
Submitted:
27 June 2024
Posted:
28 June 2024
You are already at the latest version
Abstract
The detection of early biomarkers and molecular mechanisms in Parkinson’s disease (PD) remains a challenge. Recent research has pointed to potential roles for peptidylarginine deiminases (PADs), a family of calcium activated enzymes, and associated post-translational protein citrul-lination/deimination in early stages of the disease, with a focus on hippocampal and cortex re-gions of the brain. Further analysis of brain region specific changes in PAD isozymes and identi-fication of brain-region specific citrullinomes, therefore remain to be studied. This study assessed brain-region specific PAD isozyme expression (PADs 1-4; PAD6) and associated citrullinated protein targets in the 6-hydroxydopamine (6-OHDA) induced rat model of pre-motor PD. Cortex, hippocampus, striatum, midbrain, cerebellum and olfactory bulb were compared between con-trols/shams and the PD model. In cortex, a significant increase of PAD2 and PAD3 was observed in the PD model. In hippocampus, PAD3, PAD4 and PAD6 showed strongest levels, but with no significant changes observed between PD and controls. In striatum, a significant reduction in all PAD isozymes was observed in the PD model. In the olfactory bulb, PAD3 was significantly el-evated in PD. In midbrain, PAD2, PAD4 and PAD6 showed strongest signal but with no signifi-cant changes between PD and controls. In cerebellum, a significant increase was seen in PAD3, PAD4 and PAD6 in the PD model. Citrullinated protein hits were most abundant in cortex and hippocampus, while for the PD model an increase in citrullinated hits was particularly observed in cortex and cerebellum, compared to controls. For all brain regions there was a considerable difference in citrullinated protein IDs between the PD model and the controls. Citrullinome as-sociated KEGG pathways differed in the six brain regions; some were overlapping for controls and PD, some were identified for the PD model only, and some were identified in control brains only. KEGG pathways identified in PD brains onlywere: “Axon guidance”; “Spinocerebellar ataxia”; “Hippo signalling pathway”; “NOD-like receptor signalling pathway”; “Phosphatidyl-inositol signalling system”; “Rap1 signalling pathway”; “Platelet activation”; “Yersinia infection”; “Fc gamma R-mediated phagocytosis”; “Human cytomegalovirus infection”; “Inositol phosphate metabolism”; “Thyroid hormone signalling pathway”; “Progesterone-mediated oocyte matura-tion”; “Oocyte meiosis”; and “Choline metabolism in cancer”. Our findings highlight differences in PAD isozymes, citrullinated proteins and associated KEGG pathways between brain regions, in pre-motor PD.
Keywords:
peptidylarginine deiminase (PAD)
; citrullination/deimination
; Parkinson’s disease
; brain
; cortex
; hippocampus
; midbrain
; striatum
; cerebellum
; olfactory bulb
; KEGG
1. Introduction
Molecular pathways in early stages of Parkinson’s disease (PD), including pre-motor PD, are an important area of research in the light of a lack of current treatment strategies and the need for identification of early biomarkers [1]. More than 10 million people worldwide live with PD, the second most common age-related neurodegenerative disorder, and 4% of affected people are diagnosed before the age of 50. The identification of early PD-related pathways that can serve as novel drug targets and aid early diagnosis is therefore of pivotal importance. In combination with clinical and post-mortem human patient samples, which are in limited supply, the use of early-stage PD animal models offers promising avenues for in vivo assessment of novel molecular pathways and candidate pharmacological lead compounds for therapeutic intervention.
Recent research has highlighted a role for peptidylarginine deiminases (PADs), a family of calcium activated enzymes that cause post-translational citrullination/deimination of proteins, through irreversible conversion of arginine to citrulline, and are associated with inflammatory and neurodegenerative diseases, including through altered self-epitopes, changes in protein interactions and epigenetic regulation [2], most recently also linked to PD [3,4,5].
PADs are found in 5 isoforms in mammals, show tissue specific expression and vary in preference for target proteins, with some shared targets [6]. Furthermore, beta sheets and intrinsically disordered proteins are more susceptible to citrullination/deimination and the position of the arginine also plays a role [7,8]. Importantly, a wide range of cytosolic, cytoskeletal, mitochondrial and nuclear associated proteins can undergo citrullination, leading to their modified functions, including loss of function, new acquired functions and protein moonlighting. PAD2 is considered the most ancestral and ubiquitously expressed PAD isozyme, while all isozymes have been linked to physiological and pathobiological processes [6,8,9,10,11]. The main PAD isozymes studied to date in the brain and in neurodegenerative disorders have been PAD2 and PAD4, including in development and ageing [12], Alzheimer’s disease (AD) [13,14], amyotrophic lateral sclerosis (ALS), prion diseases and in multiple sclerosis (MS) [15,16,17,18,19,20,21], as well as in acute brain injury [22,23,24,25], but importantly recently also highlighted in both human PD brain samples and in PD animal models [4,5,26]. Roles for PAD3 have been described in CNS regeneration [27], in brain cancer [28,29], in neuronal stem cells [30] and neuronal viral infection [31]. Understanding of roles for PAD3 in neurodegeneration are limited, but recent research highlighted elevated PAD3 levels in PD post-mortem human brain samples [5] as well as in pre-motor PD rat cortex and hippocampus [4]. PAD1 has been associated with skin diseases, development, and some cancers [32,33,34,35]. Importantly PAD1 was also recently identified by our group to be elevated in human postmortem PD brain samples of hippocampus and cortex [5]. PAD6 is shorter than the other isozymes, shares less sequence homology with the other PAD family members and does not seem to bind calcium, contrary to the other isozymes [36]. PAD6 has been mainly studied for roles in early embryonic development, oocyte formation and embryo implantation [37,38], and some associations have been made with cancers [39]. Importantly recent roles for PAD6 in the CNS have been highlighted, including in brains of hypoxia-challenged naked mole-rats [40] and in post-mortem human PD brain samples where PAD6 was detected in the brain vasculature by immunohistochemistry [5].
As recent studies from both animal models and human post-mortem brain samples have identified increased PADs and citrullination at early PD stages [4,5], an investigation is now warranted into the brain-region specific citrullinomes in early stages of PD. The current study therefore used the previously published toxin-induced pre-motor PD rat model [4] to assess changes in PAD isozymes and citrullinated protein targets by proteomic analysis in the following brain regions: striatum, olfactory bulb, midbrain, hippocampus, cortex, and cerebellum.
2. Results
2.1. Brain Region Related Differences in PAD Isozyme Detection
The six brain regions under study (cortex, hippocampus, midbrain, striatum, cerebellum and olfactory bulb) were assessed by western blotting for all five PAD isozymes (PAD1,2,3,4 and PAD6). Results are shown in Figure 1, Figure 2 and Figure 3 and summarised in Table 1. PAD1 was observed at very low levels in all brain regions, with only a clear signal observed in control/sham striatum. In cortex, PAD2, PAD3, PAD4 and PAD6 were all detected, although protein bands for PAD2, PAD3 and PAD4 were not clearly defined in all samples, while PAD6 showed a strong band in most samples. Based on densitometry analysis (normalized with beta-actin) of the regions for the expected PAD bands (70-75 kDa for PAD1-4, and 50-60kDa for PAD6, as indicated by arrows and brackets) a significant increase of PAD2 and PAD3 was observed in the pre-motor PD cortex (Figure 1A-E). In hippocampus PAD3, PAD4 and PAD6 showed strongest levels of the PAD isozymes, although positive protein bands were somewhat blurred for PAD2 and PAD3, while PAD4 bands were more clearly detected and PAD6 showed a strong positive band. No significant changes were observed for PADs in control versus pre-motor PD hippocampus, albeit there was some trend for reduced PAD2,3 and PAD4 levels, and increased PAD6 levels (albeit not statistically significant) (Figure 1A-E). In addition, prohibitin levels were assessed as a marker of mitochondrial housekeeping, with some trend of reduced levels (but non-significant) in cortex and hippocampus of the PD group (Figure 1F).
In cerebellum, PAD1 levels were low, PAD2, PAD3, PAD4 and PAD6 showed positive signal, although bands were only clearly defined for PAD6. Based on densitometry analysis of the 70-75 kDa region for PAD1-4 and 50-60 kDa region for PAD6, a significant increase was observed for PAD3, PAD4 and PAD6 in PD cerebellum, compared with controls/shams (Figure 2 A-E). In midbrain, PAD1 levels were negligible, while PAD2 and PAD6 were more strongly detected, and PAD3 and PAD4 detected at lower level. The protein bands for PAD1-4 were furthermore not clearly defined, while PAD6 showed a more clearly detectable band (Figure 2 A-E). While no significant changes were observed for any of the PAD isozymes between control and PD midbrains, based on densitometry analysis normalized to beta-actin, there was some trend for decreased PAD2 and PAD4 (Figure 2 B and D). In cerebellum, prohibitin levels showed some elevation for the PD group, but not significantly, while this was not observed for midbrain.
In the olfactory bulb, low detection of PAD1,2 and 3 isozymes was observed, with higher positive signal detected for PAD4 and PAD6, although PAD positive bands were all diffuse for olfactory bulb. In the PD olfactory bulb, a significant elevation for PAD3, and a trend for increased PAD4 and PAD6 (albeit non-significant) was observed based on densitometry analysis of these regions, normalized to beta-actin loading control (Figure 3 A-E). In striatum, positive signals for all PADs isozymes were observed, but with lowest detection of PAD6. Based on densitometry analysis of the PAD-positive region of the blot (as indicated by arrows and brackets) and normalized against beta-actin, a significant reduction of protein levels for PAD1, PAD2, PAD3, PAD4 and PAD6 was observed in PD striatum compared with controls/shams (Figure 2 A-E). Prohibitin protein levels were significantly increased in PD olfactory bulb, but no difference was observed in the striatum between the two groups (Figure 3F).
Findings from the western blotting analysis in Figure 1, Figure 2 and Figure 3 are summarized below in Table 1 for PAD protein levels detected in the six brain regions under study in the control (ctr) and pre-motor PD (PD) groups. It must be noted that in many of the rat brain regions, PAD isozyme detection with the human PAD-isozyme specific antibodies revealed blurred, rather than clearly defined bands, and therefore the region with expected PAD isozyme detection (70-75 kDa for PAD1-4 and 50-60 kDa for PAD6; as indicated by arrows and/or brackets) was assessed. In addition, individual variation within each sample group was observed, as reflected in the observation of a trend, rather than statistically significant differences; this is further summarized in Table 1.
2.2. Isolation of Citrullinated Target Proteins from Brain Regions and Silver Staining
Total citrullinated/deiminated proteins were isolated from brain tissue of the six brain regions, using the pan-citrulline F95 antibody and the catch-and-release immunoprecipitation kit, according to previously described methods [40]. The yield of citrullinated proteins was highest in cortex and hippocampus, as observed by silver staining of eluted fractions (Figure 4A), and citrullinated proteins were isolated at lower yield from cerebellum, midbrain, olfactory bulb, and striatum, as observed by silverstaining of the F95 enriched eluted proteins from each brain region (Figure 4B). Numbers of citrullinated proteins identified by LC-MS/MS analysis (see 3.4) per brain region, in control and PD groups, are summarised in Figure 4C. Highest numbers of citrullinated protein hits were detected in the cortex and hippocampus, while elevated numbers of total citrullinated hits were seen in the PD model for cortex and cerebellum, compared with controls.
2.3. Identification of the Brain Region Specific Citrullinomes by LC-MS/MS
Citrullinated protein hits isolated per brain region were further assessed by LC-MS/MS analysis for identification of protein target hits. All protein hits identified were compared and numbers of individual and shared hits are summarised for the different brain regions in the Venn diagrams in Figures 5C-10C. Details on individual protein hits per brain region (control and PD) are in addition provided in Supplementary Tables S1-S12. Citrullinated protein hits were then used to create protein-protein interaction networks for the citrullinome of each brain region using STRING analysis (https://string-db.org/; accessed 18th and 19th of March 2024). The resulting protein interaction networks are shown in Figure 5, Figure 6, Figure 7, Figure 8, Figure 9 and Figure 10 (A and B). All KEGG pathways identified in association to the brain region specific citrullinomes are summarised in Table 2.
2.4. Protein-Protein Interaction Network Analysis for the Brain Region Specific Citrullinomes in Pre-Motor PD versus Control Brains
Protein-protein interaction networks for the citrullinated proteins identified in each brain region were created in STRING, comparing sham/control and pre-motor PD brain regions (Figure 5, Figure 6, Figure 7, Figure 8, Figure 9 and Figure 10). The protein citrullinome networks for cortex, shams/controls and PD, are shown in Figures 5A and 5B, respectively. As summarised in the Venn diagram in Figure 5C, 195 protein hits were identified as citrullinated targets specific to PD cortex, while 74 were only identified in control cortex, and 246 citrullinated protein hits were common to both control and PD cortex. KEGG pathways associated to the cortex citrullinome were 25 for PD cortex, 7 for control cortex and further 43 KEGG pathways were identified as common between control and PD cortex; these are listed in Table 2.
The 25 KEGG pathways which were identified as associated to the citrullinome of pre-motor PD cortex, but not found associated to the control cortex citrullinome were the following metabolic pathways: “Inositol phosphate metabolism”; “2-Oxocarboxylic acid metabolism”; “Nitrogen metabolism”; “Pentose phosphate pathway”; “Fructose and mannose metabolism”; “Alanine, aspartate and glutamate metabolism”; “Cysteine and methionine metabolism”; “Arginine biosynthesis”; “Vasopressin-regulated water reabsorption”; “Gastric acid secretion”; “Phosphatidylinositol signalling system”. Pathways relating to cell adhesion, cell survival and cellular communication were: “Rap1 signalling pathway”; “SNARE interactions in vesicular transport”; “Tight junction”; “Regulation of actin cytoskeleton”. Immune related pathways were: “NOD-like receptor signalling pathway”; “Leukocyte transendothelial migration”; “Viral carcinogenesis”; “Platelet activation”; “Yersinia infection”; “Human cytomegalovirus infection”. Developmental pathways were: “Progesterone mediated oocyte maturation”; “Oocyte meiosis”. Neuronal and neurodegenerative associated pathways were: “Axon guidance”; “Spinocerebellar ataxia”.
The protein citrullinome networks for hippocampus, from sham/control and PD groups, are shown in Figures 6A and 6B, respectively. As summarised in the Venn diagram in Figure 6C, 106 protein hits were identified as citrullinated targets specific to PD hippocampus, while 144 were only identified in control cortex, and 257 citrullinated protein hits were common to both control and PD hippocampus. KEGG pathways associated to the hippocampus citrullinomes were 9 for PD hippocampus, 10 for control hippocampus and further 55 KEGG pathways were identified as common between control and PD hippocampus; these are listed in Table 2.
The 9 KEGG pathways identified in PD hippocampus, but not in the citrullinome of the sham/control hippocampus were the following metabolic pathways: “Phosphatidylinositol signalling system”; “Endocrine and other factor-regulated calcium reabsorption”; “Choline metabolism in cancer”; “Thyroid hormone signalling pathway”. Pathways relating to cell growth, development and stem cell regulation were: “cGMP-PKG signalling pathway”; “Hippo signalling pathway”; “Oocyte meiosis”. Immune related pathways were: “Fc gamma R-mediated phagocytosis”. CNS related pathways were: “Long-term depression”.
The protein citrullinome networks for cerebellum, from sham/control and PD groups, are shown in Figures 7A and 7B, respectively. As summarised in the Venn diagram in Figure 7C, 74 protein hits were identified as citrullinated targets specific to PD cerebellum, while 31 were only identified in control cerebellum, and 39 citrullinated protein hits were common to both control and PD cerebellum. KEGG pathways associated to the cerebellum citrullinomes were nine for PD, ten for control and further six KEGG pathways were identified as common between control and PD cerebellum; these are listed in Table 2.
The nine KEGG pathways identified in PD cerebellum, but not in the citrullinome of the control cerebellum were metabolic pathways: “Oxidative phosphorylation”; “Thermogenesis”. CNS and neurodegenerative linked pathways were: “Synaptic vesicle cycle”; “Alzheimer’s disease”; “Glutamatergic synapse”; “Gap junction”. Inflammatory pathways were: “Endocytosis”; “Phagosome” and “Apoptosis”.
The protein citrullinome networks for midbrain of control and PD groups, are shown in Figures 8A and 8B, respectively. As summarised in the Venn diagram in Figure 8C, 19 protein hits were identified as citrullinated targets specific to PD midbrain, while 130 were only identified in control midbrain, and further 48 citrullinated protein hits were common to both control and PD midbrain. KEGG pathways associated to the midbrain citrullinomes were 4 for PD only, 25 for control and further 12 KEGG pathways were identified as common between control and PD midbrain; these are listed in Table 2.
The four KEGG pathways identified to be specific for the citrullinome of PD midbrain, but not associated to the citrullinome of the control midbrain were inflammatory related: “Apoptosis”; “Staphylococcus aureus infection”; and “Bacterial invasion of epithelial cells”. In addition, “Arrhythmogenic right ventricular cardiomyopathy” was unique to the PD model.
The protein citrullinome networks for olfactory bulb, from sham/control and PD groups, are shown in Figures 9A and 9B, respectively. As summarised in the Venn diagram in Figure 9C, 7 protein hits were identified as citrullinated targets specific to PD olfactory bulb, while 45 were only identified in control olfactory bulb, and further 6 citrullinated protein hits were common to both control and PD olfactory bulb. KEGG pathways associated to the olfactory bulb citrullinomes were three shared between the control and PD model, while none was specific to the PD model only, and an additional 19 citrullinome associated pathways were identified for the control olfactory bulb; these are listed in Table 2.
The protein citrullinome networks for striatum, from sham/control and PD groups, are shown in Figures 10A and 10B, respectively. As summarised in the Venn diagram in Figure 10C, 10 protein hits were identified as citrullinated targets specific to PD striatum, while 20 were only identified in control striatum, and further 15 citrullinated protein hits were common to both control and PD striatum. KEGG pathways associated to the striatum citrullinomes were 3 for PD only, 2 for control and further 6 KEGG pathways were identified as common between control and PD striatum; these are listed in Table 2.
The three KEGG pathways identified in PD striatum, but not in the citrullinome of the control striatum were: “Ribosome”; “Oestrogen signalling pathway”; and “Staphylococcus aureus infection”.
