
Submitted:
26 June 2024
Posted:
27 June 2024
You are already at the latest version
Abstract
Bacillus cereus is a gram-positive bacterium widespread in the environment, especially in soil and dust. It produces two types of toxins causing vomiting and diarrhea. Nowadays, food borne outbreaks due to Bacillus cereus group bacteria (especially Bacillus cereus sensu stricto) are increased, as reported by European Food Safety Authority, representing a very huge problem in agri-food chain. In this work, we analyzed 118 strains belonging to Bacillus cereus group, isolated from several food sources (fruit and vegetables, dairy products, bakery products) concerning their susceptibility to antibiotics currently most used to treat infections. Many strains showing an intermediate susceptibility to clindamycin, erythromycin, and tetracycline were detected, suggesting an evolving acquisition of resistance against these antibiotics. Moreover, one strain showed intermediate resistance to meropenem, another antibiotic currently used to treat infections caused by Bacillus cereus. Beside the antimicrobial characterization, all strains were studied comparing their antimicrobic phenotype with the presence/absence of antimicrobial genes in their genome. The analysis showed a not complete correlation between genes carried by the strains and their phenotype, demonstrating that the antibiotic resistance is due not only to genetic factors, but also to other factors such as the inappropriate use of antibiotics that can determine an acquired resistance for bacteria.
Keywords:
Antibiotic resistance
; Bacillus cereus group
; Food poisoning
; Minimum Inhibitory Concentration
; WGS
1. Introduction
The Bacillus cereus group, also named Bacillus cereus sensu lato, is an heterogenous group of aerobic or facultative anaerobes bacteria, consisting in several species phylogenetically correlated [1,2]. They are Gram positive, ubiquitous in the environment and can grow up at optimal temperature ranging from 30 to 40°C and in the range pH between 5 and 8.8 [3]. Most strains are catalase positive and mobile [1]. One of the peculiar characteristics of these microorganisms is their ability to form spores, metabolically dormant cell types, resistant to extreme conditions including heat, freezing, drying and radiation (commonly used in food industry) [4,5].
In the environment, they populate all kinds of soils and waters, sediments, plants in the spore form, but they are also detected in animals [6,7]. Because of its widespread in environment, Bacillus spores could contaminate raw food ingredients (vegetables, milk, fruit, spices, cereals) employed during food processing. Thus, a wide variety of finished food products might contain these bacteria and germination/outgrowth during storage is also possible, causing foodstuff spoilage [8].
The B. cereus group consists of several species, including Bacillus cereus sensu scricto, Bacillus thuringensis, Bacillus weihenstephanensis, Bacillus wiedmanni, and Bacillus toyonensis [2,9,10].
B. cereus sensu scricto (B. cereus s.s.) is the model species of the Bacillus cereus group. It can complete a full saprophytic life cycle, but also be an opportunistic human pathogen [11] that causes gastrointestinal illness, bacteremia, endocarditis, respiratory and urinary tract infections, endophthalmitis and meningitis [2,10,12]. Moreover, B. cereus s.s. is one of the most common pathogens in food poisoning [13]. It causes two type of food poisoning: the diarrhea and the emetic syndrome. The diarrhea syndrome is caused by a complex of three toxins (hemolysin BL, nonhemolytic enterotoxin and cytotoxic protein), while the emetic syndrome is due to cereulide, a thermostable protein [11].
B. thuringiensis is an entomopathogen affecting many orders of insect and for this reason used as biopesticides [10]. More recently, however, B. thuringiensis was found involved in various infectious diseases in humans, such as food-poisoning-associated diarrheas and wound infections [14].
B. weihenstephanensis generally is non-pathogenic saprophyte or rarely associated with diseases in humans or other animals. Some strains of B. weihenstephanensis could produce the emetic toxin called cereulide (like B. cereus s.s.) responsible of food intoxication accompanied by vomiting [15].
Concerning B. wiedmanni, it is not considered a human pathogen; however, Miller and colleagues [16] characterized the strain FSL W8-0169T, isolated from dairy foods and dairy environments as a potential pathogen being capable of expressing the toxins HBL and the non-haemolytic toxin NHE.
At last, B. toyonensis has been isolated from pathogenic intraocular specimen, demonstrating its ability to cause endophthalmitis [17].
B. cereus s.s. and related bacteria are considered manly responsible of foodborne diseases. According to EFSA, B. cereus originates an important number of food-borne illnesses in humans [18] and the real number of cases is underestimated, at the date. In particular, B. cereus s.s. has been incriminated as a cause of toxin-induced emetic and diarrheagenic syndromes after ingestion [1] which represent the foremost worries for the public Health service [19], considering the different spectrum of diseases provoked (from gastrointestinal forms that require short time of recovery to most serious systemic diseases like bacteremia and septicemia that could have a fatal outcome). Generally, B. cereus related infections symptoms start from 0.5 to 16 hours after ingestions of contaminated food and disappear within 24 hours and do not require drug treatments [20]. Severe B. cereus infections were treated with antibiotics, but an excessive and wrong use of these molecules could lead to antibiotic resistance phenomenon [21]. Nowadays, antibiotic resistance is a huge problem in public health and a global priority because the infections caused by resistant bacteria are harder to treat than those caused by non-resistant ones, leading to higher medical costs, prolonged hospital stays, and increased mortality [22].
Hence, active surveillance of food contamination is necessary. Moreover, an antibiotic resistance surveillance plan is also important, not only concerning clinically relevant bacteria but also for other pathogens, like B. cereus group members, in order to prevent new infections and to know how dealing with them in presence of resistant bacteria.
In this study we analyzed different food sources regarding the presence of bacteria belonging to Bacillus cereus group to investigate the presence of these in different food sources and we evaluated in vitro their sensitivity to antimicrobials, also correlating these results with genetic analyses.
