toxicol Pharmacol 2018, 62, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Kile, M.L.; Houseman, E.A.; Baccarelli, A.A.; Quamruzzaman, Q.; Rahman, M.; Mostofa, G.; Cardenas, A.; Wright, R.O.; Christiani, D.C. Effect of prenatal arsenic exposure on DNA methylation and leukocyte subpopulations in cord blood. Epigenetics 2014, 9, 774–82. [Google Scholar] [CrossRef] [PubMed]
- Nygaard, U.C.; Li, Z.; Palys, T.; Jackson, B.; Subbiah, M.; Malipatlolla, M.; Sampath, V.; Maecker, H.; Karagas, M.R.; Nadeau, K.C. Cord blood T cell subpopulations and associations with maternal cadmium and arsenic exposures. PloS one 2017, 12, e0179606. [Google Scholar] [CrossRef] [PubMed]
- Farzan, S.F.; Li, Z.; Korrick, S.A.; Spiegelman, D.; Enelow, R.; Nadeau, K.; Baker, E.; Karagas, M.R. Infant Infections and Respiratory Symptoms in Relation to in Utero Arsenic Exposure in a U.S. Cohort. Environmental health perspectives 2016, 124, 840–7. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Vahter, M.; Ekstrom, E.C.; Persson, L.A. Arsenic exposure in pregnancy increases the risk of lower respiratory tract infection and diarrhea during infancy in Bangladesh. Environmental health perspectives 2011, 119, 719–24. [Google Scholar] [CrossRef] [PubMed]
- Escobar, J.; Varela-Nallar, L.; Coddou, C.; Nelson, P.; Maisey, K.; Valdes, D.; Aspee, A.; Espinosa, V.; Rozas, C.; Montoya, M.; Mandiola, C.; Rodriguez, F.E.; Acuna-Castillo, C.; Escobar, A.; Fernandez, R.; Diaz, H.; Sandoval, M.; Imarai, M.; Rios, M. Oxidative damage in lymphocytes of copper smelter workers correlated to higher levels of excreted arsenic. Mediators Inflamm 2010, 2010, 403830. [Google Scholar] [CrossRef] [PubMed]
- Luna, A.L.; Acosta-Saavedra, L.C.; Lopez-Carrillo, L.; Conde, P.; Vera, E.; De Vizcaya-Ruiz, A.; Bastida, M.; Cebrian, M.E.; Calderon-Aranda, E.S. Arsenic alters monocyte superoxide anion and nitric oxide production in environmentally exposed children. Toxicol Appl Pharmacol 2010, 245, 244–51. [Google Scholar] [CrossRef] [PubMed]
- Fry, R.C.; Navasumrit, P.; Valiathan, C.; Svensson, J.P.; Hogan, B.J.; Luo, M.; Bhattacharya, S.; Kandjanapa, K.; Soontararuks, S.; Nookabkaew, S.; Mahidol, C.; Ruchirawat, M.; Samson, L.D. Activation of inflammation/NF-kappaB signaling in infants born to arsenic-exposed mothers. PLoS Genet 2007, 3, e207. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, H.; Fu, G.; Feng, Y.; Wu, W.; Yang, H.; Zhang, Y.; Wang, S. DNA methylation analysis reveals the effect of arsenic on gestational diabetes mellitus. Genomics 2023, 115, 110674. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.D.; Lam, W.L. Health Effects Associated With Pre- and Perinatal Exposure to Arsenic. Front Genet 2021, 12, 664717. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Liu, J.; Benbrahim-Tallaa, L.; Ward, J.M.; Logsdon, D.; Diwan, B.A.; Waalkes, M.P. Aberrant DNA methylation and gene expression in livers of newborn mice transplacentally exposed to a hepatocarcinogenic dose of inorganic arsenic. Toxicology 2007, 236, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, P.; Landrigan, P.J. Neurobehavioural effects of developmental toxicity. Lancet Neurol 2014, 13, 330–8. [Google Scholar] [CrossRef] [PubMed]
