Submitted:
23 June 2024
Posted:
25 June 2024
You are already at the latest version
Abstract
In a meta-analysis, we examined the behavioral portfolio of invasive brown anoles (Anolis sagrei) and native green anoles (Anolis carolinensis) in urban and non-urban environments. We hypothesized that invasive anoles should display more agonistic and bold signals (head bobbing, dewlap extensions and pushups) than their native range counterparts, and their native competitors. We found that in urban settings, anoles of both species signaled more with dewlap extensions than with head-bobs. The brown anoles displayed significantly higher in non-urban habitats and their native range compared to urban habitats and invasive ranges. The outcome of our analysis suggests that brown anoles have plastic behavioral portfolios, whereas the green anole have relatively balanced preferences for head-bobbing irrespective of the habitat in which the populations were collected. We attribute the success of the brown anole invasion to the flexible strategy in face of higher mate competition, higher predation risks, and less resource competition in the urban environments and invasive ranges. Lastly, we observed publication biases. More studies are conducted with urban and invasive brown anoles and specifically in manipulative mesocosm experimental settings - transplanting populations from native field settings. We show this altered the display rates across all studied signals.
Keywords:
Communication
; Dewlap Extension
; Pushups
; Head-bob
; Territoriality
; Aggression
; Urban Ecology
1. Introduction
Invasive species are considered one of the major drivers of biodiversity loss in the Holocene (Anthropocene) extinction period [1]. Many researchers have tried to categorize the underpinning traits of invasive species and highlight several adaptive traits that can be generalized as the drivers of these populations. Invasive populations tend to be r-selected, exhibit density-independent growth, and grow at a much faster rate than their native-range counterparts [2,3,4]. A generalist diet has been shown to favor invasive animal species, allowing them to forage opportunistically and maximize energetic intake to increase reproductive success [5,6,7,8]. Invasives are commonly described as disturbance-loving species that can adapt to constant turnover in environmental conditions brought on by human activity [8,9,10]. Lastly, the most “successful” invasive species are ones that are characterized by bold, risk-taking, and aggressive behaviors [11,12,13,14]. It is through these behavioral strategies that they manipulate resources and gain competitive advantage over the native species in the invaded communities [15].
The characteristics of invasive species alone cannot explain the establishment of invasive populations. The properties of the invaded community and its environment will influence whether a population exhibiting some of the above-mentioned traits will successfully establish. Two patterns emerge in communities that can be invaded [16]: either open resource niches allow nonnative species to colonize, or the community’s competitors hold evolutionarily inferior resource harvesting strategies compared to the newcomers. In this review we focus on the former, specifically urban communities, where resource niches are continuously vacated by anthropogenic disturbance [17]. Human activity in urbanized zones actively favor species that are bolder and more aggressive [9]. Specifically, humans inherently pose risk to wildlife, directly managing populations and indirectly generating community-wide reverberations. Active hunting of species, perceived as direct threats and mass culling of disease vectors, changes the structure of communities [18,19]. Humans favor human-commensal and mutualistic mesocarnivores (pets) that hunt small vertebrate species [20,21,22,23]. Additionally, our technology, such as motorized vehicles and signal-interfering noise and light pollution, pose adaptive challenges to wildlife [24,25,26]. Coupled with attractive resources that humans discard into their environment, all the above risks generate ecological traps that cause native populations to decline [27]. Lastly, habitat fragmentation hinders movement of wildlife, shrinking populations, driving sub-populations into extirpation [28], and thus frees resources for bold and aggressive species. They are able to monopolize the aforementioned abundant food resources in the form of trash (rubbish) [23,29,30] and byproducts of feeding pets [29,31,32]. These factors, along with others not discussed, indicate that invasive species can utilize urbanization patterns to establish populations and expand their invasive ranges through their behavior. We set out to test this hypothesis by conducting a meta-analysis of published behavioral assays of native North American Green Anoles (Anolis carolinensis, AC) and their invasive counterparts, the Cuban Brown Anoles (Anolis sagrei, AS)— in their native and invasive ranges (Figure 1- distribution of study populations).
Anolis lizards have been used for decades as models for the study of evolutionary patterns, speciation, and the roles their behaviors play in their adaptability to habitats in varied island ecosystems [33,34,35]. Anoles have been shown to evolutionarily radiate based on microhabitat selection, especially based on perch selection [36,37], presence of different predator types [38,39,40], and in response to environmental disturbance [11,41]. The elaborate record on anole evolution [33], including their phylogenetic relationships, historic adaptive radiations [40,42], and their tendency to undergo speciation following disturbance events and isolation of subpopulations (founder effects)[43,44,45], makes them an ideal model organism for studying changes driven by anthropogenic disturbance [33]. Of all their evolutionary divergences, the strongest measures of differentiation rely on ethological (behavioral) variation in mate courting, territoriality and agonistic displays towards conspecifics. While the ability of female anoles across many species to clone themselves, parthenogenesis, allows rapid population establishment following natural catastrophes [46,47,48,49], their evolutionary radiation into various habitats depends more on their polygynous social structure [50,51,52]. This structure relies heavily on bold and aggressive male behaviors to establish territories and mate-guard.
Given these traits we must dwell on the types of behaviors established as the arsenal used by anoles in adversarial interactions. These predominantly include the establishment and guarding of a territory (territorial behavior), signaling to conspecifics of agonistic intentions (a form of honest signaling), and of course mate attracting signals that serve to display strength, virality and ability to take risks [53].
As polygynous lizards, Anolis males establish and guard territories with various females [50,54]. Territoriality refers to the agonistic behavior that anoles utilize to defend a specific area or territory, characterized by site fidelity (remaining in or returning to a fixed area) and exclusivity (the exclusion of individuals, specifically conspecifics of the same sex in that area) [54]. Male territories vary by species, but range between 50-100 m2 [55,56,57] . Meanwhile, females establish territories based on nesting, dominance to control resources and ideal locations for predator evasion [58]. While less studied, female territories, which exclude the need to defend mates from conspecifics are smaller and estimated to be half the size of their male counterparts. Anoles will often avert physical altercations by signaling strength and aggression (discussed below as the major focus of this paper), but often will physically defend their territories by biting, wrestling, and same sex copulations [59,60]. There is some debate on how territoriality and territory defense alter with captive anoles [58], but the underlying aggression remains a focus of their behavioral repertoire in both natural conditions and mesocosm experiments.
The expression of aggressive behaviors and communication among members of the Anolis species shapes their social dynamics, and evolutionarily constrain their interactions with members of the biological communities they inhabit. Anoles showcase a relatively extensive repertoire of behaviors in their communication, which has made them model organisms for both ethology and evolution [33]. For this meta-analytical review, we only focus on agonistic displays and ones that have a dual purpose with courtship and conspecific agonism: head-bobs, dewlap extensions, and pushups. The length (display time), stamina, rate of signaling (count of signals/minute), are all honest signals of strength in energy-limited ectotherms. They can communicate the size and strength of the individual, its sexual receptiveness and level of arousal, and dominance in defending territories [57,61,62,63].
Anoles use a vertical gyration, resembling a human pushup (hence the name), to signal to individuals who enter their territory to step down. Pushup displays (PU) are primarily employed by anoles as a visual communication tool to signal dominance, territorial defense, and in a lesser role of attracting potential mates. These actions convey fitness to prospective mates and provide the impression that the anole is bigger and more intimidating to predators and rivals. Male anoles exhibit pushup displays more than females [55,64]. By creating hierarchies through pushup displays, they play a critical role in intra-specific interactions— reducing the necessity for physical conflicts and injury [65]. Anoles use a vertical gyration, resembling a human pushup (hence the name), to signal to individuals who enter their territory to step down (S1). Pushup displays are primarily employed by anoles as a visual communication tool to signal dominance, territorial defense, and in a lesser role attracting potential mates. By flexing and extending their forelimbs, anoles perform pushup displays, quickly raising and lowering its body often in conjugation with head bobbing and extending their dewlaps [66]. These actions convey fitness to prospective mates and provide the impression that the anole is bigger and more intimidating to predators and rivals. Male anoles exhibit pushup displays more than females, and they are used to assert territorial boundaries and ward off intruders [55,64]. Moreover, by creating hierarchies through visual cues, pushup displays play a critical role in intra-specific interactions, hence reducing the necessity for physical conflicts [65].
Anoles unfold a dewlap, a brightly colored skin appendage from under their neck which in many species includes specs and patterns, to startle competitors and attract mates [67,68,69]. In females, dewlaps appear smaller and used less frequently than males [70]. In males, the reproductive use of the display takes precedence during the mating season. These dewlaps, which are energetically expensive to make and sustain when combined with head bobbing, are a good indicator of a male's health and vitality. Dewlap extensions (DE) in males are strongly associated with breeding season and the level of testosterone of the displayer. Higher testosterone is correlated with morphological changes, increased display rates of all signals, and greater stamina [71,72,73]. It is easy to connect this behavior specifically with the “good gene hypothesis”, where females increase the likelihood of offspring quality and survivorship by choosing mates with higher quality genes [74,75], and ones that are able to boldly take risks in the face of adversaries and predators (the handicap principle) [76,77]
Head-bobbing (HB) displays are visual signaling mechanisms to convey dominance. This display is defined as a discontinuous series of up-and-down movements of the body and tail, where each individual upward movement is referred to as a bob [78]. Head-bobbing serves to communicate the lizard's presence, size, and vigor to rivals and mates, thereby establishing dominance and minimizing the likelihood of harmful conflict. Predominantly exhibited by male anoles, these displays are essential for territorial defense and deterring intruders, indicating the male's readiness to defend his space [64,79]. Additionally, head-bobbing has a lesser role in courtship [65].Predominantly exhibited by male anoles, these displays are essential for territorial defense and deterring intruders, indicating the male's readiness to defend his space [64,79]. Additionally, head-bobbing has a lesser role in courtship [65].
