
Submitted:
26 June 2024
Posted:
26 June 2024
You are already at the latest version
Abstract
The common house mosquito (Culex pipiens) is a native vector for West Nile virus (WNV). Invasive species like the tiger mosquito (Aedes albopictus) and Asian bush mosquito (Aedes japonicus) are rapidly spreading through Europe, posing a major threat as vectors for dengue, chikungunya (CHIKV), and Japanese Encephalitis virus (JEV). These mosquitoes share a similar ecological niche as larvae, but the carry-over effects of aquatic larval interactions to terrestrial adult stage remain largely unknown and their medical relevance requires further investigation. This study examines the context-dependency of larval interactions among Aedes albopictus, Aedes japonicus and Culex pipiens. The survival, development time, growth, and energetic storage were measured in different European populations within density-response experiments (intraspecific) and replacement experiments (interspecific) at 20°C and 26°C. Overall, Ae. japonicus was the weakest competitor, while competition between Ae. albopictus and Cx. pipiens varied with temperature. Culex pipiens was infected with WNV, Ae. albopictus with CHIKV and Ae. japonicus with JEV. While no JEV infection was observed, mosquitoes experiencing interspecific interactions during their larval stages exhibited higher infection rates and viral RNA titers for CHIKV and WNV. This increased susceptibility for viral infection after larval competition suggests a higher risk of arbovirus transmission in co-occurring populations.
Keywords:
Aedes albopictus
; Aedes japonicus
; arbovirus
; viral RNA titer
; CHIKV
; Culex pipiens
; infection rate
; JEV
; WNV
1. Introduction
Over the past 50 years, the incidence of dengue has increased by 30-fold [1]. By 2023, more than 100 countries reported the circulation of chikungunya virus (CHIKV), putting an estimation of 1.3 billion people at risk of chikungunya fever globally [2]. A growing amount of autochthonous dengue cases have been observed in Europe, with a total of 74 cases reported between 2010 and 2021. In the past two years, this number increased considerably, with 71 cases in 2022 and 130 cases in 2023 ECDC [3]. The first autochthonous Chikungunya outbreak in Europe occurred in Italy in 2007, with approximately 330 cases identified. Since then, sporadic outbreaks of Chikungunya have been observed, which were often associated with extreme climate events [4]. In 2017, a total of 270 confirmed and 219 probable autochthonous Chikungunya cases have been observed [5]. An increase in autochthonous cases of WNV were recorded, with 1471 cases between 2000 and 2017, 1503 cases and 180 deaths in 2018 alone, and 1113 cases and 92 deaths in 2022 [6].
Aedes mosquitoes are the primary vectors of dengue (DENV) and CHIKV virus [7]. In Europe, these invasive species include the Asian tiger mosquito Ae. albopictus (Skuse, 1894), the Asian bush mosquito Ae. japonicus japonicus (Theobald, 1901), and the Korean bush mosquito Ae. koreicus (Edwards, 1917) [8]. There are four subspecies of Ae. japonicus, however, only Ae. japonicus japonicus is found in Europe [9], therefore it will be referred to as Ae. japonicus. Aedes albopictus and Ae. japonicus are considered to be one of the fastest spreading invasive species [9]. Their spread and establishment to new regions is largely influenced by climate change and international trade [8,10]. Additionally, anthropization of the landscape is a significant factor influencing the dynamics of vector-borne pathogens [1]. One of the main consequences, apart from nuisance due to high abundances, are outbreaks of mosquito-borne diseases. It has been observed that local outbreaks typically manifest within a time frame of 5 to 15 years subsequent to the establishment of Ae. albopictus populations [11]. This statement is supported by the recent autochthonous outbreaks of DENV, and CHIKV in Europe [12].
Additionally, Culex pipiens Linnaeus, 1758, is a known vector for WNV. This species is endemic, widespread, and abundant in Europe [13]. The species consists of two bioforms: Cx. pipiens pipiens and Cx. pipiens molestus [14]. With the arrival of Ae. albopictus (Albania: 1979 and Italy: 1990) [10], and Ae. japonicus (Belgium: 2002) [8,15], it now encounters these invaders in shared larval habitats [16]. All three species occur in artificial containers [17,18].
It is hypothesized that interspecific competition among mosquito larvae may enhance their vector competence for arboviruses [19,20,21]. Competitive stress negatively affects mosquito fitness, which in turn has a detrimental impact on the mosquito immune system and physical barriers against viral infection. This carry-over effect of the aquatic larval interactions to the terrestrial adult stage might be important to be considered when estimating the vector competence of arboviral vectors. For example, Alto, Lounibos [19] observed that Ae. albopictus females were smaller, had a higher infection rate, viral RNA titer and dissemination rate of SINV following larval competition with Ae. aegypti. Interspecific competition led to more intense competition compared to intraspecific competition. Similarly, Alto, Lounibos [20] discovered an elevated infection and dissemination rate of DENV in Ae. albopictus after larval competition with Ae. aegypti. In addition, Bevins [21] reported that Ae. triseriatus females had an increased mortality, larger size, and higher infection and dissemination rate of LACV following larval competition with Ae. albopictus.
Aedes albopictus has often been found to be a superior competitor during the larval stage in the aquatic environment [22,23,24,25,26,27,28,29]. However, other studies suggest that a balanced coexistence between Cx. pipiens and Ae. albopictus is possible when sufficient food resources are available [17,28,30]. Aedes japonicus is expected to not strongly interact with Cx. pipiens [31,32]. It is considered a weak larval competitor when compared to Ae. albopictus [24,33].
Larval density and competition are found to affect egg production, body size, energy reserves, and longevity of adult females [34,35,36,37]. Generally, larger females exhibit higher levels of protein, glycogen, and lipid content upon emergence [38]. These energy reserves are important while searching for a suitable host. They show more biting persistence, higher longevity [21] and vector capacity [39]. Additionally, lipids also regulate the immune response [35,39,40]. The innate immune system of mosquitoes consists of various lines of defense mechanics. The epithelium-lined midgut serves as the initial barrier, while hemocytes play a crucial role as cellular components of innate immunity. Additionally, Toll and Imd pathways are responsible for the signaling of the production of antimicrobial peptides (AMP), contributing to the humoral defenses of mosquitoes [35]. Lipids facilitate membrane biogenesis at infection sites and in hemocytes [40]. In addition, lipid droplets could potentially serve as an energy source for microflora, and have been linked with the activation of Toll-like receptors during DENV infection [39]. In Ae. aegypti subjected to larval nutrient stress, a decrease in the number of hemocytes was observed, however, enhanced fat body derived immune factors, such as AMPs, were found. Furthermore, transcripts of Spaetzle, a key regulator of the Toll pathway, and certain immune-related genes were less abundant but demonstrated increased expression [35].
Bevins [21] observed larger females of Ae. triseriatus after interspecific larval competition with Ae. albopictus. These larger females were more susceptible to develop LACV infections compared to females from intraspecific experiments. Larger females of Ae. albopictus have more tissue for virus replication, leading to higher viral RNA titers [19]. Controversially, a study on Ae. triseriatus females deriving from malnourished larvae reveal smaller females with a thinner basal lamina [41], a membrane that envelops the midgut and hinder virus movement [20]. These females were also associated with higher infection, dissemination and transmission rates of a bunyavirus. Telang, Qayum [35] observed that basal lamina thickness was not affected by the size of the female. It was noted that certain immune-related genes were less expressed in stressed larvae, but exhibited increased expression in females derived from these stressed larvae [35]. These findings suggest that nutritional stress during the larval stage may result in weaker immune responses in adults [35,41,42] , which potentially increase their vector competence. However, these smaller females will have a shorter longevity, and thus vector capacity, which might be too short to complete the extrinsic incubation period [21,36].
The objective of this study was to investigate the carry-over effect of larval interactions between Cx. pipiens and Ae. albopictus as well as Ae. japonicus in recently established populations from Central Europe. This carry-over effect was analyzed via intra- and interspecific larval competition on arboviral infection of Ae. albopictus, Ae. japonicus, and Cx. pipiens during their adult stage. The effect of synecological patterns at 20 and 26°C was assessed by collecting data on larval mortality, development time, behavior, pupal size, and the content of energy reserves. Afterwards, an infection experiment was conducted with the medically relevant CHIKV, JEV, and WNV to test whether significant ecological patterns from larval competition would carry over to an increased infection rate and viral RNA titer, potentially elevating the risk of arbovirus transmission.
2. Material & Methods
2.1. Larval Competition Study
2.1.1. Mosquito Material
Culex pipiens s.s./Cx. torrentium egg rafts were sampled in Frankfurt (Germany) in September 2021, June and August 2022. The egg rafts were stored for several days on humid cotton at 10°C, before they were placed in softened water at 20°C or 26°C to hatch. To analyze the biotypes, a multiplex quantitative real-time PCR (qRT-PCR) was performed as described by Rudolf, Czajka [43], with all specimens used during the infection experiments. A total of 85 Culex specimens were tested, of which 77 were Cx. pipiens pipiens, 7 Cx. torrentium, and 1 hybrid Cx. pipiens pipiens x molestus. The Ae. albopictus strain (20AAlb.DE-KABS.12) originated from egg collections in Achern (Germany) in September 2020 and was reared at 28°C with 80% relative humidity and a 16:8 L:D photoperiod. The Cx. pipiens biotype molestus strain (20CPip.BE-ITMf.6) originated from larval collections in Hove (Belgium) in 2020 and was reared as a colony with overlapping generations at 23°C with relative humidity of 80% and a 16:8 L:D photoperiod. Since the strain also contains genetically mixed forms with the bioform pipiens (nondiscriminating multiple generations hybrids and backcross, from here on called hybrids), a PCR was conducted on the used specimens [44]. A total of 182 Cx. pipiens s.l. were tested, and revealed 130 Cx. pipiens molestus, 46 hybrid Cx. pipiens pipiens x molestus, and 6 Cx. pipiens pipiens. The Ae. japonicus japonicus experiments were executed with field larvae reared from collected eggs. Oviposition traps were placed and collected in Havelange (Belgium) in June and July 2022 and June 2023. The oviposition sticks were stored at 10°C up to several months upon experimental use. All rearing and competition experiments took place at 20°C or 26°C with relative humidity of 80% and a 16:8 L:D photoperiod in climatic cupboards (CPS-P530 Climatic Cabinet, RUMED Germany) at the Merian insectary of the Institute of Tropical Medicine (ITM), Antwerp, Belgium.