2. Results
2.1. Isolation of B. cereus Group Strains
In the analyzed samples, the isolation of B. cereus colonies was evidenced by the presence of typical color morphology, being dull gray and opaque, with a rough matted surface and irregular perimeters, with zones of hemolysis surrounding colonies.
2.2. MALDI-TOF Mass Spectrometry (MS) Analysis
By MALDI-TOF MS, all the analyzed strains were identified as B. cereus using the commercial BDAL library (MBT Compass library v 7.0.0.0) with a log(score) generally between 1.7 and 2.0, accompanied by the following comment: “Bacillus anthracis, cereus, mycoides, pseudomycoides, thuringiensis and weihenstephanensis are closely related and members of the Bacillus cereus group. In particular, Bacillus cereus spectra are very similar to spectra from Bacillus anthracis. Bacillus anthracis is not included in the MALDI Biotyper database. For differentiation, an adequate identification method must be selected by an experienced professional. The quality of spectra (score) depends on the degree of sporulation: Use fresh material”.
2.3. WGS Analysis
In detail, 68 B. cereus sensu stricto, 23 B.thuringensis, 19 B. wiedmannii, 7 B. toyonensis and 1 B.weihenstephanensis were identified by WGS. As regards the food matrices from which strains were isolated, 42 B. cereus sensu stricto were detected in milk and dairy products (including ice cream), 11 in fruit and vegetables, 7 in bakery products, 3 in fish-based products, 2 in meat and 3 in mixed plates (russian salad, focaccia with scamorza, sandwich with cured meat).
We also identified other members of B. cereus s.l.: B. thuringensis, B. toyonensis, B. wiedmanni and B. weihestephanensis. These species were identified mainly in milk and its derivates (16 B. thuringensis, 13 B. wiedmanni, 3 B. toyonensis, and 1 B. weihestephanensis) and vegetables (3 B. thuringensis, 1 B. wiedmanni, 3 B. toyonensis). Moreover, they were isolated also from meat (1 B. thuringensis and 1 B. wiedmanni), fish (1 B. thuringensis and 1 B. wiedmanni) and bakery products (2 B. thuringensis, 2 B. wiedmanni and 1 B. toyonensis).
In addition, 1 B. wiedmanni was found in a box of ravioli, a kind of Italian pasta, usually filled with meat, cheese, fish, vegetables or with a mix of them.
An overview of the identified strains, and of the food sources was provided in Table S1.
The bioinformatic analysis allowed us to predict genes responsible of antimicrobic resistance (Figure 1 and Table S2).
All strains possess at least two β-lactams resistance genes. In detail, we have found bcI in 68/118 (58%), bcII in 82/118 (69%), bla in 116/118 (98%), bla1 in 116/118 (98%), bla2 in 114/118 (97%), blaP in 32/118 (27%) and blaZ in 60/118 (51%).
2% of isolates (2/118) carried fluoroquinolone genes (arlS) and 2% (2/118) carried catA gene responsible for chloramphenicol resistance.
Macrolide resistance genes Bacillus_cluster_A_intrinsic_mph, Bacillus_cluster_B_intrinsic_mph and mph(B) were found in 52/118 (44%), 4/118 (3%) and 52/118 (44%) respectively.
Moreover, 25/118 (21%) carried tetracycline resistance gene (tetL) and 58/118 (49%) possess clindamycin resistance gene (lsaB)
Others AMR genes identified were vancomycin resistance genes: vanR (118/118 strains, 100%), vanRA (15/118 strains, 16%), vanRB (10/118 strains, 8%), vanRF (17/118 strains, 14%), vanRM (101/118 strains, 86%), vanR-Pt (15/118 strains, 13%), vanS (15/118 strains, 13%), vanS-Pt2 (14/118 strains, 12%), vanW (0/118 strains, 0%), vanYA (7/118 strains, 6%), vanYF (10/118 strains, 8%), vanY-Pt (15/118 strains, 13%), vanZF (106/118 strains, 90%).
2.4. Antibiotic Susceptibility Analysis
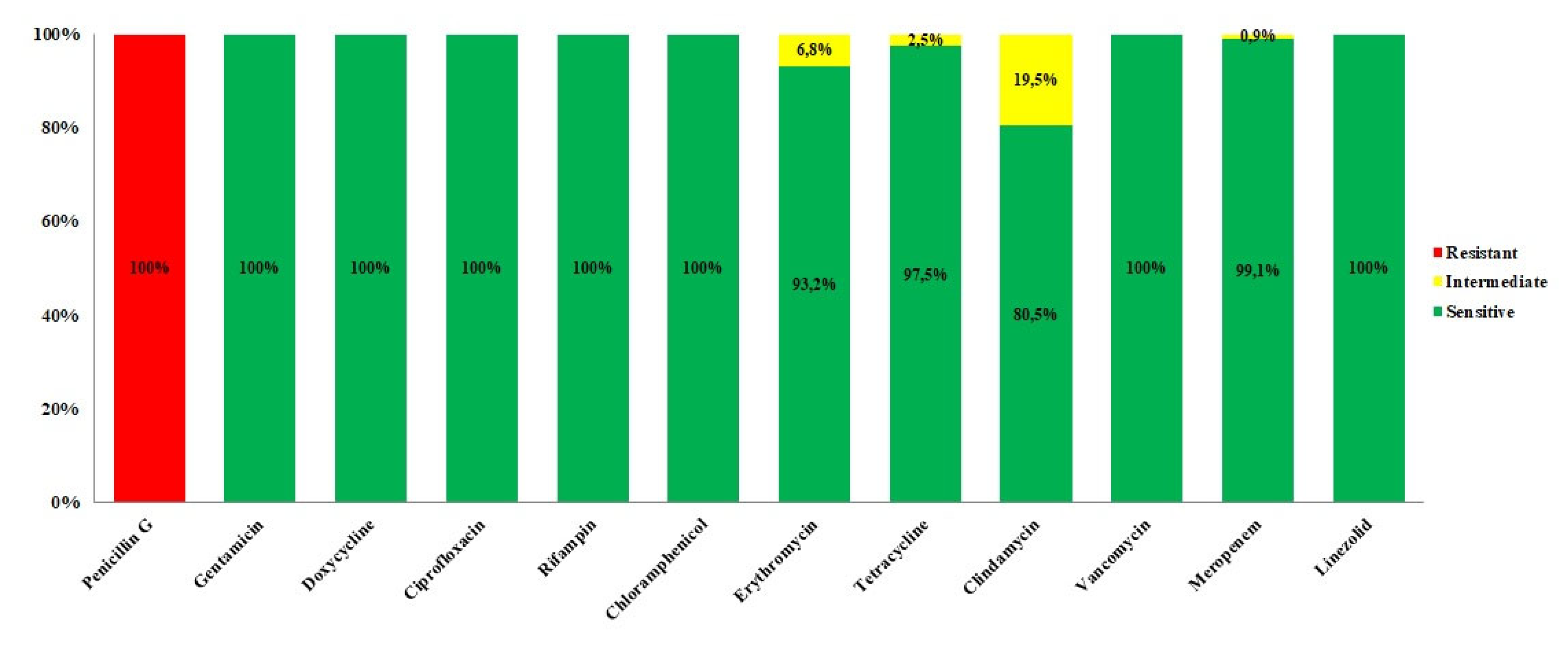
After 16h of incubation at 37°C, the 96 well plates have been read and MIC values checked. All the 118 strains (100%) showed resistance to penicillin G, according to literature [23] while were sensitive against gentamicin, doxycycline, ciprofloxacin, rifampicin, chloramphenicol, vancomycin, and linezolid. Concerning meropenem, all strains resulted to be susceptible to this antibiotic, except for one strain (0.9%) (BC399A) that showed an intermediate behavior.
