
Submitted:
19 August 2024
Posted:
21 August 2024
You are already at the latest version
Abstract
Leishmaniasis, comprising cutaneous and mucocutaneous forms, is a parasitic disease caused by Leishmania species via sandfly bites, leading to skin ulcers and mucosal damage. Traditional treatments include antiparasitic drugs such as pentavalent antimonials, amphotericin B, and miltefosine, with immunotherapy and chemotherapy being explored for enhanced efficacy and reduced relapse. Mucocutaneous leishmaniasis (ML) balances IFN-γ and IL-10, affecting severity, while cutaneous leishmaniasis (CL) shows varied IFN-γ/IL-10 ratios influencing prognosis. Cytokines and chemokines are diagnostic and prognostic markers in CL. Research into neutrophils and the NLRP3 inflammasome informs Th1/Th2 response regulation. Live attenuated LdCen-/- parasites and genetically modified vaccines show promise, with miR-21 as a potential efficacy biomarker. Understanding cytokine networks is crucial for new treatment strategies.
Keywords:
Leishmaniasis
; immune system
; vaccines
; cytokines
; biomarkers
1. Introduction
Leishmaniasis is a parasitic infection that affects people worldwide, primarily in developing countries [1]. The disease is transmitted by the bite of infected sand flies and can present in two main ways: cutaneous, and mucocutaneous. Cutaneous leishmaniasis (CL) is the most common form and causes an open sore at the bite site, which heals in a few months to a year and a half, leaving an unpleasant-looking scar. To date, there are no effective vaccines for leishmaniasis. Many of the treatments available have serious side effects and resistance mechanisms are becoming an increasingly prevalent problem [1,2,3]. Natural Killer T (NKT) cells are a unique T cell population that recognises glycolipids. They are a conserved innate-like lymphocyte population with immunomodulating effects in various settings. The role of NKT cells in the resistance or susceptibility towards Leishmania infections is still not well-defined, as there are conflicting data. However, it is proposed the activation of NKT cells for controlling Leishmania infections [1,2,3,4,5].
The behavior of these cells is influenced by various factors, including the infection site, parasite count, strain virulence, and Leishmania species. NKT cells can be activated through direct or indirect pathways. In the direct pathway, Leishmania glycolipids are presented by CD1d molecules on antigen-presenting cells (such as dendritic cells), leading to cytokine secretion by NKT cells. In the indirect pathway, Leishmania glycolipids stimulate TLR2 in dendritic cells, triggering IL-12 production and subsequent NKT cell activation. The study delves into NKT cell roles in disease development, both in humans and mouse models, and proposes NKT cell activation as a strategy for controlling Leishmania infections [4,5]. Biomarkers and vaccines are two important aspects of leishmaniasis research.
Leishmaniasis, a vector-borne disease caused by the protozoan parasite Leishmania, is endemic in various regions worldwide. Its clinical manifestations range from self-healing skin lesions to the mucocutaneous forms. Recent studies have harnessed large-scale genomic and proteomic analyses to understand infection pathways in both parasites and hosts more effectively than studying individual molecules. Additionally, researchers have identified potential pharmacodynamic biomarkers for different forms of leishmaniasis. Despite these advances, there is currently no approved vaccine for human use against this disease [6,7,8].
2. Immune Response to the Parasite
2.1. Immune Response in Cutaneous Leishmaniasis
The innate immune response serves as our initial defense against pathogens and tissue damage. In cutaneous leishmaniasis (CL), the skin’s innate response involves recognizing pathogenic molecules and activating immune cells like dendritic cells, neutrophils, macrophages, natural killer cells, and molecules like the complement system [9]. These components play a crucial role in containing the parasite, although Leishmania can manipulate them to its advantage, causing damage in the skin layers [10,11,12].
The innate immune response against Leishmania begins with the recruitment of neutrophils, essential polymorphonuclear cells responsible for phagocytosis and lysis of Leishmania promastigotes. These inflammatory cells are drawn to the infection site by soluble chemotactic factors released by Leishmania and IL-8 in the host. Neutrophils recognize the parasite through innate immune receptors (such as Toll-like receptors TLR2 and TLR4) on their membranes. This recognition is crucial for Leishmania destruction, as it activates antimicrobial substances like reactive oxygen species (ROS), nitric oxide (NO), and neutrophil extracellular traps (NETs). Additionally, neutrophils trigger cytokine release, recruit macrophages, and activate cellular immune responses against the pathogen [10,11].
Beyond neutrophils, other innate cells like inflammatory monocytes, mast cells, and NK cells also influence resistance or susceptibility to Leishmania infection. Recent research highlights how metabolic pathways in innate immune cells activate unique immune signal cascades, alongside cytokine and chemokine signals. However, the intricate interplay between metabolic pathways, epigenetic changes, and immune signaling remains a puzzle in Leishmania pathogenesis. A deeper grasp of early interactions between parasites and innate immune cells is crucial for developing effective and safe vaccines against Leishmaniasis [13].
In this study, researchers explored the impact of an empty bacterial pcDNA3 plasmid on mice infected with Leishmania major. They assessed immune mediators, including IFN-γ, IL-4, IL-10, IgG2a, IgG1, arginase activity, and nitric oxide (NO). The results revealed that pcDNA3 modulated immune responses in favor of host cells, reducing disease severity. Notably, Th2-associated mediators were downregulated, while cellular responses increased, accompanied by elevated NO and IFN-γ levels. The study highlights the potential of pcDNA3 as an immunoenhancing agent for treating zoonotic cutaneous leishmaniasis (ZCL). Further research is needed to understand the underlying mechanisms of pcDNA3’s immunogenic properties against ZCL [14].
In this study, the authors investigated the antileishmanial activity of Urtica dioica extract against zoonotic cutaneous leishmaniasis. They found that Urtica dioica extract can modulate the immune responses in favor of host cells and decrease the disease severity. By quantifying immune mediators such as IFN-γ, IL-4, IL-10, IgG2a, IgG1, arginase activity, and nitric oxide (NO), they observed significant reduction in L. major promastigotes viability. Importantly, the extract showed no toxicity to macrophages but effectively killed L. major amastigotes. These findings highlight the potential of Urtica dioica extract as an immunoenhancing agent for treating zoonotic cutaneous leishmaniasis [15]
2.2. Immune Response Against Mucocutaneous Leishmaniasis (ML)
This paper investigates the association between the balance of interferon-gamma (IFN-γ) and IL-10 and the severity of human Leishmania Viannia braziliensis infection. The authors suggest that suitable levels of IFN-γ and IL-10 are essential for the maintenance of protective responses in cutaneous leishmaniasis (CL), while high IFN-γ and low IL-10 production are associated with the severity of mucosal leishmaniasis (ML). The following study involved 33 individuals who recovered from L. braziliensis infection, and the cytokines were quantified by enzyme-linked immunosorbent assay (ELISA) in culture supernatants of L. braziliensis-stimulated peripheral blood mononuclear cells (PBMC). The results showed that cured ML cases maintained significantly lower production of IL-10 in comparison to spontaneous healing of CL or asymptomatic individuals. Thus, a high IFN-γ/IL-10 ratio observed in ML can indicate an unfavorable cytokine balance. Conversely, lower IFN-γ/IL-10 balance observed in cured CL, spontaneous healing of CL, and asymptomatic individuals can represent a better-modulated immune response associated with a favorable prognosis. The study is significant because it provides insights into the association between the balance of IFN-γ and IL-10 and the severity of human Leishmania Viannia braziliensis infection. The authors suggest that the cytokine balance is essential for the maintenance of protective responses in CL, while high IFN-γ and low IL-10 production are associated with the severity of ML. The study highlights the need for further research in this area to understand the mechanisms underlying the association between the balance of IFN-γ and IL-10 and the severity of human Leishmania Viannia braziliensis infection [16].
In this research is discussed the role of Leishmania Viannia guyanensis, LRV1 virus, and extracellular vesicles in the immune response during mucocutaneous leishmaniasis. The authors argue that the interaction between these three factors can influence the outcome of the immune response to the disease. They suggest that the LRV1 virus can modulate the host’s immune response by inhibiting inflammasome activation and inducing the production of proinflammatory cytokines such as TNFα and IL-12. The authors also highlight the role of extracellular vesicles in the pathogenesis of the disease by promoting the survival of Leishmania parasites and inhibiting the host’s immune response. The article provides a comprehensive review of the current understanding of the role of Leishmania Viannia guyanensis, LRV1 virus, and extracellular vesicles in muco-cutaneous leishmaniasis. The study is significant as it offers insights into the complex interplay between Leishmania Viannia guyanensis, the LRV1 virus, and extracellular vesicles in the pathogenesis of mucocutaneous leishmaniasis. Understanding these interactions helps elucidate the mechanisms driving the disease’s progression and severity. By highlighting the roles of these components, the research contributes to developing more effective treatments and potential strategies for managing this debilitating form of leishmaniasis, addressing current challenges of resistance and side effects in existing therapies [17]. In addition, the cells involved in the pathogenesis of the disease include macrophages, neutrophils and dendritic cells [17].