2.1.2. Larval Replacement-Series Experiments
Intraspecific interactions were studied at 20°C and 26°C with density response experiments for 3, 5, 15, 30 and 45 specimens in 1L cups with 600 ml soft water in triplicate. To identify the number of larvae that results in low mortality rates for interspecific experiments, the percentage of mortality was documented per larval density (Supplementary file 1 Figure S1). Interspecific interactions were investigated at 20°C and 26°C with replacement experiments for Aedes:Culex 30:0, 20:10, 15:15, 10:20 and 0:30 combinations in 1L cups with 600 ml soft water in triplicate. Within 24h after hatching, first instar larvae were placed in the experiments. Larvae were fed three times per week with sieved TetraMin (Tetra, Germany). A dose of 0.5 mg food per larvae was provided during the first four doses, from the fifth dose onwards 1 mg of food was provided per larvae to receive 6 mg per larvae in total based on Müller, Knautz [17], Müller, Knautz [45]. The time of pupation was registered for each experiment to measure development time and mortality. Three pupal growth parameters (area of cephalothorax, length and width of abdomen) were considered to correct for pupal size.
2.1.3. Video Tracking of Behavioral Variables
Fourth instar larvae from the same batch as the larval competition experiments were maintained at 20°C and used for behavioral observations using a high-quality video tracking system with digital image recognition. DanioVision hardware and Ethovision software were used to track social interactions, activity, and larval behavior such as total distance moved, velocity and body contact. Larvae were placed in a petri dish with soft water at room temperature and recording started after 30 seconds to allow acclimatization. Larvae were recorded for two minutes per experiment. Intraspecific experiments were conducted in triplicate for 1, 2, 5, 10 and 18 larvae per species. Intraspecific experiments were run in triplicate with Aedes:Culex ratios 18:0, 12:6, 9:9, 6:12 and 0:18.
2.1.4. Photometric Assays on Pupal Lipid, Glycogen, and Protein Content
The first five pupae per experiment were stored at -20°C to test for energy storage. The total content of glycogen, lipid and protein per pupae was analyzed via photometric assays according to Van Handel [46], Van Handel [47], and Bradford [48] respectively, as described by Bock, Kuch [49]. Pupal growth parameters (area of cephalothorax, length and width of abdomen) were taken. The abdominal width was considered the most robust metric (selection of parameter based on the lowest coefficient of variation, see Supplementary file 5 Table S1) and was used to correct for pupal size. The following equation was used:
2.1.5. Data Analysis
The Relative Crowding Coefficient (RCC) is used as a measure of competition. An RCC value of 1 shows that both species are equal competitors. Values below or greater than 1 indicate out-competition [50,51]. The RCC values were calculated for the development time, pupal size, pupal energy content, protein content, and larval behavior (total distance moved, velocity, and body contact) of the three species from the means of three replicate experiments. The formula used was described by Harper (1977) and adapted by Novak, Higley [50] and Oberg, Young [51] as follows according to Müller, Knautz [17]:
There is currently no test available to determine if these RCC results are significantly different [51], therefore we conducted two-way ANOVA’s, Kruskal-Wallis or Friedman tests on the raw data (see Supplementary files). A two-way ANOVA was conducted to test for significant differences in larval density, species or species ratio and this interaction. Tukey’s multiple comparisons test was used to compare all data. All statistical tests were conducted using Prism (version 10.1.2, GraphPad Software INC., USA). Statistical significance was defined as P < 0.05. Kolmogorov-Smirnov test and Shapiro-Wilk test were used to test for normality, and homoscedasticity was tested by curve fitting for appropriate weighting of residuals (version 10.1.2, GraphPad Software INC., USA). When assumptions for normality were violated, a Kruskal-Wallis test or Friedman test was performed instead.
2.2. Infection Study
2.2.1. Infectious Blood Feeding of Mosquitoes Which Experienced Larval Competition
Selected treatments from previously conducted larval competition experiments were repeatedly conducted at ITM in Belgium and freshly emerged adult mosquitoes were transferred to the Bernhard Nocht Institute for Tropical Medicine (BNITM) in Germany to determine the rate of infection in mosquitoes which experienced larval competition. All experiments at BNITM were performed under BSL-3 conditions. Seven to ten days after eclosion, mosquitoes were anesthetized with carbon dioxide, sorted into vials, and starved for one (Aedes) or two (Culex) night(s). Infection was performed as described by Heitmann, Jansen [53]. Blood meal, containing 50% human blood (expired banked blood), 30% of an 8% fructose solution, 10% filtrated bovine serum (FBS), and 10% virus stock; was offered with a final concentration of 107 plaque forming units (PFU)/ml for WNV (clade 1a, strain TOS-09, Genbank HM991273/HM641225), 106 PFU/ml for CHIKV (strain CNR 24/2014, supplied by the European virus Archive goes global project), and 106 PFU/ml for JEV (strain SA-14, GenBank accession number EU073992). All virus stocks were propagated using Vero cells (Chlorocebus sabaeus; CVCL 0059, obtained from ATCC, Cat# CCL-81). Culex pipiens was infected with West- Nile virus (WNV), Ae. albopictus with chikungunya virus (CHIKV) and Ae. japonicus with Japanese Encephalitis virus (JEV). Artificial blood meal was offered for Cx. pipiens via cotton stick overnight, for Ae. albopictus and Ae. japonicus in two droplets (50 µl each) per vial for 2 h. Fully engorged mosquitoes were sorted and kept at 24 +/- 5°C (mimicking fluctuating temperatures between day and night), with a relative humidity of 70%, 12:12 L:D photoperiod, and continuous fructose supply. Mosquitoes were kept for WNV and JEV for two weeks; for CHIKV for one week.
2.2.2. Quantification of Infection Rate
A total of 267 Cx. pipiens, 57 Ae. albopictus, and 47 Ae. japonicus were analyzed individually. Afterwards, specimens were homogenized in 500 µl Dulbecco’s modified eagle medium (DMEM). RNA was extracted using the MagMAX CORE nucleic acid purification kit (Applied Biosystems, Thermo Fisher Scientific Corporation, Waltham, MA, USA). By using a RT-qPCR, viral RNA titer was determined (RealStar Chikungunya RT-PCR Kit 2.0; RealStar WNV RT-PCR 2.0, both from altona Diagnostics, Hamburg, Germany; and for JEV as described by Huber, Jansen [54], using QuantiTect Probe RT-PCR Kit, Qiagen, Hilden, Germany). From these results, the infection rate was calculated as follows:
2.2.3. Statistical Analysis
A principal component analysis (PCA) was conducted on the 14 variables measured during the competition and infection experiments: larval ratio, pupal size (cephalothorax area, abdominal length and width), energy (lipid, glycogen), protein content, mortality, larval development time, behavioral variables (distance moved, velocity, and duration of the body contact), infection rate, and viral RNA titer. Statistical tests were conducted using Prism (version 10.1.2, GraphPad Software INC., USA).
3. Results
Our research shows that interspecific competition had a significant impact on all three species, although out-competition was rarely observed among the 14 variables tested. The synecological patterns were predominantly noticeable at the often overlooked metabolic and behavioral levels. Specifically, Cx. pipiens showed pronounced responses at low temperatures, Ae. albopictus at high temperatures, and Ae. japonicus at both temperatures.
3.1. Competition Study
3.1.1. Mortality
The larval mortality of these three mosquito species in the interspecific competition treatments was low. The mortality of Cx. pipiens s.s./Cx. torrentium was higher if compared to the Ae. japonicus x Cx. pipiens bioform molestus competitive treatments (Supplementary file 2 Figure S2). Friedman tests were conducted to test for difference between the species and larval densities, however, results were not significant. Low mortality after intraspecific competition in microhabitats containing 30 larvae confirmed the use of 30 larvae to be a suitable amount for interspecific experiments with the three mosquito species (Supplementary file 1 Figure S1).
3.1.2. Development Time
A two-way ANOVA conducted to compare the development time for 50 % of the larvae to emerge revealed that the factors Species, Larval ratio and its Interaction were significant for the Ae. albopictus x Cx. pipiens s.s./Cx. torrentium competitive treatments at both test temperatures, with Ae. albopictus developing faster than Cx. pipiens s.s./Cx. torrentium larvae at 26°C (Supplementary file 3 Figure S3a and c). The Ae. japonicus x Cx. p. molestus competitive treatments showed significant differences for the factors Species and Interaction at 20°C, for which Ae. japonicus developed faster compared to Cx. p. molestus (Supplementary file 3 Figure S3b). At 26°C, the factors Larval ratio and Interaction were significantly different, with Cx. p. molestus developing much faster during interspecific competition (Supplementary file 3 Figure S3d).
3.1.3. Larval Behavior
Larval behavioral data on total distance moved, velocity, and body contact showed significant differences only for the duration of the body contact between the species competitive treatments, with body contact being avoided when more Ae. japonicus were present (Supplementary file 6 Figure S5).
3.1.4. Pupal Size
There were significant differences in pupal size between the species and ratios. Generally, pupae from interspecific competition were significantly larger compared to intraspecific ones. Only the Cx. pipiens s.s./Cx. torrentium showed smaller cephalothorax size in interspecific competition (Supplementary file 4 Figure S4).
3.1.5. Energy and Protein Storage
Lipid storage in the Ae. albopictus x Cx. pipiens s.s./Cx. torrentium competitive treatments is significantly lower at the higher temperature of 26°C. At 20°C lipid content is lowest in the intraspecific treatments if compared to interspecific treatments, at 26°C a slightly higher lipid content is found in Ae. albopictus. The Ae. japonicus x Cx. p. molestus competitive treatments showed a higher lipid content at 26°C (Supplementary file 7 Figure S6a-b,g-h). The glycogen content is always higher in Ae. albopictus compared to Cx. pipiens s.s./Cx. torrentium and Ae. japonicus (Supplementary file 7 Figure S6c-d,i-j). Protein content was highest at 20°C for both Ae. albopictus and Cx. pipiens s.s./Cx. torrentium, however, no difference was observed in Ae. japonicus (Supplementary file 7 Figure S6e-f,k-l).
3.1.6. Relative Crowding Coefficient
The Relative Crowding Coefficient was computed as a metric of competition. A RCC value of one indicates equal competitiveness between both species, whereas a value below or above one suggests one species outcompeting the other. For both species combinations, the effects of the competition on the larval development were clearest at 26°C. Aedes albopictus has an advantage towards Cx. pipiens s.s./Cx. torrentium at 20°C while the opposite is true at 26°C, and Cx. p. molestus could out-compete Ae. japonicus at both temperatures (Figure 1a). The RCC for size showed a smaller difference, however, Ae. albopictus had a slight advantage towards Cx. pipiens s.s./Cx. torrentium. For the other species combination, Cx. p. molestus showed an advantage regarding larger size at 20°C and Ae. japonicus had an advantage at 26°C, respectively (Figure 1b).