Given the propensity of anoles for aggressive behaviors, they are ideal organisms to cohabitate with humans in urban zones. We hypothesize that the pressures imposed on populations of anoles by human activity would directionally select for more aggressive populations in urban zones. Therefore, we expect that studies conducted in urban zones should show higher display rates than studies conducted in “natural” environments. Additionally, given that the traits of invasive populations are expected to be more aggressive, we hypothesize that the studies conducted on the same species would find bolder and more aggressive anoles in invasive ranges, and especially the frontline of the invasion, than studies of populations in native ranges. This follows a pattern well-documented in invasive cane toads (Bufo marinus) in Australia. The toads act more aggressively along the frontline of their invasion across the continent [80]. Lastly, if aggression is facilitating biological invasions, we expect that the overall signal of aggression in invasive populations of anoles would be significantly stronger than that of native competing species in the same locations.
2. Materials and Methods
2.1. Study Organisms
In our study we compared two species of anoles, green (AC) and brown (AS). The brown anole is native to Cuba, Jamaica, and the Bahamas [36,81,82,83]. It has become invasive in several U.S. states expanding from Florida north to Georgia and as far west as Texas. Additional invasive populations, resulting from the exotic pet trade are found in Hawaii, and California and Taiwan [84,85]. The green anole is native to North America, particularly the southeastern United States [86]. It too has been introduced as an invasive to Hawaii [87]. Both AS and AC adults typically measure (SVL+ tail) between 12-22 cm with brown anoles leaning towards the upper part of the range [87]. Both species are dimorphic with males being 2-3 times larger than females [88,89]. Both species live an average of 18 months, but they can live up to five years in the wild [90,91]. Both species are insectivores, feeding primarily on small arthropods [72,92].
The species diverge in habitat preference and some morphological attributes. AS prefer sunny, exposed, and disturbed habitats with low vegetation cover [84,93], whereas AC prefer dense vegetation [94]. AC, formerly known as the American chameleon, apply color change for crypsis and mate attraction while AS are fixed in brown coloration [95]. AS lay 1-2 eggs at approximately 10-day intervals throughout the breeding season, which spans May-September [62]. AC lay single-egg clutches at approximately 7- to 14-day intervals, totaling around 15 offspring for the entire breeding season [87].
2.2. Data Collection
Conducting research from a primarily undergraduate teaching institution, we had restricted access to databases. Therefore, we relied primarily on the limited access provided by Elsevier Science Direct, JSTOR and Google Scholar. We relied heavily on interlibrary loan to supplement access to articles. We supplemented our database with studies identified in the bibliographies of articles we found helpful and searching for articles from prolific research groups in the field (Losos, Lovern, Jenssen, Lailvaux, etc.). Our searches included species names (AS, AC and the common names Brown Anole and Green Anole). We combined those with key words with focus behaviors (head-bob and dewlap extension), and environmental key terms (Urban vs. forest). The Latter was only relevant to the urban zones, identifying papers focusing on urbanization, especially in the context of the invasive AS.
2.3. Data Analyses
Using a couple of criteria, we narrowed our database to 23 publications (S1) which included peer reviewed manuscripts and theses only. We narrowed our database for the needed information on the source of animal collection, availability of all statistics for power analyses (sample sizes, mean display rates, standard deviation (SD) and or standard error (SE).
In addition, we collected descriptive information about each study: animal source (wild caught or lab bred), environment of collection site (urban zones defined as metropolitan areas with over 100,000 human inhabitants), and information regarding the experimental set up (field study or mesocosm/agonistic exposure Male-Male (MM) or reproductive scenario with Male-Female (MF) setup). Lastly, we noted the developmental state of the subjects of the studies (Juvenile/Adult) and the sex of the focal test subject.
Our data mining yielded a total of 111 comparable data. Ten papers yielded 26 data points for AC (excluding one study from the invasive range in Hawaii due to low replicability). AS literature provided 85 data points, with 23 from the native range in Cuba and the Bahamas, and the majority from the invasive range in the continental USA. To facilitate comparability between grouping variables, we made sure that the amount of data for each category combination had at least five data (Table 1, Figure 1).
2.4. Meta Analyses
Given the large scale of these meta-analyses, we ran weighted, means-based, random effect meta-analyses models (i.e. assuming different general means μ). We used grouping based on the predefined categories: environment, species, sex, and invasive status. We ran the meta-analyses based on the seven-step protocol published by Cummings [96]. To do so, we used the ESCI package [97] on the Jamovi interface [98] running on the R platform [99]. Using this protocol, we generated forest plots [100], comparative distributions and uncertainty plots [101].
3. Results
We narrowed our data based on the selection criteria and identified a subset of 23 peer-reviewed sources. These sources provided complete data, including sample sizes, observation times, behavior rates (counts/time unit), and either standard deviations or standard errors of the mean. Of the sources, 7 provided data on Anolis carolinensis and 18 for Anolis sagrei (Table 2). Despite the relatively low number of studies, together these generated 114 data (111 after removing juvenile individuals). Additionally, a publication bias was found. Significantly more publications were published on the brown anoles than on the green anoles (Table 1 and Table 2). Three additional biases were found: (1) more studies were published using urban anoles as subjects than ones collected in rural locations; (2) the studies focusing on brown anoles were overwhelmingly using populations from the invasive range over the native range; and (3) the lizards were predominantly brought into the lab for mesocosm experiments in terraria.
Observing the entire data set, the two species differed in dewlap extension rates, and in pushups but did not in their head bobbing behavior with P values of p>0.01, p=0.03, p=0.893 respectively (Figure 2, Figure 3A-C and Figure S3). Only in the pushup behaviors did the green anoles have a higher rate of signaling than the brown anoles. The anoles significantly signaled more in urban environments than in non-urban zones, with P values <0.01 for both dewlap extension and head- bobbing (Figure 3D-E). AS signaled more than AC across all behaviors (Figure 2), however the urban green anoles signaled with head-bobs at a greater rate than the urban brown anoles. Green anoles favor the head-bobbing display of aggression across all habitats. Meanwhile, brown anoles use dewlap extensions and head-bobbing at equal rates in non-urban zones, drop all signaling rates in urban zones, but favor dewlap extensions over all other displays in populations sources from those habitats.
Data for AC was significantly limited in comparison with AS, as mentioned above. We were thus limited in moderator analyses we could run. The environment from whence the animals were collected did not significantly impact dewlap extension rates (p=0.62), while sex (S4) had a marginally significant impact (pone tailed =0.028), with females signaling less than males. The rates of head-bobbing did not vary neither by urban environment (p=0.977), nor by sex (p=0.845, S4). We did not run a pushup meta-analysis for AC given the sample size we obtained was below the recommended threshold of five data per category.
Anolis sagrei’s behavior shows significantly more variable behaviors than its green counterpart. The dewlap extension rate was not as significant as the head bobbing behaviors (S5). The urban anoles bobbed their heads at a fourth of the rate of non-urban anoles (p<0.01, Figure 4A). For dewlap extension rates, the urban vs. non-urban environment comparison was not statistically significant (p=0.174), however, a clear trend was visible with urban anoles signaling more than non-urban counterparts (Figure 4B). Similar to the environment comparison, the invasive populations showed a trend of signaling more with dewlap extensions than the anoles in the Caribbean native range (p=0.189, Figure 4C). Meanwhile, aggressive head-bobbing displays were signaled by invasive populations at a third of the rate than in their native range counterparts (p<0.01, Figure 4D). Unsurprisingly, males signaled six times more than females (p=0.3) with head-bobbing, and unfortunately, we did not have enough data for sex-based comparison with dewlap extension rates.
4. Discussion
The two species of anoles varied greatly in their behaviors. Brown anoles were plastic in their response and appear to focus on signals that have dual use, mate attractions and challenger deterrence. Green anoles, on the other hand, seem to favor territorial defense signals and did not vary in behavior based on population origin.
4.1. Frontline of Invasion Compared with Established Invasive Communities
Addressing the behavior of these lizards cumulatively poses the challenge of comparing apples to oranges. The overall data suggests that AS signal more than AC, however, we suggest caution in that specific interpretation. Given the weaker data set for AC, this comparison may be influenced by data availability bias. However, when comparing the balance or proportion of signals, distinct behavioral profiles can be identified for each species. AC, a widespread species clearly focuses on head-bobbing displays, with significantly higher rates, over dewlap extension and pushups. These patterns persisted in our analysis independently of the habitat from which the populations were collected. We interpret the shape of this profile to suggest a high focus on terrestrial defense (excluding the push-up data which was not robust enough), and a secondary investment in mate attracting signaling (primary focus of dewlap extension activity). Meanwhile, AS showed flexibility in its behavioral preferences suggesting flexible energetic allocation, a signature of species that has high invasion potential [102,103].
The decrease in territorial behavior seen in the invasive AS populations was surprising. We expected to observe a shift in the invasive populations, however the significant decrease in head-bobbing rates surprised us as we predicted an opposite pattern. Current trends suggest invasive species should express aggression, as this is what gives them an upper leg in the invaded territory [16]. Thus, many studies suggest that invasives are either species that are aggressive to begin with, or the invasive range populations exhibit greater boldness and aggression than native range populations [103,104]. With that in mind, why are the AS displaying less in their invasive range? We provide three hypotheses that can explain this observed patten, all plausible and likely to play a role in the observed behavioral patterns. Thus, many studies suggest that invasive are either species that are aggressive to begin with, or the invasive range populations exhibit greater boldness and aggression than native range populations [103,104]. Then why are the AS displaying less in their invasive range? We provide three hypotheses that can explain this observed patten, all plausible and all likely play a role in the observations. Then why are the AS displaying less in their invasive range? We provide three hypotheses that can explain this observed patten, all plausible and all likely play a role in the observations.
4.1.1. Sex Over Aggression
Dewlap extension signals were higher in the invasive populations than in the native ones. This suggests that the brown anoles use this mate attracting signaling in preference over the head-bobbing. While in some cases, the dewlap extension is also a warning signal, it is used in the agonistic fashion more by females than the males [52]. We believe there is a high probability that invasive populations focus on mate attracting over aggression for a couple of reasons. First, the invasive populations are denser and thus competition for mates is lower as mate availability is greater. Second, invasive populations tend to have a higher rate of dispersal syndrome individuals which prefer dispersal over territoriality [12,105,106]. Individuals that prefer dispersal will invest energy in quickly attracting mates, many times choosing sneaky copulations over defending a territory with a harem [12,105,106]. Individuals that have a preference for dispersal, will invest energy to quickly attract mates, likely resulting in a choice for sneaky copulation over territorial defense [107].
