
Submitted:
25 June 2024
Posted:
26 June 2024
You are already at the latest version
Abstract
In the main prickly pear (Opuntia ficus-indica) producing region in the State of Mexico, fruit production (tunas) occupies the largest cultivated area with 15,800 ha, while 900 ha are cultivated for fresh consumption (“nopal” or “nopalitos”). Two composite samples consisting of prickly pear cactus cladodes (n=6) and another of nopal vegetable (n=6) showing virus-like symptoms were collected. Both sample sets were subjected to high-throughput sequencing (HTS) to identify the viruses and viroids present. The HTS results were verified using RT-PCR and Sanger sequencing. Subsequently, 86 nopal samples (nopal vegetable, prickly pear cactus, xoconostle and wild) were analyzed via RT-PCR with specific primers for the viruses and viroids previously detected via HTS. Three viruses were discovered [Opuntia virus 2 (OV2), cactus carlavirus 1 (CCV-1), and Opuntia potexvirus A (OPV-A)], along with a previously reported viroid [Opuntia viroid 1 (OPVd-1)]. Additionally, two new viroid species were identified, provisionally named Mexican opuntia viroid (genus Pospiviroid) and Opuntia viroid 2 (OVd-2). Phylogenetic analysis, pairwise identity comparison, and conserved structural elements analysis confirmed the classification of these two viroids as new species within the Pospiviroidae family. This is the first report of a pospiviroid and two apscaviroids infecting prickly pear in the world. Overall, this study enhances our understanding of the virome associated with nopal in Mexico.
Keywords:
viruses in opuntia
; Cactaceae
; new viroids
; Mexico
; Pospiviroidae
; diversity
1. Introduction
Opuntia ficus-indica (L.) Miller is native to Mexico [1] and is currently the most widely cultivated cactus in the world [2]. Globally, it is primarily cultivated as forage for livestock feed, for its fruit production, and for erosion control in desert regions [2,3]. In Mexico, in addition to its fruit, the immature cladodes are consumed as vegetables known as “nopalitos” [2,4]. This species also has applications in the food industry and serves as a host for Dactylopius coccus, which is used in the production of carminic acid, a dye employed in food, textile, and pharmaceutical products [2,5].
The State of Mexico is the leading producer of prickly pear cactus (nopal) in Mexico, with a cultivated area of approximately 17,500 ha. Of these, 15,800 ha are dedicated to the production of prickly pear fruit (tuna), 900 ha are dedicated to the production of nopal vegetable (nopalitos), and 800 ha are dedicated to the production of xoconostle nopal (Opuntia joconostle) [6]. The greatest production of nopal is concentrated in the eastern region of the State of Mexico, particularly in the municipalities of San Martín de las Pirámides, Axapusco, Otumba, Nopaltepec, Teotihuacan, Temascalapa, and Acolman [6].
High-throughput sequencing (HTS) or next-generation sequencing has significantly increased virus detection [7] and revolutionized the study of nucleic acids by allowing the sequencing of millions of nucleotides in a short period of time with very high redundancy (sequencing depth) [8]. When combined with specific bioinformatics tools, HTS can be used for the detection of known viruses as well as the discovery of new viruses or viroids [8,9]. One of the major advantages over classical detection techniques such as enzyme-linked immunosorbent assay (ELISA) and various PCR methods is that HTS does not require prior knowledge of viral genomic information, and it allows for the identification of the plant’s entire virome in a single assay [10].
Despite Mexico being the center of origin and domestication of O. ficus-indica, as well as many other species of the genus Opuntia [1], there are limited studies regarding virus detection in these plants. In studies using high-throughput sequencing for virus detection in prickly pear cacti conducted in Mexico [11,12], opuntia virus 2 (OV2, Tobamovirus genus), cactus carlavirus 1 (CCV-1, Carlavirus genus), and opuntia potexvirus A (OPV-A, Potexvirus genus) have been identified, along with a new species of viroid provisionally named Opuntia viroid 1 (OVd-1, Apscaviroid genus), the latter of which was found in nopal vegetable in Mexico City [12].
In a study conducted on various wild species of the genus Opuntia in Mexico to determine the presence of DNA genome viruses, squash leaf curl virus (SLCV, Begomovirus genus) and watermelon chlorotic stunt virus (WCSV, Begomovirus genus) were found [13]. Additionally, a new species of geminivirus tentatively named Opuntia virus 1 (OpV1) was detected in Opuntia santa-rita in the state of Sonora, Mexico [14]. Other viruses detected in prickly pear cacti in Mexico through RT-PCR include Schlumbergera virus X (SVX, Potexvirus genus) and rattail cactus necrosis-associated virus (RCNaV, Tobamovirus genus) [15,16]. To date, no study has been conducted to determine the viruses present in the State of Mexico, which is the main region where prickly pear fruit, vegetables, and xoconostle are produced in the country.
In this study, we utilized HTS to investigate the virome of both nopal vegetable and prickly pear cacti. Previously, employing this approach, a new virus and a new viroid species were detected in nopal vegetable [12]. In the present study, we leveraged the advantages of HTS technology to delve deeper and characterize the viruses and viroids in the primary production region of prickly pear cacti production regions in the State of Mexico. Our results identified various viruses and a previously reported viroid in nopal vegetable crops. Additionally, we detected two new viroid species, one provisionally named Mexican opuntia viroid (MOVd, Pospiviroid genus) in nopal vegetable, and another provisionally named Opuntia viroid 2 (OVd-2, Apscaviroid genus) in prickly pear cactus. This study sheds light on the diversity of viruses and viroids in Opuntia plants in Mexico.
2. Materials and Methods
2.1. High-Throughput Sequencing (HTS) and Bioinformatic Analysis
From June to December 2023, cladodes from prickly pear cactus (Opuntia ficus-indica) and nopal vegetable (Opuntia ficus-indica) with different putative virus-like symptoms were collected in Cuautlacingo, municipality of Otumba, and San Felipe Teotihuacán, municipality of Nopaltepec, located in the Eastern Region of the State of Mexico. Total RNA extraction from each sample was performed using the SV Total RNA Isolation System kit (Promega, USA) following the manufacturer’s instructions. The quality and quantity of RNA were verified using a NanoDrop spectrophotometer (Thermo Fisher Scientific), and RNA integrity was assessed by agarose gel electrophoresis (1%) and staining with ethidium bromide. Approximately 1.5 µg of total RNA was extracted from each collected symptom to form a composite sample (n=6) for nopal vegetable and a composite sample (n=6) for prickly pear cactus. The two composite samples were sent to Innomics Inc. (San Jose, California, USA) where the DNBSEQ Eukaryotic Strand-specific mRNA library was prepared, and sequencing was performed on the DNBSEQ platform (150 bp PE).
The sequences from each sample were de novo assembled using SPAdes v3.15.2 software with default parameters [17]. The obtained contigs were then subjected to a search in a local virus database (reference genomes, year 2023) using BLASTn and BLASTx. Subsequently, the reduced list of viral hits was compared against the complete nonredundant (nr) GenBank databases using BLASTn and BLASTx. For genome reconstruction, bowtie2 [18], SAMtools [19], and the Integrative Genomic Viewer (IGV) version 2.3 [20], were utilized. The assembly characteristics of the obtained genomes were evaluated with Qualimap 2 [21] and coverage and read depth maps were visualized and obtained through Geneious version 2024.0 (Biomatters). Finally, the genomes were annotated with the assistance of ORFinder (NCBI) and deposited into GenBank.
2.2. Genomic Analysis of New Viroid Species
For the detected viroids, a multiple sequence alignment was performed using ClustalW implemented in MEGAX [22], utilizing a representative sequence of opuntia viroid-2 (OVd-2, GenBank accession number: PP579661), Mexican opuntia viroid (MOVd, GenBank accession number: PP579660), and all RefSeq sequences of the Pospiviroidae family available in GenBank. A phylogenetic tree was constructed with 1000 bootstrap replicates using the maximum likelihood method and the Jukes-Cantor model in MEGAX [9,23]. The phylogenetic tree was visualized and edited using Interactive Tree of Life (iTOL) [24].
To compare the results with the taxonomic demarcation criteria approved by the International Committee on Taxonomy of Viruses (ICTV), pairwise sequence comparisons were performed using the Sequence Demarcation Tool (SDT) version 1.3 [25]. The pairwise identity graphs produced by SDT and the color-coded distance matrices were compared with the appropriate demarcation criteria to determine whether the viroid sequences found in the present study fell within the limits of existing species or should be considered new species.
The minimum free energy secondary structures for the two viroids were predicted using the UNAFold [26] web server (available online: www.mfold.org, accessed on January 5, 2024). The obtained secondary structures were further edited for printing using RnaViz 2 [27].
2.3. Genomic Analysis of the Detected Viruses
For the phylogenetic analyses of the viruses detected in the viral metagenomic analysis, sequences from representative members of the genus to which they belong were retrieved from GenBank. In all cases, multiple sequence alignment was performed using MAFFT [28], phylogenetic inference was conducted using the maximum likelihood method with IQ-tree 2 [29], and ModelFinder was used for evolutionary model selection [3]. Nodal support was estimated using ultrafast bootstrapping (UFBoot) (1000 replicates) [31], and phylogenetic trees were visualized and edited in iTOL [24].