In general, larval (out)competition between species became more apparent in the glycogen and protein content rather than development time and pupal size. For the lipid content, an advantage towards higher lipid content is seen for Ae. albopictus at 20°C, however, at 26°C competition is very clear with Cx. pipiens s.s./Cx. torrentium having a negative impact resulting in less the lipid storage in Ae. albopictus. For Ae. japonicus, there was a disadvantage in lipid content compared to Cx. p. molestus at both temperatures (Figure 1c). Larval competition clearly influences the glycogen content of all species at both tested temperature regimes. At 20°C, Ae. albopictus had a competitive advantage with regard to higher glycogen content over Cx. pipiens s.s./Cx. torrentium, however, at 26°C the opposite was noted with Ae. albopictus being negatively affected by Cx. pipiens s.s./Cx. torrentium. For the other species combination, Cx. p. molestus always outcompeted Ae. japonicus in glycogen content, resulting in a higher glycogen content in Cx. p. molestus in interspecific competition (Figure 1d). For protein content, Ae. albopictus had the competitive advantage over Cx. p. molestus at both temperatures, with more protein content during interspecific competition. Aedes japonicus had a competitive disadvantage at both temperatures compared to Cx. p. molestus (Figure 1e).
Both distance moved and velocity measured from larval behavior show more activity for Aedes compared to Culex. In contrast, Culex shows longer body contact compared to Aedes (Figure 2).
3.2. Infection Study
Infection was successful for Ae. albopictus and Cx. pipiens s.s./Cx. torrentium, however, no infection was found for Ae. japonicus. A difference in infection rate and viral RNA titer was found in response to the competition treatment. Assumptions for normality were violated, therefore a Kruskal-Wallis test was performed on the viral RNA titer data and a Friedman test on infection rate.
For the Ae. albopictus vs Cx. pipiens s.s./Cx. torrentium combination, the former species showed an infection rate of 100 % with CHIKV. The mean number of CHIKV RNA copies per specimen of Ae. albopictus was 6.58 x 109, 2.89 x 1010, and 1.97 x 1010 genome copies for the intraspecific combination, interspecific Culex:Aedes 10:20, and 20:10 combination respectively (Figure 3a). The difference in viral RNA copies per specimen per competitive treatment was significant (Kruskal-Wallis test, p = 0.04), Kruskal-Wallis multiple comparisons test showed a significant difference between Culex:Aedes 0:30 intraspecific and 10:20 interspecific (p = 0.04). The two other comparisons (Culex:Aedes 0:30 intraspecific vs 20:10, and 10:20 vs 20:10 interspecific) were not significantly different (p = 0.55 and p = 0.69, respectively). Culex pipiens s.s./Cx. torrentium from this combination had an infection rate of 54.8 %, 50 %, and 66.7 % for the intraspecific Culex:Aedes 30:0 combination, interspecific Culex:Aedes 20:10 and 10:20 combination respectively for WNV (Figure 3b). The mean number of WNV RNA copies per specimen of Cx. pipiens s.s./Cx. torrentium was 4.34 x 107, 1.43 x108, and 5.19 x 107 genome copies for the intraspecific Culex:Aedes 30:0 combination, interspecific Culex:Aedes 20:10 and 10:20 combination respectively (Figure 3c). Differences in viral RNA titer per competition combination were not significant (Kruskal-Wallis test, p = 0.62).
For the Ae. japonicus with Cx. p. molestus combination, no infection with JEV was found for the former species. Culex p. molestus had an infection rate of 43.9 %, 54.7 %, and 59.3 % for the intraspecific Culex:Aedes 30:0 combination, interspecific Culex:Aedes 20:10 and 10:20 combination for WNV respectively (Figure 3b). None of the infection rates were significantly different (Friedman test, p = 0.17). The mean number of WNV RNA copies per specimen was 3.90 x 108, 4.42 x 108, and 2.77 x 108 genome copies for the intraspecific Culex:Aedes 30:0 combination, interspecific Culex:Aedes 20:10 and 10:20 combination respectively (Figure 3d). Differences in the number of WNV RNA copies per specimen were not significant between mosquitoes which previously (non-)experienced larval competition (Kruskal-Wallis test, p = 0.43).
For Cx. p. pipiens, Cx. torrentium, and Cx. p. pipiens x molestus, infection rate was 51.95 %, 71.43 %, and 100% respectively (Figure 4). Viral RNA titer was 8.1 x 107, 1.4 x 108, and 2.4 x 104 genome copies respectively. Infection rate was 50 %, 47.83 %, and 66.67 % for Cx. p. molestus, Cx. p. pipiens x molestus, and Cx. p. pipiens respectively (Figure 4), viral RNA titer was similar with 3.99, 3.85, and 3.95 x 108 genome copies.
A Principal Component (PC) Analysis examined the 14 variables (Development, larval ratio, mortality, CL, AL, AW, L, P, G, distance, velocity, body contact, BT, IR) and reduced them to two PC axes, which explained about 54.71 ± 6.81 % of the variation in these variables (Figure 5, Table 1). Overall, pupal size (AL, AW, CL), energy reserves (L, G) and protein content are positively correlated. Total distance moved and velocity are also grouped together. For Ae. albopictus, the PCs explained a total of 64.68 % of the proportion of variance (Table 1). Larval ratio and development were correlated, and negatively linked with pupal size and protein content, therefore there is a link on the PC1 between interspecific larval ratio, shorter development, larger pupal size and more protein content. The PC2 showed a link between a high viral RNA titer and interspecific competition, low contents of energy storage, and less larval movement (Figure 5a, Table 1).
For the corresponding Cx. pipiens s.s./Cx. torrentium strain, the PCs explained a total of 46.74 % of the proportion of variance (Table 1). On PC1 the variables velocity, distance moved, protein content, body contact, mortality and development were correlated, meaning that lower values for these variables relate to higher viral RNA titer at interspecific competition. The velocity and distance moved was negatively correlated to cephalothorax size, larval ratio, viral RNA titer, and lipid content in PC2. This also indicates that less larval movement contributes to higher viral RNA titer, but also that larger pupae with more lipid content increase the viral RNA titer. Infection rate was negatively correlated with larval ratio, pupal size, energy reserves, protein content, mortality, and development time (Figure 5b, Table 1). For both Ae. albopictus and Cx. pipiens s.s./Cx. torrentium the viral RNA titer was higher in interspecific ratios, large pupae, and less movement. Aedes albopictus showed a higher viral RNA titer when there were less energy reserves, while Cx. pipiens s.s./Cx. torrentium had a higher viral RNA titer when more lipid was stored.
The PCs for Ae. japonicus together explained 56.93 % of the proportion of variance (Table 2). No infection with JEV was found, therefore viral RNA titer and infection rate were excluded. Size, energy and protein content were grouped together on PC1, and were negatively correlated to larval ratio and velocity. On PC2, body contact was negatively correlated with velocity, larval ratio, distance moved, and mortality. This means that interspecific ratios with lower mobility rates and mortality have a larger pupal size, more body contact, and more lipid, protein, and glycogen content (Figure 5c, Table 2).
In the corresponding Cx. p. molestus strain, 50.48 % of the proportion of variance is explained by both PCs (Table 2). On the PC1, the distance moved, velocity, cephalothorax area, and protein content were grouped and negatively correlated to larval ratio and development time. The PC2 indicated that viral RNA titer and velocity were negatively correlated to size and body contact. This indicates that within interspecific ratios, there was a fast development, larger pupae, more protein content, more velocity, and a higher viral RNA titer (Figure 5d).
4. Discussion
The present study confirmed that larval interactions between Ae. albopictus and Cx. p. molestus, and Ae. japonicus and Cx. pipiens s.s./Cx. torrentium, carry-over to the capability for arboviral infection in the adult stage, at least for Ae. albopictus and Cx. pipiens. Our findings indicate that all three species were significantly affected by interspecific competition, although out-competition become rarely evident at the level of 14 variables tested. Synecological pattern was in general most expressed at often neglected metabolic and behavioral levels, and specifically pronounces at low temperature in Cx. pipiens, at high temperature in Ae. albopictus and at both temperatures in Ae. japonicus.
4.1. Aedes Albopictus vs Culex pipiens s.s./Cx. Torrentium
Development time for Ae. albopictus and Cx. pipiens s.s./Cx. torrentium was similar at 20°C, however, at 26°C the thermophilic Ae. albopictus was faster, especially during interspecific competition, which is also found by Müller, Knautz [17]. Activity was higher in both Aedes species compared to Cx. pipiens s.s./Cx. torrentium. Aedes species are known to actively search for their food, browsing on a surface, while Cx. pipiens is a filter feeder hanging near the water surface [33,58,59]. This could imply that the foraging behavior of Cx. pipiens is well-adapted in scenarios with abundant food, in which case they conserve more energy than their competitor [17]. Aedes albopictus is known to be the strongest competitor in resource-limiting conditions due to their active search for food [16,23,25,27,28], however, in eutrophic conditions it might be less successful compared to Cx. pipiens [17,28,30]. In addition, the lipid and glycogen content of Ae. albopictus was positively affected by the presence of Cx. pipiens s.s./Cx. torrentium at 20°C, whereas at 26°C Ae. albopictus was negatively affected by its presence. During protein storage, Ae. albopictus was always positively affected by the presence of Cx. pipiens s.s./Cx. torrentium while the latter species remained unaffected.
4.2. Aedes Japonicus vs Culex Pipiens Bioform Molestus
At 20°C no influence of interspecific competition was found for larval development time, and at 26°C Cx. p. molestus developed faster in interspecific competition, outcompeting Ae. japonicus. This is in line with Andreadis and Wolfe [31] and Giunti, Becker [60], who observed that Ae. japonicus does not tolerate high temperatures and prefers colder habitats. Culex p. molestus remained unaffected by Ae. japonicus during lipid acquisition, moreover, it even benefitted from its presence during glycogen uptake, indicating having more energy reserves available in interspecific combinations. During protein uptake, Aedes japonicus was not affected by the presence of Cx. p. molestus, but the latter species benefitted from interspecific competition. This observation is in line with Andreadis and Wolfe [31], and Hardstone and Andreadis [32] who predicted that Ae. japonicus would not outcompete Cx. pipiens. It is considered a weak larval competitor compared to Ae. albopictus [24,33].
