
Submitted:
25 June 2024
Posted:
26 June 2024
You are already at the latest version
Abstract
the close phylogenetic relationship between ascidians (Tunicata) and vertebrates makes them a powerful model for studying the innate immune system. To better understand the nature and dynamics of immune responses and the mechanisms through which bacterial infections are detected and translated into inflammation in Ciona robusta, we applied an approach combining in vivo lipopolysaccharide (LPS) stimulation, immune-labelling and functional enzymatic analyses. The immunohistochemistry showed that toll-like receptor 4 (TLR4) and nuclear factor kappa B (NFκB) were expressed during the inflammatory pharynx response 4 hours post-LPS, with formation of immune-stained haemocyte nodules in the vessel lumen. Pharynx vessel endothelium also took part in the inflammatory response. Observations of histological sections from naive and MS-inoculated ascidians confirmed immuno-positive tissue response. Enzyme immune parameters – which included the activity of phenoloxidase, glutathione peroxidase, lysozyme, alkaline phosphatase and esterase – showed up-modulation 4 hours after LPS injection, confirming their involvement during ascidian inflammatory response. These findings provide new insights into the mechanisms involved in LPS-induced C. robusta response and suggest that a broad innate immune mechanism is involved in the regulation of tunicate inflammatory responses.
Keywords:
ascidian
; LPS
; inflammation
; innate immunity
; Ciona robusta
1. Introduction
Tunicates (urochordates) are generally considered to be the phylogenetically closest living relatives of vertebrates [1,2]. These organisms form a heterogeneous group spread across different marine habitats and with varied lifestyles (colonial or solitary, benthic or pelagic). Among these, the tunicate Ciona robusta, is a non-colonial ascidian that lives mainly in clusters fixed in natural and artificial substrates. C. robusta is widely considered a powerful model for studying embryological development and innate immunity, and it possesses a high degree of genetic complexity, with protein homologues involved in vertebrate immunity being described for this organism in recent decades [3,4].
Like other filter-feeding invertebrates, this species lives in environments full of infectious agents (e.g., viruses, bacteria and fungi), and are characterized by a pharynx that has respiratory and nutritional functions as well as serving as the main immune organ [4]. C. robusta have elaborated obligatory relationships with harmful agents from the microbiome that can activate defence reactions, and they possess the capacity for self/non-self-recognition [4]. Given that, one of the primary functions of the tunicate innate immune system is to recognize specific patterns of non-self-entities [4,5]. These patterns are called pathogen-associated molecular patterns (PAMPs), or microbe-associated molecular patterns (MAMPs), such as lipopolysaccharide (LPS), peptidoglycan, and mannan components of the microbial cell wall or nucleic acids. These and other PAMPs are recognized by pattern recognition receptors (PRRs), which are proteins that include both membrane-bound proteins, such as toll-like receptors (TLRs), and soluble proteins, such as lectins [3,4,6].
Several TLR genes with “hybrid” functionality have been identified in the genus Ciona [6,7], reflecting mammalian TLRs, and are generally divided into two subfamilies that mainly recognize related PAMPs: TLR1, TLR2, TLR4 and TLR6 recognize lipids, while TLR3, TLR7, TLR8 and TLR9 recognize nucleic acids [8,9]. The binding of PAMPs to TLRs activates cellular signalling cascades through myeloid differentiation primary response 88 protein (MyD88), which in turn binds members of the IL-1R-associated kinase (IRAK) family, thus leading to the activation of nuclear factor kappa B (NFκB) [10,11,12,13,14]. NFκB proteins, in turn, lead to the expression of proinflammatory cytokines (e.g., tumour necrosis factor α and interferons) [8], and are stored/sequestered in the cytoplasm by members of the κB family [13,14]. In vertebrates, two main signalling pathways are involved in NFκB activation: the canonical and non-canonical pathways, both involved in the regulation of immune and inflammatory responses [15,16,17]. In the canonical pathway, NFκB is bound to IκB in the cytoplasm without stimuli, with transcription constantly inactive. Upon stimulation, a protein kinase complex (IKK) (consisting of IKKα, IKKβ and IKKγ) is activated, leading to the degradation of the complex and allowing the translocation of NFκB to the nucleus [18,19]. For example, a gene encoding the respective proteins NfκB, IκB and Ikk has been well described in C. robusta [20,21]. The non-canonical activation of NFκB does not require the degradation of IκBα and has yet to be identified in invertebrate organisms [17].
Tunicates utilize phenoloxidase (PO) for melanin biosynthesis like other invertebrates, with the enzymatic activity participating in inflammatory processes, wound healing, sclerotization, pigmentation, and defence [4,22,23]. For example, ascidian haemocytes contain a proenzyme (proPO) which is activated by PO through the serine protease cascade, which is in turn activated by PRRs after their binding to PAMPs [22,24,25]. Two CrPO genes (CrPO-1 and CrPO-2) have already been isolated in C. robusta [22,24,25,26], and transcriptional activity in circulating haemocytes was observed as the effect of bacterial LPS inoculation [27]. Inflammatory responses also produce cytotoxic radicals, such as reactive oxygen species (ROS), which can lead to oxidative stress and cause tissue damage. Antioxidants, which readily scavenge oxygen radicals, are critical enzymes involved in functions related to cell immunity and phagocytosis during pathogen infection [28,29]. These include glutathione peroxidase (GPx), catalase, superoxide dismutase and fluorescent proteins.
Lysozyme (LYS) is a ubiquitous bacteriolytic enzyme produced by diverse groups of organisms, ranging from bacteria and bacteriophages to fungi, plants and animals [30]. It corresponds to the primary and rapid defence of organisms against attacks by pathogens and is a bactericidal hydrolytic enzyme which hydrolyses the β-1,4 glycosidic bonds of the bacterial cell wall, destabilizing the membrane [31]. Recently, a C. robusta g-type LYS, containing a GEWL domain with a GXXQ signature, was identified and characterized in various tunicate tissues [32]. In addition, the activity and kinetic characteristics of several metabolic enzymes, closely linked to immunity, are essential for maintaining invertebrate homeostasis following inflammatory activation [33,34,35]. Alkaline phosphatase (ALP) and esterase (EST) are examples of enzymes involved in a wide range of processes involving synthesis and hydrolysis reactions, as well as in various catabolic pathways in invertebrates [36,37,38].
