
Submitted:
27 June 2024
Posted:
27 June 2024
You are already at the latest version
Abstract
Cucumber (Cucumis sativus L.) is a vital economic vegetable crop, and TRM (TONNEAU1 Recruiting Motif) gene plays a key role in cucumber organ growth. However, the pan-genomic characteristics of the TRM gene family and their expression patterns under different stresses have not been reported in cucumber. In this study, we identified 29 CsTRMs from the pan-genomes of 13 cucumber accessions, with CsTRM29 existing only in PI183967. Most CsTRM proteins exhibited differences in sequence length, except five CsTRMs having consistent protein sequence lengths among the 13 accessions. All CsTRM proteins showed amino acid variations. Analysis of CsTRM gene expression patterns revealed that six CsTRM genes strongly changed in short-fruited lines compared with long-fruited lines. And four CsTRM genes strongly responded to salt and heat stress, while CsTRM14 showed responses to salt stress, powdery mildew, gray mold and downy mildew. Some CsTRM genes were induced or suppressed at different treatment timepoint, suggesting that cucumber TRM genes may play different roles in responses to different stresses, with expression patterns varying with stress changes. Remarkably, the expression of CsTRM21 showed considerable change between long and short fruit and in responses to abiotic stresses (salt stress, heat stress) as well as biotic stresses (powdery mildew, gray mold), suggesting a dual role of CsTRM21 in both fruit shape determination and stress resistance. Collectively, this study provided a base for further functional identification of CsTRM genes in cucumber plant growth and stress resistance.
Keywords:
pan-genome
; cucumber
; TRM
; fruit shape
; abiotic stress response
; biotic stress response
1. Introduction
TRM (TONNEAU1 Recruiting Motif) family genes play important roles in the growth and development of plants, exerting significant functions in various plant species. In Arabidopsis, 34 TRM proteins were identified, and half of them are putative microtubule-associated proteins [1]. AtTRM1 and AtTRM2 regulate leaf morphology by positively promoting longitudinal polar cell elongation [2]. The Attrm5 mutant causes slow leaf growth, delayed flowering, and shortened root length [3]. AtTRM61, has a conserved functional structure and possesses conserved binding motifs for cofactor S-adenosyl-L-methionine (AdoMet), affects embryo arrest and seed abortion [4]. Additionally, TRMs can interact with TON1(TONNEAU1) and PP2A (Protein Phosphatase 2A) through their M2 and M3 domains, respectively, forming the TTP (TON1-TRM-PP2A) protein complex. This complex is targeted to microtubules (MT) [5], regulating microtubule organization and preprophase band (PPB) formation, thus influencing cell division and/or growth. This regulation ultimately affects the size and shape of plant organs [5,6,7,8,9,10]. In tomatoes, TRMs can interact with OFPs through their M8 domain. The OFP-TRM protein complex undergoes relocalization between the cytoplasm and microtubules, maintaining dynamic balance to regulate cell division and organ growth, ultimately affecting fruit shape [11,12]. SlTRM5 positively regulates fruit elongation by influencing cell division [13]. In the LA1589 background, although SlTRM3/4 minimally influenced fruit shape, the absence of SlTRM5 led to a slight flattening of the fruit [14]. The fruit shape of the double mutant lacking both SlTRM3/4 and SlTRM5 closely resembles that of the single mutant lacking only SlTRM5 [14]. Introducing the non-functional versions of either SlTRM3/4 or SlTRM5 into ovate/sov1 near-isogenic lines (NILs) partially restored the pear shape of the fruit. Moreover, when both non-functional alleles of SlTRM3/4 and SlTRM5 were combined in ovate/sov1 NILs, the fruit shape index (FSI) was similar to that of wild-type (WT) fruits [14,15], indicating the additive effects of SlTRM3/4 and SlTRM5 in regulating fruit elongation. Fruit shape analyses of the null mutants of SlTRM17/20a, SlTRM19, or SlTRM26a in the LA1589 background, generated using CRISPR/Cas9, revealed an interesting finding. It suggested that SlTRM17/20a and SlTRM19 work together synergistically to regulate fruit elongation, while SlTRM26a has a minor effect on fruit shape. The null alleles of SlTRM5 and SlTRM19, whether in the LA1589 or ovate/sov1 backgrounds, were observed to counterbalance each other in the regulation of fruit elongation. This suggests that SlTRM5 and SlTRM19 have opposing effects on fruit elongation [14]. In rice, the TRM homologous genes OsGW7/GL7/SLG7 interact with TON1 and PP2A through their M2 and M3 domains, respectively, and target them to the cortical microtubules. By influencing cell length and width, they regulate grain size and quality [16,17,18]. In cucumber, CsTRM5 affects fruit shape by influencing the direction of cell division and cell expansion. Additionally, ABA participates in regulating cucumber fruit elongation through CsTRM5-mediated cell expansion [19].
TRM gene family members are often localized to microtubules [2,5,13]. Microtubules are crucial components of the plant cell skeleton, and they play vital roles in maintaining cell shape, adapting to growth, development, and environmental changes, as well as in processes such as cell division, intracellular transport, immune responses, and stress tolerance [20,21,22,23,24,25,26,27,28,29]. MICROTUBULE-DESTABILIZING PROTEIN 25 (MDP25) is a hydrophilic cation-binding protein of the plant-specific developmentally regulated plasma membrane polypeptide(DREPP) family [30]. It is postulated that AtMDP25 similarly modulates stomatal closure, root hydrotropic response, and immune responses by influencing microtubule dynamics [31,32,33]. OsDREPP2 exhibits an affinity for microtubules and, in vitro, it inhibits microtubule polymerization [34], and MtDREPP induces the fragmentation of microtubules within membrane nanodomains during rhizobial infections [35]. Ethylene signaling regulates microtubule reassembly by up-regulating microtubule-stabilizing protein WAVE-DAMPENED2-LIKE5 (WDL5) expression in response to salt stress [36]. Katanin1 (KTN1) acts as a microtubule-severing protein, aiding in the maintenance of the organized microtubule structure. Under hypersalinity, the microtubule-associated protein KTN1 regulates hypersalinity-induced microtubule disassembly/assembly, thereby enhancing salinity tolerance [37]. Microtubules under high temperature stress undergo depolymerization [38]. High temperature stress (35 °C–37 °C) primarily disrupts the formation of excessive microtubule-organizing centers, which bind to the minus end of microtubules, consequently regulating their elongation and the shortening of microtubule arrays [39]. The changes in microtubule dynamics impact vesicular transport, protein trafficking, and cell wall deposition [40,41,42,43]. Currently, there are no reports on the involvement of cucumber TRM family genes in biotic or abiotic stress.
Pangenomics seeks to capture the full spectrum of genetic variation within a species through the assembly and comparative analysis of genome sequences from multiple individuals and displayed powerful potential in discovering novel genes or gene novel function [44]. In cucumber, a graph-based pan-genome was constructed based on 12 accessions [45], which provided a resource for characterizing variations of TRM proteins. In this study, we identified a total of 29 CsTRM genes in the pan-genome of cucumber and found that most of them vary in protein length between the 13 accessions, and all CsTRM proteins showed amino acid variations. In addition, we analyzed the expression patterns of the CsTRM genes using transcriptomic data in fruit and under different stresses that may play roles in different stresses. Therefore, our study provides a reference for investigating the potential role of TRMs for fruit shape and stress resistance in cucumber.
2. Materials and Method
2.1. Identification of TRM Genes in Cucumber
To identify TRM genes in cucumber, download the cucumber pan-genome assembly and annotation files from https://www.ncbi.nlm.nih.gov/, and the ‘PI183967’ genome assembly from http://www.cucurbitgenomics.org/. Use TBtools to extract CDS sequences and translate them into protein sequences. Retrieve AtTRM family members from https://www.arabidopsis.org/, and employ these sequences as queries in TBtools to predict TRM family members in cucumber. Perform a conserved motif analysis using the online MEME tool (https://www.omicsclass.com/article/432). Visualize the results with TBtools and screen for the final CsTRM family members based on the conserved M2 motif.
2.2. Protein Length, Motif Composition and Gene Structure Analysis
The protein sequences of CsTRMs in different cucumber accessions were extracted and the proteins’ lengths were counted using TBtools. The variation of amino acids was analyzed using DNAMAN program. The conserved motifs were identified using TBtools. The location information of CDSs and UTRs was extracted from the genomic annotation database and graphed using TBtools [46].
2.3. Gene Duplication and Synteny Analysis
The genomic databases of cucumber, Arabidopsis, rice, tomato, and maize were downloaded from http://cucurbitgenomics.org/organism/20 and http://plants.ensembl.org/index.html, and then, the gene duplication events and the syntenic relationships were obtained using the Multiple Collinearity Scan toolkit (MCScanX) [47] with the default parameters. The results were constructed using TBtools [46].
2.4. Transcriptome Analysis of CsTRM Genes in Friut
The publicly available transcriptomic data of cucumber fruit carpel numbers (SPR182933) [48], long fruit 408 and short fruit 409(SPR045470) [49], WT and CsFUL1A-OX-29(SPR117025) [50] were downloaded from NCBI (https://www.ncbi.nlm.nih.gov/geo/browse) to analyze the expression patterns of CsTRMs in fruit. The genome-wide expression of the CsTRMs gene was shown on a heatmap using TBtools [46]. For the transcriptome analysis of the CsTRMs, a threshold of FDR (or p-value) ≤ 0.05 and an absolute value of log2 (fold-change) ≥ 1 or log2 (fold-change) ≤ -1 were used to define DEGs.
2.5. Transcriptome Analysis of CsTRMs in Response to Abiotic and Biotic Stresses
The publicly available transcriptomic data of cucumber treated with salt (GSE116265) [51], heat (GSE151055) [52], PM (GSE81234) [53], GM (SRP062592) [54] and DM (SRP009350) [55] were downloaded from https://www.ncbi.nlm.nih.gov/ to analyze the expression patterns of CsTRMs under different stresses. After aligning the gene IDs to the cucumber genome, the genome-wide expression of the CsTRMs gene was shown on a heatmap using TBtools [46]. For the transcriptome analysis of the CsTRMs, a threshold of FDR (or p-value) ≤ 0.05 and an absolute value of log2 (fold-change) ≥ 1 or log2 (fold-change) ≤ -1 were used to define DEGs.