4.3. Effect of Interspecific Competition On Viral Infection
This study indicates that interspecific larval competition may enhance at least the infection rates and body titers of Ae. albopictus and Cx. pipiens. Our experiments demonstrate that interspecific competition results in larger mosquitoes, a higher arboviral infection rate, and increased arboviral RNA titer. Large mosquitoes benefit of an improved longevity, blood feeding, and vector capacity (Ratnayake et al. 2023). These are crucial factors to complete the extrinsic incubation period in order to transmit a virus [21,36]. Additionally, Alto, Lounibos [19] and Bevins [21] observed that these stressed, larger females attain higher viral titers for SINDV and were more likely to disseminate LACV and develop midgut infection. Conversely, smaller and stressed females may have a weaker immune response (Grimstad and Walker 1991, Telang et al. 2012, Paige et al. 2019), potentially enhancing their vector competence. However, their shorter longevity raises uncertainty about whether they will survive long enough to complete the extrinsic incubation period [21,36]. Interspecific larval competition might therefore have an impact of expression of important factors of the immune system, influencing vector competence.
The Relative Crowding Coefficient (RCC) was calculated to use as a measure of competition. A RCC value of 1 indicates that both species are equal competitors, while a value below or greater than 1 indicates out-competition. The calculated RCC revealed that Ae. albopictus was subject to competition, which was particularly evident in its reduced lipid intake. This finding suggests a potential increase in sensitivity to viral infection, since lipids are involved in the regulation of the immune response [35,39,40].
The influence of the reduced lipid intake in Ae. albopictus was evidenced by the negative correlation with the viral RNA titer of Ae. albopictus in the PCA. Additionally, infection rate and viral RNA titer were positively correlated with interspecific competition. Similarly, infection rate in both Cx. pipiens strains were higher during interspecific competition. Consistent with previous research by Bevins [21], females subjected to intraspecific treatments were found to be less susceptible to develop LACV infections. Those findings suggest that competitive larval interactions may not directly influence DENV replication, but may contribute to a reduction in barriers that impede viral transmission [20].
In this study, we observed that interspecific competition among larvae resulted in larger pupae with elevated infection rates and increased virus-RNA titers in the female adult stage. However, interspecifically challenged Ae. albopictus had less energy storage and a higher viral RNA titer, while Cx. pipiens had a higher viral RNA titer when more lipid was stored. This might be explained by the fact that lipids are essential for flaviviruses in order to infect cells, replicate, and spread throughout the body. They facilitate its release from infected cells into new ones. This interaction between lipids and DENV in Ae. albopictus and Ae. aegypti has been well studied by Perera, Riley [61], Chotiwan, Andre [62], and Koh, Islam [63].
4.4. Implications on Bigger Scale
This study demonstrates a combination of the factors mentioned above: after interspecific competition mosquitoes were larger, had a higher infection rate, as well as a higher virus-RNA titer. This indicates that interspecific larval competition may enhance the vector competence of Ae. albopictus and Cx. pipiens. Large mosquitoes benefit of an improved longevity, blood feeding, and vector capacity [39]. These are crucial factors to complete the extrinsic incubation period in order to transmit a virus [21,36]. Additionally, these larger females attain higher viral titers [19,21]. Conversely, smaller and stressed females may have a weaker immune response [35,41,64], potentially enhancing their vector competence. However, their shorter longevity raises uncertainty about whether they will survive long enough to complete the extrinsic incubation period [21,36].
4.5. Limitations of the study
No virus transmission rate has been investigated in this study due to the large number of experimental treatment and species tested. However, the relevance of carry-over effects from larval to adult stage for the vector competence of mosquitoes merits further research. Additionally, the patterns found in laboratory environments may be different than the ones in the field [21]. Additional factors shaping larval microhabitats such as temperature, food source, water quality, larval density, species composition, physical characteristics, as well as, behavioral and immune responses of resulting adult mosquitoes of the tested strains, and finally, microbiome and viral doses, need further to be considered. A very promising avenue of research is the effect of qualitative and quantitative lipid accumulation during larval stage on arboviral vector competence to better understand the causative links in semi-aquatic environments. We observed an opposite pattern between Ae. albopictus and Cx. pipiens related to their lipid storage and infection rate. Additionally, we found that interspecific combinations develop faster and had larger pupae. This paradoxical observation requires further research.
5. Conclusions
This study provides an in-depth insight into the larval competition between Ae. albopictus, Ae. japonicus, and Cx. pipiens, and the associated carry-over effect of synecological patterns to a higher arboviral infection during their adult stage. During interspecific competition, all species developed faster, had larger pupae and more protein storage. The competition between Ae. albopictus and Cx. pipiens varied, but indicated that Ae. albopictus is the better competitor in resource limited habitats while Cx. pipiens thrives in eutrophic situations. Aedes japonicus was always a weaker competitor compared to Cx. pipiens. Aedes albopictus and Cx. pipiens were more susceptible to arboviral infection after interspecific competition and storage of lipids was lower in Ae. albopictus while it was higher in Cx. pipiens. This could suggest that lipids are involved in both the regulation of the immune response as in virus infection in the mosquito. No infection with JEV nor natural arbovirus infections were found in Ae. japonicus.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
AV: Conceptualization, Funding acquisition, Data Curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Validation, Visualization, Writing – original draft. SJ: Conceptualization: Resources, Supervision, Validation, Writing – review and editing. JDW: Resources: Writing – review and editing. CJ: Resources, Writing – review and editing. SV: Resources, Writing – review and editing. MH: Investigation, Writing – review and editing. UL: Investigation, Writing – review and editing. RL: Conceptualization, Funding acquisition, Validation, Writing – review and editing. JSC: Conceptualization, Funding acquisition, Writing – review and editing. AH: Conceptualization, Funding acquisition, Supervision, Validation, Writing – review and editing. RM: Conceptualization, Funding acquisition, Methodology, Visualization, Project administration, Supervision, Validation, Writing – review and editing.
Funding
This research was funded through the 2018-2019 BiodivERsA joint call for research proposals, under the BiodivERsA3 ERA-Net COFUND program (project DiMoC – Diversity Components of Mosquito-borne Diseases under Climate Change), and with the funding organization FWO G0G2319N. The infection study was partly funded through AIM-COST Action CA17108, supported by COST (European Cooperation in Science and Technology). Further funding was provided through the Federal Ministry of Education and Research of Germany (Grant Number 01Kl2022). This work was supported by the European Virus Archive goes Global (EVAg) project, which has received funding from the European Union’s Horizon 2020 research and innovation program under grant agreement No 653316. Adwine Vanslembrouck was supported by the Doctor Albert Dubois Foundation for tropical pathology through the Dubois-Brigué scholarship.
Acknowledgments
We would like to thank the laboratory and technical staff at the Institute of Tropical Medicine (ITM), especially Luka Wouters and Anna Schneider, for their help with the rearing of mosquitoes and collections of Ae. japonicus eggs. We thank Dr. Friederike Reuß for the German Cx. pipiens samples. We thank Artur Jöst for providing us with the Ae. albopictus colony and Dr. Giada Rossini for providing us with the WNV strain.
Data availability statement
Data supporting the conclusions of this article are included within the article. The datasets generated and analyzed during the current study are available in the Zenodo repository, DOI number.
Ethics approval and consent to participate
Not applicable.
Consent for publications
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Abbreviations
AL: Abdominal length; ANOVA: analysis of variance; AW: Abdominal width; BNITM: Bernhard Nocht Institute for Tropical Medicine; BT: body titer; CHIKV: Chikungunya virus; CT: cephalothorax area; DENV: Dengue virus; G: glycogen; IR: infection rate; ITM : Institute of Tropical Medicine; JEV: Japanese Encephalitis virus ; L : lipid ; LACV: La Crosse encephalitis virus ; P : protein ; PCA: principal component analysis; WNV: West Nile virus.
References
- Perrin, A.; Glaizot, O.; Christe, P. Worldwide impacts of landscape anthropization on mosquito abundance and diversity: A meta-analysis. Global Change Biol. 2022, 28, 6857–6871. [Google Scholar] [CrossRef] [PubMed]
- Bartholomeeusen, K.; Daniel, M.; LaBeaud, D.A.; Gasque, P.; Peeling, R.W.; Stephenson, K.E.; et al. Chikungunya fever. Nat Rev Dis Primers. 2023, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Autochthonous vectorial transmission of dengue virus in mainland EU/EEA, 2010-present [Internet]. ECDC. 2024a [cited 17/06/2024].
- Laverdeur, J.; Desmecht, D.; Hayette, M.P.; Darcis, G. Dengue and chikungunya: future threats for Northern Europe? Front Epidemiol. 2024, 4, 1342723. [Google Scholar] [CrossRef] [PubMed]
- Autochthonous transmission of chikungunya virus in mainland EU/EEA, 2007–present [Internet]. ECDC. 2024b [cited 17/06/2024].
- Historical data by year - West Nile virus seasonal surveillance [Internet]. ECDC. 2024c [cited 18/06/2024]. Available from: https://www.ecdc.europa.eu/en/west-nile-fever/surveillance-and-disease-data/historical.
- Ryan, S.J.; Carlson, C.J.; Mordecai, E.A.; Johnson, L.R. Global expansion and redistribution of Aedes-borne virus transmission risk with climate change. PLoS Negl Trop Dis. 2019, 13, e0007213. [Google Scholar] [CrossRef] [PubMed]
- Deblauwe, I.; De Wolf, K.; De Witte, J.; Schneider, A.; Verle, I.; Vanslembrouck, A.; et al. From a long-distance threat to the invasion front: a review of the invasive Aedes mosquito species in Belgium between 2007 and 2020. Parasit Vectors. 2022, 15, 206. [Google Scholar] [CrossRef] [PubMed]
- Eritja, R.; Ruiz-Arrondo, I.; Delacour-Estrella, S.; Schaffner, F.; Álvarez-Chachero, J.; Bengoa, M.; et al. First detection of Aedes japonicus in Spain: an unexpected finding triggered by citizen science. Parasites & Vectors. 2019, 12, 53. [Google Scholar]
- Sherpa, S.; Blum, M.G.B.; Capblancq, T.; Cumer, T.; Rioux, D.; Despres, L. Unravelling the invasion history of the Asian tiger mosquito in Europe. Mol Ecol. 2019, 28, 2360–2377. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.G.; Reiner, R.C.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nature Microbiology. 2019, 4, 854–863. [Google Scholar] [CrossRef]
- Worsening spread of mosquito-borne disease outbreaks in EU/EEA, according to latest ECDC figures [press release]. European Center for Disease Prevention and Control2024d. 2024d.