2.3 Immunomodulatory Plants
Drug discovery for combating leishmaniasis has primarily focused on therapeutic switching rather than developing novel drugs. It is crucial to elevate host immunity alongside directly targeting Leishmania parasites. Therefore, an effective antileishmanial approach should encompass agents capable of dual action: eliminating parasites and restoring cell-mediated immunity (CMI). Immunomodulators have shown promise in experimental leishmaniasis, exemplified by imiquimod, which activates toll-like receptor 7 on macrophages and dendritic cells, fostering a Th1 immune response. Other synthetic compounds like the S2 complex (copper chloride, ascorbic acid, and nicotinamide), acetyl salicylic acid, and an immunomodulatory peptide from cystatin, also exhibit both immune-modulating and antileishmanial activities. In addition to synthetic compounds, various herbal formulations and plant secondary metabolites such as saponins, alkaloids, flavonoids, and polysaccharides possess immunomodulatory properties. Plants, in particular, have been extensively researched for their potential as sources of bioactive compounds against Leishmania parasites and for enhancing immune responses. The diverse array of biomolecules found in plant extracts provides a natural arsenal that can combat the parasites while bolstering the host’s immune system. This dual approach not only targets the pathogen directly but also strengthens the body’s ability to resist and control leishmaniasis. As research progresses, harnessing these natural and synthetic immunomodulators could lead to more effective treatments for this challenging disease [18].
Research has shown that certain plants possess immunostimulatory properties that can be beneficial in treating cutaneous and mucocutaneous leishmaniasis. These plants include Azadirachta indica (neem), Allium sativum (garlic), and Curcuma longa (turmeric). Azadirachta indica is known for its antimicrobial and immune-boosting effects. Studies have demonstrated that neem extracts can enhance the production of cytokines and activate macrophages, which are crucial in fighting Leishmania infections. Allium sativum, commonly known as garlic, contains compounds like allicin, which exhibit strong immunomodulatory and anti-leishmanial activities. Garlic has been shown to stimulate the immune system by increasing the activity of natural killer cells and macrophages. Curcuma longa, or turmeric, is rich in curcumin, a compound with potent anti-inflammatory and immunostimulatory properties. Curcumin can modulate the immune response by enhancing the activity of T-cells and macrophages, thereby helping to control Leishmania infections. These plants, through their various active compounds, offer potential complementary therapies for cutaneous and mucocutaneous leishmaniasis by enhancing the immune system’s ability to combat the infection. Further clinical studies and trials are needed to establish their efficacy and safety profiles in humans [19,20].
2.4. Other aspects of the Immune Response in Leishmaniasis
The immune response against Leishmania begins at the site of pathogen entry in the dermis, where promastigotes interact with serum components, activating the complement system via both classical and alternative pathways. Complement opsonization of metacyclic promastigote forms leads to efficient lysis of approximately 90% of inoculated parasites, but Leishmania has developed mechanisms to resist and evade this process [1,21]. Leishmania expresses protein kinases that inhibit complement activation by phosphorylating key components [22,23].
Surface molecules like lipophosphoglycan (LPG) and glycoprotein of 63 kDa (GP63) bind to inactivated C3b (iC3b), preventing complement-mediated lysis and facilitating internalization via complement receptors (CRs) [24]. Sandfly saliva, containing exosomes, gut microbes, and various molecules with immunomodulatory effects, further influences infection outcomes [25,26,27,28]. Resident cells like macrophages, keratinocytes, mast cells, and Langerhans cells express pattern recognition receptors (PRRs), immune response cascades upon encountering Leishmania [29]. Neutrophils [30,31,32], monocytes [33], macrophages [34,35] and dendritic cells (DCs) [36,37,38] are involved in phagocytosing Leishmania parasites. Phagocytosis includes binding to specific receptors, formation of parasitophorous vacuoles, and parasite degradation via reactive oxygen species/reactive nitrogen species (ROS/RNS) [39].
2.5. Inflammasome in Leishmaniasis
Inflammasomes are important components of the innate immune system that help fight bacterial and viral infections. However, their role in regulating adaptive immunity during protozoan parasite infections is not well understood. In this study, researchers found that the NLRP3 inflammasome balances Th1/Th2 responses during leishmaniasis. Mice lacking the inflammasome components NLRP3, ASC, or caspase 1 on a Leishmania-susceptible BALB/c background were resistant to cutaneous L. major infection. The researchers also discovered that production of IL-18 promotes the Th2 cytokine IL-4, which propagates disease in susceptible BALB/c mice. “Neutralization of IL-18 in these animals reduced L. major titers and footpad swelling. The results suggest that activation of the NLRP3 inflammasome is detrimental during leishmaniasis and that IL-18 neutralization has potential as a therapeutic strategy to treat leishmaniasis patients” [40].
It is suggested that the NLRP3 inflammasome plays a dual role in leishmaniasis, both as a protector and a pathogen. The NLRP3 inflammasome is essential for the host’s defense against Leishmania infection by activating the immune system and inducing the production of pro-inflammatory cytokines. The cytokines involved in this process include IL-1β, IL-18, and IL-33. The NLRP3 inflammasome also activates the production of IL-1β, IL-18, and IL-6, which can contribute to the pathogenesis of the disease by inducing excessive inflammation and tissue damage. The cells involved in the dual role of the NLRP3 inflammasome in leishmaniasis include macrophages, neutrophils, and dendritic cells. The balance between the protective and pathogenic roles of the NLRP3 inflammasome is critical for the outcome of the disease, and further research is needed to understand the mechanisms underlying the dual role of the NLRP3 inflammasome in leishmaniasis [41,42,43].
2.6. Leishmania Infection Induces the Production of Reactive Oxygen Species (ROS) and Nitric Oxide (NO) in Macrophages
Leishmania infection in macrophages induces the production of ROS and NO, impacting various signaling pathways including MAPK, NF-κB, and PI3K/Akt. While multiple enzymes contribute to ROS production, NADPH oxidase is crucial in macrophages. Despite ROS production upon infection, Leishmania parasites inhibit ROS generation in phagolysosomes, potentially aiding their survival. However, activated macrophages can still exhibit ROS-dependent killing of Leishmania, with NO playing a role in parasite elimination as well. Studies indicate a dual role for ROS, as they contribute to both host defense and parasite survival, depending on the context of infection and the Leishmania species involved. Furthermore, ROS may influence the establishment of replicative niches within infected macrophages, facilitating long-term parasite survival. Overall, ROS emerge as key players in the intricate interplay between Leishmania parasites and host macrophages, impacting the outcome of infection [39,44,45,46,47,48,49,50,51,52,53,54,55,56,57].
2.7. Summary of the Immune Response to Leishmania spp.
Leishmania infection profoundly affects immune cells, altering their behavior and impacting both innate and adaptive immune responses. Mast cells play a significant role in disease progression by releasing IL-4 and IL-13, promoting Th2 adaptive immune responses and aiding parasite survival. Neutrophils, macrophages, and dendritic cells (innate immune responses) can either eliminate Leishmania parasites or facilitate their survival within the host. Neutrophils eliminate parasites through phagocytosis, ROS, and NETs release, but Leishmania can evade destruction by inhibiting phagolysosome biogenesis and delaying neutrophil apoptosis. Infected neutrophils also attract more immune cells, supporting parasite survival and pathology. Macrophages exhibit plasticity during leishmaniasis, differentiating into M1 or M2 phenotypes, with M1 macrophages promoting Th1 responses for disease control and M2 macrophages supporting Th2 responses and disease progression. Dendritic cells play a crucial role in regulating immune responses by presenting Leishmania antigens to naïve T cells, influencing their differentiation into either Th1 or Th2 cells. Finally, NK cells contribute to the host defense against Leishmania by secreting IFNγ, which enhances the Th1 response, thus aiding in parasite clearance [29,38,58,59]. Figure 1 shows some aspects of the immune defense in leishmaniasis.
