
Submitted:
24 June 2024
Posted:
24 June 2024
You are already at the latest version
Abstract
Polycyclic polyprenylated acylphloroglucinols (PPAPs) comprise a large group of compounds of mostly plant origin. The best-known compound is hyperforin from St. John's wort with its antidepressant, antitumor and antimicrobial properties. Chemical synthesis of PPAP variants allows the generation of compounds with improved activity and compatibility. Here we studied two synthetic PPAP-derivatives, PPAP 23 and PPAP 53, as to their antimicrobial activity. While PPAP 23 is insoluble in water, PPAP 53, the sodium salt, is water-soluble. In vitro, both compounds exhibited good and comparable activity against methicillin-resistant Staphylococcus aureus (MRSA) and vancomycin-resistant Enterococcus faecium. Both compounds had no adverse effects on Galleria mellonella wax moth larvae, even at high concentrations. However, they were unable to protect the larvae from infection with S. aureus because components of the larval coelom neutralized the antimicrobial activity; a similar neutralizing effect was also seen with serum albumin. In a septic arthritis model triggered by S. aureus PPAP23 decreased the formation of abscesses and bacterial load in kidneys, and in a mouse skin abscess model topical treatment with PPAP 53 resulted in an approximately two-fold reduction in S. aureus counts. PPAP 23 and PPAP 53 also showed high growth inhibiting activity against anaerobic Gram-positive pathogenic gut bacteria such as Clostridium perfringens and Clostridium difficile. Based on these results, we envision potential application of PPAP 23 and PPAP 53 in the topical treatment of wound infections with Gram+ pathogens, and their oral use in intestinal infections with C. difficile.
Keywords:
albumin
; antimicrobial activity
; larvae
; mouse septic arthroses
; PPAP
; gastrointestinal microbes
1. Introduction
Hyperforin is one of the main ingredients of and most likely the one responsible for the antidepressant, antitumoral and antimicrobial effects of St. John's wort [1,2,3]. Hyperforin belongs to the polycyclic polyprenylated acylphloroglucinols (PPAP) and is rather an unstable compound [4]. However, a hydrogenated hyperforin analogue was more stable and showed antimicrobial activity against various Gram-positive bacteria [5,6]. A breakthrough in PPAP research was achieved with the total synthesis of >100 defined endo-type B PPAP compounds and of non-natural PPAP analogs [7,8,9,10]. Very recently 30 additional entities based on structure-activity relationship against methicillin-resistant Staphylococcus aureus (MRSA) were synthesized [11]. All of these compounds were evaluated for their antibacterial activity and cytotoxicity.
Four of the new compounds, namely, PPAP 14, 19, 22, 23, showed high activity against multiple resistant Gram+ pathogens such as multidrug-resistant Staphylococcus aureus (MRSA) and vancomycin-resistant Enterococcus faecium, and comparatively low toxicity against HL60 cells [12,13]. Despite intensive attempts, it was not possible to isolate PPAP 23-resistant mutants, which is advantageous for a potential application.
All the results indicate that in Gram+ bacteria PPAP 23 interacts with the cytoplasmic membrane via its hydrophobic pocket. It also disrupts the respiratory chain by destroying the Fe-S cluster enzymes and has a mild iron chelating activity, thus interfering with iron metabolism [13]. Accordingly, supplementation of exogenous iron attenuated its antimicrobial activities. How this compound performs in animal studies has so far remained elusive.
Here we show that PPAP 23 and the more water-soluble sodium salt PPAP 53 have good antimicrobial properties against Gram+ pathogens, and both PPAP 23 and 53 are very well tolerated in the insect and mouse models. However, they are unable to effectively protect against S. aureus infection. The main reason for this is that the antimicrobial activity of both PPAPs is neutralized by components of the larval coelom and also by mammalian albumin. We see a possible therapeutic application in an albumin-deficient milieu, for example, in the context of certain gastrointestinal (GI) infections, as PPAP 23 has a selective efficacy against intestinal, anaerobic Gram-positive bacterial pathogens.
2. Results
2.1. The PPAP Compounds 23 and 53
Similar to hyperforin, many of the synthesized derivatives are only poorly soluble in water. For example, the well-studied PPAP 22 and 23 were soluble in 100% DMSO, 100% ethanol, 80% methanol, 8.2% cyclodextrin/18% DMSO in PBS, and 0.4% Tween 80 / 18% DMSO in PBS. Since high concentrations of DMSO, methanol, ethanol and β-cyclodextrin are cytotoxic, we selected 0.4% Tween 80 / 18% DMSO in PBS as a suitable solvent. In Figure 1 it is shown that they chemically differ in the R2 position: PPAP 23 has a prenyl residue, while in PPAP 22 the R2 position is occupied by an allyl residue [12]. When tested towards S. aureus USA300, the MIC values of PPAP 22 and PPAP 23 were 2 µg/ml and 1 µg/ml, respectively. When tested towards the human monocytic cell line HL-60, the IC50 values of PPAP 22 and PPAP 23 were 223 µg/ml and 96 µg/ml, respectively [13]. The lower MIC of PPAP 23 was accompanied by a doubled cytotoxicity compared to PPAP 22. For the investigation of the mode of action, we have previously focused on PPAP 23 [13]. To improve the solubility of PPAPs, a sodium salt derivative of PPAP 22 was generated that we named PPAP 53 (Figure 1). In this study, we mainly aimed at investigating whether the water-soluble PPAP 53 had an advantage over the less water-soluble PPAP 23.
2.2. PPAP 23 and PPAP 53 Had No Toxic Effect on Larvae But Could Not Rescue Larvae In An Infection Model
The antibacterial activity of PPAP 23 and PPAP 53 was tested in Galleria mellonella larvae infected with S. aureus USA300. A dosage of 20 mg/kg (20 x MIC) of PPAP 23 and PPAP 53 was tested on larvae. When the PPAPs were injected alone, all the larvae survived, showing that PPAP 23 and 53 are well-endured at this dosage (Figure 2A, B). However, when the larvae were infected with 106 CFU USA300, all larvae died within 3 days. Treatment with either PPAP 23 or PPAP 53 (dosage: 20 mg/kg (50 μM)) one hour after bacterial infection didn’t increase the viability of the infected larvae (Figure 2A,B). As a control we used a comparable dose of vancomycin (20 mg/kg, 13 μM). Vancomycin could completely protect the USA300 infected larvae (Figure 2C). Since PPAP 23 and PPAP 53 exhibited good antimicrobial activity in vitro, we asked ourselves why in the in vivo studies PPAPs were not effective.
2.3. The coelomic Fluid of the Larvae Antagonized the Activity of PPAP 23 and PPAP 53. We Suspected that Coelomic Fluid Neutralized the Effect of PPAPs. So, to Mimic the In Vivo Larval Experiment, 10 µg of PPAP 23 and PPAP 53 Were Added to 100 μl of Sterile Larval Coelomic Fluid and then Incubated with 106 CFU/ml S. Aureus USA300 Overnight. PPAP 23 or PPAP 53 and USA300 Incubated with PBS Served as Positive Controls. As Expected, No Antibacterial Activity of PPAP 23 and PPAP 53 Towards S. Aureus Was Observed in the Presence of the Coelomic Fluid (Figure 3A and B). This Suggests That Components in the Larval Coelomic Fluid Have an Antagonizing Effect on the PPAPs. This Raised The Question Whether Mammalian Serum and Albumin, One of the Main Proteins of Blood Plasma, Could Also Neutralize the Antibacterial Activity of PPAPs
2.4. Bovine serum and Albumin Abrogated The Bactericidal Activity of PPAP 53
The larval coelomic fluid is very similar to the mammalian blood serum (Fredrick & Ravichandran, 2012). As coelomic fluid neutralizes the antimicrobial activity of PPAP 53, we were curious to determine whether the mammalian serum and its components (mainly serum albumin, immunoglobulin (IgG) and fibrinogen (Fg)) also showed a similar effect. For this, we determined the MIC of PPAP 53 on USA300 in the presence of fetal bovine serum (FBS). We could see a 32- fold reduction in the antibacterial activity of PPAP 53 in the presence of 25% FBS (Table 1). Bovine serum albumin (BSA) (10 mg/ml) also significantly decreased the in vitro antimicrobial activity of PPAP 53, while IgG and Fg (at concentrations up to 25 mg/ml) had no effect (Table 1). Hence, albumin is the main protein in FBS that abrogates the bactericidal activity of PPAP 53. The growth kinetic studies of USA300 in the presence of PPAPs and/or 1% BSA revealed that the antimicrobial effect of the PPAPs was neutralized by 1% BSA (Figure 4A,B). The anti-bacterial activity of vancomycin remained unaffected in the presence of 1% BSA (Figure 4C).