Although the gene expressions involved in the immune response in C. robusta are well known (e.g., TLR, NFκB and PO), much remains to be understood about the wide-ranging nature and dynamics of immune activities in this ascidian during LPS exposure in vivo. In the present study, using an in vivo LPS-injection strategy, immuno-labelling techniques on pharynx tissues, and enzyme activity readout, we investigated the response of C. robusta to LPS challenge. This combined approach provided valuable additional indications about the involvement of the TLR-NFκB dependent pathway during the activation of inflammatory response following LPS injection. Furthermore, the functional activities of PO, GPX, LYS, ALP and EST enzymes were analysed for the first time in this ascidian species in LPS-mediated inflammatory response.
2. Materials and Methods
2.1. Ascidian Collection and Experimental Design
30 adult ascidians were gathered from the Palermo harbour (Sicily, Italy), maintained in tanks with filtered seawater at 18°C and fed with a marine invertebrate diet (Coraliquid, Sera Heinsberg, Germany). For the experimental plan, 10 control animals not subjected to injection were randomly sampled from the tanks and tissues were fixed for the histological experiments or snap frozen and stored at -30°C for tissue extraction. Then, 10 randomly-chosen organisms were inoculated with LPS of Escherichia coli or marine solution (MS), and 4h post injection were immediately fixed or stored at -30°C for histological experiments and tissue extraction, respectively. LPS of E. coli (ATCC 25922 strain; Chrisope Technologies, Louisiana, USA) was resuspended in MS (12 mM CaCl2.6H2O, 11 mM KCl, 26 mM MgCl2.6H2O, 43 mM TRIS HCl, 0.4 M NaCl, pH 8.0). LPS (100 µg in 100 µl MS per animal) or MS was inoculated into the body wall under the tunic at the median body wall region [4,39,40].
2.2. Immunohistochemistry
Before carrying out the immunohistochemical analyses, the antibody specificities against the selected target proteins in ascidian tissues were determined by checking the alignments between the deposited sequences of C. robusta (National Center for Biotechnology Information and UniProt databases) with the relative antibody epitopes (Fig. S1; Table S1) [34,41,42].
Body wall fragments of ascidians containing pharynx alone or both tunic and pharynx were excised at the injection site. Tissues were fixed in 4% paraformaldehyde in PBS-buffer solution (NaCl 137mM, KH2HPO4 10mM, KH2HPO4 2mM, KCl 2.7 mM, pH 7.6) at 4°C for 24 h. After dehydration in ethanol, animals were embedded in paraffin (Bio-Optica, Italy). Histological sections (7 μm thick) were cut with a rotary automatic microtome (Leica Microsystems HM350S, Wetzlar, Germany). For the immunohistochemistry assays, dewaxed sections were incubated in a blocking buffer (PBS containing 5% BSA and 1% Tween-20) for 2 h at room temperature, and then with the following primary antibodies diluted in blocking solution (PBS containing 1% BSA and 1% Tween-20): polyclonal anti-TLR4 produced in rabbit (SAB5700684, Sigma-Aldrich) (1:200); polyclonal anti-NFκB produced in rabbit (SAB4501989, Sigma-Aldrich) (1:200) overnight at 4°C. Before incubation, all slides were washed with the secondary antibodies for 90 min at room temperature. The secondary antibodies (goat anti-rabbit IgG-alkaline phosphatase; A3812, Sigma-Aldrich) were diluted 1:50 in PBS containing 1% BSA and 1% Tween-20 and incubated for 90 min. The sections were rinsed with the washing buffer (PBS containing 1% Tween-20) and stained with the BCIP/NBT chromogen substrate (Sigma-Aldrich). In all control experiments, primary antibodies were omitted, and sections were incubated only with the secondary antibodies. Slides were analysed using a light microscope (Leica DM750), and images were obtained using an ORMA-Eurotek MDH5 scientific camera (Milan, Italy). The quantification of the immune-positive stained areas in pharynx vessels (percentage of stained cells) on 6 randomly chosen fields (45000 μm2) for each slide were carried out using Image J software.
2.3. Extract Preparation and Protein Concentration
After removing the tunic from the specimens (3-5 cm length), the entire bodies were homogenized into polycarbonate tubes with 500 µl MS-buffer under ice; the resultant tissue slurry was then centrifuged (36,200 x g for 20 min at 4°C). The supernatant was collected and the protein concentration was measured according to the Bradford method [43]. Sample absorbance was read at 595 nm (RAYTO RT-2100C) with MS as a blank, and a calibration curve defined through bovine serum albumin was used to obtain the protein concentration, expressed in mg/ml. Extracts were adjusted to 0.5 mg/ml before performing enzymatic assays.
2.4. Phenoloxidase (PO)
PO activity was measured spectrophotometrically according to the Winder and Harris method [45], using L-Dopa (3,4 dihydroxy-L-phenylalanine; Sigma-Aldrich, USA) as a substrate and MBTH (3-methyl-2 benzothiazolinone hydrazone hydrochloride; Sigma-Aldrich, USA) as a specific reagent. 50 μl of sample with 50 μl of trypsin from bovine pancreas (1 mg/ml; Sigma-Aldrich, USA) or 50 μl of distilled water, as control, were incubated for 20 min at 20°C in 50 μl reaction mixture (5 mM L-DOPA and 20.7 mM MBTH in distilled water). Absorbance was read within 60 min at 5 min intervals by spectrophotometry at 505 nm (microplate reader, RAYTO RT-2100C). PO activity was expressed as units (U) per min, where 1 U = 0.001 ΔA540 min−1 mg−1 protein.
2.5. Glutathione Peroxidase (GPx)
Enzymatic activity was measured according to the Ross method [46]. In 96-well flat-bottomed plates, 50 µl of sample at a standard concentration (0.5 mg/ml) were incubated with 100 µl TMB (3,3′ 5,5′-tetramethylbenzidine; Sigma-Aldrich, USA). The reaction was stopped after 30 min of dark incubation with sulphuric acid (H2SO4) 2 M. The absorbance was read spectrophotometrically at 450 nm in a microplate reader (RAYTO RT-2100C), and the GPx produced was expressed in U/mg of protein according to the equation: U/mg = Abs * Vf / CP (Vf, final volume of the well; CP, protein concentration of the sample).
2.6. Lysozyme (LYS)
To evaluate LYS activity following the Parry method [47], 30 µl of each sample were placed in a 96-well flat-bottomed plate and incubated with 270 µl of bacterial suspension (Micrococcus lysodeikticus ATCC 4698, Sigma-Aldrich, USA) in triplicate. 30 µl MS buffer were replaced in the control sample. The reaction was carried out at 25°C, and absorbance (450 nm; microplate reader, RAYTO RT-2100C) was measured every 30 s for 10 min. A unit of LYS was defined as the amount of sample causing a decrease in absorbance of 0.001/min (U min-1), and U/ml was calculated in accordance with the formula: U/ml = (Δ abs/min-1 * dilution factor * 1000)/enzyme volume buffer.