- Emeny, R.T.; Korrick, S.A.; Li, Z.; Nadeau, K.; Madan, J.; Jackson, B.; Baker, E.; Karagas, M.R. Prenatal exposure to mercury in relation to infant infections and respiratory symptoms in the New Hampshire Birth Cohort Study. Environmental research 2019, 171, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Movassagh, H.; Halchenko, Y.; Sampath, V.; Nygaard, U.C.; Jackson, B.; Robbins, D.; Li, Z.; Nadeau, K.C.; Karagas, M.R. Maternal gestational mercury exposure in relation to cord blood T cell alterations and placental gene expression signatures. Environmental research 2021, 201, 111385. [Google Scholar] [CrossRef] [PubMed]
- Oulhote, Y.; Shamim, Z.; Kielsen, K.; Weihe, P.; Grandjean, P.; Ryder, L.P.; Heilmann, C. Children's white blood cell counts in relation to developmental exposures to methylmercury and persistent organic pollutants. Reproductive toxicology 2017, 68, 207–214. [Google Scholar] [CrossRef]
- Stratakis, N.; Conti, D.V.; Borras, E.; Sabido, E.; Roumeliotaki, T.; Papadopoulou, E.; Agier, L.; Basagana, X.; Bustamante, M.; Casas, M.; Farzan, S.F.; Fossati, S.; Gonzalez, J.R.; Grazuleviciene, R.; Heude, B.; Maitre, L.; McEachan, R.R. C.; Theologidis, I.; Urquiza, J.; Vafeiadi, M.; West, J.; Wright, J.; McConnell, R.; Brantsaeter, A.L.; Meltzer, H.M.; Vrijheid, M.; Chatzi, L. Association of Fish Consumption and Mercury Exposure During Pregnancy With Metabolic Health and Inflammatory Biomarkers in Children. JAMA Netw Open 2020, 3, e201007. [Google Scholar] [CrossRef] [PubMed]
- Nyland, J.F.; Fillion, M.; Barbosa, F. Jr.; Shirley, D.L.; Chine, C.; Lemire, M.; Mergler, D.; Silbergeld, E.K. Biomarkers of methylmercury exposure immunotoxicity among fish consumers in Amazonian Brazil. Environmental health perspectives 2011, 119, 1733–8. [Google Scholar] [CrossRef]
- Pinheiro, M.C. N.; Macchi, B.M.; Vieira, J.L. F.; Oikawa, T.; Amoras, W.W.; Guimaraes, G.A.; Costa, C.A.; Crespo-Lopez, M.E.; Herculano, A.M.; Silveira, L.C. L.; do Nascimento, J.L. M. Mercury exposure and antioxidant defenses in women: a comparative study in the Amazon. Environmental research 2008, 107, 53–9. [Google Scholar] [CrossRef]
- Boskabady, M.; Marefati, N.; Farkhondeh, T.; Shakeri, F.; Farshbaf, A.; Boskabady, M.H. The effect of environmental lead exposure on human health and the contribution of inflammatory mechanisms, a review. Environment international 2018, 120, 404–420. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.A.; McCabe, M.J. Jr. Immunomodulation by metals. Int Immunopharmacol 2002, 2, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-E.; Dietert, R.R. Developmental immunotoxicity of lead: impact on thymic function. Birth Defects Res A Clin Mol Teratol 2003, 67, 861–7. [Google Scholar] [CrossRef] [PubMed]