- Soto, A.; De Coninck, L.; Devlies, A.S.; Van De Wiele, C.; Rosales Rosas, A.L.; Wang, L.; et al. Belgian Culex pipiens pipiens are competent vectors for West Nile virus while Culex modestus are competent vectors for Usutu virus. PLoS Negl Trop Dis. 2023, 17, e0011649. [Google Scholar] [CrossRef]
- Haba, Y.; McBride, L. Origin and status of Culex pipiens mosquito ecotypes. Curr Biol. 2022, 32, R237–R46. [Google Scholar] [CrossRef]
- Versteirt, V.; Schaffner, F.; Garros, C.; Dekoninck, W.; Coosemans, M.; Van Bortel, W. Introduction and establishment of the exotic mosquito species Aedes japonicus japonicus (Diptera: Culicidae) in Belgium. J Med Entomol. 2009, 46, 1464–1467. [Google Scholar] [CrossRef] [PubMed]
- Carrieri, M.; Bacchi, M.; Bellini, R.; Maini, S. On the competition occuring between Aedes albopictus and Culex pipiens (Diptera: Culicidae) in Italy. Entomological Society of America. 2003, 32, 8. [Google Scholar]
- Müller, R.; Knautz, T.; Vollroth, S.; Berger, R.; Kress, A.; Reuss, F.; et al. Larval superiority of Culex pipiens to Aedes albopictus in a replacement series experiment: prospects for coexistence in Germany. Parasit Vectors. 2018, 11, 80. [Google Scholar] [CrossRef] [PubMed]
- Seidel, B.; Montarsi, F.; Huemer, H.P.; Indra, A.; Capelli, G.; Allerberger, F.; et al. First record of the Asian bush mosquito, Aedes japonicus japonicus, in Italy: invasion from an established Austrian population. Parasit Vectors. 2016, 9, 284. [Google Scholar] [CrossRef] [PubMed]
- Alto, B.W.; Lounibos, L.P.; Higgs, S.; Juliano, S.A. Larval Competition Differentially Affects Arbovirus Infection in Aedes Mosquitoes. Ecology. 2005, 86, 3279–3288. [Google Scholar] [CrossRef]
- Alto, B.W.; Lounibos, L.P.; Mores, C.N.; Reiskind, M.H. Larval competition alters susceptibility of adult Aedes mosquitoes to dengue infection. Proc Biol Sci. 2008, 275, 463–471. [Google Scholar] [PubMed]
- Bevins, S.N. Invasive mosquitoes, larval competition, and indirect effects on the vector competence of native mosquito species (Diptera: Culicidae). Biol Invasions. 2007, 10, 1109–1117. [Google Scholar] [CrossRef]
- Braks, M.A.H.; Honório, N.A.; Lounibos, L.P.; Lourenço-De-Oliveira, R.; Juliano, S.A. Interspecific Competition Between Two Invasive Species of Container Mosquitoes, Aedes aegypti and Aedes albopictus (Diptera: Culicidae), in Brazil. Annals of the Entomological Society of America. 2004, 97, 130–139. [Google Scholar] [CrossRef]
- Costanzo, K.S.; Mormann, K.M.; Juliano, S.A. Asymmetrical Competition and Patterns of Abundance of Aedes albopictus and Culex pipiens (Diptera: Culicidae). Journal of Medical Entomology. 2005, 42, 20. [Google Scholar] [CrossRef]
- Armistead, J.S.; Arias, J.R.; Nishimura, N.; Lounibos, L.P. Interspecific larval competition between Aedes albopictus and Aedes japonicus (Diptera: Culicidae) in northern Virginia. J Med Entomol. 2008, 45. [Google Scholar]
- Costanzo, K.S.; Muturi, E.J.; Lampman, R.L.; Alto, B.W. The effects of resource type and ratio on competition with Aedes albopictus and Culex pipiens (Diptera:Culicidae). J Med Entomol. 2011, 48, 29–38. [Google Scholar] [CrossRef]
- Bonizzoni, M.; Gasperi, G.; Chen, X.; James, A.A. The invasive mosquito species Aedes albopictus: current knowledge and future perspectives. Trends Parasitol. 2013, 29, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Marini, G.; Guzzetta, G.; Baldacchino, F.; Arnoldi, D.; Montarsi, F.; Capelli, G.; et al. The effect of interspecific competition on the temporal dynamics of Aedes albopictus and Culex pipiens. Parasit Vectors. 2017, 10, 102. [Google Scholar] [CrossRef] [PubMed]
- Leisnham, P.T.; LaDeau, S.L.; Saunders, M.E.M.; Villena, O.C. Condition-Specific Competitive Effects of the Invasive Mosquito Aedes albopictus on the Resident Culex pipiens among Different Urban Container Habitats May Explain Their Coexistence in the Field. Insects 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Giatropoulos, A.; Papachristos, D.; Michaelakis, A.; Kapranas, A.; Emmanouel, N. Laboratory study on larval competition between two related mosquito species: Aedes (Stegomyia) albopictus and Aedes (Stegomyia) cretinus. Acta Trop. 2022, 230, 106389. [Google Scholar] [CrossRef]
- Rau, J.; Fischer, S.; Werner, D.; Kampen, H. Impact of larvae of the Asian tiger mosquito Aedes albopictus on larvae of the Culex pipiens complex from Germany in laboratory co-breeding studies. Med Vet Entomol. 2023, 37, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Andreadis, T.G.; Wolfe, R.J. Evidence for reduction of native mosquitoes with increased expansion of invasive Ochlerotatus japonicus japonicus (Diptera: Culicidae) in the northeastern United States. J Med Entomol. 2010, 47, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Hardstone, M.C.; Andreadis, T.G. Weak larval competition between the invasive mosquito Aedes japonicus japonicus (Diptera: Culicidae) and three resident container-inhabiting mosquitoes in the laboratory. J Med Entomol. 2012, 49, 277–285. [Google Scholar] [CrossRef] [PubMed]
- O'Donnell, D.L.; Armbruster, P. Comparison of larval foraging behavior of Aedes albopictus and Aedes japonicus (Diptera: Culicidae). J Med Entomol. 2007, 44, 984–989. [Google Scholar] [CrossRef]
- Reiskind, M.H.; Lounibos, L.P. Effects of intraspecific larval competition on adult longevity in the mosquitoes Aedes aegypti and Aedes albopictus. Med Vet Entomol. 2009, 23, 62–68. [Google Scholar] [CrossRef]
- Telang, A.; Qayum, A.A.; Parker, A.; Sacchetta, B.R.; Byrnes, G.R. Larval nutritional stress affects vector immune traits in adult yellow fever mosquito Aedes aegypti (Stegomyia aegypti). Med Vet Entomol. 2012, 26, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Bara, J.; Rapti, Z.; Caceres, C.E.; Muturi, E.J. Effect of Larval Competition on Extrinsic Incubation Period and Vectorial Capacity of Aedes albopictus for Dengue Virus. PLoS One. 2015, 10, e0126703. [Google Scholar] [CrossRef] [PubMed]
- Muturi, E.J.; Kim, C.H.; Alto, B.W.; Berenbaum, M.R.; Schuler, M.A. Larval environmental stress alters Aedes aegypti competence for Sindbis virus. Trop Med Int Health. 2011, 16, 955–964. [Google Scholar] [CrossRef]
- Takken, W.; Klowden, M.J.; Chambers, G.M. Effect of body size on host seeking and blood meal utilization in Anopheles gambiae sensu stricto (Diptera: Culicidae): the disadvantage of being small. J Med Entomol. 1998, 35, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Ratnayake, O.C.; Chotiwan, N.; Saavedra-Rodriguez, K.; Perera, R. The buzz in the field: the interaction between viruses, mosquitoes, and metabolism. Front Cell Infect Microbiol. 2023, 13, 1128577. [Google Scholar] [CrossRef] [PubMed]
- Arrese, E.L.; Soulages, J.L. Insect fat body: energy, metabolism, and regulation. Annu Rev Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [PubMed]
- Grimstad, P.R.; Walker, E.D. Aedes triseriatus (Diptera: Culicidae) and La Crosse virus. IV. Nutritional deprivation of larvae affects the adult barriers to infection and transmission. J Med Entomol. 1991, 28, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Hauser, G.; Thievent, K.; Koella, J.C. Consequences of larval competition and exposure to permethrin for the development of the rodent malaria Plasmodium berghei in the mosquito Anopheles gambiae. Parasit Vectors. 2020, 13, 107. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, M.; Czajka, C.; Borstler, J.; Melaun, C.; Jost, H.; von Thien, H.; et al. First nationwide surveillance of Culex pipiens complex and Culex torrentium mosquitoes demonstrated the presence of Culex pipiens biotype pipiens/molestus hybrids in Germany. PLoS One. 2013, 8, e71832. [Google Scholar] [CrossRef]
- Vereecken, S.; Vanslembrouck, A.; Kramer, I.M.; Muller, R. Phenotypic insecticide resistance status of the Culex pipiens complex: a European perspective. Parasit Vectors. 2022, 15, 423. [Google Scholar] [CrossRef]
- Müller, R.; Knautz, T.; Volker, J.; Kress, A.; Kuch, U.; Oehlmann, J. Appropriate larval food quality and quantity for Aedes albopictus (Diptera: Culicidae). J Med Entomol. 2013, 50, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Van Handel, E. Rapid determination of glycogen and sugars in mosquitoes. J Am Mosq Control Assoc 1985, 1, 299–301. [Google Scholar] [PubMed]
- Van Handel, E. Rapid determination of total lipids in mosquitoes. J Am Mosq Control Assoc 1985, 1, 302–304. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Bock, F.; Kuch, U.; Pfenninger, M.; Muller, R. Standardized Laboratory Feeding of Larval Aedes japonicus japonicus (Diptera: Culicidae). J Insect Sci 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Novak, M.G.; Higley, L.G.; Christianssen, C.A.; Rowley, W.A. Evaluating Larval Competition between Aedes-Albopictus and a-Triseriatus (Diptera, Culicidae) through Replacement Series Experiments. Environ Entomol. 1993, 22, 311–318. [Google Scholar] [CrossRef]
- Oberg, A.L.; Young, L.J.; Higley, L.G. A study of the statistical properties of two measures of competition. Applied statistics in agriculture 1994. [CrossRef]
- Harper, J.L. Population biology of plants; Academic Press: New York, 1977. [Google Scholar]
- Heitmann, A.; Jansen, S.; Luhken, R.; Leggewie, M.; Schmidt-Chanasit, J.; Tannich, E. Forced Salivation As a Method to Analyze Vector Competence of Mosquitoes. J Vis Exp 2018. [Google Scholar] [CrossRef]
- Huber, K.; Jansen, S.; Leggewie, M.; Badusche, M.; Schmidt-Chanasit, J.; Becker, N.; et al. Aedes japonicus japonicus (Diptera: Culicidae) from Germany have vector competence for Japan encephalitis virus but are refractory to infection with West Nile virus. Parasitol Res. 2014, 113, 3195–3199. [Google Scholar] [CrossRef]
- Lambert, A.J.; Lanciotti, R.S. Consensus amplification and novel multiplex sequencing method for S segment species identification of 47 viruses of the Orthobunyavirus, Phlebovirus, and Nairovirus genera of the family Bunyaviridae. J Clin Microbiol. 2009, 47, 2398–2404. [Google Scholar] [CrossRef]
- Chao, D.Y.; Davis, B.S.; Chang, G.J. Development of multiplex real-time reverse transcriptase PCR assays for detecting eight medically important flaviviruses in mosquitoes. J Clin Microbiol. 2007, 45, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Eshoo, M.W.; Whitehouse, C.A.; Zoll, S.T.; Massire, C.; Pennella, T.T.; Blyn, L.B.; et al. Direct broad-range detection of alphaviruses in mosquito extracts. Virology. 2007, 368, 286–295. [Google Scholar] [CrossRef]
- Yee, D.A.; Kesavaraju, B.; Juliano, S.A. Larval feeding behavior of three co-occurring species of container mosquitoes. J Vector Ecol. 2004, 29, 315–322. [Google Scholar] [PubMed]
- Vanslembrouck, A.; Scheers, K.; Vermeersch, X.; Hendrickx, R.; Schneider, A.; De Witte, J.; et al. Exploring the efficacy of predaceous diving beetles as potential nature-based solution for combating the invasive mosquito Aedes albopictus. Neobiota. Under review.