2.7. Alkaline Phosphatase (ALP) and Esterase (EST)
For alkaline phosphatase (ALP), extracts were incubated in a 96-well flat-bottomed plate with an equal volume of 4 mM p-nitrophenyl phosphate substrate (Sigma-Aldrich, USA) liquid in 100 mM ammonium bicarbonate containing 1 mM MgCl2 (pH 7.8); esterase (EST) activity was evaluated by incubating the same volume of animal sample with 0.4 mM p-nitrophenyl myristate substrate (Sigma-Aldrich, USA) in 100 mM ammonium bicarbonate containing 0.5% of Triton X-100 (pH 7.8, 30°C; Sigma-Aldrich, USA). Enzymatic kinetics were evaluated according to the Ross method [46] at regular intervals of 5 min to 1 h at 405 nm with a microplate reader (RAYTO RT-2100C). One unit (U) of activity was defined as the amount of enzyme required to release 1 µmol of p-nitrophenol produced in 1 min.
2.8. Statistical Analyses
One-way ANOVAs were conducted in order to test differences among experimental groups. When significant differences were found, multiple comparisons using the Tukey post-hoc test were done to highlight differences between treatments. Statistical analyses were carried out using GraphPad software (Prism 8.0, San Diego, CA, USA). All experiments were performed in triplicate, and the values used were the mean ± standard deviation (SD) resulting from three independent experiments. Differences were considered significant for p < 0.05.
3. Results
3.1. TLR4 and NFκB Immunolocalization
The comparison of pharynx slides in sham ascidians revealed that very few cells expressed TLR4 (Figure 1A) or NFκB (Figure 1B) markers in the control ascidian pharynx vessels, and some positive cells were observed in haemocytes of MS-inoculated animals (Figure 1C, D). In addition, no positive expression of the examined proteins was observed in the endothelium of control ascidians. Conversely, 4h after inoculation with E. coli LPS, a large number of the vessels were densely populated with haemocytes expressing both proteins (Figure 1E, F). Quantification of the immune-positive cells (percentage) confirmed significant differences in the staining of both markers; the control and MS-inoculated ascidians showed from 8.8 to 23.4% of TLR4 stained cells and from 5.0 to 12.4% of NFκB stained cells, while significantly higher values were recorded for the LPS-inoculated ascidians compared to control and MS-inoculated specimens (74.8 and 46.5% of stained cells for TLR4 and NFκB, respectively) (Figure 1; Table 1). Immuno-positive haemocytes formed numerous large nodules inside the vessel lumen (Figure 1G, H). Positive staining was also observed in the endothelium, indicating variable expression of TLR4 and, consistently, NFκB markers within this tissue (Figure 1G, H). No staining was observed in pre-immune serum controls for either of the primary antibodies nor when the primary antibody was omitted. Figure 2
3.2. Enzymatic Response
Overall, the enzymatic activities measured in C. robusta whole-body extracts underwent a significant increase in ascidians 4 h after LPS inoculation (Figure 3; Table 2). In detail, PO activity increased in animals belonging to the MS and LPS groups with respect to controls, and even more in specimens injected with E. coli (Figure 3). However, multiple comparison analysis showed significant higher values only for ascidians injected with LPS compared to controls and MS-injected organisms (Figure 3; Table 2). The GPx trend among experimental groups was similar to PO (Figure 3), with significant higher values for the LPS-injected animals (Table 2). Regarding LYS, the one-way ANOVA evidenced significant difference between treatments (Table 2). Overall, activity was higher in the injected animals than in the control group (Figure 3). The highest LYS activity was detected in the LPS group. Also in this case, Tukey's test showed significant differences for ascidians injected with E. coli. Elevated ALP and EST activity was observed, with values 2-fold greater than in control and MS-injected animals (Figure 3). Statistical analysis revealed highly significant differences only for animals challenged with LPS compared to the other treatments, for both enzymes (Figure 3; Table 2).
4. Discussion
This study confirms that the innate immune signalling pathway activated by LPS in C. robusta is evolutionarily conserved and involves TLR-NFκB activities, in agreement with previous observations that highlighted the activation of this key immune pathway against invading pathogens and other potential threats to an ascidian host [6,7]. This is not surprising, given their key phylogenetic position in chordate evolution, generally considered a sister group of vertebrates [1,2,48,49]. In our C. robusta model, the NFκB signalling pathway appears to have been activated in pharynx vessels as a defence response against the bacterial LPS stimuli, through the involvement of TLR receptors. The upregulation of TLR4 was found at 4 h post-inoculation, when TLR4-producing haemocytes densely populated the lumen of the pharynx vessels. In addition, numerous nodules were formed in the vessels by TLR4-producing haemocytes, giving a distinctive inflammatory signature to the vessels. Consistently, an increase in circulating haemocytes expressing the key immune protein NFκB was also shown 4 h after LPS challenge, indicating the probable activation of a TLR-NFκB dependent pathway.
Nodules in the ascidian vessels were made up of tightly packed cells and were often connected or closely associated with the inner surface of the endothelium. These cells, containing TLR4 and NFκB transcripts in membranes and nuclei/cytoplasm, respectively, could be retained as activated cells engaged in inflammatory response in both the pharynx and haemolymph. The vascular endothelium was also involved in the response; although no continuous staining was observed in the endothelial tissue, several cells expressed the two proteins. Since several cells were shown to be maintained as proliferating cells, endothelium-associated haematopoietic nodules could develop following LPS stimulation [50,51,52,53]. In fact, there is the possibility that nodular stem cells differentiate into cell lines that circulate in the haemolymph and are recruited to inflamed sites [5,50]. Our results showed that the pharyngeal tissues of C. robusta can be stimulated by an LPS response and that they participate in immunity through vascular endothelium and nodules potentially acting as inflammatory haemocytes. This ascidian LPS-induced inflammatory response was also supported by the lack of pharynx inflammation observed in sham-injected animals inoculated with MS.