- Annesi-Maesano, I.; Pollitt, R.; King, G.; Bousquet, J.; Hellier, G.; Sahuquillo, J.; Huel, G. In utero exposure to lead and cord blood total IgE. Is there a connection? Allergy 2003, 58, 589–94. [Google Scholar] [CrossRef] [PubMed]
- Wells, E.M.; Bonfield, T.L.; Dearborn, D.G.; Jackson, L.W. The relationship of blood lead with immunoglobulin E, eosinophils, and asthma among children: NHANES 2005-2006. International journal of hygiene and environmental health 2014, 217, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Sekovanic, A.; Dorotic, A.; Pasalic, D.; Orct, T.; Kljakovic-Gaspic, Z.; Grgec, A.S.; Stasenko, S.; Mioc, T.; Piasek, M.; Jurasovic, J. The effects of maternal cigarette smoking on cadmium and lead levels, miRNA expression and biochemical parameters across the feto-placental unit. Heliyon 2022, 8, e12568. [Google Scholar] [CrossRef] [PubMed]
- Canepari, S.; Astolfi, M.L.; Drago, G.; Ruggieri, S.; Tavormina, E.E.; Cibella, F.; Perrino, C. PM(2.5) elemental composition in indoor residential environments and co-exposure effects on respiratory health in an industrial area. Environmental research 2023, 216, 114630. [Google Scholar] [CrossRef] [PubMed]
- Drago, G.; Perrino, C.; Canepari, S.; Ruggieri, S.; L'Abbate, L.; Longo, V.; Colombo, P.; Frasca, D.; Balzan, M.; Cuttitta, G.; Scaccianoce, G.; Piva, G.; Bucchieri, S.; Melis, M.; Viegi, G.; Cibella, F.; Group, R.C.P.; Indoor, *!!! REPLACE !!!*; Outdoor Air, Q.; Respiratory Health in, M.; Sicily, R.S. G.; Balzan, M.; Bilocca, D.; Borg, C.; Montefort, S.; Zammit, C.; Bucchieri, S.; Cibella, F.; Colombo, P.; Cuttitta, G.; Drago, G.; Ferrante, G.; L'Abbate, L.; Grutta, S.; Longo, V.; Melis, M.R.; Ruggieri, S.; Viegi, G.; Minardi, R.; Piva, G.; Ristagno, R.; Rizzo, G.; Scaccianoce, G. Relationship between domestic smoking and metals and rare earth elements concentration in indoor PM(2.5). Environmental research 2018, 165, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Breen, J.G.; Eisenmann, C.; Horowitz, S.; Miller, R.K. Cell-specific increases in metallothionein expression in the human placenta perfused with cadmium. Reproductive toxicology 1994, 8, 297–306. [Google Scholar] [CrossRef]
- Kippler, M.; Hoque, A.M.; Raqib, R.; Ohrvik, H.; Ekstrom, E.C.; Vahter, M. Accumulation of cadmium in human placenta interacts with the transport of micronutrients to the fetus. Toxicol Lett 2010, 192, 162–8. [Google Scholar] [CrossRef]
- Chandravanshi, L.; Shiv, K.; Kumar, S. Developmental toxicity of cadmium in infants and children: a review. Environ Anal Health Toxicol 2021, 36, e2021003–0. [Google Scholar] [CrossRef] [PubMed]