In addition to the KEGG analysis, brain-region specific citrullinomes were also assessed for Reactome pathways, and a comparison of total numbers of associated KEGG and Reactome pathways for all six brain regions, of sham/control and PD groups, is shown in Figure 11A. Reactome associated pathways were increased in the protein citrullinome networks of PD striatum, PD cerebellum, PD cortex and in PD hippocampus, versus controls. Reactome pathways associated to the brain region specific citrullinomes were reduced in olfactory bulb and midbrain of the PD model, compared to controls (Figure 11A).
When assessing any changes in gene ontology (GO) pathways associated to the brain specific citrullinomes, increased numbers of Biological Process GO pathways were associated to the PD cerebellum and cortex; Molecular Function GO pathways were increased in PD cortex and hippocampus; and Cellular Component GO pathways were increased in PD striatum, cerebellum, and cortex; compared with controls (Figure 11B). There was a reduction in Biological Function GO pathways associated to the citrullinomes of the PD olfactory bulb, midbrain and hippocampus, compared to controls; Molecular function GO pathways were reduced in the citrullinomes of PD olfactory bulb, cerebellum and midbrain, compared to controls; while Cellular Component GO pathways were reduced in the citrullinomes of PD olfactory bulb, midbrain and hippocampus, compared with controls (Figure 11B).
3. Discussion
The detection of early molecular mechanisms in the onset of PD remains a challenge. Pre-motor PD patients display various non-motor symptoms, which include decreased sense of smell, gastrointestinal problems, depression, sleep disturbances and autonomic dysfunction [1]. Toxin-induced rat models offer a valuable tool to investigate pre-motor aspects of PD and explore associated molecular alterations in distinct brain regions. Understanding of brain region specific changes in early PD is of considerable interest, including recent transcriptome profiling studies [41]. As previous research from our group has pointed to roles for PADs and associated post-translational protein citrullination/deimination, including findings from post-mortem human brain samples and the 6-OHDA induced rat model [4], the same pre-motor PD model was used in the current study. We assessed brain-region specific PAD isozyme expression and associated citrullinated proteins in cortex, hippocampus, striatum, midbrain, cerebellum and the olfactory bulb. The five PAD isozymes (PAD1,2,3,4 and 6) were assessed by western blotting, while immunoprecipitation in conjunction with LC-MS/MS analysis were used for identification of citrullinated protein targets in these six brain regions and downstream protein interaction network analysis was carried out for the identification of associated KEGG and GO pathways.
3.1. Brain Region Specific PAD Isozyme Detection
In all six brain regions, PAD1 detection was negligible or very low, except in striatum. Notably, it must be highlighted that for many of the rat brain samples assessed, the PAD positive bands detected using anti-human PAD isozyme specific antibodies, for PAD detection in the expected size-range of 70-75 kDa were blurred, while for some brain regions these were more defined. PAD6 overall showed the clearest positive band in the expected 50-60 kDa range. As anti-human PAD isozyme specific antibodies were used, this may have had some effect on species-cross reactive PAD isozyme detection by western blotting in these rat brains, although previously, the PAD2,3 and 4 antibodies showed positive in the same rat brains by immunohistochemistry [4]. These antibodies have though not been assessed by western blotting in rat brains before, which may explain some of the differences observed from immunohistochemistry [4] and western blotting results.
In cortex, PAD2, PAD3, PAD4 and PAD6 were all detected, with significant increase in PAD2 and PAD3 protein levels in the PD model (based on normalization with beta-actin as a loading control), compared with controls/shams. This may be correlated with the increased citrullinated hits observed for the PD group, also reflected in increased KEGG pathways associated with the PD cortex citrullinome. These findings do align with previous immunohistochemistry staining of this region in the same model [4] and in human post-mortem PD brain samples, where in anterior cingulate cortex PAD2 and PAD3 were particularly elevated at Braak stage 4, while the other PAD isozymes were though also elevated in the PD versus control brains [5].
In hippocampus, PAD3, PAD4 and PAD6 showed strongest levels, but there was some (non-significant) trend observed for reduced PAD2, PAD3 and PAD4 levels, and increased PAD6 levels in the PD group, when normalizing the band region of interest with the beta-actin loading control. Interestingly, there was a considerable difference observed in the IDs of citrullinated protein hits between PD and control hippocampus which included 106 unique citrullinated proteins in PD hippocampus and 144 unique ones for control hippocampus. This was reflected in considerable differences observed in downstream KEGG and GO pathways for the respective citrullinomes. Our findings relate somewhat to previous immunohistochemical analysis of hippocampus in the same model, where PAD2,3 and 4 were detected [4], while in human post-mortem PD brain samples, we found that PAD2 and PAD3 were elevated in the PD hippocampus, particularly at Braak stage 4, while increase in PAD4 was less. In the human PD brain samples, an increase in PAD6 was mainly linked to the brain vasculature while PAD1 was also found to be elevated in PD hippocampus [5].
Based on the western blotting analysis, in cerebellum, PAD1 levels were very low, although increased (non-significant) in the PD model, while PAD2 levels were similar in both groups. A significant increase was observed for PAD3, PAD4 and PAD6 in the PD model, indicating their possible contribution to increased citrullination hits in this brain region in the PD model and differences in the associated KEGG and GO pathway analysis of the respective cerebellum citrullinomes.
In midbrain, PAD1 protein levels were negligible, while PAD2 and PAD6 positive signals were more clearly detected, but PAD3 and PAD4 at lower level. There was some trend for decreased PAD2 and PAD4 protein levels in the PD model, which correlates with fewer citrullinated protein hits identified in PD midbrain, and this also correlated with fewer KEGG pathways unique to PD midbrain, compared with controls. It may be postulated that PAD-mediated effects on the substantia nigra (SN) at this stage may be negligible, but this will require further investigation. In a study on post-mortem human PD SN specimens, the detection of deiminated proteins in astrocytes was variable in both PD and controls SN specimens but deiminated proteins were present in the cytoplasm of SN dopamine neurones in PD samples [3].
In olfactory bulb, PAD2 and PAD6 showed a positive signal, while the other PADs were detected at lower levels, and a significant increase in PAD3 was observed in the PD group, also with a trend in increased PAD1, PAD4 and PAD6 protein levels. When assessing the citrullinomes, no specific KEGG pathway was associated with the citrullinome of the PD group only, and more citrullinated hits were overall identified in the control group, indicating that there is some modulation towards less citrullination in olfactory bulb in the early PD model; this will require further investigation.
In striatum, a positive signal for all PAD isozymes was observed, with significant reduction of protein levels for all PADs in PD striatum, when normalizing the region for PAD positive bands with the beta-actin loading control. There was indeed more citrullination observed in striatum of the control/sham group, but still some differences in target protein and associated KEGG pathways between PD and controls; with 3 unique KEGG pathways associated with the PD striatum.
The findings of our current study compare with our previous study, assessing cortex and hippocampal brain tissue sections from the same rat model by immunohistochemistry, where we identified most notable increased protein citrullination by positive F95 staining, in both cortex and hippocampus of the PD model, particularly in the brain vasculature, compared with controls [4]. This correlates to our citrullination enrichment results, showing most citrullinated hits from these two brain regions. In our previous study we had also identified by immunohistochemistry that histone H3 was particularly increased in the dentate gyrus of the rat pre-motor PD model [4] and this correlates with the findings of our current study in which LC MS/MS analysis identified histone H3 as a citrullination target in PD hippocampus, while it was not present on the hit lists for the other brain regions. Histone H3 citrullination/deimination contributes to epigenetic regulation but also extracellular trap formation (ETosis), in relation to pathogenic, autoimmune and inflammatory conditions, including in brain injury [23,42]. Inhibition of pan-citrullination, including histone H3 citrullination, with pan-PAD inhibitor Cl-amidine has been effective in reducing inflammatory responses in the CNS [23,27]. Studies from other groups have reported increased protein citrullination/deimination in post-mortem PD brains, including in surviving dopamine neurones in the SN, albeit at later stages of disease, while such staining was also reported not to be specifically restricted to Lewy bodies [3]. A study using F95 staining linked misfolded mutated alpha-synuclein protein to increased citrullination [43], and post-mortem analysis of prefrontal cortex from X-lined dystonia Parkinsonism patients showed elevation of PAD2 and PAD4 and increased histone H3 [44]. Notably, those PD related studies did not assess all five PAD isozymes.
It must furthermore be noted that in our previous study on the rat pre-motor PD model [4], only PAD 2,3 and 4 were assessed by immunohistochemistry, while in this current study, all five PAD isozymes were assessed in the same rat pre-motor PD model by western blotting. Our findings provide additional information on PAD1 and PAD6, indicating notable protein levels of PAD1 in striatum, but negligible detection in the other brain regions, and presence of PAD6 in all brain regions. Staining of post-mortem human brain samples of cortex and hippocampus, showed an increase of PAD1 levels in PD samples at Braak stage 4 and incidental Lewy body disorder [5]. No difference was observed in both regions between the two experimental groups here, however, PAD1 was shown to be increased in the cerebellum and olfactory bulb of the PD model compared with controls/shams and decreased in the striatum of the PD model. However, as the anti-PAD antibodies used in both studies are generated against human PAD isozymes, some differences in reactivity for species specificity and furthermore, differences between the pre-motor PD rat model and human post-mortem PD brain samples at different Braak stages may be likely.
In the context of dissecting the above identified changes in PAD isozyme detection by western blotting analysis and citrullinome patterns by LC-MS/MS analysis, considerations must include the fact that unchanged PAD protein levels do not necessarily reflect the calcium-catalysed activation of the PADs, which is necessary for citrullination/deimination of target proteins. In addition, a possible increase in some PAD isozymes at the protein level may allow for more availability of that specific isozyme for citrullination activity and affect the associated downstream citrullinome. Furthermore, as the PAD isozymes differ in their preference for target proteins [45], changes in individual isozymes in different brain regions may affect different downstream citrullination targets and contribute to brain-region specific citrullinomes, both linked to physiological as well as pathobiological processes. Redundancy of isozymes should also be considered; for example, PAD4 has a narrower target selection than PAD2, but with many overlapping targets, and interestingly, PAD6 has been shown to be upregulated in PAD2 and PAD4 knock-out mice [19].
In addition to assessing PADs, we used western blotting to assess changes in prohibitin, which is a mitochondrial housekeeping protein and was identified as a citrullinated protein target in PD cortex, and in both control cortex and hippocampus, by LC-MS/MS analysis. Prohibitin has previously been identified as a deimination target in the CNS [28] and while we observed some reduced levels of prohibitin in PD cortex and hippocampus, some elevation was observed in PD cerebellum and a significant increase in PD olfactory bulb. No change was seen in striatum or midbrain. Additional studies will be needed to determine whether citrullination differences in prohibitin further influence mitochondrial function in PD.
3.2. Brain Region Specific Citrullinomes
A considerable number of associated physiological and pathological KEGG and GO pathways was characterised when assessing the brain region specific citrullinomes. Some overlapped between sham/control and the PD model within each brain region, however, some were identified as common across various brain regions. In addition, some were only identified in PD brains, and others only in control brains. In addition to considering citrullinome associated pathways identified in PD brains only, citrullinated pathways that were identified in both control and PD, or in control brains only, should be taken into consideration. Our findings showed a loss of some citrullination associated KEGG pathways in PD, that were only identified in the controls, and this may indicate important roles for PADs in various physiological functions, which may be modified in PD and suggest unfavourable changes upon loss of citrullination of physiological pathways in PD. The various citrullinome associated KEGG pathways identified as shared, in PD only or in controls only, are summarised and briefly discussed in 3.2.1, 3.2.2 and 3.2.3.
3.2.1. Citrullinome KEGG Pathways Shared in Sham/Control and PD Brains
KEGG pathways that were overlapping for some of the control and PD brain region specific citrullinomes and related directly to neurodegeneration included: “Parkinson’s disease”; “Huntington disease”; “Prion disease”; “Alzheimer’s disease”; and “Amyotrophic lateral sclerosis”. PADs and citrullination have been shown in all these neurodegenerative diseases [13,14,16,17]. Additional overlapping KEGG pathways associated to neurodegeneration were “Cellular senescence”, which is an age-associated risk factor for inducing neurodegenerative diseases, including PD [46]; “Retrograde endocannabinoid signalling”, which is a lipid-based neuromodulatory system with physiological and neurodegenerative associates in the CNS, including in PD, AD, HD, MS and ALS [47,48,49,50]; “Gap junction”, which play critical roles in CNS signalling and have received considerable attention in PD [51]; “Regulation of actin cytoskeleton”, which is critical in regulating cellular morphology and function in normal physiology and is modulated in neurodegenerative diseases [52]; “cGMP-PKG signalling pathway”, which is critical for regulation of neuronal cell survival and apoptosis, including in PD [53]; “Synaptic vesicle cycle” and “SNARE interactions in vesicular transport” which are critical for the regulation of exo- and endocytosis of vesicles in neuronal communication and retrograde transport, also linked to LRRK2 in PD [54,55]; “GABAergic synapse” and “Dopaminergic synapse”, both of which are strongly involved in dopamine transmission in health and disease, and well-studied in PD models [56], as well as “Cholinergic synapse” which plays key roles in CNS synaptic function and associated to synaptic and axon degeneration and cognitive decline, including in PD [57,58]. Further shared KEGG pathways between some control and PD regions were “Serotonergic synapse”, which is associated with motor and non-motor PD symptoms and an identified risk factor for PD [59] and “Long-term depression” which is well recognised as a clinical symptom in PD [60] however, underlying mechanisms are not well understood [61]. In addition, “Cocaine addiction”, “Alcoholism”, and “Morphine addiction” were also identified, and these link to the dopaminergic system and neurodegenerative disease, including PD [62,63,64]. The “Oxytocin signalling pathway” has multifaceted roles including in brain function, in neuroinflammation and various nervous system disorders, including PD [65,66]. “HIF-1 signalling” was also a shared KEGG pathway and has been implicated in PD relating to mitochondrial dysfunction, oxidative stress and protein degradation impairment [67,68]; as well as “Endocrine and other factor-regulated calcium reabsorption” and “Calcium signalling pathways” which underly PAD-activation but also catalyse many other neurodegenerative downstream pathways, including protein aggregation, mitochondrial and other organelle crosstalk pathways in PD [69,70,71,72]. In addition, “Ferroptosis” links to glia-neuron crosstalk and has potential multifaceted roles in PD pathology [73,74,75,76]; while “Arginine biosynthesis” is associated with the gut-brain axis crosstalk, including in PD [77]. Identification of the “Ribosome” pathway in the shared citrullinome analysis highlights possible roles for PADs in local protein synthesis in the brain [78], and may also influence mitochondrial function including in PD pathology [79]; while “Thermogenesis”, is related to mitochondrial function, synaptic transmission, neurodegeneration and activation of brown fat tissue in PD [80]. In addition, shared citrulliome KEGG pathways incuded “Glycolysis/Gluconeogenesis”, and “Citrate cycle (TCA cycle)”, both of which are important for ATP production, mitochondrial and neuronal function and highlighted as a target in neurodegenerative diseases, including PD [81,82,83]. Furthermore, “Glucagon signalling pathway”, “Type II diabetes mellitus”; and “Insulin secretion” have been identified to be linked target pathways for treatment in neuroinflammation, including in PD [84] and mitochondrial dysfunction associated links have also been made to “non-alcoholic fatty liver disease” [85].
Additional metabolic pathways with critical roles in cellular function and which were identified as shared between control and PD brain citrullinomes were: “Biosynthesis of amino acids”, “Carbon metabolism”, “Nitrogen metabolism”, “Pyruvate metabolism”, “Propanoate metabolism”; “2-Oxocarboxylic acid metabolism”; “Pentose phosphate pathway”; “Fructose and mannose metabolism”; “Alanine, aspartate and glutamate metabolism”; “Cysteine and methionine metabolism”; “Glyoxylate and dicarboxylate metabolism”; “Vasopressin-regulated water reabsorption”; and “Gastric acid secretion”. Importantly, PD has been linked to various metabolic disorders, including at early stages of disease [86,87]. “Arrhythmogenic right ventricular cardiomyopathy” and “Cardiac muscle contraction” were furthermore associated to the shared KEGG citrullinomes, and this may be of interest as changes to cardiac function and cardiac dysfunction are observed in PD, with heart failure being the third leading cause of death in PD patients [88].
Shared control and PD brain citrullinome KEGG pathways linked to immunity and infection included: “Necroptosis”, “Endocytosis”, “Phagosome”; “Bacterial invasion of epithelial cells”; “Salmonella infection”; “Legionellosis”; “Staphylococcus aureus infection”; “Leukocyte transendothelial migration”; and “Viral carcinogenesis”. Phagocytic and bactericidal activities are modulated in neurodegeneration, where infection with various pathogens, including bacteria and viruses, are a topic of interest in the risk associated with developing PD, following infection [89]. This may also be of considerable interest in relation to long-term neurological outcomes from SARS-CoV-2 infection [90,91]. “Human immunodeficiency virus 1 infection (HIV-1)” was also a common shared citrullinome KEGG pathway and is associated to neurocognitive disorders (HIV-associated neurocognitive disorder (HAND)), characterised by synaptic loss and cognitive decline [92], and HIV infection is also thought to exacerbate age related brain disorders, including PD [93]. In PD, neuroprotective roles have been identified for the “Oestrogen signalling pathway”, which was an identified citrullinome KEGG pathway, and may link to observed sex differences identified in PD [94,95]. “Systemic lupus erythematosus” was also a shared KEGG citrullinome pathway for control and PD brains, and is an autoimmune multisystemic disease with complex interactions with PD and also associated to other cerebrovascular diseases [96,97,98].