3. Result
3.1. Identification of CsTRM Genes Based on the Cucumber Pan-Genome
To investigate the variation of the TRM genes across cucumber accessions, we identified CsTRM genes from the pan-genome including 13 cucumber accessions [45]. A total of 29 putative TRM genes were identified among the genomes of the 13 cucumber accessions (Table 1, Table S1). We renamed them CsTRM01-CsTRM29 based on their order on the chromosomes to avoid confusion in this study (Table 1). Additionally, CsTRM04 exhibits multiple copies in W4. There were 28 CsTRM genes identified from ‘9930’, being consistent with the previous study [56], and from PI183967, lacking CsTRM03 and possessing a unique CsTRM29 (Table 1); 27 from ‘Cu2’, ‘Cuc64’, ‘W4’, ‘Hx14’, ‘Hx17’ ‘Cuc37’, ‘Gy14’ and ‘9110gt’; 26 from ‘XTMC’; 25 from ‘Cuc80’ and ‘W8’ (Table 1). CsTRM01, 02, 06, 07, 09, 11, 12, 13, 14, 15, 17, 18, 19, 21, 23, 24, 25, 26, 27 and 28, all are present in the 13 cucumber accessions. CsTRM3 is absent in Cuc80 and PI; CsTRM04 is absent in Cuc80 and W8; CsTRM05 is absent in XTMC and W8; CsTRM08 is absent in XTMC; CsTRM10 is absent in Cu2 and Cuc80; CsTRM16 is absent in Cuc64, W4, W8, Hx14, Hx17 and Cuc37; CsTRM20 is absent in Gy14; CsTRM22 is absent in 9110gt. CsTRM29 only existing in PI183967 is identified as a new member of the CsTRM gene family in the 13 cucumber accessions
3.2. Analysis of Protein Length and Amino Acid Variations in the CsTRM Proteins
To further understand protein length variation of CsTRMs among the cucumber accessions, we showed the length of the identified CsTRM proteins in Table 2. There were 5 CsTRMs with the same protein length among 13 cucumber accessions, namely, CsTRM04, 11, 14, 15 and 21. The length of CsTRM01, 02, 05, 06, 13, 18, 22, 24 and 26 differed in only one of the accessions. And others showed differences in protein length among multiple accessions. Length difference data are marked in red in Table 2.
Among the proteins with different lengths, CsTRM01 in ‘W4’; CsTRM02 in‘9930’; CsTRM03 in ‘9930’; CsTRM05 in ‘9930’; CsTRM07 in ‘XTMC’; CsTRM09 in ‘9930’; CsTRM13 in ‘Gy14’; CsTRM18 in ‘Cuc64’; CsTRM19 in ‘9930’ and ‘XTMC’; CsTRM20 in ‘Cuc37’; CsTRM22 in ‘Gy14’; CsTRM23 and CsTRM24 in ‘9110gt’; CsTRM25 in ‘PI183967’; CsTRM27 in ‘Hx117’ and CsTRM28 in ‘Hx14’ had shorter lengths compared to those in other accessions, while CsTRM16 in ‘Cu2’, ‘Cuc80’ and ‘Gy14’; CsTRM19 in ‘Cu2’, ‘Cuc80’ and ‘Cuc64’ and CsTRM26 in ‘W4’ were longer than those in other accessions. In addition, the lengths of some proteins showed multiple polymorphism. For example, the protein length of CsTRM17 was the same in ‘9930’, ‘Cu2’, ‘Cuc80’, ‘PI’, ‘Cuc64’ and ‘Cuc37’, but totally different in other accessions, furthermore, dramatically shortened in ‘W4’, ‘W8’, ‘Hx14’, ‘Hx117’ and ‘9110gt’ (Table 2).
Besides protein length, amino acid substitution also can change a protein’s function [57]. The amino acid variations of CsTRMs in different cucumber accessions was analyzed (Table S1). Amino acid variations were annotated using CsTRMs protein sequence of 9930 as reference, and all CsTRM proteins exhibit amino acid variations. CsTRM04, 11, 14, 15 and 21 have 6, 5, 3, 7 and 2 amino acid variations, respectively, but these do not lead to changes in protein length (Table 2). Some CsTRMs exhibit amino acid insertions leading to an increase in protein length. For example, CsTRM02 has 27 amino acid insertions in accessions other than 9930. In CsTRM06, 17, 26, 27 and 28, there are frame shift leading to amino acid variations. Some amino acid variations are quite significant, such as CsTRM07, 17, and 24.
Some amino acid variations are quite significant, such as CsTRM07, 17, and 24 (Table 2). Further comparisons of CsTRM07, 17, and 24 gene structures and gene conservative motifs (Figure 1). CsTRM07 in XTMC has only 478 amino acids, which is significantly shorter than that in the other 12 accessions (Table 2), and its gene structure underwent changes along with alterations in some conserved motifs, experiencing an increase in gene length, but not leading to the loss of conserved motifs in Gy14 and PI183967 (Figure 1A). For CsTRM17, the protein length varied from 210 amino acids to 1091 amino acids across the 13 accessions (Table 2), with corresponding changes in gene structure and some conserved motifs, especially in W4, there are only two conserved motifs (Figure 1B). In CsTRM24 of 9110gt, alterations in gene structure caused the decreased protein length, but without a reduction in conserved motifs.
3.3. Synteny Analysis of CsTRM Genes
The phylogenetic relationship of the cucumber TRM family were further explored by constructing comparative syntenic maps of cucumber associated with four representative species, including two monocots (rice and maize) and two dicots (Arabidopsis and tomato) (Figure 2). 1, 3, 8, and 19 CsTRM genes showed syntenic relationships with those in the other four species: maize, rice, Arabidopsis, and tomato, respectively (Figure 2). Only 1 TRM collinear gene pairs between cucumber and maize were identified, followed by cucumber and rice (4), cucumber and Arabidopsis (9), and cucumber and tomato (20) (Table S2). It is evident that dicotyledonous plants exhibit a notably higher number of homologous genes compared to those shared between dicotyledonous and monocotyledonous plants. This observation aligns with the patterns expected in biological evolution. CsTRM18 and its collinear gene pairs with maize are observed in rice and tomato, but not in Arabidopsis, indicating differences in the evolutionary process of CsTRM18. Additionally, collinear gene pairs between cucumber and rice, maize, and Arabidopsis are observed in cucumber and tomato, suggesting that cucumber and tomato may have undergone a common evolutionary history.
3.4. Expression Profiles of CsTRM Genes in the Fruit
In cucumber and tomato, some TRM genes can regulate fruit shape [14,15,20]. To investigate the function of CsTRMs in fruit shape, we conducted expression analysis of CsTRMs using published RNA-seq data on fruits with different carpel numbers and lengths [48,49,50]. Relative to the South China type cucumber 32X (carpel number = 3), the transcription levels of CsTRMs in the mutant Gui Fei Cui (GFC, carpel number = 5) from 32X showed no significant changes (Figure 3A, Table S3), indicating that CsTRMs might not play a crucial role in regulating cucumber fruit carpel number. Compared to long fruit 408, there were 8 genes down-regulated in short fruit 409, namely CsTRM5, 6, 10, 11, 14, 21, 26 and 27 (Figure 3B). Compared to empty vector/control transgenic plants WT, CsFUL1A-OX-29 had a total of 12 genes down-regulated, namely CsTRM1, 2, 5, 6, 7, 10, 12, 13, 16, 20, 21 and 27; and 4 genes up-regulated, namely CsTRM8, 17, 25, 26 (Figure 3C). In CsFUL1A-OX-29 versus empty vector/control plants and 409 versus 408, CsTRM5, 6, 10, 21, and 27 were significantly down-regulated (Figure 3B,C), indicating that these genes play a crucial role in regulating fruit shape. However, the expression trend of CsTRM26 in the two groups of long and short fruit materials is opposite (Figure 3B,C), which may be due to different genetic backgrounds of the materials.
3.5. Expression Profiles of CsTRM Genes under Abiotic and Biotic Stresses
TRM gene family members are often localized to microtubules, microtubules are involved in immune responses and stress tolerance. We analyzed the comprehensive expression patterns of CsTRM genes under different stresses, including salt, heat, downy mildew (DM, Pseudoperonospora cubensis), gray mold (GM, Botrytis cinerea) and powdery mildew (PM, Podosphaera fusca) based on public transcriptome information [51,52,53,54,55], to further explore the roles of CsTRM genes under different stresses.
First, we analyzed the roles of CsTRM genes under salt stress (Table S4). The transcriptomic data were presented as a heatmap (Figure 4A). We observed that the expression levels of CsTRM4, 8 and 14 considerably increased in response to NaCl stress, and four genes exhibited the opposite trend with exposure to NaCl stress, they are CsTRM5, 11, 21 and 24 (Figure 4A). Under the conditions treated with Silicon (Si) only, the expression of CsTRM3 and CsTRM14 was upregulated, whereas the expression of CsTRM11, 21 and 24 was downregulated. The expression of CsTRM14 was upregulated under both individual NaCl treatment and individual Si treatment, while the expression of CsTRM11, 21 and 24 was downregulated. Previous research has demonstrated that the application of Silicon (Si) can enhance plant growth when subjected to salt stress. After treatment with Si, the gene expression levels of CsTRM11, 14 and 24, which exhibited significant changes under salt stress, returned to normal levels; CsTRM5, 8 and 21 showed only slight regression, while the expression level of upregulated CsTRM4 showed a slight increase. We also analyzed the responses of CsTRM genes to heat stress (Figure 4B, Table S4). At three hours after high-temperature treatment, CsTRM1, 11, 16, 18, 21, 22 and 26 were downregulated, while CsTRM3, 8 and 20 were upregulated. At six hours after heat stress, the expression of CsTRM16 and CsTRM22 showed no significant difference compared to the 0 hour heat treatment, while the changes in other differentially expressed genes were consistent with the 3 hour heat treatment. Specifically, the genes upregulated at three and six hours after heat stress were nearly identical (Figure 4B), indicating their potential significant roles in conferring thermostolerance. Additionally, CsTRM3, 8, 11 and 21 were differentially expressed in responding to the treatments of heat and NaCl, with consistent trends.