Moreover, 3/118 strains (2.5%) showed intermediate resistance to tetracycline (BC290A, BC329A, BC281A), 8/118 (6.8%) to erythromycin (BC2D, BC56B, BC93B, BC147A, BC166A, BC282A, BC398C, BC423A) and 23/118 (19.5%) to clindamycin (BC2A, BC4D, BC23B, BC25B, BC41A, BC56B, BC83B, BC85A, BC91C, BC156C, BC171A, BC187A, BC204A, BC223B, BC313A, BC329A, BC246A, BC266A, BC324A, BC335A, BC375A, BC391A, BC404A).
We observed some strains showing contemporary intermediate susceptibility towards two or more antibiotics. Specifically, the strain BC56B had intermediate resistance to erythromycin and clindamycin and the strain BC329A to both tetracycline and clindamycin.
3. Discussion
B. cereus s.s. can easily contaminate various types of food. It has been frequently isolated in milk and dairy products, meat and derivatives and additionally in plant origin products (i.e. vegetables ready-to-eat or to be used as ingredient) as a consequence of soil contamination [4]. In our study, we identified B. cereus s.s. as the main member of Bacillus cereus s.l. involved in food contamination. Sixty-eight strains were identified as B. cereus s.s.; most of them (42 strains) were isolated from milk and dairy products (i.e. mozzarella, scamorza, ricotta), followed by vegetables and bakery products (flour, biscuits, sweets). Also, meat and fish resulted contaminated by B. cereus s.s., even if to a lesser extent.
Moreover, B. cereus s.s. was also found in mixed sources composed by mixed elements (sandwich with cured meats, russian salad and focaccia with scamorza) that makes it difficult to determine which single food is contaminated and at what level contamination may have occurred (at primary food level or during industry processing). Other members of Bacillus cereus group identified in food matrices in this study were B. thuringensis, B. toyonensis, B. wiedmanni and B. weihestephanensis.
Bacillus species overmentioned are not commonly human pathogens and useful particularly in the agricultural field [24,25,26,27]. These bacteria, even if are underestimated as risk for human health, can represent a serious risk in food safety. However, they could cause food poisoning, since some strains are able to produce toxins and virulence factors like B. cereus s.s. [14,15,16].
Bacteria can acquire ARGs through horizontal gene transfer (HGT) that involve different type of genetic elements (plasmids, integrons and transposons) but also different way of transfer (transformation, conjugation, transduction) [28]. Transfer could undergo among the same species or different ones, and it is also possible that residual DNA carrying ARGs could persist in environment for a long time even after the death of resistant strains, with the possibility to transfer them to other strains [29]. This is the reason why the investigation of antimicrobic susceptibility is interesting also for not-highly pathogenic and not pathogenic bacteria like B. cereus group members.
Concerning antimicrobial susceptibility, all of 118 isolated strains were phenotypic sensitive to gentamicin, doxycycline, ciprofloxacin, rifampicin, chloramphenicol, vancomycin and linezolid according to data actually in literature [19,30,31,32]. Gentamicin, vancomycin and linezolid are commonly used in severe B. cereus infections [33] and our results confirmed the efficacy of these antibiotics.
Clindamycin is another antibiotic commonly used in treatment of B. cereus infections [33]. In our study, 23 strains (15 B. cereus s.s., 6 B. thuringensis and 2 B. wiedmanni) (20,3% of total strains) showed phenotypic intermediate susceptibility to clindamycin, suggesting an evolving acquisition of antibiotic resistance, in agreement with other studies [30,32,33].
Furthermore, 8 strains (4 B. cereus s.s., 2 B. thuringensis, 1 B. wiedmanni and 1 B. toyonensis), corresponding to 6,8% of strains, showed phenotypic intermediate susceptibility to erythromycin, according to current literature [9,23,32]; while a lower number of strains (3 isolates of B. cereus s.s., 2,5% of strains), showed an intermediate susceptibility to tetracycline, as previously reported [30,34,35].