3.2.2. Citrullinome KEGG Pathways in PD Brains Only
When excluding any overlap with other control brain regions, KEGG pathways only identified in PD brain region associated citrullinomes were: “Axon guidance”, which is critical to neuronal function, connectivity and repair throughout the lifespan and in neurodegeneration including in PD [99,100,101]; and “Spinocerebellar ataxia”, which is a heterogenous group of progressive neurodegenerative ataxic disorders [102] and identified here for the first time in relation to citrullination. In addition, PD specific brain citrullinome pathways included “Inositol phosphate metabolism”, which has been linked to neuronal cytotoxicity in PD, including via increased mitochondrial Ca2+ [103]; “Thyroid hormone signalling pathway”, which has been linked to neurological disorders including several models of PD [104]; “NOD-like receptor signalling pathway”, which forms part of the inflammasome axis in several neurodegenerative disease including PD [105,106,107]; “Phosphatidylinositol signalling system”, which has been linked to neuroinflammatory responses, including autophagy, and their regulation in neurodegeneration and PD [108,109]; and “Platelet activation”, which are related to synaptic plasticity and neuronal differentiation in various brain regions and also reflective of inflammatory responses in neurodegeneration, including PD [110,111,112]. Further PD citrullinome specific pathways were “Hippo signalling pathway”, which is an evolutionary conserved signalling network with crucial roles in various biological processes including proliferation and differentiation, regeneration, development and immunity [113,114], and linked to early neurodegenerative processes, including in PD, AD, HD [115,116,117]; “Rap1 signalling pathway”, which has been identified as a molecular pathway in PD patients [118], including relating to blood markers in early PD [112]; “Fc gamma R-mediated phagocytosis”, which plays roles in brain development, is linked to neurodegenerative disease development [119] and has been identified as dysregulated in AD [120] ; and “Choline metabolism in cancer”, which may be relevant as choline metabolism has been linked to the gut-brain signalling axis, which is linked to neurodegenerative disease, including PD [121,122]. In addition, citrullinome KEGG pathways in PD brains only were“Human cytomegalovirus infection”, which has been associated to numerous neurological disorders including PD, AD, HD, autism, ataxia and brain tumours [123] and identified as one of several infectious pathogens in the aetiology of PD [124]; “Yersinia infection”, which may be relevant in relation to age associated changes in gut microbiota and possible increase susceptibility to PD [125]; “Progesterone-mediated oocyte maturation”, which has been linked to molecular mechanisms involved in PD via microarray analysis [126]; as well as “Oocyte meiosis”, which may link to studies on the PD associated LLRK2 kinase family, which has also been shown to have roles in oocyte meiosis via regulation of actin assembly and mitochondrial function [127]. The detection of PAD6 in all brain regions may possibly link to some of the developmental associated KEGG pathways identified in the brain citrullinome, given its known roles in developmental processes [128,129].
3.2.3. Citrullinome KEGG Pathways in Sham/Control Brains Only
There were some KEGG pathways associated with the citrullinomes of control brain regions only, which were not identified in brains of the PD group. These were: “Phospholipase D signalling pathway”, which plays important roles in normal brain function including regulation of the synaptic vesicle cycle in neuronal communication, neuronal morphogenesis, cytoskeleton modulation, neural stem/progenitor cell differentiation, and is a suggested therapeutic target in brain disorders, including PD and AD [54,130,131,132,133]; as well as “Butanoate metabolism” and “Beta-Alanine metabolism”, both of which are associated with the gut-brain axis, including in PD [77]. Additional citrullinome KEGG pathways in control brains only included “GnRH secretion”, which is related to brain connectivity including neuron maturation, synaptic transmission, cognition and olfaction, and identified as a therapeutic target in Down syndrome [134] and reported to stimulate histone citrullination and cytoskeletal dynamics [135]; “Collecting duct acid secretion”, which plays roles in acid-base homeostasis regulation [136]; and “Apelin signalling pathway”, which is related to multifaceted cellular regulatory roles, including in the hypothalamus, in neuronal function, neuroinflammation and neurodegenerative disease, including neuroprotective effects in PD pathogenesis [137,138,139,140]. Further citrullinome KEGG pathways in control brain regions only were “Spliceosome”, which are involved in the generation of circular RNAs and influence transcription, and are related to ageing, neuroinflammation, oxidative stress, and have been suggested as diagnostic and prognostic biomarker for neurodegenerative disease including PD [141,142]; “mTOR signalling pathway”, which is critical for the regulation of autophagy, apoptosis and cell proliferation, in gut brain axis signalling and plays important roles for neuronal survival including in PD, where it has been identified as a therapeutic target [143,144,145,146]; “Antigen processing and presentation”, which may link to brain homeostasis but also inflammatory responses, and has recently been highlighted in autoimmune features in neurodegeneration, including in PD [147,148,149]; “Influenza A”, which can infect the CNS and spread through the brain, and suggested as a possible factor inducing Lewy bodies in PD [150]; and “Protein processing in endoplasmic reticulum” which is critical for biosynthesis of proteins, their folding and assembly and protein quality control, while in neurodegenerative disease, including PD, this pathway plays roles in the unfolded protein response [151,152].
Overall, our findings highlight enrichment for citrullinated proteins associated to KEGG pathways for several neurodegenerative diseases, possibly indicating shared pathogenic mechanisms which may be differently modulated by citrullination in health and disease [4,153]. Collectively the identification of the above listed KEGG pathways associated with citrullinated proteins in control and/or pre-motor PD brain regions, highlights some differences between molecular and cellular pathways modulated by this post-translational modification, in the different brain regions, and may be relevant to PD disease progression.
In our previous study on the same pre-motor PD rat model [4], circulatory citrullination signatures both in plasma and plasma extracellular vesicles (EVs), and associated KEGG pathways were assessed, and some correlation to KEGG pathways identified in the different brain regions in this current study can be made. Previously we identified “Parkinson’s disease”, “Huntington disease”, “Prion disease”, “Alzheimer’s disease”, “Retrograde endocannabinoid signalling”, “Oxidative phorphorylation”, “Oestrogen signalling pathway”, “Non-alcoholic fatty liver disease”, “SLE”, “Complement and coagulation cascades”, “Metabolic pathways” and “Apelin signalling” as citrullinome associated KEGG pathways in PD plasma and/or plasma-EVs [4]. Many of these KEGG pathways correspond to those identified for the citrullinomes of the PD brain regions in the current study, while some overlap with the control brain samples. The identification of which pathways in the circulatory plasma citrullinome can be best correlated with relevant changes in citrullinome signatures of the brain, and with respect to specific brain regions, will need further validation.
Studies from other groups have reported increased protein citrullination/deimination in post-mortem PD brains, including in surviving dopamine neurones in the SN, while such staining was reported not to be specifically restricted to Lewy bodies [3]. A study using F95 staining linked misfolded mutated alpha-synuclein protein to increased citrullination [43], and post-mortem analysis of prefrontal cortex from X-lined dystonia Parkinsonism patients showed increased PAD2 and PAD4 levels and histone H3 citrullination, [44]. Notably, those PD related studies did not assess all five PAD isozymes, contrary to our current study. Furthermore, an assessment of citrullinome signatures in PD brains and in different brain regions of PD, including PD models, has not been carried out before. In other neurodegenerative disease studies, PAD2 has been linked to Alzheimer’s disease, prion disease and amyotrophic lateral sclerosis [13,14,16,17], and in these studies other PAD isozymes have often not been assessed, partly due to previous brain studies focussing on PAD2 [12]. In the naked mole-rat, which is a hypoxia resistant animal, all PAD isozymes and the brain citrullinome were assessed, showing increased citrullination and elevated PAD1, PAD3 and PAD6 but reduced PAD2 and PAD4, and modifications in the brain citrullinome, following hypoxia challenge [40]. Increased citrullination has also been reported in the CNS in response to blast injury and traumatic brain injury [22,154], including with respect to different citrullination levels and targets between brain regions [22]. Differences in brain citrullinome protein targets between white and gray matter have been assessed in MS versus control brains [21] and sex-related differences have been identified in AD [155].
3.3. Possible Roles for Citrullination in the Gut-Brain Axis in PD
Interestingly, various infection related pathways, inflammatory, and gut-brain axis pathways were identified as citrullinated in the pre-motor PD brains in the current study. The neuro-microbiology of PD has received considerable interest [156]. Microglial activation and inflammatory responses linked to various pathogens, including parasitic, viral, and bacterial agents have been reported [89]. Several infection and pathogen linked KEGG pathways were identified in the current study linked to the brain citrulliome of both control and pre-motor PD rats. Whether and how citrullination may play roles in escalating neuroinflammatory responses and downstream neurodegeneration in relation to such infection-associated pathways may be of considerable interest in future studies. The central roles for PADs in the CNS, and their roles in various bacterial and viral infections, some of which are also related to neurological diseases [31,157,158,159] are of considerable interest in this context. This also includes long-term neurological outcomes in COVID-19 [90], which also has a gut-brain axis element, highlighting that further investigations may be of considerable interest in relation to downstream neurodegenerative pathways, including PD related ones [91,160,161,162].
The gut, nasal and oral microbiota have received considerable attention in PD [163,164], but studies in relation to PADs and citrullination are still scarce. Porphyromonas gingivalis is strongly associated with citrullination in periodontitis [165] and has indeed been identified as one of various microbes in nervous tissues from several animal models of neurodegenerative diseases, including ALS, AD, and PD [166]. Importantly, bacterial presence of P. gingivalis, which itself has citrullinating activity and may affect the host citrullinome [167], has been identified in the brain of both AD and PD based on 16S rRNA next generation sequencing, assessing early, intermediate, and late stage of the diseases. In previous studies, P. gingivalis has been reported in hippocampus and cortex from control and PD donors [168], reported to be mainly of oronasal origin. Interestingly, in AD, modification of the oral microbiome appears to be more prominent than in the gut, and AD studies have also focussed on P. gingivalis [169]. It was shown that oral gingivitis impairs gut permeability and mediates immune responses associated with neurodegeneration in LLRK2 PD mouse models of late onset PD, and chronic periodontitis is a common type of peripheral inflammation associated with PD. It remains to be established whether P. gingivalis induced dysbiosis plays a role in pathophysiology of PD. Mice receiving oral P. gingivalis showed reduced dopaminergic neurones in the SN and activation in microgial cells [170]. Interestingly, PD has been shown to alter the composition of the subgingival microbiome of periodontitis [171]. P. gingivalis has also been linked to rheumatoid arthritis (RA), one of the most extensively studied PAD-pathology to date, including in association with periodontal disease [172]. P. gingivalis has also been shown to accelerate atherosclerosis [173]. In RA, citrullination is identified in relation to intestinal permeability and microbial imbalance [174], and it must furthermore be considered that many other microbes may be able to cause citrullination of proteins in their hosts (both commensals and pathogens) due bacterial PAD-homologues (ADI) [175,176], and this may contribute to citrullination mediated inflammatory responses exacerbating various pathologies.
The composition of the fecal microbiota in the PD model treated with placebo has been shown to be different from that in the sham animals and to be correlated with increased plasma levels of inflammatory markers and neuroinflammation [177]. Further studies will be needed to characterise the possible presence and function of P.gingivalis in the PD model. In addition, it may be of interest in future studies to assess oral, nasal and gut citrullinomes in PD to establish which proteins in the brain can be modified by citrullination mediated by P. gingivalis, or other types of infectious agents or commensals. The origin of PD onset has been debated, with some indication of origin in the gut, and PD is highlighted as a systemic inflammatory disease which is accompanied by bacterial inflammagens [178]. Hence the question remains whether some of the suggested gut-bran axis involvement in PD may be linked to citrullination and while this has not been studied yet, various studies have highlighted potential roles for P. gingivalis in PD [179], including in PD with cognitive impairment [180].
3.4. Future Prospects for PAD Inhibitors in PD
Epigenetic mechanisms in PD, including post-translational modifications, are receiving increased interest as highlighted in recent studies [4,5,181,182,183,184]. As PADs may be a promising target in PD therapeutics, roles for the different isozymes must be better understood for the development of therapeutics utilising pan-PAD or PAD isozyme-specific targeting. Pharmacological PAD inhibitors include pan-PAD inhibitors Cl-amidine and BB-Cl-amidine, PAD2 inhibitor AMF30a, PAD3 inhibitor Cl4-amidine and PAD4 inhibitor GSK199 [45]; some of which have been applied in various CNS in vivo and in vitro models [23,25,27,28,29,185]. Considering choosing pan-PAD versus isozyme specific inhibitors may be important as PAD isozymes have different, and sometimes overlapping, preferences for target proteins and hence understanding of the physiological and disease related citrullinomes is also of importance. Future approaches for clinical PAD inhibitor treatment, both aimed at modulating total deimination via pan-PAD inhibitors, or a narrower range of deimination targets using PAD isozyme-specific inhibitors, still require further refinement and optimization in experimental models.
This is the first study to attempt a detailed mapping of citrullinome changes in different brain regions in PD, in this instance using a pre-motor toxin-induced PD rat model, and to identify brain region specific differences in PAD isozyme expression. Findings of this study overall align with other studies reporting modulation of PADs and citrullination in neurodegenerative diseases. It will be important, in continuation of this current study and previously published studies, to identify citrullination patterns in samples from human PD cohorts and linking possible changes in circulatory PD signatures (plasma-EVs) to citrullination changes in brains. However, this will remain challenging as the citrullinome signatures from brains are only retrievable from post-mortem human samples. Comparisons with 6-OHDA in vitro human cell cultures, in addition to other PD animal models, including the A53T alpha-synuclein mouse model which is linked to early-onset PD, may nevertheless provide some information to help move the field forwards. It will also be of interest in future studies to compare outcomes in the 6-OHDA-PD model to the LPS-PD model, which has recently highlighted the replication of crosstalk between local and systemic inflammatory response [111], which are inherent in PD pathogenesis and pathophysiology.
Based on recent studies and our current findings reported here, evidence is mounting for considerable roles for PADs and citrullination in PD, including at early stages of the disease. By increasing our understanding of PAD-mediated brain-region specific changes in disease progression, we will gain a better picture of the spatio-temporal roles for this post-translational modification in PD and its therapeutic potential.
4. Materials and Methods
4.1. Pre-Motor PD Rat Model
This study employed the 6-hydroxydopamine (6-OHDA) rat model to induce pre-motor Parkinson’s disease (PD) in male Sprague-Dawley rats (200-250g), adhering to ethical guidelines and regulatory approvals, including clearance from the Bloomsbury ethical committee and the Home Office as per the Animal Scientific Procedures Act 1986 (PPL PP3144142).
The model, displaying non-motor symptoms without motor dysfunction, has been previously described in detail [4,186]. In brief, rats were randomly allocated to two experimental groups: Sham-treated (control) and toxin-treated model (PD); n=3 animals per group were used for this study. The pre-motor PD model was established by intraperitoneal administration of either N-(2-chloroethyl)-N-ethyl-2-bromobenzylamine (DSP-4 - 25mg/kg for the PD model) or sterile saline (for sham/controls) followed by bilateral striatal injections of 6-OHDA (15mg per striatum - coordinates from Bregma: AP +1.0 mm, ML +3.0 mm, DV −6.5 mm – PD model) or saline (sham) containing 0.9% of ascorbic acid under general anaesthesia. Rats were maintained for 3 weeks after surgery and were then perfused. Brains were removed and the different brain regions of interest (cortex, hippocampus, midbrain, striatum, cerebellum, and olfactory bulb) were dissected, snap-frozen in liquid nitrogen, and stored at -80°C for subsequent protein analysis. The experimental setup is summarized in Figure 12.
4.2. Protein Isolation from Brain Tissue
Proteins were extracted from brain tissue from the different brain regions of PD induced and control animals (n=3 per group) according to previously described methods [40]. The six different brain regions under study (cortex, hippocampus, midbrain, striatum, cerebellum, and olfactory bulb) were individually homogenised in RIPA+ buffer (Sigma-Aldrich, Gillingham, UK, containing 10% protease inhibitor cocktail, Sigma-Aldrich) in 2 mL Eppendorf tubes on ice using a Mini Handheld Homogeniser (Kimble, DWK Life Sciences, VWR International). The homogenates were then gently pressed through a 23G needle into fresh Eppendorf tubes on ice, followed by gently pipetting up and down to eliminate any tissue clots. For each 100 mg of tissue, 500 µL of RIPA+ buffer was used. The brain tissue homogenates were incubated for 1.5 h at 4 °C on a rolling platform, pipetting up and down at regular intervals. For protein isolation, the homogenates were then centrifuged at 16,000× g for 30 min at 4 °C, collecting the protein containing supernatant, which was aliquoted and immediately frozen at −80 °C until further use.
4.3. Western Blotting
For Western blotting, a 100 µL aliquot of protein extract per sample was diluted with 100 µL 2× reducing Laemmli sample buffer (BioRad; containing 5% β-mercaptoethanol, Sigma-Aldrich) and boiled for 5 min at 100 °C. Samples were run by SDS-PAGE (4–20% TGX gels, BioRad, Watford, UK) at 165 V for 52 min, using a 5 µL aliquot per sample per lane. Proteins were transferred to nitrocellulose membranes using semi-dry transfer (1h at 15V), assessing even protein transfer by PonceauS red stain (Sigma-Aldrich) before blocking in 5% bovine serum albumin (BSA, Sigma-Aldrich) in TBS-T for 1 h at room temperature (RT). For detection of PAD isozymes, the membranes were incubated in primary antibodies overnight at 4 °C on a shaking platform as follows: PAD isozyme specific antibodies used were anti-human PAD1 (ab181762, Abcam Cambridge, UK,), PAD2 (ab50257), PAD3 (ab50246), PAD4 (ab50247) and PAD6 (PA5–72059, Thermo Fisher Scientific, Hemel Hempstead, UK). The mitochondrial housekeeping protein prohibitin was also assessed using anti-prohibitin (ab75771) on western blotting of the protein extracts from all six brain regions. All primary antibodies were used at 1/1000 dilution in TBS-T. Washing was carried out with TBS-T (3 × 10 min). Secondary antibody incubation was completed for 1 h at RT (using HRP-labelled anti-rabbit IgG; BioRad, diluted 1/3000 in TBS-T). Following washing (5 × 10 min in TBS-T), visualisation was carried out using ECL (Amersham Biosciences, Buckinghamshire, UK) and the UVP BioDoc-ITTM System (Thermo Fisher Scientific, Dartford, UK). All blots were re-probed with HRP-conjugated anti-β-actin antibody (ab20272, Abcam, 1/5000 in TBS-T), developed and imaged. For quantitative analysis of PAD isozymes and prohibitin, regions for protein bands in the expected size range of 70-75 kDa for PAD1-4, and 50-60 kDa for PAD6, as well as in the 27 kDa range for prohibitin,were normalised against β-actin positive bands following densitometry analysis using ImageJ.