Overall, C. robusta tissue extracts showed a marked and significant upregulation of enzymatic activity following LPS injection. PO activity 4 h post-LPS injection resulted approximately two-fold higher compared to untreated ascidians. Upon activation by PAMP detection, the PO-cascade employs several compounds to hydroxylate monophenol and diphenol substrates in melanin polymeric deposits, producing highly cytotoxic defences and barriers against foreign cells or molecules [4,22,23,54]. Comparable PO up-activation has already been documented in the haemocytes, body-wall and tunic extracts of C. robusta model species, showing its involvement in inflammatory responses following LPS challenge [22,23,54]. Based on their biochemical properties, several POs have been described among ascidian species, presumably related to various functional roles [4,22]. Although they show catecholate activity, differences in size, trypsin sensitivity, activating substances, and SDS chain sensitivity have been found between species [4,22]. Additionally, differences within the same species have been found; for example, in terms of different sizes and trypsin enzyme-sensitivity in granular haemocytes and morula cells [22,27]. Concurrent with the activation of the cytotoxic PO system, there was a corresponding heightened antioxidant activity of GPx, an enzyme that generally scavenges hydrogen peroxide [55]. The significant values greater than four-fold manifested 4 h post-LPS injection in challenged specimens, indicating this enzyme’s involvement in ascidian inflammatory response. This is consistent with the induction of oxidative stress conditions during the invertebrate immune response under LPS stimuli, as a result of the oxidative burst and as a product of the PO system’s activity [28,56].
LYS exhibited a clear and significant up-regulated activity in C. robusta extracts 4 h post-LPS challenge, three-fold greater than untreated animals. These findings further corroborate the involvement of LYS in innate immune defence and in bacterial intracellular digestion of this ascidian, a marine invertebrate that feeds by filtering seawater and which is often exposed to high concentrations of microorganisms [32]. For example, previous observations from our group showed that the spatial mRNA expression of g-type lysozymes in adult specimens of C. robusta was detected mainly in pharynx, stomach and intestine tissues from 1 to 4 h after LPS injection [32], which is consistent with the hypothesis that LYS is expressed predominantly in organ tissues exposed to the external environment or in haematopoietic tissues [57,58]. We also analysed ALP and EST activities 4 h post-LPS challenge. The results of the assays conducted on C. robusta wall-body extracts suggested a correlation between the inflammatory response and the modification of these enzymatic parameters. Involvement of ALP and EST in the innate immune response to LPS had already been observed in other marine invertebrates, such as cnidarians and annelids [34,35,59], as well as being considered among the most interesting markers during regenerative inflammatory processes [60,61].
5. Conclusions
In conclusion, our results provide convincing evidence of an involvement of the entire pharynx in the inflammatory response of C. robusta, manifesting 4 h after LPS challenge, confirming the stimulation of the TLR-NFκB-dependent pathway against pathogenic agents. Also, to our knowledge, this is the first study to provide functional indications regarding the activities of several enzymatic parameters in the innate immune response to LPS injection in wall-body extracts of ascidian animal models. To fill the knowledge gaps regarding the hierarchically organized set of molecular, cellular and organismal networks involved in immune interactions with pathogens and the subsequent evolution of immune responses, it is essential to implement new studies that use such a broad-based approach.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: Sequence alignments; Table S1: Sequence accession numbers.
Author Contributions
Conceptualization, L.B., M.C, and D.P..; formal analysis, L.B., C.L.C., M.D., and F.B.; data curation, L.B., C.L.C., M.D., and F.B.; validation, L.B., C.L.C., M.D., F.B., J.V., M.G.P., M.C., and D.P.; writing-original draft preparation, L.B.; writing-review and editing, L.B., J.V., M.G.P., M.C., and D.P.; supervision, M.G.P., M.C., and D.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Fund for University Research (FFR 2023) at the University of Palermo (M. Dara, D. Parrinello, M.G. Parisi, and M. Cammarata) and the National Biodiversity Future Center Project (NBFC), CN_00000033, D.D. MUR n. 1034-17/06/2022 grant of financing, CUP B73C22000790001 for support of this research.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author due to (specify the reason for the restriction).
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships.
References
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Delsuc, F.; Tsagkogeorga, G.; Lartillot, N.; Philippe, H. Additional molecular support for the new chordate phylogeny. Genesis 2008, 46, 592–604. [Google Scholar] [CrossRef]
- Vizzini, A. Gene expression and regulation of molecules involved in pharynx inflammatory response induced by LPS in Ciona intestinalis. Invertebrate Survival Journal 2017, 14, 119–128. [Google Scholar] [CrossRef]
- Longo, V.; Parrinello, D.; Longo, A.; Parisi, M.G.; Parrinello, N.; Colombo, P.; Cammarata, M. The conservation and diversity of ascidian cells and molecules involved in the inflammatory reaction: The Ciona robusta model. Fish & Shellfish Immunology 2021, 119, 384–396. [Google Scholar] [CrossRef]
- Parrinello, N.; Vizzini, A.; Salerno, G.; Sanfratello, M.A.; Cammarata, M.; Arizza, V.; Vazzana, M.; Parrinello, D. Inflamed adult pharynx tissues and swimming larva of Ciona intestinalis share Ci TNFα-producing cells. Cell and tissue research 2010, 341, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Arizza, V.; Bonura, A.; La Paglia, L.; Urso, A.; Pinsino, A.; Vizzini, A. Transcriptional and in silico analyses of MIF cytokine and TLR signalling interplay in the LPS inflammatory response of Ciona robusta. Scientific Reports 2020, 10, 11339. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, N.; Ogasawara, M.; Sekiguchi, T.; Kusumoto, S.; Satake, H. Toll-like receptors of the ascidian Ciona intestinalis: prototypes with hybrid functionalities of vertebrate Toll-like receptors. Journal of biological chemistry 2009, 284, 27336–27343. [Google Scholar] [CrossRef] [PubMed]