2.4. Validation of the Detected Viruses and Viroids
To validate the viral metagenome of nopal, different primers (Table 1) were designed based on sequences obtained from HTS. For cDNA synthesis, 4 μL of the composite RNA sample analyzed by HTS, 0.5 μL of random primers (Promega, Madison, USA), and 5.5 μL of nuclease-free water were used. The reaction mixture was incubated at 70 °C for 5 min and then cooled on ice for one min. Subsequently, 5 μL of MMLV buffer (5X), 2.5 μL of dNTP mix (10 mM), 0.25 μL of M-MLV reverse transcriptase (200U/μL; Promega, Madison, USA), and 5 μL of nuclease-free water were added. The tubes were incubated at 37 °C for 60 min, followed by 72 °C for 15 min. The transcribed cDNA (1 µL) was used as a template for PCR in a mixture containing 2 μL of reaction buffer (5X Green GoTaq®), 0.6 μL of MgCl2 (25 mM), 0.2 μL of dNTPs mix (10 mM), 0.6 μL (10 μM) of each primer, 4.9 μL of nuclease-free water, and 0.1 μL of GoTaq® DNA Polymerase (5 U/μL; Promega, Madison, USA). PCR conditions consisted of an initial incubation at 95 °C for 3 min, followed by 35 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, extension at 72 °C for 40 s, and a final extension cycle at 72 °C for 10 min. PCR products were sequenced using the Sanger method (Macrogen Inc.) and the sequences were compared with those obtained by HTS.
2.5. Confirmation of Two New Viroid Species
The two genome drafts of each viroid (MOVd and OVd-2), assembled by HTS, were used to design primers that would allow the amplification of the entire genome by RT-PCR in two different reactions for each of them. For this purpose, forward and reverse primers (Table 1) were designed from the 5′ end to the 5′ end in opposite directions of the genome at two different positions to generate overlapping amplicons covering the entire genome of each viroid [9].
Reverse transcription was performed as previously described, using a portion of the total RNA that underwent HTS as the template. One microliter of cDNA was added to the PCR mixture as previously indicated. The amplification conditions for the four pairs of primers were as follows: initial incubation at 95 °C for 3 min, followed by 35 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 30 s; and a final extension cycle at 72 °C for 10 min. The PCR products were purified and bidirectionally sequenced using the Sanger method (Macrogen Inc.).
2.6. RT‒PCR Detection of Viruses and Viroids
From June to December 2023, a targeted sampling was conducted in the eastern region of the State of Mexico, one of the main prickly pear cacti producing areas in the country (Table 2). Seventy-five cladodes showing putative viral symptoms were collected (Figure 1): nopal vegetable (n=36), prickly pear nopal (n=35), and xoconostle (n=4). Seven asymptomatic nopal vegetable samples, two prickly pear nopal samples, and two wild prickly pear cactus samples were collected. Total RNA extraction was performed using the SV Total RNA Isolation System kit (Promega, Madison, USA), and RNA quality and quantity were verified using a NanoDrop spectrophotometer (Thermo Fisher Scientific). RT-PCR was carried out as previously described using specific primers for the detection of the three viruses and three viroids (Table 1).
3. Results
3.1. HTS Data
The libraries from the two selected composite samples of nopal vegetable and prickly pear cactus yielded 24,089,241 and 24,123,293 paired reads of 150 bp, respectively. The reads were clean and adapter-free, and subjeted to quality control by a sequencing service provider [32]. These RNA reads were deposited in the NCBI Sequence Read Archive and assigned the accession numbers SRX24164751 and SRX24164750 (BioProject Number PRJNA1096702). Metagenomic analysis revealed different viruses and viroids in the transcriptome of the nopal vegetable and prickly pear cactus, which are described below.
3.1.2. Known Viruses and Viroids that Infect Opuntia
The presence of three previously reported viruses and one viroid was detected and confirmed [11,12]. The identified viruses were OV2, CCV-1, and OPV-A. Following genome reconstruction, they were annotated for their coding regions and deposited in GenBank (Table 3). The viroid that was detected was OVd-I, and its genome was also deposited in GenBank (Table 3). In the nopal vegetable sample, OV2, CCV-1, OPV-A, and OVd-I were detected, while in the prickly pear cactus sample, OV2, CCV-1, and OPV-A were detected. The presence of these viruses and viroids was confirmed through RT-PCR and Sanger sequencing.
3.2. Phylogenetic Analysis and Genetic Diversity of the Detected Viruses
3.2.1. Opuntia Virus 2 (Genus Tobamovirus)
In the two samples subjected to HTS, the complete genomes of OV2 and both isolates [EM_T2 (PP579659) and EM_T1 (PP579656)] had the highest nucleotide identity (98.98%) with a previously obtained isolate from nopal vegetable (NC_040685.2) in Mexico 11.
The average nucleotide coverage of the OV2 genome was 12,492x for the EM_T2 isolate and 1,712x for the EM_T1 isolate (Figures S1 and S2).
Phylogenetic analysis grouped the two OV2 isolates obtained in this study into a clade with other genomes of this virus that are phylogenetically related to tobamoviruses that naturally infect cacti (Figure 2).
3.2.2. Cactus Carlavirus 1 (Genus Carlavirus)
In the samples of prickly pear cactus and nopal vegetable, CCV-1 was detected (designated as isolates EM_C1 and EM_C2, respectively). The average nucleotide coverage over the genome of the EM_C2 isolate was 187,006x, and that of the EM_C1 isolate was 3.52x (Figures S3 and S4). Considering a 95% nucleotide identity as a tentative cutoff to determine if it is a divergent isolate, the nopal vegetable l isolate [EM_C2 (PP579657)] is considered a divergent isolate with 92.14% nucleotide identity compared to the closest available genome in GenBank (Table 3).
Phylogenetic analysis grouped this isolate into a clade with other CCV-1 isolates (Figure 3). In the case of the EM_C1 isolate obtained from prickly pear cactus, it was not possible to recover the complete genome due to the low number of reads retrieved from the transcriptome (Figure S4); therefore, it was not included in the phylogenetic analysis.
3.2.3. Opuntia Potexvirus A (Genus Potexvirus)
OPV-A was detected in both the prickly pear cactus and nopal vegetable (designated as isolates EM_A1 and EM_A2, respectively). The nucleotide identity percentages were 85.50% for EM_A2 and 90.1% for EM_A1 compared to the single reference genome available in GenBank (OQ240443.1). Based on the nucleotide identity percentage, both OPV-A isolates are considered highly divergent new isolates.
The average nucleotide coverage of the EM_A2 isolate over the reference genome was 169,648x and 33,437x for the EM_A1 isolate (Figures S5 and S6).
Phylogenetic analysis grouped the two isolates from this study into a clade with the only OPV-A sequence available in GenBank (Figure 4).
3.2.4. Opuntia Viroid 1 (Putative Genus Apscaviroid)
3.3. Description of Two New Viroid Species
In addition to the mentioned viruses and viroid, two new viroids species were also detected (Table 4), one belonging to the genus Pospiviroid and the other to the genus Apscaviroid, based on the species demarcation criteria established for each genus [33].
3.3.1. Opuntia viroid 2 (Genus Apscaviroid)
Opuntia viroid 2 (OVd-2) was detected in the prickly pear cactus sample, with an average nucleotide coverage over the genome of 831x (Figure S8). The primary structure was obtained by direct sequencing of overlapping amplicons obtained by RT-PCR with primers 90F/89R and 208F/207R (Figure 5A, Table 1), which allowed determination of the circular nature of the RNA (Figure 5b). The consensus sequence was deposited in GenBank with the accession number PP579661.1. The OVd-2 genome consists of 319 nucleotides with a G+C content of 66.6%.
The secondary structure consists of a rod-shaped molecule containing a conserved central region (CCR) (Figure 5A), a key criterion for classifying a new viroid as a member of the family Pospiviroidae [23]. Considering the type of CCR and the presence of a conserved terminal region (TCR) (Figure 5A), OVd-2 is classified as a species of the genus Apscaviroid.
In this study, autonomous replication of OVd-2 was not demonstrated; however, the molecular characteristics of the viroid RNAs reported strongly support that it is a new viroid species. A phylogenetic analysis was conducted, which included the representative sequence of OVd-2 (EDMEX-S1, GenBank accession number PP579661) and all available reference sequences (RefSeq) in GenBank for current members belonging to the family Pospiviroidae (including an OVd-1 sequence) and the sequence of the Peach latent mosaic viroid (family Avsunviroidae) as an outgroup. OVd-2 clusted with current members of the genus Apscaviroid (Figure 6). Pairwise comparison-based SDT analysis also shows that none of the viroid sequences available in GenBank were more than 80% similar across the entire genome (Figure 7).
3.3.2. Mexican Opuntia Viroid (Genus Pospiviroid)
Mexican opuntia viroid (MOVd) was found only in the nopal vegetable sample. Its genome consists of 407 nucleotides with a G+C content of 68.1%. The average nucleotide coverage across the genome was 253x (Figure S9), and by directly sequencing the overlapping amplicons obtained by RT-PCR with the primer pairs 87F/86R and 307F/306R (Table 1, Figure 8A), the circular nature of this viroid RNA was determined (Figure 8B). The consensus sequence has been deposited in GenBank with the accession number PP579660.
The molecular structures of viroid RNAs previously reported and the results of the pairwise comparison-based SDT analysis in which none of the complete genome sequences of viroids available in GenBank had more than 90% similarity (Figure 7), demonstrated that MOVd is a new viroid. Additionally, the phylogenetic analysis (Figure 6) confirmed that MOVd should be considered a new species of the genus Pospiviroid.