- Sanders, A.P.; Smeester, L.; Rojas, D.; DeBussycher, T.; Wu, M.C.; Wright, F.A.; Zhou, Y.H.; Laine, J.E.; Rager, J.E.; Swamy, G.K.; Ashley-Koch, A.; Lynn Miranda, M.; Fry, R.C. Cadmium exposure and the epigenome: Exposure-associated patterns of DNA methylation in leukocytes from mother-baby pairs. Epigenetics 2014, 9, 212–21. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Zhang, W.X.; Zheng, T.Z.; Zhou, B.; Li, J.X.; Zhang, B.; Xia, W.; Li, Y.Y.; Xu, S.Q. Prenatal and postnatal cadmium exposure and cellular immune responses among pre-school children. Environment international 2020, 134, 105282. [Google Scholar] [CrossRef] [PubMed]
- Pillet, S.; Rooney, A.A.; Bouquegneau, J.-M.; Cyr, D.G.; Fournier, M. Sex-specific effects of neonatal exposures to low levels of cadmium through maternal milk on development and immune functions of juvenile and adult rats. Toxicology 2005, 209, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, P.; Heilmann, C.; Weihe, P.; Nielsen, F.; Mogensen, U.B.; Budtz-Jorgensen, E. Serum Vaccine Antibody Concentrations in Adolescents Exposed to Perfluorinated Compounds. Environmental health perspectives 2017, 125, 077018. [Google Scholar] [CrossRef]
- Grandjean, P.; Heilmann, C.; Weihe, P.; Nielsen, F.; Mogensen, U.B.; Timmermann, A.; Budtz-Jorgensen, E. Estimated exposures to perfluorinated compounds in infancy predict attenuated vaccine antibody concentrations at age 5-years. J Immunotoxicol 2017, 14, 188–195. [Google Scholar] [CrossRef]
- Granum, B.; Haug, L.S.; Namork, E.; Stolevik, S.B.; Thomsen, C.; Aaberge, I.S.; van Loveren, H.; Lovik, M.; Nygaard, U.C. Pre-natal exposure to perfluoroalkyl substances may be associated with altered vaccine antibody levels and immune-related health outcomes in early childhood. J Immunotoxicol 2013, 10, 373–9. [Google Scholar] [CrossRef] [PubMed]
- Osuna, C.E.; Grandjean, P.; Weihe, P.; El-Fawal, H.A. Autoantibodies associated with prenatal and childhood exposure to environmental chemicals in Faroese children. Toxicol Sci 2014, 142, 158–66. [Google Scholar] [CrossRef] [PubMed]
- Ait Bamai, Y.; Goudarzi, H.; Araki, A.; Okada, E.; Kashino, I.; Miyashita, C.; Kishi, R. Effect of prenatal exposure to per- and polyfluoroalkyl substances on childhood allergies and common infectious diseases in children up to age 7years: The Hokkaido study on environment and children's health. Environment international 2020, 143, 105979. [Google Scholar] [CrossRef]
- Impinen, A.; Longnecker, M.P.; Nygaard, U.C.; London, S.J.; Ferguson, K.K.; Haug, L.S.; Granum, B. Maternal levels of perfluoroalkyl substances (PFASs) during pregnancy and childhood allergy and asthma related outcomes and infections in the Norwegian Mother and Child (MoBa) cohort. Environment international 2019, 124, 462–472. [Google Scholar] [CrossRef]
- Stein, C.R.; McGovern, K.J.; Pajak, A.M.; Maglione, P.J.; Wolff, M.S. Perfluoroalkyl and polyfluoroalkyl substances and indicators of immune function in children aged 12-19 y: National Health and Nutrition Examination Survey. Pediatr Res 2016, 79, 348–57. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.J.; Hsieh, W.-S.; Chen, C.-Y.; Fletcher, T.; Lien, G.-W.; Chiang, H.-L.; Chiang, C.-F.; Wu, T.-N.; Chen, P.-C. The effect of prenatal perfluorinated chemicals exposures on pediatric atopy. Environmental research 2011, 111, 785–91. [Google Scholar] [CrossRef] [PubMed]