4.1.2. Invasion Front and Habituated Populations
As alluded to in the previous section, dispersal takes a priority in invasive populations, and aggressive behaviors tend to be associated with these tendencies. The aggressive territorial behaviors are not always the same behaviors that would benefit the invasive populations on the front of the invasion, and the measure of risk-taking decisions in the anoles are not well surveyed in the literature. Nonetheless, the study of some of the most infamous biological invasions, namely the cane toads in Australia, suggest that the behavioral syndromes in the range expanding front of an invasion give way to shier, less aggressive, and less risk-taking populations in established habitats [108,109]. Given we do not have clear data on the age of the invasive population in each of the sampled population we do not have the ability to test whether that the pattern we observed is a result of habituation of the invasive populations, or a factor of traits associated with the range expansion. Correlating the behavioral profiles of populations with the historic record of invasion may reveal the answer to this question.
As alluded to in the previous section, dispersal takes a priority in invasive populations, and aggressive behaviors tend to be associated with these tendencies. The aggressive territorial behaviors are not always the same behaviors that would benefit the invasive populations on the front of the invasion, and the measure of risk-taking decisions in the anoles are not well surveyed in the literature. Nonetheless, the study of some of the most infamous biological invasions, namely the cane toads in Australia, suggest that the behavioral syndromes in the range expanding front of an invasion give way to shier, less aggressive, and less risk-taking populations in established habitats [108,109]. Given we do not have clear data on the age of the invasive population in each of the sampled populations, we do not have the ability to test whether the pattern we observed is a result of habituation of the invasive populations, or a factor of traits associated with the range expansion. Correlating the behavioral profiles of populations with the historic record of invasion may reveal the answer to this question. Density dependent energetics. While we think this hypothesis has merit, we are also cautious in suggesting this would be the leading cause of reduced aggression. Putnam, Pauly and Blumstein [110] suggest that brown anoles are less aggressive in California, where they have not been present as long as in some of the sampled studies in the Southeastern US.
4.1.3. Marginal Costs of territorial Defense
The final dynamic worth discussing is the tendency for invasive species to form denser populations in new ranges compared to their native ranges [111,112,113], a pattern also observed in invasive brown anoles [114,115,116]. The energetic tradeoffs in denser populations shift, driving individuals to disperse [117,118] and occupy less rich habitats that experience less competition [119,120]. The inference we draw from ideal free distribution [117] is that organisms that inhabit areas of high density adapt their behavior in various ways to maximize fitness. We hypothesize that aggressive individuals with the dispersal syndrome would be found at the invasion front, but individuals that are more appeasing, and invest less in aggressive behaviors would be the ones who stay behind in the higher density populations. In such conditions, investing in territorial defense would be a constant energetic drain, and therefore would also explain the lower rate of aggressive behaviors we observed in the literature. In lieu of energetic investment in head bobbing and pushups the established invasive brown anoles are using displays that have a dual use, defense and sexual attraction.
4.1.4. Tolerance as an adaptive Trait to Higher Densities
Green anoles hardly show differences in behavior between the urban and non-urban environments; however, brown anoles show an overall decrease in displays in the urban zones. Lailvaux et al. [11] suggests that newly established urban populations tend to be less competitive than their non-urban counterparts, specifically due to overabundance of forage and increased predatory risks from human commensal predators. As we suggest above with the pattern of biological invasions, tolerance (lower aggression) in urban zones may be driven by the necessity to coexist in overlapping and limited habitats where continuous aggressive behavior is energetically wasteful and thus maladaptive. Why is it maladaptive? Populations in urban environments are denser than in “natural” zones. Urban AS peaked at 0.97 individuals/m2 in a study in urban zones in the Bahamas [121], compared with 0.2 in forested areas [122]. The same pattern is true for AC, but at lower densities 0.14 individuals/m2 individuals compared with 0.06 in forested zones [38,121]. Given greater hazards in urban zones (predation, physiological, disturbance) the plastic response observed in AS is likely an adaptive change towards energetic efficiency (Frey et al., 2018; Seress et al., 2011). Moreover, AS success as an urban invasive is attributed to their capacity to adapt and respond to external stimuli while minimizing conflicts in already hazardous zones [123].
4.2. Sexual Signals Counter Agonistic Signals
As polygynous species, both agonistic and sexual signals have fitness tradeoffs in all anoles. The analyses we conducted allow us to gleam insight into those tradeoffs and draw some conclusions about the importance of territoriality in the different species, and across different habitats inhabited by the populations sampled in the published literature. A shift in priority we observed in the invasive populations of brown anoles suggesting that they rely more heavily on sexual signaling, and less on territoriality. We postulate that this is likely a selective pressure due to population density as discussed above. Two potential reasons may lead to the decline in territorial defense. First, a shift in life history tradeoffs in the invasive range, with supposed higher predation risks and resource competition in continental habitats [81], may increase the fitness value of sneaky copulations and outweigh the risk-taking behavior associated with agonistic territorial defense. Alternately, the abundance of competition for mates would suggest that defense of territories may become impossible with many sneaky competitor males around. In this scenario, the shift to high mate attracting signals may be the better fecundity producing strategy as the energetic payoff of territorial defense becomes an energy sink. We do not have the analytical power in this study to tease those two potential evolutionary paths, however, we can only suggest that it is these shifts in behavioral profiles, and suggest that the invasive populations attract mates, and guard them in different ways than the native populations in the Caribbean. It is likely that those shifts are responsible for the highly successful and expanding brown anole invasion across North America.
4.3. Publication Bias
One of the greatest benefits of meta-analyses is the ability to assess publication and research biases. Ecological research is always a balance of tradeoffs, and significant assumptions are made on the validity of studies under constraints.
This study identified several constraints that must be considered in interpretation of behavioral studies in anoles. First and foremost, [brown] anoles in captive conditions signal across the board at lower rates than their counterparts in field. Laboratory-based experiments offer various benefits, including the ability to source anoles from suppliers, control of the environmental noise and high level of control of experimental set-up, to name a few. However, the signal is strong that the setting greatly influences how the animals responded with weaker signals than the free-range counterparts. Additionally, two more biases were observed, specifically in the source of experimental populations. Finding data from non-urban green anoles was a challenge, with effort we were able to find some papers that represent under 16% of data. Behavioral studies of brown anoles in their native-range and represented 29% of data collected. Urban bias was also evident in the invasive AS with over 60% of data being sourced in urban zones. We connect this bias towards ease of research near university campuses, locations of animal suppliers, and lower costs of trapping in the urban environments. The native range of AS in Cuba specifically, limits access to most researchers based in US Academic institutions, which provide the major funding sources for this research.
Last, as the ecological research community, we do not stress the reporting of data relevant to power analysis in numerical form. To estimate effect sizes, we used rulers and printed versions of graphs to extract the needed information for this analysis, and excluded many publications where the relevant data could not be estimated (missing sample sizes, means or standard deviations). This reduced the accuracy of the meta-analyses. However, we trust that the signals we observe are still correct. We believe our conclusions to be representative of the patterns found in the environments where the studies were conducted by the well-established Anolis research community but would have had reduced noise given better reporting practices presently enforced by an increasing number of journals.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, S.B.; methodology, M. J. and S.B.; software, S.B.; validation, M.J. and S.B.; formal analysis, S.B.; investigation, M.J.; resources, S.B.; data curation, M.J.; writing—original draft preparation, M.J. and S.B.; writing—review and editing, S.B..; visualization, S.B..; supervision, S.B.; project administration, M.J.; All authors have read and agreed to the published version of the manuscript.
Funding
no funding was used for this publication.
Institutional Review Board Statement
As a meta-analytical study no human subjects or animals were used in this study
Data Availability Statement
We made our data available for evaluation on Mendeley Data [124] (S2).
Acknowledgments
this study would not have been initiated without the initial modular study conducted by Parker Skinner as part of the Urban Ecology course taught at Washington and Lee University in the spring of 2020, as the course shifted to a remote teaching and lab module in response to the COVID-19 Pandemic. Parker piloted the idea of running a meta-analysis on anole behavior, showed that it can be done as an undergraduate research student, and inspired this work.
Conflicts of Interest
The authors have no conflict of interest to declare
References
- Wilson, E.O. Biodiversity; The National Academy Press, 1988.