3.4. Relative Abundance of Viruses and Viroids Detected by HTS in the Eastern Nopal-Producing Area of the State of Mexico
Eighty-six samples of cladodes with different symptoms associated with viruses collected in the municipalities of Otumba, Axapusco, Temascalapa, Nopaltepec, and Teotihuacan were analyzed for the detection of OV2, CCV-1, OPVA, OVd-1, OVd-2 and MOVd. OV2 was detected in 91% of the samples (37 prickly pear cactus, 39 nopal vegetable, two xoconostle prickly pear cactus, and one wild prickly pear cactus), followed by CCV-1 in 66% (8 prickly pear cactus, 43 nopal vegetable, and two wild prickly pear cactus), and finally OPVA in 50% of the positive samples (22 prickly pear cactus, 12 nopal vegetable, two xoconostle prickly pear cactus, and one wild prickly pear cactus). For viroids, OVd-2 was detected in 12 pear cactus samples samples, OVd-1 in five nopal vegetable and one prickly pear cactus sample, and MOVd in two nopal vegetable samples. Additionally, most of the analyzed samples showed mixed infections (Supplementary Table 1).
4. Discussion
In this study, we report on the viruses and viroids detected in the nopal vegetables and prickly pear cacti using HTS in the eastern region of the State of Mexico, Mexico. Two of these viroids are new species, one belonging to the Posviroid genus and the other to the Apscaviroid genus. In 2023, it was reported that OVd-1 (from the Apscaviroid genus) infected the nopal vegetable in Mexico, making it the first viroid reported in cacti worldwide. One of the criteria used to demarcate new viroid species is having less than 90% sequence identity. Therefore, OVd-2 should be considered a new species within the Apscaviroid genus, and MOVd should be considered a new species of the Pospiviroid genus.
Additionally, the conserved structural domains of MOVd are very similar to those of the reference genomes of Potato spindle tuber viroid (PSTVd) and Iresine viroid 1 (IrVd-1) (Pospiviroid genus); the CCR of MOVd is identical to that of IrVd-1, while there are slight modifications in the TCR and CCR compared to those of PSTVd. In the case of OVd-2, the conserved structural domains were identical to those in the reference genome of the apple scar skin viroid (ASSVd) (Apscaviroid genus).
Due to the presence of mixed infections of viruses and viroids in 82% of the samples analyzed individually by RT-PCR (Supplementary Table 1), it was not possible to exclusively associate the symptoms with either of them. There is limited understanding of the role of virus/viroid coexistence, warranting a study on this interaction [34]. In Mexico, at least three cases of mixed viroid infections have been reported: Mexican papita viroid (MPVd)/tomato chlorotic dwarf viroid and MPVd/tomato severe leaf curl virus in tomatoes under greenhouse conditions and hop stunt viroid/citrus exocortis viroid, citrus tristeza virus in oranges. The symptoms of plants with mixed infections were more severe than those plants with individual infections in all cases [34]. Therefore, experiments on pathogenicity need to be performed, along with studies investigating the distribution, transmission modes, and effects on the production and quality of cladodes and prickly pears caused by these new viroids and viruses in nopal cultivation.
The detection of these new viroids in prickly pear cultivation reinforces the theory that Mexico, being the center of origin and domestication of O. ficus-indica (the most cultivated cactus species in the world) and other species of the Opuntia genus [1], is a geographical region of viroid origin and possesses considerable biodiversity including endemic species that affect important crops such as tomatoes and high-value commercial products such as avocados [35,36]. The viroids detected in this study are among the most common viroids, such as the tomato bunchy top viroid, Mexican papita viroid, and avocado sunblotch viroid, which are considered endemic to Mexico [34].
Moreover, of the detected viruses, CCV-1 and OPV-A were the most divergent (Figure 3 and Figure 4), while the OV2 isolates consistently clustered with previously reported isolates, forming a clade with viruses that naturally infect cacti (Figure 6).
It is known that tobamoviruses, such as OV2, are easily transmitted mechanically [37], so this is an important aspect to consider during crop management since many cultural practices in both prickly pear cacti and nopal vegetable involve the use of tools for pruning and harvesting that are not disinfected between plants.
OPV-A was previously detected in the nopal vegetable in Mexico City, but its range of hosts and economic impact on prickly pear cultivation are still unknown.
CCV-1 was detected alongside cactus carlavirus 2 (CCV-2) for the first time in the United States, asymptomatically, in a plant of the genus Epiphyllum (hybrid ‘Professor Ebert’) [38]. In Mexico, CCV-1 was detected in 93 out of 129 nopal vegetable samples collected in the states of Morelos, the State of Mexico, Hidalgo, and Mexico City [12]. On the other hand, it is known that carlaviruses predominantly infect herbaceous plants; many cause latent or asymptomatic infections and are transmitted by aphids in a nonpersistent manner and by whiteflies [38]. However, none of these insect groups are significant pests of prickly pear, hence transmission is not yet associated with any vector insect.
The vegetative and mechanical transmission of CCV-1, OV2 and OPV-A can contribute to their distribution in new prickly pear plantations not only in the study area but also throughout the country as the State of Mexico is the main provider of prickly pear propagative material. Therefore, phytosanitary measures should be implemented to help reduce the risk of the spread and movement of infected material carrying viruses or viroids.
Previously, a study was conducted to understand the virome of the nopal vegetable in the central region of Mexico, which included the states of Morelos, Hidalgo, Mexico City, and the State of Mexico. However, the two new viroids species reported in the present study were not detected in the State of Mexico [12]. This suggests the need to consider more sampling sites and process a greater number of samples to generate more robust information. Additionally, in this study, we report for the first time the presence of OPV-A and OV2 in xoconostle prickly pear cacti and for the first time the presence of CCV-1 and OPV-A in prickly pear cacti.
Subsequent research on OVd-1, OVd-2, and MOVd should focus on understanding their host range, environmental conditions, or crop management practices that favor asymptomatic conditions, symptoms associated with individual and mixed infections, among other aspects. Although the main characteristic of viroids is their autonomous replication, their ability to be transmitted through grafting alone is not sufficient to demonstrate this feature [9]; therefore, mechanical or biolistic transmission tests are necessary. Additionally, further investigations into other aspects of their transmission, such as the potential presence of an insect vector, will be needed to deepen our understanding of these viroids.
In this study, we confirmed the potential of HTS to provide an ideal methodology for determining the complete infection status of a plant by viruses and viroids. This technology does not depend on the availability of genomic sequence information such as RT-PCR and is limited only by the integrity of the reference database against which the sequences are compared [9], as well as the correct use of different programs for bioinformatic analysis.
5. Conclusions
Using HTS, the presence of a new viroid of the Pospiviroid genus and a second Apscaviroid in prickly pear cultivation is reported for the first time. Additionally, Opuntia virus 2 was the most abundant in the study area. Finally, mixed infection was detected in 82% of the samples analyzed. To prevent the potential spread of these viruses and viroids in new prickly pear plantations, a program for producing virus- and viroid-free mother plants should be implemented to ensure the health of the propagative material used.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1. Read coverage over the genome of Opuntia virus 2, isolate EM_T2, obtained from nopal vegetable. Open reading frame (ORF) organization and annotation are shown in different colors. *Indicates an amber stop codon in the forward reading frame. Figure S2. Read coverage over the genome of Opuntia virus 2, isolate EM_T1, obtained from prickly pear cactus. Open reading frame (ORF) organization and annotation are shown in different colors. *Indicates an amber stop codon in the forward reading frame. Figure S3. Read coverage over the genome of Cactus carlavirus 1, isolate EM_C2, obtained from nopal vegetable. Open reading frame (ORF) organization and annotation are shown in different colors. TGB, triple gene block; CP, capsid protein; NB, nucleic acid binding protein. Figure S4. Read coverage over the genome of Cactus carlavirus 1, isolate EM_C1, obtained from prickly pear cactus. Open reading frame (ORF) organization and annotation are shown in different colors. TGB, triple gene block; CP, capsid protein; NB, nucleic acid binding protein. Figure S5. Read coverage over the genome of Opuntia potexvirus A, isolate EM_A2, obtained from nopal vegetable. Open reading frame (ORF) organization and annotation are shown in different colors. TGB, triple gene block. Figure S6. Read coverage over the genome of Opuntia potexvirus A, isolate EM_A1, obtained from prickly pear cactus. Open reading frame (ORF) organization and annotation are shown in different colors. TGB, triple gene block. Figure S7. Read coverage over the genome of Opuntia viroid 1, isolate EDMEX-V1, from nopal vegetable. Figure S8. Read coverage over the genome of Opuntia viroid 2, isolate EDMEX-T1, from nopal vegetable. Figure S9. Read coverage over the genome of Mexican opuntia viroid, isolate V1, from nopal vegetable. Supplementary Table 1. Detection of viruses and viroids by RT-PCR in different prickly pear cactus samples.
Author Contributions
“Conceptualization, C.O-A and D.L.O.-M.; methodology, C.O.-A., D.L.O.-M. and E.R-L.; formal analysis, C.O-A and D.L.O.-M.; investigation, C.O.-A., D.L.O.-M. and E.R-L.; resources D.L.O.-M. and E.R-L.; writing—original draft preparation, C.O-A.; writing—review and editing, C.O.-A., D.L.O.-M. and E.R.L; supervision, D.L.O.-M. and E.R-L.; project administration, E.R.L.; funding acquisition, D.L.O.-M. and E.R.L. All authors have read and agreed to the published version of the manuscript.”
Funding
This research was partially funded by the Postgraduate College Campus Montecillo, with resources and infrastructure from the institution, and by the Consejo Mexiquense de Ciencia y Tecnología (COMECyT) through the project: Propuesta de manejo fitosanitario de nopal y agave en el oriente del Estado de México, (FICDTEM-2023-156).
