
Submitted:
24 June 2024
Posted:
25 June 2024
You are already at the latest version
Abstract
Brucellosis is a zoonoses caused by bacteria of the genus Brucella, which causes economic losses in livestock and threatens public health. A cross-sectional study was conducted to determine the prevalence of Brucella species and risk factors in smallholder dairy cattle in six regions of Tanzania from July 2019 to October 2020. Dairy cattle (n=2048) were sampled from 1371 farms. DNA ex-tracted from blood and vaginal swabs was tested for Brucella using qPCR targeting the IS711 gene and positives were tested for the alkB marker for B. abortus and BMEI1172 marker for B. melitensis. The molecular prevalence was 3.5% (95% CI: 2.8-4.4) with the highest prevalence 8.1% (95%CI: 4.6-13.0) in Njombe region. B. melitensis was the predominant species detected (66.2%). Logistic regression revealed PCR positivity in dairy cattle was higher in the animals with a history of abortion (OR: 3.17, 95%CI: 1.05-9.62) and farms without dogs around (OR: 2.68, 95%CI: 1.12-5.90). Further studies are recommended to understand the source of B. melitensis and the implications to veterinary public health. Livestock keepers should be informed of the risks and biosecurity practices to reduce introduction and control Brucella. Cattle and small ruminant vaccination pro-grams could be implemented to control brucellosis in high-risk populations in the country.
Keywords:
dairy cattle
; brucellosis
; qPCR
; molecular prevalence
; risk factors
; Brucella
; Tanzania
1. Introduction
Brucellosis is a zoonotic bacterial disease causing economic loss in dairy cattle production systems. Brucellosis is considered to be one of the most widespread zoonotic diseases globally [1]. The disease is caused by a bacterium of the Genus Brucella. Of the twelve Brucella species identified that affect mammals, the common species affecting domestic animals are B. abortus in cattle, B. melitensis in goats, B. ovis in sheep, B. suis in pigs and B. canis in dogs [2,3]. Brucella spp. are somewhat host specific, however, recent studies highlighted the importance of cross-species infection [4,5,6]. Studies have found that, brucellosis in cattle can also be caused by B. melitensis or B. suis [2,7,8]. This renders eradication through vaccination with B. abortus derived vaccines ineffective, since the efficacy of the S.19 vaccine which is widely used in endemic areas has not been fully validated against B. melitensis and those vaccines which confer cross-protection may not be available especially in LMICs [9,10,11].
In Tanzania, the first isolates of B. abortus and B. melitensis from cattle and goats respectively was done in 1967 [12], however no typing of the isolates was done [12]. In 2015, B. abortus was isolated from aborted materials of dairy cattle in Njombe region and the first typing identified B. abortus biovar 3 [13]. Around the same time B. abortus biovar 1 was detected and typed from cow’s milk [14]. Over recent years mixed farming practices have been reported to be associated with brucellosis reemergence in Tanzania [15,16]. Research in neighboring countries have identified cattle infected with other Brucella species. Studies in Kenya, Uganda and Rwanda have identified B. melitensis and B. abortus from dairy cattle [6,8,17]. Furthermore, B. melitensis the most pathogenic among the classical Brucella species has been frequently isolated from febrile human patients in northern Tanzania [18], however the possible sources of infection in humans was not established. The authors, however concluded that to control human brucellosis, vaccination should also target small ruminants by using B. melitensis vaccine [18]. In Tanzania there have not been any reports of the isolation or molecular detection of B. melitensis in cattle. Therefore, the objective of the current study was to identify Brucella species circulating in smallholder dairy cattle populations in Tanzania by using molecular techniques and establish risk factors associated with brucellosis molecular positivity.
2. Materials and Methods
2.1. Study Area and Design
A cross-sectional study was conducted from July 2019 to October 2020 to determine Brucella species circulating in smallholder dairy farming systems in two agroecological zones comprising six administrative regions of Tanzania. The study was conducted in three regions in the northern zone (Kilimanjaro, Arusha and Tanga) with 252,554 head of dairy cattle and three regions in the southern highland zone (Iringa, Njombe and Mbeya) with 103,306 head of dairy cattle (Figure 1). These regions have the highest density of smallholder dairy cattle in Tanzania [19,20]. According to the household budget survey of 2018, all six regions were above the food poverty line of TSH 33,748 9 (US$13) per person per month [21]. All study regions practice mixed farming where dairy cattle interact with other domestic animals [16].
2.2. Study Population
Dairy cattle kept under smallholder farming systems were the target for the study. Dairy cattle in these regions were mainly crossbreds of Friesian, Ayrshire, and Jersey with Tanzanian Short Horn Zebu (TSHZ) and Friesian crosses comprising the largest proportion (80%) of breeds. The feeding management systems of these dairy cattle were two (1) an intensive management system in which pastures were cut and carried to the farm for them to feed and (2) an extensive system in which cattle were left to graze in private or communal land. Dairy cattle in this study were selected from a subset of the dairy cattle registry of the Africa Dairy Genetics Gains (ADGG) program (https://data.ilri.org/portal/dataset/adgg-tanzania, accessed on 1 June 2019). The ADGG project randomly registered over 52,500 cattle across the study regions in the database, furthermore 4000 dairy cattle were randomly selected and genotyped [22]. The sample for this study was selected from the genotyped animals, although at the time of the study not all the genotyped animals were available because of a high rate of animal take off (sale, natural death and slaughter) and the time that elapsed between genotyping and sampling.
2.3. Questionnaire Administration for Risk Factors Determination
A cross-sectional survey was conducted in 1371 farms and a questionnaire tool used to collect data on possible risk factors for brucellosis. The questionnaire covered details of the respondent and their brucellosis knowledge, farm location, and herd management practices such as feeding and watering, disease control, disposal of abortion material, mixed farming practices and cow or farm abortion history. The tool was developed and piloted before a final version was uploaded using the Open Data Kit (ODK) software platform on the International Livestock Research Institute (ILRI) server in Kenya (Supplementary Material S1). Then, the forms were downloaded for completion on farm from the Google play app store onto an Android tablet (Samsung, Suwon-si, Republic of Korea). Farmers or their representative were informed on the study aims and General Data Protection Regulation (GDPR) compliance. Farmers needed to sign a consent form before being interviewed in Kiswahili, and their cattle sampled.
2.4. Blood and Swab Sampling from Dairy Cattle and Samples Storage
A total of 2049 dairy cattle were sampled, and 5mls blood was collected aseptically by venipuncture into EDTA as was explained by others [23]. Animal identification number, date of collection and field barcode were labelled on each tube. In the field, all samples were kept in a cool box containing ice packs and transported daily to the field laboratory for storage. Similarly, vaginal swabs were collected aseptically following cleaning of the vulva using chlorhexidine-soaked paper towel. The vulva lips were opened by using fingers and a long shaft swab was inserted per vaginum and the mucosa was gently swabbed by rotating the swab shaft left and right while removing. After removing, the swab was then inserted in a cryovial tube containing sterile 1ml phosphate buffered saline (PBS) and squeezed on the tube wall while mixing, the shaft was then cut-off and thrown in a waste disposal bin leaving the swab tip in the PBS cryovial. The PBS cryovial was then labeled with collection date, barcoded, and scanned into the ODK form. Both EDTA blood tubes and PBS swab tubes were stored at -20°C in upright position until they were transported to Nelson Mandela African Institution of Science and Technology laboratory in Arusha for longer term storage.