- Rypel, A.L. Do Invasive Freshwater Fish Species Grow Better When They Are Invasive? Oikos 2014, 123, 279–289. [Google Scholar] [CrossRef]
- Leishman, M.R.; Cooke, J.; Richardson, D.M. Evidence for Shifts to Faster Growth Strategies in the New Ranges of Invasive Alien Plants. J. Ecol. 2014, 102, 1451–1461. [Google Scholar] [CrossRef] [PubMed]
- Graebner, R.C.; Callaway, R.M.; Montesinos, D. Invasive Species Grows Faster, Competes Better, and Shows Greater Evolution toward Increased Seed Size and Growth than Exotic Non-Invasive Congeners. Plant Ecol. 2012, 213, 545–553. [Google Scholar] [CrossRef]
- Liao, Z.Y.; Zheng, Y.L.; Lei, Y.B.; Feng, Y.L. Evolutionary Increases in Defense during a Biological Invasion. Oecologia 2014, 174, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Stigall, A.L. Invasive Species and Evolution. Evol. Educ. Outreach 2012, 5, 526–533. [Google Scholar] [CrossRef]
- Snyder, W.E.; Evans, E.W. Ecological Effects of Invasive Arthropod Generalist Predators. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 95–122. [Google Scholar] [CrossRef]
- David, P.; Thébault, E.; Anneville, O.; Duyck, P.F.; Chapuis, E.; Loeuille, N. Impacts of Invasive Species on Food Webs: A Review of Empirical Data. Adv. Ecol. Res. 2017, 56, 1–60. [Google Scholar] [CrossRef]
- Bleicher, S.S.; Rosenzweig, M.L. Too Much of a Good Thing? A Landscape-of-Fear Analysis for Collared Peccaries ( Pecari Tajacu ) Reveals Hikers Act as a Greater Deterrent than Thorny or Bitter Food. Can. J. Zool. 2018, 96, 317–324. [Google Scholar] [CrossRef]
- Bleicher, S.S.; Dickman, C.R. On the Landscape of Fear: Shelters Affect Foraging by Dunnarts (Marsupialia, Sminthopsis Spp.) in a Sandridge Desert Environment. J. Mammal. 2020, 101, 281–290. [Google Scholar] [CrossRef]
- Lailvaux, S.P.; Edwards, J.R.; Lailvaux, S.P.; Winchell, K.M.; Reynolds, R.G.; Prado-Irwin, S.R.; Puente-Rolón, A.R.; Revell, L.J.; Stroud, J.T.; Colom, M.; et al. Hurricane Irma Induces Divergent Behavioral and Hormonal Impacts on an Urban and Forest Population of Invasive Anolis Lizards: Evidence for an Urban Resilience Hypothesis. Front. Ecol. Evol. 2020, 8, 1–13. [Google Scholar] [CrossRef]
- Gruber, J.; Brown, G.; Whiting, M.J.; Shine, R. Geographic Divergence in Dispersal-Related Behaviour in Cane Toads from Range-Front versus Range-Core Populations in Australia. Behav. Ecol. Sociobiol. 2017, 71, 1–7. [Google Scholar] [CrossRef]
- Tsutsui, N.D.; Suarez, A. V; Grosberg, R.K. Genetic Diversity, Asymmetrical Aggression, and Recognition in a Widespread Invasive Species. PNAS 2002, 100, 1078–1087. [Google Scholar] [CrossRef] [PubMed]
- Magellan, K.; Garcıa-Berthou, E. Influences of Size and Sex on Invasive Species Aggression and Native Species Vulnerability : A Case for Modern Regression Techniques. Rev. Fish Biol. Fish. 2015, 25, 537–549. [Google Scholar] [CrossRef]
- Hudina, S.; Hock, K.; Žganec, K. The Role of Aggression in Range Expansion and Biological Invasions. Curr. Zool. 2014, 60, 401–409. [Google Scholar] [CrossRef]
- Pintor, L.M.; Brown, J.S.; Vincent, T.L. Evolutionary Game Theory as a Framework for Studying Biological Invasions. Am. Nat. 2011, 177, 410–423. [Google Scholar] [CrossRef] [PubMed]
- Mckinney, M.L. Urbanization as a Major Cause of Biotic Homogenization. Biol. Conserv. 2006, 127, 246–260. [Google Scholar] [CrossRef]
- Magle, S.B.; Simoni, L.S.; Lehrer, E.W.; Brown, J.S. Urban Predator-Prey Association: Coyote and Deer Distributions in the Chicago Metropolitan Area. Urban Ecosyst. 2014, 17, 875–891. [Google Scholar] [CrossRef]
- Bradley, C. a.; Altizer, S. Urbanization and the Ecology of Wildlife Diseases. Trends Ecol. Evol. 2007, 22, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Meek, P.; Ballard, G.; Fleming, P.; Falzon, G. Are We Getting the Full Picture? Animal Responses to Camera Traps and Implications for Predator Studies. Ecol. Evol. 2016, 1, 1–10. [Google Scholar] [CrossRef]
- Avilés-Rodríguez, K.J.; Kolbe, J.J. Escape in the City: Urbanization Alters the Escape Behavior of Anolis Lizards. Urban Ecosyst. 2019, 22, 733–742. [Google Scholar] [CrossRef]
- St. Juliana, J.; Bleicher, S.S.; Mukherjee, S.; Sundararaj, V.; Brown, J.S.; Kotler, B.P. Human-Commensal Threats; Combinations of Predators Alter Gerbil Risk-Perception in Additive, Substitutable, or Antagonistic Manners. Journal: Conserv. Biol. 2023, 1–20. (In review).
- Pham, A.; Riley, J.; Bleicher, S.S. Human-Wildlife Conflicts and Sensory Disinformation, a Review of Evolutionary Trajectories Caused by Human Land- Management and Rapidly Expanding Urban Landscapes. (In Review). J. Urban Ecol. 2024, 1–33.
- Ciuti, S.; Muhly, T.B.; Paton, D.G.; McDevitt, A.D.; Musiani, M.; Boyce, M.S. Human Selection of Elk Behavioural Traits in a Landscape of Fear. Proc. Biol. Sci. 2012, 279, 4407–4416. [Google Scholar] [CrossRef] [PubMed]
- Ament, R.; Clevenger, A.P.; Yu, O.; Hardy, A. An Assessment of Road Impacts on Wildlife Populations in U. S. National Parks. Environ. Manage. 2008, 42, 480–496. [Google Scholar] [CrossRef] [PubMed]
- Seidler, R.G.; Long, R.A.; Berger, J.; Bergen, S.; Beckmann, J.P. Identifying Impediments to Long-Distance Mammal Migrations. Conserv. Biol. 2015, 29, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Zuñiga-Palacios, J.; Zuria, I.; Castellanos, I.; Lara, C.; Sánchez-Rojas, G. What Do We Know (and Need to Know) about the Role of Urban Habitats as Ecological Traps? Systematic Review and Meta-Analysis. Sci. Total Environ. 2021, 780, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Magle, S.B.; Reyes, P.; Zhu, J.; Crooks, K.R. Extirpation, Colonization, and Habitat Dynamics of a Keystone Species along an Urban Gradient. Biol. Conserv. 2010, 143, 2146–2155. [Google Scholar] [CrossRef]
- Barrett, M.A.; Telesco, D.J.; Barrett, S.E.; Katelyn, M.; Leone, E.H. Testing Bear-Resistant Trash Cans in Residential Areas of Florida. Southeast. Nat. 2014, 13, 26–39. [Google Scholar] [CrossRef]
- Kruszyk, R.; Ciach, M. White Storks, Ciconia Ciconia, Forage on Rubbish Dumps in Poland-a Novel Behaviour in Population. Eur. J. Wildl. Res. 2010, 56, 83–87. [Google Scholar] [CrossRef]
- Theimer, T.C.; Clayton, A.C.; Martinez, A.; Peterson, D.L.; Bergman, D.L. Visitation Rate and Behavior of Urban Mesocarnivores Differs in the Presence of Two Common Anthropogenic Food Sources. Urban Ecosyst. 2015, 18, 895–906. [Google Scholar] [CrossRef]
- Deatherage, N.A.; Cypher, B.; Westall, T.; Kelly, E. Spatial Relationships between Mesocarnivores and Domestic Species in an Urban Environment and Implications for Endangered San Joaquin Kit Foxes. Urban Ecosyst. 2024, 27, 321–334. [Google Scholar] [CrossRef]
- Losos, J.B. Lizards in an Evolutionary Tree: Ecology and Adaptive Radiation of Anoles; Unviersity of California Press: Berkley, CA, USA, 2009. [Google Scholar]
- Roughgarden, J. Resource Partitioning among Competing Spcecies-- A Coevolutionary Approach. Theor. Popul. Biol. 1976, 9, 388–424. [Google Scholar] [CrossRef]
- Roughgarden, J. Competition and Theory in Community Ecology. Am. Nat. 1983, 122, 583–601. [Google Scholar] [CrossRef]
- Les, A.M.; Gifford, M.E.; Parmerlee, J.S.; Powell, R. Do Polymorphic Female Brown Anoles (Anolis Sagrei ) Differ in Sprint Speed or Escape Behavior? Herpetologica 2014, 70, 47–55. [Google Scholar] [CrossRef]
- Cooper, W.E. Risk Factors Affecting Escape Behaviour by Puerto Rican Anolis Lizards. Can. J. Zool. 2006, 84, 495–504. [Google Scholar] [CrossRef]
- Bloch, N.; Irschick, D.J. An Analysis of Inter-Population Divergence in Visual Display Behavior of the Green Anole Lizard (Anolis carolinensis). Ethology 2006, 112, 370–378. [Google Scholar] [CrossRef]
- Kuo, C.Y.; Irschick, D.J.; Lailvaux, S.P. Trait Compensation between Boldness and the Propensity for Tail Autotomy under Different Food Availabilities in Similarly Aged Brown Anole Lizards. Funct. Ecol. 2015, 29, 385–392. [Google Scholar] [CrossRef]
- Losos, J.B. Integrative Approaches to Evolutionary Ecology: Anolis Lizards as Model Systems. Annu. Rev. Ecol. Systamatics 1994, 25, 467–469. [Google Scholar] [CrossRef]
- Hall, J.M.; Warner, D.A. Thermal Spikes from the Urban Heat Island Increase Mortality and Alter Physiology of Lizard Embryos. J. Exp. Biol. 2018, 221, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B. Adaptive Radiation, Ecological Opportunity, and Evolutionary Determinism. American Society of Naturalists E. O. Wilson Award Address. Am. Nat. 2010, 175, 623–639. [Google Scholar] [CrossRef] [PubMed]
- Kolbe, J.J.; Leal, M.; Schoener, T.W.; Spiller, D.A.; Losos, J.B. Founder Effects Persist Despite Adaptive Differentiation : A Field Experiment with Lizards. Science (80-. ). 2012, 335, 1086–1089. [Google Scholar] [CrossRef]
- Lovern, M.B.; Jenssen, T.A.; Orrell, K.S.; Tuchak, T.; Herpetologica, S.; Jun, N. Herpetologists’ League Comparisons of Temporal Display Structure across Contexts and Populations in Male Anolis carolinensis: Signal Stability or Lability? Herpetologica 1999, 55, 222–234. [Google Scholar]
- Eals, J.; Thorpe, R.S.; Malhotra, A. Weak Founder Effect Signal in a Recent Introduction of Caribbean Anolis. 2Molecular Ecol. 2008, 17, 1416–1426. [Google Scholar] [CrossRef]
- Manríquez-morán, Norma, L.; Villagrán-santa Cuz, M.; Méndez-de La Cruz, Fausto, R. Reproductive Biology of the Parthenogenetic Lizard, Aspidoscelis Cozumela. Herpetologica 2005, 61, 435–439. [Google Scholar] [CrossRef]
- Fujita, M.K.; Singhal, S.; Brunes, T.O.; Maldonado, J.A.; Queen, R. Evolutionary Dynamics and Consequences of Parthenogenesis in Vertebrates. Annu. Rev. Ecol. Systamatics 2020, 51, 191–216. [Google Scholar] [CrossRef]
- Jancuchova-Lanskova, J.; Landova, E.; Frynta, D. Are Genetically Distinct Lizard Species Able to Hybridize ? A Review. Curr. Zool. 2015, 61, 155–180. [Google Scholar] [CrossRef]
- Fujita, M.K.; Moritz, C. Origin and Evolution of Parthenogenetic Genomes in Lizards: Current State and Future Directions. Cytogenet. Genome Res. 2010, 127, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Kamath, A.; Losos, J. Reconsidering Territoriality Is Necessary for Understanding Anolis Mating Systems. Behav. Ecol. Sociobiol. 2018, 72, 106. [Google Scholar] [CrossRef]
- Kamath, A.; Losos, J. The Erratic and Contingent Progression of Research on Territoriality : A Case. Behav. Ecol. Sociobiol. 2017, 71, 1–13. [Google Scholar] [CrossRef]
- Jenssen, T.A.; Orrell, K.S.; Lovern, M.B. Sexual Dimorphisms in Aggressive Signal Structure and Use by a Polygynous Lizard, Anolis carolinensis. Copeia 2000, 2000, 140–149. [Google Scholar] [CrossRef]
- Jenssen, T.A.; Garrett, S.; Sydor, W.J. Complex Signal Usage by Advertising Male Green Anoles (Anolis carolinensis): A Test of Assumptions. Herpetologica 2012, 68, 345–357. [Google Scholar] [CrossRef]
- Bush, J.M.; Simberloff, D. A Case for Anole Territoriality. Behav. Ecol. Sociobiol. 2018, 72, 111. [Google Scholar] [CrossRef]
- Losos, J.B.; Schneider, C.J. Anolis Lizards. Curr. Biol. 2009, 19, R316–R318. [Google Scholar] [CrossRef] [PubMed]
- Jenssen, T.A.; Nunez, S.C. Spatial and Breeding Relationships of the Lizard, Anolis carolinensis: Evidence of Intrasexual Selection. Behaviour 1998, 135, 981–1003. [Google Scholar] [CrossRef]
- Orrell, K.S.; Jenssen, T.A. Heterosexual Signalling by the Lizard Anolis carolinensis, with Intersexual Comparisons across Contexts; 2003; Vol. 140; ISBN 1568539033221.