We also observed an interesting intermediate behavior of only 1 B. wiedmanni strain against meropenem, an antibiotic often used in clinical practice. Meropenem belongs to carbapenems drug class, β-lactam antibiotics that are active against many aerobic and anaerobic gram-positive and gram-negative bacteria. It works against extended-spectrum β-lactamases [36], but may be more susceptible to metallo-β-lactamases. Probably, its intermediate behavior is due to the presence of the bla2, a chromosomal-encoded β-lactamase, which has penicillin, cephalosporin, and carbapenem-hydrolizing abilities [37].
Alongside these results, in this study we also demonstrated that often there is a not strict correlation between genes carried by bacterial strains (Table S2) and phenotypic characteristics (Table S3). For example, most B. cereus group strains isolated in this work carried vancomycin resistance genes, but all of them resulted susceptible to this antibiotic.
Concerning the 8 strains with intermediate resistance to erythromycin, all of them carried genes responsible for resistance to this molecule, but other strains did not show an intermediate or resistant phenotype even if they possess resistance gene to macrolide class.
Likewise, only 3 strains possess the gene responsible to tetracycline (tetL) and at the same time show an intermediate susceptibility to this antibiotic.
All these phenotypic and genetic data collected during this study are not surprising, considering the so called “silent resistome” [38,39,40]. The word resistome indicates all genes able to confer single or multiple antimicrobial resistance [41] and includes constitutively expressed genes, precursors and acquired AMR genes [40].
Particularly interesting is the concept of silent antimicrobial resistance genes, also called cryptic genes. These are plasmid or chromosomal DNA sequences, carried by bacteria, that are not normally expressed or expresses at very low level, without a non-correspondence to phenotypic resistance to the antibiotics [39,40]. Expression of AMR genes, usually, is not active because of the metabolic cost of this process and for this reason from the bacterial perspective silencing is very essential to preserve fitness. This phenomenon could explain why many strains of this study did not show alteration of antibiotic susceptibility even if they possess AMR genes.
In addition, we observe a very low percentage (1.7%) of strains showing an intermediate susceptibility against two different antibiotics simultaneously (2/118 strains), suggesting that also in B. cereus group bacteria a progressive acquisition of multidrug resistance could happen, leading to antibiotic treatment failure.
4. Materials and Methods
4.1. B. cereus Group Isolated Strains
A total of 118 B. cereus group strains analyzed in this study were isolated from different types of food (dairy products, vegetables and fruit, fish and meat, bakery products), collected in different production and distribution sites of different cities of Apulia and Basilicata regions, in Southern Italy.
Cultivation was performed according to ISO 21871:2006 which specifies the method for the detection of Bacillus cereus from products intended for human consumption. Briefly, 5 g of food matrix were added to 45 mL of Buffered Peptone Water (BPW) (Biolife Italiana, Milan, Italy) and homogenized using a stomacher (PBI International, Milan, Italy) at 230 rpm for 30s. Then, 45 mL of double-strength Tryptone Soy Polymyxin Broth (TSPB) (Biolife Italiana, Milan, Italy) were added and samples were incubated at 30°C for 48 h. After incubation, 10 mL of enrichment broth were streaked onto the surface of solid selective medium Mannitol Egg Yolk Polymyxin Agar (MYP) (Biolife Italiana, Milan, Italy) and the plates were incubated at 30°C for 24-48h under aerobic condition. Then, typical presumptive Bacillus cereus group colonies for each sample were picked and subcultured on Columbia Agar with 5% sheep blood. After incubation step at 37°C for 18–24 h, the bacterial isolates were subjected to Matrix-Assisted Laser Desorption/Ionization Time-Of-Flight Mass Spectrometry (MALDI-TOF MS) [42] and subsequently to DNA extraction using the DNAeasy Blood and Tissue kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol for Gram-positive bacteria. Identification and typing of all strains were performed using Whole Genome Sequencing (WGS) method [30].
4.2. MALDI-TOF Mass Spectrometry Analysis
As first screening, MALDI-TOF MS analysis was performed for a fast identification of the Bacillus cereus group bacteria. The bacterial colony was directly transfer using a toothpick on the 96-well target plate (Bruker Daltonics, Germany), covered with 1 μL of 70% formic acid, dried at room temperature, and then overlaid with 1 μL of an α-cyano-4-hydroxycinnamic acid (CHCA) matrix solution (a saturated solution of α-cyano-4-hydroxycinnamic acid in in 50/50 [v/v] of acetonitrile/H2O containing 2.5% trifluoroacetic acid) and allowed to dry. The mass spectra were acquired using Microflex LT/SH™ mass spectrometer (Bruker Daltonik GmbH, Bremen, Germany), operating in linear positive mode covering a mass range between 2 and 20kDa. For validation of runs, the instrument was calibrated using Bruker Bacterial Test Standard (BTS, Bruker Daltonik GmbH, Bremen, Germany), an extract of the Escherichia coli DH5α strain spiked with two additional pure proteins (RNase A of 13,683.2 Da and myoglobin of 16,952.3 Da).
4.3. WGS Sequencing and Bioinformatic Analyses
Typing analysis of the 118 strains was performed by WGS, since MALDI-TOF MS is not definitively exhaustive for the identification of the bacteria belonging to Bacillus cereus group. Extracted DNA quality and concentrations were estimated by Qubit 3.0 using Qubit dsDNA HS Assay (Thermo Fisher Scientific). Libraries were prepared using the DNA Prep Illumina (Illumina, San Diego, CA, United States) and sequencing was performed. For each isolate, paired-end genomic libraries were prepared using DNA Prep Illumina. Sequencing was performed on Illumina MiSeq platform with 500 cycle chemistry (2 x 250 pair-end reads). The raw data were trimmed using Trimmomatic (Galaxy Version 0.36.6) [43] and then the draft genomes were generated by SPAdes 3.12.0 [44] de novo assembler, with default parameters. Draft genomes were submitted to BTyper 2.3.2 [45] as previously described in Bianco and colleagues [30]. The many functions of the tool include the definition of species using ANI blasts and the prediction of the major virulence genes of the studied species. Additionally, with the aim to identify the antibiotic resistance genes, the draft genomes were also analyzed using the software ABRicate (Galaxy Version 0.8), which includes different predownloaded databases: ARG-ANNOT [46]; NCBI AMRFinderPlus [47]; CARD [48]; ResFinder [49]; PlasmidFinder [50]. The draft genomes of B. cereus identified have been deposited in GenBank as BioProject PRJNA826696.