To explore the potential functions of CsTRMs in the resistance to biotic stresses, we performed expression analyses of CsTRMs using the published RNA-Seq data of cucumber seedlings inoculated with PM for 48 h, GM for 96 h and with DM for 8 days [53,54,55]. After inoculation with PM, a total of 4 genes were differentially expressed in the susceptible cucumber line D8 leaves compared with the control, the expression of CsTRM14, 21 and 27 were upregulated, while CsTRM20 were downregulated; and a total of 4 genes were differentially expressed in the resistant cucumber line SSL508-28 leaves compared with the control, the expression of CsTRM4, 14 and 27 were upregulated, while CsTRM21 were downregulated (Figure 5A). In the susceptible and the resistant cucumber line affected by PM, CsTRM14 and CsTRM27 had similar expression trends, while CsTRM21 had opposite expression trends (Figure 5A). After 96 hours of GM inoculation, cucumber seedlings showed significant downregulation of 14 CsTRM genes compared to the uninoculated control, namely CsTRM1, 2, 5, 6, 7, 10, 11, 13, 14, 16, 20, 21, 27 and 28, and significant upregulation of 3 genes, namely CsTRM3, 18 and 26 (Figure 5B). In the transcriptomic data from cucumber seedlings inoculated with DM, only 5 TRMs genes exhibited significant changes in expression (Figure 5C). CsTRM1, 7, 14 and 28 were upregulated at a minimum of one treatment timepoint, while CsTRM 8 were downregulated at 6 days post inoculation (dpi) and 8 dpi. CsTRM28 were upregulated at 2 dpi, 3 dpi, 4 dpi, 6 dpi and 8 dpi (Figure 5C), indicating its significant role in responding to the DM. In summary, the expression of CsTRM14 was significantly upregulated in cucumber seedlings inoculated with PM, BC, and DM, indicating its broad-spectrum role in responding to biotic stress.
4. Discussion
Researches demonstrated that a single reference genome is inadequate for capturing the diversity within a species [58]. Hence, we conducted a comprehensive analysis to identify and characterize the TRM family in 13 different cucumber varieties. Although in the previous study, 28 TRM family were identified [56], in this study, a novel member, CsTRM29 which is only present in PI183967, was discovered (Table 1). Moreover, only 5 CsTRMs have the same protein length among 13 cucumber accessions, and all the identified TRM proteins have amino acid variations including insertions, deletions, single amino acid changes and frame shifts (Table S1). Some CsTRMs underwent changes not only in gene structure but also in conserved motifs (Figure 1). Therefore, in this study, we found rich variations occurred in CsTRMs from the pan-genomes of 13 cucumber accessions, and these variations will provide a base for discovering TRM genes with novel functions, which will accelerate the breeding of new cucumber varieties, just as the things are performed with pan-genomics [44].
It is widely recognized that there exists a correlation between gene expression and gene function. The cucumber fruit typically have three fused carpels [59], the carpel number is an important fruit trait that affects fruit shape, size and internal quality [48]. In the lines with different carpel numbers, there were no significant differences observed in the expression of CsTRMs (Figure 3A), suggesting that CsTRMs might not play a critical role in regulating the number of carpels in cucumber fruits. However, in the short-fruited lines (409 and CsFUL1A-OX-29), CsTRM5, 6, 10, 21, and 27 were significantly down-regulated (Figure 3B,C), indicating that these genes might play crucial roles in regulating cucumber fruit length. Interestingly, the expression of CsTRM26 is lower in the short-fruited line 409 than in the long-fruited line 408, but higher in the short-fruited line CsFUL1A-OX-29 than in the wild type. This could be due to differing genetic backgrounds or the possibility that CsTRM26 does not regulate cucumber fruit length.
So far, TRMs have been reported to be functional in plant organ growth, but not in plant response to stresses. But an increasing number of researches suggested that apart from their crucial roles in mechanical architecture and cell division, microtubules are also implicated in plants adaptation to severe environmental conditions [60]. Since some TRMs are microtubule-binding proteins, they might participate in stress responses. Therefore, in this study, we analyzed the expression patterns of CsTRMs under certain stress conditions. Many CsTRM genes showed expression changes at varying degrees under different stress conditions (Figure 4 and Figure 5). Under salt and heat stress conditions, the expression of CsTRM3 and CsTRM8 was significantly upregulated, while CsTRM11 and CsTRM21 were significantly downregulated (Figure 4), however, under inoculation with PM, BC, or DM, the expression of CsTRM14 was significantly increased, while the expression of CsTRM21 showed significant changes after inoculation with PM and BC (Figure 5). These results might indicate that different CsTRMs respond to abiotic or biotic stresses. Remarkably, CsTRM21 plays a crucial role in regulating fruit shape (Figure 3B,C) and in responding to biotic stresses (Figure 4 and Figure 5). Therefore, this study provided not only a base for the function of CsTRMs in stress tolerance, but also a cross talk point between organ growth and biotic stresses.
5. Conclusions
In this study, we performed pan-genome-wide identification of the TRM gene family in cucumber. In total, 29 members were identified, including a novel member, CsTRM29 which is only present in PI183967. Only 5 of the CsTRMs have consistent protein lengths among the 13 accessions. All CsTRM proteins showed amino acid variations. Furthermore, Transcriptomic data of fruits with different shapes indicate that CsTRMs play a significant role in regulating fruit length but not in controlling carpel number. And transcriptomic data under different stress conditions revealed the differences and similarities in the stress-induced expression of CsTRMs in response to abiotic and biotic stresses, and CsTRM14 was found to response to salt stress, powdery mildew, gray mold and downy mildew. Notably, CsTRM21 plays a role in regulating both fruit shape and resistance. In conclusion, this study provides a reference for investigating the potential role of TRMs for fruit shape and stress resistance in cucumber.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Z.R., L.W. and C.C.; methodology, L.Z. and K.W.; validation, L.Z.; software, L.Z., K.W., Z.W. and S.C.; formal analysis, L.Z. and K.W.; investigation, L.Z. and K.W.; data curation, L.Z. and K.W.; writing-original draft preparation, L.Z.; writing-review and editing, Z.R.; supervision, L.W. and Z.R.; project administration, Z.R., L.W. and C.C.; funding acquisition, Z.R. and L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by fundings from the National Natural Science Foundation of China (32172605 and 31972419) and the ‘Taishan Scholar’ Foundation of the People’s Government of Shandong Province (ts20130932).
Data Availability Statement
The data presented in this study are available in this article and Supplementary Materials.
Acknowledgments
We extend our appreciation to the anonymous reviewers for their valuable suggestions to help improve this article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Drevensek, S.; Goussot, M.; Duroc, Y.; Christodoulidou, A.; Steyaert, S.; Schaefer, E.; Duvernois, E.; Grandjean, O.; Vantard, M.; Bouchez, D.; Pastuglia, M. The Arabidopsis TRM1-TON1 interaction reveals a recruitment network common to plant cortical microtubule arrays and eukaryotic centrosomes. The Plant Cell 2012, 24, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Kim, G.T.; Kim, I.J.; Park, J.; Kwak, S.S.; Choi, G.; Chung, W.I. LONGIFOLIA1 and LONGIFOLIA2, two homologous genes, regulate longitudinal cell elongation in Arabidopsis. Development 2006, 133, 4305–4314. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.Q.; Ng, P.Q.; Shi, S.S.; Fan, D.; Li, J.; Zhao, J.; Wang, H.; David, R.; Mittal, P.; Do, T.; Bock, R.; Zhao, M.; Zhou, W.B.; Searle, I. Arabidopsis TRM5 encodes a nuclear-localised bifunctional tRNA guanine and inosine-N1-methyltransferase that is important for growth. PLoS ONE 2019, 14, e0225064. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Jia, P.; Xin, P.; Chu, J.; Shi, D.Q.; Yang, W.C. The Arabidopsis TRM61/TRM6 complex is a bona fide tRNA N1-methyladenosine methyltransferase. Journal of experimental botany 2020, 71, 3024–3036. [Google Scholar] [CrossRef] [PubMed]