- Reedy, A.M.; Pope, B.D.; Kiriazis, N.M.; Giordano, C.L.; Sams, C.L.; Warner, D.A.; Cox, R.M. Female Anoles Display Less but Attack More Quickly than Males in Response to Territorial Intrusions. Behav. Ecol. 2017, 28, 1323–1328. [Google Scholar] [CrossRef]
- Evans, L.T. Courtship Behavior and Sexual Selection of Anolis. J. Comp. Psychol. 1938, 26, 475–494. [Google Scholar] [CrossRef]
- Noble, G.K.; Bradly, H.T. The Mating Behavior of Lizards; Its Bearing on the Theory of Sexual Selection. Ann. N. Y. Acad. Sci. 1993, 35, 25–100. [Google Scholar] [CrossRef]
- Wilczynski, W.; Black, M.P.; Salem, S.J.; Ezeoke, C.B. Behavioural Persistence during an Agonistic Encounter Differentiates Winners from Losers in Green Anole Lizards. Behaviour 2015, 152, 563–591. [Google Scholar] [CrossRef]
- Calsbeek, R.; Cox, R.M. Experimentally Assessing the Relative Importance of Predation and Competition as Agents of Selection. Nature 2010, 465, 613–616. [Google Scholar] [CrossRef]
- Scott, M.P. Agonistic and Courtship Displays of Male Anolis sagrei. Brevoria 1984, 479, 1–22. [Google Scholar]
- Jenssen, T.A.A. Evolution of Anoline Lizard Display Behavior. Am. Zool. 1977, 17, 203–215. [Google Scholar] [CrossRef]
- Macedonia, J.M.; Stamps, J.A. Species Recognition in Anolis grahami (Sauria, Iguanidae): Evidence from Responses to Video Playbacks of Conspecific and Heterospecific Displays. Ethology 1994, 98, 246–264. [Google Scholar] [CrossRef]
- Leal, M.; Rodriguez-Robles, J.A.; Rodri, J.A. Signalling Displays during Predator – Prey Interactions in a Puerto Rican Anole, Anolis cristatellus. Anim. Behav. 1997, 54, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Simon, V.B. Not All Signals Are Equal: Male Brown Anole Lizards (Anolis sagrei) Selectively Decrease Pushup Frequency Following a Simulated Predatory Attack. Ethology 2007, 113, 793–801. [Google Scholar] [CrossRef]
- Tokarz, R.R.; Mcmann, S.; Smith, L.C.; John-alder, H. Effects of Testosterone Treatment and Season on the Frequency of Dewlap Extensions during Male – Male Interactions in the Lizard Anolis sagrei. Horm. Behav. 2002, 41, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Tokarz, R.R.; Patterson, A. V.; McMann, S. Laboratory and Field Test of the Functional Significance of the Male’s Dewlap in the Lizard Anolis sagrei. Copeia 2003, 3, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, K.E.; Harmon, L.J.; Losos, J.B. Evolution of Anolis Lizard Dewlap Diversity. PLoS One 2007, 2, e274. [Google Scholar] [CrossRef] [PubMed]
- Tokarz, R.R. An Experimental Test of the Importance of the Dewlap Inm Male Mating Success in the Lizard Anolis aagrei. Herpetologica 2002, 58, 87–94. [Google Scholar] [CrossRef]
- Battles, A.C.; Kolbe, J.J. Miami Heat : Urban Heat Islands Influence the Thermal Suitability of Habitats for Ectotherms. Glob. Chang. Biol. 2019, 25, 562–576. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.M.; Stenquist, D.S.; Henningsen, J.P.; Calsbeek, R. Manipulating Testosterone to Assess Links between Behavior, Morphology, and Performance in the Brown Anole Anolis sagrei. Physiol. Biochem. Zool. 2009, 82, 686–698. [Google Scholar] [CrossRef]
- Trivers, R.L. Sexual Selection and Resource-Accruing Abilities in Anolis garmani. Evolution (N. Y). 1976, 30, 253–269. [Google Scholar]
- Crawford, N.G.; McGreevy, T.J.; Mullen, S.P.; Schneider, C.J. The Genetic Basis of Conspicuous Coloration in the Guadeloupean Anole: Evolution by Sexual and Ecological Selection. Ecol. Evol. 2023, 13, 1–13. [Google Scholar] [CrossRef]
- Lailvaux, S.P.; Gilbert, R.L.; Edwards, J.R. A Performance-Based Cost to Honest Signalling in Male Green Anole Lizards (Anolis carolinensis). Proc. R. Soc. B Biol. Sci. 2012, 279, 2841–2848. [Google Scholar] [CrossRef] [PubMed]
- Johnston, R.A. Sexual Selection, Honest Advertisment and the Handicape Principle: Reviewing the Evidence. Biol. Rev. 1995, 70, 1–65. [Google Scholar] [CrossRef] [PubMed]
- McMann, S.; Patterson, A.V. Display Behavior of Resident Brown Anoles (Anolis sagrei) during Close Encounters with Neighbors and Non-Neighbors. Herpetol. Conserv. Biol. 2012, 7, 27–37. [Google Scholar]
- Lovern, M.B.; Jenssen, T.A. Form Emergence and Fixation of Head Bobbing Displays in the Green Anole Lizard (Anolis carolinensis): A Reptilian Model of Signal Ontogeny. J. Comp. Psychol. 2003, 117, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Bernal, E.; Brown, G.P.; Shine, R. Invasive Cane Toads : Social Facilitation Depends upon an Individual ’ s Personality. PLoS One 2014, 9, e102880. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, D.S.; Losos, J.B.; Schoener, T.W.; Spiller, D.A.; Kolbe, J.J.; Leal, M. Predation-Associated Modulation of Movement-Based Signals by a Bahamian Lizard. Proc. Natl. Acad. Sci. 2014, 111, 9187–9192. [Google Scholar] [CrossRef] [PubMed]
- Côté, I.M.; Darling, E.S.; Malpica-Cruz, L.; Smith, N.S.; Green, S.J.; Curtis-Quick, J.; Layman, C. What Doesn’t Kill You Makes You Wary? Effect of Repeated Culling on the Behaviour of an Invasive Predator. PLoS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, S.R.; Bursey, C.R. Transport of Helminths To Hawaii Via the Brown Anole, Anolis sagrei (Polychrotidae). J. Parasitol. 2000, 86, 750. [Google Scholar] [CrossRef]
- Schwartz, A.; Henderson, R.W. Amphibians and Reptiles of the West Indies: Descriptions, Distributions, and Natural History; University of Florida Press: Gainesville, FL, USA, 1991. [Google Scholar]
- Fisher, S.R.R.; Del Pinto, L.A.A.; Fisher, R.N.N. Establishment of Brown Anoles (Anolis sagrei) across a Southern California County and Potential Interactions with a Native Lizard Species. PeerJ 2019, 8, e8937. [Google Scholar] [CrossRef]
- Horr, D.M. The Ecology of Dynamic Body Color in the Green Anole, Anolis Carolinensis (Honors Thesis), 2019.
- Lovern, M.B.; Holmes, M.M.; Wade, J. The Green Anole (Anolis carolinensis): A Reptilian Model for Laboratory Studies of Reproductive Morphology and Behavior. ILAR J. 2004, 45, 54–64. [Google Scholar] [CrossRef]
- Cox, C.L.; Peaden, R.T.; Cox, R.M. The Metabolic Cost of Mounting an Immune Response in Male Brown Anoles (Anolis sagrei). J. Exp. Zool. 2015, 323, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Mason, P.; Adkins, K. Hormones and Social Behavior in the Lizard, Anolis carolinensis. Horm. Behav. 1976, 86, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Vigil, S. Brown Anole (Anolis sagrei). In Natural History Publication Series; Mengak, M.T., Ed.; Tampa, FL, FL, 2006; pp. 1–6.