4.4. Antimicrobial Susceptibility Tests
Microdilution method was used to verify the behavior of each isolate towards the main antibiotic classes (β – lactam, aminoglycosides, tetracycline, fluoroquinolones, rifamycin, chloramphenicol, macrolides, lincosamides, glycopeptides, carbapenim and oxazolidinones), according to Clinical and Laboratory Standard Institute guidelines [51,52]. Antibiotics and related concentrations used for microdilution experiments were: penicillin G (0.031-4 µg/mL), gentamicin (0.125-16µg/mL), doxycycline (0.25-32 µg/mL), ciprofloxacin (0.25– 32 µg/mL), rifampin (0.25– 32 µg/mL), chloramphenicol (1-128 µg/mL), erythromycin (0.125-16 µg/mL), tetracycline (0.125-16 µg/mL), clindamycin (0.125-16 µg/mL), vancomycin (0.125-16 µg/mL), meropenem (0.125 - 16µg/mL) and linezolid (0.125 - 16µg/mL). Antibiotic powders were resuspended in appropriate solvent and then 100 µl of solution was pipetting into 96 well plates. Scalar dilutions of each antibiotic were set up, in a range of values included in the reference breakpoints.
After overnight incubation on Columbia Agar with 5% sheep blood, 1-2 bacterial colonies were suspended in sterile saline solution at 0.5 McFarland standard. The bacterial suspensions were further diluted 1:100 in CAMHB (cationic adjusted Muller-Hinton broth) and then inoculated into 96 well plates containing specific antibiotic. The bacterial growth was detected after 16 h of incubation at 37°C. Minimum Inhibiting Concentration (MIC) was detected for each antibiotic. It corresponds to the lowest antibiotic concentration able to inhibit bacterial growth. The obtained MIC values were interpreted using CLSI breakpoints [51,52].
The CLSI breakpoints (mg/mL) for penicillin G, meropenem, vancomycin, gentamicin, erythromycin, tetracycline, ciprofloxacin, clindamycin, chloramphenicol, and rifampin were those suggested for Bacillus spp. (not Bacillus anthracis), according to CLSI document M45 [52] whereas for linezolid and doxycycline interpretative criteria for Staphylococcus spp. were used according to CLSI M100 [51]. Staphylococcus aureus ATCC 29213 and E. coli ATCC 25922 were used as control strains, to check the experimental validity.
5. Conclusions
B. cereus infection is much more frequent than expected, and new data suggest its involvement in food contamination, even if the incidence of these foodborne diseases is also likely to be underreported because associated illness is usually self-limiting and not severe and hence, they usually remain undiagnosed [53,54]. Bacillus cereus group bacteria can be detected in food, prevalently in milk and its derivative, but also in other products (like vegetable, meat, and fish), causing food poisoning. Food contamination could happen at different stage of processing and is not possible to completely control this phenomenon. However, the management of the infections caused by these bacteria should involve also the analysis of antibiotic susceptibility of strains and their genetic makeup. Usually, in clinical practice, the most used antibiotics are meropenem, ciprofloxacin, vancomycin, erythromycin, clindamycin and linezolid. Monitoring the susceptibility of the circulating strains against these and/or other antibiotics, through minimal inhibitory concentration (MIC), could lead to an improvement of medical treatment strategies, in the selection of the most appropriate antibiotics, in order to avoid antibiotic therapy failure and the possible development of the antimicrobial resistance phenomenon. Moreover, molecular methods like WGS analysis, should be simultaneously performed to detect strains carrying “silent antimicrobial resistance genes”. These strains represent a very huge problem for public health because they represent a reservoir of ARGs that could be transmitted via HGT among species and genera and they can become active in cell host, determining a new resistance phenotype [38,39,40].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: ID strains, species of B. cereus group and source of isolation; Table S2: Presence (●) or absence of AMR genes detected in Bacillus cereus group stains analyzed in this study; Table S3: MIC value and their interpretation of the Bacillus cereus group strains tested in this study.
Author Contributions
Conceptualization, D.F., V.M., D.G.; methodology, D.F., V.M., A.B., S.C., M.C., R.F., L.S., V.R., L.P., V.V., M.L:D., D.C., M.I.; software, A.B. and S.C.; validation, A.B., S.C., D.F., V.M. and L.S.; data curation, S.C., L.P. and D.G.; writing—original draft preparation, D.F., V.M.; writing—review and editing, D.G.; supervision, M.C., D.G., A.P., A.F.; project administration, A.P.; funding acquisition, A.P., A.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Health, Italy (Rome) (Project Code: IZS PB 04/16 RC).
Data Availability Statement
The data presented in this study are openly available in GenBank (BioProject PRJNA826696). [GenBank] [https://www.ncbi.nlm.nih.gov/bioproject/?term=PRJNA826696] [BioProject PRJNA826696]
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Drobniewski, F.A. Bacillus cereus and related species. Clin Microbiol Rev. 1993 Oct;6:324-38. [CrossRef]
- Ehling-Schulz, M.; Koehler, T.M.; Lereclus, D. The Bacillus cereus Group: Bacillus species with Pathogenic Potential. Microbiol Spectr 2019, 7. [CrossRef]
- El-Arabi, T.F.; Griffiths, M.W. Bacillus cereus. In: Morris JG, Potter ME. Foodborne infections and intoxications, 4th ed. Academic Press – Elsevier, 2013, pp: 401-407.