Data Availability Statement
The genomic sequences of the viruses and viroids described in this study are available in the GenBank database under the accession numbers PP579654-PP579661 and BioProject No. PRJNA1096702.
Acknowledgments
The first author would like to thank the Consejo Mexiquense de Ciencia y Tecnología (COMECyT) for their support of this research. To the Postgraduate College Campus Montecillo and the prickly pear producers in the eastern area of the State of Mexico, for sharing their experience and allowing access to their plots for sample collection.
Conflicts of Interest
“The authors declare no conflicts of interest.”
References
- Griffith, M.P. The Origins of an Important Cactus Crop, Opuntia Ficus-Indica (Cactaceae): New Molecular Evidence. Am. J. Bot. 2004, 91, 1915–1921. [Google Scholar] [CrossRef] [PubMed]
- Nobel, P.S. Cacti: Biology and Uses; University of California Press, 2002.
- Le Houérou, H.N. The Role of Cacti (Opuntiaspp.) in Erosion Control, Land Reclamation, Rehabilitation and Agricultural Development in the Mediterranean Basin. J. Arid Environ. 1996, 33, 135–159. [Google Scholar] [CrossRef]
- Flores-Valdez, C.A. “Nopalitos” Production, Preoceáing and Marketing.
- Rodríguez, L.C.; Faúndez, E.; Seymour, J.; Escobar, C.A.; Espinoza, L.; Petroutsa, M.; Ayres, A.; Niemeyer, H.M. Biotic Factors and Concentration of Carminic Acid in Cochineal Insects (Dactylopius Coccus Costa) (Homoptera: Dactylopiidae). Agric. Téc. 2005, 65, 323–329. [Google Scholar] [CrossRef]
- SIAP. Anuario Estadístico de la Producción Agrícola; SIAP. Mex. City Mex. 2023.
- Villamor, D.E.V.; Keller, K.E.; Martin, R.R.; Tzanetakis, I.E. Comparison of High Throughput Sequencing to Standard Protocols for Virus Detection in Berry Crops. Plant Dis. 2022, 106, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Villamor, D.E.V.; Ho, T.; Al Rwahnih, M.; Martin, R.R.; Tzanetakis, I.E. High Throughput Sequencing For Plant Virus Detection and Discovery. Phytopathology® 2019, 109, 716–725. [Google Scholar] [CrossRef]
- Bester, R.; Malan, S.S.; Maree, H.J. A Plum Marbling Conundrum: Identification of a New Viroid Associated with Marbling and Corky Flesh in Japanese Plums. Phytopathology® 2020, 110, 1476–1482. [Google Scholar] [CrossRef]
- Maree, H.J.; Fox, A.; Al Rwahnih, M.; Boonham, N.; Candresse, T. Application of HTS for Routine Plant Virus Diagnostics: State of the Art and Challenges. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Ortiz, H.; De La Torre-Almaraz, R.; Sanchez-Navarro, J.A.; Pallas, V. Identification and Genomic Characterization of a Novel Tobamovirus from Prickly Pear Cactus. Arch. Virol. 2020, 165, 781–784. [Google Scholar] [CrossRef]
- Ortega-Acosta, C.; Ochoa-Martínez, D.L.; Rojas-Martínez, R.I.; Nava-Díaz, C.; Valverde, R.A. Virome of the Vegetable Prickly Pear Cactus in the Central Zone of Mexico. Rev. Mex. Fitopatol. Mex. J. Phytopathol. 2023, 41. [Google Scholar] [CrossRef]
- Fontenele, R.S.; Bhaskara, A.; Cobb, I.N.; Majure, L.C.; Salywon, A.M.; Avalos-Calleros, J.A.; Argüello-Astorga, G.R.; Schmidlin, K.; Roumagnac, P.; Ribeiro, S.G.; Kraberger, S.; Martin, D.P.; Lefeuvre, P.; Varsani, A. Identification of the Begomoviruses Squash Leaf Curl Virus and Watermelon Chlorotic Stunt Virus in Various Plant Samples in North America. Viruses 2021, 13, 810. [Google Scholar] [CrossRef]
- Fontenele, R.S.; Salywon, A.M.; Majure, L.C.; Cobb, I.N.; Bhaskara, A.; Avalos-Calleros, J.A.; Argüello-Astorga, G.R.; Schmidlin, K.; Khalifeh, A.; Smith, K.; Schreck, J.; Lund, M.C.; Köhler, M.; Wojciechowski, M.F.; Hodgson, W.C.; Puente-Martinez, R.; Van Doorslaer, K.; Kumari, S.; Vernière, C.; Filloux, D.; Roumagnac, P.; Lefeuvre, P.; Ribeiro, S.G.; Kraberger, S.; Martin, D.P.; Varsani, A. A Novel Divergent Geminivirus Identified in Asymptomatic New World Cactaceae Plants. Viruses 2020, 12, 398. [Google Scholar] [CrossRef]
- De La Torre-Almaraz, R.; Salgado-Ortiz, H.; Salazar-Segura, M.; Pallas, V.; Sanchez-Navarro, J.A.; Valverde, R.A. First Report of Schlumbergera Virus X in Prickly Pear (Opuntia Focus-Indica) in Mexico. PLANT Dis. 2016, 100, 1799–1800. [Google Scholar] [CrossRef]
- De La Torre-Almaraz, R.; Salgado-Ortiz, H.; Salazar-Segura, M.; Pallas, V.; Sanchez-Navarro, J.A.; Valverde, R.A. First Report of Rattail Cactus Necrosis-Associated Virus in Prickly Pear Fruit (Opuntia Albicarpa Scheinvar) in Mexico. PLANT Dis. 2016, 100, 2339–2339. [Google Scholar] [CrossRef]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes De Novo Assembler. Curr. Protoc. Bioinforma. 2020, 70, e102. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-Performance Genomics Data Visualization and Exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Okonechnikov, K.; Conesa, A.; García-Alcalde, F. Qualimap 2: Advanced Multi-Sample Quality Control for High-Throughput Sequencing Data. Bioinformatics 2016, 32, 292–294. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Di Serio, F.; Owens, R.A.; Li, S.-F.; Matoušek, J.; Pallás, V.; Randles, J.W.; Sano, T.; Verhoeven, J. Th. J.; Vidalakis, G.; Flores, R.; Consortium, I.R. ICTV Virus Taxonomy Profile: Pospiviroidae. Journal of General Virology 2021, 102. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A Virus Classification Tool Based on Pairwise Sequence Alignment and Identity Calculation. PLOS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Zuker, M. Mfold Web Server for Nucleic Acid Folding and Hybridization Prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- De Rijk, P.; Wuyts, J.; De Wachter, R. RnaViz 2: An Improved Representation of RNA Secondary Structure. Bioinformatics 2003, 19, 299–300. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Shi, C.; Huang, Z.; Zhang, Y.; Li, S.; Li, Y.; Ye, J.; Yu, C.; Li, Z.; Zhang, X.; Wang, J.; Yang, H.; Fang, L.; Chen, Q. SOAPnuke: A MapReduce Acceleration-Supported Software for Integrated Quality Control and Preprocessing of High-Throughput Sequencing Data. GigaScience 2018, 7, gix120. [Google Scholar] [CrossRef]
- Di Serio, F.; Flores, R.; Verhoeven, J. Th. J.; Li, S.-F.; Pallás, V.; Randles, J.W.; Sano, T.; Vidalakis, G.; Owens, R.A. Current Status of Viroid Taxonomy. Arch. Virol. 2014, 159, 3467–3478. [Google Scholar] [CrossRef]
- Aviña-Padilla, K.; Zamora-Macorra, E.J.; Ochoa-Martínez, D.L.; Alcántar-Aguirre, F.C.; Hernández-Rosales, M.; Calderón-Zamora, L.; Hammond, R.W. Mexico: A Landscape of Viroid Origin and Epidemiological Relevance of Endemic Species. Cells 2022, 11, 3487. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Soriano, J.P.; Galindo-Alonso, J.; Maroon, C.J.; Yucel, I.; Smith, D.R.; Diener, T.O. Mexican Papita Viroid: Putative Ancestor of Crop Viroids. Proc. Natl. Acad. Sci. 1996, 93, 9397–9401. [Google Scholar] [CrossRef]
- Saucedo Carabez, J.R.; Téliz Ortiz, D.; Vallejo Pérez, M.R.; Beltrán Peña, H. The Avocado Sunblotch Viroid: An Invisible Foe of Avocado. Viruses 2019, 11, 491. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, L. Epidemiology and Control of Tomato Mosaic Virus. Annu. Rev. Phytopathol. 1976, 14, 75–96. [Google Scholar] [CrossRef]
- Peng, L.; Grinstead, S.; Kinard, G.; Wu, L.-P.; Li, R. Molecular Characterization and Detection of Two Carlaviruses Infecting Cactus. Arch. Virol. 2019, 164, 1873–1876. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Symptoms associated with virosis in different species of prickly pear cactus. (A-B), Chlorosis with green islands in nopal vegetable; (C), Irregular chlorotic patterns around the spines in nopal vegetable; (D), Mottling and chlorotic spots in fig prickly pear cactus. E, Ring spot symptoms in xoconostle cactus; (F) Symptomless nopal vegetable.
Figure 1.
