
Submitted:
01 July 2024
Posted:
02 July 2024
You are already at the latest version
Abstract
Purpose of review: This review investigates the oral microbiome's composition, functions, influencing factors, connections to oral and systemic diseases, and personalised oral care strategies. Recent findings: The oral microbiome is a complex ecosystem consisting of bacteria, fungi, archaea, and viruses that contribute to oral health. Various factors, such as diet, smoking, alcohol consumption, lifestyle choices, and medical conditions, can affect the balance of the oral microbiome and lead to dysbiosis, which can result in oral health issues like dental caries, gingivitis, periodontitis, oral candidiasis, and halitosis. Importantly, our review explores novel associations between the oral microbiome and systemic diseases including gastrointestinal, cardiovascular, endocrinal, neurological conditions, autoimmune diseases, and cancer. We comprehensively review the efficacy of interventions like dental probiotics, xylitol, oral rinses, fluoride, essential oils, oil pulling, and peptides in promoting oral health by modulating the oral microbiome. Summary: This review emphasizes the critical functions of the oral microbiota in dental and overall health, providing insights into the effects of microbial imbalances on various diseases. It underlines the significant connection between the oral microbiota and general health. Furthermore, it explores the advantages of probiotics and other dental care ingredients in promoting oral health and addressing common oral issues, offering a comprehensive strategy for personalized oral care.
Keywords:
Oral Microbiome
; Oral Diseases
; Systemic Diseases
; Dysbiosis
; Probiotics
; Oral Microbiota
; Oral care
Introduction
The human oral cavity harbors a diverse and complex microbial community, collectively known as the oral microbiome [1]. It has diverse microorganisms, including bacteria, archaea, fungi, and viruses [2]. It exists in the form of a biofilm and contributes significantly to maintaining oral homeostasis by fostering a harmonious balance within the oral environment [1]. Through its natural defence mechanisms, the oral microbiome acts as a protective barrier, preventing the colonization of harmful pathogens and preventing potential infections in the oral cavity [3]. Furthermore, a balanced oral microbiome is instrumental in preventing the development of oral diseases, such as dental caries and periodontal diseases, as it helps control the growth and activity of pathogenic bacteria [1,3]. Understanding the diverse roles of the oral microbiome is essential for devising effective preventive and therapeutic strategies to promote oral health.
Interestingly, various civilizations have shown an extensive understanding of the vitality of oral health in the past. Remarkably, in ancient Egypt, the arid climate and special burial customs led to the preservation of well-kept skeletons and mummies. Researchers have learned a great deal about the historical dental practices of ancient civilizations by examining these remnants [4]. In antiquity, societies likely employed wooden toothpicks, chew sticks, tree twigs, and animal bones as implements for oral hygiene. Impressively, traces of caries and periodontal disease date back thousands of years to the upper Palaeolithic era. As opposed to their prevalent occurrence in current times, dental caries was notably infrequent in ancient Egypt due to a lack of fermentable carbohydrates in their diet, impeding plaque buildup. Additional factors, such as tooth wear, including occlusal and interproximal wear, further contributed to the scarcity of caries by smoothing surfaces and discouraging plaque proliferation [4]. This historical context enriches our understanding of the intricate interplay between oral health practices and their enduring impact on overall health.
Under healthy conditions, the oral microbiome thrives in a favourable commensal association with its environment, much like other body regions, including the skin, gut, or vagina [5]. However, it is important to acknowledge that under certain circumstances, certain opportunistic microorganisms within the oral microbiome can undergo a shift, becoming harmful pathogens. This transformation can have implications for the development of various oral and systemic diseases [5,6]. Major oral diseases such as dental caries and periodontal diseases are both caused by and contribute to disruptions in the oral microbiota balance, known as dysbiosis. This dysbiosis can extend its effects to various chronic systemic diseases, leading to the initiation or worsening of conditions such as metabolic diseases, CVD, respiratory diseases, rheumatoid arthritis (RA), adverse pregnancy outcomes, IBD, AD, autism spectrum disorders, and oral mucosal diseases [7].
In the past decade, significant progress has been made in understanding the impact of the oral microbiome on health and disease, propelled by innovative genomic technologies such as NGS and advanced bioinformatic tools. NGS is an umbrella term that encompasses various methods like 16S rRNA sequencing, metagenomics, shotgun metagenomics, and RT-PCR, all of which have replaced conventional culture-based techniques in unravelling the complexities of the oral microbiome [8,9]. In this context, stimulated saliva collection offers a controlled and comprehensive method for investigating the oral microbiome, providing valuable insights into its composition, functions, and implications for human health. (Detailed techniques for analysis of the oral microbiome is provided in Supplementary Material 1). It is important to acknowledge that the oral microbiome is moulded by various factors, such as diet, oral hygiene practices, smoking, systemic illnesses, and changes in the oral environment [10]. Therefore, emphazising the importance of promoting good oral hygiene practices, adopting a healthy diet, and making positive lifestyle changes can play a pivotal role in nurturing a balanced oral microbiome ecosystem.
This review extensively examines the diverse microbial composition and its role in maintaining oral equilibrium, outlining the significance of various bacterial genera. Additionally, it delves into the oral-gut axis and microbial translocation, unravelling the interconnection between mouth and gut. We also explore how dysbiosis can instigate a cascade of consequences, not only leading to various oral diseases such as dental caries, periodontitis, halitosis, and oral candidiasis but also exerting a profound impact on systemic health, encompassing conditions like gastrointestinal diseases, CVD, diabetes, neurological diseases, and autoimmune diseases. Lastly, understanding the pivotal role of daily dental care practices is essential for maintaining oral hygiene and preventing oral diseases, thus contributing to the preservation of a balanced oral microbiome ecosystem and good systemic health.
The Composition and Role of the Oral Microbiome
With over 700 known bacterial species, the oral microbiome is the second-most diverse microbiota within the human body [1]. Principally, this vibrant ecosystem encompasses a wide range of bacterial phyla, including Actinobacteria, Bacteroidetes, Chlamydia, Euryarchaeota, Fusobacteria, Firmicutes, Proteobacteria, Spirochaetes, and Tenericutes [11]. In a healthy oral cavity, the prevailing bacterial genera primarily encompass both Gram-positive and Gram-negative species. Within the realm of Gram-positive cocci, the genera Abiotrophia, Peptostreptococcus, Streptococcus, and Stomatococcus assume pivotal roles. Notably, some bacterial species, including Streptococcus sanguinis and salivarius, are known for producing antimicrobial compounds, making a significant contribution to the maintenance of oral health, and protecting dental structures from the damaging effects of enamel erosion. [12]. However, it is important to note that the full extent of each bacterium’s role in this complex interaction has not yet been fully elucidated.
Gram-positive rods, such as Actinomyces, Bifidobacterium, Corynebacterium, Eubacterium, Lactobacillus, Propionibacterium, Pseudoramibacter, and Rothia, contribute to the microbial consortium. Considering recent research, Actinomyces naeslundii has been discerned as proficient in utilizing urea as a nitrogen source, thereby strengthening its resistance against environmental acidification within the oral cavity. Consequently, it assumes a dominant role among dental plaque organisms, exerting a pronounced influence on the ecological processes of the plaque community. [13]. Furthermore, Lactobacillus, Bifidobacterium, Streptococcus, and Weissella are all acknowledged as helpful probiotics in treating periodontal diseases, boosting oral health, and improving oral care as a whole [14].
In addition to the Gram-positive taxa, the Gram-negative cocci genera Moraxella, Neisseria, and Veillonella are prevalent inhabitants of the healthy oral cavity. Among the Gram-negative rods, the following genera are commonly observed: Campylobacter, Capnocytophaga, Desulfobacter, Desulfovibrio, Eikenella, Fusobacterium, Hemophilus, Leptotrichia, Prevotella, Selemonas, Simonsiella, Treponema, and Wolinella [15]. The findings strongly support the idea that Fusobacterium nucleatum plays a crucial role in facilitating intergeneric coaggregation. This involves its ability to bring diverse bacteria together, enabling them to create a well-organized community known as a biofilm. Furthermore, the involvement of F. nucleatum extends to the subsequent growth and development of these biofilms [16]. Within these biofilms, Prevotella organisms have a unique role. They are capable of regulating host residence and immune cell responses without requiring direct contact with the host. While Prevotella spp. are generally considered to be aerotolerant, their ability to thrive in aerated environments appears to be significantly influenced by their coaggregation with F. nucleatum. This coaggregation not only contributes to the structural maintenance of oral biofilms but also allows Prevotella spp., to benefit from the multispecies character of these biofilms [17]. The precise functions carried out by each bacterial genus offer a field for exploration and deeper study.
Fungi are estimated to constitute less than 0.1% of the total microbial population in the oral cavity [18]. The oral cavity is frequently associated with Candida spp., making it the most prevalent fungus observed. Additionally, other notable fungi identified in this environment include species belonging to Cladosporium, Aureobasidium, Saccharomycetales, Aspergillus, Fusarium, and Cryptococcus groups [10]. When the host’s defence mechanism is weakened, fungi in the oral microbiome may still modulate immune responses or even develop into opportunistic infections [19]. Physical interactions between fungi and bacteria in the oral cavity influence the oral microbiota. Candida and other oral fungi act as bridging organisms, facilitating bacterial adhesion to their surfaces and epithelial cells. This leads to distinct sets of bacteria adhering to Candida-covered vs. non-covered cells. Additionally, these interactions contribute to bacterial resilience against antimicrobial agents, modulating the diversity of the oral microbiome [20]. Oral archaea exhibit significantly lower abundance and diversity compared to oral bacteria. In the oral cavity, all known archaeal species belong to the phylum Euryarchaeota and are characterized as methanogens, organisms that produce methane [2]. Studies have indicated that these archaea are found in higher quantities in individuals affected by periodontitis [21,22]. The composition of the oral microbiome is summarized in Table 1, showcasing the predominant phyla/genera of bacteria, fungi, and archaea, along with other notable genera.
In addition to the three primary domains of life—bacteria, fungi, and archaea—the oral microbiome also encompasses viruses. Despite viruses being the dominant infectious agents, we have limited research on the complex viral communities in the human mouth, even though some now believe they could be commensals [23,24,25]. However, it is critical to emphasize that while this hypothesis is gaining attention, substantial empirical evidence or explicit validation within the scientific literature is currently lacking to substantiate this assertion.
Factors Influencing Oral Microbiome
Diet
Emerging research brings out the profound influence of dietary macronutrients and diet types on the composition and equilibrium of the oral microbiome. The presence of nutrients plays a crucial role, as seen in the notable impact of fermentable carbohydrates like sugars and starches, which strongly influence the proliferation of acid-producing bacteria, particularly Streptococcus mutans. These microbes drive the development of dental caries-generating acids that erode tooth enamel and initiate cavities [23]. Remarkably, the oral microbiome not only responds to dietary components, indicating its ability to change in response to the types of food consumed but also actively participates in nutrient breakdown. This process involves specialised bacteria that degrade glycoproteins and sugars, a crucial step in nutrient utilisation [24].
However, the impact of diet extends beyond nutrient breakdown. The influence of food choices on oral microbiome functions is shown by the fact that a nitrate-rich diet can promote the proliferation of nitrate-reducing bacteria. Dietary nitrate changes the salivary microbiome in people of various ages, with Prevotella and Veillonella populations declining and Rothia and Neisseria populations rising [25]. Notably, this modification in the salivary microbiota can affect the control of blood pressure and nitric oxide levels [26]. Furthermore, the complex relationship intertwines with immune responses and bacterial interactions. Antioxidants and anti-inflammatory compounds found in fruits and vegetables can modulate the immune response in the oral cavity, influencing the balance of various bacterial species [27].
Smoking
The influence of smoking on the oral microbiome is a multifaceted phenomenon with far-reaching implications for oral health. The relationship between smoking and the oral microbiome involves mechanisms such as the creation of anaerobic conditions, compromised immune responses, altered salivary pH, and the antibacterial effects of cigarette smoke’s toxic compounds [28,29]. Nicotine, a substance found in tobacco leaves, has been shown to boost the virulence of oral microorganisms, possibly by stimulating the expression of virulence-related genes or promoting increased biofilm formation. Its influence on human cells is primarily through nicotine acetylcholine receptors (nAChRs) [30]. These alterations can disrupt the equilibrium within indigenous oral microbial communities, including beneficial species, while also creating a more favourable environment for the colonisation of harmful microbes associated with oral diseases. Consequently, individuals who smoke may be at an increased risk of infections and oral health complications.
Tobacco smoke has a dual impact on the oral microbiome. First, it decreases oxygen levels, creating an environment ideal for anaerobic bacteria that thrive without oxygen. Simultaneously, it increases free iron levels while inhibiting oral peroxidase activity, further promoting conditions favourable for anaerobic bacterial growth. As a result of this, exposure to tobacco smoke prompts salivary cells to switch to anaerobic glycolysis for energy production [31]. The alteration in the oral microbiota due to smoking is marked by a decrease in Proteobacteria and an increase in Firmicutes and Actinobacteria. These changes collectively contribute to the significant shifts observed in the composition of the oral microbiome in response to smoking [28]. Comprehensive studies utilizing advanced techniques are needed to illuminate the full scope of smoking’s impact, particularly on passive smokers, shedding light on potential interventions to counteract these effects and enhance oral health outcomes [10].
Alcohol Consumption
Excessive alcohol consumption can lead to an imbalance within the oral microbiome [32], causing potential repercussions for both oral and systemic health. Prolonged alcohol intake has been shown to impact various aspects of the oral environment, including the oral mucosa, salivary glands, and saliva composition [33]. Exposure to ethanol, whether acute or chronic, can result in significant changes in salivary function, characterized by decreased flow rate and impaired secretion of crucial components like total protein and amylase [34,35]. Moreover, alcohol can impair the function of neutrophils, potentially leading to bacterial overgrowth and increased bacterial infiltration [36]. It can also reduce the production of inflammatory cytokines by monocytes, creating an environment conducive to microbial proliferation [37]. Additionally, ethanol’s adverse effects extend to dental health by promoting bone resorption and suppressing bone formation [38], impacting teeth, and supporting periodontal structures [39].
Notably, research has underscored a correlation between alcohol use and alterations in the oral microbiota, characterized by diminished Lactobacillales abundance and increased presence of Neisseria, Streptococcus, and Prevotella [40,41]. Interestingly, the oral microbiota has exhibited potential as an indicator of alcohol-induced liver damage [42]. Furthermore, diurnal changes within the oral microbiota, crucial for maintaining microbial balance, are reportedly disrupted by alcohol consumption, potentially contributing to disturbances within the microbial ecosystem and subsequent functional impairments [42]. Intriguingly, studies by Thomas et al. and Liao et al. have revealed distinct variations in the oral microbiota of alcohol consumers, including reduced bacterial richness and shifts in the prevalence of specific genera such as Prevotella and Moryella. Conversely, genera like Lautropia, Haemophilus, and Porphyromonas were found to be significantly diminished [43,44].
Other Factors That Influence the Oral Microbiome
In addition to diet, smoking, and alcohol consumption, several other factors significantly influence the composition and equilibrium of the oral microbiome. Poor oral hygiene practices, medical conditions, and medications play pivotal roles in shaping this complex ecosystem. Poor oral hygiene, characterized by inadequate brushing and flossing, promotes the accumulation of dental plaque—a biofilm predominantly composed of bacteria. This plaque buildup provides a conducive environment for the proliferation of pathogenic species, leading to an imbalance in the oral microbiome [45]. Moreover, various medical conditions exert substantial effects on oral microbial communities. Systemic diseases such as diabetes, CVD, neurological conditions, gastrointestinal diseases, and autoimmune disorders can disrupt immune responses and alter oral pH, creating favourable conditions for the overgrowth of pathogenic bacteria [46]. Furthermore, medications, notably antibiotics and immunosuppressants, can disturb microbial homeostasis by directly impacting bacterial populations or weakening host defences [47]. Consequently, dysbiosis may arise, contributing to oral health issues. The use of broad-spectrum antibiotics can potentially disrupt the natural balance of beneficial oral microorganisms. In people without health issues, helpful microorganisms keep Candida in check by competing for nutrients and attaching to epithelial cells. Changes in these microorganisms might lead to the unchecked growth of Candida. Additionally, the broader effects of systemic antibiotic treatment could also contribute to oral candidiasis by affecting the immune system [48]. Moreover, the oral microbiome is a known site for the acquisition and transfer of resistance via horizontal gene transfer (HGT), which can contribute to the systematic development of antimicrobial-resistant infections [49]. HGT in the oral microbiome enables the rapid sharing of antibiotic resistance genes among microorganisms, leading to the emergence of antibiotic-resistant strains, reducing antibiotic efficacy, and contributing to the overall problem of antibiotic resistance.
Oral Microbiome Dysbiosis
Oral microbiome dysbiosis refers to an imbalance or disruption in the composition and functioning of the oral microbial community [1]. This disturbance can arise from various factors, such as poor oral hygiene, dietary habits, medical conditions, medications, and lifestyle choices. This disrupts the previously harmonic interactions between various bacteria, changing the overall microbial ecology. The consequences of oral microbiome dysbiosis are far-reaching. It can contribute to the development of common oral health problems, including dental caries (cavities), periodontal diseases (gum infections), halitosis (bad breath or oral malodor), and oral candidiasis (oral thrush). Furthermore, emerging research suggests potential links between oral microbiome dysbiosis and systemic health conditions, such as CVD, diabetes, and even cognitive disorders [7]. One of the key features of oral microbiome dysbiosis is the overgrowth of harmful bacteria and a decline in beneficial microorganisms [50]. This imbalance can create a hostile environment where pathogens thrive, leading to inflammation and damage to oral tissues. Additionally, dysbiosis can disrupt the natural processes that help maintain tissue health and repair, further exacerbating oral health problems.
Oral-Gut Axis
The oral and gut microbiomes play crucial roles in human health, each harbouring a distinct microbial community. These communities are typically separated by barriers such as the oral-gut barrier, physical distance, and chemical obstacles like gastric acid and bile [51]. However, recent research has shed light on the potential microbial translocation between these two ecosystems, highlighting the interconnected nature of the oral-gut microbiome axis. Several factors can influence the translocation of oral microbiota to the gut. Individuals at the extremes of life, such as neonates and the elderly, may have fewer effective barriers, allowing for increased microbial movement [52,53]. Certain bacteria, like Bifidobacterium, have been detected in the neonatal gut and oral fluids [54]. Additionally, decreased gastric acidity can shift the gut microbiota composition to resemble the oral microbiome. This suggests that under certain conditions, oral microbes can bypass barriers and successfully translocate to the gut [55]. Microbial translocation is not limited to physiological states. Pathological conditions, such as IBD and colon cancer, have shown evidence of oral bacteria colonising the gut mucosa [56]. This suggests that disruptions in mucosal homeostasis can lead to the invasion of oral commensals into the gut, potentially contributing to disease progression.