- Ozinsky, A.; Underhill, D.M.; Fontenot, J.D.; Hajjar, A.M.; Smith, K.D.; Wilson, C.B.; Schroeder, L.; Aderem, A. The repertoire for pattern recognition of pathogens by the innate immune system is defined by cooperation between toll-like receptors. Proceedings of the National Academy of Sciences 2000, 97, 13766–13771. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef]
- Muzio, M.; Ni, J.; Feng, P.; Dixit, V.M. IRAK (Pelle) family member IRAK-2 and MyD88 as proximal mediators of IL-1 signaling. Science 1997, 278, 1612–1615. [Google Scholar] [CrossRef]
- Wesche, H.; Henzel, W.J.; Shillinglaw, W.; Li, S.; Cao, Z. ; MyD88: an adapter that recruits IRAK to the IL-1 receptor complex. Immunity 1997, 7, 837–847. [Google Scholar] [CrossRef]
- Häcker, H.; Redecke, V.; Blagoev, B.; Kratchmarova, I.; Hsu, L.C.; Wang, G.G.; Kamps, M.P.; Raz, E.; Wagner, H.; Häcker, H.; Mann, M.; Karin, M. Specificity in Toll-like receptor signalling through distinct effector functions of TRAF3 and TRAF6. Nature 2006, 439, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Oeckinghaus, A.; Ghosh, S. The NF-κB family of transcription factors and its regulation. Cold Spring Harbor perspectives in biology 2009, 1, a000034. [Google Scholar] [CrossRef]
- Sun, S.C.; Chang, J.H.; Jin, J. Regulation of nuclear factor-κB in autoimmunity. Trends in immunology 2013, 34, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Beinke, S.; Ley, S.C. Functions of NF-κB1 and NF-κB2 in immune cell biology. Biochemical Journal 2004, 382, 393–409. [Google Scholar] [CrossRef] [PubMed]
- Vallabhapurapu, S.; Karin, M. Regulation and function of NF-κB transcription factors in the immune system. Annual review of immunology 2009, 27, 693–733. [Google Scholar] [CrossRef]
- Sun, S.C. Non-canonical NF-κB signaling pathway. Cell research 2011, 21, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.C.; Ley, S.C. New insights into NF-κB regulation and function. Trends in immunology 2008, 29, 469–478. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, S.C. NF-κB in inflammation and renal diseases. Cell Biosci 2015, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Shimada, M.; Satoh, N.; Yokosawa, H. Involvement of Rel/NF-κB in regulation of ascidian notochord formation. Development, growth & differentiation 2001, 43, 145–154. [Google Scholar] [CrossRef]
- Kawai, N.; Takahashi, H.; Nishida, H.; Yokosawa, H. Regulation of NF-κB/Rel by IκB is essential for ascidian notochord formation. Developmental biology 2005, 277, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Parrinello, N.; Arizza, V.; Chinnici, C.; Parrinello, D.; Cammarata, M. Phenoloxidases in ascidian hemocytes: characterization of the pro-phenoloxidase activating system. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology 2003, 135, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Cammarata, M.; Arizza, V.; Cianciolo, C.; Parrinello, D.; Vazzana, M.; Vizzini, A.; Salerno, G.; Parrinello, N. The prophenoloxidase system is activated during the tunic inflammatory reaction of Ciona intestinalis. Cell and Tissue Research 2008, 333, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Immesberger, A.; Burmester, T. Putative phenoloxidases in the tunicate Ciona intestinalis and the origin of the arthropod hemocyanin superfamily. Journal of Comparative Physiology B 2004, 174, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Amparyup, P.; Sutthangkul, J.; Charoensapsri, W.; Tassanakajon, A. Pattern recognition protein binds to lipopolysaccharide and β-1, 3-glucan and activates shrimp prophenoloxidase system. Journal of Biological Chemistry 2012, 287, 10060–10069. [Google Scholar] [CrossRef]
- Cammarata, M.; Parrinello, N. The ascidian prophenoloxidase activating system. Invertebrate Survival Journal 2009, 1 (Suppl)), S67–S76. [Google Scholar]
- Vizzini, A.; Parrinello, D.; Sanfratello, M.A.; Trapani, M.R.; Mangano, V.; Parrinello, N.; Cammarata, M. Upregulated transcription of phenoloxidase genes in the pharynx and endostyle of Ciona intestinalis in response to LPS. Journal of invertebrate pathology 2015, 126, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M. Free radicals in biology and medicine. Oxford university press 2015, USA. [Google Scholar]
- Lesser, M.P.; Bythell, J.C.; Gates, R.D.; Johnstone, R.W.; Hoegh-Guldberg, O. Are infectious diseases really killing corals? Alternative interpretations of the experimental and ecological data. Journal of experimental marine biology and ecology 2007, 346, 36–44. [Google Scholar] [CrossRef]
- Jollès, P.; Jollès, J. What's new in lysozyme research? Always a model system, today as yesterday. Molecular and cellular biochemistry 1984, 63, 165–189. [Google Scholar] [CrossRef]
- Li, H.; Parisi, M.G.; Toubiana, M.; Cammarata, M.; Roch, P. Lysozyme gene expression and hemocyte behaviour in the Mediterranean mussel, Mytilus galloprovincialis, after injection of various bacteria or temperature stresses. Fish & shellfish immunology 2008, 25, 143–152. [Google Scholar] [CrossRef]
- Di Falco, F.; Cammarata, M.; Vizzini, A. Molecular characterisation, evolution and expression analysis of g-type lysozymes in Ciona intestinalis. Developmental & Comparative Immunology 2017, 67, 457–463. [Google Scholar] [CrossRef]
- Parisi, M.G.; Lentini, A.; Cammarata, M. Seasonal changes in morpho-functional aspects of two Anemonia sulcata (Pennant, 1777) wild populations. Marine Biodiversity 2017, 47, 561–573. [Google Scholar] [CrossRef]
- La Corte, C.; Dara, M.; Bertini, F.; Parrinello, D.; Piazzese, D.; Parisi, M.G. Response of Sabella spallanzanii to multiple stressors. The combined effect of infection and copper sulphate. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2023, 263, 109475. [Google Scholar] [CrossRef]
- Bisanti, L.; La Corte, C.; Dara, M.; Bertini, F.; Parrinello, D.; Chemello, R.; Cammarata, M.; Parisi, M.G. How does warmer sea water change the sensitivity of a Mediterranean thermophilic coral after immune-stimulation? Coral Reefs 2024, 43, 137–150. [Google Scholar] [CrossRef]
- Stamatis, H.; Christakopoulos, P.; Kekos, D.; Macris, B.J.; Kolisis, F.N. Studies on the synthesis of short-chain geranyl esters catalysed by Fusarium oxysporum esterase in organic solvents. Journal of Molecular Catalysis B: Enzymatic 1998, 4, 229–236. [Google Scholar] [CrossRef]