2.5. PPAP 53 Reduced Growth of S. Aureus USA300 in Subcutaneous Abscesses
Since BSA inhibits the activity of PPAP53, we also analyzed whether topical application of PPAP53 could inhibit bacterial growth in skin abscesses. To this end, we used a mouse skin abscess model that closely resembles staphylococcal skin infections in humans [14]. Since USA300 is known to induce severe skin infections, we induced skin abscesses by subcutaneous injection of USA300 into the flanks of mice. Interestingly, topical application of PPAP53 on filter paper discs two hours and 24h after infection significantly reduced bacterial loads in skin abscesses (Figure 5), indicating that PPAP53 can penetrate skin tissues and reach subcutaneous abscesses to limit bacterial growth.
2.6. PPAP 23 Showed A Beneficial But Not Fully Protective Effect on S. Aureus Septic Arthritis Mouse Model
Since we obtained different results in the insect and mammalian model, we checked whether PPAP 23 had a protective effect in the mouse model of septic arthritis caused by S. aureus Newman. PPAP 23 was injected intraperitoneally at a dose of 100 μg PPAP 23/mouse twice daily from day 2 of infection with S. aureus Newman. Visible symptoms of septic arthritis started on day 2 post Newman infection and worsened till day 7. We didn’t observe any difference between the clinical severity of septic arthritis in mice treated with PPAP 23 and the control group (mice treated with vehicle (0.5% Tween 80 in PBS) (Figure 6A). During the span of the study, we also didn’t observe any significant difference in the loss of body weight of mice in two groups after infection (Figure 6B). Nevertheless, the abscesses in the kidneys of mice treated with PPAP 23 were significantly less with respect to abscess score and cfu/kidney as compared to that of the mice treated with the vehicle (P < 0.05; Figure 6C). The bacterial load in the kidneys of PPAP 23 treated mice was >25-fold lower than the one treated with the vehicle (P<0.05; Figure 6D). The bacterial load and the abscess score in the kidneys were significantly correlated (r=0.95; P<0.001). This indicates that PPAP 23 treatment had a slight, but favorable effect on the clearance of S. aureus in septic arthritic mice.
2.7. PPAP 23 and PPAP 53 Showed Activity Against Gut, Gram+ Anaerobic Pathogens
We previously showed that PPAP 23 and 53 has good antibacterial activity against aerobic and facultative aerobic Gram+ bacteria[13]. The effect of PPAPs on anaerobic gut microbiota has not been studied so far. Here we found that PPAP 23 and 53 had good bactericidal activity against various Gram+ anaerobic pathogenic gut bacteria such as Clostridium perfringens, Clostridium difficile, and other disease associated species (Table 2); while some commensal anaerobic gut species were unaffected by PPAP 23 and 53.
3. Discussion
The polycyclic polyprenylated acylphloroglucinols (PPAPs) represent a large group of natural products. In particular, the diverse biological activity of these compounds has spurred chemical synthesis and concomitant derivatization. The current database comprises more than 850 structures with different bioactivities [3,15,16,17,18]. The development of a seven-step synthetic approach enabled the total synthesis of a large variety of structurally diverse endo type B PPAPs [7,8]. In previous studies, we mainly focused on the antimicrobial mode of action of PPAP 23. We could show that PPAP 23 mainly targets the bacterial cytoplasmic membrane of Gram+ bacteria, it inactivates Fe-S cluster enzymes and induces a breakdown of the bacterial membrane potential [12,13] . Because of their small size (~400 Da) PPAP 23 and 53 makes them good candidates for drugs.
Here, we focused more on the in vivo activity of PPAP 23 and we included in our study the new water-soluble sodium salt derivative of PPAP 22, named PPAP 53 (Structures in Figure 1). Through its water solubility, we expected an increased antibacterial activity, and indeed, the MIC (µg/ml) against S. aureus USA300 was lowered from 1.0 to 0.5 (Table 1). The first animal model in which we tested the two PPAPs was the wax moth larval model of Galleria mellonella, which is widely used to study the infectivity of pathogenic bacteria or the in vivo efficacy of antibiotics [19]. Both PPAPs were well tolerated by larvae at the concentration used suggesting that the PPAPs do not induce toxic side effects.
However, when the larvae were challenged with USA300, both PPAPs failed to protect the infected larvae from killing by USA300 because of their neutralization by components of the coelomic fluid. Unfortunately, BSA also caused neutralization of our PPAPs, while other common serum proteins such as fibrinogen or immunoglobulins showed no neutralizing effect (Table 1). Despite this not-so-promising perspective, and because the insect system and the mammalian system are significantly different, we investigated the efficacy of PPAPs in two different mouse models: In the mouse skin abscess model S. aureus simulates a typical skin disease which allows the testing of antimicrobial agents by targeted application to mouse skin. First, we showed that topical treatment of the skin with PPAP 53 did not cause local or systemic signs of toxicity, indicating that PPAP 53 was skin tolerable. In the USA300 abscess model, USA300 was injected subcutaneously into the shaved flanks of mice after adsorption to silica nanospheres to hold the bacteria in a specific location in the skin tissue and allow abscess formation. The topical treatment with PPAP 53 caused a twofold reduction in bacterial counts (Figure 5). Since the bacteria were administered subcutaneously but PPAP 53 was applied on the surface of the skin, it can be assumed that PPAP 53 can also penetrate and show activity into deeper layers of the skin. In the septic arthritis mouse model, we investigated the antibacterial activity of PPAP 23. This compound did not lead to a full protection from USA300 infection; however, the formation of kidney abscesses was decreased and also the bacterial load was reduced (Figure 6). In these two mouse models we demonstrated that both PPAPs are well tolerated and significantly reduced bacterial load.
Despite these positive effects in the animal model, one must assume that neutralization by serum albumin undermines the efficacy of the two PPAPs. With a concentration of 35-50 mg/ml in the human blood, serum albumin is the most abundant protein in vertebrates. Albumin is well-known to bind to a broad spectrum of small molecules or compounds mainly via two binding sites in the subdomain IIA (site 1) and subdomain IIIA (site 2) [20,21]. Because of their small size and/or their hydrophobicity, PPAPs belong precisely to the class of compounds that are bound by albumin. Although PPAP 53 is water soluble in contrast to PPAP 23, its activity is also neutralized by albumin, most likely by being bound by albumin.
While we have relatively high albumin levels in most body regions and organs, in the gastrointestinal tract the albumin level is comparatively low with only 0.01-0.24 mg/g wet weight of feces, which is 100 to 1000 times lower than the blood levels [22]. Furthermore, it has been demonstrated that when added to feces, albumin is degraded within a few hours [23]; and the albumin concentration in the gastrointestinal tract of patients with inflammatory bowel disease (IBD) is even lower than in healthy individuals [13,24]. Therefore, it is likely that the antimicrobial activity of PPAPs is not impaired at such a low albumin concentration in the gastrointestinal tract.
With this in mind, we asked to what extent the PPAPs inhibits the growth of anaerobic intestinal bacteria. Intriguingly, some of the most pathogenic anaerobic gastrointestinal (GI) bacteria were susceptible to PPAP 23/53. These include Clostridium difficile causing colitis and Clostridium perfringens causing diarrhea and food poisoning [25,26]. Other GI bacteria sensitive to PPAP 23 were Parabacteroides distasonis, Clostridium ramosum, and Ruminococcus gnavus. Clostridium ramosum is sometimes observed to cause bacteremia [27]. Reports show emerging antimicrobial resistance in Parabacteroides distasonis, an aerotolerant gut anaerobe with pathogenic and probiotic effects on human health [28]. Ruminococcus gnavus is associated with inflammatory bowel disease, particularly Crohn's disease, and the production of inflammatory polysaccharides [29] (Table 2). Our results suggest the potential application of the PPAPs as a nutritional supplement for the treatment of intestinal dysbiosis caused by Clostridium difficile. This possible application of PPAP 23/53 is not so far-fetched if we compare it with the application of hyperforin. Hyperforin also binds with high affinity to serum albumin which affects its anti-inflammatory effect [30,31]. Hyperforin or St. John's wort extract are administered orally. This means that the active compound reaches the intestine and passes the intestinal epithelial cells as an interactive interface. For instance, hyperforin is similar to PPAP 23 in its poor water solubility and high hydrophobicity. The advantage of PPAP 23 over hyperforin is that it has lower cytotoxicity and is photochemically stable; PPAP 23 possessed an IC50 value of only 96 µg/ml, which is 15-fold less toxic than hyperforin [12].
4. Materials and Methods
4.1. Bacterial Strains, Growth Conditions and Antibiotics
S. aureus USA300 was grown in Tryptic Soy Broth (TSB, Difco) at 37°C with continuous shaking. MIC studies were carried out in Mueller Hinton Broth (MHB). Bovine serum albumin (BSA), fetal bovine serum (FBS), IgG from bovine serum, and fibronectin from bovine plasma were purchased from Sigma. For the MIC studies, FBS and BSA were dissolved in the MHB to attain the desired concentrations, followed by sterile filtration. Bovine IgG and fibronectin were dissolved in 0.9% NaCl and sterile filtered before further dilution in MHB. PPAP 22, PPAP 23 and PPAP 53 were synthesized in the research group of Bernd Plietker. For BSA-interaction studies we used FDA-approved ligands known to bind human serum albumin.