4.4. Isolation of Citrullinated Proteins from Brain Tissue
To identify the brain-region specific citrullinomes, immunoprecipitation was carried out to isolate citrullinated/deiminated proteins from the different brain regions’ protein isolates using the F95 pan-citrulline antibody (MABN328, Merck, Watford, UK) [187]. For a representative citrullinome of each brain region (cortex, hippocampus, striatum, midbrain, cerebellum, olfactory bulb), protein extracts from 3 brains per experimental group (PD versus sham) were pooled (3 x 20 µL). Immunoprecipitation was carried out using the Catch and Release® v2.0 Immunoprecipitation Kit (17-500M, Merck) together with the F95 pan-citrulline antibody and the affinity ligand provided with the kit, according to the manufacturer’s instructions (Merck). F95 enrichment was carried out overnight, incubating the mini-IP columns at 4 °C on a rotating platform. Thereafter the citrullinated F95 bound proteins were eluted with the elution buffer provided with the kit, according to the manufacturer’s instructions (Merck) and assessed by SDS-PAGE and silver staining (BioRad Silver Stain Plus Kit) for protein yield and by LC-MS/MS analysis for the identification of individual citrullinated protein hits.
4.5. Liquid Chromatography with Tandem Mass Spectrometry (LC-MS/MS)
For LC-MS/MS analysis, the F95-enriched eluates from each brain region were run 0.5 cm into a 12% TGX gel (BioRad) and thereafter cut out as one band each, respectively; followed by in-gel digestion (Cambridge Proteomics, Cambridge, UK), according to previously described methods [4,40]. In brief, automated LC-MS/MS analysis was carried out using a Dionex Ultimate 3000 RSLC nanoUPLC (Thermo Fisher Scientific Inc., Waltham, MA, USA) system in conjunction with a QExactive Orbitrap mass spectrometer (Thermo Fisher Scientific Inc., Waltham, MA, USA). Peptide separation was carried out using reverse-phase chromatography and a Thermo Scientific reverse-phase nano Easy-spray column (Thermo Fisher Scientific Inc). The LC eluent was sprayed into the mass spectrometer using an Easy-Spray source (Thermo Fisher Scientific Inc.). The m/z values of all eluting ions were measured in an Orbitrap mass analyzer, data dependent scans (selecting top 20) were employed for automatic isolation and generation of fragment ions using the HCD collision cell, measured using the Orbitrap analyser. Both singly charged ions as well as ions with unassigned charge states were excluded from selection for MS/MS. A dynamic exclusion window of 20 sec was also applied. Data were processed post-run using Protein Discoverer (version 2.1., Thermo Scientific), converted to mgf files and submitted Mascot (Mascot search algorithm; Matrix Science, London, UK). Search for hits was carried out against the UniProt Rattus_norvegicus_20181203 (31,558 sequences; 17,280,660 residues) database with peptide and fragment mass tolerances respectively set at 20 ppm and 0.1 Da. The threshold value for significance was set at p < 0.05, and the peptide cut-off score was set at 35.
4.6. Protein-Protein Interaction Network Analysis
To identify protein–protein interaction networks for rat specific citrullinated/deiminated proteins hits from the six different brain regions (brain-region specific citrullinomes), STRING analysis (Search Tool for the Retrieval of Interacting Genes/Proteins; https://string-db.org/) was used (accessed on 18th and 19th March 2024). The following functions were applied in STRING: “search multiple proteins”, the species database chosen was “Rattus norvegicus”, and “basic settings and medium confidence were applied. Colour lines between the nodes indicate the following evidence-based interactions for network edges: “known interactions” (based on curated databases, experimentally determined), as well as “predicted interactions” (based on gene neighbourhood, gene fusion, gene co-occurrence, or via text mining, co-expression, or protein homology). Data for the pathway analysis of the protein networks were exported as STRING network images and as Excel files for KEGG and GO pathways and compared between brain regions from PD and sham rats, respectively.
4.7. Statistical Analysis
For comparison between datasets from PD versus control brains, GraphPad Prism version 10 was used. T-tests were used to determine significance between groups for densitometry readings from western blotting analysis, showing mean and standard deviation (n = 3 per experimental group). Statistical significance was regarded as p < 0.05. STRING analysis was carried out with medium confidence in STRING (https://string-db.org/, accessed on 18th and 19th March 2024).
5. Conclusions
This is the first study to attempt a detailed mapping of citrullinome changes in different brain regions in PD, using a pre-motor toxin-induced PD rat model. Findings of the study highlight some brain region specific differences in PAD isozyme expression and the respective citrullinomes in cortex, hippocampus, striatum, midbrain and olfactory bulb of control versus pre-motor PD rat brains. We report both overlapping, control/sham and PD specific KEGG and Gene ontology (GO) pathways associated with the brain region specific citrullinomes relating to metabolic, immune, cell signalling and neurodegenerative disease related pathways. Our findings identify roles for PAD-mediated citrullination in physiological and pathobiological processes, including in early stages of PD, highlighting their potential for future therapeutic avenues.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1. LC-MS/MS analysis of F95-enriched proteins in cortex of sham-treated (control) rats. Table S2. LC-MS/MS analysis of F95-enriched proteins in cortex of pre-motor PD-model rats. Table S3. LC-MS/MS analysis of F95-enriched proteins in hippocampus of sham-treated (control) rats. Table S4. LC-MS/MS analysis of F95-enriched proteins in hippocampus of pre-motor PD-model rats. Table S5. LC-MS/MS analysis of F95-enriched proteins in cerebellum of sham-treated (control) rats. Table S6. LC-MS/MS analysis of F95-enriched proteins in cerebellum of pre-motor PD-model rats. Table S7. LC-MS/MS analysis of F95-enriched proteins in midbrain of sham-treated (control) rats. Table S8. LC-MS/MS analysis of F95-enriched proteins in midbrain of pre-motor PD-model rats. Table S9. LC-MS/MS analysis of F95-enriched proteins in olfactory bulb of sham-treated (control) rats. Table S10. LC-MS/MS analysis of F95-enriched proteins in olfactory bulb of pre-motor PD-model rats. Table S11. LC-MS/MS analysis of F95-enriched proteins in striatum of sham-treated (control) rats. Table S12. LC-MS/MS analysis of F95-enriched proteins in striatum of pre-motor PD-model rats.
Author Contributions
Conceptualization, S.L.; methodology, M.S., A.M., A.Mac. and S.L.; validation, S.L., A. Mac. and A.M.; formal analysis, S.L. M.S. and A.Mac.; investigation, A.M., M.S., A.Mac and S.L.; resources, A.M. and S.L.; data curation, S.L., A.Mac, M.S. and A.M.; writing—original draft preparation,S.L.; writing—review and editing, S.L., A.Mac and A.M.; visualization, S.L. and A.Mac.; supervision, S.L. and A.M.; project administration, S.L. and A.M.; funding acquisition, S.L. and A.M.. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partly funded by Quality-related Research (QR) funds from the University of Westminster (S.L.) and by internal funds from the UCL School of Pharmacy (A.M.).
Institutional Review Board Statement
All procedures in the study were approved by the Bloomsbury ethical committee and the Home Office and followed the British Home Office regulations with regard to the Animal Scientific Procedures Act 1986 (PPL PP3144142). All experiments were in compliance with the ARRIVE guidelines (Animal Research: Reporting in Vivo Experiments) for how to REPORT animal experiments. The animal study protocol was approved by the Bloomsbury AWERB of University College London (protocol 1 – PPL number PP3144142 granted 14 Jan 2020).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data supporting the study are included in the article and supplementary materials.
Acknowledgments
Thanks to Michael Deery and Yagnesh Umrania at the Cambridge Centre for Proteomics for performing the LC-MS/MS analysis.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Siderowf, A.; Lang, A.E. Premotor Parkinson’s disease: Concepts and definitions. Mov. Disord. 2012, 27, 608. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ciesielski, O.; Biesiekierska, M.; Panthu, B.; Soszyński, M.; Pirola, L.; Balcerczyk, A. Citrullination in the pathology of inflammatory and autoimmune disorders: Recent advances and future perspectives. Cell Mol Life Sci. 2022, 79. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, A.P. Dual immunofluorescence study of citrullinated proteins in Parkinson diseased substantia nigra. Neurosci Lett. 2011, 495, 26. [Google Scholar] [CrossRef] [PubMed]
- Sancandi, M.; Uysal-Onganer, P.; Kraev, I.; Mercer, A.; Lange, S. Protein Deimination Signatures in Plasma and Plasma-EVs and Protein Deimination in the Brain Vasculature in a Rat Model of Pre-Motor Parkinson’s Disease. Int J Mol Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Mercer, A.; Jaunmuktane, Z.; Hristova, M.; Lange, S. Differential, Stage Dependent Detection of Peptidylarginine Deiminases and Protein Deimination in Lewy Body Diseases-Findings from a Pilot Study. Int J Mol Sci. 2022, 23. [Google Scholar] [CrossRef]
- Witalison, E.E.; Thompson, P.R.; Hofseth, L.J. Protein arginine deiminases and associated citrullination: Physiological functions and diseases associated with dysregulation. Curr. Drug Targets 2015, 16, 700–710. [Google Scholar] [CrossRef] [PubMed]
- György, B.; Tóth, E.; Tarcsa, E.; Falus, A.; Buzás, E.I. Citrullination: A posttranslational modification in health and disease. Int. J. Biochem. Cell Biol. 2006, 38, 1662–1677. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, M.; Alasmari, D.; Assiri, A.; Mattar, E.; Aljaddawi, A.A.; Alattas, S.G.; Redwan, E.M. An Overview of the Intrinsic Role of Citrullination in Autoimmune Disorders. J. Immunol. Res. 2019, 2019, 7592851. [Google Scholar] [CrossRef]
- Vossenaar, E.R.; Zendman, A.J.; van Venrooij, W.J.; Pruijn, G.J. PAD, a growing family of citrullinating enzymes: Genes, features and involvement in disease. Bioessays. 2003, 25(11), 1106. [Google Scholar] [CrossRef]
- Magnadóttir, B.; Hayes, P.; Hristova, M.; Bragason, B.T.; Nicholas, A.P.; Dodds, A.W.; Guðmundsdóttir, S.; Lange, S. Post-translational protein deimination in cod (Gadus morhua L.) ontogeny novel roles in tissue remodelling and mucosal immune defences? Dev Comp Immunol. 2018, 87, 157. [Google Scholar] [CrossRef]
- Christophorou, M.A. The virtues and vices of protein citrullination. R Soc Open Sci. 2022, 9. [Google Scholar] [CrossRef]
- Shimada, N.; Handa, S.; Uchida, Y.; Fukuda, M.; Maruyama, N.; Asaga, H.; Choi, E.K.; Lee, J.; Ishigami, A. Developmental and age-related changes of peptidylarginine deiminase 2 in the mouse brain. J. Neurosci. Res. 2010, 88, 798–806. [Google Scholar] [CrossRef]
- Ishigami, A.; Ohsawa, T.; Hiratsuka, M.; Taguchi, H.; Kobayashi, S.; Saito, Y.; Murayama, S.; Asaga, H.; Toda, T.; Kimura, N.; Maruyama, N. Abnormal accumulation of citrullinated proteins catalyzed by peptidylarginine deiminase in hippocampal extracts from patients with Alzheimer’s disease. J. Neurosci. Res. 2005, 80, 120–128. [Google Scholar] [CrossRef]
- Ishigami, A.; Masutomi, H.; Handa, S.; Nakamura, M.; Nakaya, S.; Uchida, Y.; Saito, Y.; Murayama, S.; Jang, B.; Jeon, Y.C.; Choi, E.K.; Kim, Y.S.; Kasahara, Y.; Maruyama, N.; Toda, T. Mass spectrometric identification of citrullination sites and immunohistochemical detection of citrullinated glial fibrillary acidic protein in Alzheimer’s disease brains. J. Neurosci. Res. 2015, 93, 1664–1674. [Google Scholar] [CrossRef]
- Jang, B.; Jin, J.K.; Jeon, Y.C.; Cho, H.J.; Ishigami, A.; Choi, K.C.; Carp, R.I.; Maruyama, N.; Kim, Y.S.; Choi, E.K. Involvement of peptidylarginine deiminase-mediated post-translational citrullination in pathogenesis of sporadic Creutzfeldt-Jakob disease. Acta Neuropathol. 2009, 119, 199–210. [Google Scholar] [CrossRef]
- Jang, B.; Ishigami, A.; Maruyama, N.; Carp, R.I.; Kim, Y.S.; Choi, E.K. Peptidylarginine deiminase and protein citrullination in prion diseases: Strong evidence of neurodegeneration. Prion 2013, 7, 42–46. [Google Scholar] [CrossRef]
- Yusuf, I.O.; Qiao, T.; Parsi, S.; Tilvawala, R.; Thompson, P.R.; Xu, Z. Protein citrullination marks myelin protein aggregation and disease progression in mouse ALS models. Acta Neuropathol Commun. 2022, 10. [Google Scholar] [CrossRef]
- Raijmakers, R.; Vogelzangs, J.; Raats, J.; Panzenbeck, M.; Corby, M.; Jiang, H.; Thibodeau, M.; Haynes, N.; van Venrooij, W.J.; Pruijn, G. J.; Werneburg, B. Experimental autoimmune encephalomyelitis induction in peptidylarginine deiminase 2 knockout mice. J Comp Neurol. 2006, 498, 217. [Google Scholar] [CrossRef]
- van Beers, J.J.; Zendman, A.J.; Raijmakers, R.; Stammen-Vogelzangs, J.; Pruijn, G.J. Peptidylarginine deiminase expression and activity in PAD2 knock-out and PAD4-low mice. Biochimie. 2013, 95, 299. [Google Scholar] [CrossRef]
- Mastronardi, F.G.; Wood, D.D.; Mei, J.; Raijmakers, R.; Tseveleki, V.; Dosch, H.M.; Probert, L.; Casaccia-Bonnefil, P.; Moscarello, M.A. Increased citrullination of histone H3 in multiple sclerosis brain and animal models of demyelination: A role for tumor necrosis factor-induced peptidylarginine deiminase 4 translocation. J. Neurosci. 2006, 26, 11387–11396. [Google Scholar] [CrossRef]
- Faigle, W.; Cruciani, C.; Wolski, W.; Roschitzki, B.; Puthenparampil, M.; Tomas-Ojer, P.; Sellés-Moreno, C.; Zeis, T.; Jelcic, I.; Schaeren-Wiemers, N.; Sospedra, M.; Martin, R. Brain Citrullination Patterns and T Cell Reactivity of Cerebrospinal Fluid-Derived CD4+ T Cells in Multiple Sclerosis. Front. Immunol. 2019, 10, 540. [Google Scholar] [CrossRef]