- Yasumiba, K.; Okada, A.; Okochi, I.; Iwai, N. Minimum Longevity and Growth of the Invasive Green Anole, Anolis carolinensis, in Chichi-Jima of the Ogasawara Islands, Japan. Curr. Herpetol. 2016, 35, 101–105. [Google Scholar] [CrossRef]
- Battles, A.C.; Whittle, T.K.; Stehle, C.M.; Johnson, M.A.; Attles, A.N.C.B.; Hittle, T.A.R.A.K.W.; Tehle, C.H.M.S.; Ohnson, M.I.A.J. Effects of Human Land Use on Prey Availability and Body Condition in the Green Anole Lizard, Anolis carolinensis. Herpetol. Conserv. Biol. 2013, 8, 16–26. [Google Scholar]
- Norval, G.; Chen, C.K.; Hsiao, W.-F.; Huang, S.-C.; Chen, C.-K. The Diet of an Introduced Lizard Species, the Brown Anole (Anolis sagrei), in Chiayi County, Taiwan. Russ. J. Herpetol. 2010, 17, 131–138. [Google Scholar]
- Schaefer, R.R.; Fleet, R.R.; Rudolph, D.C.; Koerth, N.E. Habitat Selection by Anolis carolinensis (Green Anole) in Open Pine Forests in Eastern Texas. In Proceedings of the Proceedings of the 4th Big Thicket Science Conference Southeastern Naturalist; 2009; pp. 63–76.
- Horr, D.M.; Payne, A.A.; Mcentire, K.D.; Johnson, M.A. Sexual Dimorphism in Dynamic Body Color in the Green Anole Lizard. Behav. Ecol. Sociobiol. 2023. [Google Scholar] [CrossRef]
- Cumming, G. Understanding the New Statistics; Effect Sizes, Confidence Intervals, and Meta Analyses; Routledge: New York, New York, 2012.
- Bonnet, D.G.; Calin-Jageman, R.J. ESCI (Effect Sizes and Confidence Intervals); Statpsych: Statistical Methods for Psychologists 2023, 1.5.0.
- Jamovi The Jamovi Project 2023, Version 2.4.
- R Core Team R: A Langauage and Environment for Statistical Computing 2022, Version 4.1.
- Bonnet, D.G. Confidence Intervals for Standardized Linear Contrast Means. Psychol. Methods 2008, 13, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Kay, M. Ggdist: Visualization of Distributions and Uncertinty in the Grammar of Graphics. IEEE Trans. Visulaization Comput. Graph. 2002, 30, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Heavener, S.J.; Carthey, A.J.R.; Banks, P.B. Competitive Naïveté between a Highly Successful Invader and a Functionally Similar Native Species. Oecologia 2014, 175, 73–84. [Google Scholar] [CrossRef]
- Sih, A.; Bell, A.; Johnson, J.C. Behavioral Syndromes: An Ecological and Evolutionary Overview. Trends Ecol. Evol. 2004, 19, 372–378. [Google Scholar] [CrossRef]
- Carpintero, S.; Reyes, J. Capter 7: The Role of Agression in the Success of the Invasive Argentine Ant. In Handbook of Aggressive Behavior Research Ed:; Tawse, C.Q. and S., Ed.; Nova Science Publishers, Inc, 2009; pp. 241–268 ISBN 9781607415831.
- Molina-Montenegro, M. a.; Cleland, E.E.; Watts, S.M.; Broitman, B.R. Can a Breakdown in Competition-Colonization Tradeoffs Help Explain the Success of Exotic Species in the California Flora? Oikos 2012, 121, 389–395. [Google Scholar] [CrossRef]
- Mason, R.A.B.; Cooke, J.; Moles, A.T.; Leishman, M.R. Reproductive Output of Invasive versus Native Plants. Glob. Ecol. Biogeogr. 2008, 17, 633–640. [Google Scholar] [CrossRef]
- Kamath, A.; Losos, J.B. Estimating Encounter Rates as the First Step of Sexual Selection in the Lizard Anolis sagrei. Proc. R. Soc. B Biol. Sci. 2018, 285. [Google Scholar] [CrossRef] [PubMed]
- Hudina, S.; Žganec, K.; Hock, K. Differences in Aggressive Behaviour along the Expanding Range of an Invasive Crayfish: An Important Component of Invasion Dynamics. Biol. Invasions 2015, 17, 3101–3112. [Google Scholar] [CrossRef]
- Michelangeli, M.; Smith, C.R.; Wong, B.B.M.; Chapple, D.G. Aggression Mediates Dispersal Tendency in an Invasive Lizard. Anim. Behav. 2017, 133, 29–34. [Google Scholar] [CrossRef]
- Putman, B.J.; Pauly, G.B.; Blumstein, D.T. Urban Invaders Are Not Bold Risk-Takers: A Study of 3 Invasive Lizards in Southern California. Curr. Zool. 2020, 66, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.C.; Ruiz-Navarro, A.; Britton, J.R. Population Density Modifies the Ecological Impacts of Invasive Species. Oikos 2015, 124, 880–887. [Google Scholar] [CrossRef]
- Čuda, J.; Skálová, H.; Janovský, Z.; Pyšek, P. Competition among Native and Invasive Impatiens Species: The Roles of Environmental Factors, Population Density and Life Stage. AoB Plants 2015, 7, 1–12. [Google Scholar] [CrossRef]
- Akin-Fajiye, M.; Gurevitch, J. The Influence of Environmental Factors on the Distribution and Density of Invasive Centaurea Stoebe across Northeastern USA. Biol. Invasions 2018, 20, 3009–3023. [Google Scholar] [CrossRef]
- Angetter, L.S.; Lötters, S.; Rödder, D. Climate Niche Shift in Invasive Species: The Case of the Brown Anole. Biol. J. Linn. Soc. 2011, 104, 943–954. [Google Scholar] [CrossRef]
- Campbell, T.S.; Echternacht, A.C. Introduced Species as Moving Targets: Changes in Body Sizes of Introduced Lizards Following Experimental Introductions and Historical Invasions. Biol. Invasions 2003, 5, 193–212. [Google Scholar] [CrossRef]
- Chejanovski, Z.A.; Giery, S.T.; Kolbe, J.J. Effects of Urbanization on the Trophic Niche of the Brown Anole, a Widespread Invasive Lizard. Food Webs 2022, 33, e00257. [Google Scholar] [CrossRef]
- Morris, D.W. Density-Dependent Habitat Selection : Testing the Theory with Fitness Data. Evol. Ecol. 1989, 3, 80–94. [Google Scholar] [CrossRef]
- Grand, T.C.; Dill, L.M. Predation Risk, Unequal Competitors and the Ideal Free Distribution. Evol. Ecol. Res. 1999, 1, 389–409. [Google Scholar]
- Morris, D.W. How Can We Apply Theories of Habitat Selection to Wildlife Conservation and Management? Wildl. Res. 2003, 30, 303–319. [Google Scholar] [CrossRef]
- Brown, J.S.; Kotler, B.P. Hazardous Duty Pay and the Foraging Cost of Predation. Ecol. Lett. 2004, 7, 999–1014. [Google Scholar] [CrossRef]
- Schoener, T.W.; Schoener, A. Ecological and Demographic Correlates of Injury Rates in Some Bahamian Anolis Lizards. Copeia 1980, 1980, 839–850. [Google Scholar] [CrossRef]
- Losos, J.B. Detective Work in the West Indies : Integrating Historical and Experimental Approaches to Study Island Lizard Evolution. Teach. Biol. 2007, 57, 585–597. [Google Scholar] [CrossRef]
- Kolbe, J.J.; VanMiddlesworth, P.; Battles, A.C.; Stroud, J.T.; Buffum, B.; Forman, R.T.T.; Losos, J.B. Determinants of Spread in an Urban Landscape by an Introduced Lizard. Landsc. Ecol. 2016, 31, 1795–1813. [Google Scholar] [CrossRef]
- Bleicher, S.S. Anole Behavior Meta-Analysis. Mendeley Data 2024, V1, 1. [Google Scholar] [CrossRef]
Figure 1.
Map displaying sources of populations used in publications used for this meta-analysis generated on Google Earth*. Markers represent locations where populations were sampled and may represent various publications. Bahamian brown anoles were considered native species in this review.
Figure 1.
Map displaying sources of populations used in publications used for this meta-analysis generated on Google Earth*. Markers represent locations where populations were sampled and may represent various publications. Bahamian brown anoles were considered native species in this review.