4.2. Synthesis of PPAP 53
In a heated 5 ml finger schlenk under air-atmosphere, 1.0 eq. of PPAP 22 or PPAP 23 was dissolved in dry tetrahydrofuran THF (1.0 M) and treated with NaH (1.0 eq.) at 0°C if not stated otherwise. The resulting solution was stirred for one hour at room temperature until complete dissolution as monitored via TLC. The solution was then layered with n-pentane, the precipitation filtered off and washed with n-pentane. Yield: PPAP22 (77 mg, 0.2 mmol, 1.0 eq.) was treated with NaH (60% in mineral oil, 8.0 mg, 0.2 mmol, 1.0 eq.) to yield PPAP 53 as a white powder (49.6 mg, 0.12 mmol, 61 %).
4.3. Bacterial Growth Kinetics
S. aureus grown overnight in TSB were adjusted to OD=0.01 in a 48 well plate and 1X MIC PPAP 53 and/or 1% BSA were added to the culture. The bacterial growth was measured using Varioskan LUX Multimode Microplate Reader. The instrument carries out a kinetic measurement at optical density 578 nm, every 1 h for a total of 24 hr, at 37°C with continuous shaking.
4.4. Antibiotic Susceptibility Testing
The susceptibility of the microbe towards an antimicrobial compound was measured through the minimal inhibitory concentration (MIC) test [32]. The MIC values were determined by the method of microdilution, following the Clinical and Laboratory Standards Institute document M07-A9 guidelines [33]. Here, PPAP 23/53 were serially diluted in MHB to a volume of 50 μl in a 96-well microtiter plate, to which 50 μl of 106 CFU/ml USA300 were added. USA300 without PPAP served as the positive control and MHB alone served as the negative control. The plate was then incubated for 18 hours at 37°C with continuous shaking. The lowest concentration of either PPAPs that inhibited the visible growth of a microorganism was considered the MIC [34] .
4.5. Galleria Mellonella Infection Model
The larvae of Galleria mellonella purchased from Reptilienkosmos (Germany) were grouped based on their weights. S. aureus USA300 cells were grown overnight in TSB, washed with PBS and adjusted to an OD578 of 0.1. Ten μl of bacterial suspension corresponding to 106 CFU of USA300 was injected into the last right proleg of each larva; and 10 μl of either PPAP 23 or PPAP 53 corresponding to a dose of 20 mg/kg was injected into the left proleg of each larva in the treatment group. Untreated larvae and larvae injected with PBS served as control groups. Ten larvae in each group were monitored over 5 days at 37 ºC. The experiment was carried out three times and the survival curves of the larvae were plotted by GraphPad Prism [35].
4.6. Ex Vivo Killing Assay of Galleria Mellonella Larvae
The Galleria mellonella larvae were crushed, coelomic fluid was extracted and sterile filtered. A single larva yielded approximately 100 μl of larval liquid. To mimic the in vivo larva infection studies, the same quantity of PPAP and same CFU of bacterial suspension were used for the ex vivo killing assay. Here, 106 CFU of USA300 and 10 μg of PPAP were co-incubated with 100 μl of coelomic fluid in a 48 well plate. Untreated coelomic fluid and coelomic fluid treated with phosphate buffered saline (PBS) served as controls. The plate was incubated overnight at 37 ºC, with a shaking speed of 20 rpm resembling the movement of larvae. The bacteria in each group were quantified by the drop plate method[36].
4.7. Mouse Model for Hematogenous S. Aureus Arthritis
Female NMRI mice (6- 7 weeks) were used for the experiment. Mice were kept under standard environmental conditions of temperature and light, and were fed laboratory chow and water ad libitum. Pre-made batches of bacteria (Newman strain) were thawed, washed and diluted to desired concentration. Ten NMRI mice (5 mice/group) were inoculated intravenously (i.v.) into the tail vein with 0.2 ml of S. aureus Newman with expected dose (4x106cfu/mouse). PPAP 23 was dissolved in 0.5% Tween 80 in sterile PBS. The mice were injected with 0.2 mL of the PPAP 23 (100µg/mouse) or same volume of vehicle intraperitoneally every 12 hours, starting at day 2 and continuing until day 7 after infection. The mice were regularly weighed and examined for clinical arthritis by observers blinded to the groups. Observers blinded to the treatment groups visually inspected all 4 limbs of each mouse [37]. The arthritis index was constructed by adding the scores from all 4 limbs for each animal as described before. On day 10, the mice were sacrificed and kidneys were aseptically removed and blindly assessed by one investigator (T.J.) for abscesses. A scoring system ranging from 0-3 was used (0- healthy kidneys; 1- 1 to 2 small abscesses on kidneys without structure changes; 2- more than 2 abscesses, but < 75% kidney tissue involved; and 3- large amounts of abscesses with >75% kidney tissue involved)[37]. Thereafter, the kidneys were homogenized and then up to 5 serial dilutions 1:10 in PBS were performed followed by spreading of 100 µl of bacterial suspension onto horse blood agar plates. The plates were cultured for 24 hours at 37°C and the bacteria quantified as CFUs.
4.8. Mouse Model of Subcutaneous Abscess Formation
Six-eight week-old, female C57B1/6J mice were obtained from Envigo. 105 CFUs of S. aureus USA300 were mixed with sterile dextran beads (Cytodex 1, Sigma) and the mixture (0.2 ml) was injected subcutaneously (s.c.) into the shaved flanks of mice as previously described [14]. Two hours and 24 hours after infection, mice were treated either with PBS or with PPAP53 (500 µg/kg) in PBS using filter paper discs, Fixomull stretch and Finn chambers under anesthesia (isofluorane). Mice were euthanized 48 hours after infection. The abscesses were isolated, homogenized, plated on agar plates and cfu was determined.
4.9. IC25 Values of PPAP 23 Against Some of The Anaerobic Bacterial Strains
PPAP 23 was diluted in dimethyl sulfoxide (DMSO) to a concentration of 4.5, 2.2, 0.4 and 0.2 mM (≤100x) in a v-bottom 96-well plate (Greiner Bio-One) and stored at -20°C. Modified Gifu Anaerobic Medium broth (mGAM) (produced by Nissui Pharmaceuticals) was used for preparing drug plates (u-bottom plates, Nunclon delta surface, Thermo Fisher), due to robust growth of our selected species of the gut microbiome and enteropathogens. The drug plates contained 50µl/well of 2x concentrated PPAP 23, were sealed with Aluminum Foil Lids and stored at -20°C max. 4 weeks until use. Consumables and medium were pre-reduced at least 2 days before inoculation in an anaerobic chamber (Coy Laboratory Products Inc., 2% H2, 12% CO2, rest N2). Bacteria were grown twice overnight in mGAM under anaerobic conditions, drug plates were thawed and brought in the anaerobic chamber overnight. The second overnight culture was diluted in mGAM to an OD of 0.02 and 50µl was added to all wells of the drug plate to reach a total volume of 100µl /well containing 1%DMSO, bacteria with an OD of 0.01 and 45, 22, 4 and 2µM PPAP23, respectively. In the case of Bilophila wadsworthia, mGAM was supplemented with 60mM sodium formate and 10 mM taurine. Plates were sealed with breathable membranes and incubated at 37°C. We measured the OD at 578nm after a 60sec shaking step for 20h by using a microplate spectrophotometer (Epoch2, BioTek, Gen5 software, version 3.05) and an automated microplate stacker (BioStack 4, BioTek). Three biological replicates were tested and growth curves were analyzed according to [38] with a cutoff of 25% (drug concentration inhibiting 25% of bacterial growth).
5. Conclusion
Here we show that PPAPs 23 and 53 have the potential to be used in the future in the treatment of topical and gastrointestinal infections and the decolonizing unwanted Gram+ gut pathogens. The selective activity of the PPAPs will be an advantage over other conventional antibiotics. However, we still need to know more about their antidepressant and anxiolytic properties.
Author Contributions
FG: AVA and MM conceived the idea and designed the study. AVA, MM, HW and MP performed most of the experiments. BP, FK and FI Synthesized the PPAPs. TJ and Mam performed mice hematogenous arthritis studies. DK and AP studied subcutaneous abscess formation in mice model. LM and AG tested the activity of PPAP against anaerobic bacterial strains. AVA, MM analyzed the data. AVA, TJ, FG wrote the manuscript.
Acknowledgments
This work was supported by grants from the Ministry for Science, Research and the Arts of Baden-Wuerttemberg (MWK) “AntibioPPAP” and the Deutsche Forschungsgemeinschaft GO 371/18-1. The funders had no role in the study design, data collection and interpretation, or the decision to submit work for publication. We acknowledge the support by Open Access Publishing Fund of University of Tübingen.