- Lazarus, R.C.; Buonora, J.E.; Flora, M.N.; Freedy, J.G.; Holstein, G.R.; Martinelli, G.P.; Jacobowitz, D.M.; Mueller, G.P. Protein Citrullination: A Proposed Mechanism for Pathology in Traumatic Brain Injury. Front. Neurol. 2015, 6, 204. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.; Rocha-Ferreira, E.; Thei, L.; Mawjee, P.; Bennett, K.; Thompson, P.R.; Subramanian, V.; Nicholas, A.P.; Peebles, D.; Hristova, M.; et al. Peptidylarginine deiminases: Novel drug targets for prevention of neuronal damage following hypoxic ischemic insult (HI) in neonates. J. Neurochem. 2014, 130, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Lange, S. Peptidylarginine deiminases and extracellular vesicles: Prospective drug targets and biomarkers in central nervous system diseases and repair. Neural. Regen. Res. 2021, 16, 934–938. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Liu, L.; Cao, Y.; et al. Inhibition of neutrophil extracellular trap formation ameliorates neuroinflammation and neuronal apoptosis via STING-dependent IRE1α/ASK1/JNK signaling pathway in mice with traumatic brain injury. J Neuroinflammation. 2023, 20. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.; Wray, S.; Devine, M.; Matarin, M.; Hardy, J. Protein Deimination in Protein Misfolding Disorders–Modelled in Human Induced Pluripotent Stem Cells (iPSCs). In Protein Deimination in Human Health and Disease; Nicholas, A.P., Bhattacharya, S.K., Eds.; Springer Science and Business Media: New York, NY, USA, 2017; Volume 2. [Google Scholar]
- Lange, S.; Gögel, S.; Leung, K.Y.; Vernay, B.; Nicholas, A.P.; Causey, C.P.; Thompson, P.R.; Greene, N.D.; Ferretti, P. Protein deiminases: New players in the developmentally regulated loss of neural regenerative ability. Dev. Biol. 2011, 355, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Kosgodage, U.S.; Uysal-Onganer, P.; MacLatchy, A.; Kraev, I.; Chatterton, N.P.; Nicholas, A.P.; Inal, J.M.; Lange, S. Peptidylarginine Deiminases Post-Translationally Deiminate Prohibitin and Modulate Extracellular Vesicle Release and MicroRNAs in Glioblastoma Multiforme. Int. J. Mol. Sci. 2018, 20, 103. [Google Scholar] [CrossRef]
- Uysal-Onganer, P.; MacLatchy, A.; Mahmoud, R.; Kraev, I.; Thompson, P.R.; Inal, J.M.; Lange, S. Peptidylarginine Deiminase Isozyme-Specific PAD2, PAD3 and PAD4 Inhibitors Differentially Modulate Extracellular Vesicle Signatures and Cell Invasion in Two Glioblastoma Multiforme Cell Lines. Int. J. Mol. Sci. 2020, 21, 1495. [Google Scholar] [CrossRef]
- U, K.P.; Subramanian, V.; Nicholas, A.P.; Thompson, P.R.; Ferretti, P. Modulation of calcium-induced cell death in human neural stem cells by the novel peptidylarginine deiminase-AIF pathway. Biochim Biophys Acta. 2014, 1843, 1162. [Google Scholar] [CrossRef]
- Pasquero, S.; Gugliesi, F.; Biolatti, M.; et al. Citrullination profile analysis reveals peptidylarginine deaminase 3 as an HSV-1 target to dampen the activity of candidate antiviral restriction factors. PLoS Pathog. 2023, 19. [Google Scholar] [CrossRef]
- Méchin, M.C.; Takahara, H.; Simon, M. Deimination and peptidylarginine deiminases in skin physiology and diseases. Int. J. Mol. Sci. 2020, 21, 566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, X.; Zhang, M.; Li, T.; Muth, A.; Thompson, P.R.; Coonrod, S.A.; Zhang, X. Peptidylarginine deiminase 1-catalyzed histone citrullination is essential for early embryo development. Sci. Rep. 2016, 6, 38727. [Google Scholar] [CrossRef]
- Qin, H.; Liu, X.; Li, F.; Miao, L.; Li, T.; Xu, B.; An, X.; Muth, A.; Thompson, P.R.; Coonrod, S.A.; Zhang, X. PAD1 promotes epithelial-mesenchymal transition and metastasis in triple-negative breast cancer cells by regulating MEK1-ERK1/2-MMP2 signaling. Cancer Lett. 2017, 409, 30–41. [Google Scholar] [CrossRef]
- Coassolo, S.; Davidson, G.; Negroni, L.; Gambi, G.; Daujat, S.; Romier, C.; Davidson, I. Citrullination of pyruvate kinase M2 by PADI1 and PADI3 regulates glycolysis and cancer cell proliferation. Nat. Commun. 2021, 12, 1718. [Google Scholar] [CrossRef]
- Williams, J.P.C.; Walport, L.J. PADI6: What we know about the elusive fifth member of the peptidyl arginine deiminase family. Philos Trans R Soc Lond B Biol Sci. 2023, 378. [Google Scholar] [CrossRef]
- Yurttas, P.; Vitale, A.M.; Fitzhenry, R.J.; Cohen-Gould, L.; Wu, W.; Gossen, J.A.; Coonrod, S.A. Role for PADI6 and the cytoplasmic lattices in ribosomal storage in oocytes and translational control in the early mouse embryo. Development 2008, 135, 2627–2636. [Google Scholar] [CrossRef]
- Xu, Y.; Shi, Y.; Fu, J.; Yu, M.; Feng, R.; Sang, Q.; Liang, B.; Chen, B.; Qu, R.; Li, B.; Yan, Z.; Mao, X.; Kuang, Y.; Jin, L.; He, L. , Sun, X.; Wang, L. Mutations in PADI6 Cause Female Infertility Characterized by Early Embryonic Arrest. Am. J. Hum. Genet. 2016, 99, 744–752. [Google Scholar] [CrossRef]
- Inal, J.M.; Hristova, M.; Lange, S. A Pilot Study on Peptidylarginine Deiminases and Protein Deimination in Animal Cancers across Vertebrate Species. Int. J. Mol. Sci. 2022, 23, 8697. [Google Scholar] [CrossRef]
- D’Alessio, S.; Cheng, H.; Eaton, L.; Kraev, I.; Pamenter, M.E.; Lange, S. Acute Hypoxia Alters Extracellular Vesicle Signatures and the Brain Citrullinome of Naked Mole-Rats (Heterocephalus glaber). Int J Mol Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Y. , Huang, Y., Shi, G., Lei, X., Li, K., Zhou, R., Bai, L., Qin, C. Transcriptome profiling of five brain regions in a 6-hydroxydopamine rat model of Parkinson’s disease. CNS Neurosci Ther. 2021, 27, 1289. [Google Scholar] [CrossRef] [PubMed]
- Vaibhav, K.; Braun, M.; Alverson, K.; Khodadadi, H.; Kutiyanawalla, A.; Ward, A.; Banerjee, C.; Sparks, T.; Malik, A.; Rashid, M.H.; et al. Neutrophil extracellular traps exacerbate neurological deficits after traumatic brain injury. Sci. Adv. 2020, 6, eaax8847. [Google Scholar] [CrossRef]
- Nicholas, A.P.; Lu, L.; Heaven, M.; Kadish, I.; van Groen, T.; Accavitti-Loper, M.A.; Wewering, S.; Kofskey, D.; Gambetti, P.; Brenner, M. Ongoing studies of deimination in neurodegenerative diseases using the F95 antibody. In Protein Deimination in Human Health and Disease; Nicholas, A.P., Bhattacharya, S.K., Eds.; Springer-Verlag: New York, NY, USA, 2014; pp. 257–280. [Google Scholar]
- Petrozziello, T.; Mills, A.N.; Vaine, C.A.; Penney, E.B.; Fernandez-Cerado, C.; Legarda, G.; Velasco-Andrada, M.S.; Acuña, P.J.; Ang, M.A.; Muñoz, E.L.; Diesta, C. C. E.; Macalintal-Canlas, R.; Acuña-Sunshine, G.; Ozelius, L. J.; Sharma, N.; Bragg, D. C.; Sadri-Vakili, G. Neuroinflammation and histone H3 citrullination are increased in X-linked Dystonia Parkinsonism post-mortem prefrontal cortex. Neurobiol. Dis. 2020, 144, 105032. [Google Scholar] [CrossRef]
- Mondal, S.; Thompson, P.R. Protein Arginine Deiminases (PADs): Biochemistry and Chemical Biology of Protein Citrullination. Acc Chem Res. 2019, 52, 818. [Google Scholar] [CrossRef]
- Verma, D.K.; Seo, B.A.; Ghosh, A.; Ma, S.X.; Hernandez-Quijada, K.; Andersen, J.K.; Ko, H.S.; Kim, Y.H. Alpha-Synuclein Preformed Fibrils Induce Cellular Senescence in Parkinson’s Disease Models. Cells. 2021, 10. [Google Scholar] [CrossRef]
- Melis, M.; Pistis, M. Endocannabinoid signaling in midbrain dopamine neurons: More than physiology? Curr. Neuropharmacol. 2007, 5, 268–277. [Google Scholar] [CrossRef]
- Orgado, J.M.; Fernández-Ruiz, J.; Romero, J. The endocannabinoid system in neuropathological states. Int. Rev. Psychiatry 2009, 21, 172–180. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Q.J.; Wang, H.S.; Wang, T.; Liu, J. Genome-wide microarray analysis identifies a potential role for striatal retrograde endocannabinoid signaling in the pathogenesis of experimental L-DOPA-induced dyskinesia. Synapse 2014, 68, 332–343. [Google Scholar] [CrossRef]
- Aymerich, M.S.; Aso, E.; Abellanas, M.A.; Tolon, R.M.; Ramos, J.A.; Ferrer, I.; Romero, J.; Fernández-Ruiz, J. Cannabinoid pharmacology/therapeutics in chronic degenerative disorders affecting the central nervous system. Biochem. Pharmacol. 2018, 157, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.P.; Bano, S.; Sen, S.; Suchal, K.; Kumar, S.; Nikolajeff, F.; Dey, S.K; Sharma, V. Altered neural cell junctions and ion-channels leading to disrupted neuron communication in Parkinson’s disease. NPJ Parkinsons Dis. 2022, 8. [Google Scholar] [CrossRef]
- Sen, S.; Lagas, S.; Roy, A.; Kumar, H. Cytoskeleton saga: Its regulation in normal physiology and modulation in neurodegenerative disorders. Eur J Pharmacol. 2022, 925. [Google Scholar] [CrossRef]
- Calabresi, P.; Di Filippo, M.; Gallina, A.; Wang, Y.; Stankowski, J.N.; Picconi, B.; Dawson, V.L.; Dawson, T.M. New synaptic and molecular targets for neuroprotection in Parkinson’s disease. Mov Disord. 2013, 28, 51. [Google Scholar] [CrossRef] [PubMed]
- Barber, C.N.; Huganir, R.L.; Raben, D.M. Phosphatidic acid-producing enzymes regulating the synaptic vesicle cycle: Role for PLD? Adv Biol Regul. 2018, 67, 141. [Google Scholar] [CrossRef]
- Beilina, A.; Bonet-Ponce, L.; Kumaran, R.; et al. The Parkinson’s Disease Protein LRRK2 Interacts with the GARP Complex to Promote Retrograde Transport to the trans-Golgi Network. Cell Rep. 2020, 31. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Hwang, B.; Reva, M.; Lee, J.; Lee, B. E.; Lee, Y.; Cho, E. J.; Jeong, M.; Lee, S.E.; Myung, K.; Baik, J.H.; Park, J.H.; Kim, J.I. GABAergic-like dopamine synapses in the brain. Cell Rep. 2023, 42. [Google Scholar] [CrossRef] [PubMed]
- Bohnen, N.I.; Roytman, S.; Kanel, P.; et al. Progression of regional cortical cholinergic denervation in Parkinson’s disease. Brain Commun. 2022, 4, fcac. [Google Scholar] [CrossRef] [PubMed]
- Gcwensa, N.Z.; Russell, D.L.; Cowell, R.M.; Volpicelli-Daley, L.A. Molecular Mechanisms Underlying Synaptic and Axon Degeneration in Parkinson’s Disease. Front Cell Neurosci. 2021, 15. [Google Scholar] [CrossRef]
- de Natale, E.R.; Wilson, H.; Politis, M. Serotonergic imaging in Parkinson’s disease. Prog Brain Res. 2021, 261, 303. [Google Scholar] [CrossRef]
- Richard, I.H. Depression in Parkinson’s Disease. Curr Treat Options Neurol. 2000, 2, 263. [Google Scholar] [CrossRef] [PubMed]
- Weintraub, D.; Aarsland, D.; Chaudhuri, K.R.; Dobkin, R.D.; Leentjens, A.F.; Rodriguez-Violante, M.; Schrag, A. The neuropsychiatry of Parkinson’s disease: Advances and challenges. Lancet Neurol. 2022, 21, 89. [Google Scholar] [CrossRef]
- Reumann, D.; Krauditsch, C.; Novatchkova, M.; Sozzi, E.; Wong, S.N.; Zabolocki, M.; Priouret, M.; Doleschall, B.; Ritzau-Reid, K. I.; Piber, M.; Morassut, I.; Fieseler, C.; Fiorenzano, A.; Stevens, M.M.; Zimmer, M.; Bardy, C.; Parmar, M.; Knoblich, J.A. In vitro modeling of the human dopaminergic system using spatially arranged ventral midbrain-striatum-cortex assembloids. Nat Methods. 2023, 20, 2034. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.; Yang, Q.; B Joshi, R.; Liu, Y.; Akbar, M.; Song, B. J.; Zhou, S.; Wang, X. Role of Alcohol Drinking in Alzheimer’s Disease, Parkinson’s Disease, and Amyotrophic Lateral Sclerosis. Int J Mol Sci. 2020, 21. [Google Scholar] [CrossRef]
- Jang, Y.; Pletnikova, O.; Troncoso, J.C.; Pantelyat, A.Y.; Dawson, T.M.; Rosenthal, L.S.; Na, C.H. Mass Spectrometry-Based Proteomics Analysis of Human Substantia Nigra From Parkinson’s Disease Patients Identifies Multiple Pathways Potentially Involved in the Disease. Mol Cell Proteomics. 2023, 22. [Google Scholar] [CrossRef]
- Iovino, M.; Messana, T.; Tortora, A.; Giusti, C.; Lisco, G.; Giagulli, V.A.; Guastamacchia, E.; De Pergola, G.; Triggiani, V. Oxytocin Signaling Pathway: From Cell Biology to Clinical Implications. Endocr Metab Immune Disord Drug Targets. 2021, 21, 91. [Google Scholar] [CrossRef]
- Higashida, H.; Hashii, M.; Tanaka, Y.; et al. CD38, CD157, and RAGE as Molecular Determinants for Social Behavior. Cells. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Lestón Pinilla, L.; Ugun-Klusek, A.; Rutella, S.; De Girolamo, L.A. Hypoxia Signaling in Parkinson’s Disease: There Is Use in Asking “What HIF?”. Biology (Basel). 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Trist, B.G.; Hare, D.J.; Double, K.L. Oxidative stress in the aging substantia nigra and the etiology of Parkinson’s disease. Aging Cell. 2019, 18. [Google Scholar] [CrossRef] [PubMed]
- Peggion, C.; Barazzuol, L.; Poggio, E.; Calì, T.; Brini, M. Ca2+ signalling: A common language for organelles crosstalk in Parkinson’s disease. Cell Calcium. 2023, 115. [Google Scholar] [CrossRef]
- Barazzuol, L.; Giamogante, F.; Brini, M.; Calì, T. PINK1/Parkin Mediated Mitophagy, Ca2+ Signalling, and ER-Mitochondria Contacts in Parkinson’s Disease. Int J Mol Sci. 2020, 21. [Google Scholar] [CrossRef]
- Virdi, G.S.; Choi, M.L.; Evans, J.R.; et al. Protein aggregation and calcium dysregulation are hallmarks of familial Parkinson’s disease in midbrain dopaminergic neurons. NPJ Parkinsons Dis. 2022, 8. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, J.; Zhao, K.; Wu, S.; Chen, X.; Hu, W. The Role of Calcium and Iron Homeostasis in Parkinson’s Disease. Brain Sci. 2024, 14. [Google Scholar] [CrossRef]
- Wang, Z.L.; Yuan, L.; Li, W.; Li, J.Y. Ferroptosis in Parkinson’s disease: Glia-neuron crosstalk. Trends Mol Med. 2022, 28, 258. [Google Scholar] [CrossRef]
- Mahoney-Sánchez, L.; Bouchaoui, H.; Ayton, S.; Devos, D.; Duce, J.A.; Devedjian, J.C. Ferroptosis and its potential role in the physiopathology of Parkinson’s Disease. Prog Neurobiol. 2021, 196. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.S.; Gao, L.; Han, Z.; et al. Ferroptosis in Parkinson’s disease: Molecular mechanisms and therapeutic potential. Ageing Res Rev. 2023, 91. [Google Scholar] [CrossRef] [PubMed]
- Thapa, K.; Khan, H.; Kanojia, N.; Singh, T.G.; Kaur, A.; Kaur, G. Therapeutic Insights on Ferroptosis in Parkinson’s disease. Eur J Pharmacol. 2022, 930. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Gao, X.; Zhao, L.; Wang, Y.; Zhang, J.; Liu, S. Effects of Cannabidiol on Parkinson’s Disease in a Transgenic Mouse Model by Gut-Brain Metabolic Analysis. Evid Based Complement Alternat Med. 2022, 2022. [Google Scholar] [CrossRef] [PubMed]