2.5. DNA Extraction from EDTA Blood and PBS Swabs
EDTA tubes containing blood and PBS tubes containing a swab were allowed to thaw at room temperature on a table. After thawing each tube was briefly mixed by vortexing. Three hundred microliter (300µl) of blood/PBS was aliquoted and placed in a sample autoplate of TANBeads®. Total genomic DNA extraction was done by using TANBeads® Nucleic Acid Extraction Validation Kit (OptiPure Blood DNA Auto Plate) designed for use with the Maelstrom 9600 (Taiwan Advanced Nanotech Inc, Taoyuan City, Taiwan), a robotic system in ILRI laboratories in Nairobi, Kenya. The extraction kit was suitable for isolating DNA from whole blood including deep frozen blood and it used the silicone dioxide layer coated on the magnetic beads. A genomic DNA extract was provided in 100µl. After the extraction, random DNA samples were tested for quality and degradation by using Nanodrop spectrophotometer and 1% agar-rose gel electrophoresis respectively (Supplementary material S2).
2.6. Real Time PCR for Brucella Genus Detection and Species Characterization
The QuantiStudio 5 qPCR machine (Applied Biosystems, Woodlands, Singapore) with the 96 well plate format and 0.2ml block installed with QuantiStudio TM Design and Analysis software v1.5 was used for the analysis.
For Brucella genera detection, the DNA samples were tested for the presence of insertion sequence IS711 by using the IS711 primer pair and probe Table 1 in a uniplex assay. The qPCR conditions and reaction volumes used were adopted from Akoko et al., [5]. The reaction volumes were prepared as follows; 5µl of ready to use Master Mix (Lunar Universal Probe qPCR Master Mix, New England BioLabs, MA, USA), 1.25µl of IS711 Primer and Probe mix (Macrogen, Netherlands), 1.75µl molecular grade water and 3µl DNA template. The total reaction mix volume (cocktail) of 11µl was then thoroughly mixed up gentle vortexing for 1minute before setting the 96 wells PCR plate into the qPCR machine.
The thermo cycler conditions were set as for pretreatment with Uracil-DNA Glycosylases (UDGs) at 50°C for 2min, polymerase activation and DNA denaturation at 95°C for 10min, amplification step at 95°C for 15s, then 1min for annealing at 60 °C and 42 cycles.
Positive DNA samples based on the IS711 gene marker (positive to Brucella genus) were further characterized for B. abortus and B. melitensis using alkB and BMEI1172 primers and probe respectively Table 1.
The reaction volumes used for B. abortus and B. melitensis specific assays were 7.5µl Master mix (Perfecta qPCR ToughMix UNG Low ROX), 0.75µl Primer and probe (species-specific), 1.75µl Molecular grade water, and 5µl of DNA template. The final reaction mix (cocktail) of 15µl was thoroughly mixed by vortexing for 1 minutes before setting the 96 well PCR plate into the qPCR machine.
The PCR conditions for Brucella species-specific assays were as follows; pretreatment stage with Uracil-N-Glycosylase (UNG- step) was set at 45°C for 5min to cleave all contaminating templates containing U bases, DNA denaturation at 95°C for 5min, amplification at 95°C for 15 seconds, annealing at 60°C for 30 seconds and 42 number of cycles.
Positive controls for Brucella strains, B. melitensis 16M and B. abortus 544 both sourced from Friedrich-Loeffler-Institute, Germany. For negative controls, a mixture of RNAase free molecular grade water and master mix was used. The assay efficiency statistics and limit of detection of the ten-fold serial dilution of the reference materials for genus and species detection are provided in the Supplementary material S3.
2.7. Data Analysis for Calculation of Molecular Prevalence and Risk Factors
Data analysis was carried out using R software (version 4.2.3) [24] with individual animals as primary sampling unit and district as the clustering unit rather than herd because so many herds were small and had only 1 animal sampled. The prevalence was estimated as ratio of PCR positives (numerator) and total number of animals tested (denominator) for overall and regional prevalences and binom.test function was used to generate the 95% binomial confidence interval. The design adjusted overall prevalence was estimated after incorporating the different sampling weights (Table 3) into the estimation using the svydesign, confint and svyby functions in the survey package [25]. This allowed the stratified study design to be accounted for in the prevalence estimates.
The univariable analysis was done by using the R packages epitools and epitab to calculate the odds ratio between PCR positivity (combined blood and swab results) and independent variables (risk factors) and the unadjusted 95% confidence interval and p-value. Independent variables with p value ≤0.2 were carried forward to multivariable analysis. None of the risk factors was significantly associated with the outcome of interest when the full data set for all six regions was used, therefore the dataset was sub-setted to include only four regions with the highest molecular prevalence (Njombe, Kilimanjaro, Mbeya and Arusha Regions). Collinearity was tested by calculating the Spearman’s correlation coefficient rho from a pair of continuous variables by using cor.test function of the stat package, highly correlated variables (rho >0.29) were not included (years of experience in dairy farming and animal age) in the models. A multivariable logistic regression was fitted using the R package glmmTMB, a saturated model was fitted, and variables selected by backward elimination to get the final model. Model comparisons were done using Akaike information criterion (AIC) (AIC function in the MuMIn package) and lrtest function (lmtest package) to estimate the best fitting model. The final model validation and diagnostics was assessed using the DHARMa package.
3. Results
3.1. Description of Sampled Dairy Cattle
A total of 2049 dairy cattle were sampled from 1371 farms across the six study regions. The median herd size was 2 cattle. The majority of sampled cattle were female (97.2%). The predominant breed was SHZ-Friesian crosses (68.7%), with other breeds being SHZ-Ayrshire (20.8%), SHZ-Jersey (6.9%) and indigenous breeds (3.6%).
3.2. Brucellosis Molecular Prevalence of Dairy Cattle in Selected Regions of Tanzania
There were 35/2046 blood and 37/1893 swab samples Brucella genus-positive (Table 2). There was no agreement between sample types.
The overall unadjusted animal molecular (PCR) prevalence (being positive by either blood or vagina swab) was 3.5% (95% CI: 2.8-4.4) and the overall design adjusted prevalence was 3.7% (96%CI: 2.7-3.7). Among the study regions Njombe region had the highest molecular (PCR) prevalence of 8.1%, followed by Arusha 4.7%, Mbeya 3.8% and Kilimanjaro region 3.7% (Table 3 and Figure 2).
3.3. Brucella Species Circulating in Dairy Cattle Population Identified from Brucella Genus Positive Swab and Blood Samples
The majority of blood samples 19/35 (54.3%) and vaginal swabs 29/37 (78.4%) were PCR-positive for B. melitensis only, with a further 10/35 (28.6%) blood samples and 7/37 (18.9%) swabs PCR-positive for both (B. melitensis and B. abortus), meaning the vast majority of infections involved B. melitensis (Table 4). B. abortus occurred on its own in 2/35 (5.7%) blood samples. There were 4 blood and 1 swab samples where no species was identified (Table 4).