- Giunti, G.; Becker, N.; Benelli, G. Invasive mosquito vectors in Europe: From bioecology to surveillance and management. Acta Trop. 2023, 239, 106832. [Google Scholar] [CrossRef]
- Perera, R.; Riley, C.; Isaac, G.; Hopf-Jannasch, A.S.; Moore, R.J.; Weitz, K.W.; et al. Dengue virus infection perturbs lipid homeostasis in infected mosquito cells. PLoS Pathog. 2012, 8, e1002584. [Google Scholar] [CrossRef] [PubMed]
- Chotiwan, N.; Andre, B.G.; Sanchez-Vargas, I.; Islam, M.N.; Grabowski, J.M.; Hopf-Jannasch, A.; et al. Dynamic remodeling of lipids coincides with dengue virus replication in the midgut of Aedes aegypti mosquitoes. PLoS Pathog. 2018, 14, e1006853. [Google Scholar] [CrossRef] [PubMed]
- Koh, C.; Islam, M.N.; Ye, Y.H.; Chotiwan, N.; Graham, B.; Belisle, J.T.; et al. Dengue virus dominates lipid metabolism modulations in Wolbachia-coinfected Aedes aegypti. Commun Biol. 2020, 3, 518. [Google Scholar] [CrossRef]
- Paige, A.S.; Bellamy, S.K.; Alto, B.W.; Dean, C.L.; Yee, D.A. Linking nutrient stoichiometry to Zika virus transmission in a mosquito. Oecologia. 2019, 191, 1–10. [Google Scholar] [CrossRef]
Figure 1.
Relative Crowding Coefficient with advantage or disadvantage. a) development time for 50% of the pupae to emerge, b) the pupal size, c) lipid, d) glycogen, e) protein content (size corrected) of combination 1) Ae. albopictus vs Cx. pipiens s.s./Cx. torrentium, and 2) Ae. japonicus vs Cx. p. molestus during interspecific competition at 20°C and 26°C. In red are the differences with one of the species having a RCC below 1 indicating out-competition, according to Oberg, Young [51].
Figure 1.
Relative Crowding Coefficient with advantage or disadvantage. a) development time for 50% of the pupae to emerge, b) the pupal size, c) lipid, d) glycogen, e) protein content (size corrected) of combination 1) Ae. albopictus vs Cx. pipiens s.s./Cx. torrentium, and 2) Ae. japonicus vs Cx. p. molestus during interspecific competition at 20°C and 26°C. In red are the differences with one of the species having a RCC below 1 indicating out-competition, according to Oberg, Young [51].
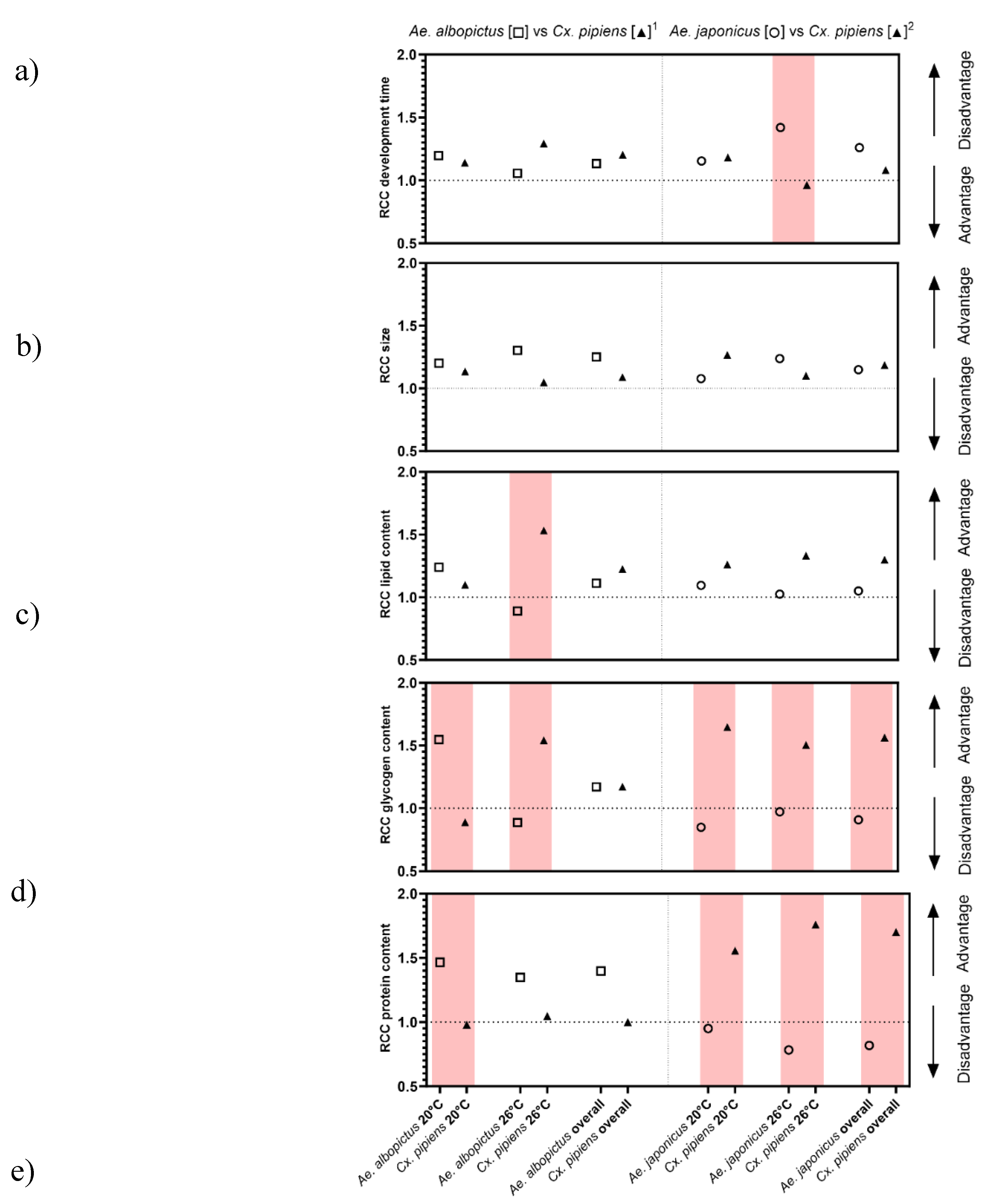
Figure 2.
Relative Crowding Coefficient on activity or inactivity. Behavior variables total distance moved, velocity and duration of the body contact between intra- and interspecific larval competition ratios of Ae. albopictus, Cx. pipiens s.s./Cx. torrentium, Ae. japonicus, and Cx. p. molestus. In red are the differences with one of the species having a RCC below 1 indicating significant differences in behavior in a competitive environment.
Figure 2.
Relative Crowding Coefficient on activity or inactivity. Behavior variables total distance moved, velocity and duration of the body contact between intra- and interspecific larval competition ratios of Ae. albopictus, Cx. pipiens s.s./Cx. torrentium, Ae. japonicus, and Cx. p. molestus. In red are the differences with one of the species having a RCC below 1 indicating significant differences in behavior in a competitive environment.