Two main hypotheses have been proposed for the transmission of oral bacteria to the gut: the hematogenous route and the enteral route (Figure 1). The hematogenous route suggests that oral bacteria enter the bloodstream, circulate, and colonise the gut mucosa. Meanwhile, the enteral route proposes that bacteria from the mouth travel through the stomach to the intestines. Although the human body possesses defences against microbial invasion, instances can arise where these barriers are compromised, allowing for bacterial movement [57]. The findings of oral-to-gut microbial translocation implicate human health. Antibiotic use and conditions like achlorhydria (absence of hydrochloric acid in the gastric secretions) can diminish barriers, facilitating the colonisation of oral bacteria in the gut [58,59]. Certain pathogens, like Klebsiella spp. and Porphyromonas gingivalis, possess antibiotic resistance and acid-resistant properties, respectively, enhancing their ability to translocate to the gut [58,60]. The occurrence of oral commensal bacteria in the gut is an uncommon and abnormal event, resulting from a process known as ectopic colonisation, where these bacteria are found in a location where they do not typically belong. This presence of oral commensals in the gut is considered a characteristic feature of various diseases, including IBD [61]. Several known oral bacterial species belonging to genera such as Staphylococcus, Porphyromonas, Veillonella, Fusobacterium, Actinomyces, and Parvimonas have been detected within the gut of patients with gastrointestinal disorders [59]. The emerging understanding of oral-gut microbial translocation deepens the complex interplay between these two microbial ecosystems.
Oral Microbiome and Oral Diseases
Dental Caries
Dental caries stands as the most prevalent infectious disease within the oral environment [62], primarily driven by bacterial pathogens and characterised by progressive dental hard tissue deterioration [63]. Recent investigations have unveiled the pivotal role of the oral microbiome in caries onset and progression. Prevotella spp., Lactobacillus spp., Dialister spp., and Filifactor spp. have surfaced as potential contributors to caries pathogenesis and advancement [64]. Caries development is attributed to the acidic by-products of sugar metabolism by acidogenic bacteria, inducing demineralization and cavitation [65]. All these factors, such as ecological imbalances, trends towards acidogenic environments, and the acidification of biofilms, contribute to the emergence of caries [66]. The caries biofilm, a complex and highly active ecosystem, initiates the accumulation of dental plaque on the tooth’s surface. Within this environment, various microorganisms colonise, with S. mitis and S. mutans being among the initial colonisers [67,68].
The formation of the biofilm begins when a salivary glycoprotein film (dental pellicle) coats a tooth surface [69]. These initial colonisers create extracellular polymers (EPS) that enhance the adherence of other microorganisms to the biofilm. EPS-rich environments provide ideal binding sites for acid-producing bacteria. Acid-producing bacterial species, such as Veillonella, Lactobacillus, and Propionibacterium, become part of the dental biofilm. They contribute to cariogenic conditions in the mouth by producing acids [70]. S. mutans has cariogenic potential, excels in acid production, allowing it to metabolise a wide range of carbohydrates into organic acids, and exhibits remarkable acid resistance, enabling it to thrive even in low pH environments. Additionally, it can synthesize EPS that not only promotes its growth but also offers protection to its cells in challenging and harsh conditions. These attributes collectively underscore the significant role of S. mutans in the pathogenesis of dental caries [71]. S. mutans utilises a crucial virulence factor, specifically three enzymes known as glucosyltransferases (GtfBCD), to initiate and establish cariogenic biofilms [72]. Bacterial diversity within caries-associated microbiota is modulated by competitiveness, as evidenced by varying compositions across different carious sites [72,73]. S. mutans [74], long considered a key player, has been joined by acidogenic species like Bifidobacterium [75] and Lactobacillus [76]. The multifaceted cariogenic consortia include Prevotella, Corynebacterium, Veillonella, and Capnocytophaga, collectively contributing to local pH reduction and tissue demineralization [75,77].
Gingivitis
Gingivitis is described as inflammation of the gums, typically caused by the accumulation of microbial plaque, or bacteria, on the surface of the teeth due to ineffective tooth brushing. Thus, the importance of effective tooth brushing cannot be overstated, as it is essential for removing food debris and preventing the further growth of plaque [78]. Bacteria present in accumulated plaque on the tooth surface can penetrate the gingival tissue, particularly the gingival sulcus, making the marginal area prone to microbial infection. Common microbial species associated with gingivitis include P. gingivalis , A. actinomycetemcomitans, Streptococcus sp., Fusobacterium sp., Actinomyces sp., Veillonella sp., Treponema denticola, and Prevotella intermedia [79]. If untreated, gingivitis has the potential to advance to periodontitis, a condition that can lead to irreversible damage not only to the gums but also to the surrounding bone that supports the teeth [78].
Periodontitis
The oral microbiome plays a pivotal role in the development and progression of periodontal diseases, encompassing gingival diseases and periodontitis [80]. Chronic periodontitis, a prevalent form of periodontal disease, spans a wide age range and evolves from gingivitis to deeply affecting periodontal tissues. Dental plaque bacteria emerge as the principal culprits in the onset of periodontal disease [81]. Plaque exists in both supragingival and subgingival areas, fostering microbial biofilms that thrive on tooth surfaces. The crevicular epithelium and gingival crevice act as pivotal habitats for microbial initiation, supported by gingival crevicular fluid and a favourable anaerobic environment [8]. The subgingival plaque, comprising more than 500 species, undergoes compositional shifts during disease progression [82]. In healthy conditions, Gram-positive cocci and rods dominate as early colonisers, while F. nucleatum bridges bacterial co-aggregation [82]. However, as gingivitis advances, Gram-negative bacteria like Prevotella spp. and F. nucleatum ss. polymorphum surge, associated with elevated inflammatory cytokines such as IL-1, IL-6, IL-17, TNF-α, and IL-23 [83,84,85].
IL-17 plays a stimulatory role in activating T helper (Th)-17 and B cells, which, when activated, elevate the expression of receptor activator of nuclear factor kappa-B ligand (RANKL). RANKL is also directly activated by the neutrophils that have migrated to the site. RANKL, in turn, triggers the activation and maturation of osteoclast precursors into active osteoclasts, which are responsible for bone resorption. Simultaneously, the recruited neutrophils contribute to tissue degradation by inducing the production of enzymes like matrix metalloproteinases (MMP) and cytotoxic substances, including reactive oxygen species (ROS). These interactions among microbes, the innate immune response, and the adaptive immune response illustrate key mechanisms involved in the persistence of inflammation, which, if left unresolved, leads to tissue destruction [86]. The red complex bacteria—P. gingivalis, Treponema denticola, and Tannerella forsythia [11]—emerge as keystone pathogens, triggering an imbalance in host-bacterial interactions and resulting in bone loss [87,88]. Furthermore, recent investigations highlight the systemic implications of periodontal diseases. Periodontitis not only poses local health risks but also acts as a potential risk factor for systemic conditions like CVD, diabetes, and cancer [9].
Halitosis
Halitosis, commonly referred to as oral malodor or bad breath, is a frequent oral condition marked by an unpleasing or disagreeable odour that originates within an individual’s mouth [89]. This widespread condition impacts a significant portion of the worldwide population, ranging from 15% to 60%, and is characterised by the emission of an offensive odour from the mouth, nasal passages, and throat regions [90]. Intra-oral halitosis (IOH), closely tied to microbial interactions within the oral environment, is a prominent facet of this condition. Anaerobic bacteria give rise to volatile compounds, with key volatile sulfur compounds (VSCs) including hydrogen sulfide, dimethyl sulfide, dimethyl disulfide, and methyl mercaptan [90,91]. Oral bacteria, such as Prevotella spp. [92]. Bacteroides spp. and Actinomyces spp. play vital roles in halitosis by producing VSCs during the biodegradation of sulfur-containing amino acids [91,92]. These sulfur-containing amino acids, such as L-cysteine and L-methionine, originate from saliva, blood, the oral mucosa, and epithelial cell debris. The enzymatic reactions involving L-cysteine and L-methionine are crucial in sulfur-containing amino acid metabolism. L-cysteine undergoes desulfhydration, catalysed by the L-cysteine desulfhydrase enzyme, resulting in the formation of pyruvate, ammonia, and hydrogen sulfide. On the other hand, L-methionine is acted upon by the L-methionine α-deamino-γ-mercaptomethane-lyase enzyme, leading to the generation of α-ketobutyrate, ammonia, and methyl mercaptan [93]. In addition, VSCs, particularly hydrogen sulfide and methyl mercaptan, exhibit significant toxicity. These observations imply that VSCs not only play a role in causing bad breath but could also be involved in the development of periodontitis [94].
Gram-negative bacteria, including Prevotella (Bacteroides) melaninogenica, Treponema denticola, Porphyromonas gingivalis, Porphyromonas endodontalis, and others like Bacteroides loescheii, Enterobacteriaceae, Tannerella forsythensis, Centipeda periodontii, Eikenella corrodens, and various Fusobacterium species, are implicated as likely contributors to oral malodor. However, the lack of a clear association between a specific bacterial infection and halitosis suggests that malodor is the result of intricate interactions among multiple oral bacterial species [95].
Earlier studies by Persson and collaborators highlighted the production of H2S by bacteria such as Treponema denticola and Bacteroides intermedius, alongside Peptostreptococcus, Eubacterium, and Fusobacterium [96]. Additionally, investigations by other researchers revealed the dominance of Veillonella spp., Actinomyces spp., and Prevotella spp. in patients experiencing oral malodor [97]. Conversely, Neisseria spp., Fusobacterium spp., and Porphyromonas spp. were noted to be most prevalent in individuals with elevated H2S concentrations, while Veillonella spp., Prevotella spp, Megasphaera, and Selenomonas predominated in those exhaling primarily CH3SH [98]. Notably, lower levels of Streptococcus and Granulicatella were evident, while higher proportions of Leptotrichia, Peptostreptococcus, Eubacterium, and Fusobacterium were detected across both (H2S and CH3SH) malodor groups [98].
Taste Impairment
Taste ability is notably compromised in acutely hospitalized elderly individuals who exhibit decayed teeth, elevated levels of oral bacteria linked to caries, poor oral hygiene, and dry mouth [99]. It has been proposed that the loss of taste associated with poor oral health may be attributed to toxins and inflammatory products produced by oral bacteria [100]. A study revealed that patients with high Lactobacilli growth experienced particularly impaired sour taste perception [99]. Lactobacilli thrive in acidic environments and produce acid themselves, potentially causing an adaptation in sour taste perception and increasing the taste threshold for sour flavours. Additionally, we found that inadequate oral hygiene correlated with a reduced total taste score and diminished perception of salty taste [101,102]. Saliva plays a crucial role in transporting food particles and taste stimuli to the taste buds in the oral cavity, and reduced saliva flow has been linked to taste impairment [103]. Hospitalized elderly individuals frequently consume a high number of medications, which can induce xerostomia (dry mouth) and hypo-salivation [104], resulting in loss of taste. Findings by Kamel et al. [105] and Weiffenbach et al. [106], reveal reduced taste sensitivity in individuals with Sjøgren’s syndrome, characterized by oral dryness, compared to controls.
Burning Mouth Syndrome
Burning Mouth Syndrome (BMS) is a condition characterized by a persistent burning sensation in the mouth without any apparent lesions [107]. Recent research suggests a potential association between BMS and the oral microbiota, shedding light on the role of oral bacteria in the etiology of this syndrome. A study investigating the oral microbiota signature of BMS patients found marked differences in microbial profiles between BMS patients and healthy controls [108]. The composition of bacterial strains, including dominant genera, was distinct in BMS patients compared to controls. Streptococcus, Rothia, Bergeyella, and Granulicatella were prevalent in the BMS group, while Prevotella, Haemophilus, Fusobacterium, Campylobacter, and Allorevotella were more common in healthy controls [108]. This correlation between oral microbiome composition and BMS suggests a potential role of oral bacteria in the pathogenesis of BMS.
Oral Thrush
Oral thrush, also known as oral candidiasis, is a fungal infection that affects the oral mucosa. It is caused by an overgrowth of Candida albicans [109]. Candida krusei, Candida stellatoidea, Candida tropicalis, Candida glabrata, Candida guilliermondii, and Candida dubliniensis are other species seen in oral candidiasis lesions [110]. The symptoms of oral thrush can include white patches on the tongue, inner cheeks, and roof of the mouth, as well as redness and soreness in the affected areas. In severe cases, it can cause difficulty swallowing and a burning sensation in the mouth [111]. Factors such as poor oral hygiene, a weakened immune system, or certain medical conditions like diabetes or HIV/AIDS can increase the risk of oral thrush [112]. These factors can create an imbalance in the natural flora of the mouth, allowing the Candida fungus to overgrow and cause an infection. Several processes contribute to Candida albicans pathogenicity (Figure 2). Initially, it adheres to host surfaces through weak and reversible interactions that are influenced by both hydrophobic and electrostatic forces [113]. The process of adhesion is facilitated through the presence of specific host tissue receptors, notably the glycoproteins ALS (agglutinin-like sequence) [114] and HWP1 (hyphal wall protein) [100]. C. albicans can adhere to epithelial cells using various host cell receptors, such as EphA2 (through β-glucan) and E-cadherin (through ALS3).
Additionally, host cell transglutaminases create direct links between C. albicans and the epithelial surface by interacting with HWP1. Among the family of eight glycosylphosphatidylinositol (GPI)-linked Als proteins, ALS3 stands out as a pivotal player in mediating the attachment of fungal hyphae to epithelial cells and facilitating their subsequent penetration into host cells [115,116]. HWP1 exhibits elevated expression levels during oral infections in humans [117], and it serves as a target for epithelial transglutaminases, facilitating robust and covalent connections with various epithelial proteins [118]. Interaction with oral bacteria can promote biofilm formation. Candida spp. readily forms biofilms, microbial communities encased in an extracellular matrix. Biofilms contribute to increased virulence and resistance to antimicrobial agents. In oral candidiasis, biofilms on dentures are a major factor [119]. For instance, Streptococcus gordonii is a common resident that interacts with C. albicans in the oral cavity, promoting the formation of hyphal structures and biofilms. This interaction is facilitated by the direct binding of ALS3 on C. albicans to the surface protein SspB (cell wall-associated protein) on S. gordonii, highlighting the complex interactions between microbes in the oral environment that impact biofilm formation and colonization [120]. C. albicans can change into an invasive filamentous form after adhering to host surfaces, which significantly improves its ability to penetrate epithelial tissue [121,122].
Candida can phenotypically switch between its yeast and hyphal states. C. albicans worsens host tissue damage by releasing enzymes outside the cell. Candida secretes enzymes like secreted aspartyl proteinases (SAPs), phospholipases, and lipases. These enzymes can degrade host immune factors, such as antibodies and antimicrobial peptides, reducing the effectiveness of host defences [123]. For tissue invasion, the shift to the hyphal form is crucial [124]. Hyphae can infiltrate and harm the epithelial cells, resulting in inflammation and distinctive white patches that appear in oral thrush. Thigmotropism, a directional growth response to surface contact, is a characteristic of hyphal cells and aids in their ability to successfully explore and infiltrate host tissues [125]. Candida can release candidalysin, a hypha-specific toxin that promotes immunological activation and tissue destruction [126]. Candidalysin, when acting upon oral epithelial cells, triggers the entry of calcium ions and the release of lactate dehydrogenase (LDH), both of which are indicative of cellular harm and destabilization of the cell membrane [127]. In addition to this, Candida engages in a complementary, passive process called endocytosis. ALS3 plays a dual role as both an adhesin and an invasion, and it collaborates with Ssa1, a heat shock protein. Together, they facilitate the endocytosis of C. albicans into epithelial cells through their interaction with E-cadherin. In this process, fungal enzymes and invasins on the hyphae bind to and degrade inter-epithelial cell junctional proteins like E-cadherin, enabling the yeast to penetrate and enter host epithelial cells [128].
Oral Microbiome and Systemic Diseases
Gastrointestinal Disorder: Inflammatory Bowel Disease
Inflammatory bowel disease (IBD), a chronic inflammatory condition affecting the gut, is known to be associated with oral microbial dysbiosis [56]. In healthy conditions, the mucosal barrier largely prevents microbial translocation from the oral cavity to the gut [129]. However, IBD’s altered gut epithelial permeability allows oral-resident bacterial strains to infiltrate the gut microbiome [130,131]. F. nucleatum, typically an oral inhabitant, has been detected in the intestines of IBD patients, underscoring the potential oral-gut axis in disease pathogenesis [132]. This notion is supported by in vivo studies wherein oral microbiota transplantation led to gut dysbiosis and exacerbated visceral hypersensitivity, further accentuating the role of oral microbes in influencing gut health [85,133]. Periodontitis exhibits a strong association with IBD. Keystone pathogen P. gingivalis disrupts intestinal barrier function, impacting the gut microbiome composition and inciting inflammation [83,87]. P. gingivalis enters the gastrointestinal tract and triggers an inflammatory response in the ileum. This inflammation is a localised immune reaction intended to combat the invading pathogen. During the inflammatory response, there is an activation and proliferation of a particular subset of immune cells known as IL9+ CD4+ lamina propria T cells. These T cells are known for their production of interleukin-9 (IL-9), a cytokine that plays a role in immune responses [134] (Figure 1).
Moreover, paediatric Crohn’s Disease (CD) patients, a type of IBD, exhibited enrichment of resident oral bacteria, including Enterobacteriaceae, in the gut [133]. Klebsiella spp. derived from the oral cavity have been identified as potent inducers of Th1-mediated inflammation in the gut. This implies a potential contribution of oral bacteria to CD pathogenesis [59]. Collectively, the oral microbiome manifests in IBD’s complex aetiology. The oral-gut axis suggests that oral dysbiosis can influence IBD pathogenesis while alterations in gut health reciprocally impact the oral microbiome [56]. It is promising that further research into the precise mechanisms and microbes responsible for these interactions will lead to new strategies for treating IBD.