- Tirloni, E.; Stella, S.; Celandroni, F.; Mazzantini, D.; Bernardi, C.; Ghelardi, E. Bacillus Cereus in Dairy Products and Production Plants. Foods 2022, 11 (17), 2572. [CrossRef]
- Abee, T.; Groot, M. N.; Tempelaars, M.; Zwietering, M.; Moezelaar, R.; Voort, M. van der. Germination and Outgrowth of Spores of Bacillus Cereus Group Members: Diversity and Role of Germinant Receptors. Food Microbiology 2011, 28 (2), 199–208. [CrossRef]
- Logan, N.A. Bacillus and relatives in foodborne illness. Journal of Applied Microbiology, 2012, 112: 417-429. [CrossRef]
- Majed, R.; Faille, C.; Kallassy, M.; Gohar, M. Bacillus cereus Biofilms — Same, Only Different. Front. Microbiol. 2016, 7, 1054. [CrossRef]
- Ceuppens, S., Boon, N., Uyttendaele, M. Diversity of Bacillus cereus group strains is reflected in their broad range of pathogenicity and diverse ecological lifestyles. FEMS microbiology ecology, 2013, 84, 433–450. [CrossRef]
- Ikeda, M.; Yagihara, Y.; Tatsuno, K.; Okazaki, M.; Okugawa, S.; Moriya, K. Clinical characteristics and antimicrobial susceptibility of Bacillus cereus blood stream infections. Ann. Clin. Microbiol. Antimicrob., 2015, 14:43. [CrossRef]
- Rasko, D.A.; Altherr, M.R.; Han, S.C.; Ravel, J. Genomics of the Bacillus cereus group of organisms. FEMS Microbiology Reviews. 2005, 29 303–329. [CrossRef]
- Senesi, S; Ghelardi, E. Production, Secretion and Biological Activity of Bacillus cereus Enterotoxins. Toxins 2010,2, 1690-1703; [CrossRef]
- Bottone, E.J. Bacillus cereus, a Volatile Human Pathogen. Clinical Microbiology Reviews, 2010, 382–398 Vol. 23, No. 2 0893-8512/10/$12.00. [CrossRef]
- Dietrich, R.; Jessberger, N.; Ehling-Schulz, M.; Märtlbauer, E.; Granum, P.E. The Food Poisoning Toxins of Bacillus cereus. Toxins 2021, 13, 98. [CrossRef]
- Celandroni, F.; Salvetti, S.; Senesi, S.; Ghelardi, E. Bacillus Thuringiensis Membrane-Damaging Toxins Acting on Mammalian Cells. FEMS Microbiol Lett 2014, 361 (2), 95–103. [CrossRef]
- Guérin, A.; Rønning, H. T.; Dargaignaratz, C.; Clavel, T.; Broussolle, V.; Mahillon, J.; Granum, P. E.; Nguyen-The, C. Cereulide Production by Bacillus Weihenstephanensis Strains during Growth at Different PH Values and Temperatures. Food Microbiology 2017, 65, 130–135. [CrossRef]
- Miller, R.A.; Beno, S.M.; Kent, D.J.; Carroll, L.M.; Martin, N.H.; Boor, K.J.; Kovac, J. Bacillus Wiedmannii Sp. Nov., a Psychrotolerant and Cytotoxic Bacillus Cereus Group Species Isolated from Dairy Foods and Dairy Environments. Int J Syst Evol Microbiol 2016, 66 (11),4744–4753. [CrossRef]
- Yuan, J.; Li, Y.-Y.; Xu, Y.; Sun, B.-J.; Shao, J.; Zhang, D.; Li, K.; Fan, D.-D.; Xue, Z.-B.; Chen, W.-H.; Pak, C.; Lou, Y.-L.; Su, J.-Z.; Zheng, M.-Q. Molecular Signatures Related to the Virulence of Bacillus Cereus Sensu Lato, a Leading Cause of Devastating Endophthalmitis. mSystems 2019, 4 (6), e00745-19. [CrossRef]
- EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards). Scientific opinion on the risks for public health related to the presence of Bacillus cereus and other Bacillus spp. including Bacillus thuringiensis in foodstuffs. EFSA Journal 2016; 14:4524, 93 pp. [CrossRef]
- Shawish, R.; Tarabees, R. Prevalence and antimicrobial resistance of Bacillus cereus isolated from beef products in Egypt. Open Vet J. 2017;7:337-341. [CrossRef]
- Granum, P.E.; Lund, T. Bacillus cereus and its food poisoning toxins, FEMS Microbiology Letters. 1997, Volume 157, Issue 2, Pages 223–228. [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: part 1: causes and threats. P & T: a peer-reviewed journal for formulary management 2015, 40(4), 277–283.