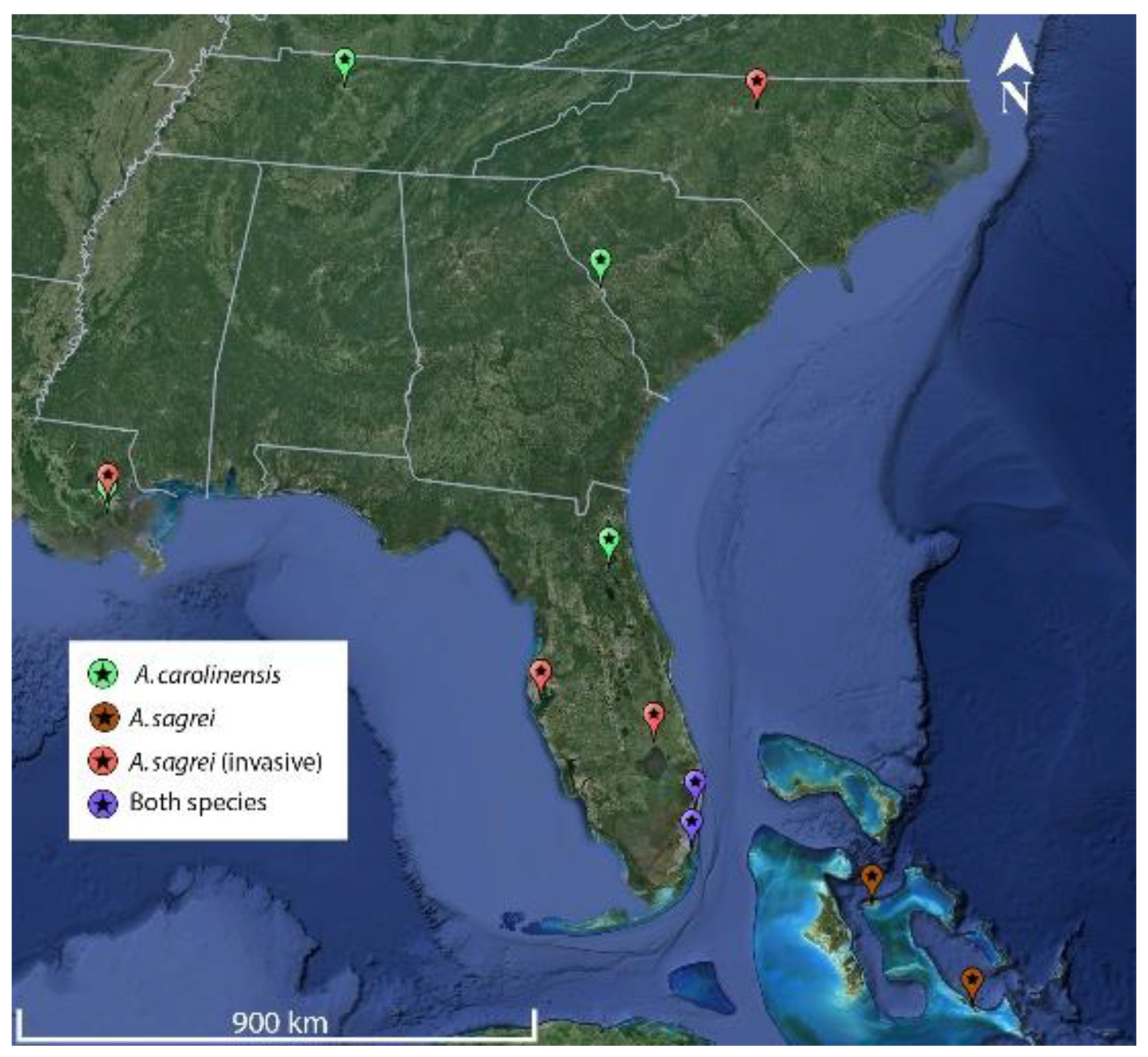
Figure 2.
Visual comparison of mean display rates (per minute) generated by meta-analyses for each of the display behaviors, for each species of anole in urban and non-urban populations. The * represents a significant power analysis for generated meta-analytical mean ±SE. ‡ represents an un-refined estimate for the purpose of visualization, not the outcome of statistical analysis. Overall display rates are greater for Anolis sagrei than Anolis carolinensis, however the former show greater behavioral plasticity and reduction of display rates in urban populations.
Figure 2.
Visual comparison of mean display rates (per minute) generated by meta-analyses for each of the display behaviors, for each species of anole in urban and non-urban populations. The * represents a significant power analysis for generated meta-analytical mean ±SE. ‡ represents an un-refined estimate for the purpose of visualization, not the outcome of statistical analysis. Overall display rates are greater for Anolis sagrei than Anolis carolinensis, however the former show greater behavioral plasticity and reduction of display rates in urban populations.
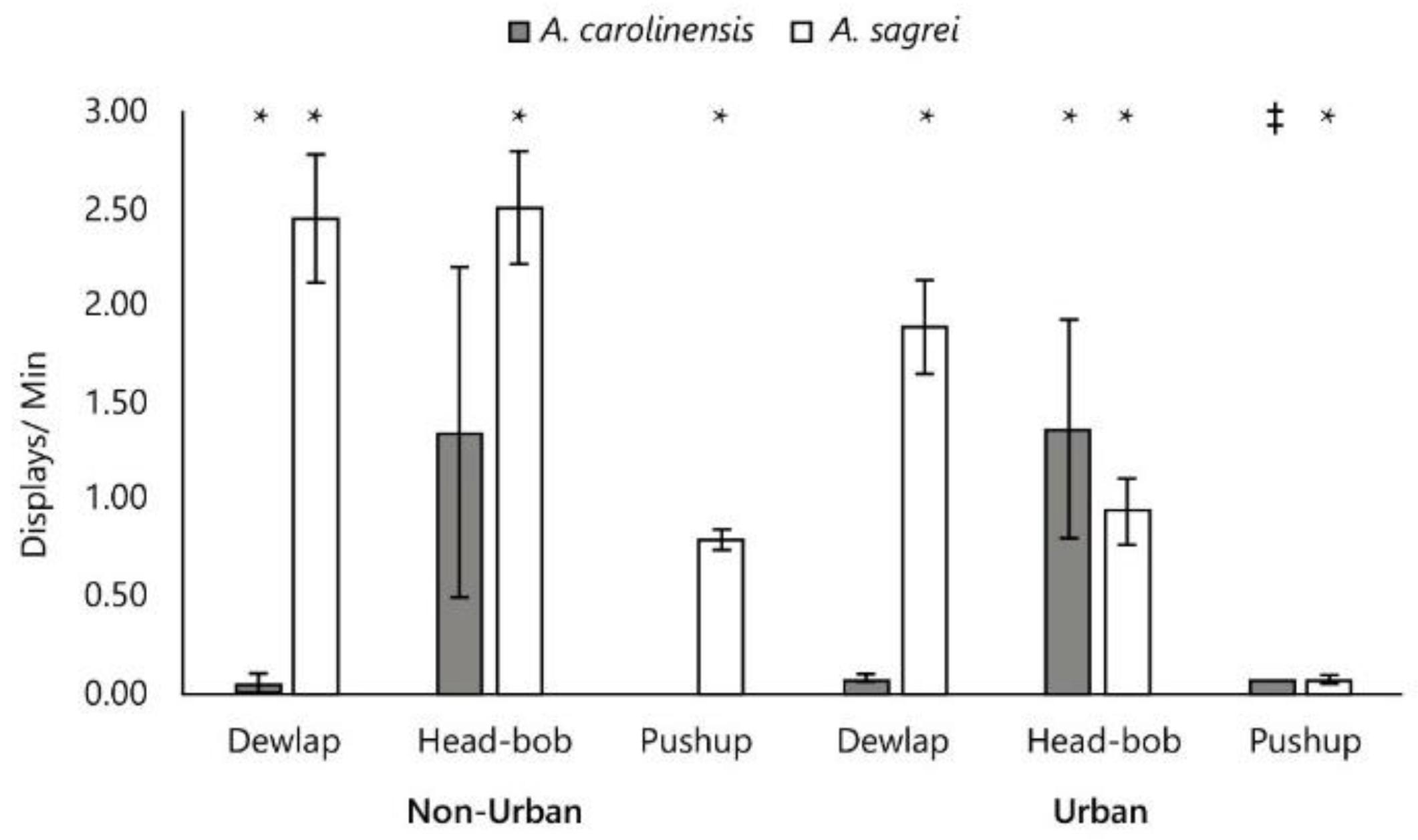
Figure 3.
Distribution of means based on 95% CI generated from a large-scale meta-analysis for the combined dataset (N=111) as expressed in dichotomous moderator analyses of: (A-C) Display rates (per second) by species, (D-E) Display rates/ population environment. Anolis sagrei are shown to display more with dewlap extensions and head-bobs while Anolis carolinensis perform more pushups. Head-bobbing rates are lower while dewlap extension rates are slightly elevated in urban dwelling lizards. .
Figure 3.
Distribution of means based on 95% CI generated from a large-scale meta-analysis for the combined dataset (N=111) as expressed in dichotomous moderator analyses of: (A-C) Display rates (per second) by species, (D-E) Display rates/ population environment. Anolis sagrei are shown to display more with dewlap extensions and head-bobs while Anolis carolinensis perform more pushups. Head-bobbing rates are lower while dewlap extension rates are slightly elevated in urban dwelling lizards. .
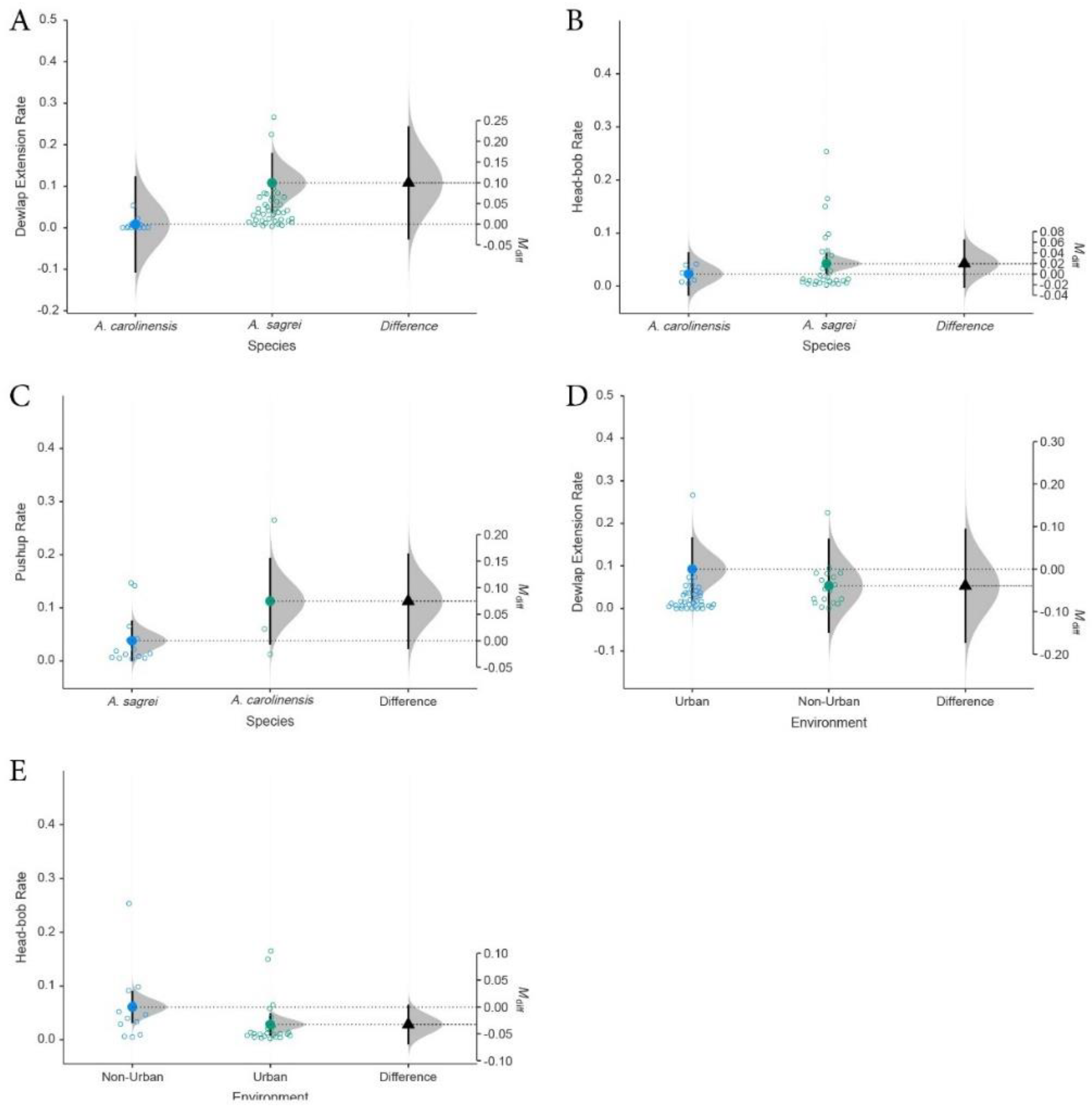
Figure 4.