Cardiovascular Diseases: Atherosclerosis
Studies have revealed the connection between oral dysbiosis and cardiovascular diseases (CVD), as well as the impact of CVD on the oral microbiota. In the context of periodontal diseases, an increase in gram-negative bacterial populations, such as P. gingivalis, T. denticola, and T. forsythia, can exacerbate CVD by triggering local inflammation, which in turn stimulates systemic inflammatory responses, oxidative stress, immune activation, and platelet aggregation factors associated with CVD [135,136]. Pathogens such as S. mutans [137] and P. gingivalis [138] are increased in patients with both periodontal disease and atherosclerosis. Emerging evidence suggests that the relationship between oral microbiota and CVD is bidirectional, with CVD impacting the oral microbiota through systemic inflammation and immune activation. Microbial invasion of the bloodstream [139], facilitated by oral-gut microbiota transfer [61,140] and bacteremia, may exacerbate systemic inflammation, leading to a cascade of effects that further disrupt the oral microbial balance.
Metabolites produced by the oral microbiota are gaining recognition for their potential to influence cardiometabolic health. Elevated concentrations of bacterial surface molecules like lipopolysaccharides (LPS) and bacterial flagellins stimulate the production of pro-inflammatory mediators and cytokines [141,142]. These molecules trigger inflammatory pathways such as the nuclear factor kappa-B (NF-κB) and matrix metalloproteinase 9 (MMP9) pathways, contributing to inflammation and immunoreactivity [138]. Aggregatibacter actinomycetemcomitans, linked to chronic periodontitis, releases leukotoxin A (LtxA), which kills white blood cells by causing cofilin dephosphorylation and actin depolymerization [143]. LtxA disrupts enzyme function in neutrophils, causing hypercitrullination and releasing altered proteins that could trigger autoimmune responses, possibly contributing to atherosclerosis [144]. Several additional bacterial species have also been implicated in the interaction between disrupted oral microbial balance and CVD. These species, including Prevotella intermedia, Prevotella nigrescens, Campylobacter rectus, Parvimonas micra, P. endodontalis, Eubacterium timidum, Eubacterium brachy, and Eubacterium saphenum, have been linked to both oral dysbiosis and the development of CVD [145,146,147].
Apart from this, autoimmune processes constitute a crucial mechanism in the development of atherosclerosis. One intriguing aspect of this immune response involves heat shock proteins (HSPs), which are part of a family of molecular chaperones that protect cells during stress [148]. These proteins are present in the host as well as in bacteria, including P. gingivalis. In the case of P. gingivalis, its HSP60 equivalent, known as GroEL, can trigger both a humoral and cellular immune response in humans [149]. This immune response can lead to chronic activation, as antibodies originally primed for bacterial HSPs may cross-react with human HSP60 due to structural similarities. This chronic immune activation fuels inflammation within arterial walls, promoting the accumulation of plaque composed of cholesterol, immune cells, and debris. Over time, plaque buildup narrows arteries, reducing blood flow and raising the risk of cardiovascular events like heart attacks and strokes when unstable plaques rupture [144].
Endocrine Disorders: Diabetes Mellitus
Diabetes mellitus, marked by hyperglycemia and inflammation coupled with elevated oxidative stress, creates an environment conducive to microbial dysbiosis. Subsequently, alterations within the oral microbiome have been reported, with distinct bacterial shifts observed in diabetic subjects [150]. Elevated proportions of Capnocytophaga [151], P. gingivalis, T. forsythia [152,153], and others have been noted, hinting at potential microbial contributors to diabetic-associated oral complications. However, findings remain diverse and inconsistent, partly due to limited bacterial profiling and confounding factors. A newly gained understanding of the underlying mechanisms has revealed how diabetes-induced changes in host responses play a significant role [154]. Individuals with diabetes experience elevated inflammatory reactions, impaired neutrophil activity, and a diminished ability to clear bacteria. This cascade of events contributes to exacerbated periodontal inflammation and tissue damage. Moreover, the dysregulated cytokine environment, including tumor necrosis factor (TNF), fuels inflammation, exacerbating periodontal degradation [155,156]. Oral microbes in diabetic individuals exhibit high virulence, potentially fueling disease progression. Diabetes-induced IL-17 elevation further amplifies oral microbial pathogenicity, intensifying the risk of periodontal deterioration [150]. Insightfully, interventions targeting IL-17 have shown promise in mitigating the pathogenic potential of the oral microbiome in diabetes.
Obesity
Obesity is characterised by chronic low-grade inflammation, recognised as a key link between obesity and metabolic dysfunction [157,158]. This interaction, termed “meta inflammation,” is particularly notable in adipose tissue [159]. Recent research indicates a connection between obesity and the oral microbiome, suggesting oral bacteria might play a role in obesity’s development and progression [160]. Distinct shifts in the composition of the oral microbiome have been observed in individuals with obesity compared to non-obese individuals [161]. Intriguingly, lean (non-obese) individuals showcase elevated populations of Firmicutes, Bacteroidetes, and Spirochaetes [162,163] in sublingual plaque, while their obese counterparts exhibit increased levels of Proteobacteria, Chloroflexi, and Firmicutes [164,165]. Alterations in bacterial populations have been associated with changes in metabolic homeostasis. An area of investigation focuses on the migration of oral bacteria to the gut, subsequently influencing the composition of the gut microbiome. This “oral-gut-axis” hypothesis suggests that oral microbes can influence distant organs, potentially contributing to systemic inflammation and metabolic perturbations. Additionally, the concept of an “oral-blood-axis” highlights the potential for oral bacteria and inflammatory molecules to enter the bloodstream, further propagating inflammation and metabolic dysfunction [158]. Moreover, studies have identified a significant link between periodontal disease, oral dysbiosis, and obesity-associated inflammation [166]. Intriguingly, overweight individuals exhibit altered salivary bacterial composition, possibly serving as early indicators of developing obesity.
Neurological Disorders: Alzheimer’s Disease
Alzheimer’s disease (AD) is a progressive and neurodegenerative disorder predominantly affecting individuals aged over 65 years, characterized by significant cognitive decline that impairs daily activities [167]. Recent research has unveiled a robust connection between AD and the composition of the oral microbiota. Notably, specific pathogens within this microbial array, namely P. gingivalis, T. forsythia, and T. denticola, play central roles in periodontitis, inciting a cascade of inflammatory responses that have been implicated in AD progression [168,169,170,171]. The oral microbiota’s production of inflammatory agents introduces a novel perspective on systemic infection and inflammation. These mediators can traverse the bloodstream, influencing remote tissues and organs, including the brain. This suggests that the oral microbiota may act as a potential primary agent for AD, provoking neuroinflammation and contributing to the disease’s aetiology [172]. Interestingly, P. gingivalis, an oral pathogen, has been detected in brain tissue [167]. It can directly impact AD through various mechanisms. These microbes gain access to the brain via routes such as transient bacteremia, root canal pathways, or gingival crevices, where they provoke direct damage to the central nervous system (CNS), initiating an immune cascade characterised by microglial activation and inflammatory responses [173]. This process may induce the production of Aβ protein, which, while normally protective, could be overproduced in response to oral microbes, potentially contributing to the formation of Aβ plaques in AD [174].
Furthermore, the blood-brain barrier (BBB) may become more permeable in elderly AD patients, aiding oral microbe entry. Some oral microorganisms and their secreted substances can even directly alter BBB permeability. Indirectly, chronic oral diseases weaken the oral mucosal barrier, permitting cytotoxins and pro-inflammatory factors to enter the bloodstream and trigger systemic inflammation. These factors can then reach the CNS, indirectly influencing AD by inciting neuroinflammation and cognitive impairment associated with the disease [175]. While Aβ peptides have demonstrated antimicrobial efficacy against a range of bacteria and Candida albicans, their prolonged accumulation due to persistent colonisation by pathogenic oral bacteria might lead to the formation of amyloid plaques and tau protein hyperphosphorylation, ultimately culminating in brain tissue damage [173,176].
Parkinson’s Disease
Parkinson’s Disease (PD), a prevalent neurodegenerative disorder primarily recognised for its impact on movement control, poses significant challenges to the ageing population [177,178]. It is a disorder that affects memory. PD’s core manifestation lies in compromised fine and gross motor skills. The onset of PD, frequently late in life, may not exclusively stem from genetic inheritance but rather a combination of multifactorial contributors, including genetic predisposition, environmental risks, and ageing [179]. Recently, many studies have shown a correlation between P. gingivalis infection and PD [180,181,182,183]. Studies have revealed that gingipain R1 (RgpA), a virulence factor produced by P. gingivalis, can be detected in the bloodstream, indicating systemic dissemination of the bacterium [180]. Additionally, PD patients exhibit a hypercoagulable state characterized by hyperactivated platelets and fibrinogen amyloid features, which may be influenced by P. gingivalis and its products [180,184]. Furthermore, P. gingivalis is implicated in inducing systemic inflammation, as evidenced by the presence of its lipopolysaccharides (LPS) and gingipain in circulation [184]. This systemic inflammation is known to play a role in the pathogenesis of PD. Animal studies have provided further insights into the potential mechanisms linking P. gingivalis to PD. Oral administration of live P. gingivalis to mice with a genetic mutation associated with PD leads to neurodegeneration in the substantia nigra, activation of microglia, and upregulation of IL-17A secretion and its receptor IL-17RA [183]. These findings suggest that P. gingivalis may contribute to PD pathogenesis by inducing systemic inflammation, promoting hypercoagulability, and exacerbating neurodegeneration. However, further research is needed to fully elucidate the complex interplay between oral microbiota, systemic inflammation, and neurodegenerative diseases like PD.
Autoimmune Conditions: Rheumatoid Arthritis
Rheumatoid arthritis (RA), a chronic autoimmune disease characterised by symmetrical inflammation and joint pain, has a complex aetiology involving genetic and environmental factors [185]. Recent research has unveiled a compelling connection between the oral microbiota and RA pathogenesis, shedding light on the role of specific microorganisms and their molecular mechanisms in triggering and exacerbating the disease. Among the key players, P. gingivalis stands out as a significant pathogen implicated in both periodontitis and RA [186]. P. gingivalis is equipped with virulent factors, including the bacterial protein arginine deiminase (PPAD), capable of citrullinating host proteins [187]. This citrullination has been postulated to initiate an autoimmune response, culminating in the production of anticitrullinated protein antibodies (ACPAs) that are characteristic of RA. Neutrophil extracellular traps (NETs) generated during infection further contribute to the citrullination of proteins catalysed by PAD4 enzymes and ACPA production [188,189]. The presence of ACPAs has been correlated with P. gingivalis exposure and has been proposed as a crucial link between periodontal inflammation and RA onset (Figure 3).
Beyond P. gingivalis, a broader dysbiosis of the oral microbiota has appeared as a potential factor in RA development. Elevated levels of Prevotella, Veillonella, and anaerobic species, such as Lactobacillus salivarius have been observed in RA patients [190]. Conversely, decreased levels of beneficial microbes, such as Haemophilus spp., have been noted. This dysbiosis can lead to altered immune responses, including the overactivation of T helper 17 (Th17) cells, which play a pivotal role in autoimmune processes. Th17 cells produce pro-inflammatory cytokines like IL-17 [191]. Elevated levels of IL-17 have been found in the synovium of RA patients and are associated with disease severity [192]. Moreover, the interplay between microbial communities and immune cells can exacerbate joint inflammation and bone destruction in RA [193]. Recent investigations have also implicated A. actinomycetemcomitans, a periodontal bacterium, in RA pathogenesis. Leukotoxin A (LtxA) secreted by A. actinomycetemcomitans triggers neutrophil hypercitrullination and the subsequent release of citrullinated autoantigens, further fueling the ACPA response [194]. Some oral microbiome molecules structurally resemble human self-antigens, potentially leading to cross-reactivity with host proteins. For instance, P. gingivalis’ α-enolase protein shares homology with human α-enolase, an autoantigen in RA. Antibodies against citrullinated α-enolase (anti-CEP-1) have been found in individuals with chronic PD, suggesting a connection [195].
Systemic Lupus Erythematosus
Systemic Lupus Erythematosus (SLE) is a complex autoimmune disease that impacts various organs and tissues, disproportionately affecting women of reproductive age [190]. Although its aetiology remains elusive, recent research has shed light on the potential involvement of the oral microbiome in SLE pathogenesis. Numerous studies have demonstrated alterations in the oral microbiota composition in individuals with SLE. This microbial dysbiosis extends beyond the oral cavity, possibly contributing to the systemic inflammation characteristic of SLE [196]. A study conducted by Pessoa et al. made a significant observation regarding the oral microbiota in individuals with SLE compared to healthy controls. 14 distinct subgingival bacteria in SLE patients were enriched, suggesting a distinct microbial profile associated with the disease. Interestingly, this elevated microbial abundance was particularly evident in certain bacteria, specifically T. denticola and T. forsythia [197]. Moreover, an increased diversity of oral microbiota was observed in SLE patients’ saliva, with Veillonella, Streptococcus, and Prevotella as the dominant species [198].
Periodontitis may exacerbate SLE through activation the of Toll-like receptors (TLRs), such as TLR2 and TLR4. These receptors are pivotal in recognising microbial components and initiating immune responses [199]. The interesting concept of molecular mimicry arises in the context of SLE. Cross-recognition between self-antigens, like Ro60, and homologous peptides from oral bacteria suggests a potential role for bacterial antigens in triggering autoimmune responses. These observations highlight the oral microbiota’s capacity to induce and perpetuate aberrant immune reactions in SLE [199]. Elevated levels of pro-inflammatory cytokines, such as interleukin-6 (IL-6), IL-17A, and IL-33, have been detected in the saliva of SLE patients with chronic periodontitis. These cytokines play a pivotal role in promoting immune cell activation and differentiation, thereby intensifying autoimmune responses and tissue damage [200,201,202]. The complicated relationship between oral microbial dysbiosis, periodontitis-induced inflammation, and systemic immune dysregulation emphasises the potential impact of the oral microbiota on SLE progression.
Cancer
Within the realm of cancer research, specific bacterial strains have emerged as influential contributors to cancer progression, shedding light on the intricate connections between the oral microbiome and this complex disease. Notably, two bacterial strains, particularly F. nucleatum and P. gingivalis, have been recognised as agents that contribute to cancer progression. F. nucleatum employs a range of surface proteins to infiltrate diverse cell types, potentially initiating tumor development in the mouth, colon, and oesophagus. Similarly, P. gingivalis is connected to periodontal harm and can support tumor formation by invading neighbouring tissues. This bacterium can also travel to other sections of the oral and digestive systems, and its association with pancreatic cancer has been observed [203]. Certain bacteria produce specific substances with potent cancer-causing properties. For instance, lipopolysaccharide, which is generated by numerous anaerobic oral microorganisms, including S. oralis, S. mitis, S. sanguinis, Lactobacillus fermentum, Lactobacillus acidophilus, and Bifidobacterium adolescentis, can induce inflammatory mechanisms linked to the development of cancer [204].
A comprehensive overview of the associations between the oral microbiome and various diseases is mentioned in Table 2. It is noteworthy that the organisms discussed are referred to as commensal when they are present at low levels, typical of a healthy microbiome.
Oral Health and Dental Care Practices
Probiotics
Dental probiotics contribute to the balance of the oral microbiota by colonising the oral mucosa, producing specialised metabolites to maintain the host microbiota’s ecological equilibrium, and promoting a healthy level of immunity [205]. Scientifically studied probiotic strains such as Streptococcus salivarius M18, Streptococcus salivarius K12, Lactobacillus plantarum, Bifidobacterium lactis, Lactobacillus reuteri, and Lactobacillus salivarius have demonstrated their efficacy in supporting oral health. These strains are known to combat the bacteria responsible for bad breath and gum inflammation. S. salivarius can generate powerful antibacterial proteins known as bacteriocins, which can be harmful for the growth of various bacterial species [206]. Furthermore, laboratory investigations have revealed that S. salivarius engages directly with the gingival epithelium [207], exhibiting anti-inflammatory characteristics and fostering a harmonious immune response [208,209].
Specifically, two strains of L. reuteri (DSM 17938 and ATCC PTA 5289) have been shown to significantly reduce halitosis [210]. L. salivarius (WB21 and TI 2711) may reduce dental caries through mechanisms including coaggregation and growth inhibition of S. mutans, as well as reducing plaque acidogenicity [211]. The probiotic strain Bifidobacterium animalis subsp. lactis (B. lactis) HN019 showed significant improvement in clinical periodontal indicators, a more efficient reduction of periodontal pathogens, and a decrease in proinflammatory cytokine levels within the gingival crevicular fluid [212]. Moreover, L. plantarum exhibited notable effectiveness in inhibiting the growth of S. mutans and C. albicans, disrupting the formation of virulent biofilms, and influencing gene expression related to crucial metabolic pathways, fungal resistance, and oxidative stress. These findings underscore the potential of L. plantarum as a promising candidate for controlling cariogenic biofilms and, consequently, for the prevention of dental caries [213].
Peptides in Oral Health and Their Role in Oral Care
Peptides, short chains of amino acids, play critical roles in various biological processes, including promoting health and healing [214]. In the past twenty years, peptides have gained significant attention in the dental field [215,216]. These peptides have been applied across various dental specialties, including endodontic therapy, restorative dentistry, caries control, bone and dental tissue remineralization, and the enhancement of dental materials [215,217]. The integration of peptides in oral care is an emerging area of research with the potential to revolutionize oral health practices.
Antimicrobial peptides (AMPs) such as defensins and cathelicidins are known for their ability to inhibit the growth of oral bacteria, fungi, and viruses [218]. These peptides disrupt the integrity of microbial cell membranes, providing a natural defense against oral pathogens [219]. AMPs exhibit a range of mechanisms to target and inhibit microbes. Primarily, they interact electrostatically with negatively charged microbial membranes due to their cationic nature. This leads to the formation of pores, disrupting membrane integrity and increasing permeability. AMPs allow leakage of ions, metabolites, and other vital cellular components, ultimately causing microbial cell death. The interfacial activity of AMPs enables them to partition into the membrane-water interfaces, modifying lipid structures. While eukaryotic cell membranes remain unaffected due to their neutral charge, the specific interactions with microbial membranes make AMPs selective and effective in combating pathogens [218].