- von Holst, H.; Nayak, P.; Dembek, Z.; Buehler, S.; Echeverria, D.; Fallacara, D.; John, L. Perfluoroalkyl substances exposure and immunity, allergic response, infection, and asthma in children: review of epidemiologic studies. Heliyon 2021, 7, e08160. [Google Scholar] [CrossRef] [PubMed]
- Heilmann, C.; Grandjean, P.; Weihe, P.; Nielsen, F.; Budtz-Jorgensen, E. Reduced antibody responses to vaccinations in children exposed to polychlorinated biphenyls. PLoS Med 2006, 3, e311. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Hertz-Picciotto, I.; Petrik, J.; Palkovicova, L.; Kocan, A.; Trnovec, T. Prenatal PCB exposure and thymus size at birth in neonates in Eastern Slovakia. Environmental health perspectives 2008, 116, 104–9. [Google Scholar] [CrossRef] [PubMed]
- Horvathova, M.; Jahnova, E.; Palkovicova, L.; Trnovec, T.; Hertz-Picciotto, I. The kinetics of cell surface receptor expression in children perinatally exposed to polychlorinated biphenyls. J Immunotoxicol 2011, 8, 367–80. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; Singh, U.P.; Guan, H.; Nagarkatti, P.; Nagarkatti, M. Prenatal exposure to TCDD triggers significant modulation of microRNA expression profile in the thymus that affects consequent gene expression. PloS one 2012, 7, e45054. [Google Scholar] [CrossRef] [PubMed]
- Gaspari, L.; Paris, F.; Kalfa, N.; Soyer-Gobillard, M.O.; Sultan, C.; Hamamah, S. Experimental Evidence of 2,3,7,8-Tetrachlordibenzo-p-Dioxin (TCDD) Transgenerational Effects on Reproductive Health. International journal of molecular sciences 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- O'Boyle, N.M.; Delaine, T.; Luthman, K.; Natsch, A.; Karlberg, A.T. Analogues of the epoxy resin monomer diglycidyl ether of bisphenol F: effects on contact allergenic potency and cytotoxicity. Chem Res Toxicol 2012, 25, 2469–78. [Google Scholar] [CrossRef] [PubMed]
- Donohue, K.M.; Miller, R.L.; Perzanowski, M.S.; Just, A.C.; Hoepner, L.A.; Arunajadai, S.; Canfield, S.; Resnick, D.; Calafat, A.M.; Perera, F.P.; Whyatt, R.M. Prenatal and postnatal bisphenol A exposure and asthma development among inner-city children. J Allergy Clin Immunol 2013, 131, 736–42. [Google Scholar] [CrossRef]
- Gascon, M.; Casas, M.; Morales, E.; Valvi, D.; Ballesteros-Gomez, A.; Luque, N.; Rubio, S.; Monfort, N.; Ventura, R.; Martinez, D.; Sunyer, J.; Vrijheid, M. Prenatal exposure to bisphenol A and phthalates and childhood respiratory tract infections and allergy. J Allergy Clin Immunol 2015, 135, 370–8. [Google Scholar] [CrossRef] [PubMed]