- Fusco, C.M.; Desch, K.; Dörrbaum, A.R.; Wang, M.; Staab, A.; Chan, I.C.W.; Vail, E.; Villeri, V.; Langer, J.D.; Schuman, E.M. Neuronal ribosomes exhibit dynamic and context-dependent exchange of ribosomal proteins. Nat Commun. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Tresse, E.; Marturia-Navarro, J.; Sew, W.Q.G.; et al. Mitochondrial DNA damage triggers spread of Parkinson’s disease-like pathology. Mol Psychiatry. 2023, 28, 4902. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Meng, X.; Yang, C.; Fang, D.; Wang, X.; An, J.; Zhang, J.; Wang, L.; Lu, T.; Ruan, H.B.; et al. Brown adipose tissue activation in a rat model of Parkinson’s disease. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E731–E736. [Google Scholar] [CrossRef]
- Naeem, U.; Arshad, A.R.; Jawed, A.; et al. Glycolysis: The Next Big Breakthrough in Parkinson’s Disease. Neurotox Res. 2022, 40, 1707. [Google Scholar] [CrossRef]
- Tang, B.L. Glucose, glycolysis, and neurodegenerative diseases. J Cell Physiol. 2020, 235, 7653. [Google Scholar] [CrossRef] [PubMed]
- Pathania, A.; Garg, P.; Sandhir, R. Impaired mitochondrial functions and energy metabolism in MPTP-induced Parkinson’s disease: Comparison of mice strains and dose regimens. Metab Brain Dis. 2021, 36, 2343. [Google Scholar] [CrossRef] [PubMed]
- Kopp, K.O.; Glotfelty, E.J.; Li, Y.; Greig, N.H. Glucagon-like peptide-1 (GLP-1) receptor agonists and neuroinflammation: Implications for neurodegenerative disease treatment. Pharmacol Res. 2022, 186. [Google Scholar] [CrossRef] [PubMed]
- Jha, S.K.; Jha, N.K.; Kumar, D.; Ambasta, R.K.; Kumar, P. Linking mitochondrial dysfunction, metabolic syndrome and stress signaling in Neurodegeneration. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1132–1146. [Google Scholar] [CrossRef]
- Limphaibool, N.; Iwanowski, P.; Holstad, M.J.V.; Perkowska, K. Parkinsonism in Inherited Metabolic Disorders: Key Considerations and Major Features. Front. Neurol. 2018, 9, 857. [Google Scholar] [CrossRef] [PubMed]
- Özcan, E.; Çakır, T. Genome-Scale Brain Metabolic Networks as Scaffolds for the Systems Biology of Neurodegenerative Diseases: Mapping Metabolic Alterations. Adv. Neurobiol. 2018, 21, 195–217. [Google Scholar]
- Gonçalves, V.C.; Cuenca-Bermejo, L.; Fernandez-Villalba, E.; Martin-Balbuena, S.; da Silva Fernandes, M.J.; Scorza, C.A.; Herrero, M.T. Heart Matters: Cardiac Dysfunction and Other Autonomic Changes in Parkinson’s Disease. Neuroscientist. 2022, 28, 530. [Google Scholar] [CrossRef] [PubMed]
- Smeyne, R.J.; Noyce, A.J.; Byrne, M.; Savica, R.; Marras, C. Infection and Risk of Parkinson’s Disease. J Parkinsons Dis. 2021, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Arisan, E.D., Uysal-Onganer. Uysal-Onganer, P., Lange, S. Putative Roles for Peptidylarginine Deiminases in COVID-. Int J Mol Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Leta, V.; Boura, I.; van Wamelen, D.J.; Rodriguez-Violante, M.; Antonini, A.; Chaudhuri, K.R. Covid-19 and Parkinson’s disease: Acute clinical implications, long-COVID and post-COVID-19 parkinsonism. Int Rev Neurobiol. 2022, 165, 63. [Google Scholar] [CrossRef]
- Priyanka, *!!! REPLACE !!!*; Seth, P. Insights Into the Role of Mortalin in Alzheimer’s Disease, Parkinson’s Disease, and HIV-1-Associated Neurocognitive Disorders. Front Cell Dev Biol. 2022, 10. [Google Scholar] [CrossRef]
- Olson, K.E.; Bade, A.N.; Namminga, K.L.; Potash, M.J.; Mosley, R.L.; Poluektova, L.Y.; Volsky, D.J.; Gendelman, H.E. Persistent EcoHIV infection induces nigral degeneration in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-intoxicated mice. J Neurovirol. 2018, 24, 398. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.M.; Dahodwala, N. Sex differences in Parkinson’s disease and other movement disorders. Exp Neurol. 2014, 259, 44. [Google Scholar] [CrossRef] [PubMed]
- Al-Sweidi, S.; Morissette, M.; Bourque, M.; Di Paolo, T. Estrogen receptors and gonadal steroids in vulnerability and protection of dopamine neurons in a mouse model of Parkinson’s disease. Neuropharmacology. 2011, 61, 583. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.K.; Chan, L.L.; Auchus, A.P. Reversible Parkinsonism in systemic lupus erythematosus. J. Neurol. Sci. 2001, 193, 53–57. [Google Scholar] [CrossRef]
- Kivity, S.; Agmon-Levin, N.; Zandman-Goddard, G.; Chapman, J.; Shoenfeld, Y. Neuropsychiatric lupus: A mosaic of clinical presentations. BMC Med. 2015, 13, 43. [Google Scholar] [CrossRef]
- Li, M.; Wan, J.; Xu, Z.; Tang, B. The association between Parkinson’s disease and autoimmune diseases: A systematic review and meta-analysis. Front Immunol. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Kalaani, J.; Roche, J.; Hamade, E.; Badran, B.; Jaber, M.; Gaillard, A.; Prestoz, L. Axon guidance molecule expression after cell therapy in a mouse model of Parkinson’s disease. Restor Neurol Neurosci. 2016, 34, 877. [Google Scholar] [CrossRef]
- Lesnick, T.G.; Sorenson, E.J.; Ahlskog, J.E.; Henley, J.R.; Shehadeh, L.; Papapetropoulos, S.; Maraganore, D.M. Beyond Parkinson disease: Amyotrophic lateral sclerosis and the axon guidance pathway. PLoS ONE. 2008, 3. [Google Scholar] [CrossRef]
- Lin, L.; Lesnick, T.G.; Maraganore, D.M.; Isacson, O. Axon guidance and synaptic maintenance: Preclinical markers for neurodegenerative disease and therapeutics. Trends Neurosci. 2009, 32, 142. [Google Scholar] [CrossRef]
- Sullivan, R.; Yau, W.Y.; O’Connor, E.; Houlden, H. Spinocerebellar ataxia: An update. J Neurol. 2019, 266, 533. [Google Scholar] [CrossRef]
- Horvath, J.D.; Casas, M.; Kutchukian, C.; Sánchez, S.C.; Pergande, M.R.; Cologna, S.M.; Simó, S.; Dixon, R.E.; Dickson, E.J. α-Synuclein-dependent increases in PIP5K1γ drive inositol signaling to promote neurotoxicity. Cell Rep. 2023, 42. [Google Scholar] [CrossRef]
- Napolitano, F.; D’Angelo, L.; de Girolamo, P.; Avallone, L.; de Lange, P.; Usiello, A. The Thyroid Hormone-target Gene Rhes a Novel Crossroad for Neurological and Psychiatric Disorders: New Insights from Animal Models. Neuroscience. 2018, 384, 419. [Google Scholar] [CrossRef]
- Li, Y.; Xia, Y.; Yin, S.; Wan, F.; Hu, J.; Kou, L.; Sun, Y.; Wu, J.; Zhou, Q.; Huang, J.; Xiong, N.; Wang, T. Targeting Microglial α-Synuclein/TLRs/NF-kappaB/NLRP3 Inflammasome Axis in Parkinson’s Disease. Front Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Piancone, F.; La Rosa, F.; Marventano, I.; Saresella, M.; Clerici, M. The Role of the Inflammasome in Neurodegenerative Diseases. Molecules. 2021, 26. [Google Scholar] [CrossRef]
- Panicker, N.; Kanthasamy, A.; Kanthasamy, A.G. Fyn amplifies NLRP3 inflammasome signaling in Parkinson’s disease. Aging (Albany NY). 2019, 11, 5871. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, B.; Tian, T.; Zhang, B.; Shi, G.; Zhang, C.; Li, G. , Huang, M. Taurine protects dopaminergic neurons in paraquat-induced Parkinson’s disease mouse model through PI3K/Akt signaling pathways. Amino Acids. 2022, 54, 1. [Google Scholar] [CrossRef]
- Deneubourg, C.; Ramm, M.; Smith, L.J.; Baron, O.; Singh, K.; Byrne, S.C.; Duchen, M.R.; Gautel, M.; Eskelinen, E.L.; Fanto, M.; Jungbluth, H. The spectrum of neurodevelopmental, neuromuscular and neurodegenerative disorders due to defective autophagy. Autophagy. 2022, 18, 496. [Google Scholar] [CrossRef]
- Ferrer-Raventós, P.; Beyer, K. Alternative platelet activation pathways and their role in neurodegenerative diseases. Neurobiol Dis. 2021, 159. [Google Scholar] [CrossRef]
- Oliynyk, Z.; Rudyk, M.; Dovbynchuk, T.; Dzubenko, N.; Tolstanova, G.; Skivka, L. Inflammatory hallmarks in 6-OHDA- and LPS-induced Parkinson’s disease in rats. Brain Behav Immun Health. 2023, 30. [Google Scholar] [CrossRef]
- Liu, X.; Chen, J.; Guan, T.; Yao, H.; Zhang, W.; Guan, Z.; Wang, Y. miRNAs and target genes in the blood as biomarkers for the early diagnosis of Parkinson’s disease. BMC Syst Biol. 2019, 13. [Google Scholar] [CrossRef]
- Moya, I.M.; Halder, G. Hippo-YAP/TAZ signalling in organ regeneration and regenerative medicine. Nat Rev Mol Cell Biol. 2019, 20, 211. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Hu, Y.; Lan, T.; Guan, K.L.; Luo, T.; Luo, M. The Hippo signalling pathway and its implications in human health and diseases. Signal Transduct Target Ther. 2022, 7. [Google Scholar] [CrossRef] [PubMed]
- Eddin, L.B.; Azimullah, S.; Jha, N.K.; Nagoor Meeran, M.F.; Beiram, R.; Ojha, S. Limonene, a Monoterpene, Mitigates Rotenone-Induced Dopaminergic Neurodegeneration by Modulating Neuroinflammation, Hippo Signaling and Apoptosis in Rats. Int J Mol Sci. 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Varela, L.; Garcia-Rendueles, M.E.R. Oncogenic Pathways in Neurodegenerative Diseases. Int J Mol Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Huang, G.; Liu, J.; Ge, J.; Zhang, W.; Mei, Z. An update on the role of Hippo signaling pathway in ischemia-associated central nervous system diseases. Biomed Pharmacother. 2023, 162. [Google Scholar] [CrossRef]
- Xu, D.D.; Li, G.Q.; Wu, Z.S.; Liu, X.Q.; Yang, X.X.; Wang, J.H. Bioinformatics analysis and identification of genes and molecular pathways involved in Parkinson’s disease in patients with mutations in the glucocerebrosidase gene. Neuroreport. 2021, 32, 918. [Google Scholar] [CrossRef] [PubMed]
- Stamou, M.; Grodzki, A.C.; van Oostrum, M.; Wollscheid, B.; Lein, P.J. Fc gamma receptors are expressed in the developing rat brain and activate downstream signaling molecules upon cross-linking with immune complex. J Neuroinflammation 2018, 15. [Google Scholar] [CrossRef]
- Park, Y.H.; Hodges, A.; Risacher, S.L.; Lin, K.; Jang, J.W.; Ahn, S.; Kim, S.; Lovestone, S.; Simmons, A.; Weiner, M.W.; Saykin, A.J.; Nho, K. AddNeuroMed consortium and the Alzheimer’s Disease Neuroimaging Initiative. Dysregulated Fc gamma receptor-mediated phagocytosis pathway in Alzheimer’s disease: Network-based gene expression analysis. Neurobiol Aging 2020, 88, 24. [Google Scholar] [CrossRef] [PubMed]
- Massey, W.J.; Kay, K.E.; Jaramillo, T.C.; et al. Metaorganismal choline metabolism shapes olfactory perception. J Biol Chem. 2023, 299. [Google Scholar] [CrossRef]
- Yang, Y.; Qian, J.; Mei, J.; Zhong, K.; Niu, C. In vivo detection of metabolic changes in the striatum of proteasomal inhibition-induced Parkinson’s disease in rats using proton MR spectroscopy at 9.4 T. Int J Neurosci. 2020, 130, 153. [Google Scholar] [CrossRef]
- Sanami, S.; Shamsabadi, S.; Dayhimi, A.; Pirhayati, M.; Ahmad, S.; Pirhayati, A.; Ajami, M.; Hemati, S.; Shirvani, M.; Alagha, A.; Abbarin, D.; Alizadeh, A.; Pazoki-Toroudi, H. Association between cytomegalovirus infection and neurological disorders: A systematic review. Rev Med Virol. 2024, 34, 34. [Google Scholar] [CrossRef]
- Bu, X.L.; Wang, X.; Xiang, Y.; Shen, L.L.; Wang, Q.H.; Liu, Y.H.; Jiao, S.S.; Wang, Y.R.; Cao, H.Y.; Yi, X.; Liu, C.H.; Deng, B.; Yao, X.Q.; Xu, Z.Q.; Zhou, H.D.; Wang, Y.J. The association between infectious burden and Parkinson’s disease: A case-control study. Parkinsonism Relat Disord. 2015, 21, 877. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, Y.; Kashyap, P.C. Parkinson’s disease: Are gut microbes involved? Am J Physiol Gastrointest Liver Physiol. 2020, 2020 319, G529. [Google Scholar] [CrossRef]
- Tan, C.; Liu, X.; Chen, J. Microarray Analysis of the Molecular Mechanism Involved in Parkinson’s Disease. Parkinsons Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.N.; Liu, J.C.; Ju, J.Q.; Wang, Y.; Sun, S.C. LRRK2 regulates actin assembly for spindle migration and mitochondrial function in mouse oocyte meiosis. J Mol Cell Biol. 2022, 2022 14, mjab. [Google Scholar] [CrossRef]
- Esposito, G.; Vitale, A.M.; Leijten, F.P.; Strik, A.M.; Koonen-Reemst, A.M.; Yurttas, P.; Robben, T.J.; Coonrod, S.; Gossen, J.A. Peptidylarginine deiminase (PAD) 6 is essential for oocyte cytoskeletal sheet formation and female fertility. Mol Cell Endocrinol. 2007, 273, 25. [Google Scholar] [CrossRef] [PubMed]
- Horibata, S.; Coonrod, S.A.; Cherrington, B.D. Role for peptidylarginine deiminase enzymes in disease and female reproduction. J Reprod Dev. 2012, 58, 274. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, T.G.; Di Paolo, G. Phospholipase D in brain function and Alzheimer’s disease. Biochim Biophys Acta. 2010, 1801, 799. [Google Scholar] [CrossRef]
- Lindsley, C.W.; Brown, H.A. Phospholipase D as a therapeutic target in brain disorders. Neuropsychopharmacology. 2012, 37, 301. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, F.; Zhu, J.; Liu, K. Phospholipase D, a Novel Therapeutic Target Contributes to the Pathogenesis of Neurodegenerative and Neuroimmune Diseases. Anal Cell Pathol (Amst). 2024, 2024. [Google Scholar] [CrossRef]
- Bottero, V.; Powers, D.; Yalamanchi, A.; Quinn, J.P.; Potashkin, J.A. Key Disease Mechanisms Linked to Alzheimer’s Disease in the Entorhinal Cortex. Int J Mol Sci. 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Manfredi-Lozano, M.; Leysen, V.; Adamo, M.; et al. GnRH replacement rescues cognition in Down syndrome. Science. 2022, 377, eabq. [Google Scholar] [CrossRef] [PubMed]
- Quigley, E.B.; DeVore, S.B.; Khan, S.A.; Geisterfer, Z.M.; Rothfuss, H.M.; Sequoia, A.O.; Thompson, P.R.; Gatlin, J.C. , Cherrington, B.D., Navratil, A.M. GnRH Induces Citrullination of the Cytoskeleton in Murine Gonadotrope Cells. Int J Mol Sci. 2024, 25. [Google Scholar] [CrossRef]
- Roy, A.; Al-bataineh, M.M.; Pastor-Soler, N.M. Collecting duct intercalated cell function and regulation. Clin J Am Soc Nephrol. 2015, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Reaux-Le Goazigo, A.; Bodineau, L.; De Mota, N.; Jeandel, L.; Chartrel, N.; Knauf, C.; Raad, C.; Valet, P.; Llorens-Cortes, C. Apelin and the proopiomelanocortin system: A new regulatory pathway of hypothalamic α-MSH release. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E955–E966. [Google Scholar] [CrossRef]
- Masoumi, J.; Abbasloui, M.; Parvan, R.; Mohammadnejad, D.; Pavon-Djavid, G.; Barzegari, A.; Abdolalizadeh, J. Apelin, a promising target for Alzheimer disease prevention and treatment. Neuropeptides 2018, 70, 76–86. [Google Scholar] [CrossRef]