Naturally occurring peptides in saliva, such as histatins, play a key role in oral health. They possess antimicrobial properties and contribute to maintaining oral tissue integrity, helping to prevent infections and inflammation [220]. Histatins interact with the plasma membrane of pathogens, forming pores that disrupt membrane integrity. This action allows for the leakage of ions such as potassium, as well as other intracellular components, ultimately impairing the pathogen’s cellular functions and leading to cell death. Histatins bind to the Trk1 potassium transporter on yeast cells, causing potassium efflux and further destabilizing the cell’s internal environment. Histatins exhibit both fungistatic (inhibiting fungal growth) and fungicidal (killing fungi) effects, particularly against Candida albicans [221]. Among the histatins, Histatin-5 is the most effective, followed by Histatin-3 and then Histatin-1 [222].
Enamel strengthening peptides, such as peptide P11-4, are an innovative development in dental health, designed to support the remineralization and strengthening of tooth enamel [223]. The biomimetic nature of P11-4 allows it to closely resemble the mineral structure of enamel, aiding the regeneration and remineralization of dental hard tissues [223]. Once applied to the tooth surface, P11-4 forms scaffold-like structures with negative charge domains that attract calcium ions, inducing the precipitation of hydroxyapatite, the mineral essential for tooth enamel [224]. Combined with fluoride, P11-4 enhances the remineralization process by incorporating calcium and phosphate ions into the enamel structure [223]. Studies, including randomized clinical controlled trials, have demonstrated the safety and efficacy of P11-4 in promoting enamel health [223].
Personalised Oral Care Approaches
In addition to probiotics, a comprehensive oral care regimen can support the natural healing of teeth and gums, aligning with the body’s inherent mouth chemistry for enhanced oral health. In this regimen xylitol, is a vital component that helps rebalance mouth acidity. Regular use of xylitol mints and gums has been associated with decreased counts of S. mutans, potentially reducing the adherence of plaque [225]. These products consist of xylitol, natural peppermint flavour, magnesium stearate, and natural menthol. Xylitol being the crucial component, is the one that decreases the presence of S. mutans, which is responsible for tooth decay. Xylitol, once taken up by S. mutans, significantly disrupts the bacterium’s usual energy production pathways, fundamentally altering its growth and survival mechanisms. Unlike other sugars, xylitol cannot be processed by S. mutans for energy, initiating a futile energy cycle that derails the bacterium’s normal metabolic processes. This disruption results in the conversion of xylitol to xylitol-5-phosphate within the bacterial cell, causing the formation of vacuoles and damaging the cell membrane. These alterations impede the bacterium’s growth and viability. Furthermore, the attempt by S. mutans to expel the dephosphorylated xylitol-5-phosphate from the cell, with no energy yield from xylitol metabolism, essentially starves the bacterium, hindering its growth and contributing to its eventual death [226].
Following in the regimen is the mouth rinse, comprising stabilized chlorine dioxide (ClO2), trisodium phosphate, and citric acid. This distinct formulation highlights its capacity to offer therapeutic advantages, making it an attractive option for patients concerned about taste preferences and tooth discolouration during oral health treatment [227]. ClO2, being the compound of interest acts on volatile sulfur compounds (VSCs), the chief contributors to oral malodor, through a sequential mechanism. Initially, ClO2 reacts with thiol compounds like L-cysteine, generating ClO2- and thiol radicals (RS.), subsequently converting thiol compounds into disulfides like cystine. This reduction of thiol compounds progresses through reactions involving the formation of disulfides, addressing malodorous thiols. Simultaneously, ClO2 showcases bactericidal properties by permeating bacterial cells and disrupting cytoplasmic amino acids and membrane proteins, leading to microbial cell death. The chemical reactions involved underline the efficacy of ClO2 in neutralizing malodor-causing compounds and eradicating odour-producing bacteria, essential for combating oral malodor [228].
Chemical reactions:
Reaction 1:
RSH (e.g., CH3SH) + ClO2 → RS. + ClO2.- + H+
Thiol compounds Chlorine Thiol Chlorine dioxide Hydrogen (e.g., L-cysteine) dioxide radical radical ion
Reaction 2:
2RS. → RSSR (e.g., CH3SSCH3)
Thiol radical Disulfides (e.g., cystine)
Reaction 3:
4RSH + ClO2.- → 2RSSR + Cl- + 2H2O Thiol compounds Chlorine dioxide Disulfides Chloride Water
(e.g., L-cysteine) radical (e.g., cystine) ions
Results from Shinada et al.‘s study [228], confirm that a seven-day regimen using a ClO2 mouth rinse significantly reduced morning oral malodor and lowered VSCs in healthy subjects. Additionally, it effectively reduced plaque, tongue coating, and F. nucleatum counts in saliva.
Alternatively, individuals can incorporate a mouth rinse into their oral care routine, which includes thymol, eucalyptol, menthol, and methyl salicylate dissolved in ethanol (27%) [229]. Thymol, derived from Thyme essential oil, is an essential component in antiseptic mouthwashes due to its potent antifungal properties. Its mechanism involves altering membrane lipids’ fluidity and permeability, disrupting the production of hyphae, which are essential structures for the pathogenicity of C. albicans. Additionally, thymol significantly reduces adhesion of C. albicans to epithelial cells, an initial step in infection development [230]. Eucalyptol, a primary constituent of Eucalyptus essential oil, features prominently in oral hygiene products due to its potent antimicrobial properties. Its efficacy extends to combating various oral pathogens like P. gingivalis and S. mutans, commonly associated with periodontitis and dental issues. Eucalyptol operates by disrupting bacterial cell membranes, notably the cytoplasmic membrane. Components like 1.8-cineole and terpinolene modify membrane permeability, compromising its integrity and allowing increased passage of ions and molecules, leading to cellular dysfunction and eventual bacterial demise [231]. The combined action of these ingredients aims to support oral health by targeting bacteria and promoting a clean and refreshed feeling in the mouth after use [232].
A fluoride toothpaste is an essential part of oral care, featuring sodium fluoride among other vital components like hydrated silica, cellulose gum, and glycerin. These ingredients collectively contribute to fortifying tooth enamel, preventing cavities, and promoting oral hygiene. Fluoride’s action in preventing tooth decay involves multiple mechanisms and chemical reactions. Fluoride ions (F-) can replace hydroxyl ions (OH-) in the hydroxyapatite crystal structure of enamel, forming fluorapatite (FAP) [233]. This substitution creates a more acid-resistant structure, making the enamel less susceptible to acids produced by plaque bacteria.
The chemical reaction can be represented as:
Hydroxyapatite (HA) + Fluoride ions (F-) → Fluorapatite (FAP) + Hydroxyl ions (OH-)
Fluoride deposition is higher in the outer enamel layers, enhancing their acid resistance and conferring caries resistance to the tooth. In an acidic environment, fluoride can form hydrogen fluoride (HF), an undissociated weak acid that can penetrate bacterial cell membranes. HF’s penetration into bacterial cells leads to two physiological impacts: interference with fluoride-sensitive enzymes and intracellular acidification, hindering essential enzymes necessary for cell growth. Fluoride, when present, combines with calcium to form calcium fluoride (CaF2) on plaque and enamel surfaces. CaF2 acts as a reservoir for fluoride. When pH drops due to acid, fluoride and calcium ions are released into plaque fluid. Fluoride then diffuses into enamel pores, forming FAP. FAP is more acid-resistant than hydroxyapatite, reducing demineralization and enhancing remineralization of enamel between pH 4.5-5.5. This process shortens the demineralization period and helps in maintaining enamel health [233,234]. Combinations such as using toothpaste containing fluoride concentrations ranging from 1100 ppm to 1500 ppm, either on its own or paired with a 0.05% sodium fluoride mouth rinse, have demonstrated considerable effectiveness in reducing the progression of root caries [235].
Fluoride has long been relied upon for its role in preventing dental cavities, yet the growing concerns regarding public acceptance and the potential risk of fluorosis in children highlight the need for alternative solutions. Toothpastes incorporating hydroxyapatite (HA) have emerged as fluoride-free options and have demonstrated promising efficacy as anti-cavity agents in recent research [236]. HA toothpaste works by replenishing lost minerals in enamel and restoring its structure. It binds to damaged enamel, filling in surface irregularities and promoting deeper penetration into lesions for comprehensive repair. Mimicking natural enamel, HA particles effectively remineralize teeth, offering an alternative to fluoride-based products [236].
Cetylpyridinium chloride (CPC), a quaternary ammonium compound, and a prevalent antiseptic found in various oral care formulations like mouthwashes and toothpaste [237]. CPC serves as a potent antimicrobial in oral care by engaging in a multifaceted role on bacterial membranes and cellular functions, ensuring its efficacy in combating plaque-forming bacteria for maintaining oral hygiene. Its positive charge enables interaction with negatively charged bacterial membranes, disrupting the balance of ions like Mg2+ and Ca2+ and integrating its hydrophobic tail into the lipid bilayer. At lower concentrations, CPC disrupts bacterial osmoregulation, causing leakage of vital cellular components and potentially activating autolytic enzymes. Higher concentrations lead to extensive membrane disintegration, resulting in leakage of cytoplasmic contents, and damage to proteins, nucleic acids, and cell walls. CPC’s effectiveness extends to Gram-negative bacteria, bypassing their complex cell walls due to their molecular size and leveraging damaged cell walls to enhance their penetration. Additionally, CPC’s surfactant properties facilitate uniform coverage of irregular surfaces, aiding in biofilm disruption [237].
Clove oil, primarily composed of eugenol, eugenyl acetate, carvacrol, and other compounds, has gained attention in dental health due to its therapeutic properties. Clove oil demonstrates potent antibacterial effects against multi-resistant Staphylococcus species. Its action inhibits the growth and proliferation of these bacteria, aiding in controlling oral infections. With its antifungal activity, clove oil impedes the growth of fungi like C. albicans. It reduces ergosterol levels, a crucial component of fungal cell membranes, and inhibits germ tube formation by C. albicans, essential for its pathogenicity. The oil exhibits strong radical scavenging activity, indicating its antioxidant potential. This property helps combat oxidative stress in the oral cavity, contributing to overall oral health [238]. In dentistry, clove oil finds application in various forms, including mouth rinses or gels. Its mode of action involves disrupting microbial cell membranes, inhibiting essential cellular processes, and interfering with the structural integrity of pathogens. These actions collectively contribute to its efficacy in preventing and managing oral infections and inflammation.
Oil pulling is a traditional oral hygiene technique involving swishing oil, typically coconut, sesame, or sunflower oil, in the mouth for around 15–20 minutes [239]. Oil pulling generates antioxidants that damage the cell walls of microorganisms, killing them effectively. The oils attract the lipid layer of bacterial cell membranes, causing them to stick to the oil and eventually get removed from the oral cavity. The oil gets emulsified during the swishing process, increasing its surface area and efficacy. The oil coats the teeth and gums, hindering bacterial co-aggregation and plaque formation. It effectively removes plaque-building bacteria responsible for various oral issues like dental caries, gingivitis, and bad breath. Coconut oil, particularly due to its lauric acid content, exhibits potent antimicrobial properties. It effectively fights Streptococcus mutans and Candida albicans in biofilm models. The lauric acid in coconut oil reacts with saliva’s alkalis to form a substance that reduces plaque adhesion and accumulation, aiding in cleansing the oral cavity. Lauric acid also prevents dental caries, possesses anti-inflammatory properties, and promotes overall oral health. Its pleasant taste adds to its appeal [239,240] (Table 3).
Practicing daily dental care (Supplementary Material 2) with the above-mentioned key components, and strategies may ensure the removal of plaque, resolution of gingivitis, prevention of cavities, treatment of periodontal disease, and management of gum issues, all while fostering a harmonious and healthy oral microbiome.
Conclusion
In conclusion, the oral microbiome is a diverse and complex ecosystem that plays a significant role in maintaining oral and systemic health. Comprising a plethora of bacterial, fungal, and archaeal species, the oral microbiome’s composition and dynamics are intricately linked to various physiological processes. Disruptions in this delicate balance, leading to oral dysbiosis, can precipitate a spectrum of oral ailments like dental caries, gingivitis, periodontitis, halitosis, and oral candidiasis, while also exerting profound effects on systemic health, including gastrointestinal disorders, cardiovascular diseases, endocrine imbalances, and neurological conditions. The interplay between the oral and gut microbiomes, underscored by microbial translocation, emphasize the bidirectional relationship between oral and systemic health, wherein disturbances in either ecosystem can cascade into the other, exacerbating disease states. Recent research illuminating connections between specific oral bacteria, such as P. gingivalis, and systemic diseases like IBD, cancer, Alzheimer’s, and RA, bring out the far-reaching impact of oral health on overall health outcomes.
As our understanding of the oral microbiome expands, it becomes increasingly evident that fostering a balanced oral microbial ecosystem is paramount for holistic health. Embracing evidence-based dental care practices, such as integrating dental probiotics like S. salivarius and Lactobacillus strains, alongside utilizing oral ingredients like xylitol, fluoride toothpaste, and essential oil-based mouth rinses, presents a comprehensive approach to oral health maintenance. One could test for these problems and arrive at a personalized dental care plan to address specific needs. These interventions, rooted in scientific validation, offer promising avenues for preventing oral dysbiosis and tackling common oral health issues like bad breath, gum inflammation, and dental caries. By adopting these strategies, individuals can actively contribute to nurturing a balanced oral microbiome, thus safeguarding not only their oral health but also their systemic well-being.
Authors’ contributions
Conception: HKK, JJR, and VJ. Manuscript preparation: HKK, JB, VJ, KK, TW, KB and JJR.
Funding
Vibrant America provided funding for this study in the form of salaries for authors [HKK, JB, VJ, KK, TW, KB and JJR]. The specific roles of these authors are articulated in the ‘author contributions’ section. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Availability of data and material
The data used to support the findings of this study can be acquired from Vibrant America LLC.
Consent for publication
NA
Acknowledgement
We acknowledge Vibrant America LLC for supporting this research.
Competing interests
The authors have read the journal’s policy and the authors of this manuscript have the following competing interests: HKK, VJ, KK, TW, KB, and JJR are paid employees of Vibrant Sciences LLC. JB is paid employee of Vibrant America LLC. Vibrant Sciences or Vibrant America is a commercial lab which performs saliva testing for oral micro-organisms. Vibrant Sciences or Vibrant America could benefit from increased testing based on the observations. There are no patents, products in development, or marketed products to declare. This does not alter our adherence to Microorganisms policies on sharing data and materials.
Abbreviations
| ACPAs | Anticitrullinated Protein Antibodies |
| α-synuclein | Alpha-synuclein |
| ALS | Agglutinin-like sequence |
| AD | Alzheimer’s Disease |
| BBB | Blood-Brain Barrier |
| CaF2 | Calcium Fluoride |
| CD | Crohn’s Disease |
| CH3SH | Methyl mercaptan |
| CH3SSCH3 | Dimethyl sulfide |
| CPC | Cetylpyridinium chloride |
| CVD | Cardiovascular Diseases |
| EPS | Extracellular polymers |
| F- | Fluoride ions |
| FAP | Fluorapatite |
| GPI | Glycosylphosphatidylinositol |
| HA | Hydroxyapatite |
| HF | Hydrogen Fluoride |
| HWP1 | Hyphal wall protein |
| HSPs | Heat Shock Proteins |
| IBD | Inflammatory Bowel Disease |
| IL | Interleukin |
| LDH | Lactate Dehydrogenase |
| LPS | Lipopolysaccharides |
| LtxA | Leukotoxin A |
| MMP9 | Matrix Metalloproteinase 9 |
| nAChRs | Nicotine Acetylcholine Receptors |
| NETs | Neutrophil Extracellular Traps |
| NF-κB | Nuclear Factor Kappa-B |
| NGS | Next-Generation Sequencing |
| OH- | Hydroxyl ions |
| PD | Parkinson’s Disease |
| PPAD | Porphyromonas gingivalis peptidylarginine deiminase |
| RA | Rheumatoid Arthritis |
| RS. | Thiol radicals |
| RSH | Thiol compounds |
| RSSR | Disulfides |
| RT-PCR | quantitative Real-Time PCR |
| SAPs | Secreted Aspartyl Proteinases |
| SLE | Systemic Lupus Erythematosus |
| SspB | Surface protein SspB |
| Th | T helper |
| VSCs | Volatile Sulfur Compounds |
References
- Deo, P.; Deshmukh, R. Oral microbiome: Unveiling the fundamentals. J. Oral Maxillofac. Pathol. 2019, 23, 122–128. [Google Scholar] [CrossRef] [PubMed]
- The oral microbiome in health and disease - PubMed [Internet]. [cited 2023 Sep 19]. Available from: https://pubmed.ncbi.nlm.nih.gov/23201354/.
- Kilian, M.; Chapple, I.L.C.; Hannig, M.; Marsh, P.D.; Meuric, V.; Pedersen, A.M.L.; Tonetti, M.S.; Wade, W.G.; Zaura, E. The oral microbiome – an update for oral healthcare professionals. Br. Dent. J. 2016, 221, 657–666. [Google Scholar] [CrossRef]
- Forshaw, R.J. Dental health and disease in ancient Egypt. Br. Dent. J. 2009, 206, 421–424. [Google Scholar] [CrossRef] [PubMed]
- The oral microbiome in health and disease and the potential impact on personalized dental medicine - PubMed [Internet]. [cited 2023 Sep 19]. Available from: https://pubmed.ncbi.nlm.nih.gov/21902769/.
- He, J.; Li, Y.; Cao, Y.; Xue, J.; Zhou, X. The oral microbiome diversity and its relation to human diseases. Folia Microbiol. 2014, 60, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Minty, M.; Vinel, A.; Canceill, T.; Loubières, P.; Burcelin, R.; Kaddech, M.; Blasco-Baque, V.; Laurencin-Dalicieux, S. Oral Microbiota: A Major Player in the Diagnosis of Systemic Diseases. Diagnostics 2021, 11, 1376. [Google Scholar] [CrossRef] [PubMed]
- Exploring the Human Microbiome: The Potential Future Role of Next-Generation Sequencing in Disease Diagnosis and Treatment - PubMed [Internet]. [cited 2023 Sep 19]. Available from: https://pubmed.ncbi.nlm.nih.gov/30666248/.
- Lee, Y.-H.; Chung, S.W.; Auh, Q.-S.; Hong, S.-J.; Lee, Y.-A.; Jung, J.; Lee, G.-J.; Park, H.J.; Shin, S.-I.; Hong, J.-Y. Progress in Oral Microbiome Related to Oral and Systemic Diseases: An Update. Diagnostics 2021, 11, 1283. [Google Scholar] [CrossRef]
- Sharma, N.; Bhatia, S.; Sodhi, A.S.; Batra, N. Oral microbiome and health. AIMS Microbiol. 2018, 4, 42–66. [Google Scholar] [CrossRef]
- The human oral microbiome - PubMed [Internet]. [cited 2023 Sep 19]. Available from: https://pubmed.ncbi.nlm.nih.gov/20656903/.