3.4. Brucellosis Hotspot Areas
Figure 2 is the spatial choropleth map of Tanzania (main) with the right bottom inset shows the study regions and their molecular prevalence with the highest prevalence in Njombe region (dark blue) and the main map shows molecular prevalence of the study districts (local authorities) for each study region. The main map shows that PCR-positive animals were clustered within a small number of local authorities in the Kilimanjaro, Mbeya, Arusha and Njombe regions.
3.5. Univariable Analysis Results to Combined PCR Positivity
The univariable analysis is presented in Table 5. There was a significant association between keeping a bull and reduced Brucella PCR positivity (p<0.05). Dairy cattle kept on a farm that kept a bull were 0.46 times less likely to be Brucella PCR-positive (OR: 046, 95%CI: 0.21-1.03) when compared to dairy farms which did not keep bull. There was an association between dairy breeds and Brucella PCR-positivity, with Jersey and indigenous SHZ more likely to be Brucella PCR positive compared to Ayrshire (OR: 2.62, 95%CI; 0.97-7.16). There was a positive association between Brucella PCR positivity and history of abortion at the animal level (OR: 2.51, 95%CI: 0.86-7.38), and history of abortion on the farm level (OR: 1.51, 95%CI 0.58-3.89). Further associations were shown between Brucella PCR positivity and keeping pigs dogs or having pigs around and reduced risk of an animal being Brucella PCR positive ((OR: 0.55, 95%CI: 0.32-0.93), or with having contact with dogs (OR:0.5, 95%CI:0.25-0.99) and increased risk with introduction of new animals (OR: 1.53, 95%CI: 0.71-3.31). Regionally there does also appear to be a much higher risk of an animal being Brucella PCR positive if it is in Njombe compared to Arusha (OR:1.76, 95%CI: 0.84-3.69).
3.6. Multivariable Logistic Regression Model for Association with Brucella PCR Positivity
The final logistic regression model is presented in Table 6. The model revealed that history of abortion of the animal was significantly associated with Brucella PCR positivity (p>0.05). Dairy cattle with a history of abortion were 3.17 times more likely to be Brucella PCR positive (OR: 3.17, 95%CI: 1.05-9.62) compared to those with no history of abortion. Dairy cattle in a farm with dogs around on the farm were less likely to be Brucella PCR positive (OR: 0.37, 95%CI: 0.18-0.77) than those in a farm which has dogs around on the farm or the neighbours farm. Even though there was no statistical significance (p>005), results showed that dairy cattle in a farm which introduced a new animal (dairy cattle) were 1.87 times more likely to be Brucella PCR positive (OR: 1.87, 95%CI: 0.82-4.24) than those in a farm which did not introduce a new animal. Keeping pigs or contact with neighbor’s pigs (like with dogs) was protective with Brucella PCR positivity (p<0.05) and dairy cattle in a farm which keeps pigs or having pigs around were less likely to be Brucella PCR positive (OR: 0.46, 95%CI: 0.25-0.83) compared to those in a farm which do not keep pigs or have pigs around.
Few dairy farms keep bulls, the regression model revealed that, dairy cattle in farms which kept a bull were less likely (OR:0.39, 95%CI: 0.16-0.96) to be Brucella PCR positive compared to those farms which did not keep a bull. Njombe region was significantly associated with Brucella PCR positivity, and dairy cattle in Njombe was 2.72 times more likely of being Brucella PCR positive (OR: 2.72, 95%CI: 1.21-6.10) than those in Arusha region.
4. Discussion
Brucellosis is a globally neglected bacterial zoonosis. It was characterized for the first time in Tanzania from domestic animals in 1967 and later in 2015 [12,13,14]. In Tanzania, brucellosis in dairy cattle is endemic and has been affecting dairy production and public health apart from short period in the late 1990’s when it was controlled [26]. In Tanzania, most brucellosis studies have depended on the use of serological tests to make recommendations and conclusions on the best way to control the disease in animals and have assumed Brucella host-specificity due to lack of serological tools to differentiate them [27]. However, recent studies have shown that host-specificity of Brucella species no longer applies, as cross-infections have been recently reported globally [6,28]. Therefore, molecular characterization of Brucella species is becoming increasingly important to understand the different epidemiology and make appropriate recommendations for control and eradication [29].
The current study reports the overall animal level adjusted PCR prevalence (molecular prevalence) of 3.5% across the study regions. The molecular prevalence reported in this study is lower than the molecular prevalence of 18.9% reported in Kenya [5] and similar to the 5.6% reported in Rwanda [6]. The discrepancy in prevalence is likely due to differences in study population and study locations as brucellosis prevalence has been reported to be lower in highland areas in Kenya [30] which have a similar agroecology to the areas in this study. To control the disease in the country, high-risk regions such as Njombe, Kilimanjaro, Mbeya and Arusha need to be prioritized for disease interventions [16].
The current study has identified that B. abortus, B. melitensis and unidentified Brucella species are circulating in this dairy cattle population. The current study revealed that dairy cattle are predominantly PCR-positive with B. melitensis, which is generally considered to be a pathogen of sheep and goats. Although, we did not find a statistically significant association between keeping goats and Brucella PCR positivity in this study, our previous work has demonstrated an association with seropositivity, with the odds of cattle being seropositive on a farm that keeps goats being 3.02 times compared to cattle on a farm that do not keep goats [16]. The role of goats in the epidemiology of cattle brucellosis in SSA has also been reported in previous studies [15].
The current study also reports dairy cattle PCR-positive for two Brucella species, B. abortus and B. melitensis and an unidentified Brucella species. Co-infections with more than one Brucella species have been reported in neighboring Rwanda and other African countries [6,28,31,32]. Furthermore, PCR-positivity for two Brucella species was attributed to the mingling of cattle and small ruminants [33,34]. The detection of an unidentified Brucella species highlights the potential of infection with other Brucella species such as B. ovis, B. canis, and B. suis which have been identified in dairy cattle following natural infection in different countries [35,36,37] and may be related to the presence of other domestic animals such as sheep, dogs and pigs in the dairy farms.
The multivariable model developed for the regions with the highest number of PCR positive animals (Njombe, Kilimanjaro, Mbeya and Arusha) demonstrated that the odds of Brucella PCR positivity in a dairy farm with dogs was 0.37 times when compared to farms that do not have dogs around. Earlier studies reported that feeding dogs fetal membranes and aborted material was found to be a common practice in 62.5% of cattle farmers [38]. This is further evidenced by detection of B. melitensis and B. abortus from herding dogs [39,40]. It is likely that dogs eating placenta and other aborted material reduce the chance to spread the bacteria in the environment, however this finding was contrary to the finding of a recent study in India which found that the odds of disease was 2.55 times in a farm that have dogs around compared to those with no dogs around [41]. This study also revealed that the odds of Brucella PCR positivity in a farms that do have pigs was 0.46 times when compared to the farms not having pigs around. This finding disagreed with the finding of other studies which found that presence of pigs in a farm was associated with the risk of brucellosis in cattle [42]. Although dogs and pigs may clean up the infected placenta and aborted materials and hence reduce the risk of infection to dairy cattle, this should not be encouraged and alternative control methods such as burning or burying the placenta are preferred.