- Zhu, J.; Gao, W.; Shan, X.; Wang, C.; Wang, H.; Shao, Z.; Dou, S.; Jiang, Y.; Wang, C.; Cheng, B. Apelin-36 mediates neuroprotective effects by regulating oxidative stress, autophagy and apoptosis in MPTP-induced Parkinson’s disease model mice. Brain Res. 2020, 1726, 146493. [Google Scholar] [CrossRef]
- Angelopoulou, E.; Paudel, Y.N.; Bougea, A.; Piperi, C. Impact of the apelin/APJ axis in the pathogenesis of Parkinson’s disease with therapeutic potential. J Neurosci Res. 2021, 99, 2117. [Google Scholar] [CrossRef]
- Doxakis, E. Insights into the multifaceted role of circular RNAs: Implications for Parkinson’s disease pathogenesis and diagnosis. NPJ Parkinsons Dis. 2022, 8. [Google Scholar] [CrossRef]
- Shvetcov, A.; Thomson, S.; Spathos, J.; et al. Blood-Based Transcriptomic Biomarkers Are Predictive of Neurodegeneration Rather Than Alzheimer’s Disease. Int J Mol Sci. 2023, 24. [Google Scholar] [CrossRef]
- Lan, A.P.; Chen, J.; Zhao, Y.; Chai, Z.; Hu, Y. mTOR Signaling in Parkinson’s Disease. Neuromolecular Med. 2017, 19, 1. [Google Scholar] [CrossRef] [PubMed]
- Pena-Leon, V.; Perez-Lois, R.; Seoane, L.M. mTOR Pathway is Involved in Energy Homeostasis Regulation as a Part of the Gut-Brain Axis. Int J Mol Sci. 2020, 21. [Google Scholar] [CrossRef]
- Tan, C.; Ai, J.; Zhu, Y. mTORC1-Dependent Protein and Parkinson’s Disease: A Mendelian Randomization Study. Brain Sci. 2023, 13. [Google Scholar] [CrossRef]
- Khan, M.R.; Yin, X.; Kang, S.U.; et al. Enhanced mTORC1 signaling and protein synthesis in pathologic α-synuclein cellular and animal models of Parkinson’s disease. Sci Transl Med. 2023, 15, eadd. [Google Scholar] [CrossRef] [PubMed]
- Fahmy, A.M.; Boulais, J.; Desjardins, M.; Matheoud, D. Mitochondrial antigen presentation: A mechanism linking Parkinson’s disease to autoimmunity. Curr Opin Immunol. 2019, 58, 31. [Google Scholar] [CrossRef] [PubMed]
- Hobson, B.D.; Sulzer, D. Neuronal Presentation of Antigen and Its Possible Role in Parkinson’s Disease. J Parkinsons Dis. 2022, 12, S137–S. [Google Scholar] [CrossRef]
- Schonhoff, A.M.; Figge, D.A.; Williams, G.P.; et al. Border-associated macrophages mediate the neuroinflammatory response in an alpha-synuclein model of Parkinson disease. Nat Commun. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Yamada, T. Viral etiology for Parkinson’s disease--a possible role of influenza A virus infection. Jpn J Infect Dis. 1999, 52, 89–98. [Google Scholar] [CrossRef]
- Ghemrawi, R.; Khair, M. Endoplasmic Reticulum Stress and Unfolded Protein Response in Neurodegenerative Diseases. Int J Mol Sci. 2020, 21, Published–2020. [Google Scholar] [CrossRef]
- Mou, Z.; Yuan, Y.H.; Zhang, Z.; Song, L.K.; Chen, N.H. Endoplasmic reticulum stress, an important factor in the development of Parkinson’s disease. Toxicol Lett. 2020, 324, 20. [Google Scholar] [CrossRef]
- Li, P.; Nie, Y.; Yu, J. An Effective Method to Identify Shared Pathways and Common Factors among Neurodegenerative Diseases. PLoS ONE 2015, 10, e0143045. [Google Scholar] [CrossRef]
- Attilio, P.J.; Flora, M.; Kamnaksh, A.; Bradshaw, D.J.; Agoston, D.; Mueller, G.P. The Effects of Blast Exposure on Protein Deimination in the Brain. Oxid. Med. Cell Longev. 2017, 2017, 8398072. [Google Scholar] [CrossRef] [PubMed]
- Gallart-Palau, X.; Lee, B.S.; Adav, S.S.; et al. Gender differences in white matter pathology and mitochondrial dysfunction in Alzheimer’s disease with cerebrovascular disease. Mol Brain. 2016, 9. [Google Scholar] [CrossRef]
- Munoz-Pinto, M.F.; Empadinhas, N.; Cardoso, S.M. The neuromicrobiology of Parkinson’s disease: A unifying theory. Ageing Res Rev. 2021, 70. [Google Scholar] [CrossRef]
- Griffante, G.; Gugliesi, F.; Pasquero, S.; et al. Human cytomegalovirus-induced host protein citrullination is crucial for viral replication. Nat Commun. 2021, 12. [Google Scholar] [CrossRef]
- Casanova, V.; Sousa, F.H.; Shakamuri, P.; et al. Citrullination Alters the Antiviral and Immunomodulatory Activities of the Human Cathelicidin LL-37 During Rhinovirus Infection. Front Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Martens, C.P.; Peetermans, M.; Vanassche, T.; Verhamme, P.; Jacquemin, M.; Martinod, K. Peptidylarginine deiminase 4 and ADAMTS13 activity in Staphylococcusaureus bacteraemia. Philos Trans R Soc Lond B Biol Sci. 2023, 378. [Google Scholar] [CrossRef]
- Sinha, S.; Mittal, S.; Roy, R. Parkinson’s Disease and the COVID-19 Pandemic: A Review Article on the Association between SARS-CoV-2 and α-Synucleinopathy. J Mov Disord. 2021, 14, 184. [Google Scholar] [CrossRef]
- Boura, I.; Qamar, M.A.; Daddoveri, F.; et al. SARS-CoV-2 and Parkinson’s Disease: A Review of Where We Are Now. Biomedicines. 2023, 11. [Google Scholar] [CrossRef]
- Calculli, A.; Bocci, T.; Porcino, M.; et al. Parkinson disease following COVID-19: Report of six cases. Eur J Neurol. 2023, 30, 1272. [Google Scholar] [CrossRef]
- Li, Z.; Lu, G.; Luo, E.; et al. Oral, Nasal, and Gut Microbiota in Parkinson’s Disease. Neuroscience. 2022, 480, 65. [Google Scholar] [CrossRef]
- Lazarini, F.; Roze, E.; Lannuzel, A.; Lledo, P.M. The microbiome-nose-brain axis in health and disease. Trends Neurosci. 2022, 45, 718. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, A.; Pirracchio, L.; Imber, J.C.; et al. Citrullination in periodontium is associated with Porphyromonas gingivalis. Arch Oral Biol. 2020, 114. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.P. Jr.; Keeney, P.M.; Brohawn, D.G. RNA Sequencing Reveals Small and Variable Contributions of Infectious Agents to Transcriptomes of Postmortem Nervous Tissues From Amyotrophic Lateral Sclerosis, Alzheimer’s Disease and Parkinson’s Disease Subjects, and Increased Expression of Genes From Disease-Activated Microglia. Front Neurosci. 2019, 13. [Google Scholar] [CrossRef]
- Gawron, K.; Bereta, G.; Nowakowska, Z.; et al. Peptidylarginine deiminase from Porphyromonas gingivalis contributes to infection of gingival fibroblasts and induction of prostaglandin E2 -signaling pathway. Mol Oral Microbiol. 2014, 29, 321. [Google Scholar] [CrossRef] [PubMed]
- Emery, D.C.; Davies, M.; Cerajewska, T.L.; et al. High resolution 16S rRNA gene Next Generation Sequencing study of brain areas associated with Alzheimer’s and Parkinson’s disease. Front Aging Neurosci. 2022, 14. [Google Scholar] [CrossRef]
- Cirstea, M.S.; Kliger, D.; MacLellan, A.D.; et al. The Oral and Fecal Microbiota in a Canadian Cohort of Alzheimer’s Disease. J Alzheimers Dis. 2022, 87, 247. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.K.; Wu, Q.L.; Peng, Y.W.; et al. Oral P. gingivalis impairs gut permeability and mediates immune responses associated with neurodegeneration in LRRK2 R1441G mice. J Neuroinflammation. 2020, 17. [Google Scholar] [CrossRef]
- Yay, E.; Yilmaz, M.; Toygar, H.; et al. Parkinson’s disease alters the composition of subgingival microbiome. J Oral Microbiol. 2023, 15. [Google Scholar] [CrossRef]
- Perricone, C.; Ceccarelli, F.; Saccucci, M.; Di Carlo, G.; Bogdanos, D.P.; Lucchetti, R.; Pilloni, A.; Valesini, G.; Polimeni, A.; Conti, F. Porphyromonas gingivalis and rheumatoid arthritis. Curr Opin Rheumatol. 2019, 31, 517. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Kurita-Ochiai, T.; Kobayashi, R.; Suzuki, T.; Ando, T. Activation of the NLRP3 inflammasome in Porphyromonas gingivalis-accelerated atherosclerosis. Pathog Dis. 2015, 73, ftv. [Google Scholar] [CrossRef] [PubMed]
- Blenkinsopp, H.C.; Seidler, K.; Barrow, M. Microbial Imbalance and Intestinal Permeability in the Pathogenesis of Rheumatoid Arthritis: A Mechanism Review with a Focus on Bacterial Translocation, Citrullination, and Probiotic Intervention. J Am Nutr Assoc. 2024, 43, 59. [Google Scholar] [CrossRef] [PubMed]
- Bielecka, E.; Scavenius, C.; Kantyka, T.; Jusko, M.; Mizgalska, D.; Szmigielski, B.; Potempa, B.; Enghild, J.J.; Prossnitz, E.R.; Blom, A.M.; Potempa, J. Peptidyl arginine deiminase from Porphyromonas gingivalis abolishes anaphylatoxin C5a activity. J. Biol. Chem. 2014, 289, 32481–32487. [Google Scholar] [CrossRef]
- Kosgodage, U.S.; Matewele, P.; Mastroianni, G.; Kraev, I.; Brotherton, D.; Awamaria, B.; Nicholas, A.P.; Lange, S.; Inal, J.M. Peptidylarginine Deiminase Inhibitors Reduce Bacterial Membrane Vesicle Release and Sensitize Bacteria to Antibiotic Treatment. Front. Cell Infect. Microbiol. 2019, 9, 227. [Google Scholar] [CrossRef]
- Sancandi, M.; De Caro, C.; Cypaite, N.; Marascio, N.; Avagliano, C.; De Marco, C.; Russo, E.; Constanti, A.; Mercer, A. Effects of a probiotic suspension Symprove™ on a rat early-stage Parkinson’s disease model. Front Aging Neurosci. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Adams, B.; Nunes, J.M.; Page, M.J.; Roberts, T.; Carr, J.; Nell, T.A.; Kell, D.B.; Pretorius, E. Parkinson’s Disease: A Systemic Inflammatory Disease Accompanied by Bacterial Inflammagens. Front Aging Neurosci. 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Olsen, I.; Kell, D.B.; Pretorius, E. Is Porphyromonas gingivalis involved in Parkinson’s disease? Eur J Clin Microbiol Infect Dis. 2020, 39, 2013. [Google Scholar] [CrossRef]
- Li, D.; Ren, T.; Li, H.; Liao, G.; Zhang, X. Porphyromonas gingivalis: A key role in Parkinson’s disease with cognitive impairment? Front Neurol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Ramazi, S.; Allahverdi, A.; Zahiri, J. Evaluation of post-translational modifications in histone proteins: A review on histone modification defects in developmental and neurological disorders. J Biosci 2020, 45, 135. [Google Scholar] [CrossRef]
- Antunes, A.S.L.M. Post-translational Modifications in Parkinson’s Disease. Adv Exp Med Biol. 2022, 1382, 1382, 85. [Google Scholar] [CrossRef]
- Zhang, S.; Zhu, R.; Pan, B.; et al. Post-translational modifications of soluble α-synuclein regulate the amplification of pathological α-synuclein. Nat Neurosci. 2023, 26, 213. [Google Scholar] [CrossRef] [PubMed]
- Bogetofte, H.; Ryan, B.J.; Jensen, P.; et al. Post-translational proteomics platform identifies neurite outgrowth impairments in Parkinson’s disease GBA-N370S dopamine neurons. Cell Rep. 2023, 42. [Google Scholar] [CrossRef]
- Kim, S.; Song, Y.K.; Cho, C.S.; Kim, H.J.; Fang, S.; Jo, D.H.; Kim, H. Inhibition of protein arginine deiminase II suppresses retinoblastoma in orthotopic transplantation in mice. Oncol Rep. 2023, 50. [Google Scholar] [CrossRef] [PubMed]
- Sancandi, M.; Schul, E.V.; Economides, G.; Constanti, A.; Mercer, A. Structural Changes Observed in the Piriform Cortex in a Rat Model of Pre-motor Parkinson’s Disease. Front. Cell Neurosci. 2018, 12, 479. [Google Scholar] [CrossRef]
- Nicholas, A.P.; Whitaker, J.N. Preparation of a monoclonal antibody to citrullinated epitopes: Its characterization and some applications to immunohistochemistry in human brain. Glia 2002, 37, 328–336. [Google Scholar] [CrossRef]
Figure 1.
PAD isozyme and prohibitin protein detection in cortex (Ctx) and hippocampus (Hip) of sham/control (Ctr) and pre-motor PD (PD) rats. A) PAD1; B) PAD2; C) PAD3; D) PAD4; E) PAD6; F) Prohibitin (PHB). Protein levels were assessed in n = 3 brains per group and normalised against beta-actin protein levels (fold-changes are shown); exact p-values are shown with significant differences indicated with a star *(p<0.05); mean and standard deviation are presented. Arrows (and brackets) indicate the area of the blot assessed for the predicted PAD protein bands (at 70-75 kDa for PAD1-4 and 50-60 kDa for PAD6) and prohibitin (PHB, 27 kDa); note low detection of PAD1 in both cortex and hippocampus.
Figure 1.
PAD isozyme and prohibitin protein detection in cortex (Ctx) and hippocampus (Hip) of sham/control (Ctr) and pre-motor PD (PD) rats. A) PAD1; B) PAD2; C) PAD3; D) PAD4; E) PAD6; F) Prohibitin (PHB). Protein levels were assessed in n = 3 brains per group and normalised against beta-actin protein levels (fold-changes are shown); exact p-values are shown with significant differences indicated with a star *(p<0.05); mean and standard deviation are presented. Arrows (and brackets) indicate the area of the blot assessed for the predicted PAD protein bands (at 70-75 kDa for PAD1-4 and 50-60 kDa for PAD6) and prohibitin (PHB, 27 kDa); note low detection of PAD1 in both cortex and hippocampus.
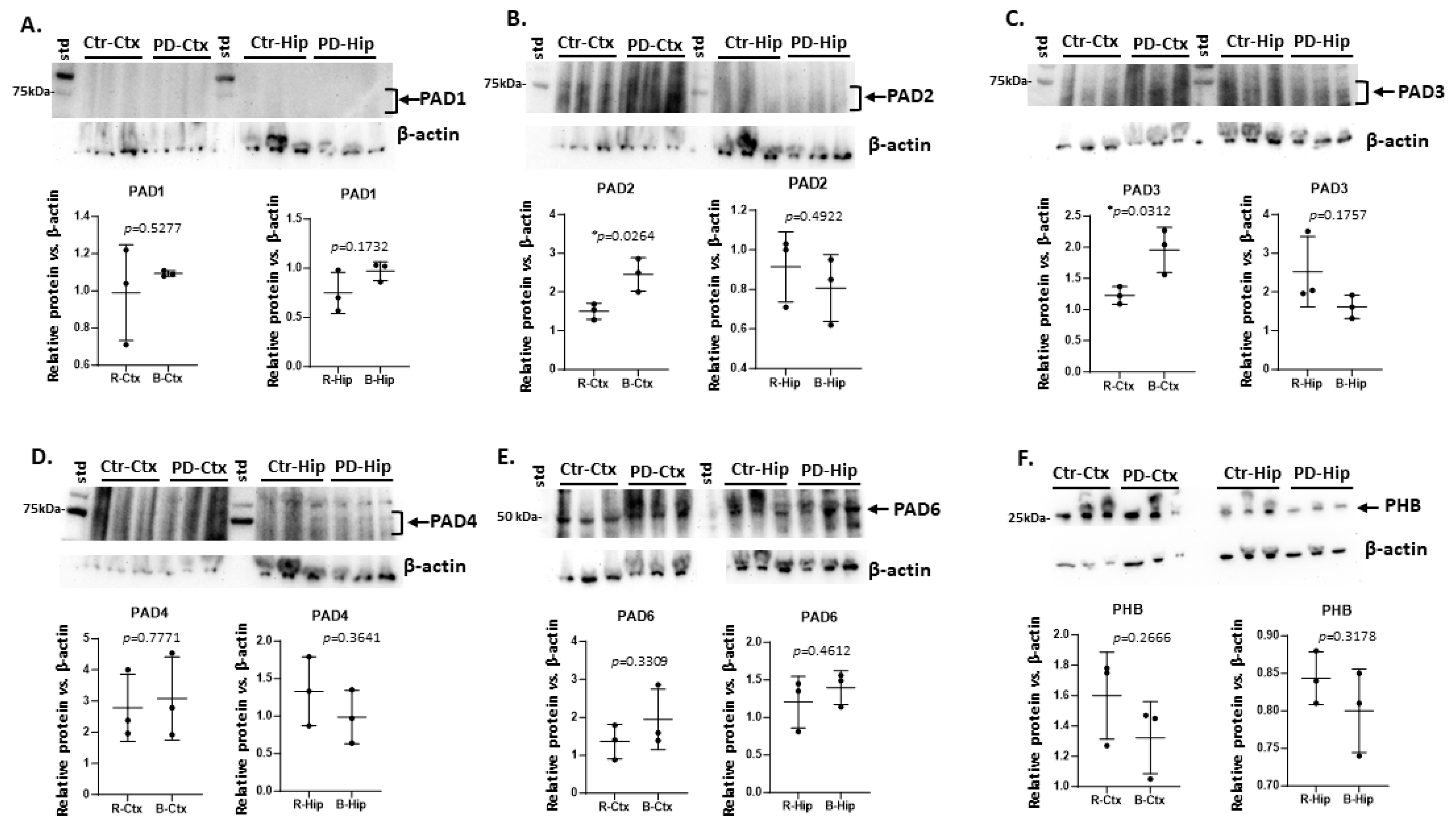
Figure 2.
PAD isozyme and prohibitin detection in cerebellum (Cer) and midbrain (Mid) of sham/control (Ctr) and pre-motor PD (PD) rats. A) PAD1; B) PAD2; C) PAD3; D) PAD4; E) PAD6; F) Prohibitin (PHB). Protein levels were assessed in n = 3 brains per group and normalised against beta-actin protein levels (fold-changes are shown); exact p-values are shown with significant differences indicated with a star *(p<0.05); mean and standard deviation are presented. Arrows (and brackets) indicate the area of the blot assessed for the predicted PAD protein bands (at 70-75 kDa for PAD1-4 and 50-60 kDa for PAD6) and prohibitin (PHB, 27 kDa).
Figure 2.
PAD isozyme and prohibitin detection in cerebellum (Cer) and midbrain (Mid) of sham/control (Ctr) and pre-motor PD (PD) rats. A) PAD1; B) PAD2; C) PAD3; D) PAD4; E) PAD6; F) Prohibitin (PHB). Protein levels were assessed in n = 3 brains per group and normalised against beta-actin protein levels (fold-changes are shown); exact p-values are shown with significant differences indicated with a star *(p<0.05); mean and standard deviation are presented. Arrows (and brackets) indicate the area of the blot assessed for the predicted PAD protein bands (at 70-75 kDa for PAD1-4 and 50-60 kDa for PAD6) and prohibitin (PHB, 27 kDa).
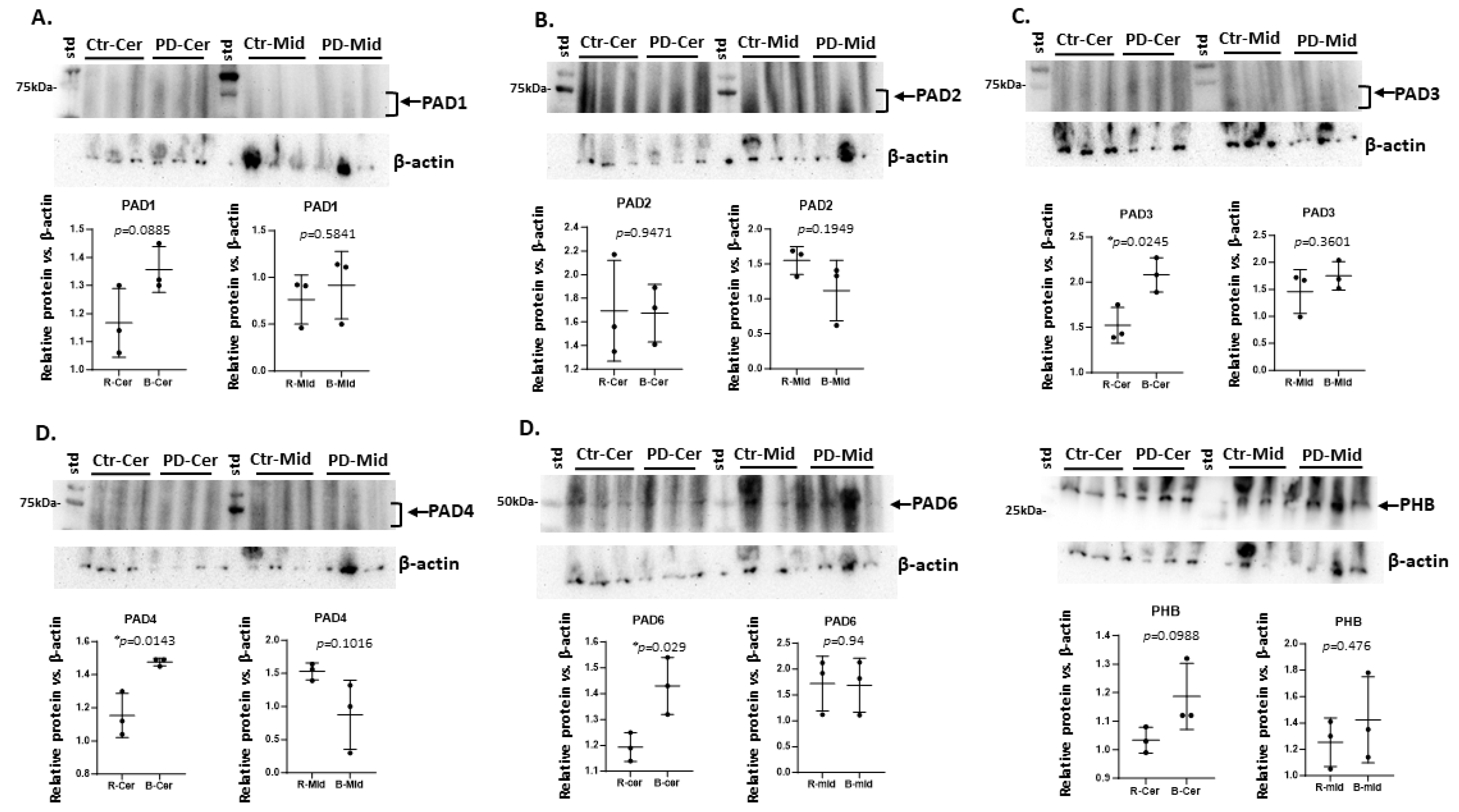
Figure 3.
PAD isozyme and prohibitin protein detection in olfactory bulb (Ob) and striatum (Str) of sham/control (Ctr) and pre-motor PD (PD) rats. A) PAD1; B) PAD2; C) PAD3; D) PAD4; E) PAD6; F) Prohibitin (PHB). Protein levels were assessed in n = 3 brains per group and normalised against beta-actin protein levels (fold-changes are shown); exact p-values are shown with significant differences indicated with a star *(p<0.05); mean and standard deviation are presented. Arrows (and brackets) indicate the area of the blot assessed for the predicted PAD protein bands (at 70-75 kDa for PAD1-4 and 50-60 kDa for PAD6) and prohibitin (PHB, 27 kDa).
Figure 3.