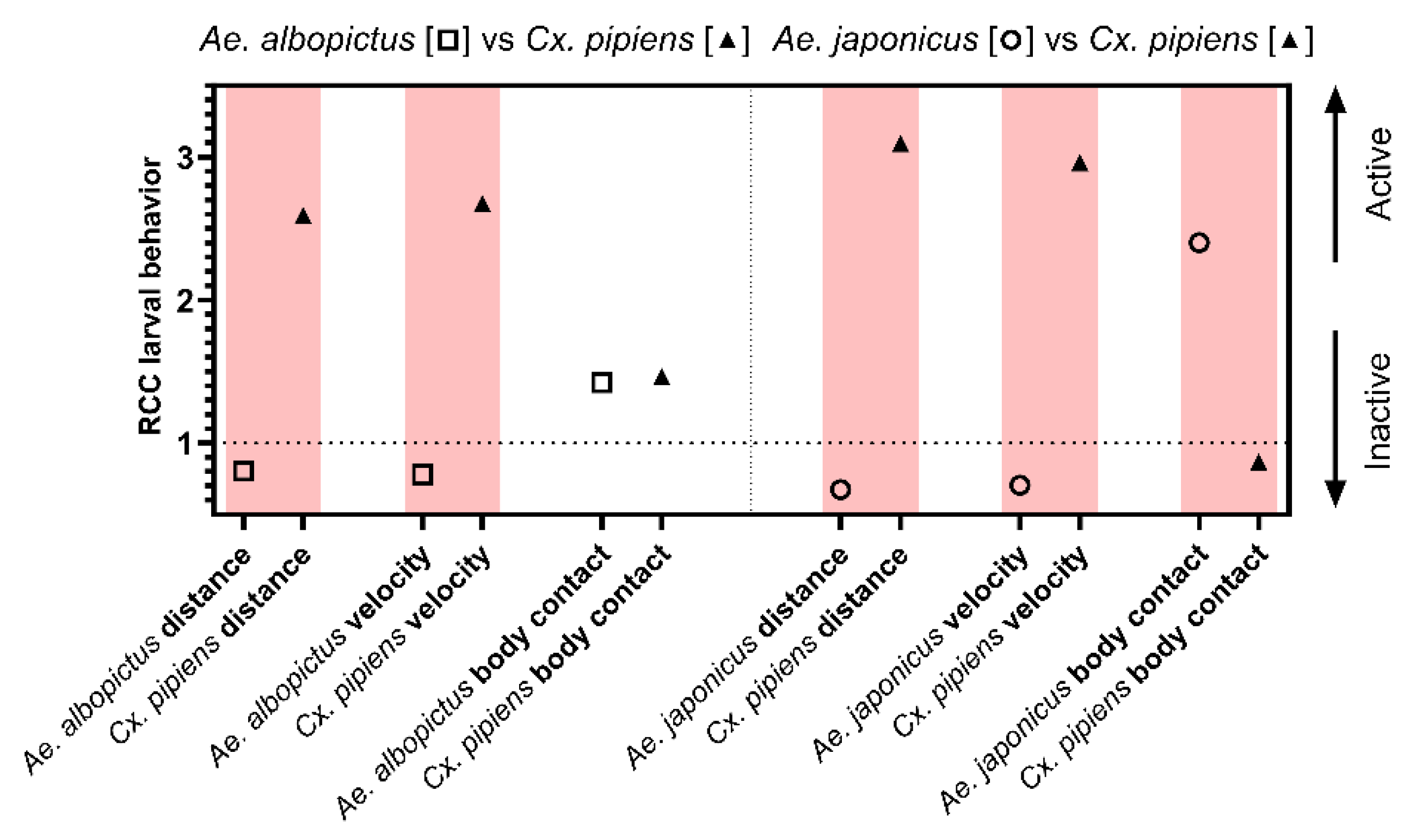
Figure 3.
Mean viral RNA body titer per specimen and infection rate per species ratio. a) Mean CHIKV RNA body titer per specimen of Ae. albopictus, b) Infection rate of WNV of both Cx. pipiens strains, c) Mean WNV RNA body titer per specimen of Cx. pipiens s.s./Cx. torrentium (see Zenodo repository in data availability statement for separated results for bioforms) from Ae. albopictus combination, d) Mean WNV RNA body titer per specimen of Cx. p. molestus from Ae. japonicus combination. Infection rate was 100% and 0% for all Ae. albopictus and Ae. japonicus respectively, body titer was 0 genome copies per mosquito for Ae. japonicus.
Figure 3.
Mean viral RNA body titer per specimen and infection rate per species ratio. a) Mean CHIKV RNA body titer per specimen of Ae. albopictus, b) Infection rate of WNV of both Cx. pipiens strains, c) Mean WNV RNA body titer per specimen of Cx. pipiens s.s./Cx. torrentium (see Zenodo repository in data availability statement for separated results for bioforms) from Ae. albopictus combination, d) Mean WNV RNA body titer per specimen of Cx. p. molestus from Ae. japonicus combination. Infection rate was 100% and 0% for all Ae. albopictus and Ae. japonicus respectively, body titer was 0 genome copies per mosquito for Ae. japonicus.
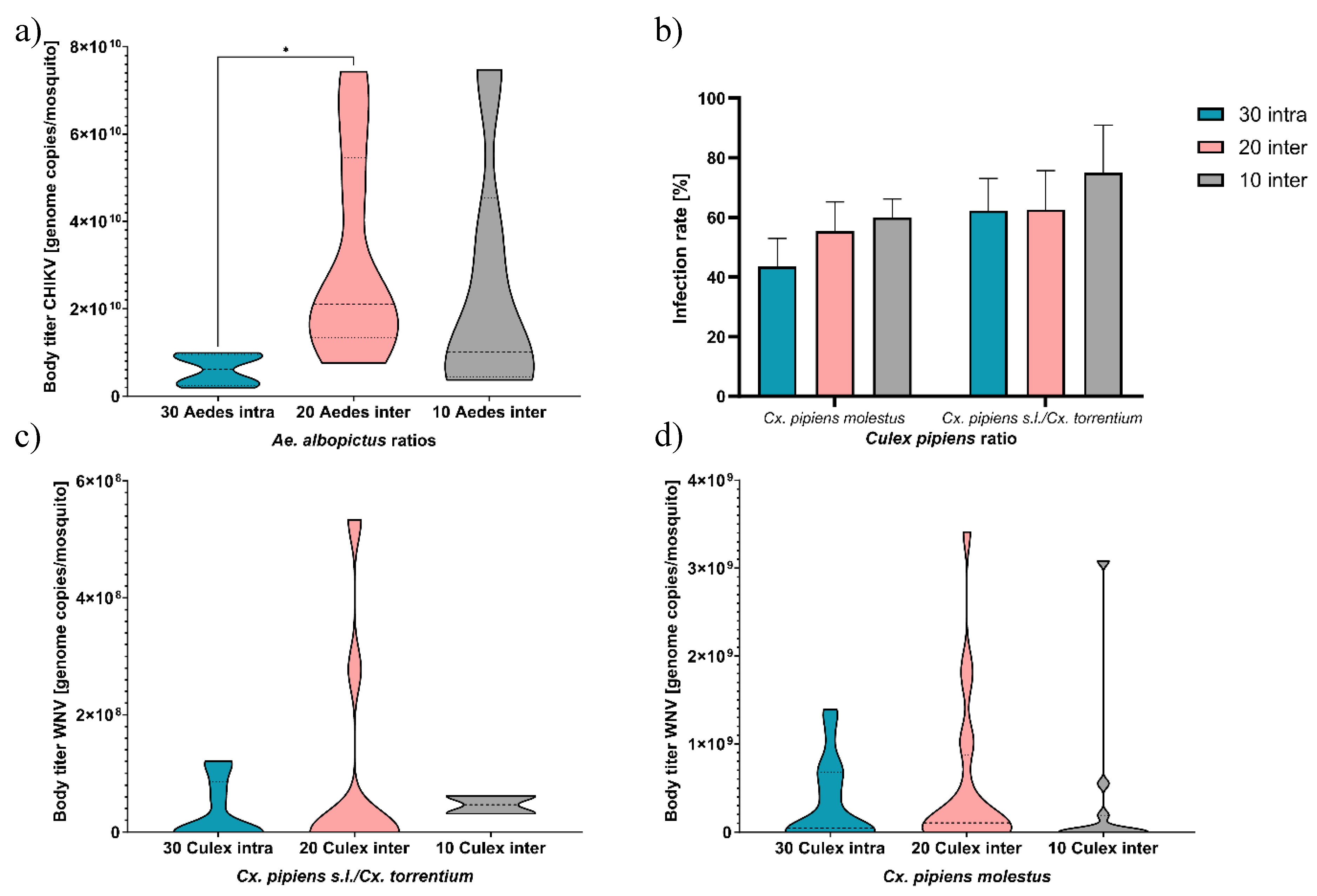
Figure 4.
Culex species and Cx. pipiens biotypes used during infection study.
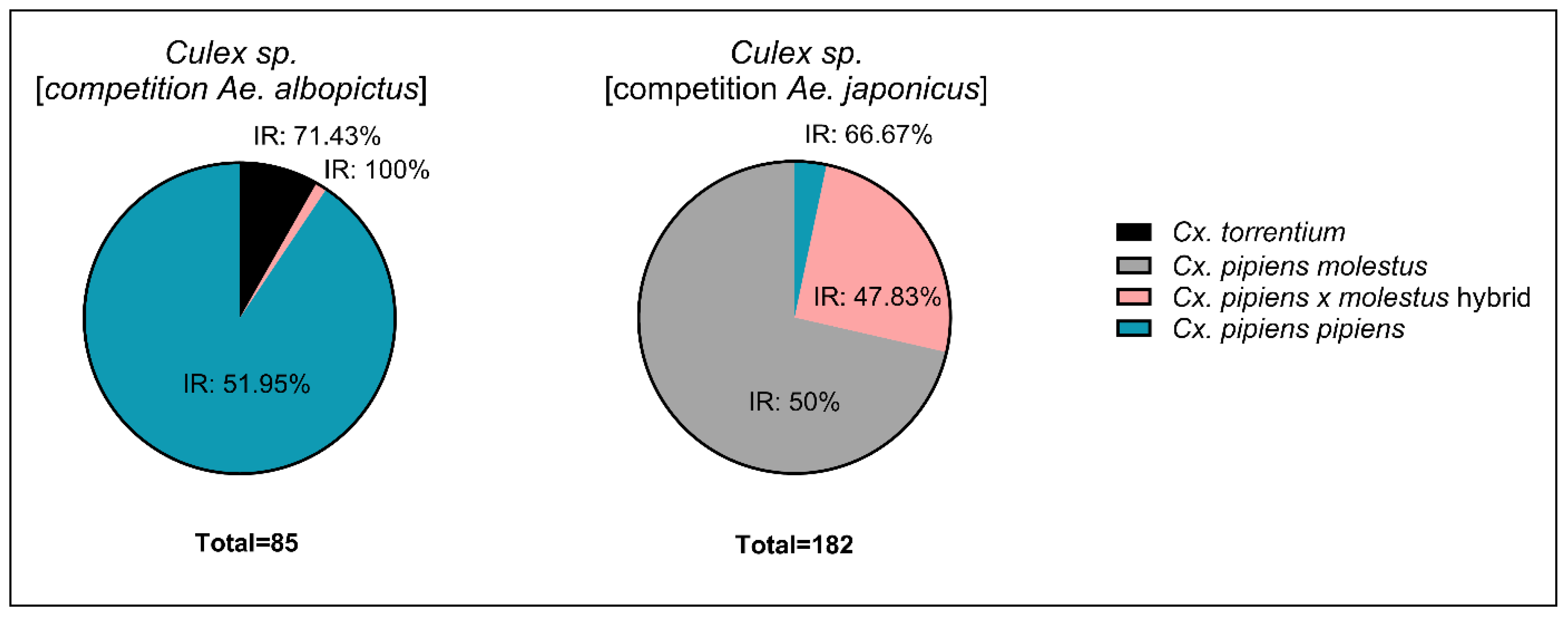
Figure 5.