History of abortion of an animal was significantly associated with Brucella PCR positivity, the odds of Brucella PCR positivity was 3.17 in dairy cattle with a history of abortion when compared to animals with no history of abortion. This finding was in agreement with the finding of other studies that the odds of the disease was higher in cattle with history of abortion [43,44,45]. Brucellosis is characterized by late-term abortion, even though is not pathognomonic sign, abortion may still be indication to implement strategies for brucellosis control in a dairy farm. Even though it was not statistically significant, introductions of new animals (cattle) in a farm was associated with Brucella PCR positivity in dairy cattle. However, other studies found that introduction of new animals to a farm was significantly associated with the risk of brucellosis in cattle [46,47] regardless of the source of the animal [48]. Testing new animals for brucellosis should be prioritized as an effort toward the control brucellosis in the region. This study revealed that the odds of Brucella PCR positivity of a dairy cattle on a farm that kept its own bull was 0.39 times less than dairy cattle on a farm that did not keep a bull. This finding was in agreement with the finding of other studies which found that farms which used own bull were at low risk of brucellosis [46]. Unlike borrowing a bull for breeding, keeping a bull reduces the risk of Brucella infections among dairy cows on a farm. Njombe region was significantly associated with Brucella PCR positivity, and dairy cattle in Njombe was 2.72 times more likely to be Brucella PCR positive than dairy cattle in Arusha. This suggests that Njombe is a brucellosis high-risk region and should be a focus for rolling-out strategies to control the disease.
The presence of B. melitensis and mixed B. abortus and B. melitensis PCR positives in dairy cattle pose a challenge to controlling the disease by vaccination. The monovalent B. abortus S19 vaccine which is produced in Tanzania may not be effective in controlling the disease under these scenarios (B. melitensis and co-infections) as the vaccine has not been fully validated for conferring cross-protection and alternative vaccines which may confer protection may not be available for use in LMICs [9,10,11]. Further validation of the currently available vaccines under this scenario is required, furthermore production of a bivalent vaccine (containing B. abortus and B. melitensis) might assist in control and eradication of the disease in cattle [49,50].
There were limitations of the study as there was no agreement in PCR positivity between vaginal swab and blood from the same animal which could be attributed by the short and transient bacteremia in cattle and that shedding of bacteria in vaginal samples tends to be post calving. Furthermore, poor agreement could have been attributed by long storage of samples for over one year in a deep freezing temperature (-20°C) which is likely to degrade the samples. There are no studies that have looked at the different sample types in cattle, however, a study in dogs compared blood and vaginal swabs and found moderate agreement between vaginal swab PCR and blood PCR, in addition there were more blood PCR positive and no vaginal swab PCR positive in a bacteremic group of bitches [51]. Finally, there was not sufficient DNA to allow sequencing for further characterization of the pathogens.
5. Conclusions
The current study confirms that bacteria of the genus Brucella are circulating in smallholder dairy cattle suggesting that brucellosis is present and likely to be causing clinical disease in dairy cattle. The importance of B. melitensis infections in smallholder dairy cattle is not clear and further understanding of the clinical significance and veterinary public health implications is needed.
This study recommends further studies on Brucella species circulating in dairy cattle and studies on the roles of small ruminants and other domestic animals in the epidemiology of brucellosis in dairy cattle. Training of famers in good biosecurity and control methods is recommended, as well as vaccination of dairy cattle in high-risk populations.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, I.J.M., G.M.S., E.A.J.C. and B.M.d.C.B.; methodology, I.J.M., G.M.S., E.A.J.C., B.M.d.C.B., S.K.M., S.F.B. and L.E.H.-C.; formal analysis, I.J.M., B.M.d.C.B., and L.E.H.-C.; resources, E.A.J.C. and B.M.d.C.B., data curation, I.J.M., S.K.M., S.F.B., B.M.d.C.B. and L.E.H.-C.; writing—original draft preparation I.J.M.; writing—review and editing, I.J.M., G.M.S., E.A.J.C., B.M.d.C.B., L.E.H.-C., E.L. and D.M.K.; supervision, G.M.S., B.M.d.C.B. and E.A.J.C.; project administration, G.M.S., B.M.d.C.B. and E.A.J.C.; funding acquisition, B.M.d.C.B. and E.A.J.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was conducted as part of the CGIAR Research Program on Livestock. ILRI is supported by contributors to the CGIAR Trust Fund. CGIAR is a global research partnership for a food-secure future. Its science is carried out by 15 Research Centres in close collaboration with hundreds of partners across the globe (www.cgiar.org, accessed on 10 December 2018). This research was funded in part by the Bill & Melinda Gates Foundation and with UK aid from the UK Foreign, Commonwealth and Development Office (Grant Agreement OPP1127286) under the auspices of the Centre for Tropical Livestock Genetics and Health (CTLGH), established jointly by the University of Edinburgh, SRUC (Scotland’s Rural College), and the International Livestock Research Institute (ILRI). This work was also supported by funding from the BBSRC (BBS/E/D/30002275).
Institutional Review Board Statement
Ethics of the study for animal subjects was reviewed and approved by the International Livestock Research Institute Institutional Animal Care and Use Committee (IL-RI-IACUC2018-27) and the research permit was granted by the Tanzania Commission for Science and Technology (COSTECH), Ref. (2019-207-NA-2019-95).
Informed Consent Statement
Consent forms were signed by cattle owners prior to the interview and sample collection.
Data Availability Statement
All relevant data are presented within the manuscript.
Acknowledgments
B.M.d.C.B.: E.A.J.C., I.J.M., S.F.B., S.K.M. and L.H.C were supported by the Bill & Melinda Gates Foundation and by a UK aid from the UK Foreign, Commonwealth, and Development Office (Grant Agreement OPP1127286) under the auspices of the Centre for Tropical Live-stock Genetics and Health established jointly by the University of Edinburgh, Scotland’s Rural College, and the ILRI. Tanzania Livestock Research Institute (TALIRI) and the Local Government Authorities offices are recognized for their support. Thank you to the participants for their time. For the purpose of open access, the author has applied a Creative Commons Attribution (CC BY) license to any Author Accepted Manuscript version arising from this submission.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schelling, E., et al., Brucellosis and Q-fever seroprevalences of nomadic pastoralists and their livestock in Chad. Preventive Veterinary Medicine 2003, 61, 279–293. [CrossRef] [PubMed]
- World Organization for Animal Health (WOAH)., Brucellosis. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, twelfth edition 2023. Chapter 3.1.4, 2023: p. 1-35.