- Copeland, R.A. Enzymes: a practical introduction to structure, mechanism, and data analysis; John Wiley & Sons, 2023. [Google Scholar]
- Lopes, D.B.; Fraga, L.P.; Fleuri, L.F.; Macedo, G.A. Lipase and esterase: to what extent can this classification be applied accurately? Food Science and Technology 2011, 31, 603–613. [Google Scholar] [CrossRef]
- Vizzini, A.; Pergolizzi, M.; Vazzana, M.; Salerno, G.; Di Sano, C.; Macaluso, P.; Arizza, V.; Parrinello, D.; Cammarata, M.; Parrinello, N. FACIT collagen (1α-chain) is expressed by hemocytes and epidermis during the inflammatory response of the ascidian Ciona intestinalis. Developmental & Comparative Immunology 2008, 32, 682–692. [Google Scholar] [CrossRef]
- Parrinello, N.; Vizzini, A.; Arizza, V.; Salerno, G.; Parrinello, D.; Cammarata, M.; Giaramita, F.T.; Vazzana, M. Enhanced expression of a cloned and sequenced Ciona intestinalis TNFα-like (CiTNFα) gene during the LPS-induced inflammatory response. Cell and tissue research 2008, 334, 305–317. [Google Scholar] [CrossRef]
- Parisi, M.G.; Baranzini, N.; Dara, D.; La Corte, C.; Vizioli, J.; Cammarata, M. AIF-1 and RNASET2 are involved in the inflammatory response in the Mediterranean mussel Mytilus galloprovincialis following Vibrio infection. Fish & Shellfish Immunology 2022, 127, 109–118. [Google Scholar] [CrossRef]
- Bisanti, L.; La Corte, C.; Dara, M.; Bertini, F.; Parisi, M.G.; Chemello, R.; Cammarata, M.; Parrinello, D. Global warming-related response after bacterial challenge in Astroides calycularis, a Mediterranean thermophilic coral. Scientific Reports 2024, 14, 8495. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical biochemistry 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Winder, A.J.; Harris, H. New assays for the tyrosine hydroxylase and dopa oxidase activities of tyrosinase. European Journal of Biochemistry 1991, 198, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Ross, N.W.; Firth, K.J.; Wang, A.; Burka, J.F.; Johnson, S.C. Changes in hydrolytic enzyme activities of naive Atlantic salmon Salmo salar skin mucus due to infection with the salmon louse Lepeophtheirus salmonis and cortisol implantation. Diseases of aquatic organisms 2000, 41, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Parry Jr, R.M.; Chandan, R.C.; Shahani, K.M. A rapid and sensitive assay of muramidase. Proceedings of the Society for Experimental Biology and Medicine 1965, 119, 384–386. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Swalla, B.J. Molecular phylogeny of the protochordates: chordate evolution. Canadian Journal of Zoology 2005, 83, 24–33. [Google Scholar] [CrossRef]
- Tsagkogeorga, G.; Turon, X.; Hopcroft, R.R.; Tilak, M.K.; Feldstein, T.; Shenkar, N.; Loya, Y.; Huchon, D.; Douzery, E.J.P.; Delsuc, F. An updated 18S rRNA phylogeny of tunicates based on mixture and secondary structure models. BMC evolutionary biology 2009, 9, 1–16. [Google Scholar] [CrossRef]
- Ermak, T.H. The renewing cell populations of ascidians. American Zoologist 1982, 22, 795–805. [Google Scholar] [CrossRef]
- Sawada, T.; Zhang, J.; Cooper, E.L. Sustained viability and profileration of hemocytes from the cultured pharynx of Styela clava. Marine Biology 1994, 119, 597–603. [Google Scholar] [CrossRef]
- Peddie, C.M.; Smith, V.J. “Lymphocyte-like” cells in ascidians: Precursors for vertebrate lymphocytes? Fish & Shellfish Immunology 1995, 5, 613–629. [Google Scholar] [CrossRef]
- Peddie, C.M.; Richest, A.C.; Smith, V.J. Proliferation of undifferentiated blood cells from the solitary ascidian, Ciona intestinalis in vitro. Developmental & Comparative Immunology 1995, 19, 377–387. [Google Scholar] [CrossRef]
- Trapani, M.R.; Sanfratello, M.A.; Mangano, V.; Parrinello, D.; Vizzini, A.; Cammarata, M. Phenoloxidases of different sizes are modulated by LPS inoculation into Ciona intestinalis tunic and pharynx. Invertebrate Survival Journal 2015, 12, 75–81. [Google Scholar]
- Nappi, A.J.; Ottaviani, E. Cytotoxicity and cytotoxic molecules in invertebrates. BioEssays 2000, 22, 469–480. [Google Scholar] [CrossRef]
- Sadd, B.M.; Schmid-Hempel, P. PERSPECTIVE: principles of ecological immunology. Evolutionary applications 2009, 2, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Hikima, J.I.; Minagawa, S.; Hirono, I.; Aoki, T. Molecular cloning, expression and evolution of the Japanese flounder goose-type lysozyme gene, and the lytic activity of its recombinant protein. Biochimica et Biophysica Acta (BBA)-Gene Structure and Expression 2001, 1520, 35–44. [Google Scholar] [CrossRef]
- Zheng, W.; Tian, C.; Chen, X. Molecular characterization of goose-type lysozyme homologue of large yellow croaker and its involvement in immune response induced by trivalent bacterial vaccine as an acute-phase protein. Immunology Letters 2007, 113, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Trapani, M.R.; Parisi, M.G.; Parrinello, D.; Sanfratello, M.A.; Benenati, G.; Palla, F.; Cammarata, M. Specific inflammatory response of Anemonia sulcata (Cnidaria) after bacterial injection causes tissue reaction and enzymatic activity alteration. Journal of invertebrate pathology 2016, 135, 15–21. [Google Scholar] [CrossRef] [PubMed]
- De Petrocellis, L.; Orlando, P.; Pierobon, P.; De Falco, M.; Ruggiero, A.M.; Stefano, G.S.; Tino, A.; Grippo, P. Kelletinin A, from the marine mollusc Buccinulum corneum, promotes differentiation in Hydra vulgaris. Research communications in molecular pathology and pharmacology 1999, 103, 17–28. [Google Scholar]
- Parisi, M.G.; Grimaldi, A.; Baranzini, N.; La Corte, C.; Dara, M.; Parrinello, D.; Cammarata, M. Mesoglea extracellular matrix reorganization during regenerative process in Anemonia viridis (Forskål, 1775). International Journal of Molecular Sciences 2021, 22, 5971. [Google Scholar] [CrossRef]
Figure 1.
Representative sections of C. robusta pharynx at 4h post-LPS inoculation showing immunohistochemistry with anti-TLR4 and anti-NFκB antibodies. (A, B) control ascidians (not injected); (C, D) sham-injected ascidians inoculated with MS; (E, F) pharynx vessels at 4 h post-LPS inoculation showing densely populated haemocytes and nodules (nd) marked by the anti-TLR4 and anti-NFκB antibodies, respectively; (G, H) magnification of marked haemocyte nodules and endothelium (end) in the vessels.
Figure 1.