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare (Basel). 2023;11(13):1946. [CrossRef]
- Turnbull, P.C.; Sirianni, N.M.; LeBron, C.I.; Samaan, M.N.; Sutton, F.N.; Reyes, A.E.; Peruski, L.F.Jr. MICs of selected antibiotics for Bacillus anthracis, Bacillus cereus, Bacillus thuringiensis, and Bacillus mycoides from a range of clinical and environmental sources as determined by the Etest. J Clin Microbiol. 2004 Aug;42(8):3626-34. [CrossRef]
- Ghelardi, E.; Celandroni, F.; Salvetti, S.; Fiscarelli, E.; Senesi, S. Bacillus thuringiensis pulmonary infection: critical role for bacterial membrane-damaging toxins and host neutrophils. Microbes Infect. 2007;9(5):591-8. [CrossRef]
- Roca, A.; Cabeo, M.; Enguidanos, C.; Martínez-Checa, F.; Sampedro, I.; Llamas, I. Potential of the quorum-quenching and plant-growth promoting halotolerant Bacillus toyonensis AA1EC1 as biocontrol agent. Microb Biotechnol. 2024;17(3):e14420. [CrossRef]
- Andriūnaitė, E.; Tamošiūnė. I; Aleksandravičiūtė, M.; Gelvonauskienė, D.; Vinskienė, J.; Rugienius, R.; Baniulis, D. Stimulation of Nicotiana tabacum L. In Vitro Shoot Growth by Endophytic Bacillus cereus Group Bacteria. Microorganisms. 2021;9(9):1893. [CrossRef]
- Hollensteiner, J.; Wemheuer, F.; Harting, R.; Kolarzyk, A.M.; Diaz, Valerio S.M.; Poehlein, A.; Brzuszkiewicz, E.B.; Nesemann, K.; Braus-Stromeyer, S.A.; Braus, G.H.; Daniel, R.; Liesegang, H. Bacillus thuringiensis and Bacillus weihenstephanensis Inhibit the Growth of Phytopathogenic Verticillium Species. Front Microbiol. 2017 Jan 18;7:2171. [CrossRef]
- Hinnekens, P.; Fayad, N.; Gillis, A.; Mahillon, J. Conjugation across Bacillus cereus and kin: A review. Front Microbiol. 2022 Nov 4;13:1034440. [CrossRef]
- Jian, Z.; Zeng, L.; Xu, T.; Sun, S.; Yan, S.; Yang, L.; Huang, Y.; Jia, J.; Dou, T. Antibiotic Resistance Genes in Bacteria: Occurrence, Spread, and Control. J Basic Microbiol 2021, 61 (12), 1049–1070. [CrossRef]
- Bianco, A.; Capozzi, L.; Monno, M.R.; Del Sambro, L.; Manzulli, V.; Pesole, G.; Loconsole, D.; Parisi, A. Characterization of Bacillus cereus Group Isolates from Human Bacteremia by Whole-Genome Sequencing. Front Microbiol. 2021 Jan 12;11:599524. [CrossRef]
- Zhai, Z.; Cui, C.; Li ,X.; Yan, J.; Sun, E.; Wang, C.; Guo, H.; Hao, Y. Prevalence, antimicrobial susceptibility, and antibiotic resistance gene transfer of Bacillus strains isolated from pasteurized milk. J Dairy Sci. 2023;106(1):75-83. [CrossRef]
- Bartoszewicz, M. and Czyżewska, U. Comparison of the antibiotic resistance between genetically diverse and toxigenic Bacillus cereus sensu lato from milk, pepper and natural habitats. J Appl Microbiol. 2021;130(2):370-381. [CrossRef]
- Mills, E.; Sullivan, E.; Kovac, J. Comparative Analysis of Bacillus cereus Group Isolates' Resistance Using Disk Diffusion and Broth Microdilution and the Correlation between Antimicrobial Resistance Phenotypes and Genotypes. Appl Environ Microbiol. 2022;88(6):e0230221. [CrossRef]
- Fiedler, G.; Schneider, C.; Igbinosa, E. O.; Kabisch, J.; Brinks, E.; Becker, B.; Stoll, D. A.; Cho, G.-S.; Huch, M.; Franz, C. M. A. P. Antibiotics Resistance and Toxin Profiles of Bacillus Cereus-Group Isolates from Fresh Vegetables from German Retail Markets. BMC Microbiol. 2019;19(1):250. [CrossRef]
- Yim, J.-H.; Kim, K.-Y.; Chon, J.-W.; Kim, D.-H.; Kim, H.-S.; Choi, D.-S.; Choi, I.-S.; Seo, K.-H. Incidence, Antibiotic Susceptibility, and Toxin Profiles of Bacillus Cereus Sensu Lato Isolated from Korean Fermented Soybean Products. Journal of food science 2015, 80. [CrossRef]
- Nicoletti, G.; Russo, G.; Bonfiglio, G. Recent Developments in Carbapenems. Expert Opinion on Investigational Drugs 2002, 11 (4), 529–544. [CrossRef]
- Materon, I.C.; Queenan, A.M.; Koehler, T.M.; Bush, K.; Palzkill, T. Biochemical characterization of beta-lactamases Bla1 and Bla2 from Bacillus anthracis. Antimicrob Agents Chemother. 2003;47(6):2040-2. [CrossRef]
- Wagner, T.M.; Howden, B.P.; Sundsfjord, A.; Hegstad, K. Transiently silent acquired antimicrobial resistance: an emerging challenge in susceptibility testing. J Antimicrob Chemother. 2023;78(3):586-598. [CrossRef]
- Deekshit, V.K.; Srikumar, S. 'To be, or not to be'-The dilemma of 'silent' antimicrobial resistance genes in bacteria. J Appl Microbiol. 2022;133(5):2902-2914. [CrossRef]
- Stasiak, M.; Maćkiw, E.; Kowalska, J.; Kucharek, K.; Postupolski, J. Silent Genes: Antimicrobial Resistance and Antibiotic Production. Pol J Microbiol. 2021;70(4):421-429. [CrossRef]
- Bravo, A.; Moreno-Blanco, A.; Espinosa, M. One Earth: The Equilibrium between the Human and the Bacterial Worlds. Int J Mol Sci. 2023;24(20):15047. [CrossRef]
- Manzulli, V.; Rondinone, V.; Buchicchio, A.; Serrecchia, L.; Cipolletta, D.; Fasanella, A.; Parisi, A.; Difato, L.; Iatarola, M.; Aceti, A.; Poppa, E.; Tolve, F.; Pace, L.; Petruzzi, F.; Rovere, I.D.; Raele, D.A.; Del Sambro, L.; Giangrossi, L.; Galante, D. Discrimination of Bacillus cereus Group Members by MALDI-TOF Mass Spectrometry. Microorganisms. 2021 Jun 2;9(6):1202. [CrossRef]
- Bolger, A.M., Lohse, M., Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 2014,30, 2114–2120. [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012 ;19(5):455-77. [CrossRef]
- Carroll, L.M.; Kovac, J.; Miller, R.A.; Wiedmann, M. Rapid, High-throughput identification of anthrax-causing and emetic Bacillus cereus group genome assemblies via BTyper, a computational tool for virulence-based classification of Bacillus cereus group isolates by using nucleotide sequencing data. Appl. Environ. Microbiol. 2017, 83:e1096-17. [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; et al. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [CrossRef]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I., et al. Validating the AMRFinder tool and resistance gene database by using antimicrobial resistance genotype-phenotype correlations in a collection of isolates. Antimicrob. Agents Chemother. 2019,63:e483-19. [CrossRef]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K., et al. CARD 2017: expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2017,45,D566–D573. [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; et al. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; et al. In silico detection and typing of plasmids using plasmidfinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria. CLSI Guideline M45. 2016, 3rd ed. Wayne: CLSI.