- Pappas, G. The changing Brucella ecology: novel reservoirs, new threats. International Journal of Antimicrobial Agents 2010, 36, S8–S11. [Google Scholar] [CrossRef] [PubMed]
- Corbel, M.J. Brucellosis in Humans and Animals. 2006, Switzeland: World Health Organization (WHO), Food and Agriculture Organization (FAO) and World Organization for Animal Health (OIE).
- Akoko, J.M., et al., Molecular epidemiology of Brucella species in mixed livestock-human ecosystems in Kenya. Scientific Reports 2021, 11, 2045–2322. [CrossRef]
- Ntivuguruzwa, J.B., et al., Seroprevalence of brucellosis and molecular characterization of Brucella spp. from slaughtered cattle in Rwanda. PLoS ONE 2022, 17, e0261595. [CrossRef]
- ElTahir, Y., et al., Investigation on Brucella infection in farm animals in Saham, Sultanate of Oman with reference to human brucellosis outbreak. BMC Veterinary Research 2019, 15, 1–8. [CrossRef]
- Muendo, E.N., et al., Infection of cattle in Kenya with Brucella abortus biovar 3 and Brucella melitensis biovar 1 genotypes. Tropical Animal Health and Production 2012, 44, 17–20. [CrossRef] [PubMed]
- Moriyón, I., et al., Rough vaccines in animal brucellosis: structural and genetic basis and present status. Veterinary Research 2004, 35, 1–38. [CrossRef] [PubMed]
- Schurig, G.G., N. Sriranganathan, and M.J. Corbel, Brucellosis vaccines: past, present and future. Veterinary Microbiology 2002, 90, 479–496. [CrossRef] [PubMed]
- van Straten, M., et al., Brucella abortus S19 vaccine protects dairy cattle against natural infection with Brucella melitensis. Vaccine 2016, 34, 5837–5839. [CrossRef] [PubMed]
- Mahlau, E. Further brucellosis surveys in Tanzania. Bulletin of Epizootic Diseases of Africa. Bulletin des épizooties en Afrique 1967, 15, 373–378. [Google Scholar] [PubMed]
- Mathew, C., et al., First isolation, identification, phenotypic and genotypic characterization of Brucella abortus biovar 3 from dairy cattle in Tanzania. BMC Veterinary Research 2015, 11, 1–9. [CrossRef]
- Assenga, J.A., et al., Epidemiology of Brucella infection in the human, livestock and wildlife interface in the Katavi-Rukwa ecosystem, Tanzania. BMC Veterinary Research 2015, 11, 1–11. [CrossRef]
- Ducrotoy, M., et al., Brucellosis in Sub-Saharan Africa: Current challenges for management, diagnosis and control. Acta Tropica 2017, 165, 179–193. [CrossRef] [PubMed]
- Mengele, I.J., et al., The status and risk factors of brucellosis in smallholder dairy cattle in selected regions of Tanzania. Veterinary Sciences 2023, 10, 155. [CrossRef]
- Makita, K., et al., Herd prevalence of bovine brucellosis and analysis of risk factors in cattle in urban and peri-urban areas of the Kampala economic zone, Uganda. BMC Veterinary Research 2011, 7, 1–8.
- Bodenham, R.F., et al., Prevalence and speciation of brucellosis in febrile patients from a pastoralist community of Tanzania. Scientific Reports 2020, 10, 7081. [CrossRef]
- Njombe, A., et al. Dairy Industry Status in Tanzania. Ministry of Livestock Development and Fisheries. Available online: https://dairyafrica.com/africadairyportal/wp-content/uploads/2020/06/Dairy_Industry_Status_in_Tanzania_2011.pdf (accessed on 10 May 2021). in 7th African Dairy Conference and Exhibition. 2011. Dar Es Salaam, Tanzania, 25-27 May..
- National Bureau of Statistics, National Sample Census of Agriculture 2019-2020. National Report. Ministry of Finance and Planning, United Republic of Tanzania, 2021: p. 1-317.
- Ministry of Finance and Planning (MoFP), Tanzania Mainland Household Budget Survey 2017-2018, Key indicators Report: Poverty Eradication Division. National Bureau of Statistics, United Republic of Tanzania. Poverty Eradication Division. National Bureau of Statistics, United Republic of Tanzania, 2019.
- Mrode, R., et al., Genomic prediction of crossbred dairy cattle in Tanzania: A route to productivity gains in smallholder dairy systems. Journal of Dairy Science 2021, 104, 11779–11789. [CrossRef] [PubMed]
- Shirima, G.M. The epidemiology of brucellosis in animals and humans in Arusha and Manyara regions in Tanzania. PhD Thesis, University of Glasgow, UK, 2005. [Google Scholar]
- R Core Team, R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2021. Available online: https://www.R-project.org/.
- Lumley, T. Analysis of complex survey samples. Journal of Statistical Software 2004, 9, 1–19. [Google Scholar] [CrossRef]
- Shirima, G., B. Lyimo, and N. Kanuya, Re-emergence of Bovine Brucellosis in Smallholder Dairy Farms in Urban Settings of Tanzania. Journal of Applied Life Sciences International 2018, 17, 1–7. [Google Scholar] [CrossRef]
- Mengele, I.J., et al., Diagnostic challenges of brucellosis in humans and livestock in Tanzania: A thematic review. CABI One Health 2023, ohcs20230001. [CrossRef]
- Aliyev, J., et al., Identification and molecular characterization of Brucella abortus and Brucella melitensis isolated from milk in cattle in Azerbaijan. BMC Veterinary Research 2022, 18, 71. [CrossRef]
- Oliveira, M.S., et al., Molecular epidemiology of Brucella abortus isolated from cattle in Brazil, 2009–2013. Acta Tropica 2017, 166, 106–113. [CrossRef] [PubMed]
- Akoko, J.M., et al., Mapping brucellosis risk in Kenya and its implications for control strategies in sub-Saharan Africa. Scientific Reports 2023, 13, 20192. [CrossRef] [PubMed]
- Mitterran, K.N.R., et al., Detection of Brucella abortus and Brucellla melitensis in cattle and sheep from southern Cameroon. Research Square 2020, 2, 1–13. [CrossRef]
- Abnaroodheleh, F., et al., Shedding rate of Brucella spp. in the milk of seropositive and seronegative dairy cattle. Heliyon 2023, 9, 1–8.