- Drevensek, S.; Goussot, M.; Duroc, Y.; Christodoulidou, A.; Steyaert, S.; Schaefer, E.; Duvernois, E.; Grandjean, O.; Vantard, M.; Bouchez, D.; Pastuglia, M. The Arabidopsis TRM1-TON1 interaction reveals a recruitment network common to plant cortical microtubule arrays and eukaryotic centrosomes. The Plant Cell 2012, 24, 178–191. [Google Scholar] [CrossRef]
- Camilleri, C.; Azimzadeh, J.; Pastuglia, M.; Bellini, C.; Grandjean, O.; Bouchez, D. The Arabidopsis TONNEAU2 gene encodes a putative novel protein phosphatase 2A regulatory subunit essential for the control of the cortical cytoskeleton. The Plant Cell 2002, 14, 833–845. [Google Scholar] [CrossRef]
- Azimzadeh, J.; Nacry, P.; Christodoulidou, A.; Drevensek, S.; Camilleri, C.; Amiour, N.; Parcy, F.; Pastuglia, M.; Bouchez, D. Arabidopsis TONNEAU1 proteins are essential for preprophase band formation and interact with centrin. The Plant Cell 2008, 20, 2146–2159. [Google Scholar] [CrossRef]
- Spinner, L.; Gadeyne, A.; Belcram, K.; Goussot, M.; Moison, M.; Duroc, Y.; Eeckhout, D.; De Winne, N.; Schaefer, E.; Van De Slijke, E.; Persiau, G.; Witters, E.; Gevaert, K.; Jaeger, G.D.; Bouchez, D.; Van Damme, D.; Pastuglia, M. A protein phosphatase 2A complex spatially controls plant cell division. Nature Communications 2013, 4, 1863. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, E.; Belcram, K.; Uyttewaal, M.; Duroc, Y.; Goussot, M.; Legland, D.; Laruelle, E.; Tauzia-Moreau, M.D.; Pastuglia, M.; Bouchez, D. The preprophase band of microtubules controls the robustness of division orientation in plants. Science 2017, 356, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Q.; Chen, B.Q.; Dang, X.; Zhu, L.L.; Rao, J.Q.; Ren, H.B.; Lin, C.T.; Qin, Y.; Lin, D.S. Arabidopsis IPGA1 is a microtubule-associated protein essential for cell expansion during petal morphogenesis. Journal of Experimental Botany 2019, 70, 5231–5243. [Google Scholar] [CrossRef]
- Van der Knaap, E.; Chakrabarti, M.; Chu, Y.H.; Clevenger, J.P.; Illa-Berenguer, E.; Huang, Z.J.; Keyhaninejad, N.; Mu, Q.; Sun, L.; Wang, Y.P.; Wu, S. What lies beyond the eye: the molecular mechanisms regulating tomato fruit weight and shape. Frontiers in Plant Science 2014, 5, 227. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, B.Y.; Keyhaninejad, N.; Rodríguez, G.R.; Kim, H.J.; Chakrabarti, M.; Illa-Berenguer, E.; Taitano, N.K.; Gonzalo, M.J.; Díaz, A.; Pan, Y.P.; Leisner, C.P.; Halterman, D.; Buell, C.R.; Weng, Y.Q.; Jansky, S.H.; van Eck, H.; Willemsen, J.; Monforte, A.J.; Meulia, T.; van der Knaap, E. A common genetic mechanism underlies morphological diversity in fruits and other plant organs. Nature Communications 2018, 9, 4734. [Google Scholar] [CrossRef] [PubMed]
- Lazzaro, M.D.; Wu, S.; Snouffer, A.; Wang, Y.P.; Van der Knaap, E. Plant organ shapes are regulated by protein interactions and associations with microtubules. Front Plant Science 2018, 9, 1766. [Google Scholar] [CrossRef]
- Zhang, B.; Li, Q.; Keyhaninejad, N.; Taitano, N.; Sapkota, M.; Snouffer, A.; van der Knaap, E. A combinatorial TRM-OFP module bilaterally fine-tunes tomato fruit shape. New Phytologist 2023, 238, 2393–2409. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, B.; Keyhaninejad, N.; Rodríguez, G.R.; Kim, H.J.; Chakrabarti, M.; Illa-Berenguer, E.; Taitano, N.K.; Gonzalo, M.J.; Díaz, A.; Pan, Y.; Leisner, C.P.; Halterman, D.; Buell, C.R.; Weng, Y.; Jansky, S.H.; van Eck, H.; Willemsen, J.; Monforte, A.J.; Meulia, T.; van der Knaap, E. A common genetic mechanism underlies morphological diversity in fruits and other plant organs. Nature communications 2018, 9, 4734. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.K.; Li, S.; Liu, Q.; Wu, K.; Zhang, J.Q.; Wang, S.S.; Wang, Y.; Chen, X.B.; Zhang, Y.; Gao, C.X.; Wang, F.; Huang, H.X.; Fu, X.D. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality. Nature Genetics 2015, 47, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Xiong, G.S.; Hu, J.; Jiang, L.; Yu, H.; Xu, J.; Fang, Y.X.; Zeng, L.J.; Xu, E.; Xu, J.; Ye, W.J.; Meng, X.B.; Liu, R.F.; Chen, H.Q.; Jing, Y.H.; Wang, Y.H.; Zhu, X.D.; Li, J.Y.; Qian, Q. Copy number variation at the GL7 locus contributes to grain size diversity in rice. Nature Genetics 2015, 47, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Miao, J.; Gu, H.Y.; Peng, X.R.; Leburu, M.; Yuan, F.H.; Gu, H.W.; Gao, Y.; Tao, Y.J.; Zhu, J.Y.; Gong, Z.Y.; Yi, C.D.; Gu, M.H.; Yang, Z.F.; Liang, G.H. Natural Variations in SLG7 Regulate Grain Shape in Rice. Genetics 2015, 201, 1591–1599. [Google Scholar] [CrossRef]
- Xie, Y.; Liu, X.F.; Sun, C.Z.; Song, X.F.; Li, X.L.; Cui, H.N.; Guo, J.Y.; Liu, L.; Ying, A.; Zhang, Z.Q.; Zhu, X.Y.; Yan, L.Y.; Zhang, X.L. CsTRM5 regulates fruit shape via mediating cell division direction and cell expansion in cucumber. Horticulture Research 2023, 10, uhad007. [Google Scholar] [CrossRef]
- Wade, R.H. On and around microtubules: an overview. Mol Biotechnol 2009, 43, 177–191. [Google Scholar] [CrossRef]
- Landrein, B.; Hamant, O. How mechanical stress controls microtubule behavior and morphogenesis in plants: history; experiments and revisited theories. Plant Journal 2013, 75, 324–338. [Google Scholar] [CrossRef] [PubMed]
- Nick, P. Microtubules, signalling and abiotic stress. Plant Journal 2013, 75, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Sampathkumar, A.; Yan, A.; Krupinski, P.; Meyerowitz, E.M. Physical forces regulate plant development and morphogenesis. Curr Biol 2014, 24, R475–R483. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, W. Regulation of developmental and environmental signaling by interaction between microtubules and membranes in plant cells. Protein Cell 2016, 7, 81–88. [Google Scholar] [CrossRef]
- Zhou, S.; Chen, Q.; Sun, Y.; Li, Y. Histone H2B monoubiquitination regulates salt stress-induced microtubule depolymerization in Arabidopsis. Plant Cell Environ 2017, 40, 1512–1530. [Google Scholar] [CrossRef]
- Yang, P.Z.; Jin, J.W.; Zhang, J.R.; Wang, D.; Bai, X.C.; Xie, W.F.; Hu, T.M.; Zhao, X.; Mao, T.L.; Qin, T. MDP25 mediates the fine-tuning of microtubule organization in response to salt stress. Journal of integrative plant biology 2022, 64, 1181–1195. [Google Scholar] [CrossRef]
- McNally, F.J.; Roll-Mecak, A. Microtubule-severing enzymes: From cellular functions to molecular mechanism. Journal of Cell Biology 2018, 217, 4057–4069. [Google Scholar] [CrossRef] [PubMed]
- Bao, Z.R.; Xu, Z.J.; Zang, J.Z.; Bürstenbinder, K.; Wang, P.W. The morphological diversity of plant organs: manipulating the organization of microtubules may do the trick. Frontiers in Cell and Developmental Biology 2021, 9, 691. [Google Scholar] [CrossRef]
- Bao, Z.R.; Guo, Y.; Deng, Y.L.; Zang, J.Z.; Zhang, J.H.; Deng, Y.T.; Ouyang, B.; Qu, X.L.; Bürstenbinder, K.; Wang, P.W. Microtubule-associated protein SlMAP70 interacts with IQ67-domain protein SlIQD21a to regulate fruit shape in tomato. The Plant Cell 2023, 35, 4266–4283. [Google Scholar] [CrossRef]
- Gantet, P.; Masson, F.; Domergue, O.; Marquis-Mention, M.; Bauw, G.; Inze, D.; Rossignol, M.; de la Serve, B.T.; et al. Cloning of a cDNA encoding a developmentally regulated 22 kDa polypeptide from tobacco leaf plasma membrane. Biochemistry and molecular biology international 1996, 40, 469–477. [Google Scholar] [CrossRef]