- Mendy, A.; Salo, P.M.; Wilkerson, J.; Feinstein, L.; Ferguson, K.K.; Fessler, M.B.; Thorne, P.S.; Zeldin, D.C. Association of urinary levels of bisphenols F and S used as bisphenol A substitutes with asthma and hay fever outcomes. Environmental research 2020, 183, 108944. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.; Hong, X.; Wang, X.; Tang, W.Y. Aberrant 5'-CpG Methylation of Cord Blood TNFalpha Associated with Maternal Exposure to Polybrominated Diphenyl Ethers. PloS one 2015, 10, e0138815. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.; Drago, G.; Longo, A.; Ruggieri, S.; Sprovieri, M.; Cibella, F.; Colombo, P. A multipollutant low-grade exposure regulates the expression of miR-30b, Let-7a and miR-223 in maternal sera: Evidence from the NEHO cohort. The Science of the total environment 2022, 844, 157051. [Google Scholar] [CrossRef] [PubMed]
- Shaddick, G.; Thomas, M.L.; Amini, H.; Broday, D.; Cohen, A.; Frostad, J.; Green, A.; Gumy, S.; Liu, Y.; Martin, R.V.; Pruss-Ustun, A.; Simpson, D.; van Donkelaar, A.; Brauer, M. Data Integration for the Assessment of Population Exposure to Ambient Air Pollution for Global Burden of Disease Assessment. Environmental science & technology 2018, 52, 9069–9078. [Google Scholar]
- Chen, Y.; Liu, S.; Xu, H.; Zheng, H.; Bai, C.; Pan, W.; Zhou, H.; Liao, M.; Huang, C.; Dong, Q. Maternal exposure to low dose BDE209 and Pb mixture induced neurobehavioral anomalies in C57BL/6 male offspring. Toxicology 2019, 418, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Mesa, M.; Narduzzi, L.; Ouzia, S.; Soetart, N.; Jaillardon, L.; Guitton, Y.; Le Bizec, B.; Dervilly, G. Metabolomics and lipidomics to identify biomarkers of effect related to exposure to non-dioxin-like polychlorinated biphenyls in pigs. Chemosphere 2022, 296, 133957. [Google Scholar] [CrossRef] [PubMed]
- Naville, D.; Gaillard, G.; Julien, B.; Vega, N.; Pinteur, C.; Chanon, S.; Vidal, H.; Le Magueresse-Battistoni, B. Chronic exposure to a pollutant mixture at low doses led to tissue-specific metabolic alterations in male mice fed standard andhigh-fat high-sucrose diet. Chemosphere 2019, 220, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, J.P.; Parent, A.S.; Kleinjans, J.C. S.; Nawrot, T.S.; Schoeters, G.; Van Larebeke, N. Rationale for Environmental Hygiene towards global protection of fetuses and young children from adverse lifestyle factors. Environmental health : a global access science source 2018, 17, 42. [Google Scholar] [CrossRef]
- Cori, L.; Bianchi, F.; Sprovieri, M.; Cuttitta, A.; Ruggieri, S.; Alessi, A.L.; Biondo, G.; Gorini, F. Communication and Community Involvement to Support Risk Governance. International journal of environmental research and public health 2019, 16. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the structure of the review.
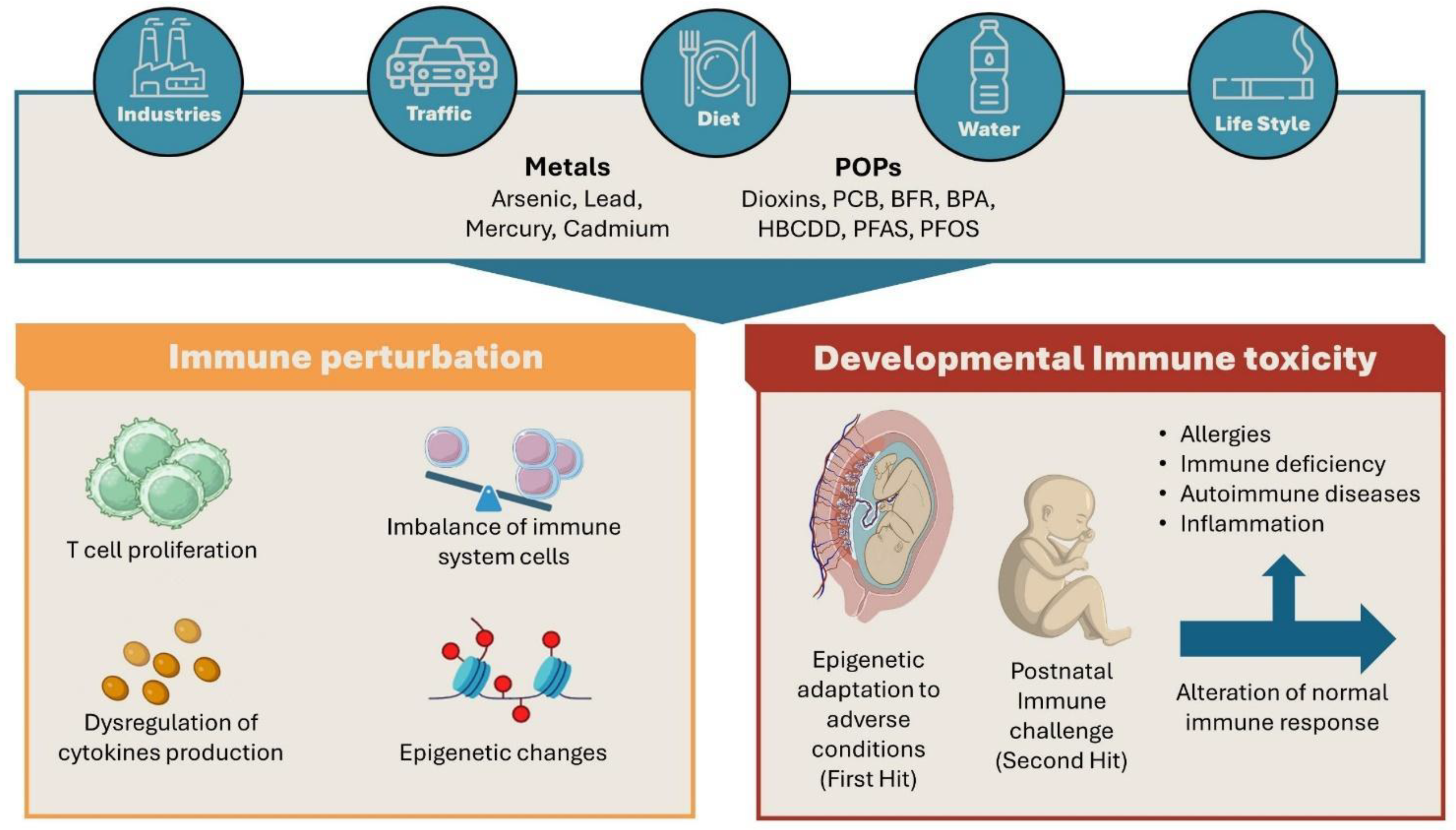
Figure 2.
Schematic representation of the data obtained by in vitro, ex vivo and in vivo studies about the effects of the main classes of environmental pollutants on immune response.
Figure 2.
Schematic representation of the data obtained by in vitro, ex vivo and in vivo studies about the effects of the main classes of environmental pollutants on immune response.
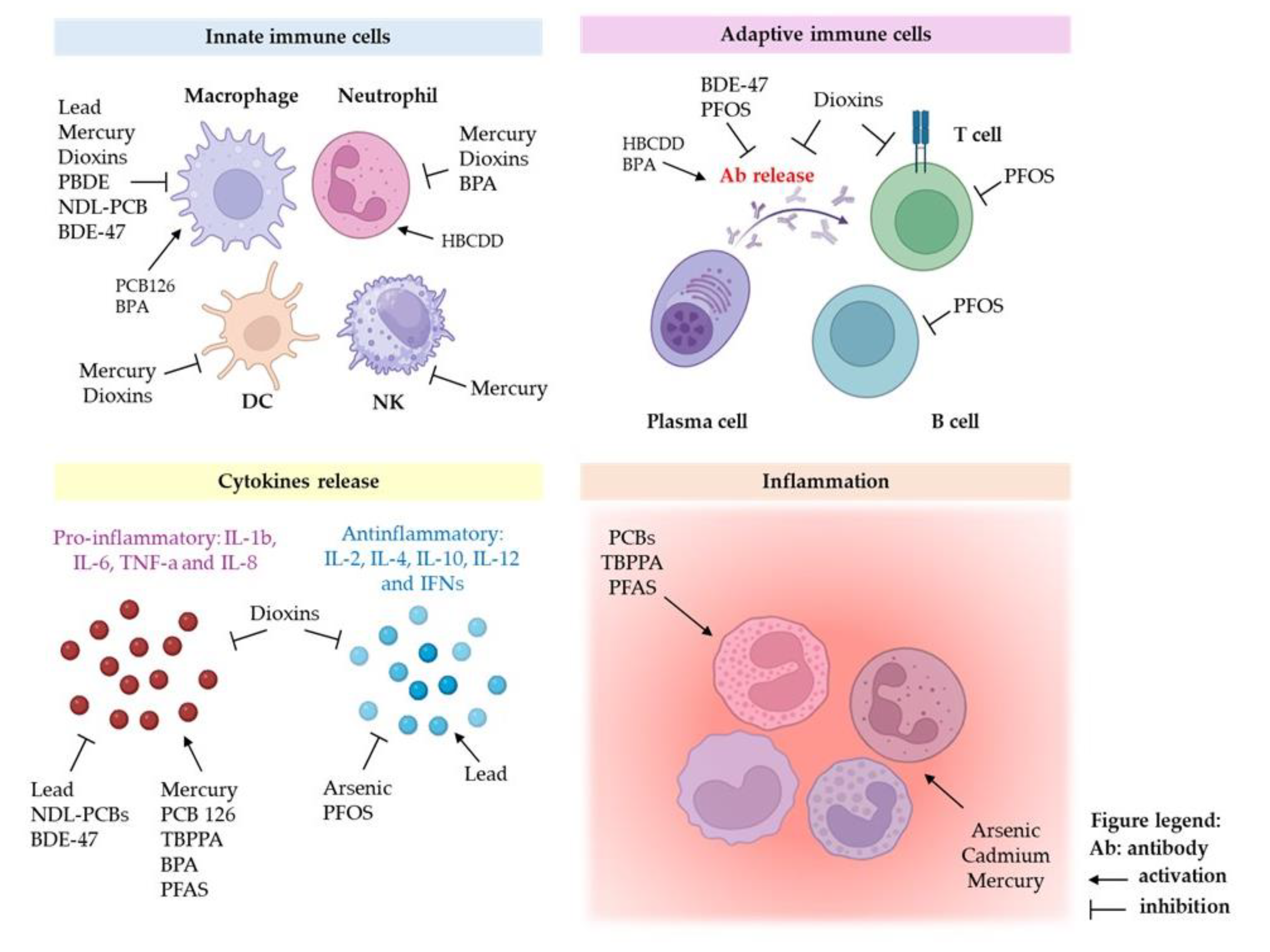

Table 1.