- Kolo, F.B., et al., Seroprevalence and characterization of Brucella species in cattle slaughtered at Gauteng abattoirs, South Africa. Veterinary Medicine and Science 2019, 5, 545–555. [CrossRef]
- Morales-Estrada, A.I., et al., Characterization of Brucella species in Mexico by Bruce-Ladder polymerase chain reaction (PCR). African Journal of Microbiology Research 2012, 6, 2793–2796. [CrossRef]
- Ewalt, D.R., et al., Brucella suis biovar 1 in naturally infected cattle: a bacteriological, serological, and histological study. Journal of Veterinary Diagnostic Investigation 1997, 9, 417–420. [CrossRef]
- Fretin, D., et al., Unexpected Brucella suis biovar 2 infection in a dairy cow, Belgium. Emerging Infectious Diseases 2013, 19, 2053. [CrossRef] [PubMed]
- Baek, B., et al., The first detection of Brucella canis in cattle in the Republic of Korea. Zoonoses and Public Health 2012, 59, 77–82. [CrossRef] [PubMed]
- Segwagwe, B.E., et al., Prevalence and risk factors for brucellosis seropositivity in cattle in Nyagatare District, Eastern Province, Rwanda. Journal of the South African Veterinary Association 2018, 89, 1–8. [CrossRef]
- Alamian, S. and M. Dadar, Brucella melitensis infection in dog: a critical issue in the control of brucellosis in ruminant farms. Comparative Immunology, Microbiology and Infectious Diseases 2020, 73, 101554. [CrossRef]
- Mol, J.P., et al., Diagnosis of canine brucellosis: comparison of various serologic tests and PCR. Journal of Veterinary Diagnostic Investigation 2020, 32, 77–86. [CrossRef]
- Shome, R., et al., Management of bovine brucellosis in organized dairy herds through the identification of risk factors: A cross-sectional study from Karnataka, India. Veterinary World 2023, 16.
- Aguiar, D.M.d., et al., Risk factors and seroprevalence of Brucella spp. in cattle from western Amazon, Brazil. J Arquivos do Instituto Biológico, 2022, 74, 301–305. [CrossRef]
- Bagenda, I., et al., Case Study of Impact and Risk Factors of Brucellosis (Brucella abortus) in Beef Cattle. Hasanuddin Journal of Animal Science 2023, 5, 66–83. [CrossRef]
- Chagunda, M.G., et al., Risk, knowledge and preventive measures of smallholder dairy farmers in northern Malawi with regard to zoonotic brucellosis and bovine tuberculosis. Onderstepoort Journal of Veterinary Research 2014, 81, 1–6.
- Getahun, T.K., B. Urge, and G. Mamo, Seroprevalence of bovine brucellosis and associated factors among dairy cows with recent cases of abortion in Ethiopia. Public Health Challenges 2023, 2, 1–8. [CrossRef]
- Cárdenas, L., et al., Risk factors for new bovine brucellosis infections in Colombian herds. BMC Veterinary Research 2019, 15, 81. [CrossRef] [PubMed]
- Shome, R., et al., Management of bovine brucellosis in organized dairy herds through the identification of risk factors: A cross-sectional study from Karnataka, India. Veterinary World 2023, 16, 1122–1130. [CrossRef]
- de Alencar Mota, A.L.A., et al., Large-scale study of herd-level risk factors for bovine brucellosis in Brazil. Acta Tropica 2016, 164, 226–232. [CrossRef] [PubMed]
- Sanz, C., et al., Mass vaccination as a complementary tool in the control of a severe outbreak of bovine brucellosis due to Brucella abortus in Extremadura, Spain. Preventive Veterinary Medicine 2010, 97, 119–125. [CrossRef] [PubMed]
- Lord, V.R., et al., Field study of vaccination of cattle with Brucella abortus strains RB51 and 19 under high and low disease prevalence. American Journal of Veterinary Research 1998, 59, 1016–1020. [CrossRef]
- Keid, L.B., et al., A polymerase chain reaction for detection of Brucella canis in vaginal swabs of naturally infected bitches. Theriogenology 2007, 68, 1260–1270. [CrossRef] [PubMed]
Figure 1.
Map of Tanzania showing study regions (in purple color) with large populations of smallholder dairy cattle and unstudied regions in gray (right). Black dots indicate the locations of cattle sampled. Inset shows the location of Tanzania in Africa.
Figure 1.
Map of Tanzania showing study regions (in purple color) with large populations of smallholder dairy cattle and unstudied regions in gray (right). Black dots indicate the locations of cattle sampled. Inset shows the location of Tanzania in Africa.
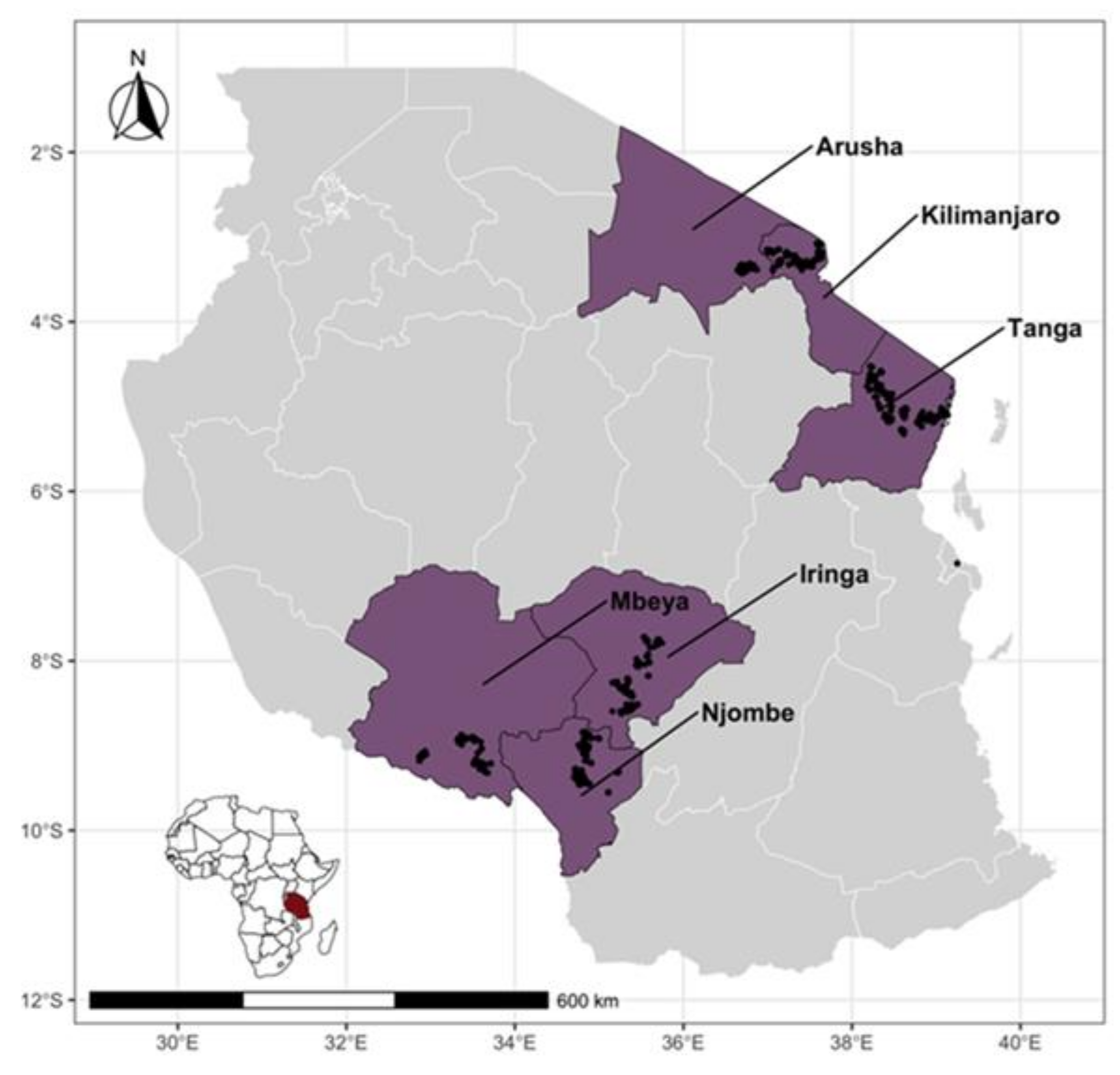
Figure 2.