- Nagasaki-Takeuchi, N.; Miyano, M.; Maeshima, M. A plasma membrane-associated protein of Arabidopsis thaliana AtPCaP1 binds copper ions and changes its higher order structure. J Biochem 2008, 144, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Tanaka-Takada, N.; Kobayashi, A.; Takahashi, H.; Kamiya, T.; Kinoshita, T.; Maeshima, M. Plasma Membrane-Associated Ca2+-Binding Protein PCaP1 is Involved in Root Hydrotropism of Arabidopsis thaliana. Plant Cell Physiol 2019, 60, 1331–1341. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, M.; Marti, L.; Ferrari, S.; Tanaka-Takada, N.; Maeshima, M.; Ott, T.; De Lorenzo, G.; Mattei, B. The plasma membrane-associated Ca2+ -binding protein, PCaP1, is required for oligogalacturonide and flagellin-induced priming and immunity. Plant Cell Environment 2021, 44, 3078–3093. [Google Scholar] [CrossRef] [PubMed]
- Yamada, N.; Theerawitaya, C.; Kageyama, H.; Cha-Um, S.; Takabe, T. Expression of developmentally regulated plasma membrane polypeptide (DREPP2) in rice root tip and interaction with Ca(2+)/CaM complex and microtubule. Protoplasma 2015, 252, 1519–1527. [Google Scholar] [CrossRef]
- Su, C.; Klein, M.L.; Hernández-Reyes, C.; Batzenschlager, M.; Ditengou, F.A.; Lace, B.; Keller, J.; Delaux, P.M.; Ott, T. The Medicago truncatula DREPP Protein Triggers Microtubule Fragmentation in Membrane Nanodomains during Symbiotic Infections. The Plant Cell 2020, 32, 1689–1702. [Google Scholar] [CrossRef] [PubMed]
- Dou, L.; He, K.; Higaki, T.; Wang, X.; Mao, T. Ethylene Signaling Modulates Cortical Microtubule Reassembly in Response to Salt Stress. Plant Physiol 2018, 176, 2071–2081. [Google Scholar] [CrossRef]
- Yang, J.; An, B.; Luo, H.; He, C.; Wang, Q. AtKATANIN1 Modulates Microtubule Depolymerization and Reorganization in Response to Salt Stress in Arabidopsis. Int J Mol Sci 2019, 21, 138. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Jeevaraj, T.; Yunus, M.H.; Chakraborty, S.; Chakraborty, N. The plant cytoskeleton takes center stage in abiotic stress responses and resilience. Plant Cell Environ 2023, 46, 5–22. [Google Scholar] [CrossRef]
- Smirnova, E.A.; Bajer, A.S. Microtubule converging centers and reorganization of the interphase cytoskeleton and the mitotic spindle in higher plant Haemanthus. Cell Motility and the Cytoskeleton 1994, 27, 219–233. [Google Scholar] [CrossRef]
- Parrotta, L.; Faleri, C.; Cresti, M.; Cai, G. Heat stress affects the cytoskeleton and the delivery of sucrose synthase in tobacco pollen tubes. Planta 2016, 243, 43–63. [Google Scholar] [CrossRef]
- Parveen, S.; Rahman, A. Actin isovariant ACT7 modulates root thermomor-phogenesis by altering intracellular auxin homeostasis. International Journal of Molecular Sciences 2021, 22, 7749. [Google Scholar] [CrossRef] [PubMed]
- Pressman, E.; Harel, D.; Zamski, E.; Shaked, R.; Althan, L.; Rosenfeld, K.; Firon, N. The effect of high temperatures on the expression and activity of sucrose-cleaving enzymes during tomato (Lycopersicon esculentum) anther development. The Journal of Horticultural Science and Biotechnology 2006, 81, 341–348. [Google Scholar] [CrossRef]
- Zheng, Y.; Anderson, S.; Zhang, Y.; Garavito, R.M. The structure of sucrose synthase-1 from Arabidopsis thaliana and its functional implications. Journal of Biological Chemistry 2011, 286, 36108–36118. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, M.; Jayakodi, M.; Stein, N.; Mascher, M. Plant pangenomes for crop improvement; biodiversity and evolution. Nature reviews Genetics 2024, 10, 1038. [Google Scholar] [CrossRef]
- Wang, C.; Han, J.; Wang, T.; Chen, C.; Liu, J.; Xu, Z.; Zhang, Q.; Wang, L.; Ren, Z. Pan-Genome-Wide Identification and Transcriptome-Wide Analysis of DREB Genes That Respond to Biotic and Abiotic Stresses in Cucumber. Agriculture 2022, 12, 1879. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; Kissinger, J.C.; Paterson, A.H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Che, G.; Gu, R.; Zhao, J.; Liu, X.; Song, X.; Zi, H.; Cheng, Z.; Shen, J.; Wang, Z.; Liu, R.; Yan, L.; Weng, Y.; Zhang, X. Gene regulatory network controlling carpel number variation in cucumber. Development 2020, 147, dev184788. [Google Scholar] [CrossRef]
- Jing, L.; Yan, S.; Yang, W.; Li, Y.; Xia, M.; Chen, Z.; Wang, Q.; Yan, L.; Song, X.; Liu, R.; Zhang, X. Transcriptomic analysis reveals the roles of microtubule-related genes and transcription factors in fruit length regulation in cucumber (Cucumis sativus L.). Sci. Rep. 2015, 5, 8031. [Google Scholar] [CrossRef]
- Zhao, J.; Jiang, L.; Che, G.; Pan, Y.; Li, Y.; Hou, Y.; Zhao, W.; Zhong, Y.; Ding, L.; Yan, S.; Sun, C.; Liu, R.; Yan, L.; Wu, T.; Li, X.; Weng, Y.; Zhang, X. A Functional Allele of CsFUL1 Regulates Fruit Length through Repressing CsSUP and Inhibiting Auxin Transport in Cucumber. The Plant cell 2019, 31, 1289–1307. [Google Scholar] [CrossRef]
- Zhu, Y.; Yin, J.; Liang, Y.; Liu, J.; Jia, J. Transcriptomic dynamics provide an insight into the mechanism for silicon-mediated alleviation of salt stress in cucumber plants. Ecotoxicol Environ Saf 2019, 174, 245–254. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Tang, R.; Wang, L.; Chen, C.; Ren, Z. Genome-Wide identification and expression analysis of Hsf and Hsp gene families in cucumber (Cucumis sativus L.). Plant Growth Regul 2021, 95, 223–239. [Google Scholar] [CrossRef]
- Xu, Q.; Xu, X.; Shi, Y.; Qi, X.; Chen, X. Elucidation of the molecular responses of a cucumber segment substitution line carrying Pm5.1 and its recurrent parent triggered by powdery mildew by comparative transcriptome profiling. BMC Genom 2017, 18, 21. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Chen, N.; Liu, T.; Zhu, J.; Wang, J.; He, X.; Jin, Y. Large-Scale Transcriptome Analysis of Cucumber and Botrytis cinerea during Infection. PLoS One 2015, 10, e0142221. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B.N.; Savory, E.A.; Vaillancourt, B.; Childs, K.L.; Hamilton, J.P.; Day, B.; Buell, C.R. Expression Profiling of Cucumis sativus in Response to Infection by Pseudoperonospora cubensis. PLoS ONE 2012, 7, e34954. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, S.; Chai, S.; Yang, Z.; Zhang, Q.; Xin, H.; Xu, Y.; Lin, S.; Chen, X.; Yao, Z.; Yang, Q.; Fei, Z.; Huang, S.; Zhang, Z. Graph-based pan-genome reveals structural and sequence variations related to agronomic traits and domestication in cucumber. Nat Commun 2022, 13, 682. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hu, Y.J.; Zhang, Q.X.; Wang, L.N.; Ren, Z.H. Identification and Analysis on TRM Family in Cucumber. Journal of Shandong Agricultural University 2021, 52, 358–363. [Google Scholar]
- Yin, S.; Zhao, L.; Liu, J.; Sun, Y.; Li, B.; Wang, L.; Ren, Z.; Chen, C. Pan-genome Analysis of WOX Gene Family and Function Exploration of CsWOX9 in Cucumber. Int. J. Mol. Sci. 2023, 24, 17568. [Google Scholar] [CrossRef]
- Bai, S.L.; Peng, Y.-B.; Cui, J.X.; Gu, H.T.; Xu, L.Y.; Li, Y.Q.; Xu, Z.H.; Bai, S.N. Developmental analyses reveal early arrests of the spore-bearing parts of reproductive organs in unisexual flowers of cucumber (Cucumis sativus L.). Planta 2004, 220, 230–240. [Google Scholar] [CrossRef]
- Ma, H.; Liu, M. The microtubule cytoskeleton acts as a sensor for stress response signaling in plants. Mol Biol Rep 2019, 46, 5603–5608. [Google Scholar] [CrossRef]
- Liu, L.; Chen, J.; Gu, C.; Wang, S.; Xue, Y.; Wang, Z.; Han, L.; Song, W.; Liu, X.; Zhang, J.; Li, M.; Li, C.; Wang, L.; Zhang, X.; Zhou, Z. The exocyst subunit CsExo70B promotes both fruit length and disease resistance via regulating receptor kinase abundance at plasma membrane in cucumber. Plant Biotechnol J 2024, 22, 347–362. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Comparison of the conserved motifs and gene structures of CsTRM07 (A), CsTRM17 (B) and CsTRM24 (C) in the 13 cucumber accessions.
Figure 1.
Comparison of the conserved motifs and gene structures of CsTRM07 (A), CsTRM17 (B) and CsTRM24 (C) in the 13 cucumber accessions.
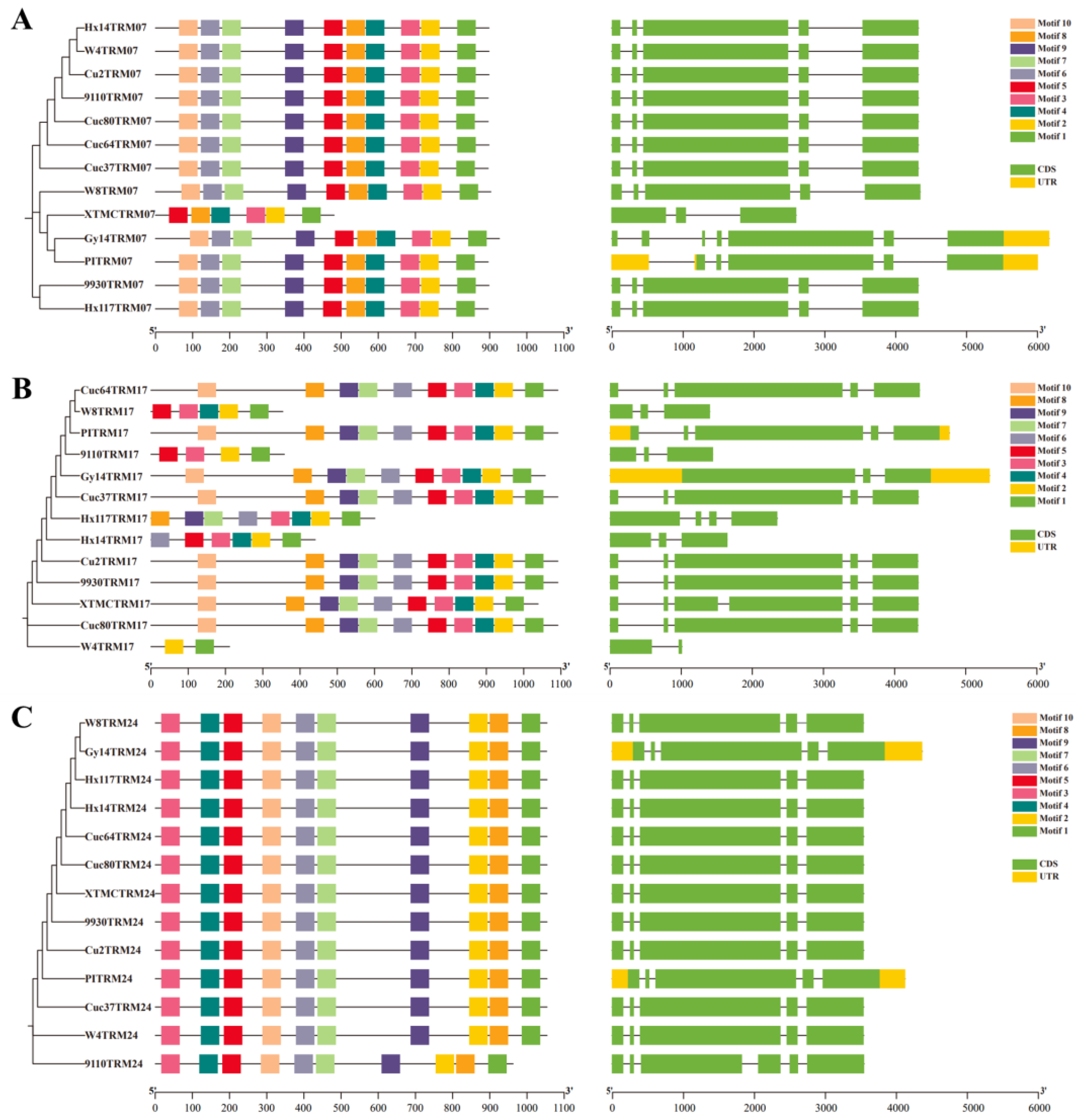
Figure 2.
Synteny analysis of TRMs between cucumber and other plant species: The collinear blocks are marked by gray lines, while the collinear gene pairs with TRM genes are highlighted by red lines. ‘C. sativus’, ‘Z. mays’, ‘O. sativa’, ‘A. thaliana’, and ‘S. lycopersicum’ indicate Cucumis sativus, Zea mays, Oryza sativa, Arabidopsis thaliana, and Solanum lycopersicum, respectively.
Figure 2.