Distribution of mean display rates based on 95% CI generated from a large-scale meta-analysis Anolis sagrei showing dichotomous moderator analyses with dependent variables of each behavioral display rate (per second): (A-B) comparing urban and non-urban populations, (C-D) comparing population from native and invasive ranges. The patterns of elevated dewlap extension rates and lowered head-bobbing are duplicated in urban environments and invasive populations.
Figure 4.
Distribution of mean display rates based on 95% CI generated from a large-scale meta-analysis Anolis sagrei showing dichotomous moderator analyses with dependent variables of each behavioral display rate (per second): (A-B) comparing urban and non-urban populations, (C-D) comparing population from native and invasive ranges. The patterns of elevated dewlap extension rates and lowered head-bobbing are duplicated in urban environments and invasive populations.
Figure 5.
Distribution of mean display rates based on 95% CI generated from a large-scale meta-analysis Anolis sagrei showing dichotomous moderator analyses with dependent variables of behavioral display rate (per second) comparing mesocosm experimental setups with field studies. (A) for head-bobbing display and (B) dewlap extension rates. Field studies record higher display rates than mesocosm experiments suggesting experimental bias in experimental outcomes.
Figure 5.
Distribution of mean display rates based on 95% CI generated from a large-scale meta-analysis Anolis sagrei showing dichotomous moderator analyses with dependent variables of behavioral display rate (per second) comparing mesocosm experimental setups with field studies. (A) for head-bobbing display and (B) dewlap extension rates. Field studies record higher display rates than mesocosm experiments suggesting experimental bias in experimental outcomes.
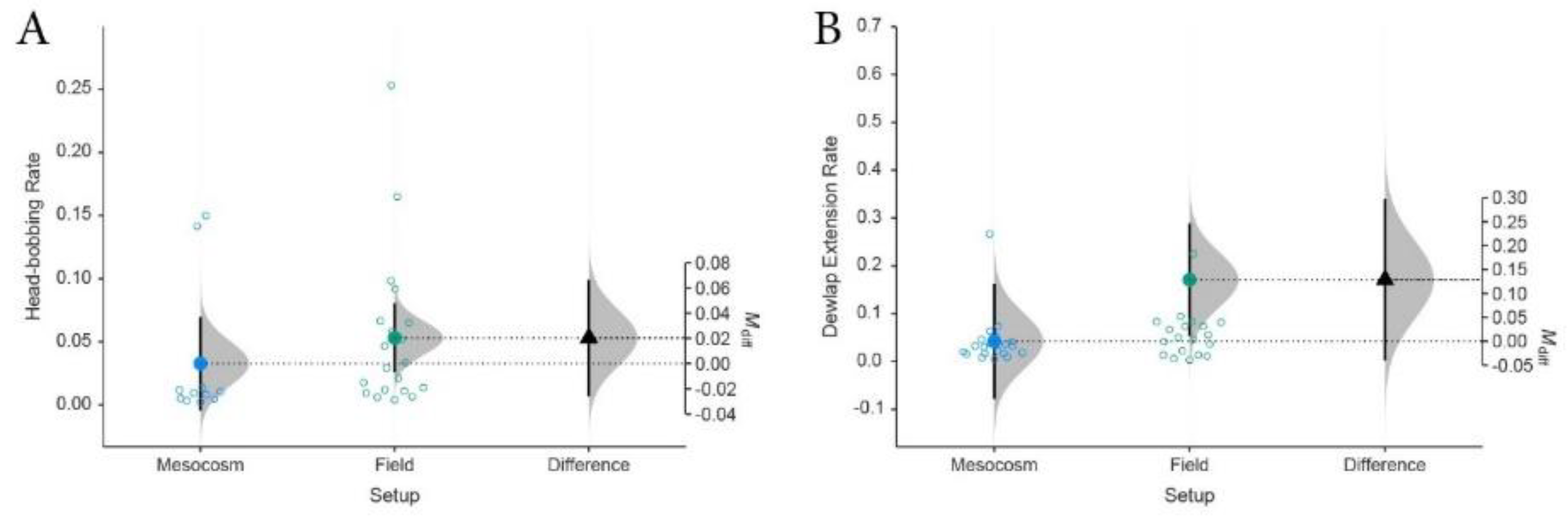

Table 1.
Datamining results, exhibiting data collected by species, population source environment, sex of subjects, invasive status and experimental set up. Summed and organized based on behavior observed.
Table 1.
Datamining results, exhibiting data collected by species, population source environment, sex of subjects, invasive status and experimental set up. Summed and organized based on behavior observed.
| Category | Factor | Total | A. carolinensis | A. sagrei | |||||||||
| (N) | DE | HB | PU | N | DE | HB | PU | N | DE | HB | PU | ||
| Species | AC | 26 | 16 | 7 | 3 | ||||||||
| AS | 88 | 42 | 31 | 15 | |||||||||
| Environment | Non-Urban | 36 | 18 | 12 | 16 | 4 | 2 | 2 | ND | 32 | 16 | 10 | 6 |
| Urban | 78 | 40 | 26 | 12 | 22 | 14 | 5 | 3 | 56 | 26 | 21 | 9 | |
| Sex | Female | 15 | 7 | 5 | 3 | 7 | 5 | 2 | ND | 8 | 2 | 3 | 3 |
| Male | 99 | 51 | 33 | 15 | 19 | 11 | 5 | 3 | 80 | 40 | 28 | 12 | |
| Invasive status | Invasive | 65 | 31 | 24 | 10 | ND | ND | ND | ND | 65 | 31 | 24 | 10 |
| Native | 49 | 27 | 14 | 8 | 26 | 16 | 7 | 3 | 23 | 11 | 7 | 5 | |
| Experimental Setup |
Field | 49 | 22 | 20 | 7 | 1 | 1 | ND | ND | 48 | 21 | 20 | 7 |
| Mesocosm | 65 | 36 | 18 | 11 | 25 | 15 | 7 | 3 | 40 | 21 | 11 | 8 | |
| Abbreviations: N- Data collected, DE- Dewlap Extensions, HB – Head-Bobs, PU- Pushups, ND – No Data, AC- Anolis carolinensis, AS – Anolis sagrei | |||||||||||||
Table 2.
Data collected by publication designated by species, sex, behavior types and the number of data mined from each publication (correlated with S1 – alphabetical bibliography).
Table 2.
Data collected by publication designated by species, sex, behavior types and the number of data mined from each publication (correlated with S1 – alphabetical bibliography).
| Publication | Species |
Sex |
Display | |||
|---|---|---|---|---|---|---|
| DE | HB | PU | # of data | |||
| Calsbeek & Marnocha, 2006* | AS | M | √ | √ | 4 | |
| Cox et al., 2009* | AS | M | √ | 2 | ||
| Decourcy & Jenssen, 1994 | AC | M | √ | 2 | ||
| Driessens et al., 2014* | AS | Both | √ | √ | 10 | |
| Edwards & Lailvaux, 2012 | AS | M | √ | √ | 4 | |
| Farrell et al., 2016 | AC | M | √ | √ | 4 | |
| Johnson & Wade, 2010* | AC | M | √ | 1 | ||
| Magaña, 2017*‡ | Both | Both | √ | √ | 19 | |
| McMann & Paterson, 2003 | AS | M | √ | 1 | ||
| McMann & Patterson, 2012 | AS | M | √ | √ | 4 | |
| Orrell, 2002‡ | AC | Both | √ | 6 | ||
| Partan et al., 2011* | AS | Both | √ | √ | √ | 6 |
| Paterson, 1999‡ | AS | M | √ | √ | 10 | |
| Paterson, 2002 | AS | M | √ | √ | 6 | |
| Simon, 2002*‡ | AS | M | √ | √ | √ | 10 |
| Simon, 2007* | AS | M | √ | √ | √ | 3 |
| Simon, 2011* | AS | M | √ | √ | 8 | |
| Stroud et al., 2019 | AS | M | √ | √ | 4 | |
| Tokarz & Beck, 1987 | Both | M | √ | 3 | ||
| Tokarz et al., 2002* | AS | M | √ | 1 | ||
| Tokarz et al., 2003 | AS | M | √ | √ | 3 | |
| Tokarz et al., 2005 | AS | M | √ | 1 | ||
| Yang & Wilczynski, 2002* | AS | M | √ | √ | 2 | |
| Total | 111 | |||||
Abbreviations: DE- dewlap extension, HB – head-bob, PU – pushup, * - data estimated from figures, ‡ - signifies a thesis, AS – Anolis sagrei, AC – Anolis carolinensis, M- Male.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