- Bacterial interactions in dental biofilm - PubMed [Internet]. [cited 2023 Sep 19]. Available from: https://pubmed.ncbi.nlm.nih.gov/21778817/.
- Genetic and physiologic characterization of urease of Actinomyces naeslundii - PubMed [Internet]. [cited 2023 Sep 19]. Available from: https://pubmed.ncbi.nlm.nih.gov/9916052/.
- Zhang, Y.; Ding, Y.; Guo, Q. Probiotic Species in the Management of Periodontal Diseases: An Overview. Front. Cell. Infect. Microbiol. 2022, 12, 806463. [Google Scholar] [CrossRef] [PubMed]
- Marsh, PD. Role of the Oral Microflora in Health. Microb Ecol Health Dis. 2000 Jan 1;12(3):130–7.
- Bradshaw, D.J.; Marsh, P.D.; Watson, G.K.; Allison, C. Role of Fusobacterium nucleatum and Coaggregation in Anaerobe Survival in Planktonic and Biofilm Oral Microbial Communities during Aeration. Infect. Immun. 1998, 66, 4729–32. [Google Scholar] [CrossRef]
- Könönen, E.; Fteita, D.; Gursoy, U.K.; Gursoy, M. Prevotella species as oral residents and infectious agents with potential impact on systemic conditions. J. Oral Microbiol. 2022, 14, 2079814. [Google Scholar] [CrossRef]
- Ecology of the Oral Microbiome: Beyond Bacteria - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/28089325/.
- Diaz, P.; Dongari-Bagtzoglou, A. Critically Appraising the Significance of the Oral Mycobiome. J. Dent. Res. 2020, 100, 133–140. [Google Scholar] [CrossRef]
- Candida and other fungal species: forgotten players of healthy oral microbiota - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/24487378/.
- Diversity and quantitative analysis of Archaea in aggressive periodontitis and periodontally healthy subjects - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/21539593/.
- Methanogenic Archaea and human periodontal disease - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/15067114/.
- Moye, Z.D.; Zeng, L.; Burne, R.A. Fueling the caries process: carbohydrate metabolism and gene regulation byStreptococcus mutans. J. Oral Microbiol. 2014, 6. [Google Scholar] [CrossRef]
- A Cross-Talk between Diet and the Oral Microbiome: Balance of Nutrition on Inflammation and Immune System’s Response during Periodontitis - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/35745156/.
- Schlagenhauf, U. On the Role of Dietary Nitrate in the Maintenance of Systemic and Oral Health. Dent. J. 2022, 10, 84. [Google Scholar] [CrossRef]
- Nitrate-responsive oral microbiome modulates nitric oxide homeostasis and blood pressure in humans - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/29807159/.
- Antioxidant-Rich Natural Raw Materials in the Prevention and Treatment of Selected Oral Cavity and Periodontal Diseases - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/34829719/.
- Cigarette smoking and the oral microbiome in a large study of American adults - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/27015003/.
- Jia, Y.-J.; Liao, Y.; He, Y.-Q.; Zheng, M.-Q.; Tong, X.-T.; Xue, W.-Q.; Zhang, J.-B.; Yuan, L.-L.; Zhang, W.-L.; Jia, W.-H. Association Between Oral Microbiota and Cigarette Smoking in the Chinese Population. Front. Cell. Infect. Microbiol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Wagenknecht, D.R.; BalHaddad, A.A.; Gregory, R.L. Effects of Nicotine on Oral Microorganisms, Human Tissues, and the Interactions between Them. Curr. Oral Heal. Rep. 2018, 5, 78–87. [Google Scholar] [CrossRef]
- Beghini, F.; Renson, A.; Zolnik, C.P.; Geistlinger, L.; Usyk, M.; Moody, T.U.; Thorpe, L.; Dowd, J.B.; Burk, R.; Segata, N.; et al. Tobacco exposure associated with oral microbiota oxygen utilization in the New York City Health and Nutrition Examination Study. Ann. Epidemiology 2019, 34, 18–25. [Google Scholar] [CrossRef]
- Yussof, A.; Yoon, P.; Krkljes, C.; Schweinberg, S.; Cottrell, J.; Chu, T.; Chang, S.L. A meta-analysis of the effect of binge drinking on the oral microbiome and its relation to Alzheimer’s disease. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Alcohol-related diseases of the mouth and throat - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/16508283/.
- Enberg, N.; Alho, H.; Loimaranta, V.; Lenander-Lumikari, M. Saliva flow rate, amylase activity, and protein and electrolyte concentrations in saliva after acute alcohol consumption. Oral Surgery, Oral Med. Oral Pathol. Oral Radiol. Endodontology 2001, 92, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Maier, H.; A Born, I.; Mall, G. Effect of chronic ethanol and nicotine consumption on the function and morphology of the salivary glands.. 1988, 140–50.
- Consequences of alcohol consumption on host defence - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/10659718/.
- Szabo G, Mandrekar P, Girouard L, Catalano D. Regulation of human monocyte functions by acute ethanol treatment: decreased tumor necrosis factor-alpha, interleukin-1 beta and elevated interleukin-10, and transforming growth factor-beta production. Alcohol Clin Exp Res. 1996 Aug;20(5):900–7.
- Pepersack, T.; Fuss, M.D.; Otero, J.; Bergmann, P.; Valsamis, J.; Corvilain, J. Longitudinal study of bone metabolism after ethanol withdrawal in alcoholic patients. J. Bone Miner. Res. 1992, 7, 383–387. [Google Scholar] [CrossRef]
- Ogden, G.R.; Wight, A.J.; Rice, P. Effect of alcohol on the oral mucosa assessed by quantitative cytomorphometry. J. Oral Pathol. Med. 1999, 28, 216–220. [Google Scholar] [CrossRef]
- Fan, X.; Peters, B.A.; Jacobs, E.J.; Gapstur, S.M.; Purdue, M.P.; Freedman, N.D.; Alekseyenko, A.V.; Wu, J.; Yang, L.; Pei, Z.; et al. Drinking alcohol is associated with variation in the human oral microbiome in a large study of American adults. Microbiome 2018, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- The oral microbiome in alcohol use disorder: a longitudinal analysis during inpatient treatment - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/34880965/.
- Diurnal changes of the oral microbiome in patients with alcohol dependence - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/36579346/.
- Alcohol and tobacco consumption affects bacterial richness in oral cavity mucosa biofilms - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/25278091/.
- The Effects of Alcohol Drinking on Oral Microbiota in the Chinese Population - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/35565124/.
- Gm A. Oral Biofilm and Its Impact on Oral Health, Psychological and Social Interaction. [cited 2023 Sep 20]; Available from: https://clinmedjournals.org/articles/ijodh/international-journal-of-oral-and-dental-health-ijodh-7-127.php?jid=ijodh.
- Peng, X.; Cheng, L.; You, Y.; Tang, C.; Ren, B.; Li, Y.; Xu, X.; Zhou, X. Oral microbiota in human systematic diseases. Int. J. Oral Sci. 2022, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Patangia, D.V.; Ryan, C.A.; Dempsey, E.; Ross, R.P.; Stanton, C. Impact of antibiotics on the human microbiome and consequences for host health. Microbiologyopen 2022, 11, e1260. [Google Scholar] [CrossRef] [PubMed]
- Farah, C.; Lynch, N.; McCullough, M. Oral fungal infections: an update for the general practitioner. Aust. Dent. J. 2010, 55, 48–54. [Google Scholar] [CrossRef] [PubMed]
- The impact of horizontal gene transfer on the adaptive ability of the human oral microbiome - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/25250243/.
- DeGruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current Understanding of Dysbiosis in Disease in Human and Animal Models. Inflamm. Bowel Dis. 2016, 22, 1137–1150. [Google Scholar] [CrossRef] [PubMed]
- Composition of the adult digestive tract bacterial microbiome based on seven mouth surfaces, tonsils, throat and stool samples - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/22698087/.
- Gut microbiome and aging: Physiological and mechanistic insights - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/29951588/.
- Age-associated Impairment of the Mucus Barrier Function is Associated with Profound Changes in Microbiota and Immunity - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/30723224/.
- Neonatal oral fluid as a transmission route for bifidobacteria to the infant gut immediately after birth - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/31213639/.
- Proton pump inhibitors affect the gut microbiome - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/26657899/.
- Park SY, Hwang BO, Lim M, Ok SH, Lee SK, Chun KS, et al. Oral-Gut Microbiome Axis in Gastrointestinal Disease and Cancer. Cancers. 2021 Apr 28;13(9):2124.
- Interconnections Between the Oral and Gut Microbiomes: Reversal of Microbial Dysbiosis and the Balance Between Systemic Health and Disease - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/33652903/.
- Ectopic colonization of oral bacteria in the intestine drives TH1 cell induction and inflammation - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/29051379/.
- Kitamoto, S.; Nagao-Kitamoto, H.; Hein, R.; Schmidt, T.M.; Kamada, N. The Bacterial Connection between the Oral Cavity and the Gut Diseases. J. Dent. Res. 2020, 99, 1021–1029. [Google Scholar] [CrossRef]
- Walker, M.Y.; Pratap, S.; Southerland, J.H.; Farmer-Dixon, C.M.; Lakshmyya, K.; Gangula, P.R. Role of oral and gut microbiome in nitric oxide-mediated colon motility. Nitric Oxide 2018, 73, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Extensive transmission of microbes along the gastrointestinal tract - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/30747106/.
- Oral Biofilms: Pathogens, Matrix, and Polymicrobial Interactions in Microenvironments - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/29097091/.
- Identification of bistable populations of Porphyromonas gingivalis that differ in epithelial cell invasion - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/20576685/.
- Tanner, A.; Kressirer, C.; Rothmiller, S.; Johansson, I.; Chalmers, N. The Caries Microbiome: Implications for Reversing Dysbiosis. Adv. Dent. Res. 2018, 29, 78–85. [Google Scholar] [CrossRef]
- Selwitz RH, Ismail AI, Pitts NB. Dental caries. Lancet Lond Engl. 2007 Jan 6;369(9555):51–9.
- Caries ecology revisited: microbial dynamics and the caries process - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/18832827/.
- The oral microbiota: dynamic communities and host interactions - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/30301974/.
- Seminario, A.; Broukal, Z.; Ivancaková, R. Mutans streptococci and the development of dental plaque. 2005, 106, 349–58. [Google Scholar]
- Madléna, M.; Dombi, C.; Gintner, Z.; Bánóczy, J. Effect of amine fluoride/stannous fluoride toothpaste and mouthrinse on dental plaque accumulation and gingival health. Oral Dis. 2004, 10, 294–297. [Google Scholar] [CrossRef]
- Microbial Etiology and Prevention of Dental Caries: Exploiting Natural Products to Inhibit Cariogenic Biofilms - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/32674310/.
- Structure-Based Discovery of Small Molecule Inhibitors of Cariogenic Virulence - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/28729722/.
- Simón-Soro, A.; Guillen-Navarro, M.; Mira, A. Metatranscriptomics reveals overall active bacterial composition in caries lesions. J. Oral Microbiol. 2014, 6, 25443–25443. [Google Scholar] [CrossRef] [PubMed]
- Solving the etiology of dental caries - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/25435135/.
- Beyond Streptococcus mutans: dental caries onset linked to multiple species by 16S rRNA community analysis - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/23091642/.
- Association between Bifidobacteriaceae and the clinical severity of root caries lesions - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/19121067/.
- Badet, C.; Thebaud, N.B. Ecology of Lactobacilli in the Oral Cavity: A Review of Literature. Open Microbiol. J. 2008, 2, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Microbial geography of the oral cavity - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/23674263/.
- Intan Suhana MA, Farha A, Hassan BM. Inflammation of the Gums. Malays Fam Physician Off J Acad Fam Physicians Malays. 2020;15(1):71–3.
- Rathee M, Jain P. Gingivitis. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2024 [cited 2024 May 3]. Available from: http://www.ncbi.nlm.nih.gov/books/NBK557422/.
- Oral microbiomes: more and more importance in oral cavity and whole body - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/29736705/.
- Lu, M.; Xuan, S.; Wang, Z. Oral microbiota: A new view of body health. Food Sci. Hum. Wellness 2019, 8, 8–15. [Google Scholar] [CrossRef]
- Moore, W.E.C.; Moore, L.V.H. The bacteria of periodontal diseases. Periodontol. 2000 1994, 5, 66–77. [Google Scholar] [CrossRef] [PubMed]
- The role of the microbiota in periodontal disease - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/32385883/.
- Kistler, J.O.; Booth, V.; Bradshaw, D.J.; Wade, W.G. Bacterial Community Development in Experimental Gingivitis. PLOS ONE 2013, 8, e71227. [Google Scholar] [CrossRef] [PubMed]
- Clinical, Immune, and Microbiome Traits of Gingivitis and Peri-implant Mucositis - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/28033066/.
- Abdulkareem, A.A.; Al-Taweel, F.B.; Al-Sharqi, A.J.; Gul, S.S.; Sha, A.; Chapple, I.L. Current concepts in the pathogenesis of periodontitis: from symbiosis to dysbiosis. J. Oral Microbiol. 2023, 15, 2197779. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Darveau, R.P.; Curtis, M.A. The keystone-pathogen hypothesis. Nat. Rev. Microbiol. 2012, 10, 717–725. [Google Scholar] [CrossRef] [PubMed]
- The Role of Oral Pathobionts in Dysbiosis during Periodontitis Development - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/24646638/.
- Kapoor, U.; Sharma, G.; Juneja, M.; Nagpal, A. Halitosis: Current concepts on etiology, diagnosis and management. Eur. J. Dent. 2016, 10, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Halitosis: a review of associated factors and therapeutic approach - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/19838550/.
- Hampelska, K.; Jaworska, M.M.; Babalska, Z.Ł.; Karpiński, T.M. The Role of Oral Microbiota in Intra-Oral Halitosis. J. Clin. Med. 2020, 9, 2484. [Google Scholar] [CrossRef]
- Microbial basis of oral malodor development in humans - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/24101743/.
- Nakano, Y.; Yoshimura, M.; Koga, T. Correlation between oral malodor and periodontal bacteria. Microbes Infect. 2002, 4, 679–683. [Google Scholar] [CrossRef]
- The relationship between oral malodor, gingivitis, and periodontitis. A review - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/10368052/.
- Porter SR, Scully C. Oral malodour (halitosis). BMJ. 2006 Sep 23;333(7569):632–5.
- Persson, S.; Edlund, M.; Claesson, R.; Carlsson, J. The formation of hydrogen sulfide and methyl mercaptan by oral bacteria. Oral Microbiol. Immunol. 1990, 5, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, T.; Suzuki, N.; Nakano, Y.; Yasui, M.; Yoneda, M.; Shimazaki, Y.; Hirofuji, T.; Yamashita, Y. Discrimination of the oral microbiota associated with high hydrogen sulfide and methyl mercaptan production. Sci. Rep. 2012, 2, 215. [Google Scholar] [CrossRef] [PubMed]
- Washio, J.; Sato, T.; Koseki, T.; Takahashi, N. Hydrogen sulfide-producing bacteria in tongue biofilm and their relationship with oral malodour. J. Med Microbiol. 2005, 54, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Solemdal, K.; Sandvik, L.; Willumsen, T.; Mowe, M.; Hummel, T. The Impact of Oral Health on Taste Ability in Acutely Hospitalized Elderly. PLOS ONE 2012, 7, e36557. [Google Scholar] [CrossRef] [PubMed]
- A Finkelstein, J.; Schiffman, S.S. Workshop on Taste and Smell in the Elderly: An Overview. Physiol. Behav. 1999, 66, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, B. Receptors and transduction in taste. Nature 2001, 413, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Spielman, A. Interaction of Saliva and Taste. J. Dent. Res. 1990, 69, 838–843. [Google Scholar] [CrossRef]
- Matsuo, R. Role of Saliva in the Maintenance of Taste Sensitivity. Crit. Rev. Oral Biol. Med. 2000, 11, 216–229. [Google Scholar] [CrossRef]
- Nederfors T, Isaksson R, Mörnstad H, Dahlöf C. Prevalence of perceived symptoms of dry mouth in an adult Swedish population--relation to age, sex and pharmacotherapy. Community Dent Oral Epidemiol. 1997 Jun;25(3):211–6.
- Impact of primary Sjogren’s syndrome on smell and taste: effect on quality of life - PubMed [Internet]. [cited 2024 May 3]. Available from: https://pubmed.ncbi.nlm.nih.gov/19752179/.
- Weifenbach, J.M.; Schwartz, L.K.; Atkinson, J.C.; Fox, P.C. Taste performance in Sjogren's syndrome. Physiol. Behav. 1995, 57, 89–96. [Google Scholar] [CrossRef]
- Aravindhan, R.; Vidyalakshmi, S.; Kumar, M.; Satheesh, C.; Balasubramanium, A.; Prasad, V. Burning mouth syndrome: A review on its diagnostic and therapeutic approach. J. Pharm. Bioallied Sci. 2014, 6, S21–S25. [Google Scholar] [CrossRef]
- Lee, B.-M.; Park, J.W.; Jo, J.H.; Oh, B.; Chung, G. Comparative analysis of the oral microbiome of burning mouth syndrome patients. J. Oral Microbiol. 2022, 14, 2052632. [Google Scholar] [CrossRef] [PubMed]
- Singh A, Verma R, Murari A, Agrawal A. Oral candidiasis: An overview. J Oral Maxillofac Pathol JOMFP. 2014 Sep;18(Suppl 1):S81-85.
- Rajendra Santosh AB, Muddana K, Bakki SR. Fungal Infections of Oral Cavity: Diagnosis, Management, and Association with COVID-19. SN Compr Clin Med. 2021;3(6):1373–84.
- Candidiasis - StatPearls - NCBI Bookshelf [Internet]. [cited 2023 Sep 20]. Available from: https://www.ncbi.nlm.nih.gov/books/NBK560624/.
- Oral thrush: Overview - InformedHealth.org - NCBI Bookshelf [Internet]. [cited 2023 Sep 20]. Available from: https://www.ncbi.nlm.nih.gov/books/NBK367586/.
- Cotter, G.; Kavanagh, K. Adherence mechanisms of Candida albicans. 2000, 57, 241–9. [Google Scholar] [PubMed]
- The ALS gene family of Candida albicans - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/11286882/.