Synteny analysis of TRMs between cucumber and other plant species: The collinear blocks are marked by gray lines, while the collinear gene pairs with TRM genes are highlighted by red lines. ‘C. sativus’, ‘Z. mays’, ‘O. sativa’, ‘A. thaliana’, and ‘S. lycopersicum’ indicate Cucumis sativus, Zea mays, Oryza sativa, Arabidopsis thaliana, and Solanum lycopersicum, respectively.
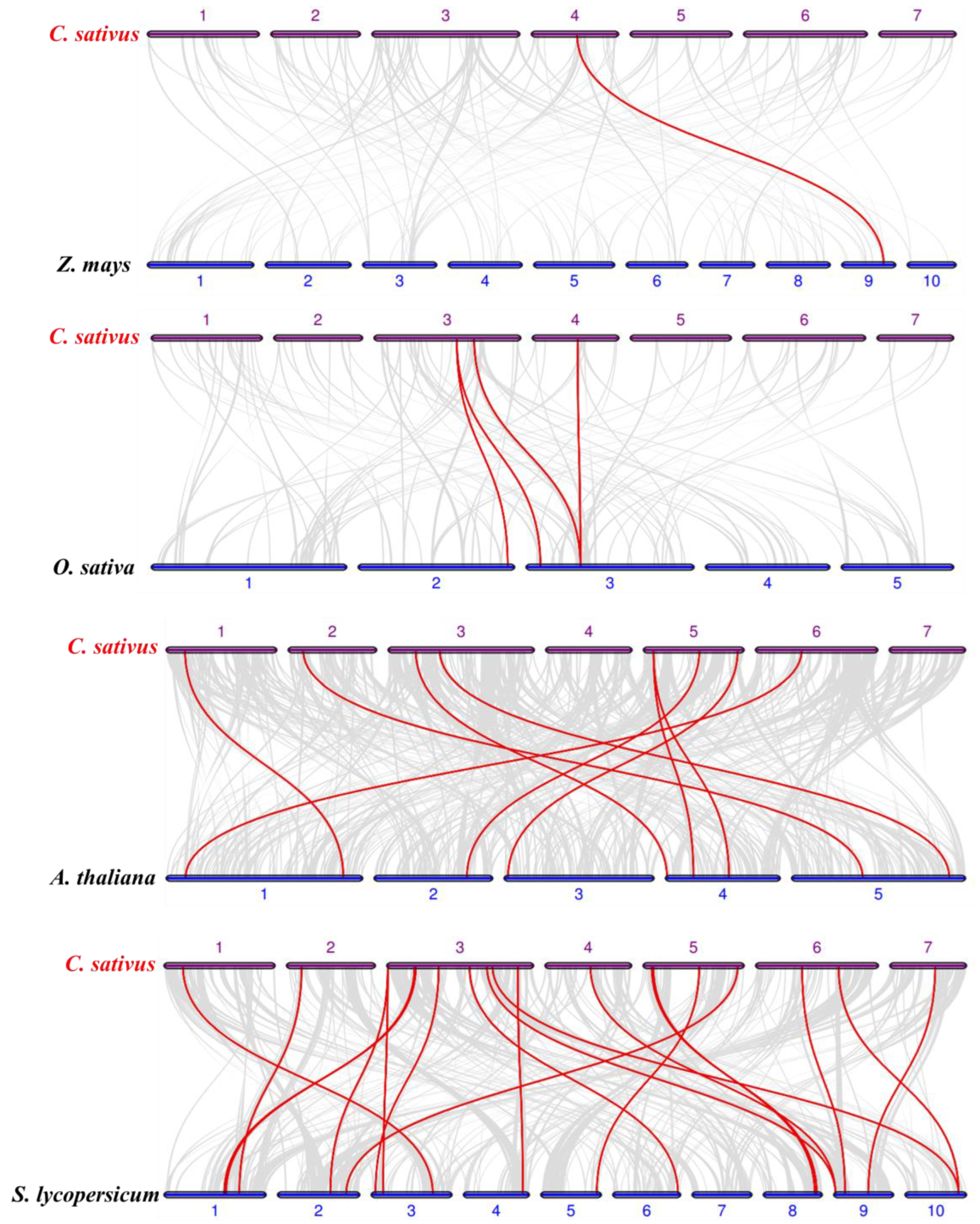
Figure 3.
Expression analysis of CsTRMs in the fruit: The transcriptional levels of CsTRM genes in GFC (carpel number = 5) and 32X (carpel number = 3) (A), 408 (long fruit) and 409 (short fruit) (B), and WT and CsFUL1A-OX (C) are shown on the heatmaps. A range of -2.0 to 2.0 and -1.5 to 1.5 was artificially set with the color scale limits according to the normalized values. The color scale shows increasing expression levels from blue to red. GFC, mutant Gui Fei Cui (GFC) from South China type cucumber 32X. The carpel number changed from 3 in 32X to 5 in GFC, despite the number of other floral organs, such as sepal, petal and stamen remain unchanged. WT, empty vector/control transgenic plants. FC, fold-change.
Figure 3.
Expression analysis of CsTRMs in the fruit: The transcriptional levels of CsTRM genes in GFC (carpel number = 5) and 32X (carpel number = 3) (A), 408 (long fruit) and 409 (short fruit) (B), and WT and CsFUL1A-OX (C) are shown on the heatmaps. A range of -2.0 to 2.0 and -1.5 to 1.5 was artificially set with the color scale limits according to the normalized values. The color scale shows increasing expression levels from blue to red. GFC, mutant Gui Fei Cui (GFC) from South China type cucumber 32X. The carpel number changed from 3 in 32X to 5 in GFC, despite the number of other floral organs, such as sepal, petal and stamen remain unchanged. WT, empty vector/control transgenic plants. FC, fold-change.
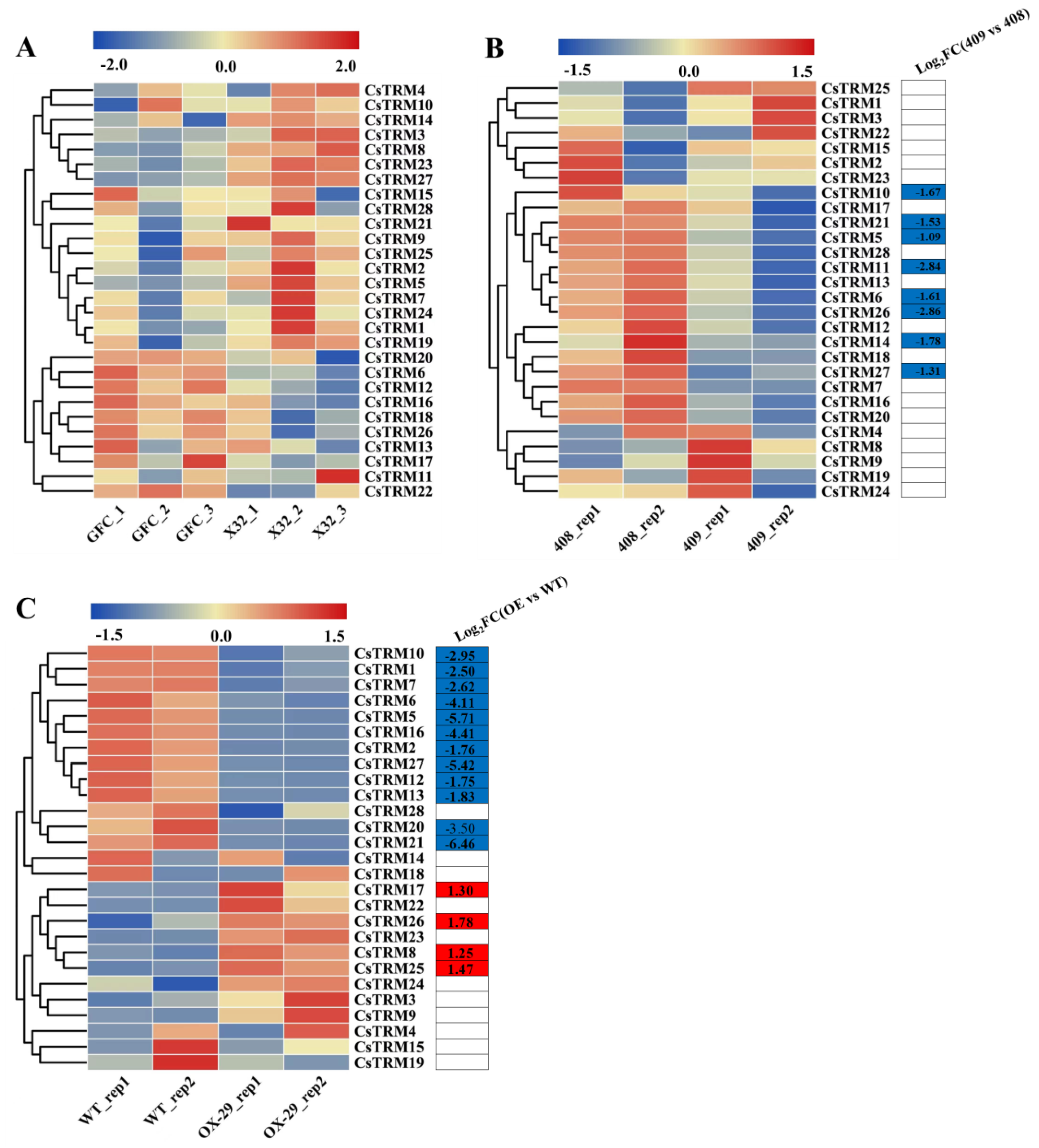
Figure 4.
Expression profiles of CsTRM genes in response to various abiotic stress treatments: The transcriptional levels of CsTRM genes in response to salt (A) and heat (B) stresses are shown on the heatmap. A range of -3.0 to 3.0 was artificially set with the color scale limits according to the normalized values. The color scale shows increasing expression levels from blue to red. CT, control treatment; HT, heat treatment; HT0h, heat treatment for 0 h (hours); HT3h, heat treatment for 3 h; HT6h, heat treatment for 6 h; FC, fold-change.
Figure 4.