Representative sections of C. robusta pharynx at 4h post-LPS inoculation showing immunohistochemistry with anti-TLR4 and anti-NFκB antibodies. (A, B) control ascidians (not injected); (C, D) sham-injected ascidians inoculated with MS; (E, F) pharynx vessels at 4 h post-LPS inoculation showing densely populated haemocytes and nodules (nd) marked by the anti-TLR4 and anti-NFκB antibodies, respectively; (G, H) magnification of marked haemocyte nodules and endothelium (end) in the vessels.
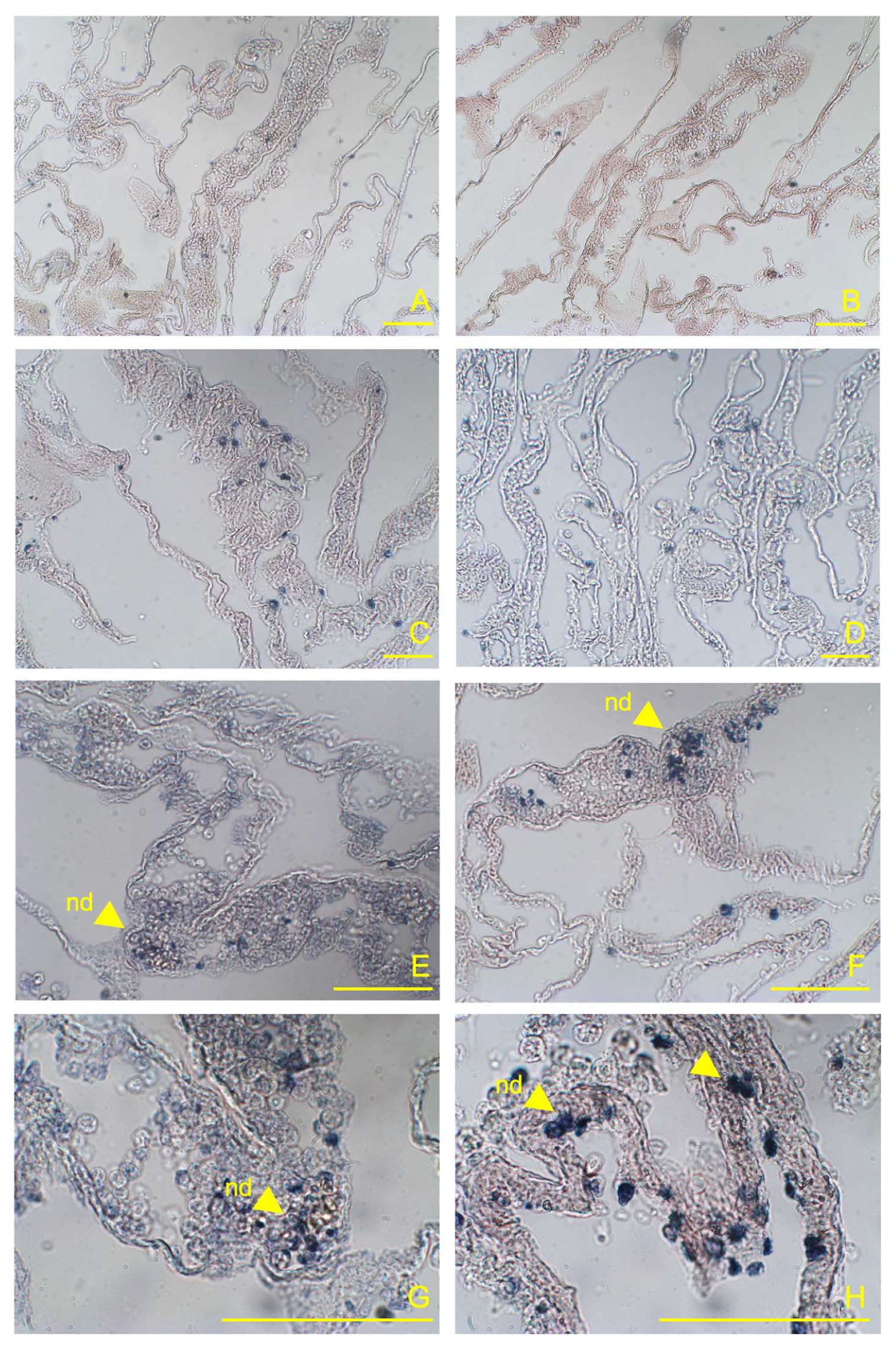
Figure 2.
Quantification of the immune-positive stained areas in pharynx vessels (percentage of stained cells; mean values ± SD) from slides belonging to experimental treatments. The letters indicate statistically significant differences (p < 0.05) between experimental groups.
Figure 2.
Quantification of the immune-positive stained areas in pharynx vessels (percentage of stained cells; mean values ± SD) from slides belonging to experimental treatments. The letters indicate statistically significant differences (p < 0.05) between experimental groups.
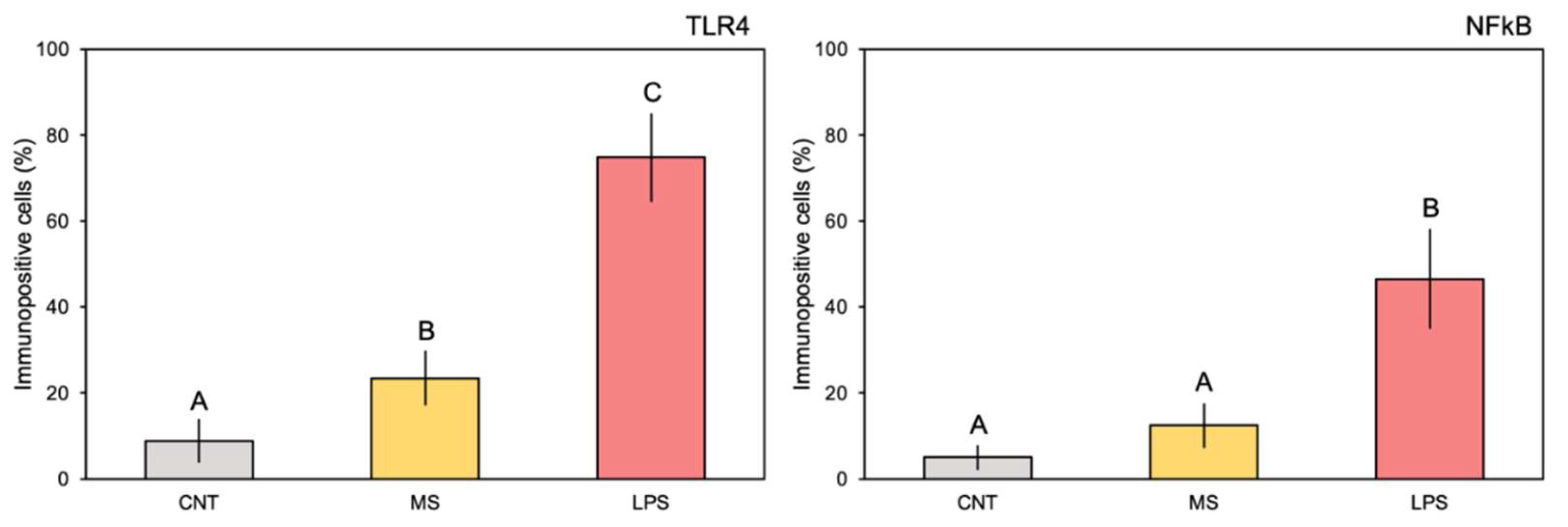
Figure 3.