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. CLSI Supplement M100. 2017, 27th ed. Wayne: CLSI.
- Glasset, B.; Herbin, S.; Guillier, L.; Cadel-Six, S.; Vignaud, M.L.; Grout, J.; Pairaud, S.; Michel, V.; Hennekinne, J.A.; Ramarao, N.; Brisabois, A. Bacillus cereus-induced food-borne outbreaks in France; 2007 to 2014: epidemiology and genetic characterisation. Euro Surveill. 2016;21(48):30413. [CrossRef]
- Kotiranta, A.; Lounatmaa, K.; Haapasalo, M. Epidemiology and pathogenesis of Bacillus cereus infections. Microbes Infect. 2000;2(2):189-98. [CrossRef]
Figure 1.
The histogram shows the percentages of presence (in red) or absence (in green) of AMR genes detected in Bacillus cereus group stains analyzed in this study.
Figure 1.
The histogram shows the percentages of presence (in red) or absence (in green) of AMR genes detected in Bacillus cereus group stains analyzed in this study.
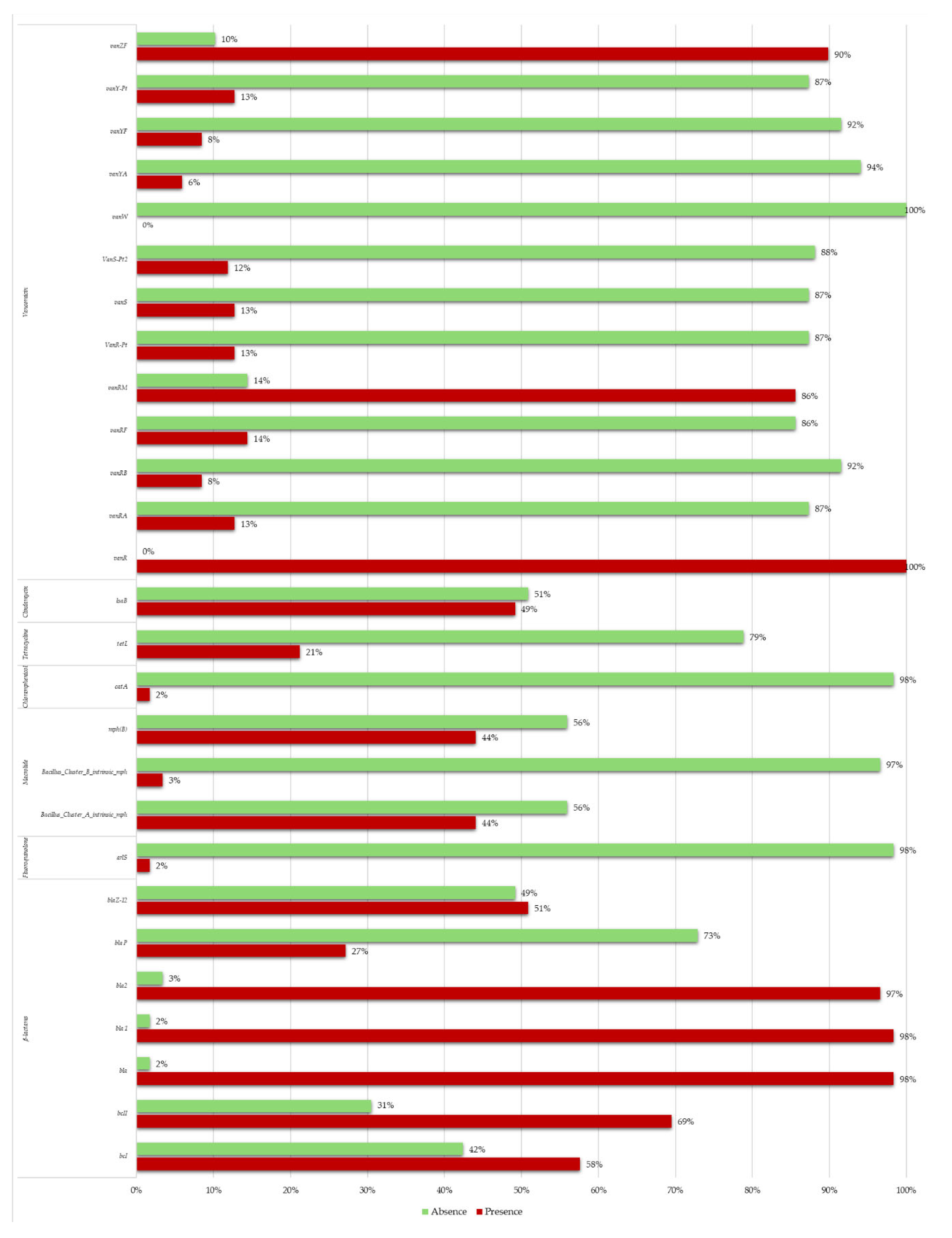
Figure 2.
In this graphic are showed the percentages of sensitivity (in green), resistance (in red) and intermediate behavior (in yellow) of the Bacillus cereus group strains tested in this study against 12 clinically relevant antibiotics.
Figure 2.
In this graphic are showed the percentages of sensitivity (in green), resistance (in red) and intermediate behavior (in yellow) of the Bacillus cereus group strains tested in this study against 12 clinically relevant antibiotics.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