- Nikou, S.-A.; Kichik, N.; Brown, R.; Ponde, N.O.; Ho, J.; Naglik, J.R.; Richardson, J.P. Candida albicans Interactions with Mucosal Surfaces during Health and Disease. Pathogens 2019, 8, 53. [Google Scholar] [CrossRef] [PubMed]
- Discovering the secrets of the Candida albicans agglutinin-like sequence (ALS) gene family--a sticky pursuit - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/17852717/.
- Naglik JR, Fostira F, Ruprai J, Staab JF, Challacombe SJ, Sundstrom P. Candida albicans HWP1 gene expression and host antibody responses in colonization and disease. J Med Microbiol. 2006 Oct;55(Pt 10):1323–7.
- Sundstrom P, Balish E, Allen CM. Essential role of the Candida albicans transglutaminase substrate, hyphal wall protein 1, in lethal oroesophageal candidiasis in immunodeficient mice. J Infect Dis. 2002 Feb 15;185(4):521–30.
- Vila, T.; Sultan, A.S.; Montelongo-Jauregui, D.; Jabra-Rizk, M.A. Oral Candidiasis: A Disease of Opportunity. J. Fungi 2020, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- Silverman, R.J.; Nobbs, A.H.; Vickerman, M.M.; Barbour, M.E.; Jenkinson, H.F. Interaction of Candida albicans Cell Wall Als3 Protein with Streptococcus gordonii SspB Adhesin Promotes Development of Mixed-Species Communities. Infect. Immun. 2010, 78, 4644–4652. [Google Scholar] [CrossRef] [PubMed]
- Lewis M, a. O, Williams DW. Diagnosis and management of oral candidosis. Br Dent J. 2017 Nov 10;223(9):675–81.
- Candida albicans Hyphae: From Growth Initiation to Invasion - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/29371503/.
- Candida albicans secreted aspartyl proteinases in virulence and pathogenesis - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/12966142/.
- Candida albicans pathogenicity mechanisms - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/23302789/.
- Höfs, S.; Mogavero, S.; Hube, B. Interaction of Candida albicans with host cells: virulence factors, host defense, escape strategies, and the microbiota. J. Microbiol. 2016, 54, 149–169. [Google Scholar] [CrossRef] [PubMed]
- Candidalysin Is Required for Neutrophil Recruitment and Virulence During Systemic Candida albicans Infection - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/31401652/.
- andidalysin is a fungal peptide toxin critical for mucosal infection - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/27027296/.
- Phan, Q.T.; Myers, C.L.; Fu, Y.; Sheppard, D.C.; Yeaman, M.R.; Welch, W.H.; Ibrahim, A.S.; Edwards, J.E., Jr.; Filler, S.G. Als3 Is a Candida albicans Invasin That Binds to Cadherins and Induces Endocytosis by Host Cells. PLoS Biol. 2007, 5, e64. [Google Scholar] [CrossRef] [PubMed]
- Bacteria from diverse habitats colonize and compete in the mouse gut - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/25284151/.
- Peeters, M.; Geypens, B.; Claus, D.; Nevens, H.; Ghoos, Y.; Verbeke, G.; Baert, F.; Vermeire, S.; Vlietinck, R.; Rutgeerts, P. Clustering of increased small intestinal permeability in families with Crohn's disease. Gastroenterology 1997, 113, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Altered tight junction structure contributes to the impaired epithelial barrier function in ulcerative colitis - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/9922310/.
- Fusobacterium nucleatum - symbiont, opportunist and oncobacterium - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/30546113/.
- Atarashi K, Suda W, Luo C, Kawaguchi T, Motoo I, Narushima S, et al. Ectopic colonization of oral bacteria in the intestine drives TH1 cell induction and inflammation. Science. 2017 Oct 20;358(6361):359–65.
- Porphyromonas gingivalis indirectly elicits intestinal inflammation by altering the gut microbiota and disrupting epithelial barrier function through IL9-producing CD4+ T cells - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/34958712/.
- The subgingival microbiome in health and periodontitis and its relationship with community biomass and inflammation - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/23303375/.
- Li, Y.; Zhu, M.; Liu, Y.; Luo, B.; Cui, J.; Huang, L.; Chen, K.; Liu, Y. The oral microbiota and cardiometabolic health: A comprehensive review and emerging insights. Front. Immunol. 2022, 13, 1010368. [Google Scholar] [CrossRef]
- Lucchese, A. Streptococcus mutans antigen I/II and autoimmunity in cardiovascular diseases. Autoimmun. Rev. 2017, 16, 456–460. [Google Scholar] [CrossRef]
- Xie, M.; Tang, Q.; Nie, J.; Zhang, C.; Zhou, X.; Yu, S.; Sun, J.; Cheng, X.; Dong, N.; Hu, Y.; et al. BMAL1-Downregulation Aggravates Porphyromonas Gingivalis -Induced Atherosclerosis by Encouraging Oxidative Stress. Circ. Res. 2020, 126, E15–E29. [Google Scholar] [CrossRef]
- Stinson, M.W.; Alder, S.; Kumar, S. Invasion and Killing of Human Endothelial Cells by Viridans Group Streptococci. Infect. Immun. 2003, 71, 2365–2372. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Wong, L.-Y.; Jia, L.T.; Kuklenyik, Z.; Calafat, A.M. Trends in Exposure to Polyfluoroalkyl Chemicals in the U.S. Population: 1999−2008. Environ. Sci. Technol. 2011, 45, 8037–8045. [Google Scholar] [CrossRef] [PubMed]
- Ramos, H.C.; Rumbo, M.; Sirard, J.-C. Bacterial flagellins: mediators of pathogenicity and host immune responses in mucosa. Trends Microbiol. 2004, 12, 509–517. [Google Scholar] [CrossRef]
- Zindel, J.; Kubes, P. DAMPs, PAMPs, and LAMPs in Immunity and Sterile Inflammation. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 493–518. [Google Scholar] [CrossRef]
- Aggregatibacter actinomycetemcomitans leukotoxin (LtxA; Leukothera) induces cofilin dephosphorylation and actin depolymerization during killing of malignant monocytes - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/25169107/.
- Aarabi, G.; Heydecke, G.; Seedorf, U. Roles of Oral Infections in the Pathomechanism of Atherosclerosis. Int. J. Mol. Sci. 2018, 19, 1978. [Google Scholar] [CrossRef] [PubMed]
- Linkages between oral commensal bacteria and atherosclerotic plaques in coronary artery disease patients - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/28649401/.
- Oral Microbiome in Relation to Periodontitis Severity and Systemic Inflammation - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/34070915/.
- Lourenço, T.G.B.; Heller, D.; Silva-Boghossian, C.M.; Cotton, S.L.; Paster, B.J.; Colombo, A.P.V. Microbial signature profiles of periodontally healthy and diseased patients. J. Clin. Periodontol. 2014, 41, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, R.I. Cells in Stress: Transcriptional Activation of Heat Shock Genes. Science 1993, 259, 1409–1410. [Google Scholar] [CrossRef]
- Goulhen, F.; Grenier, D.; Mayrand, D. Oral Microbial Heat-shock Proteins and Their Potential Contributions to Infections. Crit. Rev. Oral Biol. Med. 2003, 14, 399–412. [Google Scholar] [CrossRef]
- Xiao, E.; Mattos, M.; Vieira, G.H.A.; Chen, S.; Corrêa, J.D.; Wu, Y.; Albiero, M.L.; Bittinger, K.; Graves, D.T. Diabetes Enhances IL-17 Expression and Alters the Oral Microbiome to Increase Its Pathogenicity. Cell Host Microbe 2017, 22, 120–128. [Google Scholar] [CrossRef]
- Mashimo, P.A.; Yamamoto, Y.; Slots, J.; Park, B.H.; Genco, R.J. The Periodontal Microflora of Juvenile Diabetics: Culture, Immunofluorescence, and Serum Antibody Studies. J. Periodontol. 1983, 54, 420–430. [Google Scholar] [CrossRef]
- Diabetes and periodontal disease: a case-control study - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/15857077/.
- da Cruz, G.A.; de Toledo, S.; Sallum, E.A.; Sallum, A.W.; Ambrosano, G.M.B.; Sardi, J.d.C.O.; da Cruz, S.E.B.; Gonçalves, R.B. Clinical and Laboratory Evaluations of Non-Surgical Periodontal Treatment in Subjects With Diabetes Mellitus. J. Periodontol. 2008, 79, 1150–1157. [Google Scholar] [CrossRef]
- The Oral Microbiota Is Modified by Systemic Diseases - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/30359170/.
- Wu, Y.-Y.; Xiao, E.; Graves, D.T. Diabetes mellitus related bone metabolism and periodontal disease. Int. J. Oral Sci. 2015, 7, 63–72. [Google Scholar] [CrossRef]
- Priming of neutrophil oxidative burst in diabetes requires preassembly of the NADPH oxidase - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/18390927/.
- Benahmed, A.G.; Gasmi, A.; Doşa, A.; Chirumbolo, S.; Mujawdiya, P.K.; Aaseth, J.; Dadar, M.; Bjørklund, G. Association between the gut and oral microbiome with obesity. Anaerobe 2020, 70, 102248. [Google Scholar] [CrossRef]
- Schamarek, I.; Anders, L.; Chakaroun, R.M.; Kovacs, P.; Rohde-Zimmermann, K. The role of the oral microbiome in obesity and metabolic disease: potential systemic implications and effects on taste perception. Nutr. J. 2023, 22, 1–13. [Google Scholar] [CrossRef]
- Gut Microbiota as a Trigger for Metabolic Inflammation in Obesity and Type 2 Diabetes - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/33178196/.
- Goodson JM, Groppo D, Halem S, Carpino E. Is obesity an oral bacterial disease? J Dent Res. 2009 Jun;88(6):519–23.
- Metabolic endotoxemia initiates obesity and insulin resistance - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/17456850/.
- Salivary inflammatory markers and microbiome in normoglycemic lean and obese children compared to obese children with type 2 diabetes - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/28253297/.
- Profiling the Oral Microbiome and Plasma Biochemistry of Obese Hyperglycemic Subjects in Qatar - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/31816998/.
- Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/28532414/.
- Mathur R, Barlow GM. Obesity and the microbiome. Expert Rev Gastroenterol Hepatol. 2015;9(8):1087–99.
- Experimental periodontitis induces gene expression of proinflammatory cytokines in liver and white adipose tissues in obesity - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/20367095/.
- Evidence and Therapeutic Perspectives in the Relationship between the Oral Microbiome and Alzheimer’s Disease: A Systematic Review - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/34769677/.
- Molecular and immunological evidence of oral Treponema in the human brain and their association with Alzheimer’s disease - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/11929559/.
- Determining the presence of periodontopathic virulence factors in short-term postmortem Alzheimer’s disease brain tissue - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/23666172/.
- Miklossy, J. Emerging roles of pathogens in Alzheimer disease. Expert Rev. Mol. Med. 2011, 13, e30. [Google Scholar] [CrossRef]
- Miklossy, J. Alzheimer’s disease - a neurospirochetosis. Analysis of the evidence following Koch’s and Hill’s criteria. J Neuroinflammation. 2011 Aug 4;8:90.
- Sureda A, Daglia M, Argüelles Castilla S, Sanadgol N, Fazel Nabavi S, Khan H, et al. Oral microbiota and Alzheimer’s disease: Do all roads lead to Rome? Pharmacol Res. 2020 Jan;151:104582.
- Microbial involvement in Alzheimer disease development and progression - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/32709243/.
- Amyloid beta is an early responder cytokine and immunopeptide of the innate immune system - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/33163614/.
- Liu S, Butler CA, Ayton S, Reynolds EC, Dashper SG. Porphyromonas gingivalis and the pathogenesis of Alzheimer’s disease. Crit Rev Microbiol. 2023 Jan 4;1–11.
- Soscia SJ, Kirby JE, Washicosky KJ, Tucker SM, Ingelsson M, Hyman B, et al. The Alzheimer’s disease-associated amyloid beta-protein is an antimicrobial peptide. PloS One. 2010 Mar 3;5(3):e9505.
- von Campenhausen, S.; Bornschein, B.; Wick, R.; Bötzel, K.; Sampaio, C.; Poewe, W.; Oertel, W.; Siebert, U.; Berger, K.; Dodel, R. Prevalence and incidence of Parkinson's disease in Europe. Eur. Neuropsychopharmacol. 2005, 15, 473–490. [Google Scholar] [CrossRef] [PubMed]
- Weintraub D, Comella CL, Horn S. Parkinson’s disease--Part 1: Pathophysiology, symptoms, burden, diagnosis, and assessment. Am J Manag Care. 2008 Mar;14(2 Suppl):S40-48.
- Parkinson disease - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/28332488/.
- Parkinson’s Disease: A Systemic Inflammatory Disease Accompanied by Bacterial Inflammagens - PMC [Internet]. [cited 2024 Apr 12]. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6718721/.
- Chen, C.-K.; Wu, Y.-T.; Chang, Y.-C. Periodontal inflammatory disease is associated with the risk of Parkinson’s disease: a population-based retrospective matched-cohort study. PeerJ 2017, 5, e3647. [Google Scholar] [CrossRef] [PubMed]
- Jeong, E.; Park, J.-B.; Park, Y.-G. Evaluation of the association between periodontitis and risk of Parkinson’s disease: a nationwide retrospective cohort study. Sci. Rep. 2021, 11, 1–8. [Google Scholar] [CrossRef]
- Oral P. gingivalis impairs gut permeability and mediates immune responses associated with neurodegeneration in LRRK2 R1441G mice - PMC [Internet]. [cited 2024 Apr 12]. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7677837/.
- The Potential of LPS-Binding Protein to Reverse Amyloid Formation in Plasma Fibrin of Individuals With Alzheimer-Type Dementia - PMC [Internet]. [cited 2024 Apr 12]. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6113936/.
- The oral microbiome in autoimmune diseases: friend or foe? - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/36949458/.
- Exploring the Oral Microbiome in Rheumatic Diseases, State of Art and Future Prospective in Personalized Medicine with an AI Approach - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/34209167/.
- Hypothesis: the humoral immune response to oral bacteria provides a stimulus for the development of rheumatoid arthritis - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/16245073/.
- Guo, Q.; Wang, Y.; Xu, D.; Nossent, J.; Pavlos, N.J.; Xu, J. Rheumatoid arthritis: pathological mechanisms and modern pharmacologic therapies. Bone Res. 2018, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Microbial complexes in subgingival plaque - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/9495612/.
- The possible role of oral microbiome in autoimmunity - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/33898698/.
- Marwaha AK, Leung NJ, McMurchy AN, Levings MK. TH17 Cells in Autoimmunity and Immunodeficiency: Protective or Pathogenic? Front Immunol. 2012;3:129.
- Identification of a cytokine network sustaining neutrophil and Th17 activation in untreated early rheumatoid arthritis - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/20961415/.
- Periodontal disease and influence of periodontal treatment on disease activity in patients with rheumatoid arthritis and spondyloarthritis - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/31701185/.
- Konig, M.F.; Abusleme, L.; Reinholdt, J.; Palmer, R.J.; Teles, R.P.; Sampson, K.; Rosen, A.; Nigrovic, P.A.; Sokolove, J.; Giles, J.T.; et al. Aggregatibacter actinomycetemcomitans –induced hypercitrullination links periodontal infection to autoimmunity in rheumatoid arthritis. Sci. Transl. Med. 2016, 8, 369ra176–369ra176. [Google Scholar] [CrossRef] [PubMed]
- Rooney, C.M.; Mankia, K.; Emery, P. The Role of the Microbiome in Driving RA-Related Autoimmunity. Front. Cell Dev. Biol. 2020, 8, 538130. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Cheng, Z.; Zhu, F.; Bi, C.; Shi, Q.; Chen, X. The Oral Microbiome and Its Role in Systemic Autoimmune Diseases: A Systematic Review of Big Data Analysis. Front. Big Data 2022, 5, 927520. [Google Scholar] [CrossRef] [PubMed]
- Host-Microbial Interactions in Systemic Lupus Erythematosus and Periodontitis - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/31781106/.
- Liu, F.; Ren, T.; Li, X.; Zhai, Q.; Xu, X.; Zhang, N.; Jiang, P.; Niu, Y.; Lv, L.; Shi, G.; et al. Distinct Microbiomes of Gut and Saliva in Patients With Systemic Lupus Erythematous and Clinical Associations. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Salivary dysbiosis and the clinical spectrum in anti-Ro positive mothers of children with neonatal lupus - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/31677965/.
- Interleukin-17 in systemic lupus erythematosus: A comprehensive review - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/25894789/.
- IL-33 induces Th17-mediated airway inflammation via mast cells in ovalbumin-challenged mice - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/22180658/.
- The paradigm of IL-6: from basic science to medicine - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/12110143/.
- Yamamura K, Izumi D, Kandimalla R, Sonohara F, Baba Y, Yoshida N, et al. Intratumoral Fusobacterium Nucleatum Levels Predict Therapeutic Response to Neoadjuvant Chemotherapy in Esophageal Squamous Cell Carcinoma. Clin Cancer Res Off J Am Assoc Cancer Res. 2019 Oct 15;25(20):6170–9.
- Sun, J.; Tang, Q.; Yu, S.; Xie, M.; Xie, Y.; Chen, G.; Chen, L. Role of the oral microbiota in cancer evolution and progression. Cancer Med. 2020, 9, 6306–6321. [Google Scholar] [CrossRef]
- Sarao, L.K.; Arora, M. Probiotics, prebiotics, and microencapsulation: A review. Crit. Rev. Food Sci. Nutr. 2015, 57, 344–371. [Google Scholar] [CrossRef] [PubMed]
- A concerted probiotic activity to inhibit periodontitis-associated bacteria - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/33667279/.
- Cosseau, C.; Devine, D.A.; Dullaghan, E.; Gardy, J.L.; Chikatamarla, A.; Gellatly, S.; Yu, L.L.; Pistolic, J.; Falsafi, R.; Tagg, J.; et al. The Commensal Streptococcus salivarius K12 Downregulates the Innate Immune Responses of Human Epithelial Cells and Promotes Host-Microbe Homeostasis. Infect. Immun. 2008, 76, 4163–4175. [Google Scholar] [CrossRef]
- Streptococcus salivarius inhibits immune activation by periodontal disease pathogens - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/33962608/.
- Anti-inflammatory properties of Streptococcus salivarius, a commensal bacterium of the oral cavity and digestive tract - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/24271166/.