References
- Barnes, J.; Anderson, L.A.; Phillipson, J.D. St John’s Wort (Hypericum Perforatum L.): A Review of Its Chemistry, Pharmacology and Clinical Properties. J Pharm Pharmacol 2001, 53, 583–600. [Google Scholar] [CrossRef]
- Chatterjee, S.S.; Bhattacharya, S.K.; Wonnemann, M.; Singer, A.; Müller, W.E. Hyperforin as a Possible Antidepressant Component of Hypericum Extracts. Life Sci 1998, 63, 499–510. [Google Scholar] [CrossRef]
- Ciochina, R.; Grossman, R.B. Polycyclic Polyprenylated Acylphloroglucinols. Chem Rev 2006, 106, 3963–3986. [Google Scholar] [CrossRef]
- Bystrov, N.S.; Dobrynin, V.N.; Kolosov, M.N.; Chernov, B.K.; Chervin, I.I. Structure of the Antibiotic Hyperforin. Dokl Akad Nauk SSSR 1976, 226, 88–90. [Google Scholar]
- Beerhues, L. Hyperforin. Phytochemistry 2006, 67, 2201–2207. [Google Scholar] [CrossRef]
- Schiavone, B.I.P.; Rosato, A.; Marilena, M.; Gibbons, S.; Bombardelli, E.; Verotta, L.; Franchini, C.; Corbo, F. Biological Evaluation of Hyperforin and Its Hydrogenated Analogue on Bacterial Growth and Biofilm Production. J Nat Prod 2013, 76, 1819–1823. [Google Scholar] [CrossRef]
- Biber, N.; Möws, K.; Plietker, B. The Total Synthesis of Hyperpapuanone, Hyperibone L, Epi-Clusianone and Oblongifolin A. Nat Chem 2011, 3, 938–942. [Google Scholar] [CrossRef]
- Horeischi, F.; Biber, N.; Plietker, B. The Total Syntheses of Guttiferone A and 6-Epi-Guttiferone A. J Am Chem Soc 2014, 136, 4026–4030. [Google Scholar] [CrossRef]
- Socolsky, C.; Plietker, B. Total Synthesis and Absolute Configuration Assignment of MRSA Active Garcinol and Isogarcinol. Chemistry – A European Journal 2015, 21, 3053–3061. [Google Scholar] [CrossRef] [PubMed]
- Verotta, L.; Appendino, G.; Belloro, E.; Bianchi, F.; Sterner, O.; Lovati, M.; Bombardelli, E. Synthesis and Biological Evaluation of Hyperforin Analogues. Part I. Modification of the Enolized Cyclohexanedione Moiety. J Nat Prod 2002, 65, 433–438. [Google Scholar] [CrossRef]
- Peslalz, P.; Vorbach, A.; Bleisch, A.; Liberini, E.; Kraus, F.; Izzo, F.; Brötz-Oesterhelt, H.; Götz, F.; Plietker, B.; Peslalz, P.; et al. Chemical Predictive Modelling and Natural Product-Based Divergent Synthesis – Design of Type B PPAPs with Nanomolar Activities against MRSA. Chemistry – A European Journal, 2024. [Google Scholar]
- Guttroff, C.; Baykal, A.; Wang, H.; Popella, P.; Kraus, F.; Biber, N.; Krauss, S.; Götz, F.; Plietker, B. Polycyclic Polyprenylated Acylphloroglucinols: An Emerging Class of Non-Peptide-Based MRSA- and VRE-Active Antibiotics. Angew Chem Int Ed Engl 2017, 56, 15852–15856. [Google Scholar] [CrossRef]
- Wang, H.; Kraus, F.; Popella, P.; Baykal, A.; Guttroff, C.; François, P.; Sass, P.; Plietker, B.; Götz, F. The Polycyclic Polyprenylated Acylphloroglucinol Antibiotic PPAP 23 Targets the Membrane and Iron Metabolism in Staphylococcus Aureus. Front Microbiol 2019, 10. [Google Scholar] [CrossRef]
- Wanner, S.; Schade, J.; Keinhörster, D.; Weller, N.; George, S.E.; Kull, L.; Bauer, J.; Grau, T.; Winstel, V.; Stoy, H.; et al. Wall Teichoic Acids Mediate Increased Virulence in Staphylococcus Aureus. Nature Microbiology 2017 2:4 2017, 2, 1–12. [Google Scholar] [CrossRef]
- Bailly, C.; Vergoten, G. Anticancer Properties and Mechanism of Action of Oblongifolin C, Guttiferone K and Related Polyprenylated Acylphloroglucinols. Nat Prod Bioprospect 2021, 11, 629. [Google Scholar] [CrossRef]
- Richard, J.A.; Pouwer, R.H.; Chen, D.Y.K. The Chemistry of the Polycyclic Polyprenylated Acylphloroglucinols. Angewandte Chemie International Edition 2012, 51, 4536–4561. [Google Scholar] [CrossRef]
- Yang, X.W.; Grossman, R.B.; Xu, G. Research Progress of Polycyclic Polyprenylated Acylphloroglucinols. Chem Rev 2018, 118, 3508–3558. [Google Scholar] [CrossRef]
- Grossman, R.B. , Table of PPAPs Available online:. Available online: https://organicchemistrydata.org/grossman/ppap/allPPAPs.html (accessed on 11 June 2024).
- Tsai, C.J.Y.; Loh, J.M.S.; Proft, T. Galleria Mellonella Infection Models for the Study of Bacterial Diseases and for Antimicrobial Drug Testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef]
- Obradović, D.; Radan, M.; Đikić, T.; Nikolić, M.P.; Oljačić, S.; Nikolić, K. The Evaluation of Drug-Plasma Protein Binding Interaction on Immobilized Human Serum Albumin Stationary Phase, Aided by Different Computational Approaches. J Pharm Biomed Anal 2022, 211. [Google Scholar] [CrossRef]
- Zsila, F.; Bikadi, Z.; Malik, D.; Hari, P.; Pechan, I.; Berces, A.; Hazai, E. Evaluation of Drug-Human Serum Albumin Binding Interactions with Support Vector Machine Aided Online Automated Docking. Bioinformatics 2011, 27, 1806–1813. [Google Scholar] [CrossRef] [PubMed]
- Morrow, R.J.; Lawson, N.; Hussaini, S.H.; Asquith, P. The Usefulness of Faecal Haemoglobin, Albumin and Alpha-1-Antitrypsin in the Detection of Gastrointestinal Bleeding. Ann Clin Biochem 1990, 27 ( Pt 3) Pt 3, 208–212. [Google Scholar] [CrossRef]
- Schmidt, P.N.; Blirup-Jensen, S.; Svendsen, P.J.; Wandall, J.H. Characterization and Quantification of Plasma Proteins Excreted in Faeces from Healthy Humans. Scand J Clin Lab Invest 1995, 55, 35–45. [Google Scholar] [CrossRef]
- Su, Q.; Li, X.; Mo, W.; Yang, Z. Low Serum Bilirubin, Albumin, and Uric Acid Levels in Patients with Crohn’s Disease. Medicine 2019, 98. [Google Scholar] [CrossRef]