Principal Component Analysis (PCA) of total data set with variables. Development time, larval ratio, mortality, larval ratio, cephalothorax length (CL), abdominal length (AL), abdominal width (AW), lipid (L), protein (P), glycogen (G), mortality, total distance moved, velocity, body contact, viral RNA body titer (BT), and infection rate (IR) for different species competition treatments in a, b) Ae. albopictus and Cx. pipiens s.s./Cx. torrentium and c, d) Ae. japonicus and Cx. p. molestus combinations at 26°C. Infection rate is not included for Ae. albopictus and Ae. japonicus, viral RNA body titer is not included for Ae. japonicus.
Figure 5.
Principal Component Analysis (PCA) of total data set with variables. Development time, larval ratio, mortality, larval ratio, cephalothorax length (CL), abdominal length (AL), abdominal width (AW), lipid (L), protein (P), glycogen (G), mortality, total distance moved, velocity, body contact, viral RNA body titer (BT), and infection rate (IR) for different species competition treatments in a, b) Ae. albopictus and Cx. pipiens s.s./Cx. torrentium and c, d) Ae. japonicus and Cx. p. molestus combinations at 26°C. Infection rate is not included for Ae. albopictus and Ae. japonicus, viral RNA body titer is not included for Ae. japonicus.
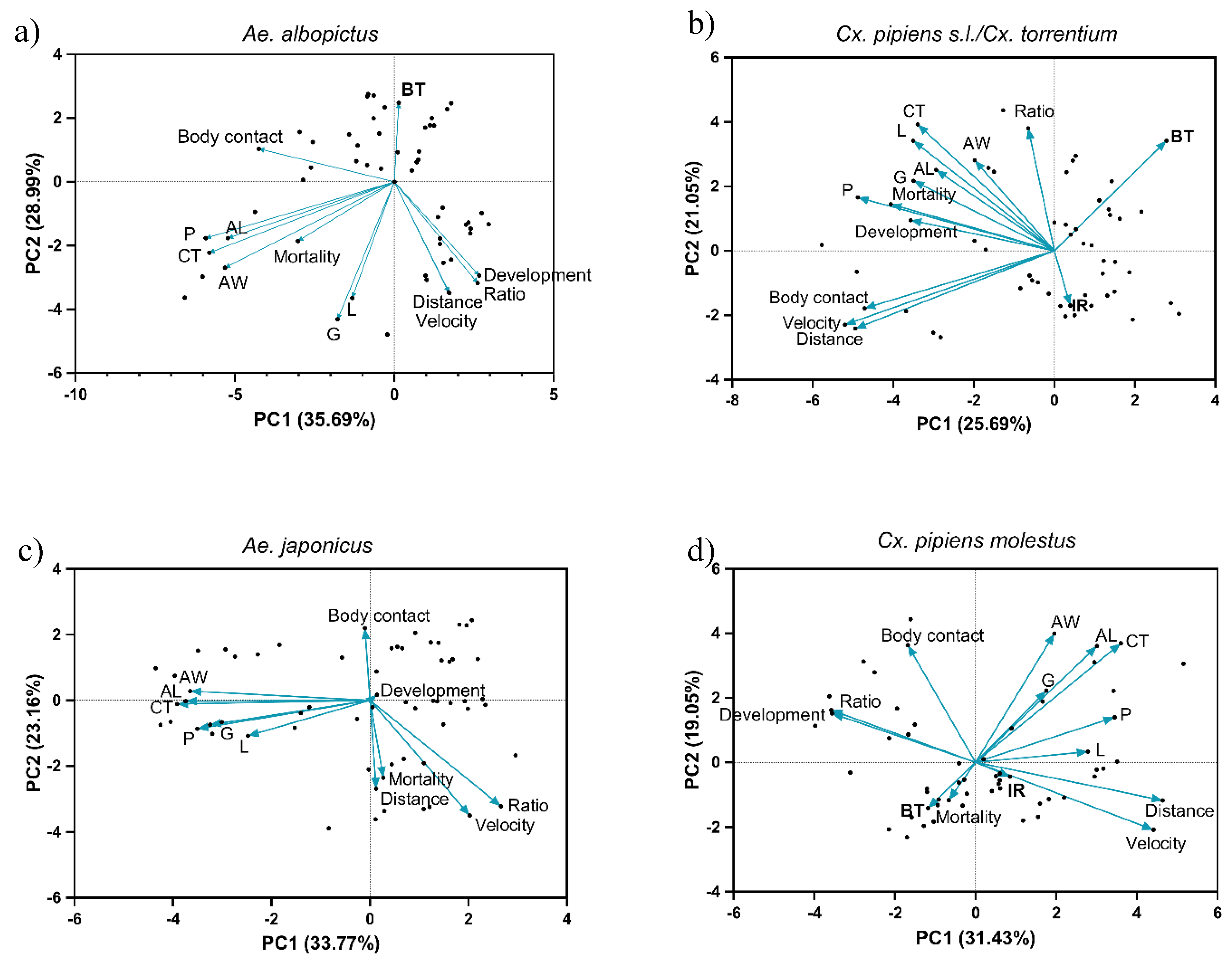

Table 1.
Infection rates (IR), viral RNA copy number/body (mean log10 RNA copies/specimen), and statistical significance for Ae. albopictus, Cx. pipiens s.s./Cx. torrentium, and Cx. p. molestus.
Table 1.
Infection rates (IR), viral RNA copy number/body (mean log10 RNA copies/specimen), and statistical significance for Ae. albopictus, Cx. pipiens s.s./Cx. torrentium, and Cx. p. molestus.
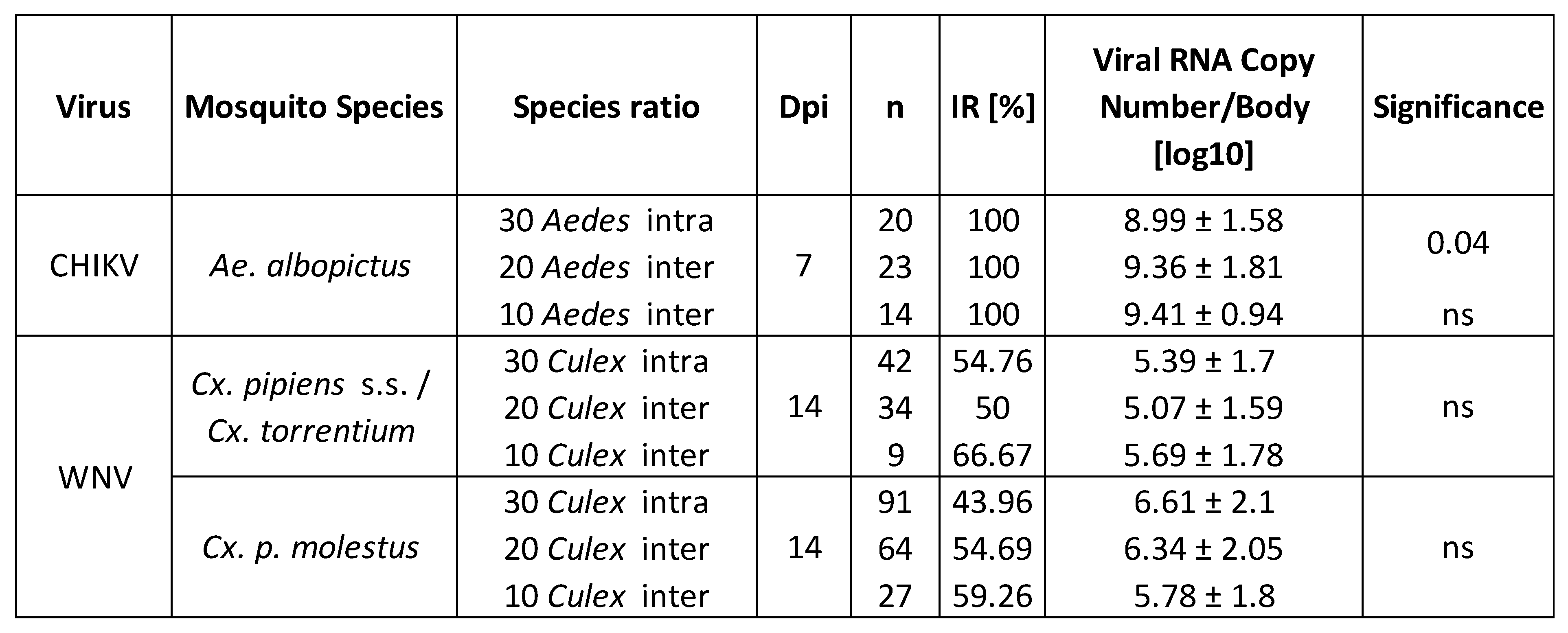
Table 2.
Overview of the loadings per variable for each species and principal component (PC). The six highest variables (development time, larval ratio, mortality, larval ratio, cephalothorax length (CL), abdominal length (AL), abdominal width (AW), lipid (L), protein (P), glycogen (G), mortality, total distance moved, velocity, body contact, viral RNA body titer (BT), and infection rate (IR)) per species and principal component are included in this overview.
| PC1 | |||||||
| Ae. albopictus | Cx. pipienss.s./Cx. torrentium | Ae. japonicus | Cx. p. molestus | ||||
| Development | 0.78 | Development | -0.51 | L | -0.58 | Distance | 0.81 |
| Ratio | 0.77 | Mortality | -0.58 | G | -0.76 | Velocity | 0.77 |
| AL | -0.71 | Body contact | -0.67 | P | -0.82 | CT | 0.63 |
| AW | -0.73 | P | -0.69 | AW | -0.86 | P | 0.60 |
| CT | -0.79 | Distance | -0.70 | AL | -0.88 | Development | -0.83 |
| P | -0.81 | Velocity | -0.74 | CT | -0.92 | Ratio | -0.84 |
| PC2 | |||||||
| Ae. albopictus | Cx. pipienss.s./Cx. torrentium | Ae. japonicus | Cx. p. molestus | ||||
| BT | 0.58 | CT | 0.57 | Body contact | 0.61 | AW | 0.68 |
| Ratio | -0.59 | Ratio | 0.55 | L | -0.27 | CT | 0.63 |
| Velocity | -0.65 | BT | 0.50 | Mortality | -0.59 | Body contact | 0.62 |
| Distance | -0.65 | L | 0.50 | Distance | -0.68 | AL | 0.62 |
| L | -0.68 | Velocity | -0.63 | Ratio | -0.81 | BT | -0.38 |
| G | -0.80 | Distance | -0.66 | Velocity | -0.88 | Velocity | -0.56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