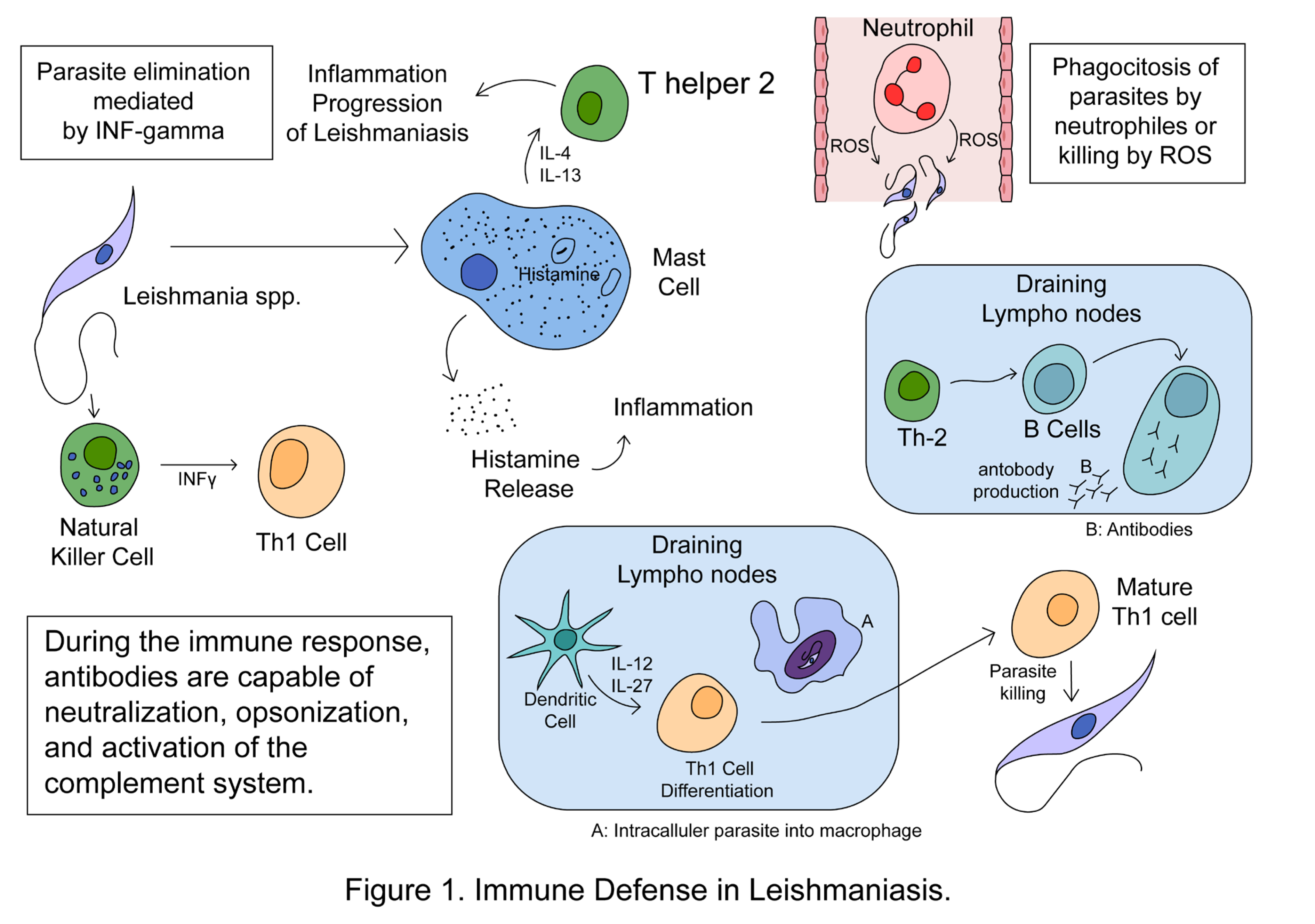
3. Leishmaniasis Biomarkers
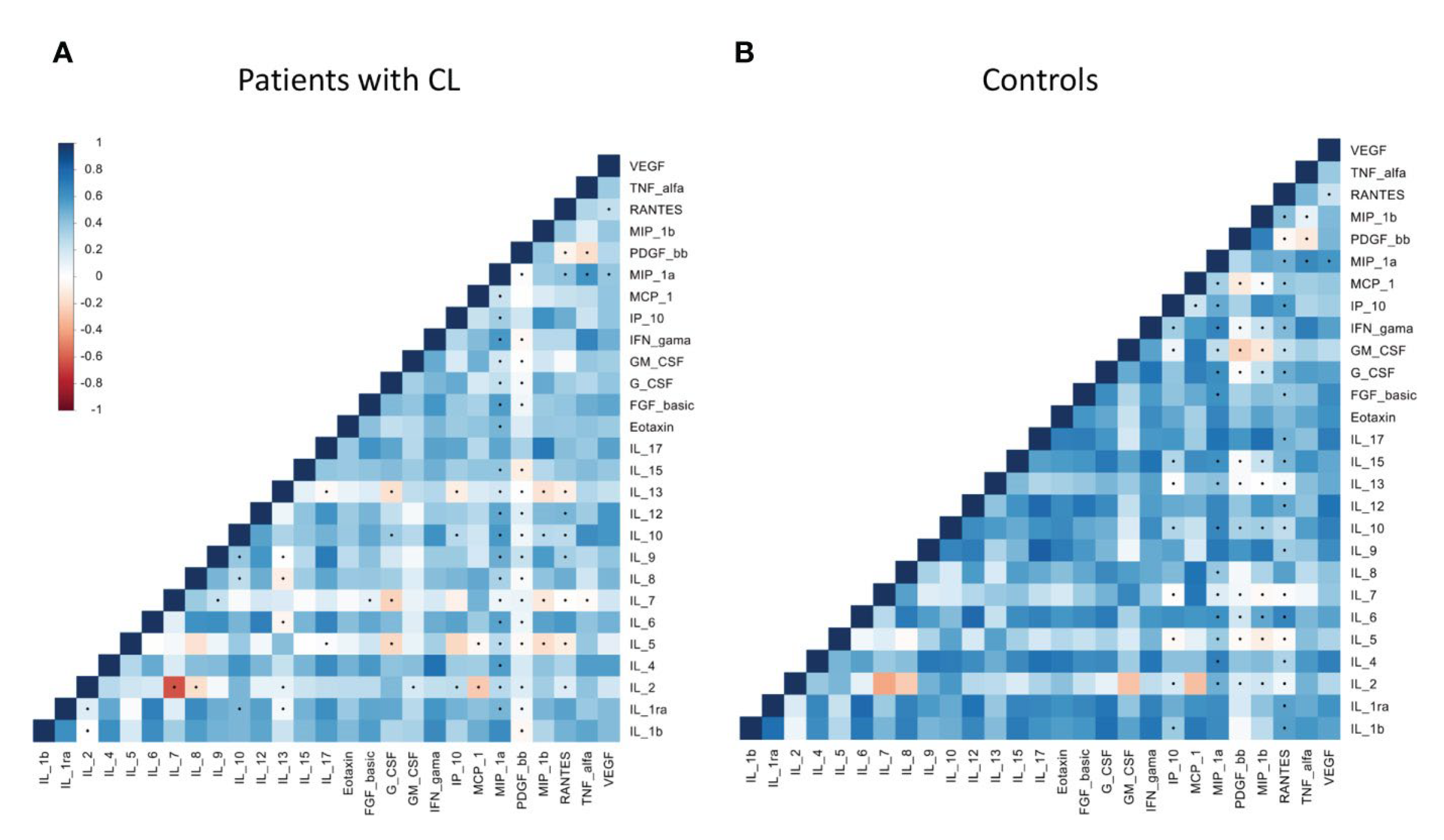
A new study aimed to identify the chemokines and cytokines present in the plasma of patients infected with Leishmania guyanensis, one of the causative agents of cutaneous leishmaniasis. The authors used a multiplex assay to measure the levels of 25 cytokines and chemokines in the plasma of 30 patients with cutaneous leishmaniasis and 30 healthy controls. They found that the levels of several cytokines and chemokines were significantly different between the two groups. For example, the levels of interleukin-1 receptor antagonist (IL-1RA), interleukin-6 (IL-6), and interferon-gamma (IFN-γ) were significantly higher in the plasma of patients with cutaneous leishmaniasis than in healthy controls. Conversely, the levels of interleukin-10 (IL-10) and interleukin-13 (IL-13) were significantly lower in the plasma of patients with cutaneous leishmaniasis than in healthy controls. The authors also performed a principal component analysis (PCA) to identify the cytokines and chemokines that best distinguished between the two groups. They found that the first principal component (PC1) was mainly composed of IL-1RA, IL-6, and IFN-γ, while the second principal component (PC2) was mainly composed of IL-10 and IL-13. The authors suggest that these cytokines and chemokines could be used as biomarkers for the diagnosis and prognosis of cutaneous leishmaniasis. This study provides valuable insights into the cytokine and chemokine profiles of patients with cutaneous leishmaniasis. The study found that cytokines IL-17 and IL1β had the highest positive pair correlations among patients. Biomarker analysis revealed that IP-10, IL-2, and RANTES were the most discriminatory variables in the patient group, while PDGF, IL-1ra, and eotaxin were more prominent in control subjects. The biomarkers IP-10, IL-2, IL-1β, RANTES, and IL-17 demonstrated predictive value for disease progression in Lg-infected individuals. This suggests that these specific cytokines and chemokines could serve as potential indicators for monitoring the development of the disease in infected patients [60]. Table 2 shows Correlation matrices within chemokines, cytokines and growth factors among patients with cutaneous leishmaniasis (A) and healthy controls (B) groups.
Figure 2.
Correlation matrices within chemokines, cytokines and growth factors among patients with cutaneous leishmaniasis (A) and healthy controls (B) groups. Increasing values are represented by colors codes: blue for positive correlation and red for negative correlation. Significance: p< 0.05. Figure taken from open access journal [60].
Figure 2.
Correlation matrices within chemokines, cytokines and growth factors among patients with cutaneous leishmaniasis (A) and healthy controls (B) groups. Increasing values are represented by colors codes: blue for positive correlation and red for negative correlation. Significance: p< 0.05. Figure taken from open access journal [60].