- Abt, M.C.; McKenney, P.T.; Pamer, E.G. Clostridium Difficile Colitis: Pathogenesis and Host Defence. Nature Reviews Microbiology 2016 14:10 2016, 14, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Eichner, M.; Augustin, C.; Fromm, A.; Piontek, A.; Walther, W.; Bücker, R.; Fromm, M.; Krause, G.; Schulzke, J.D.; Günzel, D.; et al. In Colon Epithelia, Clostridium Perfringens Enterotoxin Causes Focal Leaks by Targeting Claudins Which Are Apically Accessible Due to Tight Junction Derangement. J Infect Dis 2017, 217, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Legaria, M.C.; García, S.D.; Tudanca, V.; Barberis, C.; Cipolla, L.; Cornet, L.; Famiglietti, A.M.R.; Stecher, D.; Vay, C.A. Clostridium Ramosum Rapidly Identified by MALDI-TOF MS. A Rare Gram-Variable Agent of Bacteraemia. Access Microbiol 2020, 2. [Google Scholar] [CrossRef]
- Ezeji, J.C.; Sarikonda, D.K.; Hopperton, A.; Erkkila, H.L.; Cohen, D.E.; Martinez, S.P.; Cominelli, F.; Kuwahara, T.; Dichosa, A.E.K.; Good, C.E.; et al. Parabacteroides Distasonis: Intriguing Aerotolerant Gut Anaerobe with Emerging Antimicrobial Resistance and Pathogenic and Probiotic Roles in Human Health. Gut Microbes 2021, 13. [Google Scholar] [CrossRef]
- Henke, M.T.; Kenny, D.J.; Cassilly, C.D.; Vlamakis, H.; Xavier, R.J.; Clardy, J. Ruminococcus Gnavus, a Member of the Human Gut Microbiome Associated with Crohn’s Disease, Produces an Inflammatory Polysaccharide. Proc Natl Acad Sci U S A 2019, 116, 12672–12677. [Google Scholar] [CrossRef]
- Traeger, A.; Voelker, S.; Shkodra-Pula, B.; Kretzer, C.; Schubert, S.; Gottschaldt, M.; Schubert, U.S.; Werz, O. Improved Bioactivity of the Natural Product 5-Lipoxygenase Inhibitor Hyperforin by Encapsulation into Polymeric Nanoparticles. Mol Pharm 2020, 17, 810–816. [Google Scholar] [CrossRef]
- Füller, J.; Kellner, T.; Gaid, M.; Beerhues, L.; Müller-Goymann, C.C. Stabilization of Hyperforin Dicyclohexylammonium Salt with Dissolved Albumin and Albumin Nanoparticles for Studying Hyperforin Effects on 2D Cultivation of Keratinocytes in Vitro. Eur J Pharm Biopharm 2018, 126, 115–122. [Google Scholar] [CrossRef]
- Gemmell, C.G.; Edwards, D.I.; Fraise, A.P.; Gould, F.K.; Ridgway, G.L.; Warren, R.E. Guidelines for the Prophylaxis and Treatment of Methicillin-Resistant Staphylococcus Aureus (MRSA) Infections in the UK. J Antimicrob Chemother 2006, 57, 589–608. [Google Scholar] [CrossRef]
- Rennie, R.P.; Turnbull, L.; Brosnikoff, C.; Cloke, J. First Comprehensive Evaluation of the M.I.C. Evaluator Device Compared to Etest and CLSI Reference Dilution Methods for Antimicrobial Susceptibility Testing of Clinical Strains of Anaerobes and Other Fastidious Bacterial Species. J Clin Microbiol 2012, 50, 1153–1157. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nature Protocols 2008 3:2 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Popella, P.; Krauss, S.; Ebner, P.; Nega, M.; Deibert, J.; Götz, F. VraH Is the Third Component of the Staphylococcus Aureus VraDEH System Involved in Gallidermin and Daptomycin Resistance and Pathogenicity. Antimicrob Agents Chemother 2016, 60, 2391. [Google Scholar] [CrossRef]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to Optimize the Drop Plate Method for Enumerating Bacteria. J Microbiol Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Ali, A.; Welin, A.; Schwarze, J.C.; Svensson, M.N.D.; Na, M.; Jarneborn, A.; Magnusson, M.; Mohammad, M.; Kwiecinski, J.; Josefsson, E.; et al. CTLA4 Immunoglobulin but Not Anti-Tumor Necrosis Factor Therapy Promotes Staphylococcal Septic Arthritis in Mice. J Infect Dis 2015, 212, 1308–1316. [Google Scholar] [CrossRef]
- Maier, L.; Pruteanu, M.; Kuhn, M.; Zeller, G.; Telzerow, A.; Anderson, E.E.; Brochado, A.R.; Fernandez, K.C.; Dose, H.; Mori, H.; et al. Extensive Impact of Non-Antibiotic Drugs on Human Gut Bacteria. Nature 2018, 555, 623–628. [Google Scholar] [CrossRef]
Figure 1.
Structures of PPAP 23, 22 and 53 used in this work. The PPAPs have similar MIC values (1 to 2 µg/ml) for the multi resistant S. aureus USA300. PPAP 53 is the Na salt of PPAP 22 and is therefore more water soluble and was dissolved in water. PPAP 23 was dissolved in DMSO.
Figure 1.
Structures of PPAP 23, 22 and 53 used in this work. The PPAPs have similar MIC values (1 to 2 µg/ml) for the multi resistant S. aureus USA300. PPAP 53 is the Na salt of PPAP 22 and is therefore more water soluble and was dissolved in water. PPAP 23 was dissolved in DMSO.
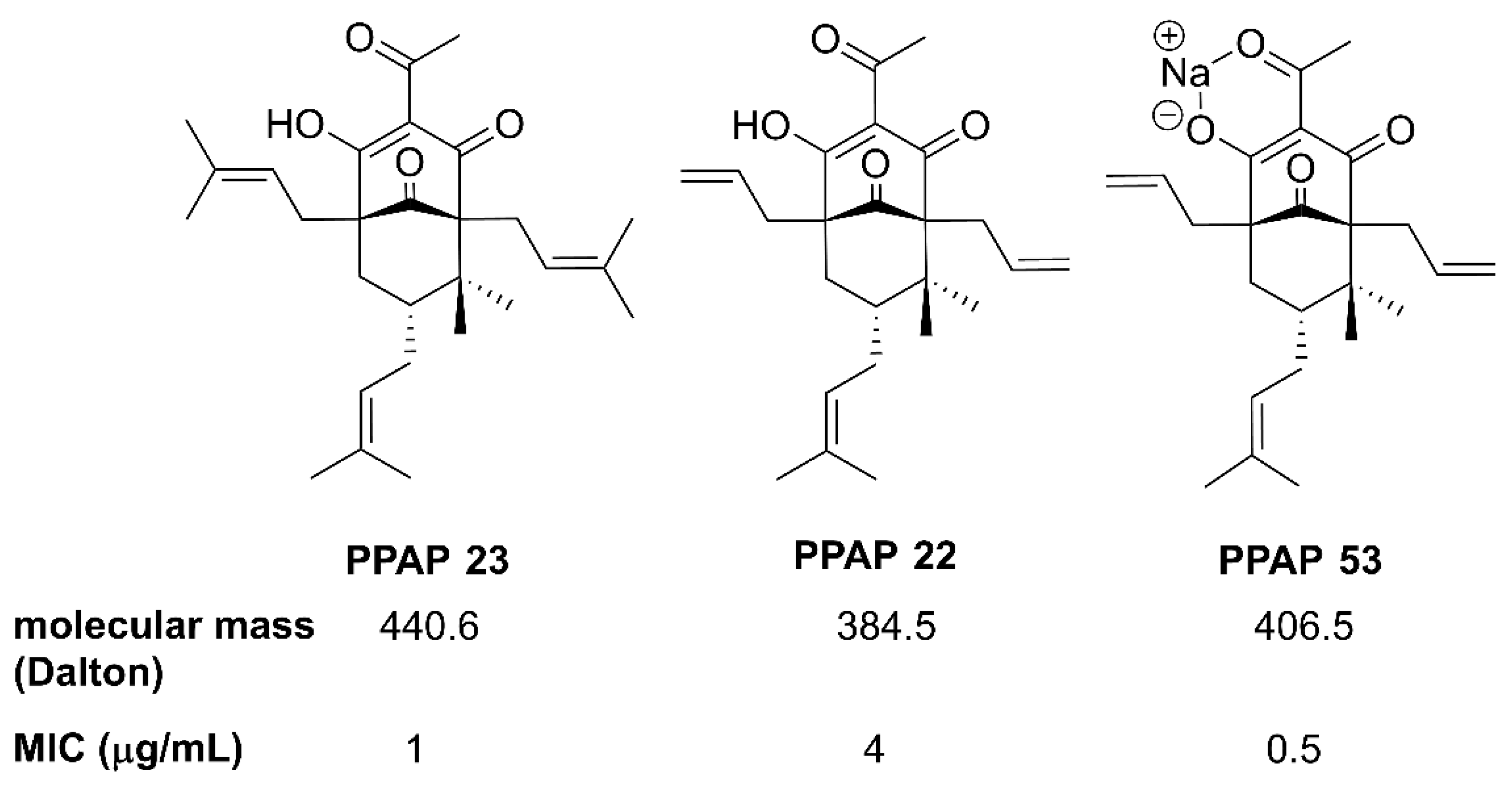
Figure 2.
PPAP 23 and PPAP 53 are non-toxic to larvae, but failed to protect larvae from infection with S. aureus USA300. Ten Galleria mellonella larvae per group, with weight average of 500 mg/larvae, were either non-treated, injected with 106 colony forming units (cfu) S. aureus USA300 (last right proleg). 1 h after administration of bacteria larvae were treated with 20 mg/kg (45 µM) PPAP 23 (A), or 20 mg/kg (50 µM) PPAP 53 (B), or vancomycin (C) at a comparable dose (20 mg/kg, 13 µM). Infected larvae without treatment were normally killed by S. aureus after 3 days. The larvae were maintained at 37 °C and observed for mortality every day over the course of 5 days. A total of three biological replicates are represented in the graph.
Figure 2.
PPAP 23 and PPAP 53 are non-toxic to larvae, but failed to protect larvae from infection with S. aureus USA300. Ten Galleria mellonella larvae per group, with weight average of 500 mg/larvae, were either non-treated, injected with 106 colony forming units (cfu) S. aureus USA300 (last right proleg). 1 h after administration of bacteria larvae were treated with 20 mg/kg (45 µM) PPAP 23 (A), or 20 mg/kg (50 µM) PPAP 53 (B), or vancomycin (C) at a comparable dose (20 mg/kg, 13 µM). Infected larvae without treatment were normally killed by S. aureus after 3 days. The larvae were maintained at 37 °C and observed for mortality every day over the course of 5 days. A total of three biological replicates are represented in the graph.