Symptoms associated with virosis in different species of prickly pear cactus. (A-B), Chlorosis with green islands in nopal vegetable; (C), Irregular chlorotic patterns around the spines in nopal vegetable; (D), Mottling and chlorotic spots in fig prickly pear cactus. E, Ring spot symptoms in xoconostle cactus; (F) Symptomless nopal vegetable.

Figure 2.
A phylogenetic tree was constructed using the maximum likelihood method with complete genome sequences of the Tobamovirus genus obtained from GenBank (accession numbers shown). The tree was built with IQtree 2.3.1 using a color code to represent the different botanical families that are natural hosts of each virus. The identified OV2 sequences in this study are highlighted in bold. The sequence of soil-borne cereal mosaic virus (genus Furovirus) was used as an outgroup. Circles on branches indicate UFBoot support values > 70%.
Figure 2.
A phylogenetic tree was constructed using the maximum likelihood method with complete genome sequences of the Tobamovirus genus obtained from GenBank (accession numbers shown). The tree was built with IQtree 2.3.1 using a color code to represent the different botanical families that are natural hosts of each virus. The identified OV2 sequences in this study are highlighted in bold. The sequence of soil-borne cereal mosaic virus (genus Furovirus) was used as an outgroup. Circles on branches indicate UFBoot support values > 70%.
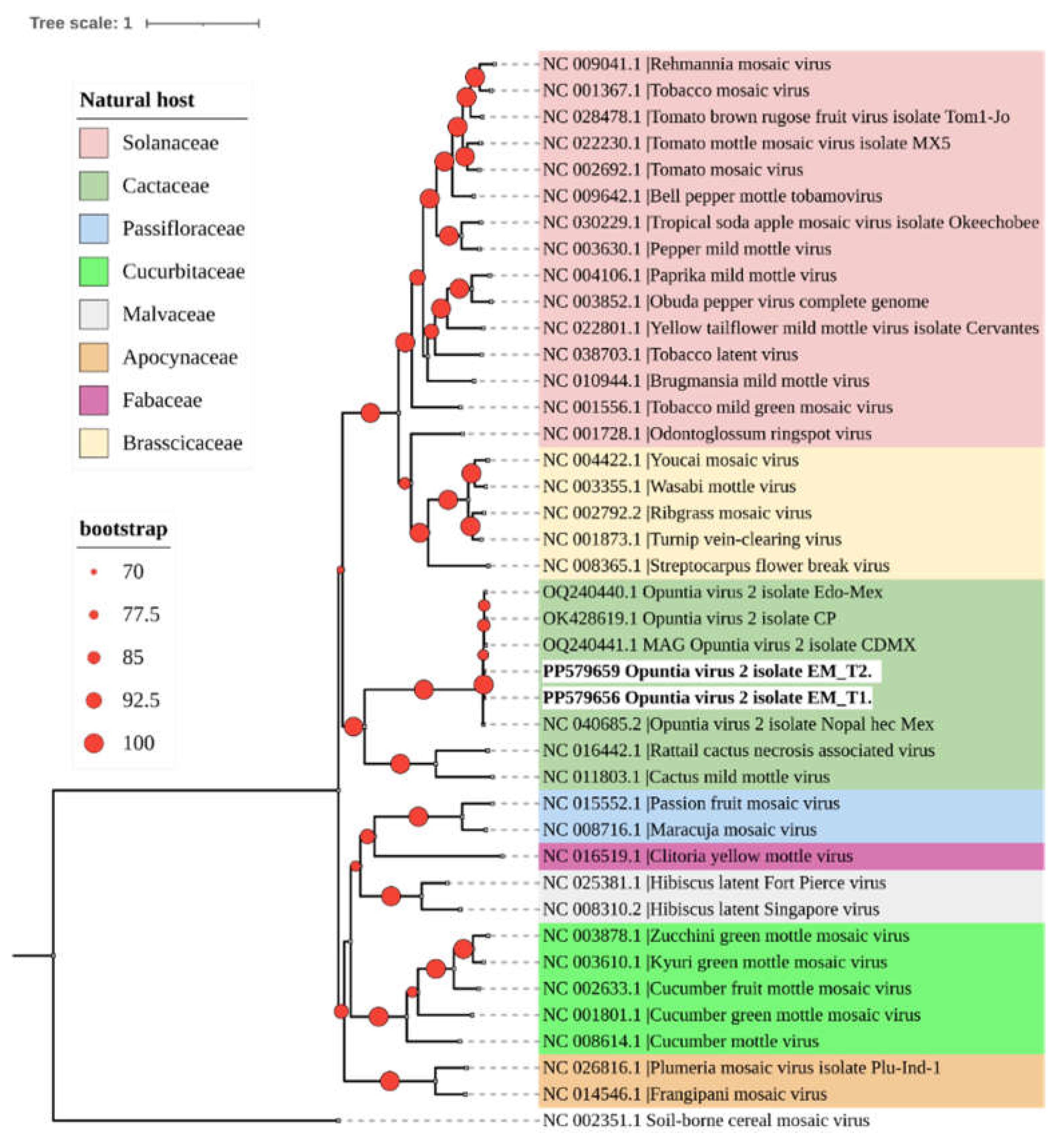
Figure 3.
A phylogenetic tree was constructed using the maximum likelihood method with complete genome sequences of the Carlavirus genus obtained from GenBank (accession numbers shown). The tree was built with IQtree 2.3.1. The CCV-1 sequence identified in this study is highlighted in bold. The sequence of Apricot vein clearing associated virus (genus Prunevirus) was used as an outgroup. Circles on branches indicate UFBoot support values > 70%.
Figure 3.
A phylogenetic tree was constructed using the maximum likelihood method with complete genome sequences of the Carlavirus genus obtained from GenBank (accession numbers shown). The tree was built with IQtree 2.3.1. The CCV-1 sequence identified in this study is highlighted in bold. The sequence of Apricot vein clearing associated virus (genus Prunevirus) was used as an outgroup. Circles on branches indicate UFBoot support values > 70%.

Figure 4.
A phylogenetic tree was constructed using the maximum likelihood method with complete genome sequences of the Potexvirus genus obtained from GenBank (accession numbers shown). The tree was built with IQtree 2.3.1. The identified OPV-A sequences in this study are highlighted in bold. The sequence of Senna severe yellow mosaic virus (genus Allexivirus) was used as an outgroup. Circles on branches indicate UFBoot support values > 70%.
Figure 4.
A phylogenetic tree was constructed using the maximum likelihood method with complete genome sequences of the Potexvirus genus obtained from GenBank (accession numbers shown). The tree was built with IQtree 2.3.1. The identified OPV-A sequences in this study are highlighted in bold. The sequence of Senna severe yellow mosaic virus (genus Allexivirus) was used as an outgroup. Circles on branches indicate UFBoot support values > 70%.
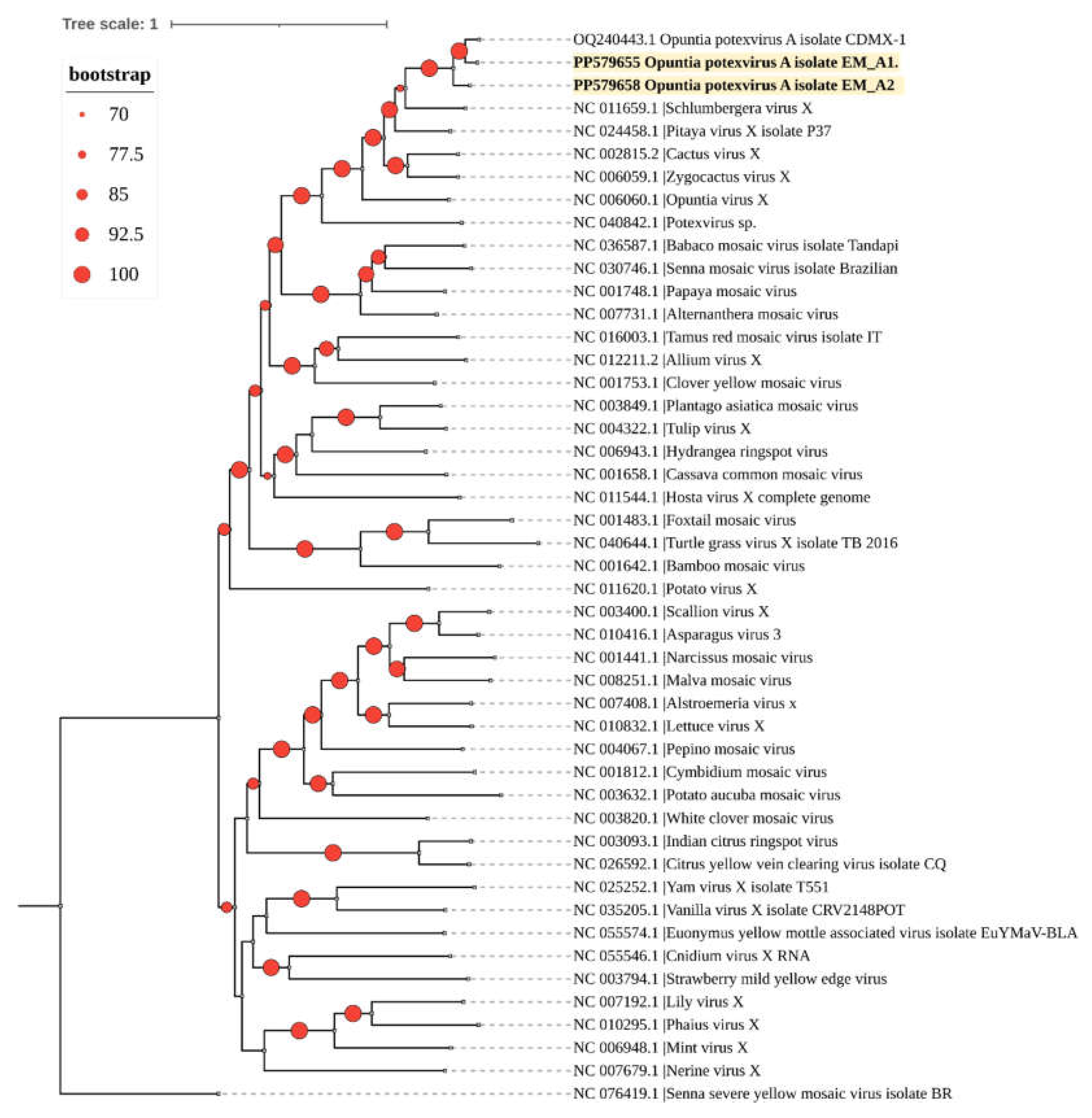
Figure 5.