The authors discuss the limitations of current vaccines against CVL and the need for new approaches. The main focus of the article is on the potential of biomarkers to improve the efficacy of vaccines against CVL. The authors describe the various types of biomarkers that have been studied, including those related to the immune response, parasite load, and genetic factors. Biomarkers are measurable indicators that can be used to diagnose, predict, or monitor the progression of a disease. In the context of canine visceral leishmaniasis (CVL), researchers have studied various types of biomarkers that could improve the efficacy of vaccines against this disease. One type of biomarker that has been studied is related to the immune response. Researchers have identified several immune-related biomarkers that could be used to predict the efficacy of vaccines against CVL. For example, the levels of certain cytokines, such as IFN-γ and IL-10, have been shown to be associated with protection against CVL. Another type of biomarker that has been studied is related to parasite load. Researchers have investigated the use of parasite-specific biomarkers, such as the K39 antigen, to monitor the progression of CVL and assess the efficacy of vaccines. Finally, genetic factors have also been studied as potential biomarkers for CVL. Researchers have identified several genetic polymorphisms that are associated with susceptibility to CVL, including those related to the MHC class II genes. They call for further research in this area and suggest that a better understanding of the immune response to Leishmania infection will be critical to the development of effective vaccines. In summary, this study provides a comprehensive overview of the current state of research on CVL and the potential of biomarkers to improve the efficacy of vaccines against this disease. Their work highlights the need for continued research in this area and provides a valuable resource for researchers and clinicians working to combat CVL [61].
4. Investigation on The effects of Treatment of Human Leishmaniasis
In this study, researchers explored the impact of amphotericin B treatment on patients with leishmaniasis, a parasitic disease. By analyzing cytokine production in peripheral blood mononuclear cells (PBMCs), they found that patients with mucocutaneous leishmaniasis (MCL) exhibited higher IFN-γ levels compared to those with cutaneous leishmaniasis (CL). Additionally, MCL patients showed increased IL-12 synthesis. These findings highlight potential avenues for controlling protozoan infections through combined use of immunoenhancing agents and antiprotozoal drugs, emphasizing the need for further research in this area [62].
In Sudanese patients with post-kala-azar dermal leishmaniasis (PKDL), two treatments were compared: paromomycin plus miltefosine (Arm 1) and liposomal amphotericin B plus miltefosine (Arm 2). Arm 1 (98.2% cure rate) triggered Th1/Th2/Th17 responses, while Arm 2 (80% cure rate) induced Th1/Th2 responses. Lower IFN-γ, TNF, and IL-1β levels at baseline were linked to relapses in Arm 2. Arm 1 was more effective in low IFN-γ producers. Assessing IFN-γ, TNF, and IL-1β levels before treatment could predict relapse risk and treatment success [63].
5. Vaccines against Leishmaniasis
Table 1 shows different types of vaccines against leishmaniasis mainly cutaneous and mucocutaneous leishmaniasis (Tegumentary).
The research revealed that neutrophils play a crucial role in initial host responses to pathogens and can independently activate T cell responses or collaborate indirectly with dendritic cells (DCs). Additionally, direct interactions between neutrophils and T cells were observed following immunization with a live attenuated Leishmania donovani centrin-deleted parasite vaccine (LdCen-/-). However, the specific role of neutrophil-DC interactions in T cell priming during vaccine immunity remains less understood. The study compared LdCen-/- infection with wild-type parasite (LdWT) both in vitro and in vivo, concluding that LdCen-/- parasites induced increased expression of CCL3 in neutrophils, leading to enhanced recruitment of DCs and a robust proinflammatory response, as well as elevated co-stimulatory molecule expression compared to LdWT infection [84].
In a recent immunology study, researchers explored the dynamic interactions between neutrophils and dendritic cells (DCs) during immune responses. The LdCen-/- parasite induced increased CCL3 expression in neutrophils, leading to enhanced DC recruitment and robust proinflammatory responses, ultimately shaping vaccine-induced immunity [85].
This paper discusses the development of biomarkers for genetically modified live attenuated Leishmania vaccines against leishmaniasis. The study highlights the need for clear biomarkers associated with protection and safety for live attenuated parasite vaccines [86]. The article discusses the development of a vaccine for leishmaniasis. The authors provide a comprehensive overview of the studies conducted by Dr. Nakhasi’s group in developing a vaccine against leishmaniasis based on genetically modified Leishmania lines lacking a centrin-coding gene. They describe the improvements introduced over time and the challenges faced in developing a vaccine for leishmaniasis [67].
This study was conducted by Fiuza JA et al. and aimed to evaluate the efficacy of a live attenuated vaccine against L. infantum in dogs. The researchers used a centrin gene-deleted L. donovani parasite as a vaccine candidate and tested it on dogs. The results showed that the vaccine was able to induce a protective immune response against L. infantum in dogs. The study also found that the vaccine was safe and did not cause any adverse effects in the dogs. However, further research is needed to evaluate the long-term efficacy of the vaccine and its safety in different dog breeds and ages [87].
The study aimed to evaluate the efficacy of a live attenuated vaccine against Leishmania major and Leishmania infantum in BALB/c mice. The researchers used a p27 gene-deleted L. major parasite as a vaccine candidate and tested it on mice. The results showed that the vaccine was able to induce a protective immune response against L. major and L. infantum in mice. The study also found that the vaccine was safe and did not cause any adverse effects in the mice. The study suggests that vaccination using live attenuated L. major parasites with p27 gene deletion can provide protection against L. major and L. infantum in mice. However, further research is needed to evaluate the long-term efficacy of the vaccine and its safety in different animal models and humans [68].
Th1 concomitant immune response mediated by IFN-γ protects against sand fly delivered Leishmania infection [88]. A novel vaccination approach combining recombinant L. tarentolae (a nonpathogenic lizard protozoan parasite) with a sand fly salivary antigen (PpSP15) shows promise as a protective vaccine against major infection [70]. Figure 3 displays cytokines involved in host protection and disease progression.

Figure 3.
Cytokines involved in host protection (yellow), and disease progression (blue). Adapted from [89].
Figure 3.
Cytokines involved in host protection (yellow), and disease progression (blue). Adapted from [89].
| Cytokine | Function |
Reference |
| IFN-γ |
Activates monocytes and macrophages to produce oxygen radicals | [90,91,92,93,94] |
| IL-2 | Activates T cells and NK cells |
[95,96] |
| TNF-α | Activates the macrophages to kill amastigotes |
[97] |
| IL-12 | Drives Th-1 response and IFN-γ production |
[98] |
| IL-10 | Inhibits interferon-gamma, IL-1, IL-6, IL-12, and TNF-α production |
[99,100] |
| TGF-β | Enhances arginase expression and polyamines synthesis |
[101,102,103] |
| IL-5 | Favours the parasite growth and dissemination |
[104] |
| IL-6 | Favours Th2 response and suppresses macrophage activation |
[105] |
Leishmaniasis, a vector-borne disease prevalent in over 100 countries, lacks an available vaccine for human use. Despite evaluating various candidate antigens, comprehensive biomarkers that serve as reliable indicators of protection or exacerbation remain elusive. While host immune factors associated with disease progression have been identified, no definitive biomarkers have emerged. Researchers propose that combining data from animal models and patients could unveil specific immunity biomarkers, potentially guiding vaccine and drug development against leishmaniasis. Four candidate vaccines have been developed to or near to the clinic [106,107,108]. These are:
- LEISHF3+ GLA-SE: A recombinant fusion protein delivered with strong Th1-inducing adjuvants.
- LeishDNAvax: A naked multi-epitope DNA vaccine.
- ChAd63-KH: An adenovirus-based vaccine.
These vaccines have been proposed to counter cutaneous and also visceral leishmaniasis, which is not the object of study in this review (CL and VL, respectively). However, the authors of the review suggest that more research is needed to determine the safety and efficacy of these vaccines. In addition to these candidate vaccines, researchers are also exploring other vaccine strategies such as the use of sand fly saliva proteins as vaccine candidates [81,109].
6. Conclusion
The immune response to Leishmania spp. varies significantly between mucocutaneous leishmaniasis (ML) and cutaneous leishmaniasis (CL), reflecting distinct disease mechanisms and outcomes. In ML, the balance between interferon-gamma (IFN-γ) and interleukin-10 (IL-10) levels is critical; high IFN-γ and low IL-10 are associated with severe disease, whereas a lower IFN-γ/IL-10 balance correlates with a better prognosis in CL. Factors like the Leishmania RNA virus 1 (LRV1) and extracellular vesicles influence immune responses in ML, complicating disease progression. Diagnostic and prognostic biomarkers, such as cytokines and chemokines, are pivotal in CL. They aid in distinguishing active infections from latent ones and guide treatment decisions. Neutrophils play a dual role in leishmaniasis, contributing to parasite clearance but also potentially exacerbating disease. The NLRP3 inflammasome, crucial for regulating Th1/Th2 responses, can promote disease propagation in susceptible hosts, though its exact role requires further elucidation. Efforts to develop vaccines against cutaneous and mucocutaneous leishmaniasis have employed various strategies. Subunit vaccines, focusing on specific parasite antigens like Leish-111f from Leishmania major, aim to induce protective immunity and have shown promise in trials. Alternatively, whole-parasite vaccines, using live attenuated or killed parasites, mimic natural infection without causing disease. Despite successes in animal models, challenges remain in translating these vaccines to safe and effective human use, especially across diverse Leishmania species and geographical settings. Advancements in vaccine technologies, including recombinant DNA techniques and novel adjuvants, offer hope for overcoming these challenges. Continued research is crucial for developing durable immunity and effective vaccines against these complex and debilitating diseases.