- Keller MK, Bardow A, Jensdottir T, Lykkeaa J, Twetman S. Effect of chewing gums containing the probiotic bacterium Lactobacillus reuteri on oral malodour. Acta Odontol Scand. 2012 May;70(3):246–50.
- Keller MK, Hasslöf P, Stecksén-Blicks C, Twetman S. Co-aggregation and growth inhibition of probiotic lactobacilli and clinical isolates of mutans streptococci: an in vitro study. Acta Odontol Scand. 2011 Sep;69(5):263–8.
- Invernici, M.M.; Salvador, S.L.; Silva, P.H.F.; Soares, M.S.M.; Casarin, R.; Palioto, D.B.; Souza, S.L.S.; Taba, M., Jr.; Novaes, A.B., Jr.; Furlaneto, F.A.C.; et al. Effects of Bifidobacterium probiotic on the treatment of chronic periodontitis: A randomized clinical trial. J. Clin. Periodontol. 2018, 45, 1198–1210. [Google Scholar] [CrossRef] [PubMed]
- Lactobacillus plantarum Disrupts S. mutans-C. albicans Cross-Kingdom Biofilms - PubMed [Internet]. [cited 2023 Sep 20]. Available from: https://pubmed.ncbi.nlm.nih.gov/35392605/.
- Biochemistry, Peptide - StatPearls - NCBI Bookshelf [Internet]. [cited 2024 May 2]. Available from: https://www.ncbi.nlm.nih.gov/books/NBK562260/.
- Frontiers | Bioactive Synthetic Peptides for Oral Tissues Regeneration [Internet]. [cited 2024 May 2]. Available from: https://www.frontiersin.org/articles/10.3389/fmats.2021.655495/full.
- The antimicrobial peptides secreted by the chromaffin cells of the adrenal medulla link the neuroendocrine and immune systems: From basic to clinical studies - PMC [Internet]. [cited 2024 May 2]. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9452953/.
- Alkilzy, M.; Santamaria, R.; Schmoeckel, J.; Splieth, C. Treatment of Carious Lesions Using Self-Assembling Peptides. Adv. Dent. Res. 2018, 29, 42–47. [Google Scholar] [CrossRef]
- The antimicrobial peptides and their potential clinical applications - PMC [Internet]. [cited 2024 May 2]. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6684887/.
- Cathelicidins, multifunctional peptides of the innate immunity - PubMed [Internet]. [cited 2024 May 2]. Available from: https://pubmed.ncbi.nlm.nih.gov/12960280/.
- Khurshid, Z.; Najeeb, S.; Mali, M.; Moin, S.F.; Raza, S.Q.; Zohaib, S.; Sefat, F.; Zafar, M.S. Histatin peptides: Pharmacological functions and their applications in dentistry. Saudi Pharm. J. 2016, 25, 25–31. [Google Scholar] [CrossRef]
- Oppenheim, F.G.; Xu, T.; McMILLIAN, F.M.; Levitz, S.M.; Diamond, R.D.; Offner, G.D.; Troxler, R.F. Histatins, a novel family of histidine-rich proteins in human parotid secretion. Isolation, characterization, primary structure, and fungistatic effects on Candida albicans. J. Biol. Chem. 1988, 263, 7472–7477. [Google Scholar] [CrossRef] [PubMed]
- Anticandidal activity of major human salivary histatins. - PMC [Internet]. [cited 2024 May 2]. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC258054/.
- Wang, Q.-Q.; Wang, S.; Zhao, T.; Li, Y.; Yang, J.; Liu, Y.; Zhang, H.; Miao, L.; Sun, W. Biomimetic oligopeptide formed enamel-like tissue and dentin tubule occlusion via mineralization for dentin hypersensitivity treatment. J. Appl. Biomater. Funct. Mater. 2021, 19. [Google Scholar] [CrossRef] [PubMed]
- Physico-chemical properties of crystal surfaces in matrix–mineral interactions during mammalian biomineralisation - ScienceDirect [Internet]. [cited 2024 May 3]. Available from: https://www.sciencedirect.com/science/article/abs/pii/S1359029402000171?via%3Dihub.
- Effects of xylitol chewing gum and candies on the accumulation of dental plaque: a systematic review - PubMed [Internet]. [cited 2024 Jan 2]. Available from: https://pubmed.ncbi.nlm.nih.gov/34677696/.
- Nayak, P.A.; Nayak, U.A.; Khandelwal, V. The effect of xylitol on dental caries and oral flora. Clin. Cosmet. Investig. Dent. 2014, 6, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Drake, D.; Villhauer, A.L. An in vitro comparative study determining bactericidal activity of stabilized chlorine dioxide and other oral rinses. . 2011, 22, 1–5. [Google Scholar] [PubMed]
- Shinada, K.; Ueno, M.; Konishi, C.; Takehara, S.; Yokoyama, S.; Zaitsu, T.; Ohnuki, M.; Wright, F.A.C.; Kawaguchi, Y. Effects of a mouthwash with chlorine dioxide on oral malodor and salivary bacteria: a randomized placebo-controlled 7-day trial. Trials 2010, 11, 14–14. [Google Scholar] [CrossRef]
- C V, S CH, E H, A AA. A Preliminary Investigation on the Antimicrobial Activity of Listerine®, Its Components, and of Mixtures Thereof. Phytother Res PTR [Internet]. 2015 Oct [cited 2024 Jan 8];29(10). Available from: https://pubmed.ncbi.nlm.nih.gov/26104602/.
- Al-Ani, E.; Heaselgrave, W. The Investigation of Thymol Formulations Containing Poloxamer 407 and Hydroxypropyl Methylcellulose to Inhibit Candida Biofilm Formation and Demonstrate Improved Bio-Compatibility. Pharmaceuticals 2022, 15, 71. [Google Scholar] [CrossRef]
- Wińska K, Mączka W, Łyczko J, Grabarczyk M, Czubaszek A, Szumny A. Essential Oils as Antimicrobial Agents—Myth or Real Alternative? Molecules. 2019 Jun 5;24(11):2130.
- The use of mouthwash containing essential oils (LISTERINE®) to improve oral health: A systematic review - PubMed [Internet]. [cited 2024 Jan 2]. Available from: https://pubmed.ncbi.nlm.nih.gov/30166864/.
- Current concept on the anticaries fluoride mechanism of the action - PubMed [Internet]. [cited 2024 Jan 8]. Available from: https://pubmed.ncbi.nlm.nih.gov/11811302/.
- Vranić, E.; Lačević, A.; Mehmedagić, A.; Uzunović, A. Formulation ingredients for toothpastes and mouthwashes. Bosn. J. Basic Med Sci. 2004, 4, 51–58. [Google Scholar] [CrossRef]
- Zhang, J.; Sardana, D.; Li, K.Y.; Leung, K.C.M.; Lo, E.C.M. Topical Fluoride to Prevent Root Caries: Systematic Review with Network Meta-analysis. J. Dent. Res. 2020, 99, 506–513. [Google Scholar] [CrossRef]
- O’hagan-Wong, K.; Enax, J.; Meyer, F.; Ganss, B. The use of hydroxyapatite toothpaste to prevent dental caries. Odontology 2021, 110, 223–230. [Google Scholar] [CrossRef]
- Mao, X.; Auer, D.L.; Buchalla, W.; Hiller, K.-A.; Maisch, T.; Hellwig, E.; Al-Ahmad, A.; Cieplik, F. Cetylpyridinium Chloride: Mechanism of Action, Antimicrobial Efficacy in Biofilms, and Potential Risks of Resistance. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Dagli, N.; Dagli, R.; Mahmoud, R.S.; Baroudi, K. Essential oils, their therapeutic properties, and implication in dentistry: A review. J. Int. Soc. Prev. Community Dent. 2015, 5, 335–40. [Google Scholar] [CrossRef] [PubMed]
- Shanbhag, V.K.L. Oil pulling for maintaining oral hygiene – A review. J. Tradit. Complement. Med. 2016, 7, 106–109. [Google Scholar] [CrossRef] [PubMed]
- The effect of oil pulling with coconut oil to improve dental hygiene and oral health: A systematic review - PMC [Internet]. [cited 2024 Jan 8]. Available from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7475120/.
Figure 1.
Oral-Gut Pathways to IBD and Systemic Diseases: Oral microbes such as Porphyromonas gingivalis can take two distinct routes to enter the body. The hematogenous route (shown on the left), often associated with dental problems like tooth decay, periodontitis, gingivitis, oral thrush and halitosis, which can further lead to systemic conditions such as CVD, neurological disorders, autoimmune diseases, diabetes, IBD, and cancer. On the other hand, the enteral route (shown in the right) allows these oral microbes to travel from the stomach to the intestines. P. gingivalis, which possesses resistance to antibiotics and can survive stomach acid, moves from the stomach into the gut. Changes in gastric acidity can alter the gut microbiota, making it resemble the oral microbiome. Upon entering the gastrointestinal tract, P. gingivalis disrupts the intestinal barrier, compromising gut integrity. This disruption, along with changes in the microbiome, initiates inflammation, typically occurring in the ileum. Within this inflammatory response, specific immune cells known as IL9+ CD4+ lamina propria T cells become active and produce IL-9, a cytokine that fuels immune responses and inflammation. While inflammation serves as a defense against invaders, excessive or chronic inflammation can lead to conditions like IBD. Abbreviations: CVD: cardiovascular disease, IL-9: Interleukin-9, IL9+ CD4+ lamina propria T cells: Immune cells producing IL-9, IBD: Inflammatory Bowel Disease.
Figure 1.
Oral-Gut Pathways to IBD and Systemic Diseases: Oral microbes such as Porphyromonas gingivalis can take two distinct routes to enter the body. The hematogenous route (shown on the left), often associated with dental problems like tooth decay, periodontitis, gingivitis, oral thrush and halitosis, which can further lead to systemic conditions such as CVD, neurological disorders, autoimmune diseases, diabetes, IBD, and cancer. On the other hand, the enteral route (shown in the right) allows these oral microbes to travel from the stomach to the intestines. P. gingivalis, which possesses resistance to antibiotics and can survive stomach acid, moves from the stomach into the gut. Changes in gastric acidity can alter the gut microbiota, making it resemble the oral microbiome. Upon entering the gastrointestinal tract, P. gingivalis disrupts the intestinal barrier, compromising gut integrity. This disruption, along with changes in the microbiome, initiates inflammation, typically occurring in the ileum. Within this inflammatory response, specific immune cells known as IL9+ CD4+ lamina propria T cells become active and produce IL-9, a cytokine that fuels immune responses and inflammation. While inflammation serves as a defense against invaders, excessive or chronic inflammation can lead to conditions like IBD. Abbreviations: CVD: cardiovascular disease, IL-9: Interleukin-9, IL9+ CD4+ lamina propria T cells: Immune cells producing IL-9, IBD: Inflammatory Bowel Disease.
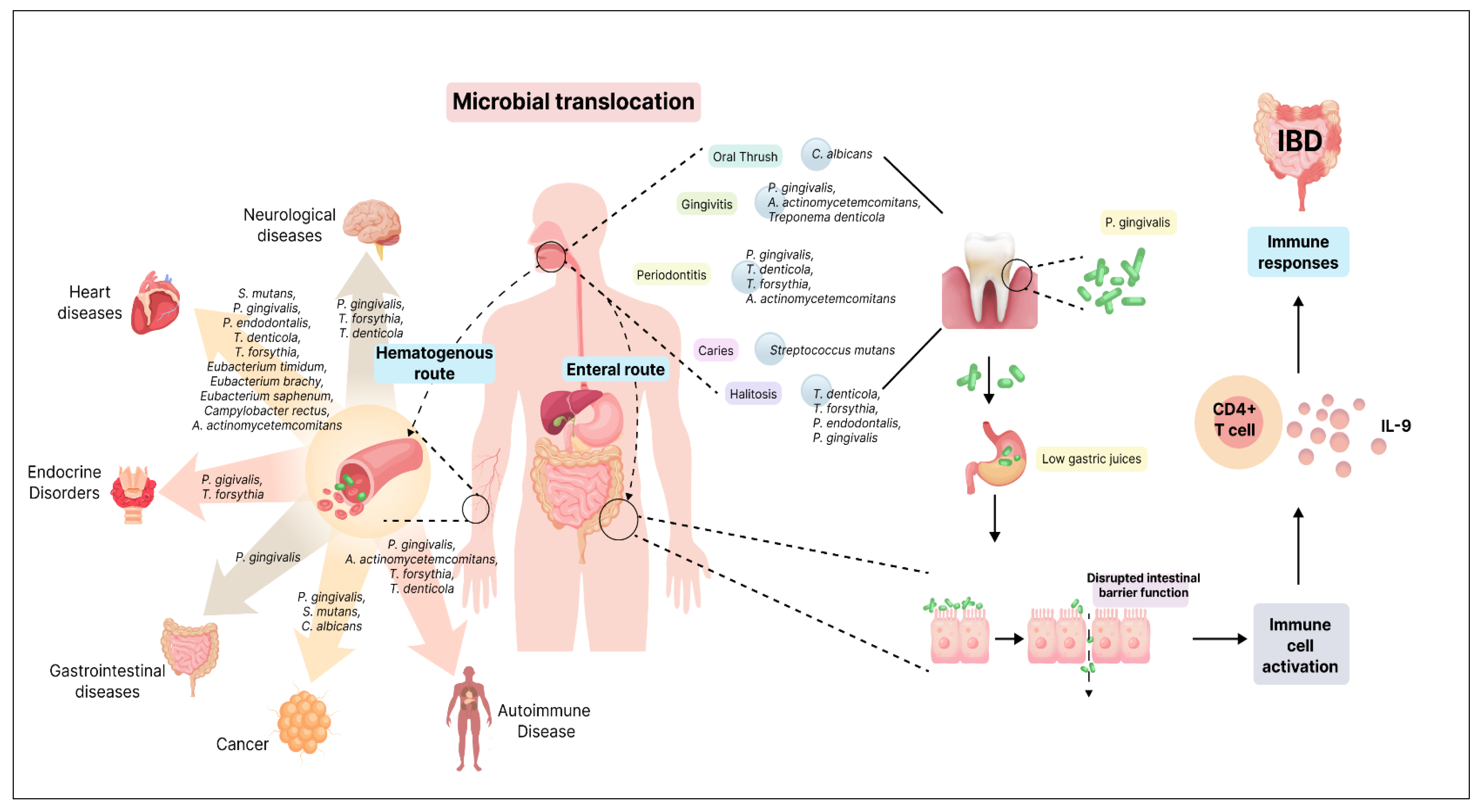
Figure 2.
Mechanisms of Candida albicans Pathogenicity: A) Adhesion to Host Surfaces: Candida albicans initially adheres to host surfaces through weak and reversible interactions influenced by hydrophobic and electrostatic forces, facilitated by host tissue receptors glycoproteins ALS and HWP1. B) Phenotypic Switch: C. albicans can phenotypically switch between yeast and hyphal states. C) Host Cell Receptor Interaction: C. albicans can adhere to epithelial cells using various host cell receptors, such as EphA2 (through β-glucan) and E-cadherin (through ALS3). Host cell transglutaminases create a cross-linking between C. albicans and the epithelial surface by interacting with HWP1. D) Biofilm Formation: Interaction with oral bacteria, such as Streptococcus gordonii, promotes the formation of biofilms. ALS3 on C. albicans binds to surface protein SspB on S. gordonii, facilitating biofilm formation. E) Hyphal penetration into epithelial cells: The shift to the hyphal form is crucial for tissue invasion. Hyphae infiltrate and harm epithelial cells, causing inflammation and white patches in oral thrush. Abbreviations: ALS: Agglutinin-like sequence, HWP1: Hyphal wall protein, SspB: Surface protein SspB, EphA2: Eph receptor A2.
Figure 2.
Mechanisms of Candida albicans Pathogenicity: A) Adhesion to Host Surfaces: Candida albicans initially adheres to host surfaces through weak and reversible interactions influenced by hydrophobic and electrostatic forces, facilitated by host tissue receptors glycoproteins ALS and HWP1. B) Phenotypic Switch: C. albicans can phenotypically switch between yeast and hyphal states. C) Host Cell Receptor Interaction: C. albicans can adhere to epithelial cells using various host cell receptors, such as EphA2 (through β-glucan) and E-cadherin (through ALS3). Host cell transglutaminases create a cross-linking between C. albicans and the epithelial surface by interacting with HWP1. D) Biofilm Formation: Interaction with oral bacteria, such as Streptococcus gordonii, promotes the formation of biofilms. ALS3 on C. albicans binds to surface protein SspB on S. gordonii, facilitating biofilm formation. E) Hyphal penetration into epithelial cells: The shift to the hyphal form is crucial for tissue invasion. Hyphae infiltrate and harm epithelial cells, causing inflammation and white patches in oral thrush. Abbreviations: ALS: Agglutinin-like sequence, HWP1: Hyphal wall protein, SspB: Surface protein SspB, EphA2: Eph receptor A2.