Characterization of the predicted secondary structure and conserved structural elements of OVd-2. (A), Nucleotides in the upper and lower conserved central region (CCR) are indicated in red and blue, respectively. The nucleotides in green indicate the terminal conserved region (TCR) structural element. The two pairs of arrows pointing in opposite directions represent the primers used for amplifying the complete genome of the viroid; (B), The circular RNA structure of OVd-2 was determined using RT-PCR with the primer sets F90/89R and 208F/207R, each adjacent to the other in opposite directions (‘F’ = forward primers; ‘R’ = reverse primers). A healthy nopal vegetable sample was used as a negative control (NC). M, 100 bp molecular marker (Promega, Madison, USA).
Figure 5.
Characterization of the predicted secondary structure and conserved structural elements of OVd-2. (A), Nucleotides in the upper and lower conserved central region (CCR) are indicated in red and blue, respectively. The nucleotides in green indicate the terminal conserved region (TCR) structural element. The two pairs of arrows pointing in opposite directions represent the primers used for amplifying the complete genome of the viroid; (B), The circular RNA structure of OVd-2 was determined using RT-PCR with the primer sets F90/89R and 208F/207R, each adjacent to the other in opposite directions (‘F’ = forward primers; ‘R’ = reverse primers). A healthy nopal vegetable sample was used as a negative control (NC). M, 100 bp molecular marker (Promega, Madison, USA).
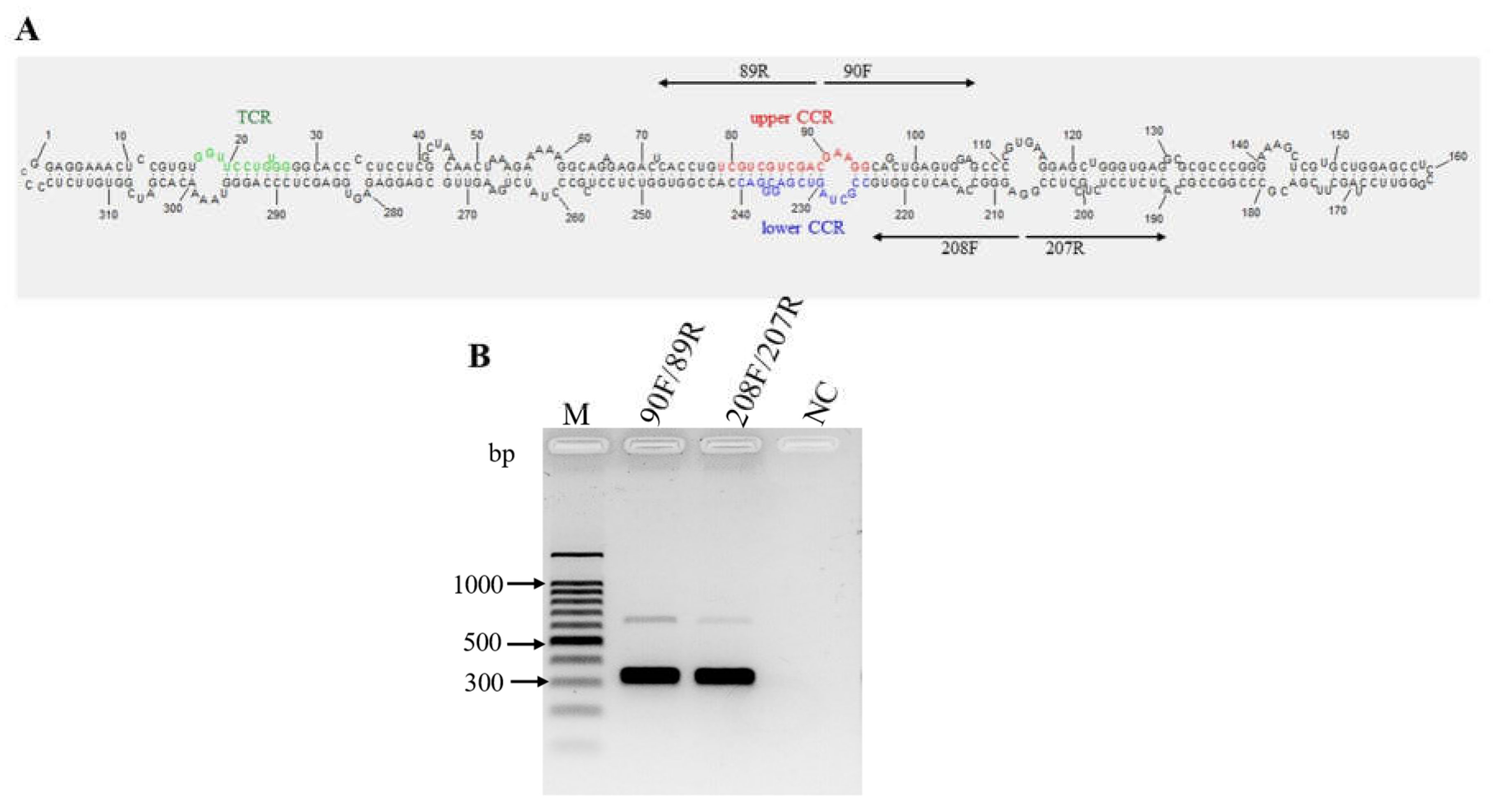
Figure 6.
Phylogenetic relationships of Opuntia viroid 2 (OPVd-2), Mexican opuntia viroid (MOVd) (highlighted in bold), and all current RefSeq viroid sequences available in GenBank for the family Pospiviroidae. The phylogenetic tree was built with 1000 bootstrap replicates using maximum likelihood method and the Jukes-Cantor model in MEGA X [9,23]. A color code was used to represent the different genera within this family. Peach latent mosaic viroid (PLMVd) (family Avsunviroidae) was included as an outgroup. Circles on branches indicate bootstrap values > 70% (generated from 1000 replicates).
Figure 6.
Phylogenetic relationships of Opuntia viroid 2 (OPVd-2), Mexican opuntia viroid (MOVd) (highlighted in bold), and all current RefSeq viroid sequences available in GenBank for the family Pospiviroidae. The phylogenetic tree was built with 1000 bootstrap replicates using maximum likelihood method and the Jukes-Cantor model in MEGA X [9,23]. A color code was used to represent the different genera within this family. Peach latent mosaic viroid (PLMVd) (family Avsunviroidae) was included as an outgroup. Circles on branches indicate bootstrap values > 70% (generated from 1000 replicates).
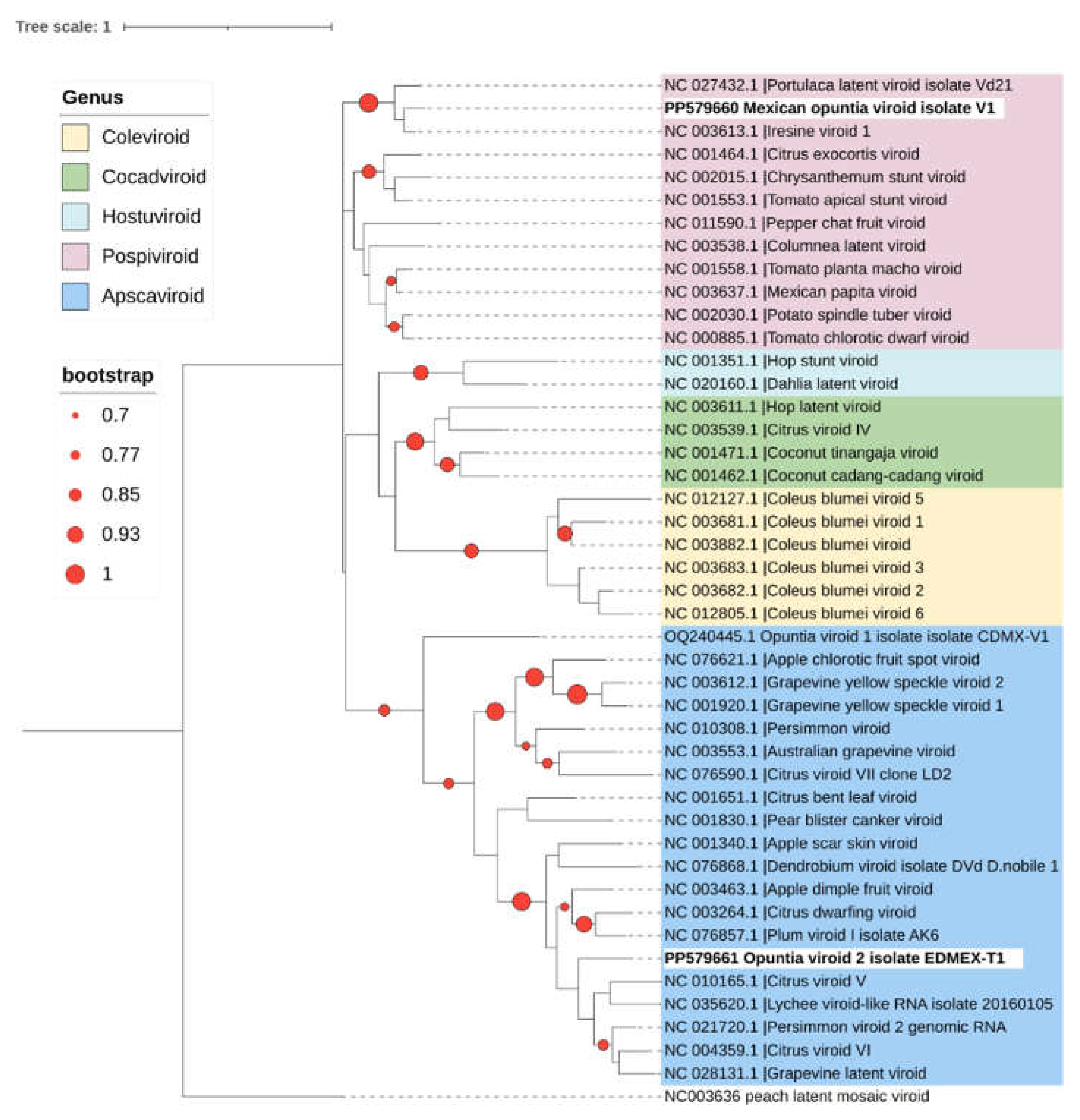
Figure 7.