Author Contributions
The manuscript was conceptualized by A.J.-V. and P.E.A. Planning and discussion were conducted by all authors. A.J.-V. wrote the initial draft of the manuscript. A.J.-V. and D.G. reviewed the manuscript. All authors investigated, participated in software, writing or review of the final draft. All authors have read and agreed to the published version of the manuscript.
Funding
This study did not receive any external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The dataset supporting the findings of this study is included within the manuscript and its referenced sources, ensuring comprehensive access to the relevant data for further examination and analysis.
Acknowledgments
The authors would like to sincerely thank West Indian Immunology Society (WIIS) for their assistance.
Conflicts of Interest
The authors declare no conflict of interest
References
- von Stebut, E. Cutaneous Leishmania Infection: Progress in Pathogenesis Research and Experimental Therapy. Exp. Dermatol. 2007, 16, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Marín, M.; López, M.; Gallego-Yerga, L.; Álvarez, R.; Peláez, R. Experimental Structure Based Drug Design (SBDD) Applications for Anti-Leishmanial Drugs: A Paradigm Shift? Med. Res. Rev. 2024, 44, 1055–1120. [Google Scholar] [CrossRef] [PubMed]
- Pace, D. Leishmaniasis. J. Infect. 2014, 69 Suppl 1, S10–S18. [Google Scholar] [CrossRef]
- Bendelac, A.; Savage, P.B.; Teyton, L. The Biology of NKT Cells. Annu. Rev. Immunol. 2007, 25, 297–336. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Chimal, J.; Hernández-Ruiz, J.; Becker, I. NKT Cells in Leishmaniasis. Immunobiology 2017, 222, 641–646. [Google Scholar] [CrossRef]
- Jain, K.; Jain, N.K. Vaccines for Visceral Leishmaniasis: A Review. J. Immunol. Methods 2015, 422, 1–12. [Google Scholar] [CrossRef]
- Veras, P.S.T.; Ramos, P.I.P.; de Menezes, J.P.B. In Search of Biomarkers for Pathogenesis and Control of Leishmaniasis by Global Analyses of Leishmania-Infected Macrophages. Front. Cell. Infect. Microbiol. 2018, 8, 326. [Google Scholar] [CrossRef]
- Seyed, N., Taheri, T., Rafati, S., & Taslimi, Y. Identification of Leishmania Vaccine Candidates: A Proteome-Wide Immunoinformatics Approach. Parasitol. Res. 2018.
- Liese, J.; Schleicher, U.; Bogdan, C. The Innate Immune Response against Leishmania Parasites. Immunobiology 2008, 213, 377–387. [Google Scholar] [CrossRef]
- Serrano-Coll, H.; Cardona-Castro, N.; Ramos, A.P.; Llanos-Cuentas, A. Innate Immune Response: Ally or Enemy in Cutaneous Leishmaniasis? Pathog. Dis. 2021, 79. [Google Scholar] [CrossRef]
- Feijó, D.; Tibúrcio, R.; Ampuero, M.; Brodskyn, C.; Tavares, N. Dendritic Cells and Leishmania Infection: Adding Layers of Complexity to a Complex Disease. J Immunol Res 2016, 2016, 3967436. [Google Scholar] [CrossRef]
- Carlsen, E.D.; Liang, Y.; Shelite, T.R.; Walker, D.H.; Melby, P.C.; Soong, L. Permissive and Protective Roles for Neutrophils in Leishmaniasis. Clin. Exp. Immunol. 2015, 182, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Volpedo, G.; Pacheco-Fernandez, T.; Bhattacharya, P.; Oljuskin, T.; Dey, R.; Gannavaram, S.; Satoskar, A.R.; Nakhasi, H.L. Determinants of Innate Immunity in Visceral Leishmaniasis and Their Implication in Vaccine Development. Front. Immunol. 2021, 12, 748325. [Google Scholar] [CrossRef] [PubMed]
- Montakhab-Yeganeh, H.; Shafiei, R.; Najm, M.; Masoori, L.; Aspatwar, A.; Badirzadeh, A. Immunogenic Properties of Empty pcDNA3 Plasmid against Zoonotic Cutaneous Leishmaniasis in Mice. PLoS One 2022, 17, e0263993. [Google Scholar] [CrossRef] [PubMed]
- Badirzadeh, A.; Heidari-Kharaji, M.; Fallah-Omrani, V.; Dabiri, H.; Araghi, A.; Salimi Chirani, A. Antileishmanial Activity of Urtica Dioica Extract against Zoonotic Cutaneous Leishmaniasis. PLoS Negl. Trop. Dis. 2020, 14, e0007843. [Google Scholar] [CrossRef]
- Gomes-Silva, A.; de Cássia Bittar, R.; Dos Santos Nogueira, R.; Amato, V.S.; da Silva Mattos, M.; Oliveira-Neto, M.P.; Coutinho, S.G.; Da-Cruz, A.M. Can Interferon-Gamma and Interleukin-10 Balance Be Associated with Severity of Human Leishmania (Viannia) Braziliensis Infection? Clin. Exp. Immunol. 2007, 149, 440–444. [Google Scholar] [CrossRef]
- Olivier, M.; Zamboni, D.S. Leishmania Viannia Guyanensis, LRV1 Virus and Extracellular Vesicles: A Dangerous Trio Influencing the Faith of Immune Response during Muco-Cutaneous Leishmaniasis. Curr. Opin. Immunol. 2020, 66, 108–113. [Google Scholar] [CrossRef]
- Chouhan, G.; Islamuddin, M.; Sahal, D.; Afrin, F. Exploring the Role of Medicinal Plant-Based Immunomodulators for Effective Therapy of Leishmaniasis. Front. Immunol. 2014, 5, 193. [Google Scholar] [CrossRef]
- Carneiro, S.M.P.; Carvalho, F.A.A.; Santana, L.C.L.R.; Sousa, A.P.L.; Neto, J.M.M.; Chaves, M.H. The Cytotoxic and Antileishmanial Activity of Extracts and Fractions of Leaves and Fruits of Azadirachta Indica (A Juss.). Biol. Res. 2012, 45, 111–116. [Google Scholar] [CrossRef]
- Oryan, A. Plant-Derived Compounds in Treatment of Leishmaniasis. Iran J Vet Res 2015, 16, 1–19. [Google Scholar]
- Regli, I.B.; Passelli, K.; Hurrell, B.P.; Tacchini-Cottier, F. Survival Mechanisms Used by Some Leishmania Species to Escape Neutrophil Killing. Front. Immunol. 2017, 8, 1558. [Google Scholar] [CrossRef]
- Salei, N.; Hellberg, L.; Köhl, J.; Laskay, T. Enhanced Survival of Leishmania Major in Neutrophil Granulocytes in the Presence of Apoptotic Cells. PLoS One 2017, 12, e0171850. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Mandal, A.; Ansari, M.Y.; Kumar, A.; Abhishek, K.; Ghosh, A.K.; Kumar, A.; Kumar, V.; Das, S.; Das, P. Leishmania Donovani Inhibitor of Serine Peptidases 2 Mediated Inhibition of Lectin Pathway and Upregulation of C5aR Signaling Promote Parasite Survival inside Host. Front. Immunol. 2018, 9, 63. [Google Scholar] [CrossRef]
- Hermoso, T.; Fishelson, Z.; Becker, S.I.; Hirschberg, K.; Jaffe, C.L. Leishmanial Protein Kinases Phosphorylate Components of the Complement System. EMBO J. 1991, 10, 4061–4067. [Google Scholar] [CrossRef] [PubMed]
- Francesquini, F.C.; Silveira, F.T.; Passero, L.F.D.; Tomokane, T.Y.; Carvalho, A.K.; Corbett, C.E.P.; Laurenti, M.D. Salivary Gland Homogenates from Wild-Caught Sand Flies Lutzomyia Flaviscutellata and Lutzomyia (Psychodopygus) Complexus Showed Inhibitory Effects on Leishmania (Leishmania) Amazonensis and Leishmania (Viannia) Braziliensis Infection in BALB/c Mice. Int. J. Exp. Pathol. 2014, 95, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Lestinova, T.; Vlkova, M.; Votypka, J.; Volf, P.; Rohousova, I. Phlebotomus Papatasi Exposure Cross-Protects Mice against Leishmania Major Co-Inoculated with Phlebotomus Duboscqi Salivary Gland Homogenate. Acta Trop. 2015, 144, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Thiakaki, M.; Rohousova, I.; Volfova, V.; Volf, P.; Chang, K.-P.; Soteriadou, K. Sand Fly Specificity of Saliva-Mediated Protective Immunity in Leishmania Amazonensis-BALB/c Mouse Model. Microbes Infect. 2005, 7, 760–766. [Google Scholar] [CrossRef]