In vitro, ex vivo and in vivo effects of Heavy Metals.
| Pollutant | Effects | Model systems (in vitro/ex vivo/in vivo) | Reference |
|---|---|---|---|
| Arsenic | Reduction of (IFN-γ, IL-4 and IL-10) | Murine splenocytes, mouse activated T cell (ex vivo) |
[34,35] |
| Macrophages, neutrophil, B and T cell apoptosis induction |
Human primary cultures (ex vivo) |
[36] |
|
| Modulation of T cell Receptor, cell cycle and apoptosis |
Human PBMCs (ex vivo) |
[37,38] |
|
| Involvement in fibrinogenic processes and chronic hepatic fibrosis | Co-cultures of THP-1 macrophages or BMDM and primary lung fibroblasts from C57BL/6 mice (in vivo/in vitro/ex vivo); co-cultures of THP-1 monocytes and LX2 cell lines (in vitro) |
[39,40] |
|
| Hypomethylation of leukocyte DNA |
Human blood samples (ex vivo) | [42] |
|
| Oxidative stress signalling, DNA damage and cytotoxicity in T cells and in human polymorphonuclear neutrophils | Human primary cell cultures and human cell lines (ex vivo, in vitro) | [44,45] | |
| Lead | Induction of allergies, infectious and autoimmune diseases in human |
Human ex vivo studies | [46,47] |
| Dysregulation of proinflammatory cytokine and impairment of THP-1 monocyte/macrophage cell viability |
Human THP-1 Monocyte/Macrophage cultures (in vitro) |
[48,49,50] | |
| Quiescence of haematopoietic stem cells |
C57BL/6 murine model (in vivo and ex vivo); | [51] | |
| Induction of higher levels of IFN-γ, IL-2, IL-12 and IL-17 in exposed workers | Human THP-1 Monocyte/Macrophage (in vitro); Human serum samples and primary T cell cultures (ex vivo) |
[52,53] |
|
| Mercury | Genetic damage, neurological, kidney, cardiac and immunological diseases in human | In vitro, ex vivo and in vivo model systems | [56] |
| Superoxide ion production and cytotoxic effect in neutrophils |
Human neutrophils (ex vivo) |
[57] |
|
| Impairment of macrophage migratory and phagocytic activity; NO and pro-inflammatory cytokines production |
BALB/cABOM peritoneal macrophages (ex vivo); Monocytes from Human PBMCs; co-cultures of Caco-2, HT29-MTX intestinal epithelial cells and THP-1 macrophages (in vitro). |
[58,59,60] |
|
| Genotoxic effects. Suppression both tumoricidal activity of blood and splenic NK cells and T and B cells proliferation |
Human cell lines (in vitro) and blood samples (ex vivo); Balb/c mouse and rat primary cell cultures (in vivo and ex vivo) | [61,62,63] | |
| Cadmium | Inflammation and oxidative damage induction in neutrophils and macrophages | Rat liver and kidney primary cell cultures (in vivo and ex vivo); various cells and tissues in vitro and ex vivo models. | [66,67] |
|
Down regulation of TNF-α, IL-12p40, TLRs, CD14, MD2, BD2, MyD88, p65 and NOS2 |
Wild boars’ macrophages (ex vivo) |
[69] |
|
|
Impairment of adaptive immunity cell populations in offsprings |
C57Bl/6 mice (in vivo and ex vivo) |
[70,71] |
|
| Modulation of miRNAs associated with inflammation and carcinogenesis and alteration oh Th17 and Treg lymphocytes subpopulation in exposed workers | Human blood samples (ex vivo) | [72] |
Table 2.