Expression profiles of CsTRM genes in response to various abiotic stress treatments: The transcriptional levels of CsTRM genes in response to salt (A) and heat (B) stresses are shown on the heatmap. A range of -3.0 to 3.0 was artificially set with the color scale limits according to the normalized values. The color scale shows increasing expression levels from blue to red. CT, control treatment; HT, heat treatment; HT0h, heat treatment for 0 h (hours); HT3h, heat treatment for 3 h; HT6h, heat treatment for 6 h; FC, fold-change.
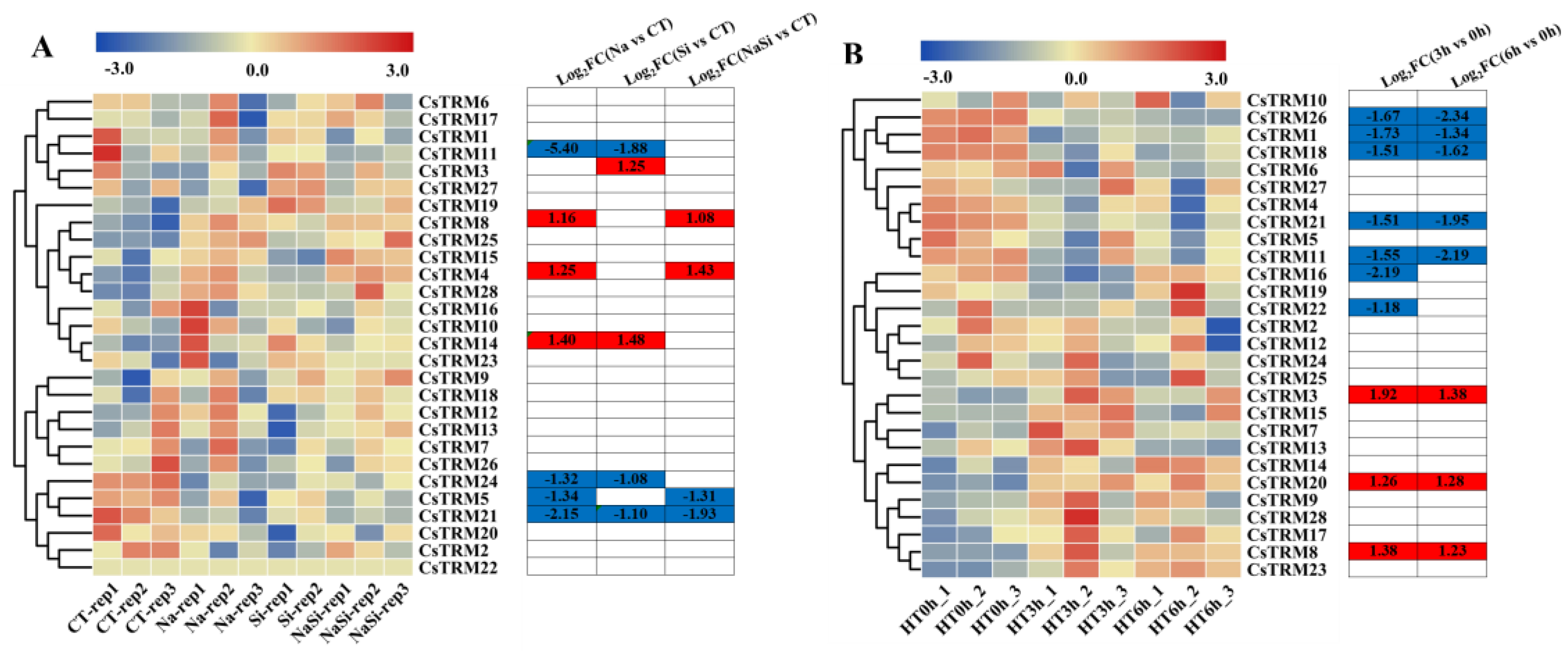
Figure 5.
Expression analysis of CsTRMs under biotic stresses: The transcriptional levels of CsTRM genes after infection with powdery mildew (PM) for 48 h (A), gray mold (GM) for 96 h (B), and with downy mildew (DM) for 1–8 days post-inoculation (C) are shown on the heatmaps. A range of -3.0 to 3.0, -2.5 to 2.5 and 0.0 to 7.0 was artificially set with the color scale limits according to the normalized values. The color scale shows increasing expression levels from blue to red. ID, PM-inoculated susceptible cucumber line D8 leaves; NID, non-inoculated D8 leaves; IS, PM-inoculated resistant cucumber line SSL508-28 leaves; NIS, non-inoculated SSL508-28 leaves; CT, without inoculation; DPI, days post inoculation; FC, fold-change.
Figure 5.
Expression analysis of CsTRMs under biotic stresses: The transcriptional levels of CsTRM genes after infection with powdery mildew (PM) for 48 h (A), gray mold (GM) for 96 h (B), and with downy mildew (DM) for 1–8 days post-inoculation (C) are shown on the heatmaps. A range of -3.0 to 3.0, -2.5 to 2.5 and 0.0 to 7.0 was artificially set with the color scale limits according to the normalized values. The color scale shows increasing expression levels from blue to red. ID, PM-inoculated susceptible cucumber line D8 leaves; NID, non-inoculated D8 leaves; IS, PM-inoculated resistant cucumber line SSL508-28 leaves; NIS, non-inoculated SSL508-28 leaves; CT, without inoculation; DPI, days post inoculation; FC, fold-change.
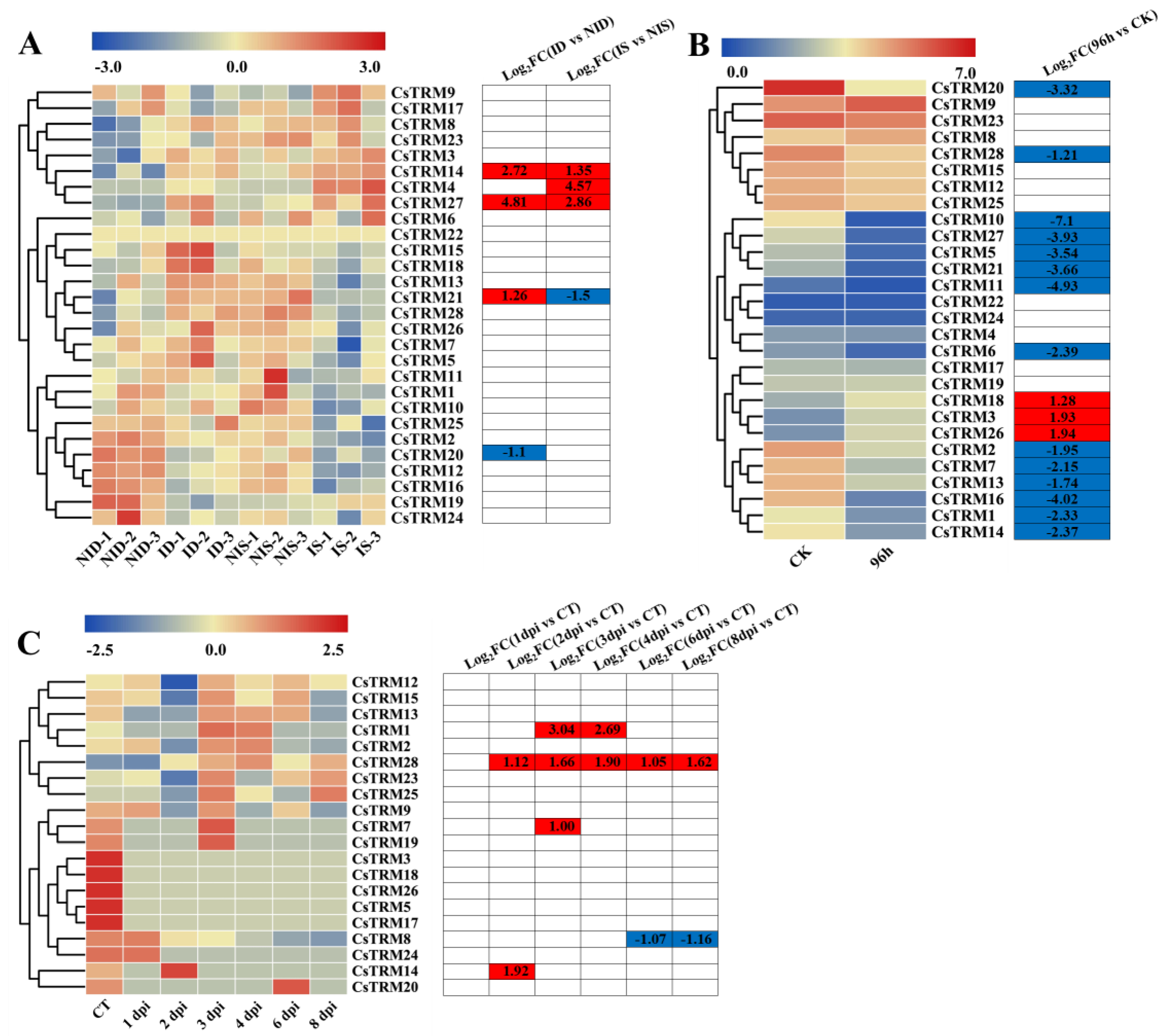

Table 1.