- Abdeladhim, M.; Kamhawi, S.; Valenzuela, J.G. What’s behind a Sand Fly Bite? The Profound Effect of Sand Fly Saliva on Host Hemostasis, Inflammation and Immunity. Infect. Genet. Evol. 2014, 28, 691–703. [Google Scholar] [CrossRef]
- Pacheco-Fernandez, T.; Volpedo, G.; Verma, C.; Satoskar, A.R. Understanding the Immune Responses Involved in Mediating Protection or Immunopathology during Leishmaniasis. Biochem. Soc. Trans. 2021, 49, 297–311. [Google Scholar] [CrossRef]
- Charmoy, M.; Auderset, F.; Allenbach, C.; Tacchini-Cottier, F. The Prominent Role of Neutrophils during the Initial Phase of Infection by Leishmania Parasites. J. Biomed. Biotechnol. 2010, 2010, 719361. [Google Scholar] [CrossRef]
- Kupani, M.; Pandey, R.K.; Mehrotra, S. Neutrophils and Visceral Leishmaniasis: Impact on Innate Immune Response and Cross-Talks with Macrophages and Dendritic Cells. J. Cell. Physiol. 2021, 236, 2255–2267. [Google Scholar] [CrossRef]
- Passelli, K.; Billion, O.; Tacchini-Cottier, F. The Impact of Neutrophil Recruitment to the Skin on the Pathology Induced by Leishmania Infection. Front. Immunol. 2021, 12, 649348. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.M.; Taye, T.; Akuffo, H.O. Mechanisms of Resistance to Leishmania Aethiopica. I. Interferon-Gamma in Combination with a Cytokine (not Tumor Necrosis Factor-Alpha) Is Required, but Cannot Act Alone in the Inhibition of Intracellular Forms of L. Aethiopica in THP1 Cells. Eur. J. Immunol. 1992, 22, 2331–2337. [Google Scholar] [CrossRef]
- Siqueira-Neto, J.L.; Moon, S.; Jang, J.; Yang, G.; Lee, C.; Moon, H.K.; Chatelain, E.; Genovesio, A.; Cechetto, J.; Freitas-Junior, L.H. An Image-Based High-Content Screening Assay for Compounds Targeting Intracellular Leishmania Donovani Amastigotes in Human Macrophages. PLoS Negl. Trop. Dis. 2012, 6, e1671. [Google Scholar] [CrossRef] [PubMed]
- Corradin, S.B.; Buchmüller-Rouiller, Y.; Mauël, J. Phagocytosis Enhances Murine Macrophage Activation by Interferon-Gamma and Tumor Necrosis Factor-Alpha. Eur. J. Immunol. 1991, 21, 2553–2558. [Google Scholar] [CrossRef]
- Argueta-Donohué, J.; Wilkins-Rodríguez, A.A.; Aguirre-García, M.; Gutiérrez-Kobeh, L. Differential Phagocytosis of Leishmania Mexicana Promastigotes and Amastigotes by Monocyte-Derived Dendritic Cells. Microbiol. Immunol. 2016, 60, 369–381. [Google Scholar] [CrossRef]
- Rivera-Fernández, I.; Argueta-Donohué, J.; Wilkins-Rodríguez, A.A.; Gutiérrez-Kobeh, L. Effect of Two Different Isolates of Leishmania Mexicana in the Production of Cytokines and Phagocytosis by Murine Dendritic Cells. J. Parasitol. 2019, 105, 359–370. [Google Scholar] [CrossRef]
- Kautz-Neu, K.; Schwonberg, K.; Fischer, M.R.; Schermann, A.I.; von Stebut, E. Dendritic Cells in Leishmania Major Infections: Mechanisms of Parasite Uptake, Cell Activation and Evidence for Physiological Relevance. Med. Microbiol. Immunol. 2012, 201, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Filardy, A.A.; Costa-da-Silva, A.C.; Koeller, C.M.; Guimarães-Pinto, K.; Ribeiro-Gomes, F.L.; Lopes, M.F.; Heise, N.; Freire-de-Lima, C.G.; Nunes, M.P.; DosReis, G.A. Infection with Leishmania Major Induces a Cellular Stress Response in Macrophages. PLoS One 2014, 9, e85715. [Google Scholar] [CrossRef] [PubMed]
- Gurung, P.; Karki, R.; Vogel, P.; Watanabe, M.; Bix, M.; Lamkanfi, M.; Kanneganti, T.-D. An NLRP3 Inflammasome-Triggered Th2-Biased Adaptive Immune Response Promotes Leishmaniasis. J. Clin. Invest. 2015, 125, 1329–1338. [Google Scholar] [CrossRef]
- Paget, C.; Doz-Deblauwe, E.; Winter, N.; Briard, B. Specific NLRP3 Inflammasome Assembling and Regulation in Neutrophils: Relevance in Inflammatory and Infectious Diseases. Cells 2022, 11. [Google Scholar] [CrossRef]
- Meyers, A.K.; Zhu, X. The NLRP3 Inflammasome: Metabolic Regulation and Contribution to Inflammaging. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Raneros, A.B.; Bernet, C.R.; Flórez, A.B.; Suarez-Alvarez, B. An Epigenetic Insight into NLRP3 Inflammasome Activation in Inflammation-Related Processes. Biomedicines 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Wanasen, N.; Soong, L. L-Arginine Metabolism and Its Impact on Host Immunity against Leishmania Infection. Immunol. Res. 2008, 41, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Gandarilla, J.A., Osorio-Trujillo, C., & Hernández-Ramírez, V.I Leishmanicidal Activity of NO and H2O2 Alone or in Combination against Leishmania Mexicana. Parasitol. Res. 2013.
- Bedard, K.; Krause, K.-H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Nathan, C.; Cunningham-Bussel, A. Beyond Oxidative Stress: An Immunologist’s Guide to Reactive Oxygen Species. Nat. Rev. Immunol. 2013, 13, 349–361. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric Oxide and the Immune Response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef]
- Antoine, J.C.; Prina, E.; Jouanne, C.; Bongrand, P. Parasitophorous Vacuoles of Leishmania Amazonensis-Infected Macrophages Maintain an Acidic pH. Infect. Immun. 1990, 58, 779–787. [Google Scholar] [CrossRef]
- Nandan, D.; Reiner, N.E. Attenuation of Gamma Interferon-Induced Tyrosine Phosphorylation in Mononuclear Phagocytes Infected with Leishmania Donovani: Selective Inhibition of Signaling through Janus Kinases and Stat1. Infect. Immun. 1995, 63, 4495–4500. [Google Scholar] [CrossRef]
- Green, S.J.; Meltzer, M.S.; Hibbs, J.B., Jr; Nacy, C.A. Activated Macrophages Destroy Intracellular Leishmania Major Amastigotes by an L-Arginine-Dependent Killing Mechanism. J. Immunol. 1990, 144, 278–283. [Google Scholar] [CrossRef]
- Assreuy, J.; Cunha, F.Q.; Epperlein, M.; Noronha-Dutra, A.; O’Donnell, C.A.; Liew, F.Y.; Moncada, S. Production of Nitric Oxide and Superoxide by Activated Macrophages and Killing of Leishmania Major. Eur. J. Immunol. 1994, 24, 672–676. [Google Scholar] [CrossRef]
- Díaz-Gandarilla, J.A., Osorio-Trujillo, C., Hernández-Ramírez, V.I., & Salaiza-Suazo, N. Human Monocytes Infected with Leishmania Mexicana Are Primed for IL-12p70 Production. Parasite Immunology, 33(12), 643-651. 2011.
- Vieira, L.Q., Goldschmidt, M., Nashleanas, M., Pfeffer, K., & Mak, T. Association of LMP-7 with MHC Class I Molecules Accounts for a Maturation-Dependent Increase in Their Peptide Presentation in Leishmania Amazonensis-Infected Cells. European Journal of Immunology, 26(4), 1001-1010 1996.
- Mohapatra, S. Drug Resistance in Leishmaniasis: Newer Developments. Trop. Parasitol. 2014, 4, 4–9. [Google Scholar] [CrossRef]
- Mukbel, R.M.; Patten, C., Jr; Gibson, K.; Ghosh, M.; Petersen, C.; Jones, D.E. Macrophage Killing of Leishmania Amazonensis Amastigotes Requires Both Nitric Oxide and Superoxide. Am. J. Trop. Med. Hyg. 2007, 76, 669–675. [Google Scholar] [CrossRef]
- Farias, L.H.S., Rodrigues, A.P.D., & Silveira, F.T New Approaches on Leishmania Therapy and Prevention Based on Understanding of Leishmania Interactions within the Mammalian Host. Veterinary Parasitology, 258, 54-64. 2018.