Enzymatic response of phenoloxidase (PO), glutathione peroxidase (GPx), lysozyme (LYS), alkaline phosphatase (ALP) and esterase (EST) in ascidians at 4 h post inoculation with E. coli LPS. The letters indicate statistically significant differences (p < 0.05) between experimental groups.
Figure 3.
Enzymatic response of phenoloxidase (PO), glutathione peroxidase (GPx), lysozyme (LYS), alkaline phosphatase (ALP) and esterase (EST) in ascidians at 4 h post inoculation with E. coli LPS. The letters indicate statistically significant differences (p < 0.05) between experimental groups.
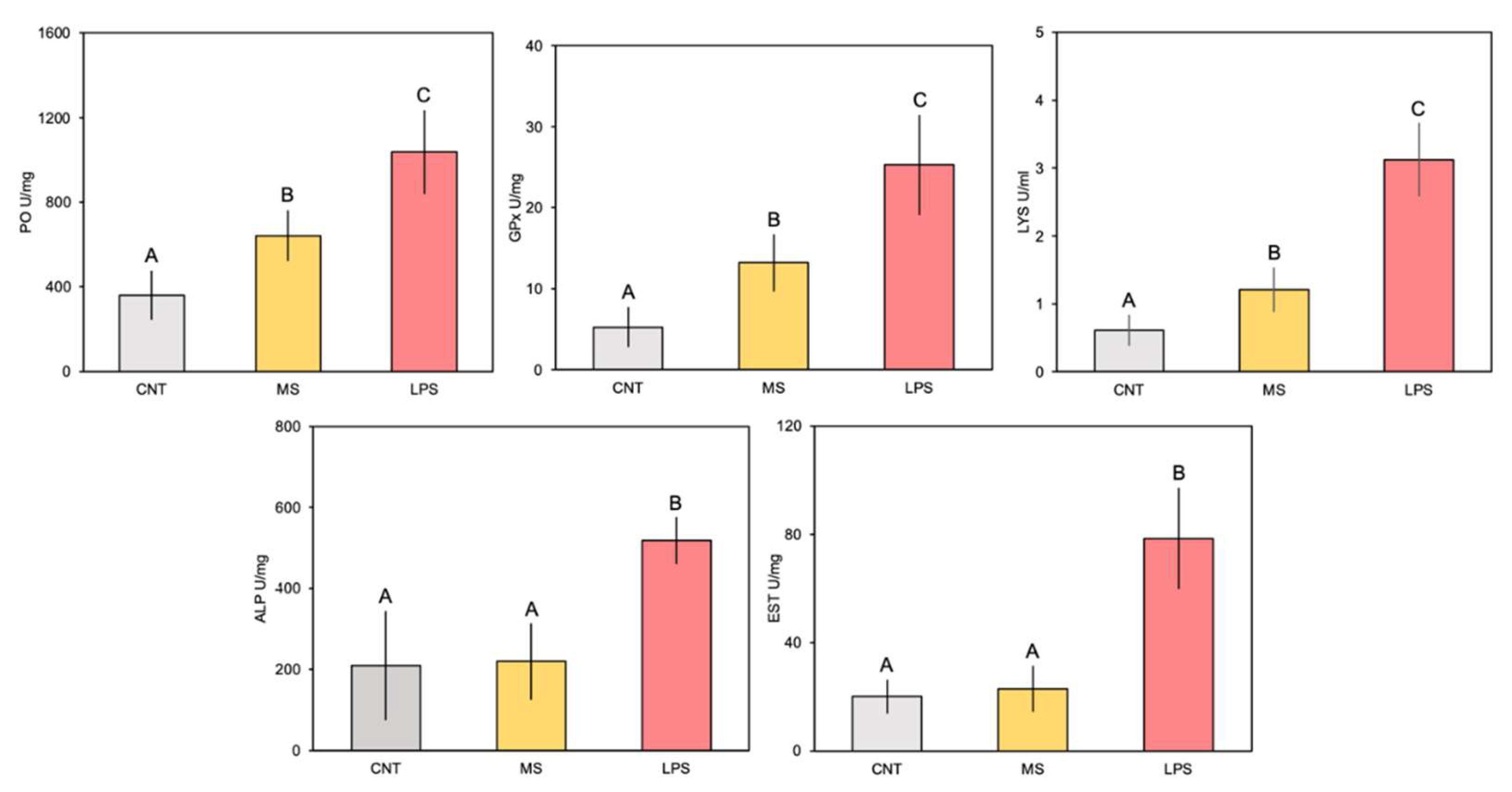

Table 1.
Summary of the one-way ANOVAs carried out on marker expression (TLR4 and NFκB) among experimental treatments on immune-positive stained areas in pharynx vessels (percentage of stained cells).
Table 1.
Summary of the one-way ANOVAs carried out on marker expression (TLR4 and NFκB) among experimental treatments on immune-positive stained areas in pharynx vessels (percentage of stained cells).
| Ordinary one-way ANOVA |
F | P value | R square |
|---|---|---|---|
| TLR4 | 170.00 | < 0.0001 | 0.95 |
| NFκB | 51.24 | < 0.0001 | 0.87 |
Table 2.
Summary of the ordinary one-way ANOVAs carried out on enzymatic activities among experimental groups.
Table 2.
Summary of the ordinary one-way ANOVAs carried out on enzymatic activities among experimental groups.
| Ordinary one-way ANOVA |
F | P value | R square |
|---|---|---|---|
| PO | 15.29 | 0.0044 | 0.83 |
| GPx | 16.08 | 0.0039 | 0.91 |
| LYS | 30.52 | 0.0007 | 0.91 |
| EST | 42.11 | < 0.0001 | 0.84 |
| ALP | 12.09 | 0.0028 | 0.72 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