Identification of TRM genes in the 13 cucumber accessions.
| Gene name | 9930 | XTMC | Cu2 | Cuc80 | PI | Cuc64 | W4 | W8 | Hx14 | Hx117 | Cuc37 | Gy14 | 9110gt |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CsTRM01 | 1G003000 | 1G002960 | 1G003190 | 1G003020 | 1G02950 | 1G003020 | 1G003020 | 1G003050 | 1G008150 | 1G003070 | 1G003050 | 1G002970 | 1G003150 |
| CsTRM02 | 1G006080 | 1G006040 | 1G006230 | 1G006090 | 1G06220 | 1G006110 | 1G006160 | 1G006130 | 1G012280 | 1G009330 | 1G006130 | 1G005930 | 1G006390 |
| CsTRM03 | 1G024390 | 1G023100 | 1G019380 | 1G034690 | 1G022250 | 1G022640 | 1G033380 | 1G035450 | 1G025870 | 1G016710 | 1G021460 | ||
| CsTRM04 | 1G036240 | 1G038300 | 1G031670 | 1G23930 | 1G045200 |
1G039460 1G050980 |
1G048780 | 1G052690 | 1G036240 | 1G023410 | 1G034790 | ||
| CsTRM05 | 2G002210 | 2G001130 | 2G002170 | 2G01120 | 2G001150 | 2G001150 | 2G002120 | 2G002150 | 2G001120 | 2G001120 | 2G002200 | ||
| CsTRM06 | 2G006910 | 2G004780 | 2G004780 | 2G005680 | 2G04550 | 2G004690 | 2G004760 | 2G005690 | 2G006680 | 2G005730 | 2G004660 | 2G004680 | 2G005790 |
| CsTRM07 | 2G013800 | 2G013420 | 2G014430 | 2G016150 | 2G11310 | 2G012220 | 2G015170 | 2G022290 | 2G022190 | 2G018160 | 2G012230 | 2G011350 | 2G015370 |
| CsTRM08 | 3G000320 | 3G000290 | 3G000270 | 3G00310 | 3G000310 | 3G000300 | 3G000290 | 3G000310 | 3G000300 | 3G000310 | 3G000260 | 3G000300 | |
| CsTRM09 | 3G008900 | 3G014120 | 3G011330 | 3G009320 | 3G08770 | 3G009130 | 3G011320 | 3G009390 | 3G013030 | 3G018440 | 3G009280 | 3G008870 | 3G011230 |
| CsTRM10 | 3G009320 | 3G014570 | 3G09200 | 3G009570 | 3G011790 | 3G009840 | 3G013470 | 3G018890 | 3G009740 | 3G009280 | 3G011660 | ||
| CsTRM11 | 3G016640 | 3G023990 | 3G019120 | 3G016980 | 3G16440 | 3G027380 | 3G019160 | 3G017460 | 3G023810 | 3G029290 | 3G017050 | 3G016550 | 3G018960 |
| CsTRM12 | 3G020250 | 3G028160 | 3G024300 | 3G021230 | 3G20290 | 3G031530 | 3G023320 | 3G021650 | 3G030910 | 3G038450 | 3G021080 | 3G020040 | 3G025120 |
| CsTRM13 | 3G028590 | 3G044970 | 3G034490 | 3G039640 | 3G27110 | 3G050790 | 3G034230 | 3G032590 | 3G049730 | 3G053340 | 3G043310 | 3G025270 | 3G034790 |
| CsTRM14 | 3G033690 | 3G052230 | 3G039760 | 3G045880 | 3G31210 | 3G055180 | 3G039400 | 3G038880 | 3G057040 | 3G059810 | 3G049600 | 3G029050 | 3G041170 |
| CsTRM15 | 3G035160 | 3G053700 | 3G041160 | 3G047320 | 3G32570 | 3G056620 | 3G040870 | 3G040290 | 3G058490 | 3G061290 | 3G050990 | 3G030380 | 3G042680 |
| CsTRM16 | 3G036950 | 3G056500 | 3G043950 | 3G049050 | 3G34290 | 3G032070 | 3G044470 | ||||||
| CsTRM17 | 3G045060 | 3G067760 | 3G055880 | 3G057270 | 3G42630 | 3G066680 | 3G051990 | 3G050610 | 3G070610 | 3G071560 | 3G061110 | 3G040150 | 3G052920 |
| CsTRM18 | 4G024630 | 4G030170 | 4G024030 | 4G078840 | 4G14290 | 4G027840 | 4G018900 | 4G021930 | 4G026900 | 4G030090 | 4G084710 | 4G013800 | 4G026010 |
| CsTRM19 | 4G031780 | 4G042910 | 4G034540 | 4G090510 | 4G21450 | 4G044030 | 4G027440 | 4G033570 | 4G040430 | 4G039790 | 4G095540 | 4G020000 | 4G035410 |
| CsTRM20 | 5G002760 | 5G003630 | 5G003640 | 5G002610 | 5G05360 | 5G003650 | 5G002590 | 5G005490 | 5G006540 | 5G003570 | 5G002660 | 5G003770 | |
| CsTRM21 | 5G003260 | 5G004130 | 5G004140 | 5G003110 | 5G05880 | 5G003020 | 5G003100 | 5G004990 | 5G007040 | 5G004090 | 5G003170 | 5G003160 | 5G004310 |
| CsTRM22 | 5G005590 | 5G007580 | 5G007530 | 5G005560 | 5G08200 | 5G000650 | 5G006510 | 5G001620 | 5G009400 | 5G009680 | 5G005630 | 5G005580 | |
| CsTRM23 | 5G026190 | 5G042130 | 5G041190 | 5G050680 | 5G17200 | 5G021470 | 5G028290 | 5G024190 | 5G042910 | 5G054980 | 5G043620 | 5G016900 | 5G034200 |
| CsTRM24 | 5G038540 | 5G060730 | 5G054610 | 5G063070 | 5G29400 | 5G043920 | 5G040890 | 5G049340 | 5G063180 | 5G067580 | 5G056820 | 5G028860 | 5G046740 |
| CsTRM25 | 6G016870 | 6G024320 | 6G018060 | 6G025320 | 6G14470 | 6G015450 | 6G015250 | 6G019390 | 6G025200 | 6G018200 | 6G015300 | 6G014340 | 6G017280 |
| CsTRM26 | 6G022550 | 6G035270 | 6G023850 | 6G053560 | 6G17180 | 6G022100 | 6G019950 | 6G023190 | 6G032860 | 6G030040 | 6G019540 | 6G016650 | 6G023980 |
| CsTRM27 | 6G040450 | 6G052040 | 6G035430 | 6G079450 | 6G25260 | 6G035100 | 6G032700 | 6G033070 | 6G052400 | 6G045570 | 6G036810 | 6G024690 | 6G035970 |
| CsTRM28 | 7G025430 | 7G031600 | 7G024250 | 7G035470 | 7G13640 | 7G025890 | 7G021920 | 7G034950 | 7G031600 | 7G031470 | 7G037050 | 7G012470 | 7G023340 |
| CsTRM29 | UnG00530 |
Table 2.
The predicted lengths of TRM proteins (amino acid residues) in the 13 cucumber accessions.
| Protein number | 9930 | XTMC | Cu2 | Cuc80 | PI | Cuc64 | W4 | W8 | Hx14 | Hx117 | Cuc37 | Gy14 | 9110gt |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CsTRM01 | 1048 | 1048 | 1048 | 1048 | 1048 | 1048 | 1043 | 1048 | 1048 | 1048 | 1048 | 1063 | 1048 |
| CsTRM02 | 1040 | 1067 | 1067 | 1067 | 1067 | 1067 | 1067 | 1067 | 1067 | 1067 | 1067 | 1067 | 1067 |
| CsTRM03 | 780 | 788 | 788 | 788 | 781 | 781 | 781 | 781 | 781 | 788 | 781 | ||
| CsTRM04 | 402 | 402 | 402 | 402 | 402 | 402/402 | 402 | 402 | 402 | 402 | 402 | ||
| CsTRM05 | 776 | 803 | 803 | 803 | 803 | 803 | 803 | 803 | 803 | 803 | 803 | 803 | 803 |
| CsTRM06 | 722 | 722 | 722 | 722 | 722 | 722 | 722 | 750 | 722 | 722 | 722 | ||
| CsTRM07 | 893 | 478 | 893 | 891 | 891 | 893 | 893 | 899 | 893 | 891 | 891 | 922 | 891 |
| CsTRM08 | 893 | 879 | 879 | 893 | 879 | 879 | 879 | 893 | 879 | 879 | 904 | 879 | |
| CsTRM09 | 930 | 933 | 933 | 933 | 932 | 933 | 933 | 933 | 933 | 933 | 933 | 932 | 933 |
| CsTRM10 | 346 | 346 | 344 | 344 | 344 | 344 | 346 | 344 | 346 | 305 | 346 | ||
| CsTRM11 | 616 | 616 | 616 | 616 | 616 | 616 | 616 | 616 | 616 | 616 | 616 | 616 | 616 |
| CsTRM12 | 953 | 953 | 954 | 954 | 953 | 953 | 953 | 953 | 953 | 952 | 953 | 954 | 953 |
| CsTRM13 | 963 | 963 | 963 | 963 | 963 | 963 | 963 | 963 | 963 | 963 | 963 | 927 | 963 |
| CsTRM14 | 353 | 353 | 353 | 353 | 353 | 353 | 353 | 353 | 353 | 353 | 353 | 353 | 353 |
| CsTRM15 | 888 | 888 | 888 | 888 | 888 | 888 | 888 | 888 | 888 | 888 | 888 | 888 | 888 |
| CsTRM16 | 472 | 472 | 550 | 550 | 472 | 472 | 550 | ||||||
| CsTRM17 | 1091 | 1038 | 1091 | 1091 | 1091 | 1091 | 210 | 353 | 440 | 600 | 1091 | 1058 | 357 |
| CsTRM18 | 961 | 961 | 961 | 961 | 961 | 922 | 961 | 961 | 961 | 961 | 961 | 961 | 961 |
| CsTRM19 | 903 | 903 | 987 | 987 | 906 | 987 | 906 | 906 | 906 | 906 | 906 | 906 | 906 |
| CsTRM20 | 785 | 785 | 785 | 785 | 781 | 781 | 781 | 781 | 785 | 785 | 745 | 785 | |
| CsTRM21 | 476 | 476 | 476 | 476 | 476 | 476 | 476 | 476 | 476 | 476 | 476 | 476 | 476 |
| CsTRM22 | 495 | 495 | 495 | 495 | 495 | 495 | 495 | 495 | 495 | 495 | 495 | 449 | |
| CsTRM23 | 794 | 794 | 794 | 848 | 795 | 795 | 794 | 795 | 794 | 794 | 795 | 794 | 736 |
| CsTRM24 | 1049 | 1049 | 1049 | 1049 | 1049 | 1049 | 1049 | 1049 | 1049 | 1049 | 1049 | 1049 | 958 |
| CsTRM25 | 1011 | 1011 | 1011 | 1011 | 902 | 959 | 1009 | 1011 | 1022 | 1009 | 1011 | 940 | 1011 |
| CsTRM26 | 936 | 936 | 936 | 936 | 936 | 936 | 938 | 936 | 936 | 936 | 936 | 936 | 936 |
| CsTRM27 | 505 | 505 | 505 | 505 | 505 | 505 | 505 | 736 | 505 | 473 | 505 | 505 | 505 |
| CsTRM28 | 960 | 960 | 960 | 959 | 959 | 994 | 995 | 1047 | 895 | 995 | 995 | 976 | 978 |
| CsTRM29 | 788 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