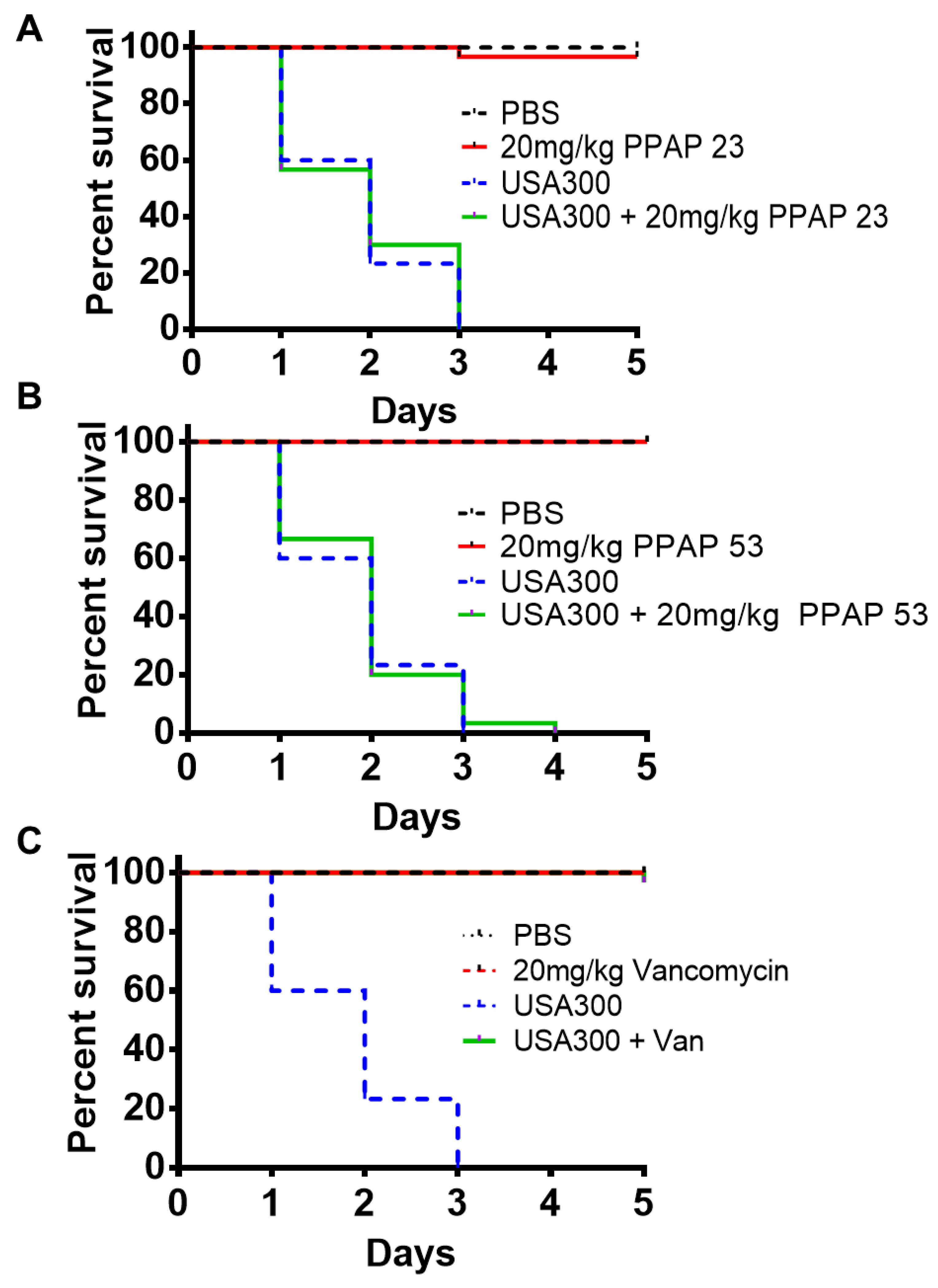
Figure 3.
The bactericidal activity of PPAP 23 and PPAP 53 is reversed by the larval coelomic fluid. To mimic the in vivo larva infection assay, the ex vivo killing assay was adopted. Bacterial inoculum of 105 CFU and 10 μg of (A) PPAP 23 and (B) PPAP53 was added to 100 μl of larva coelomic fluid as the treatment group. Untreated larval liquid and larval liquid treated with PBS were used as controls. The viability of bacteria in each group was determined by the drop plate method. The bactericidal effect of PPAP 23 and PPAP 53 on S. aureus is reversed by coelomic fluid.
Figure 3.
The bactericidal activity of PPAP 23 and PPAP 53 is reversed by the larval coelomic fluid. To mimic the in vivo larva infection assay, the ex vivo killing assay was adopted. Bacterial inoculum of 105 CFU and 10 μg of (A) PPAP 23 and (B) PPAP53 was added to 100 μl of larva coelomic fluid as the treatment group. Untreated larval liquid and larval liquid treated with PBS were used as controls. The viability of bacteria in each group was determined by the drop plate method. The bactericidal effect of PPAP 23 and PPAP 53 on S. aureus is reversed by coelomic fluid.
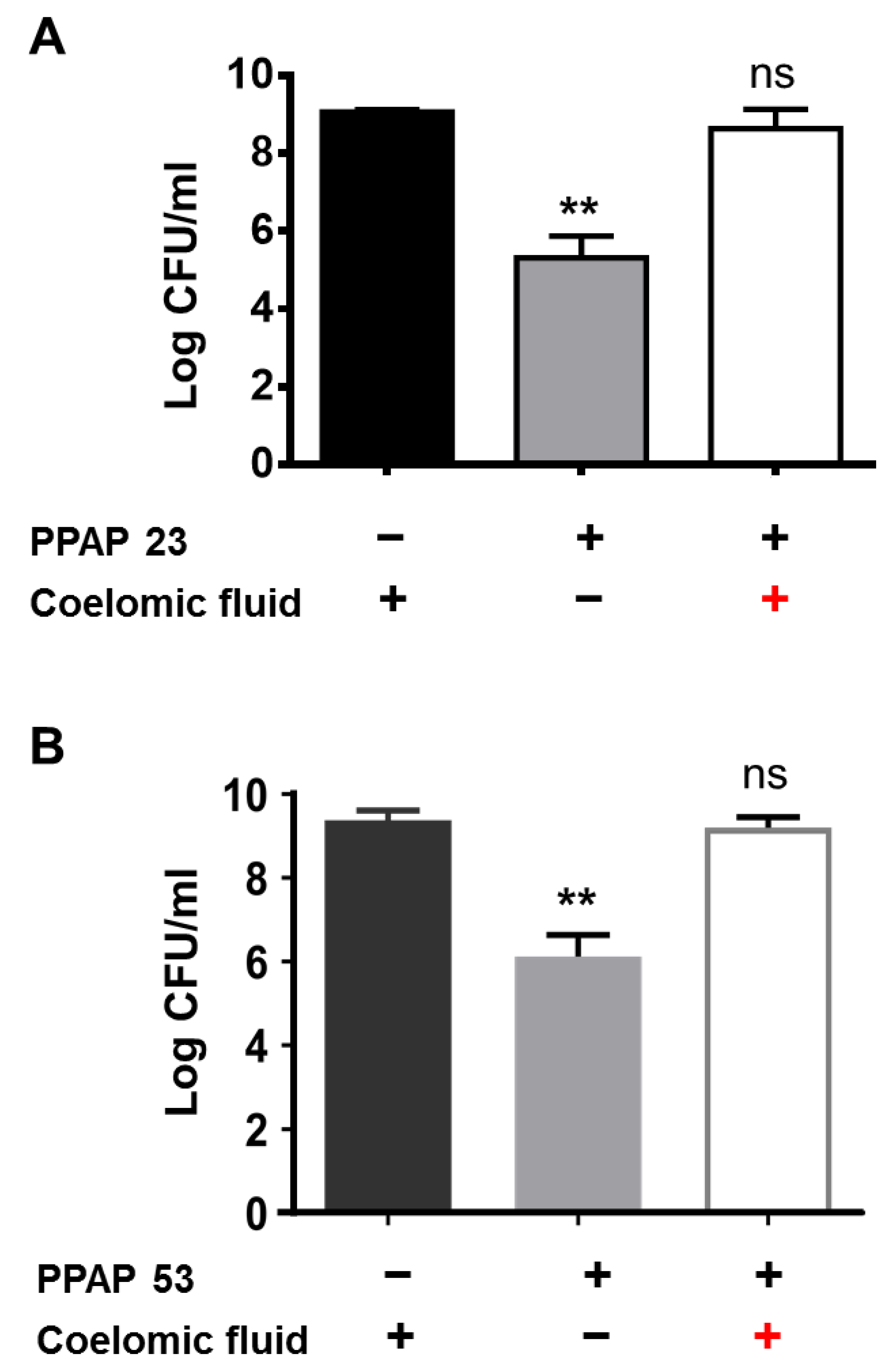
Figure 4.