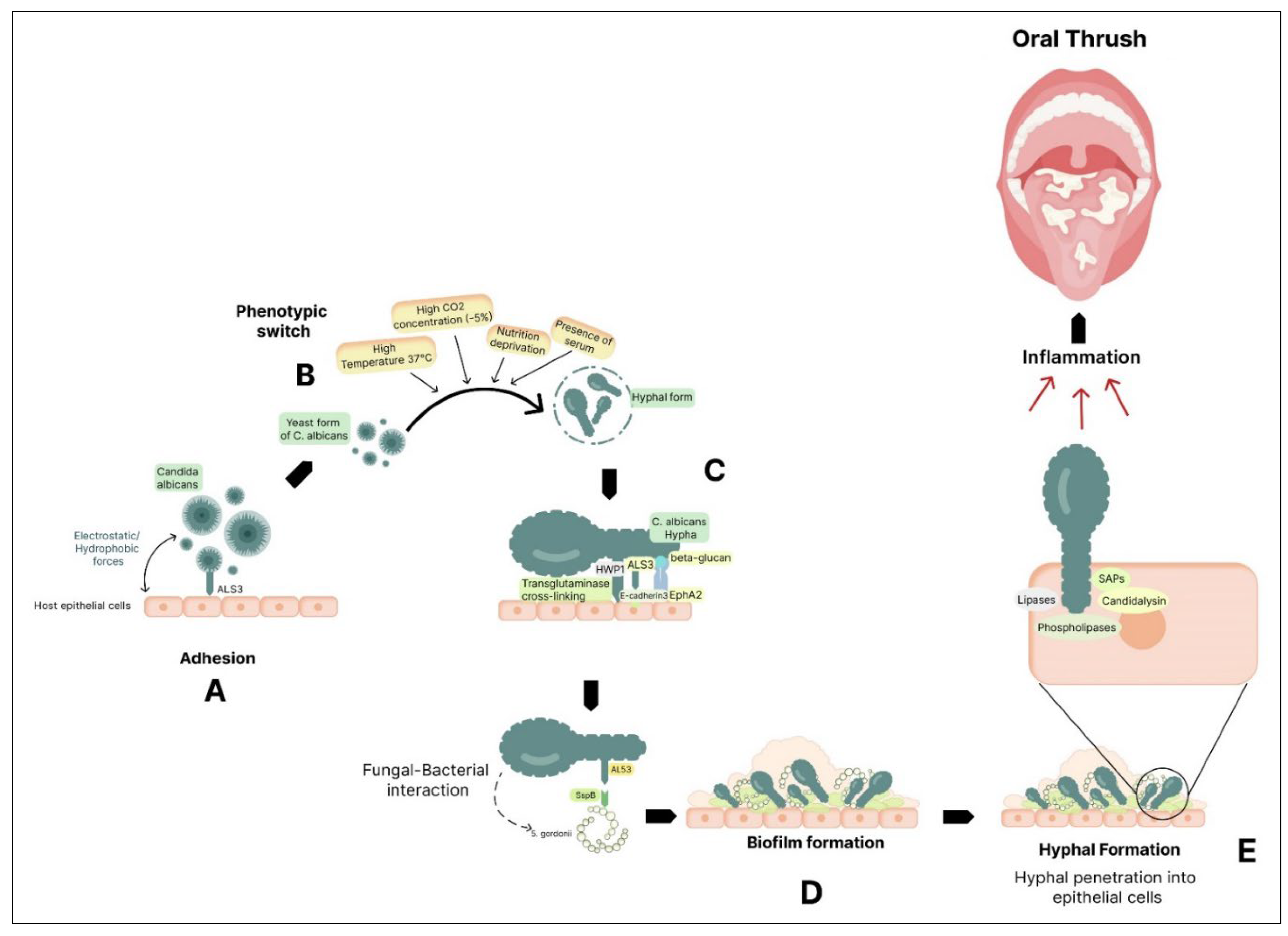
Figure 3.
Mechanisms Linking Oral Pathogens to RA:Porphyromonas gingivalis and RA Link: P. gingivalis possesses virulent factors, including the PPAD, which citrullinates host proteins. This citrullination is thought to trigger autoimmune responses, leading to the production of ACPAs characteristic of RA. NETs and ACPA Production: NETs formed during periodontitis contribute to protein citrullination catalysed by PAD4 enzymes, further enhancing ACPA production. Aggregatibacter actinomycetemcomitans and RA: A. actinomycetemcomitans secretes LtxA, triggering neutrophil hypercitrullination and the release of citrullinated autoantigens, further fuelling the ACPA response. Citrullinated autoantigens can trigger an autoimmune response, wherein, Th17 cells in the immune system become overactivated. Th17 cells are known for producing pro-inflammatory cytokines, notably IL-17. IL-17 stimulates B cells to produce antibodies, including ACPAs. These antibodies can target citrullinated proteins, contributing to the autoimmune processes and inflammation seen in RA. Abbreviations: PPAD: Protein Arginine Deiminase of P. gingivalis, ACPAs: Anticitrullinated Protein Antibodies, NETs: Neutrophil Extracellular Traps, PAD4: Peptidylarginine Deiminase 4, LtxA: Leukotoxin A, Th17: T Helper 17, IL 17: Interleukin-17, RA: Rheumatoid Arthritis.
Figure 3.
Mechanisms Linking Oral Pathogens to RA:Porphyromonas gingivalis and RA Link: P. gingivalis possesses virulent factors, including the PPAD, which citrullinates host proteins. This citrullination is thought to trigger autoimmune responses, leading to the production of ACPAs characteristic of RA. NETs and ACPA Production: NETs formed during periodontitis contribute to protein citrullination catalysed by PAD4 enzymes, further enhancing ACPA production. Aggregatibacter actinomycetemcomitans and RA: A. actinomycetemcomitans secretes LtxA, triggering neutrophil hypercitrullination and the release of citrullinated autoantigens, further fuelling the ACPA response. Citrullinated autoantigens can trigger an autoimmune response, wherein, Th17 cells in the immune system become overactivated. Th17 cells are known for producing pro-inflammatory cytokines, notably IL-17. IL-17 stimulates B cells to produce antibodies, including ACPAs. These antibodies can target citrullinated proteins, contributing to the autoimmune processes and inflammation seen in RA. Abbreviations: PPAD: Protein Arginine Deiminase of P. gingivalis, ACPAs: Anticitrullinated Protein Antibodies, NETs: Neutrophil Extracellular Traps, PAD4: Peptidylarginine Deiminase 4, LtxA: Leukotoxin A, Th17: T Helper 17, IL 17: Interleukin-17, RA: Rheumatoid Arthritis.
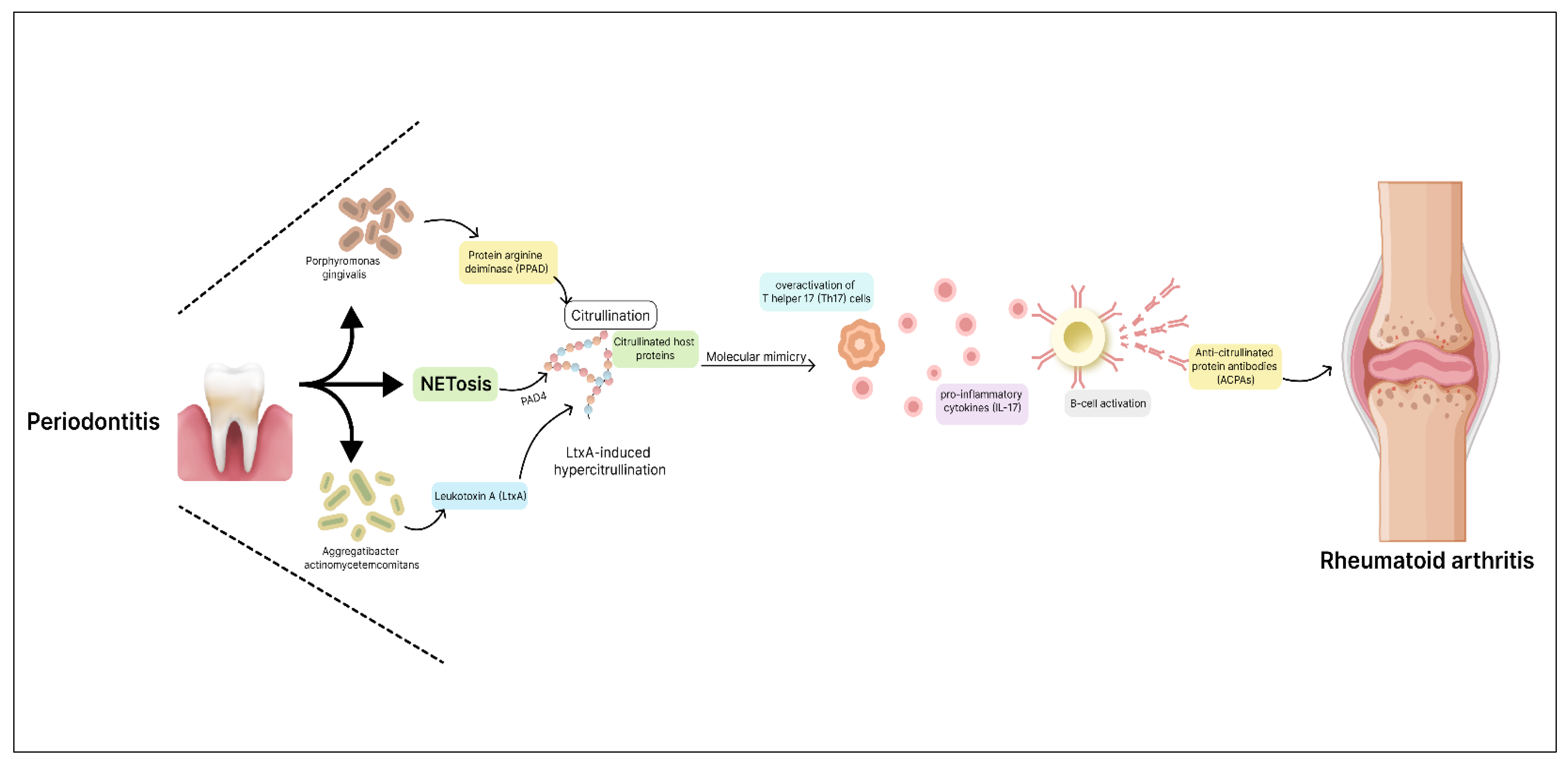

Table 1.
The composition of the oral microbiome.
| Microbial Component | Predominant Phyla/Genera | Other Notable Genera |
|---|---|---|
| Bacteria | Actinobacteria, Bacteroidetes, Chlamydia, Euryarchaeota, Fusobacteria, Firmicutes, Proteobacteria, Spirochaetes, Tenericutes |
- Gram-positive cocci: Abiotrophia, Peptostreptococcus, Streptococcus, Stomatococcus - Gram-positive rods: Actinomyces, Bifidobacterium, Corynebacterium, Eubacterium, Lactobacillus, Propionibacterium, Pseudoramibacter, Rothia - Gram-negative cocci: Moraxella, Neisseria, Veillonella - Gram-negative rods: Campylobacter, Capnocytophaga, Desulfobacter, Desulfovibrio, Eikenella, Fusobacterium, Hemophilus, Leptotrichia, Prevotella, Selemonas, Simonsiella, Treponema, Wolinella |
| Fungi | Candida | Cladosporium, Aureobasidium, Saccharomycetales, Aspergillus, Fusarium, Cryptococcus |
| Archaea | Euryarchaeota | - |
Table 2.
Oral microbes and disease associations.
| Disease | Commensals | Pathogens | Directionality | Mechanisms | References |
|---|---|---|---|---|---|
| Oral Diseases | |||||
| Dental Caries |
Lactobacillus spp., Veillonella, Propionibacterium, Bifidobacterium, Corynebacterium, Capnocytophaga |
S. mutans | ↑ | Acidogenic bacteria produce acidic by-products, leading to demineralization and cavitation; Dysbiosis-driven disorder | [64,65,66,71,72,73,74,75,76,77] |
| Gingivitis | Streptococcus sp., Actinomyces sp., Veillonella sp. |
P. gingivalis, T. denticola, A. actinomycetemcomitans, Fusobacterium sp., P. intermedia |
↑ | Caused by the accumulation of microbial plaque on the tooth surface, which penetrates the gingival tissue and leads to inflammation. | [79] |
| Periodontitis | Prevotella melaninogenica |
P. gingivalis, T. denticola, T. forsythia, F. nucleatum ss. polymorphum, P. intermedia |
↑ | Pathogenic bacteria induce inflammation, oxidative stress, immune activation, and tissue damage; Potential systemic implications | [11,83,84,85] |
| Halitosis |
Prevotella melaninogenica, Veillonella spp. Peptostreptococcus Actinomyces spp. Eubacterium, Megasphaera, Selenomonas, Leptotrichia, Fusobacterium, Eikenella corrodens |
Treponema denticola, Porphyromonas gingivalis, Porphyromonas endodontalis, Tannerella forsythensis, Bacteroides loescheii, Centipeda periodontii |
↑ | Anaerobic bacteria produce VSCs causing malodor; Bacterial degradation of sulfur-containing amino acids | [92,93,94,95] |
| Taste impairment | Lactobacilli | - | ↑ | High levels of acid produced by the bacteria impair taste, affecting taste perception. | [99] |
| Burning mouth syndrome (BMS) |
Streptococcus, Rothia, Bergeyella, Granulicatella |
- | ↑ | Alteration in bacterial strains may contribute to the development of BMS influencing pathways involved in inflammation, immune responses, and sensory perception | [108] |
| Oral Thrush |
Candida parapsilosis, Candida krusei, Candida stellatoidea, Candida tropicalis |
Candida albicans, Candida glabrata, Candida dubliniensis, Candida guilliermondii |
↑ | Overgrowth of Candida species due to factors like poor oral hygiene, weakened immune system, or underlying medical conditions; Adhesion to host surfaces and tissue invasion through various mechanisms; Hyphal formation, biofilm production, and secretion of enzymes that degrade host immune factors | [109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128] |
| Systemic diseases | |||||
| IBD | - |
P. gingivalis, F. nucleatum |
↑ | Oral-resident bacteria may infiltrate the gut microbiome; Gut dysbiosis due to altered gut epithelial permeability | [59,132,134] |
| Atherosclerosis |
Prevotella nigrescens, Parvimonas micra, |
S. mutans, P. gingivalis, P. endodontalis, T. denticola, T. forsythia, Prevotella intermedia, Aggregatibacter actinomycetemcomitans, Eubacterium timidum, Eubacterium brachy, Eubacterium saphenum, Campylobacter rectus, |
↑ | Oral dysbiosis triggers local inflammation, systemic inflammatory responses, oxidative stress, immune activation, platelet aggregation | [135,136,137,138,145,146,147] |
| Diabetes | Capnocytophaga, |
P. gingivalis, T. forsythia |
↑ | Hyperglycemia and oxidative stress create a conducive environment for microbial dysbiosis; Elevated inflammatory reactions |
[151,152,153,154,155,156] |
| Obesity |
Proteobacteria, Chloroflexi, Firmicutes |
- | ↑ | Potential migration of oral bacteria to the gut; Alterations in bacterial populations impact metabolic homeostasis | [162,163,164,165,166] |
| AD | - |
P. gingivalis, T. forsythia, T. denticola, |
↑ | Oral microbiota’s production of inflammatory agents potentially contributes to neuroinflammation and AD progression | [168,169,170,171,173,176] |
| PD | - | P. gingivalis | ↑ | P. gingivalis infection is correlated with PD, with studies demonstrating the presence of gingipain R1 (RgpA) in the bloodstream, indicating systemic dissemination. P. gingivalis may contribute to PD pathogenesis by inducing systemic inflammation, promoting hypercoagulability, and exacerbating neurodegeneration. | [180,183,184] |
| RA |
Prevotella, Veillonella, Lactobacillus salivarius |
P. gingivalis, A. actinomycetemcomitans, |
↑ | P. gingivalis implicated in RA onset via citrullination and ACPA production; Dysbiosis of oral microbiota exacerbates joint inflammation. | [186,188,189,190] |
| SLE |
Veillonella, Streptococcus, Prevotella |
T. forsythia, T. denticola |
↑ | Oral microbial dysbiosis and periodontitis may exacerbate SLE via immune activation; Potential contribution to autoimmune responses | [197,198,199,200,201,202] |
| Cancer |
S. oralis, S. mitis, S. sanguinis, Lactobacillus fermentum, Lactobacillus acidophilus, Bifidobacterium adolescentis |
P. gingivalis, F. nucleatum, |
↑ | Specific bacteria infiltrate cells, initiate tumor development, produce cancer-promoting substances like lipopolysaccharide | [203,204] |
Table 3.
Summary of oral health compounds: Mechanisms of action and formulations.
| Component | Role/Mechanism | Formulation | References |
|---|---|---|---|
| Probiotics | Support balance of oral microbiota, produce specialized metabolites for maintaining microbiota equilibrium, promote healthy immunity |
Probiotic supplements containing strains such as Streptococcus salivarius M18, Streptococcus salivarius K12, Lactobacillus plantarum, Bifidobacterium lactis, Lactobacillus reuteri, Lactobacillus salivarius, help maintain oral microbiota balance, combat bad breath and gum inflammation, and enhance immune responses. | [205] |
| Xylitol | Rebalances mouth acidity, reduces S. mutans counts, and disrupts S. mutans energy production. | Xylitol mints and gums, contains xylitol, natural peppermint flavour, magnesium stearate, natural menthol |
[225,226] |
| Chlorine Dioxide Mouth Rinse | Neutralizes volatile sulfur compounds (VSCs), kills odour-producing bacteria, reduces plaque and F. nucleatum counts | Stabilized chlorine dioxide (ClO2), trisodium phosphate, citric acid; typically found in commercially available mouth rinses | [228] |
| Thymol Mouth Rinse | Antifungal properties, disrupts C. albicans hyphae production and adhesion to epithelial cells | Thymol, eucalyptol, menthol, methyl salicylate dissolved in ethanol (27%); available in various commercial mouthwash formulations | [230] |
| Fluoride Toothpaste | Strengthens enamel, prevents cavities, promotes oral hygiene | Sodium fluoride, hydrated silica, cellulose gum, glycerin, available in various brands and formulations of toothpaste | [233] |
| Hydroxyapatite Toothpaste | Replenishes lost minerals in enamel, restores enamel structure, alternative to fluoride-based products | Hydroxyapatite particles, available in various brands and formulations of toothpaste | [236] |
| Cetylpyridinium Chloride (CPC) | Antimicrobial properties, disrupts bacterial membranes and cellular functions, effective against plaque-forming bacteria | Found in various oral care formulations such as mouthwashes and toothpaste | [237] |
| Clove Oil | Antibacterial and antifungal effects, inhibits multi-resistant Staphylococcus species growth, reduces C. albicans ergosterol levels | Clove oil containing eugenol, eugenyl acetate, carvacrol, available in various oral care products such as mouth rinses and gels | [238] |
| Oil Pulling | Generates antioxidants to damage microbial cell walls, removes bacteria by attracting them to oil, hinders bacterial co-aggregation and plaque formation | Coconut, sesame, or sunflower oil, typically used as a standalone oral hygiene technique | [239] |
| Antimicrobial Peptides | Antimicrobial peptides (AMPs) such as defensins and cathelicidins disrupt microbial cell membranes through electrostatic interactions, forming pores that lead to ion leakage and cell death, providing natural defense against oral pathogens. | Antimicrobial peptides (AMPs) like defensins and cathelicidins are integrated into oral care products to combat oral bacteria, fungi, and viruses. | [218,219] |
| Enamel Strengthening Peptides | Enamel strengthening peptides, exemplified by peptide P11-4, mimic the mineral structure of enamel. They facilitate remineralization by attracting calcium ions, promoting the formation of hydroxyapatite essential for enamel regeneration and strengthening. | P11-4: dental gels, pastes, some toothpastes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