- Conde, L.; Maciel, G.; de Assis, G.M.; Freire-de-Lima, L.; Nico, D.; Vale, A.; Freire-de-Lima, C.G.; Morrot, A. Humoral Response in Leishmaniasis. Front. Cell. Infect. Microbiol. 2022, 12, 1063291. [Google Scholar] [CrossRef]
- Volpedo, G.; Pacheco-Fernandez, T.; Holcomb, E.A.; Cipriano, N.; Cox, B.; Satoskar, A.R. Mechanisms of Immunopathogenesis in Cutaneous Leishmaniasis And Post Kala-Azar Dermal Leishmaniasis (PKDL). Front. Cell. Infect. Microbiol. 2021, 11, 685296. [Google Scholar] [CrossRef]
- de Mesquita, T.G.R.; Junior, J. do E.S.; da Silva, L.D.O.; Silva, G.A.V.; de Araújo, F.J.; Pinheiro, S.K.; Kerr, H.K.A.; da Silva, L.S.; de Souza, L.M.; de Almeida, S.A.; et al. Distinct Plasma Chemokines and Cytokines Signatures in Leishmania Guyanensis-Infected Patients with Cutaneous Leishmaniasis. Front. Immunol. 2022, 13, 974051. [Google Scholar]
- Giunchetti, R.C.; Silveira, P.; Resende, L.A.; Leite, J.C.; Melo-Júnior, O.A. de O.; Rodrigues-Alves, M.L.; Costa, L.M.; Lair, D.F.; Chaves, V.R.; Soares, I.D.S.; et al. Canine Visceral Leishmaniasis Biomarkers and Their Employment in Vaccines. Vet. Parasitol. 2019, 271, 87–97. [Google Scholar] [CrossRef]
- Cuna, W.R.; Velasquez, R.; Riva, J.; Guachalla, I.; Rodríguez, C. Enhancement of a TH1 Immune Response in Amphotericin B-Treated Mucocutaneous Leishmaniasis. J. Biomed. Biotechnol. 2007, 2007, 96410. [Google Scholar] [CrossRef]
- Torres, A., Younis, B. M., Alamin, M., Tesema, S., Bernardo, L., Solana, J. C., Moreno, J., Mustafa, A. A., Alves, F., Musa, A. M., & Carrillo, E. Differences in the Cellular Immune Response during and after Treatment of Sudanese Patients with Post-kala-azar Dermal Leishmaniasis, and Possible Implications for Outcome. Journal of epidemiology and global health, 2024.
- Mendonça, S.C.F. Differences in Immune Responses against Leishmania Induced by Infection and by Immunization with Killed Parasite Antigen: Implications for Vaccine Discovery. Parasit. Vectors 2016, 9, 492. [Google Scholar] [CrossRef]
- Hugentobler, F.; Di Roberto, R.B.; Gillard, J.; Cousineau, B. Oral Immunization Using Live Lactococcus Lactis Co-Expressing LACK and IL-12 Protects BALB/c Mice against Leishmania Major Infection. Vaccine 2012, 30, 5726–5732. [Google Scholar] [CrossRef] [PubMed]
- Hugentobler, F.; Yam, K.K.; Gillard, J.; Mahbuba, R.; Olivier, M.; Cousineau, B. Immunization against Leishmania Major Infection Using LACK- and IL-12-Expressing Lactococcus Lactis Induces Delay in Footpad Swelling. PLoS One 2012, 7, e30945. [Google Scholar] [CrossRef] [PubMed]
- Volpedo, G.; Bhattacharya, P.; Gannavaram, S.; Pacheco-Fernandez, T.; Oljuskin, T.; Dey, R.; Satoskar, A.R.; Nakhasi, H.L. The History of Live Attenuated Centrin Gene-Deleted Leishmania Vaccine Candidates. Pathogens 2022, 11. [Google Scholar] [CrossRef]
- Elikaee, S.; Mohebali, M.; Rezaei, S.; Eslami, H.; Khamesipour, A.; Keshavarz, H.; Eshraghian, M.R. Leishmania Major p27 Gene Knockout as a Novel Live Attenuated Vaccine Candidate: Protective Immunity and Efficacy Evaluation against Cutaneous and Visceral Leishmaniasis in BALB/c Mice. Vaccine 2019, 37, 3221–3228. [Google Scholar] [CrossRef]
- Katebi, A.; Riazi-Rad, F.; Varshochian, R.; Ajdary, S. PLGA nanoparticle-delivered Leishmania antigen and TLR agonists as a therapeutic vaccine against cutaneous leishmaniasis in BALB/c mice. International immunopharmacology. 2024, 138, 112538. [Google Scholar] [PubMed]
- Zahedifard, F.; Gholami, E.; Taheri, T.; Taslimi, Y.; Doustdari, F.; Seyed, N.; Torkashvand, F.; Meneses, C.; Papadopoulou, B.; Kamhawi, S.; et al. Enhanced Protective Efficacy of Nonpathogenic Recombinant Leishmania Tarentolae Expressing Cysteine Proteinases Combined with a Sand Fly Salivary Antigen. PLoS Negl. Trop. Dis. 2014, 8, e2751. [Google Scholar] [CrossRef]
- Valenzuela, J.G.; Belkaid, Y.; Garfield, M.K.; Mendez, S.; Kamhawi, S.; Rowton, E.D.; Sacks, D.L.; Ribeiro, J.M. Toward a Defined Anti-Leishmania Vaccine Targeting Vector Antigens: Characterization of a Protective Salivary Protein. J. Exp. Med. 2001, 194, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Requena, J. M.; Iborra, S.; Carrión, J.; Alonso, C.; Soto, M. Recent advances in vaccines for leishmaniasis. Expert opinion on biological therapy. 2004, 4, 1505–1517. [Google Scholar]
- Collin, N.; Gomes, R.; Teixeira, C.; Cheng, L.; Laughinghouse, A.; Ward, J.M.; Elnaiem, D.-E.; Fischer, L.; Valenzuela, J.G.; Kamhawi, S. Sand Fly Salivary Proteins Induce Strong Cellular Immunity in a Natural Reservoir of Visceral Leishmaniasis with Adverse Consequences for Leishmania. PLoS Pathog. 2009, 5, e1000441. [Google Scholar] [CrossRef] [PubMed]
- Tavares, N.M.; Silva, R.A.; Costa, D.J.; Pitombo, M.A.; Fukutani, K.F.; Miranda, J.C.; Valenzuela, J.G.; Barral, A.; de Oliveira, C.I.; Barral-Netto, M.; et al. Lutzomyia Longipalpis Saliva or Salivary Protein LJM19 Protects against Leishmania Braziliensis and the Saliva of Its Vector, Lutzomyia Intermedia. PLoS Negl. Trop. Dis. 2011, 5, e1169. [Google Scholar] [CrossRef]
- Xu, X.; Oliveira, F.; Chang, B.W.; Collin, N.; Gomes, R.; Teixeira, C.; Reynoso, D.; my Pham, V.; Elnaiem, D.-E.; Kamhawi, S.; et al. Structure and Function of a “Yellow” Protein from Saliva of the Sand Fly Lutzomyia Longipalpis That Confers Protective Immunity against Leishmania Major Infection. J. Biol. Chem. 2011, 286, 32383–32393. [Google Scholar] [CrossRef]
- Martin-Martin, I.; Chagas, A.C.; Guimaraes-Costa, A.B.; Amo, L.; Oliveira, F.; Moore, I.N.; DeSouza-Vieira, T.S.; Sanchez, E.E.; Suntravat, M.; Valenzuela, J.G.; et al. Immunity to LuloHya and Lundep, the Salivary Spreading Factors from Lutzomyia Longipalpis, Protects against Leishmania Major Infection. PLoS Pathog. 2018, 14, e1007006. [Google Scholar] [CrossRef]
- Lerner, E.A.; Ribeiro, J.M.; Nelson, R.J.; Lerner, M.R. Isolation of Maxadilan, a Potent Vasodilatory Peptide from the Salivary Glands of the Sand Fly Lutzomyia Longipalpis. J. Biol. Chem. 1991, 266, 11234–11236. [Google Scholar] [CrossRef]
- Gomes, R.; Oliveira, F.; Teixeira, C.; Meneses, C.; Gilmore, D.C.; Elnaiem, D.-E.; Kamhawi, S.; Valenzuela, J.G. Immunity to Sand Fly Salivary Protein LJM11 Modulates Host Response to Vector-Transmitted Leishmania Conferring Ulcer-Free Protection. J. Invest. Dermatol. 2012, 132, 2735–2743. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.; Rowton, E.; Aslan, H.; Gomes, R.; Castrovinci, P.A.; Alvarenga, P.H.; Abdeladhim, M.; Teixeira, C.; Meneses, C.; Kleeman, L.T.; et al. A Sand Fly Salivary Protein Vaccine Shows Efficacy against Vector-Transmitted Cutaneous Leishmaniasis in Nonhuman Primates. Sci. Transl. Med. 2015, 7, 290ra90. [Google Scholar] [CrossRef] [PubMed]
- Cunha, J.M.; Abbehusen, M.; Suarez, M.; Valenzuela, J.; Teixeira, C.R.; Brodskyn, C.I. Immunization with LJM11 Salivary Protein Protects against Infection with Leishmania Braziliensis in the Presence of Lutzomyia Longipalpis Saliva. Acta Trop. 2018, 177, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Gholami, E.; Oliveira, F.; Taheri, T.; Seyed, N.; Gharibzadeh, S.; Gholami, N.; Mizbani, A.; Zali, F.; Habibzadeh, S.; Bakhadj, D.O.; et al. DNA Plasmid Coding for Phlebotomus Sergenti Salivary Protein PsSP9, a Member of the SP15 Family of Proteins, Protects against Leishmania Tropica. PLoS Negl. Trop. Dis. 2019, 13, e0007067. [Google Scholar] [CrossRef]