The bactericidal activity of PPAP 23 and PPAP 53 is reversed 1% Bovine serum albumin (BSA).S. aureus grown overnight in TSB were adjusted to OD= 0.01 in a 48 well plate and 1X MIC PPAPs and vancomycin with or without 1% BSA were added to the culture. The bacterial growth was measured using Varioskan LUX Multimode Microplate Reader for every 2h. The bactericidal effect of (A) PPAP 23 and (B) PPAP 53 (MIC: 0.5-1µg/ml) on S. aureus USA300 is reversed by 1% BSA whereas the effect of (C) vancomycin remained unchanged.
Figure 4.
The bactericidal activity of PPAP 23 and PPAP 53 is reversed 1% Bovine serum albumin (BSA).S. aureus grown overnight in TSB were adjusted to OD= 0.01 in a 48 well plate and 1X MIC PPAPs and vancomycin with or without 1% BSA were added to the culture. The bacterial growth was measured using Varioskan LUX Multimode Microplate Reader for every 2h. The bactericidal effect of (A) PPAP 23 and (B) PPAP 53 (MIC: 0.5-1µg/ml) on S. aureus USA300 is reversed by 1% BSA whereas the effect of (C) vancomycin remained unchanged.
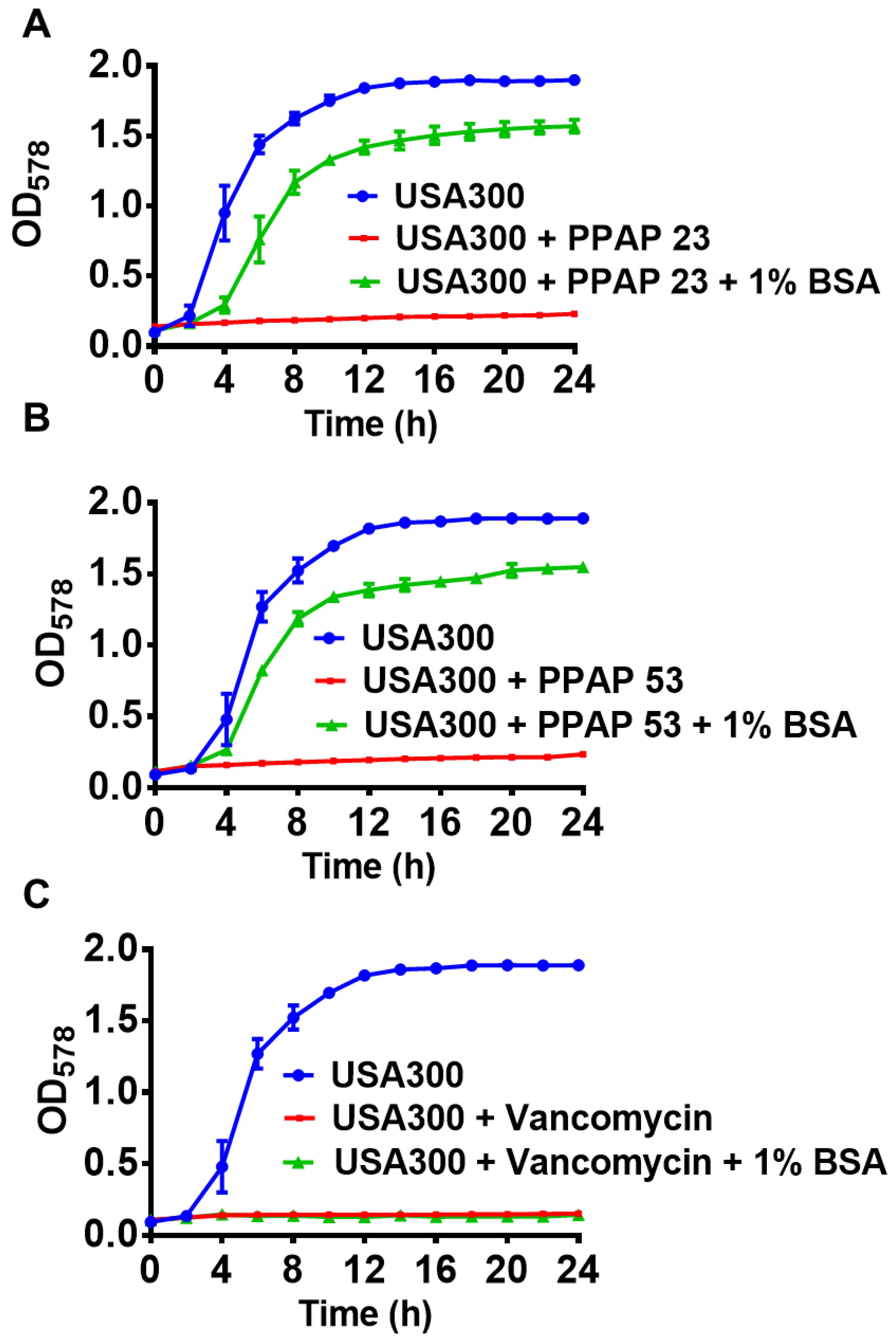
Figure 5.
PPAP 53 treatment significantly reduced bacterial load in skin abscesses. C57BL6J mice s.c. inoculated with S. aureus USA300 strain (105 colony-forming units/mouse) and sterile dextran beads were treated twice, two hours and 24 hours after infection with PPAP 53 dissolved in PBS (500µg/kg) or with PBS (n = 9). After 48 hours the abscesses were excised, homogenized and cfu was determined. Statistical evaluations were performed using the Mann–Whitney U test. Data are mean values ± standard error of the mean. *P < 0.05.
Figure 5.
PPAP 53 treatment significantly reduced bacterial load in skin abscesses. C57BL6J mice s.c. inoculated with S. aureus USA300 strain (105 colony-forming units/mouse) and sterile dextran beads were treated twice, two hours and 24 hours after infection with PPAP 53 dissolved in PBS (500µg/kg) or with PBS (n = 9). After 48 hours the abscesses were excised, homogenized and cfu was determined. Statistical evaluations were performed using the Mann–Whitney U test. Data are mean values ± standard error of the mean. *P < 0.05.
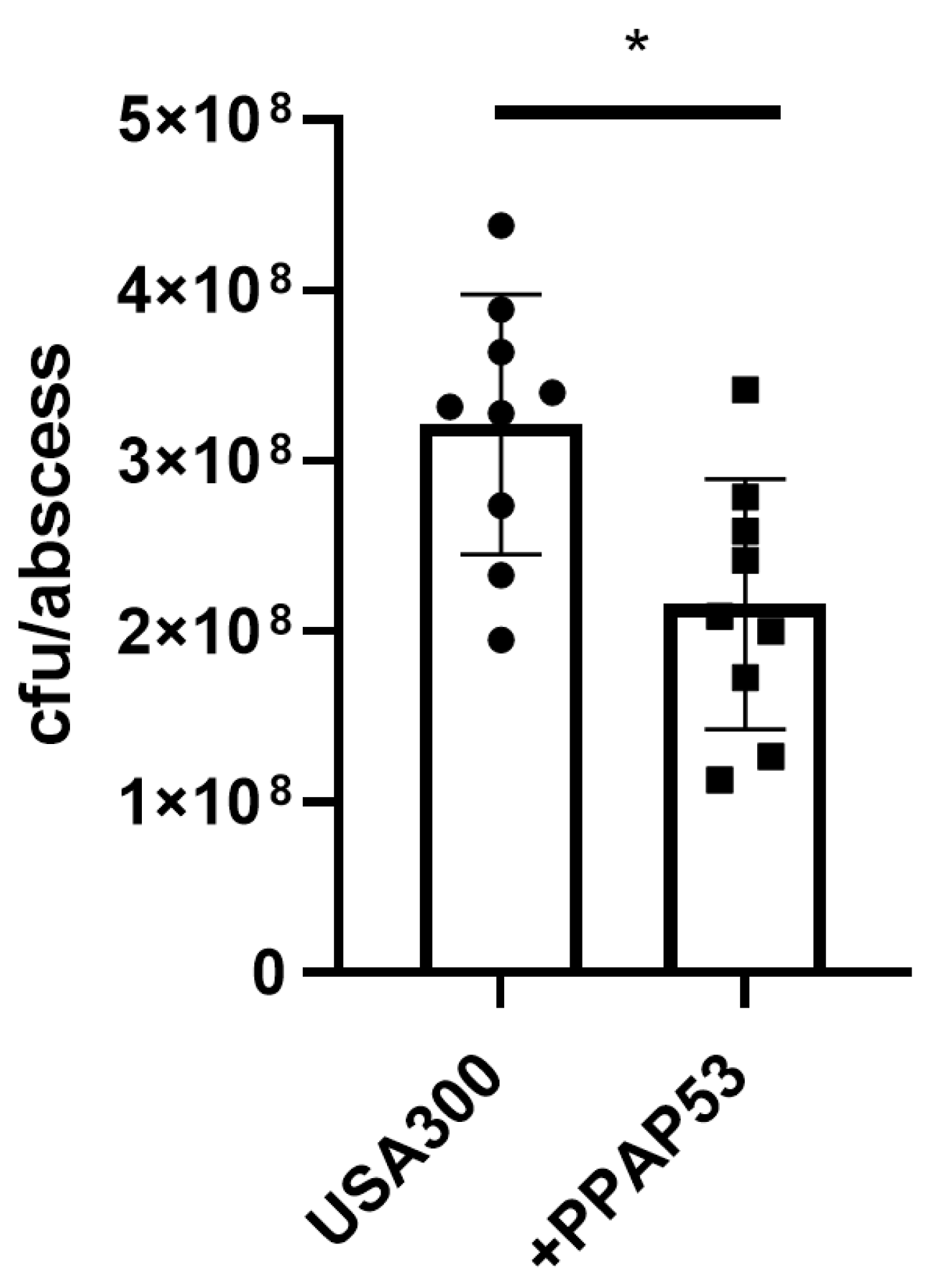
Figure 6.