Choropleth map showing the regional molecular prevalence (insets) and the detailed molecular prevalence by local authority sampled in each region.
Figure 2.
Choropleth map showing the regional molecular prevalence (insets) and the detailed molecular prevalence by local authority sampled in each region.
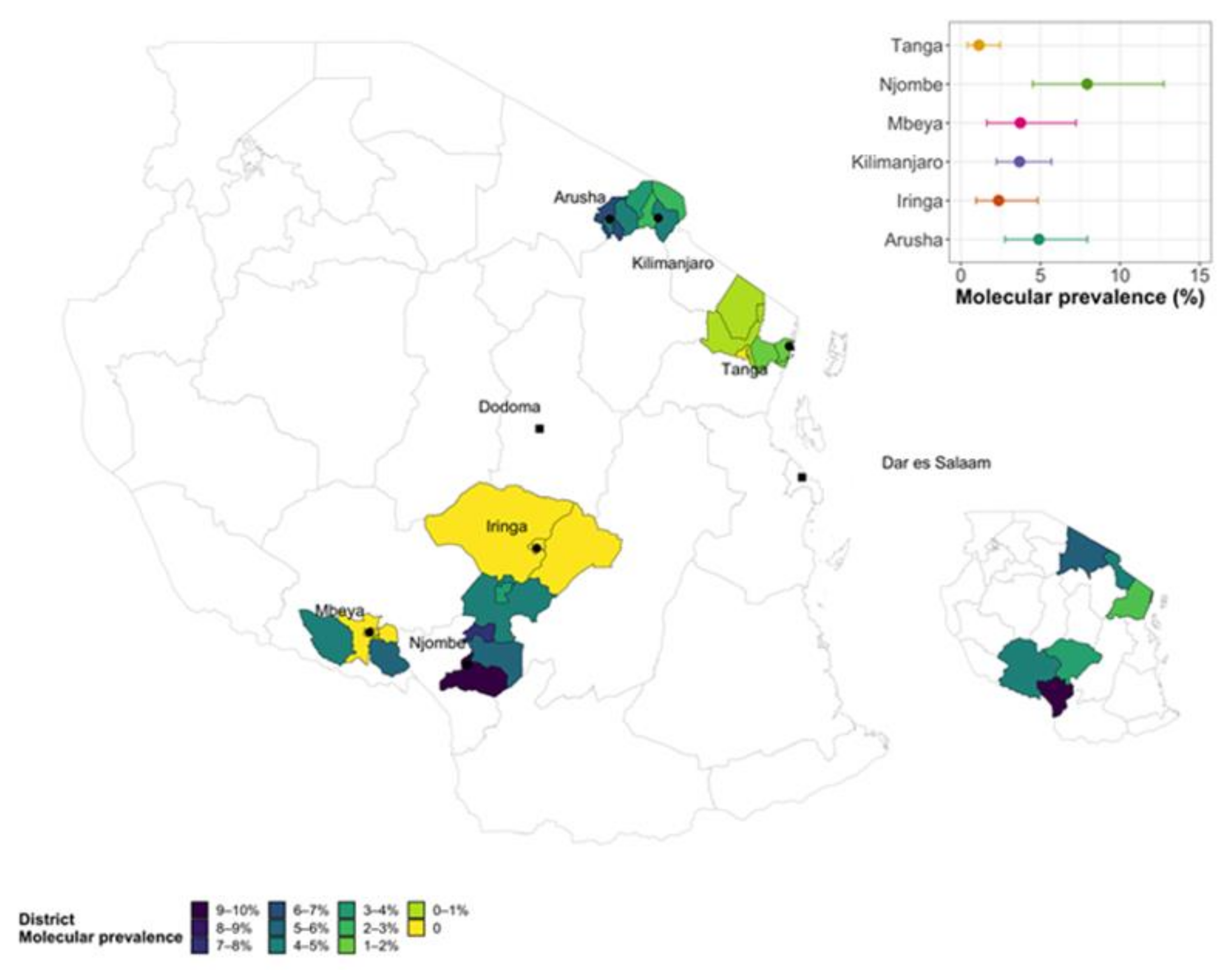

Table 1.
Oligonucleotide primers and probes used to perform qPCR assays.
| Target | Targeted gene | Sequences of primers and probes (5’ -3’) | Fluorophore/ quencher | Reference |
|---|---|---|---|---|
| Genus Brucella | IS711 | Probe: AAG CCA ACA CCC GGC Forward: GGC CTA CCG CTG CGA AT Reverse: TTG CGG ACA GTC ACC ATA ATG |
FAM/-MGBNFQ | Matero, 2011 |
| B. melitensis | IS711 downstream of BMEI1162 | Probe CAGGAGTGTTTCGGCTCAGAATAATCCACA Forward AACAAGCGGCACCCCTAAAA Reverse CATGCGCTATGATCTGGTTACG |
Texas Red/BHQ2 | Probert. 2004 |
| B. abortus | IS711 downstream of alkB | Probe: CGCTCATGCTCGCCAGACTTCAATG Forward: GCGGCTTTTCTATCACGGTATTC Reverse: CATGCGCTATGATCTGGTTACG |
JOE/BHQ1 |
Table 2.
Regional distribution of Brucella genus positive blood and swab DNA samples from individual cattle in Tanzania.
Table 2.
Regional distribution of Brucella genus positive blood and swab DNA samples from individual cattle in Tanzania.
| Region | Total Animal Sampled | Number of positive_blood samples (%) | Number of positive_swab samples (%) |
|---|---|---|---|
| Arusha | 318 | 5/318 (1.6%) | 10/294 (3.4%) |
| Tanga | 524 | 6/524 (1.0%) | 1/412 (0.2%) |
| Kilimanjaro | 521 | 11/519 (2.1%) | 8/513 (1.6%) |
| Iringa | 281 | 7/281 (2.5%) | 1/273 (0.4%) |
| Njombe | 187 | 1/186 (1.1%) | 14/186 (7.5%) |
| Mbeya | 218 | 5/218 (2.3%) | 3/215 (1.4%) |
| Total | 2049 | 35/2046 (1.7%) | 37/1893 (2.0%) |
Table 3.