In vitro, ex vivo and in vivo effects of POPs.
| Pollutant | Effects | Model systems (in vitro/ex vivo/in vivo) | Reference | |||
|---|---|---|---|---|---|---|
| Dioxins | Impairment of macrophages, NK, neutrophils and dendritic cells |
in vivo murine models; in vitro and ex vivo cell cultures |
[76,77,78] |
|||
| Reduction both of antibody’s production by B cell and cytotoxic activity of T lymphocytes |
In vitro and ex vivo cell cultures; in vivo murine models |
[79] | ||||
| Attenuation of IgE mediated hypersensitivity response |
In vivo and ex vivo studies |
[80] |
||||
| Alterations of THP-1 macrophages adherence, adhesion molecule expression, morphology, multiple cytokine/chemokine production and total mRNA expression |
THP-1 monocyte/macrophage cell line (in vitro) | [81] |
||||
| Effects on human reproductive health | Murine models; Human ex vivo studies |
[82] | ||||
| PCBs | Proinflammatory activity in macrophages |
THP-1 monocyte/macrophage cell line (in vitro) |
[83] |
|||
| In vitro immunosuppressive effects and expression of reactive species | J774A.1 cell line and primary murine macrophages (in vitro and ex vivo) | [84] |
||||
|
In vivo inflammatory effects and impairment of immune system functions in mouse model |
Wild-type mice C57BL/6J and SVJ129 (in vivo and ex vivo) |
[86] |
||||
| DNA methylation differences in PCB-exposed populations | Human PBMCs (ex vivo) |
[87] | ||||
| PBDEs | Liver, kidney, gut, and thyroid toxicity |
In vitro, ex vivo and in vivo studies |
[91] |
|||
| Neurotoxic, cardiotoxic, hepatotoxic and teratogenic effects on zebrafish and fish |
Zebrafish embryos (in vivo and ex vivo); | [92,93,94] | ||||
| Impairment of proinflammatory response modulation of small extracellular vesicle biogenesis and miRNA cargo and exacerbation of LPS-induced pro-inflammatory response in macrophage cell line. |
THP-1 monocyte/macrophage cell line (in vitro) |
[95,96,97] |
||||
| Alteration of tight junctions, adhesion molecules, cytokines and EMT (epithelial-mesenchymal transition) markers expression in epithelial lung cells |
ALI cultures of human A549 cell line (in vitro) | [98] |
||||
| Cardiovascular toxicity |
THP-1 monocyte/macrophage cell line (in vitro) |
[99] | ||||
| Destruction of macrophage functional activity in animal model systems |
RTG-2 cell line (in vitro) |
[100,101,102] | ||||
| Reduction of antibody response, histopathological effects on liver, spleen, small intestine, and thyroid |
BALB/c murine model (in vivo and ex vivo) | [103] | ||||
| HBCDD | Bioaccumulation in human blood, adipose tissue and breast milk |
Human blood, adipose tissue and maternal milk (ex vivo) |
[109,110,111] | |||
| TBBPA | Genotoxic effects | Human PBMCs (ex vivo) |
[112,113,114] |
|||
| Induction of inflammatory phenotype in human dendritic cells from healthy subjects | Human monocyte-derived dendritic cells (ex vivo) | [115] |
||||
|
Neurotoxic, nephrotoxic, hepatotoxic and immunotoxic effects |
[116] |
|||||
| Correlation with human thyroid and neurological disorders, reproductive health, immunological, oncological, and cardiovascular diseases |
Laboratory animals and human samples (in vivo and ex vivo) | [117,118] | ||||
| BPA | Impairment of neutrophils chemotactic function |
Neutrophils isolated from human blood | [119] |
|||
| Induction of M1 macrophages polarization | Peritoneal macrophages from C57BL/6 J mice (ex vivo) |
[120] |
||||
| Upregulation of proinflammatory cytokines | RAW264.7 murine macrophage cell line (in vitro) |
[121,122] |
||||
| Immunotoxic effects on adaptive immune response and effect on IgM reactivity against tumour antigens | Guinea pig model system (in vivo and ex vivo) |
[123] | ||||
| Inhibition of T cell proliferation | BALB/c murine model (in vivo and ex vivo) |
[102] |
||||
| PFAS (PFOS and PFOA) |
PFOS bioaccumulation in human liver, kidneys, lungs, hair, breast milk urine and blood |
Human blood, urine, milk, hair, nails and tissue samples |
[125,126] | |||
| In vitro reduction of IL-2 expression in human T cells | Jurkat cell line and primary T cell (in vitro and ex vivo) | [127] |
||||
| De novo localization of immune cell populations in organs like the spleen and liver | C57BL/6 murine model (in vivo and ex vivo) |
[128] | ||||
| Reduction of B-cell subtypes and IgM antibody primary response in female mice induced by PFOA | C57BL/6 murine model (in vivo and ex vivo) |
[129] | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