- Chagas, A.C.; Oliveira, F.; Debrabant, A.; Valenzuela, J.G.; Ribeiro, J.M.C.; Calvo, E. Lundep, a Sand Fly Salivary Endonuclease Increases Leishmania Parasite Survival in Neutrophils and Inhibits XIIa Contact Activation in Human Plasma. PLoS Pathog. 2014, 10, e1003923. [Google Scholar] [CrossRef]
- Morris, R.V.; Shoemaker, C.B.; David, J.R.; Lanzaro, G.C.; Titus, R.G. Sandfly Maxadilan Exacerbates Infection with Leishmania Major and Vaccinating against It Protects against L. Major Infection. J. Immunol. 2001, 167, 5226–5230. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Dey, R.; Saxena, A.; Karmakar, S.; Ismail, N.; Gannavaram, S.; Dagur, P.K.; Satoskar, M.; Satoskar, S.; De Paoli, S.; et al. Essential Role of Neutrophils in the Protective Immune Response Induced by a Live Attenuated Leishmania Vaccine. J. Immunol. 2020, 205, 3333–3347. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Ismail, N.; Saxena, A.; Gannavaram, S.; Dey, R.; Oljuskin, T.; Akue, A.; Takeda, K.; Yu, J.; Karmakar, S.; et al. Neutrophil-Dendritic Cell Interaction Plays an Important Role in Live Attenuated Leishmania Vaccine Induced Immunity. PLoS Negl. Trop. Dis. 2022, 16, e0010224. [Google Scholar] [CrossRef]
- Gannavaram, S.; Dey, R.; Avishek, K.; Selvapandiyan, A.; Salotra, P.; Nakhasi, H.L. Biomarkers of Safety and Immune Protection for Genetically Modified Live Attenuated Leishmania Vaccines against Visceral Leishmaniasis - Discovery and Implications. Front. Immunol. 2014, 5, 241. [Google Scholar] [CrossRef]
- Fiuza, J.A.; Gannavaram, S.; Santiago, H. da C.; Selvapandiyan, A.; Souza, D.M.; Passos, L.S.A.; de Mendonça, L.Z.; Lemos-Giunchetti, D. da S.; Ricci, N.D.; Bartholomeu, D.C.; et al. Vaccination Using Live Attenuated Leishmania Donovani Centrin Deleted Parasites Induces Protection in Dogs against Leishmania Infantum. Vaccine 2015, 33, 280–288. [Google Scholar] [PubMed]
- Seyed, N.; Rafati, S. Th1 Concomitant Immune Response Mediated by IFN-γ Protects against Sand Fly Delivered Leishmania Infection: Implications for Vaccine Design. Cytokine 2021, 147, 155247. [Google Scholar] [CrossRef] [PubMed]
- Dayakar, A.; Chandrasekaran, S.; Kuchipudi, S.V.; Kalangi, S.K. Cytokines: Key Determinants of Resistance or Disease Progression in Visceral Leishmaniasis: Opportunities for Novel Diagnostics and Immunotherapy. Front. Immunol. 2019, 10, 670. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, E.M.; Bacellar, O.; Brownell, C.; Regis, T.; Coffman, R.L.; Reed, S.G. Restoration of IFN-Gamma Production and Lymphocyte Proliferation in Visceral Leishmaniasis. J. Immunol. 1994, 152, 5949–5956. [Google Scholar] [CrossRef]
- Taylor, A.P.; Murray, H.W. Intracellular Antimicrobial Activity in the Absence of Interferon-Gamma: Effect of Interleukin-12 in Experimental Visceral Leishmaniasis in Interferon-Gamma Gene-Disrupted Mice. J. Exp. Med. 1997, 185, 1231–1239. [Google Scholar] [CrossRef]
- Nathan, C.F.; Hibbs, J.B., Jr. Role of Nitric Oxide Synthesis in Macrophage Antimicrobial Activity. Curr. Opin. Immunol. 1991, 3, 65–70. [Google Scholar] [CrossRef]
- Hart, P.H.; Whitty, G.A.; Piccoli, D.S.; Hamilton, J.A. Control by IFN-Gamma and PGE2 of TNF Alpha and IL-1 Production by Human Monocytes. Immunology 1989, 66, 376–383. [Google Scholar]
- de Waal Malefyt, R.; Haanen, J.; Spits, H.; Roncarolo, M.G.; te Velde, A.; Figdor, C.; Johnson, K.; Kastelein, R.; Yssel, H.; de Vries, J.E. Interleukin 10 (IL-10) and Viral IL-10 Strongly Reduce Antigen-Specific Human T Cell Proliferation by Diminishing the Antigen-Presenting Capacity of Monocytes via Downregulation of Class II Major Histocompatibility Complex Expression. J. Exp. Med. 1991, 174, 915–924. [Google Scholar] [CrossRef]
- Kelly, C.D.; Welte, K.; Murray, H.W. Antigen-Induced Human Interferon-Gamma Production. Differential Dependence on Interleukin 2 and Its Receptor. J. Immunol. 1987, 139, 2325–2328. [Google Scholar] [CrossRef]
- Smith, K.A. Interleukin-2: Inception, Impact, and Implications. Science 1988, 240, 1169–1176. [Google Scholar] [CrossRef]
- Titus, R.G.; Sherry, B.; Cerami, A. Tumor Necrosis Factor Plays a Protective Role in Experimental Murine Cutaneous Leishmaniasis. J. Exp. Med. 1989, 170, 2097–2104. [Google Scholar] [CrossRef] [PubMed]
- Manetti, R.; Parronchi, P.; Giudizi, M.G.; Piccinni, M.P.; Maggi, E.; Trinchieri, G.; Romagnani, S. Natural Killer Cell Stimulatory Factor (interleukin 12 [IL-12]) Induces T Helper Type 1 (Th1)-Specific Immune Responses and Inhibits the Development of IL-4-Producing Th Cells. J. Exp. Med. 1993, 177, 1199–1204. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T.R. Cytokine Secretion Patterns and Cross-Regulation of T Cell Subsets. Immunol. Res. 1991, 10, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Holaday, B.J.; Pompeu, M.M.; Jeronimo, S.; Texeira, M.J.; Sousa, A. de A.; Vasconcelos, A.W.; Pearson, R.D.; Abrams, J.S.; Locksley, R.M. Potential Role for Interleukin-10 in the Immunosuppression Associated with Kala Azar. J. Clin. Invest. 1993, 92, 2626–2632. [Google Scholar] [CrossRef]
- Corraliza, I.M.; Soler, G.; Eichmann, K.; Modolell, M. Arginase Induction by Suppressors of Nitric Oxide Synthesis (IL-4, IL-10 and PGE2) in Murine Bone-Marrow-Derived Macrophages. Biochem. Biophys. Res. Commun. 1995, 206, 667–673. [Google Scholar] [CrossRef]
- Omer, F.M.; Kurtzhals, J.A.; Riley, E.M. Maintaining the Immunological Balance in Parasitic Infections: A Role for TGF-Beta? Parasitol. Today 2000, 16, 18–23. [Google Scholar] [CrossRef]
- Iniesta, V.; Gómez-Nieto, L.C.; Corraliza, I. The Inhibition of Arginase by N(omega)-Hydroxy-L-Arginine Controls the Growth of Leishmania inside Macrophages. J. Exp. Med. 2001, 193, 777–784. [Google Scholar] [CrossRef]
- Brombacher, F. The Role of Interleukin-13 in Infectious Diseases and Allergy. Bioessays 2000, 22, 646–656. [Google Scholar] [CrossRef]
- Tripathi, P.; Singh, V.; Naik, S. Immune Response to Leishmania: Paradox rather than Paradigm. FEMS Immunol. Med. Microbiol. 2007, 51, 229–242. [Google Scholar] [CrossRef]
- Coutinho De Oliveira, B.; Duthie, M.S.; Alves Pereira, V.R. Vaccines for Leishmaniasis and the Implications of Their Development for American Tegumentary Leishmaniasis. Hum. Vaccin. Immunother. 2020, 16, 919–930. [Google Scholar] [CrossRef]
- Sacks, D.; Kamhawi, S. Molecular Aspects of Parasite-Vector and Vector-Host Interactions in Leishmaniasis. Annu. Rev. Microbiol. 2001, 55, 453–483. [Google Scholar] [CrossRef]
- Melikishvili, M. KEY IMMUNOLOGICAL CONSIDERATIONS FOR LEISHMANIASIS VACCINE DEVELOPMENT. 2023.
- Tlili, A.; Marzouki, S.; Chabaane, E.; Abdeladhim, M.; Kammoun-Rebai, W.; Sakkouhi, R.; Belhadj Hmida, N.; Oliveira, F.; Kamhawi, S.; Louzir, H.; et al. Phlebotomus Papatasi Yellow-Related and Apyrase Salivary Proteins Are Candidates for Vaccination against Human Cutaneous Leishmaniasis. J. Invest. Dermatol. 2018, 138, 598–606. [Google Scholar] [CrossRef]
Table 1.
Some types of vaccines against Leishmaniasis.
| Type of vaccine |
Reference |
| Killed parasite antigen |
[64] |
| Oral immunization using live Lactococcus lactis co-expressing LACK and IL-12 | [65,66] |
| Live attenuated Centrin gene-deleted Leishmania vaccine | [67] |
| Live attenuated L. major parasites with p27 gene deletion | [68] |
| Therapeutic vaccine candidate using Leishmania antigens (SLA) and Toll-like receptor (TLR) 7/8 agonist (R848) encapsulated into the poly (lactic-co-glycolic acid) (PLGA) nanoparticles (NP) | [69] |
| Recombinant protein derived from sandfly saliva against Leishmania infection | [70,71,72,73,74,75,76,77,78,79,80,81,82,83] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