The pairwise identity frequency distributions obtained with the Sequence Demarcation Tool (SDT) [25] indicated that Opuntia viroid 2 (OVd-2) and the Mexican opuntia viroid (MOVd) are new viroid species. The sequences of OVd-2 and MOVd had less than 80% and 90% identity, respectively, with any other known viroid sequence across the entire genome. Different genera within the family Pospiviroidae are indicated. Peach latent mosaic viroid (PLMVd, NC_003636) (family Avsunviroidae) was included as an outgroup.
Figure 7.
The pairwise identity frequency distributions obtained with the Sequence Demarcation Tool (SDT) [25] indicated that Opuntia viroid 2 (OVd-2) and the Mexican opuntia viroid (MOVd) are new viroid species. The sequences of OVd-2 and MOVd had less than 80% and 90% identity, respectively, with any other known viroid sequence across the entire genome. Different genera within the family Pospiviroidae are indicated. Peach latent mosaic viroid (PLMVd, NC_003636) (family Avsunviroidae) was included as an outgroup.
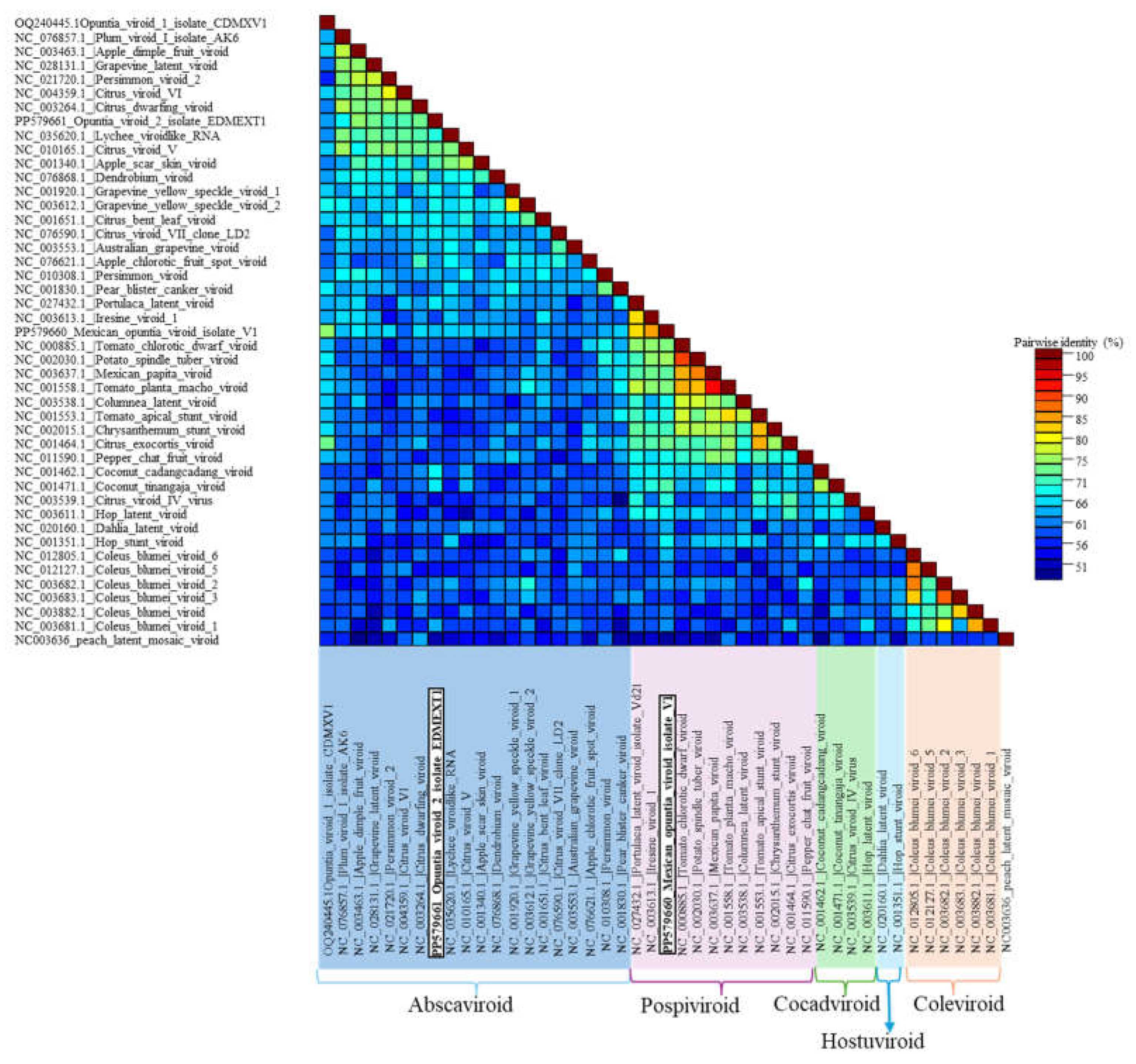
Figure 8.
Characterization of the predicted secondary structure and conserved structural elements of Mexican opuntia viroid (MOVd). (A), Nucleotides in the upper and lower conserved central region (CCR) are indicated in red and blue, respectively. The green nucleotides indicate the terminal conserved region (TCR) structural elements. The two pairs of arrows pointing in opposite directions represent the primers used for full genome amplification; (B) The circular nature of MOVd RNA was confirmed through RT-PCR with the primer sets F87/86R and 307F/306R, each adjacent to the other in opposite directions (‘F’ = forward primers; ‘R’ = reverse primers). A healthy nopal vegetable sample was used as a negative control (NC). M, 100 bp molecular marker (Promega, Madison, USA).
Figure 8.
Characterization of the predicted secondary structure and conserved structural elements of Mexican opuntia viroid (MOVd). (A), Nucleotides in the upper and lower conserved central region (CCR) are indicated in red and blue, respectively. The green nucleotides indicate the terminal conserved region (TCR) structural elements. The two pairs of arrows pointing in opposite directions represent the primers used for full genome amplification; (B) The circular nature of MOVd RNA was confirmed through RT-PCR with the primer sets F87/86R and 307F/306R, each adjacent to the other in opposite directions (‘F’ = forward primers; ‘R’ = reverse primers). A healthy nopal vegetable sample was used as a negative control (NC). M, 100 bp molecular marker (Promega, Madison, USA).