The combined molecular (PCR) prevalence based on Brucella genus positive results.
| Region | Negative | Positive | Total | PCR Prevalence % | 95% CI | Dairy cattle Population |
|---|---|---|---|---|---|---|
| Arusha | 303 | 15 | 318 | 4.7 | 2.7-7.7 | 78,637 |
| Tanga | 517 | 7 | 524 | 1.3 | 0.5-2.7 | 41,639 |
| Kilimanjaro | 500 | 19 | 519 | 3.7 | 2.2-5.7 | 161,984 |
| Iringa | 273 | 8 | 281 | 2.8 | 1.2-5.5 | 7,081 |
| Njombe | 171 | 15 | 186 | 8.1 | 4.6-13.0 | 7,177 |
| Mbeya | 210 | 8 | 218 | 3.8 | 1.7-7.4 | 72,724 |
| Total | 1974 | 72 | 2046 | 3.5 | 2.8-4.4 | 369,242 |
Table 4.
Real-Time Polymerase chain reaction (qPCR) results for Brucella species identified from genus-positive swab and blood samples.
Table 4.
Real-Time Polymerase chain reaction (qPCR) results for Brucella species identified from genus-positive swab and blood samples.
| Region | Sample | B. abortus | B. melitensis | Mixed | Unidentified |
|---|---|---|---|---|---|
| Arusha | Blood n=5 | 0 | 3 | 2 | 0 |
| Swabs n=10 | 0 | 9 | 1 | 0 | |
| Kilimanjaro | Blood n=11 | 1 | 9 | 0 | 1 |
| Swabs n=8 | 0 | 6 | 2 | 0 | |
| Tanga | Blood n=6 | 0 | 2 | 3 | 1 |
| Swabs n=1 | 0 | 1 | 0 | 0 | |
| Njombe | Blood n=1 | 0 | 1 | 0 | 0 |
| Swabs n=14 | 0 | 11 | 2 | 1 | |
| Iringa | Blood n=7 | 0 | 3 | 2 | 2 |
| Swabs n=1 | 0 | 1 | 0 | 0 | |
| Mbeya | Blood n=5 | 1 | 1 | 3 | 0 |
| Swabs n=3 | 0 | 1 | 2 | 0 | |
| Total | Blood n=35 | 2 | 19 | 10 | 4 |
| Swabs n=37 | 0 | 29 | 7 | 1 |
Mixed=PCR-positive for both B. abortus and B. melitensis.
Table 5.
Univariable analysis of combined blood and vaginal swab PCR positive dairy animals.
| Variable | Level | Negative | Positive | OR | 95% Confid. Interval | P_value | |
|---|---|---|---|---|---|---|---|
| Farmer’s gender | female | 541 | 26 | ref | |||
| male | 643 | 31 | 1.003 | 0.59 | 1.71 | 1 | |
| Herd size | 1-2animals | 589 | 31 | ref | |||
| 3-4animals >4animals |
429 163 |
22 4 |
0.97 0.47 |
0.53 0.12 |
1.77 1.35 |
0.341 0.353 |
|
| Farmer’s experience | ≤5years | 105 | 6 | ref | |||
| in dairy farming | >5years | 1079 | 51 | 0.83 | 0.35 | 1.97 | 0.63 |
| Farmer’s education | basic | 969 | 49 | ref | |||
| secondary_plus | 215 | 8 | 0.74 | 0.34 | 1.57 | 0.59 | |
| Farm or neighbor | have_goat | 795 | 38 | ref | |||
| keep goat | no_goat | 387 | 19 | 1.03 | 0.58 | 1.80 | 1 |
| Farm or neighbor | no_dog | 773 | 36 | ref | |||
| keep dog | have_dog | 411 | 21 | 1.09 | 0.63 | 1.90 | 0.78 |
| Farm or neighbor | have_sheep | 271 | 12 | ref | |||
| keep sheep Farm or neighbor keep pig |
no_sheep no_pig have_pig |
911 891 293 |
45 47 10 |
1.11 ref 0.65 |
0.58 0.32 |
2.14 1.29 |
0.87 0.26 |
| Keeping bull | no_bull | 909 | 50 | ref | |||
| have_bull | 275 | 7 | 0.46 | 0.21 | 1.03 | 0.05 | |
| Body condition score | good | 630 | 29 | ref | |||
| poor | 554 | 28 | 1.09 | 0.64 | 1.86 | 0.78 | |
| Introduction of new | no | 1070 | 49 | ref | |||
| animal | yes | 114 | 8 | 1.53 | 0.71 | 3.31 | 0.26 |
| Placenta management | correct_dispose (bury/burn) |
956 | 44 | ref | |||
| Incorrect_dispose | 228 | 13 | 1.24 | 0.65 | 2.34 | 0.49 | |
| Farm’s distance | <100 | 946 | 41 | ref | |||
| ≥100 | 238 | 16 | 1.55 | 0.85 | 2.81 | 0.17 | |
| Cattle breed | SHZxAyrshire | 264 | 10 | ref | |||
| SHZxFriesian | 850 | 40 | 1.24 | 0.6 | 2.8 | 0.550 | |
| SHZxJersey+SHZ | 70 | 7 | 2.62 | 0.82 | 7.95 | 0.058 | |
| Animal sex | female | 1147 | 57 | ref | |||
| male | 37 | 0 | 0 | 0 | NaN | 0.41 | |
| Animal age | 1-4years | 501 | 21 | ref | |||
| 5-7years >7years |
488 192 |
27 9 |
1.32 1.12 |
0.71 0.44 |
2.49 2.60 |
0.351 0.784 |
|
| Region | Arusha Kilimanjaro Mbeya Njombe |
303 500 210 172 |
15 19 8 15 |
ref 0.77 1.77 1.80 |
0.38 0.32 0.85 |
1.53 1.85 3.71 |
0.47 0.66 0.17 |
| History of abortion (Animal) |
no yes |
1049 32 |
53 4 |
ref 2.51 |
0.86 |
7.38 |
0.056 |
| History of abortion | no | 1110 | 52 | ref | |||
| (Farm) Feeding management |
yes pasture intensive |
71 166 1915 |
5 6 51 |
1.51 ref 1.39 |
0.58 0.59 |
3.89 3.29 |
0.39 0.452 |
Table 6.
Final multivariable logistic regression model for Brucella PCR-positivity for dairy cattle in Tanzania.
Table 6.
Final multivariable logistic regression model for Brucella PCR-positivity for dairy cattle in Tanzania.
| Variable | Level | OR | Lower | Upper | P-value |
|---|---|---|---|---|---|
| History of abortion (Animal) | no | ref | |||
| yes | 3.17 | 1.05 | 9.62 | 0.04 | |
| Farm or neighbour | have dog | ref | |||
| keep Dog | no dog | 2.68 | 1.30 | 5.55 | 0.008 |
| Introduction of | no | ref | |||
| new animal | yes | 1.87 | 0.82 | 4.24 | 0.14 |
| Farm or neighbor | have_pig | ref | |||
| keep pig | no_pig | 2.19 | 1.21 | 3.98 | 0.01 |
| Keeping bull | no | ref | |||
| yes | 0.39 | 0.16 | 0.96 | 0.04 | |
| Region | Arusha | ref | |||
| Kilimanjaro | 1.20 | 0.56 | 2.56 | 0.6 | |
| Mbeya Njombe |
1.39 2.72 |
0.50 1.21 |
3.86 6.10 |
0.5 0.016 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