PPAP 23 treatment significantly reduced the abscess formation and bacterial load in kidneys in mice with S. aureus septic arthritis. NMRI mice inoculated with S. aureus Newman strain (4 × 106 colony-forming units/mouse) were treated with PPAP 23 dissolved in 0.5% tween 80 in PBS (100µg/mouse; n = 5) or same volume of 0.5% tween 80 in PBS (n = 5) twice a day starting on day 2 after inoculation with bacteria and continuing until the animals were euthanized on day 7. The severity of clinical arthritis (A) and the body weight development (B) in the mice were observed for 7 days after infection. Kidney abscess scores (C) and persistence of S. aureus in kidneys (D) from the mice euthanized 7 days after infection. Statistical evaluations were performed using the Mann–Whitney U test. Data are mean values ± standard error of the mean. *P < 0.05.
Figure 6.
PPAP 23 treatment significantly reduced the abscess formation and bacterial load in kidneys in mice with S. aureus septic arthritis. NMRI mice inoculated with S. aureus Newman strain (4 × 106 colony-forming units/mouse) were treated with PPAP 23 dissolved in 0.5% tween 80 in PBS (100µg/mouse; n = 5) or same volume of 0.5% tween 80 in PBS (n = 5) twice a day starting on day 2 after inoculation with bacteria and continuing until the animals were euthanized on day 7. The severity of clinical arthritis (A) and the body weight development (B) in the mice were observed for 7 days after infection. Kidney abscess scores (C) and persistence of S. aureus in kidneys (D) from the mice euthanized 7 days after infection. Statistical evaluations were performed using the Mann–Whitney U test. Data are mean values ± standard error of the mean. *P < 0.05.
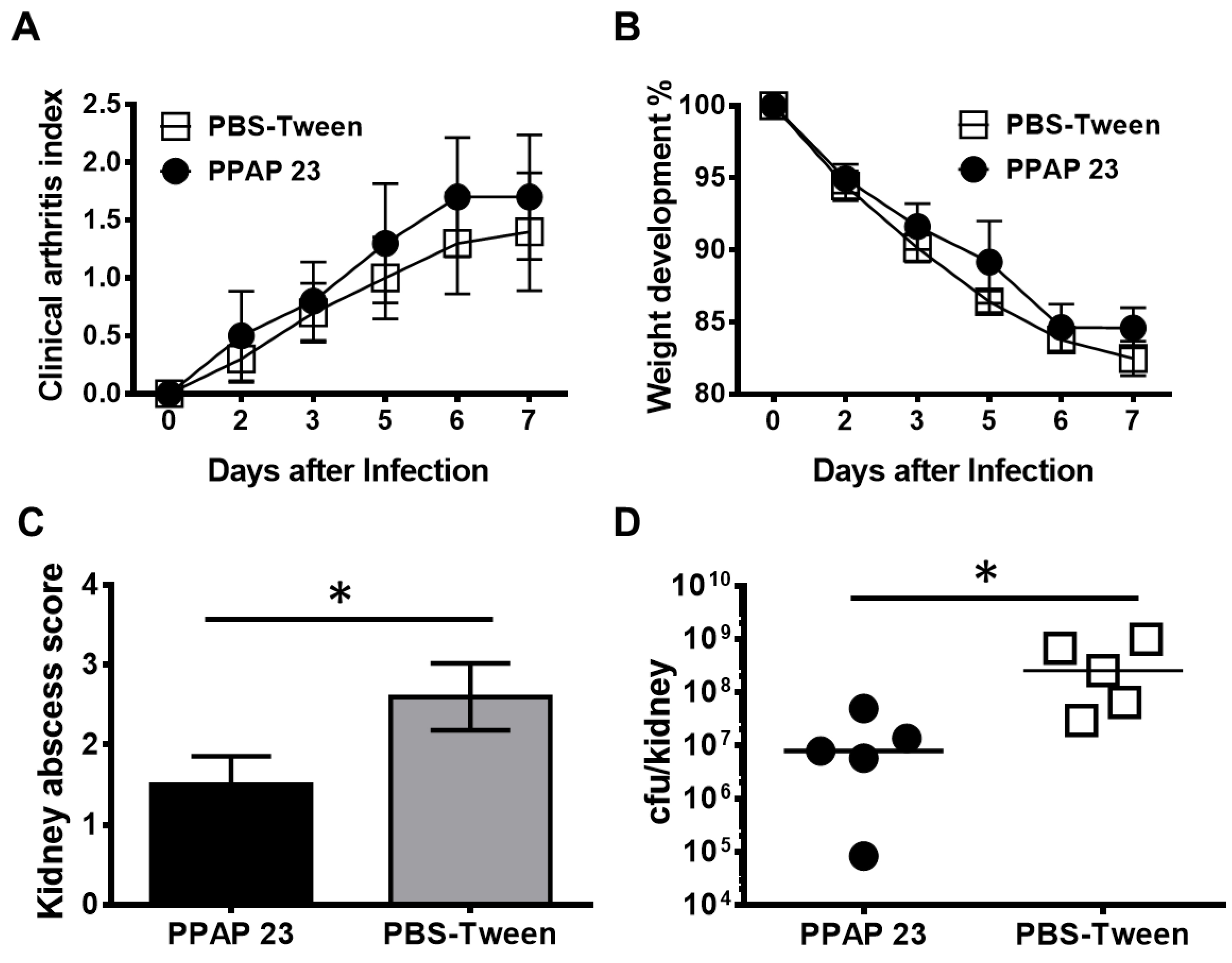

Table 1.
Impact of serum components on PPAP 53 activity against S. aureus USA300.
| Medium | MIC (µg/ml) | ||
|---|---|---|---|
| PPAP 23 | PPAP 53 | Vancomycin | |
| MHB | 1 | 0.5 | 1 |
| MHB + 25% FBS | 32 | 32 | 2 |
| MHB + 0.5% BSA | 4 | 8 | 1 |
| MHB +1% BSA | 8 | 8 | 1 |
| MHB + 2.5% BSA | 16 | 16 | 1 |
| MHB + 5% BSA | 32 | 32 | 2 |
| MHB + 1% IgG | 1 | 0.5 | 1 |
| MHB + 2.5% IgG | 1 | 0.5 | 1 |
| MHB + 1% Fg | 1 | 0.5 | 1 |
| MHB + 2.5% Fg | 1 | 0.5 | 1 |
Table 2.
IC25 values of PPAP 23 and PPAP 53 against some of the anaerobic bacterial strains.
| Strains | PPAP 23 IC25 (µM) | PPAP 53 IC53 (µM) |
|
|---|---|---|---|
| Pathogenic G (+) gut anaerobes |
Clostridium difficile | < 2 | > 80 |
| Clostridium perfringens | < 2 | 5 | |
| Ruminococcus gnavus | < 2 | 5 | |
| Clostridium ramosum | < 2 | 10 | |
| Commensal gut anaerobes (+/-) | Streptococcus salivarius | < 2 | 5 |
| Dorea formicigenerans | 4 | 10 | |
| Streptococcus parasanguinis | 4 | 10 | |
| Roseburia intestinalis | 4 | 10 | |
| Coprococcus comes | 4 | 10 | |
| Collinsella aerofaciens | 4 | 10 | |
| Eubacterium rectale | 4 | 5 | |
| Clostridium bolteae | 22 | 10 | |
| Parabacteroides merdae | 22 | 5 | |
| Clostridium saccharolyticum | 22 | 10 | |
| Fusobacterium nucleatum subsp. Nucleatum | > 45 | 40 | |
| Bacteroides vulgatus | 22 | 5 | |
| Bacteroides uniformis | > 45 | 10 | |
| Bacteroides thetaiotaomicron | > 45 | 20 | |
| Bacteroides fragilis NT | 22 | 10 | |
| Pathogenic G (–) gut anaerobes |
Yersinia pseudotuberculosis | > 22 | 2.5 |
| Yersinia enterocolitica WA-314 | > 45 | > 80 | |
| Vibrio cholerae | > 45 | 80 | |
| Shigella sonnei 53G | > 45 | > 80 | |
| Shigella flexneri | > 45 | > 80 | |
| Salmonella enterica typhimurium LT2 | > 45 | > 80 | |
| Salmonella enterica typhimurium | > 45 | > 80 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