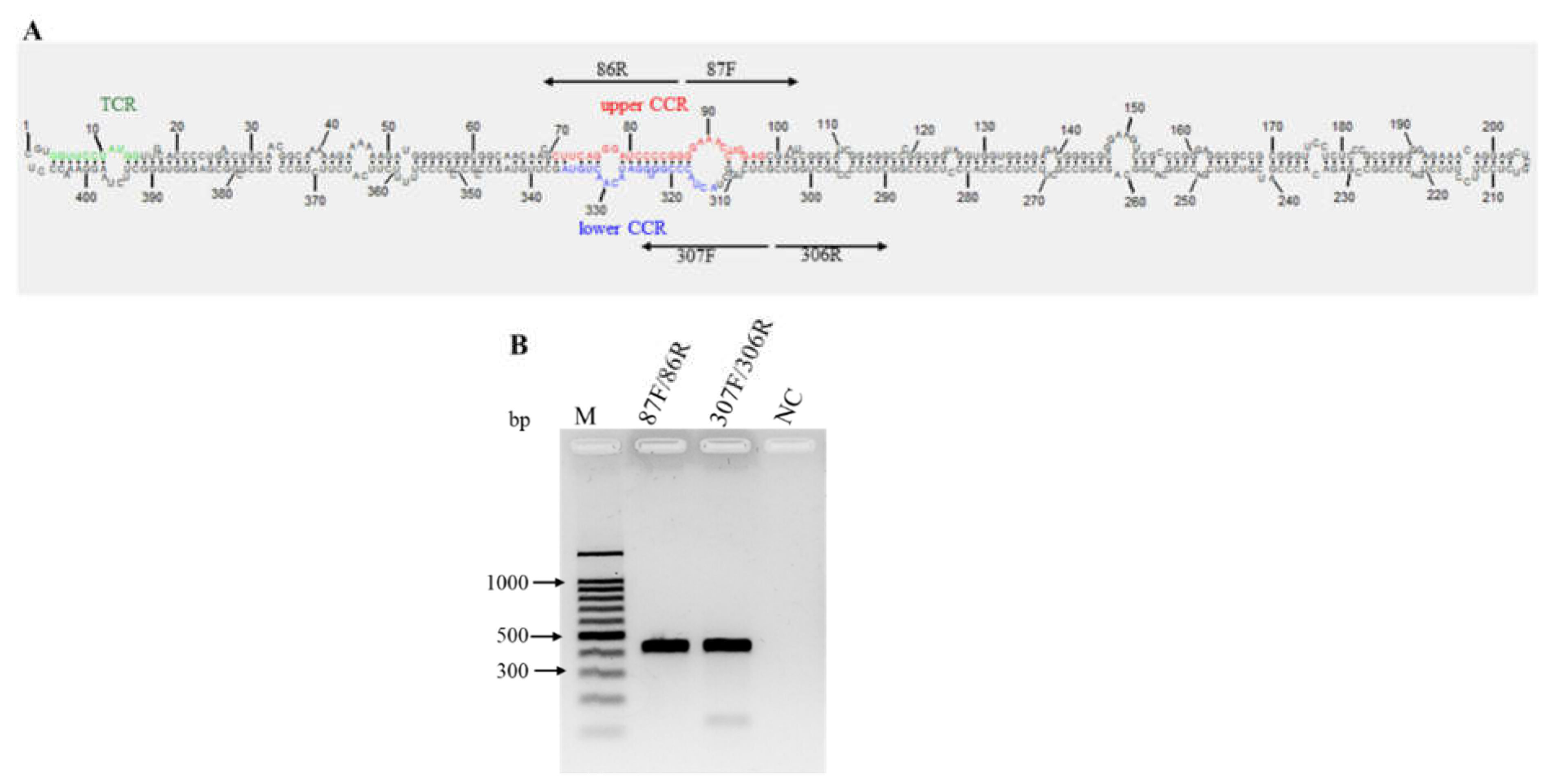

Table 1.
Primers used for the validation and detection of viruses/viroids in prickly pear cactus (fruit and nopal vegetable) from the eastern region of the State of Mexico.
Table 1.
Primers used for the validation and detection of viruses/viroids in prickly pear cactus (fruit and nopal vegetable) from the eastern region of the State of Mexico.
| Virus/Viroid | Primer | Sequence (5′-3′) | Amplicon (bp) | Region |
|---|---|---|---|---|
| Opuntia potexvirus A | OPVA-RepF | AAGCTCGCAGCATCCATCAA | 482 | Viral replicase |
| Opuntia potexvirus A | OPVA-RepR | GGGTGAAGGGACGGTAGTTG | ||
| Cactus carlavirus 1 | CCV-1F | AATGGGCGCCTTTAGGTTCA | 559 | Capsid protein |
| Cactus carlavirus 1 | CCV-1R | AATTCCAAGCTCCCGTCAGG | ||
| Opuntia virus 2 | OV2-F | CTTCCAAGAGTTCTAGCGCCT | 609 | Capsid protein |
| Opuntia virus 2 | OV2-R | ACCTGCAGGATTACCACCAC | ||
| Opuntia viroid I | OPVd_IF | GACGGAGCGTCGAGAAGTAG | 412 | Complete genome |
| Opuntia viroid I | OPVd_IR | GCC GGC GCC GAA GCC CGA G | ||
| Opuntia viroid II | Opuntia_viroid_II_307F | TCTGGCTACTACCCGGTGG | 407 | |
| Opuntia viroid II | Opuntia_viroid_II_306R | GCGACCAGCAGGGGAAG | Complete genome | |
| Opuntia viroid II | Opuntia_viroid_II_86R | CCGGGGATCCCTGAAG | 407 | |
| Opuntia viroid II | Opuntia_viroid_II_87F | GGAAACCTGGAGCGAACTC | Complete genome | |
| Opuntia viroid III | Opuntia_viroid_III_F90 | GAAGGCAGCTGAGTGGAG | 319 | |
| Opuntia viroid III | Opuntia_viroid_III_R89 | GTCGACGACGACAGGTGA | Complete genome | |
| Opuntia viroid III | Opuntia_viroid_III_208F | AGGGCCACACTCGGTG | 319 | Complete genome |
| Opuntia viroid III | Opuntia_viroid_III_207R | CCGGAGGCAGAGGAGAG | ||
Table 2.
Locations sampled for the detection of viruses and viroids in different types of prickly pear cactus in the eastern region of the State of Mexico.
Table 2.
Locations sampled for the detection of viruses and viroids in different types of prickly pear cactus in the eastern region of the State of Mexico.
| Municipality | Location | Type of Nopal | |||
|---|---|---|---|---|---|
| Fruit | Nopal Vegetable | Xoconostle | wild prickly pear cactus | ||
| Axapusco | Cuautlacingo | 3 | 38 | 0 | 0 |
| Axapusco | 4 | 0 | 0 | 0 | |
| Temascalapa | Santa Ana Tlachiahualpa | 11 | 1 | 0 | 0 |
| Nopaltepec | San Felipe Teotitlán | 20 | 0 | 4 | 2 |
| Teotihuacán | Teotihuacán | 0 | 4 | 0 | 0 |
| Total | 37 | 43 | 4 | 2 | |
Table 3.
Comparison of total or partial genome sequences of the viruses and viroids detected through HTS in nopal vegetable and prickly pear cactus samples with the most similar reference sequence available in NCBI’s GenBank.
Table 3.
Comparison of total or partial genome sequences of the viruses and viroids detected through HTS in nopal vegetable and prickly pear cactus samples with the most similar reference sequence available in NCBI’s GenBank.
| HTS Sample | Virus/Viroid Detected* | GenBank Accession Number | Isolation | Genome Segment | Reference Sequence Accession Number (NCBI GenBank) | % Identity with Reference |
|---|---|---|---|---|---|---|
| nopal vegetable | CCV-1 | PP579657 | EM_C2 | complete | KU854930.4 | 92.14% |
| OPV- A | PP579658 | EM_A2 | complete | OQ240443.1 | 85.50% | |
| OV2 | PP579659 | EM_T2 | complete | NC_040685.2 | 98.98% | |
| OVd 1 | PP579662 | EDMEX-V1 | complete | OQ240445.1 | 98.54% | |
| prickly pear cactus | CCV-1 | PP579654 | EM_C1. | **RdRp | KU854930.4 | 88.83% |
| OPV-A | PP579655 | EM_A1 | complete | OQ240443.1 | 90.16% | |
| OV2 | PP579656 | EM_T1 | complete | NC_040685.2 | 98.98% |
*CCV-1: Cactus carlavirus 1; OPV-A: Opuntia potexvirus A; OVd-1: Opuntia viroid 1; ** RNA polymerase-RNA dependent.
Table 4.
New viroid species detected by HTS in prickly pear cactus and nopal vegetable.
| HTS Sample | Viroid | GenBank Accession Number | Isolate | Closest Viroid in BLASTn Analysis | Reference Sequence, Accession Number (NCBI GenBank) | % Identity with Reference (% Query Cover) |
|---|---|---|---|---|---|---|
| nopal vegetable | Mexican opuntia viroid | PP579660 | V1 | Iresine viroid 1 | OM108483.1 | 83.33 (99) |
| prickly pear cactus | Opuntia viroid 2 | PP579661 | EDMEX-T1 | Grapevine latent viroid | MG770884.1 | 82.81 (38) |
Table 5.
Location of conserved domains of viroids in the reference sequence of the Pospiviroid and Apscaviroid genera compared to the new viroid species detected in this study. Discrepancies in the sequences are underlined and in bold.
Table 5.
Location of conserved domains of viroids in the reference sequence of the Pospiviroid and Apscaviroid genera compared to the new viroid species detected in this study. Discrepancies in the sequences are underlined and in bold.
| Viroid* | TCR | CCR Upper Strand | CCR Lower Strand |
|---|---|---|---|
| MOVd (PP579660.1) | GGUUCCUGUGG | CUUCAGGGAUCCCCGGGGAAACCUGGAG | ACUACCCGGUGGAUACAACUGUAGCU |
| PSTVd (NC_002030.1) | GGUUCCUAUGG | CUUCAGGGAUCCCCGGGGAAACCUGGAG | ACUACCCGGUGGAAACAACUGAAGCU |
| IrVd-1 (NC_003613.1) | GGUUCCAAUGG | CUUCAGGGAUCCCCGGGGAAACCUGGAG | ACUACCCGGUGGAUACAACUGUAGCU |
| OVd-2 (PP579661.1) | GGUUCCUGUGG | UCGUCGUCGACGAAGG | CCGCUAGUCGAGCGGAC |
| ASSVd (NC_001340) | GGUUCCUGUGG | UCGUCGUCGACGAAGG | CCGCUAGUCGAGCGGAC |
* MOVd: Mexican opuntia viroid; PSTVd: potato spindle tuber viroid; IrVd-1: Irasine viroid 1; OVd-2: opuntia viroid-2; ASSVd: apple scar skin viroid.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