PAD isozyme and prohibitin protein detection in olfactory bulb (Ob) and striatum (Str) of sham/control (Ctr) and pre-motor PD (PD) rats. A) PAD1; B) PAD2; C) PAD3; D) PAD4; E) PAD6; F) Prohibitin (PHB). Protein levels were assessed in n = 3 brains per group and normalised against beta-actin protein levels (fold-changes are shown); exact p-values are shown with significant differences indicated with a star *(p<0.05); mean and standard deviation are presented. Arrows (and brackets) indicate the area of the blot assessed for the predicted PAD protein bands (at 70-75 kDa for PAD1-4 and 50-60 kDa for PAD6) and prohibitin (PHB, 27 kDa).
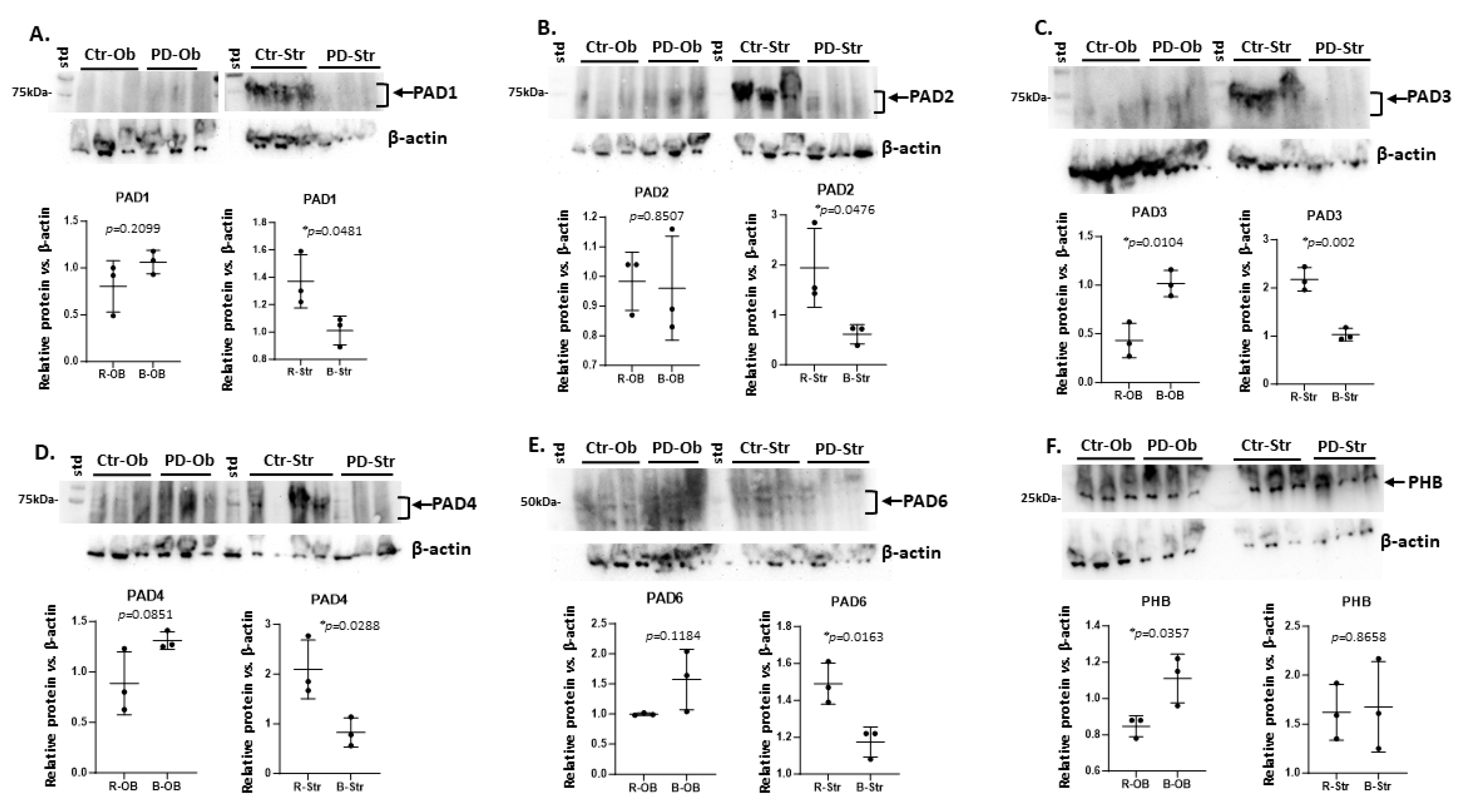
Figure 4.
Citrullinated proteins, isolated by F95 enrichment from: A) cortex (ctx) and hippocampus (hip); B) cerebellum (cer), midbrain (mid), olfactory bulb (OB) and striatum (str). F95 enriched fractions are shown from sham/control (ctr) and pre-motor PD (PD) brain regions on a silver stained SDS-PAGE TGX 4-20% gel. C) A summary of numbers of citrullinated proteins (#F95) identified per brain region and with respect to experimental group (control/sham – ctr; PD-group) by LC-MS/MS analysis.
Figure 4.
Citrullinated proteins, isolated by F95 enrichment from: A) cortex (ctx) and hippocampus (hip); B) cerebellum (cer), midbrain (mid), olfactory bulb (OB) and striatum (str). F95 enriched fractions are shown from sham/control (ctr) and pre-motor PD (PD) brain regions on a silver stained SDS-PAGE TGX 4-20% gel. C) A summary of numbers of citrullinated proteins (#F95) identified per brain region and with respect to experimental group (control/sham – ctr; PD-group) by LC-MS/MS analysis.
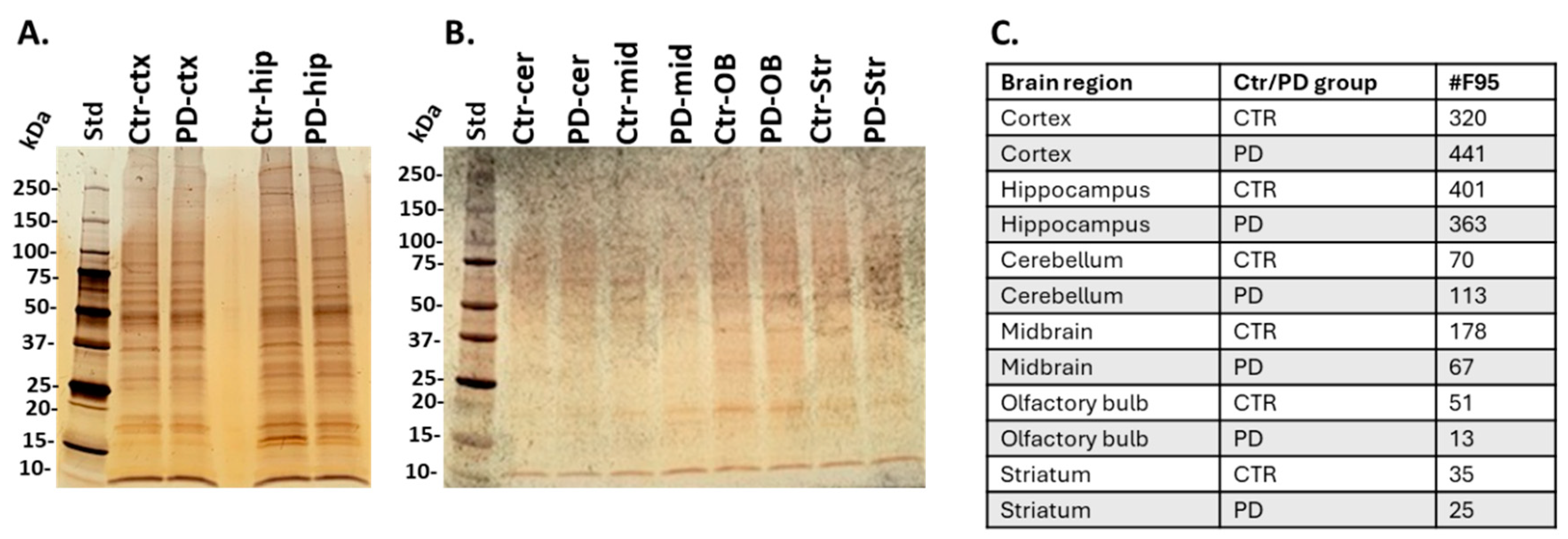
Figure 5.
The citrullinomes of control and pre-motor PD rat cortex. A) Protein interaction networks of the protein citrullinome of sham/control rat cortex. B) Protein interaction network of the protein citrullinome of PD rat cortex. C) A Venn diagram representing number (#) of citrullinated protein hits (F95) and of associated KEGG pathways in sham/control and pre-motor PD cortex. .
Figure 5.
The citrullinomes of control and pre-motor PD rat cortex. A) Protein interaction networks of the protein citrullinome of sham/control rat cortex. B) Protein interaction network of the protein citrullinome of PD rat cortex. C) A Venn diagram representing number (#) of citrullinated protein hits (F95) and of associated KEGG pathways in sham/control and pre-motor PD cortex. .

Figure 6.
The citrullinomes of control and pre-motor PD rat hippocampus. A) Protein interaction networks of the protein citrullinome of sham/control rat hippocampus. B) Protein interaction network of the protein citrullinome of PD rat hippocampus. C) A Venn diagram representing number (#) of citrullinated protein hits (F95) and of associated KEGG pathways in control and pre-motor PD hippocampus.
Figure 6.
The citrullinomes of control and pre-motor PD rat hippocampus. A) Protein interaction networks of the protein citrullinome of sham/control rat hippocampus. B) Protein interaction network of the protein citrullinome of PD rat hippocampus. C) A Venn diagram representing number (#) of citrullinated protein hits (F95) and of associated KEGG pathways in control and pre-motor PD hippocampus.
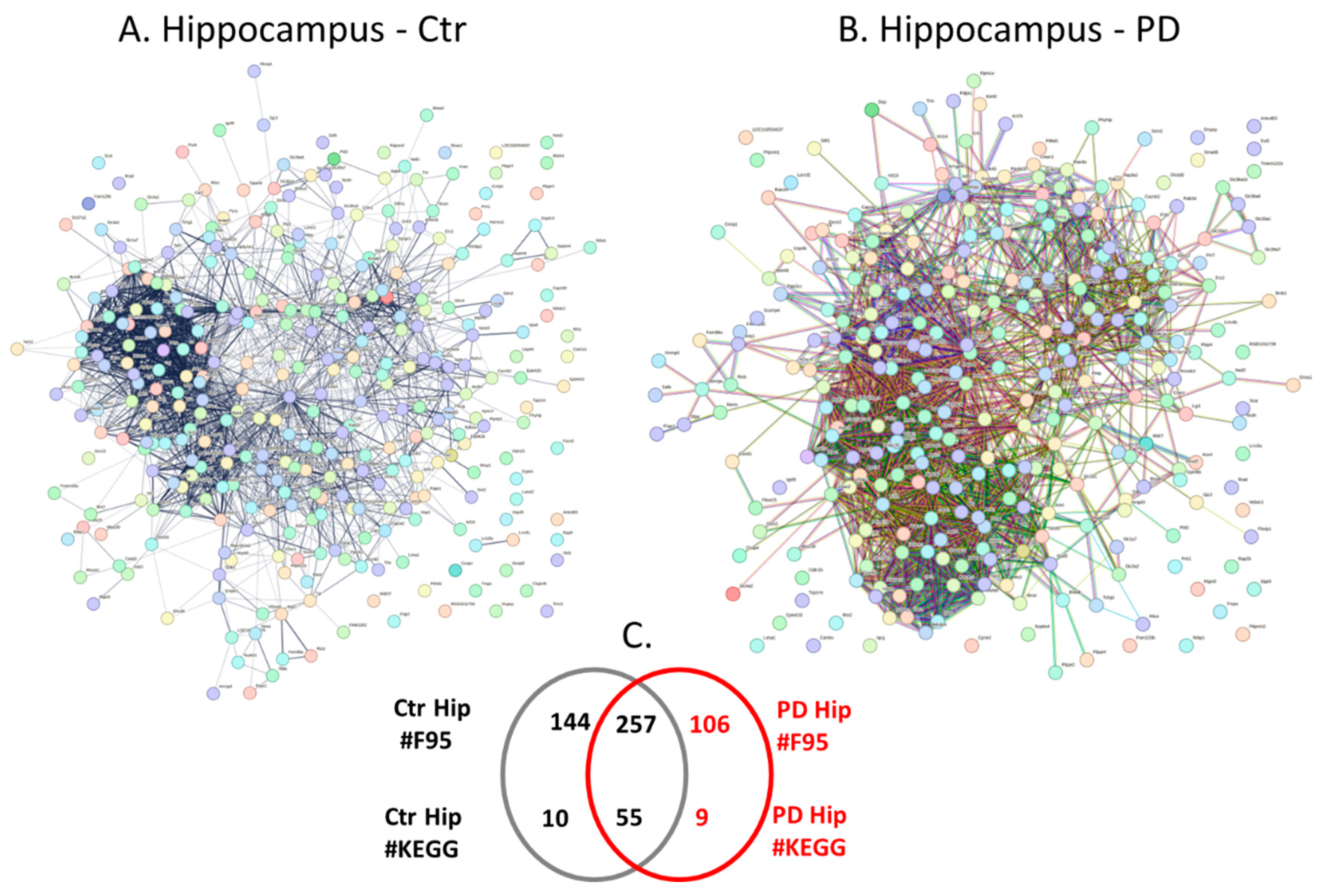
Figure 7.
The citrullinomes of sham/control and pre-motor PD rat cerebellum. A) Protein interaction networks of the protein citrullinome of control rat cerebellum. B) Protein interaction network of the protein citrullinome of PD rat cerebellum. C) A Venn diagram representing number (#) of citrullinated protein hits (F95) and associated KEGG pathways in control and pre-motor PD cerebellum.
Figure 7.
The citrullinomes of sham/control and pre-motor PD rat cerebellum. A) Protein interaction networks of the protein citrullinome of control rat cerebellum. B) Protein interaction network of the protein citrullinome of PD rat cerebellum. C) A Venn diagram representing number (#) of citrullinated protein hits (F95) and associated KEGG pathways in control and pre-motor PD cerebellum.
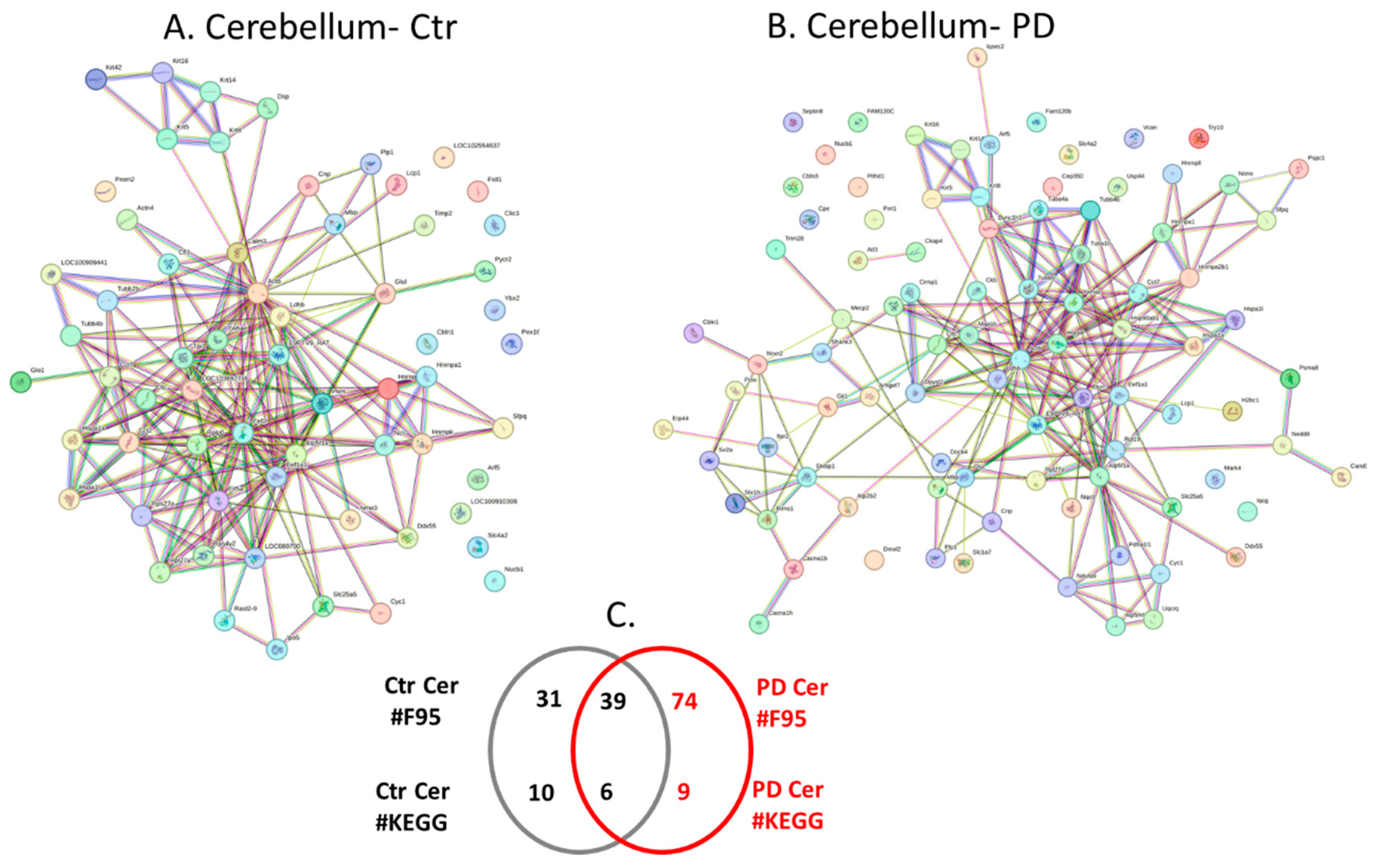
Figure 8.
The citrullinomes of sham/control and pre-motor PD rat midbrain. A) Protein interaction networks of the protein citrullinome of control rat midbrain. B) Protein interaction network of the protein citrullinome of PD rat midbrain. C) A Venn diagram representing number (#) of citrullinated protein hits (F95) and associated KEGG pathways in control and pre-motor PD midbrain.
Figure 8.
The citrullinomes of sham/control and pre-motor PD rat midbrain. A) Protein interaction networks of the protein citrullinome of control rat midbrain. B) Protein interaction network of the protein citrullinome of PD rat midbrain. C) A Venn diagram representing number (#) of citrullinated protein hits (F95) and associated KEGG pathways in control and pre-motor PD midbrain.
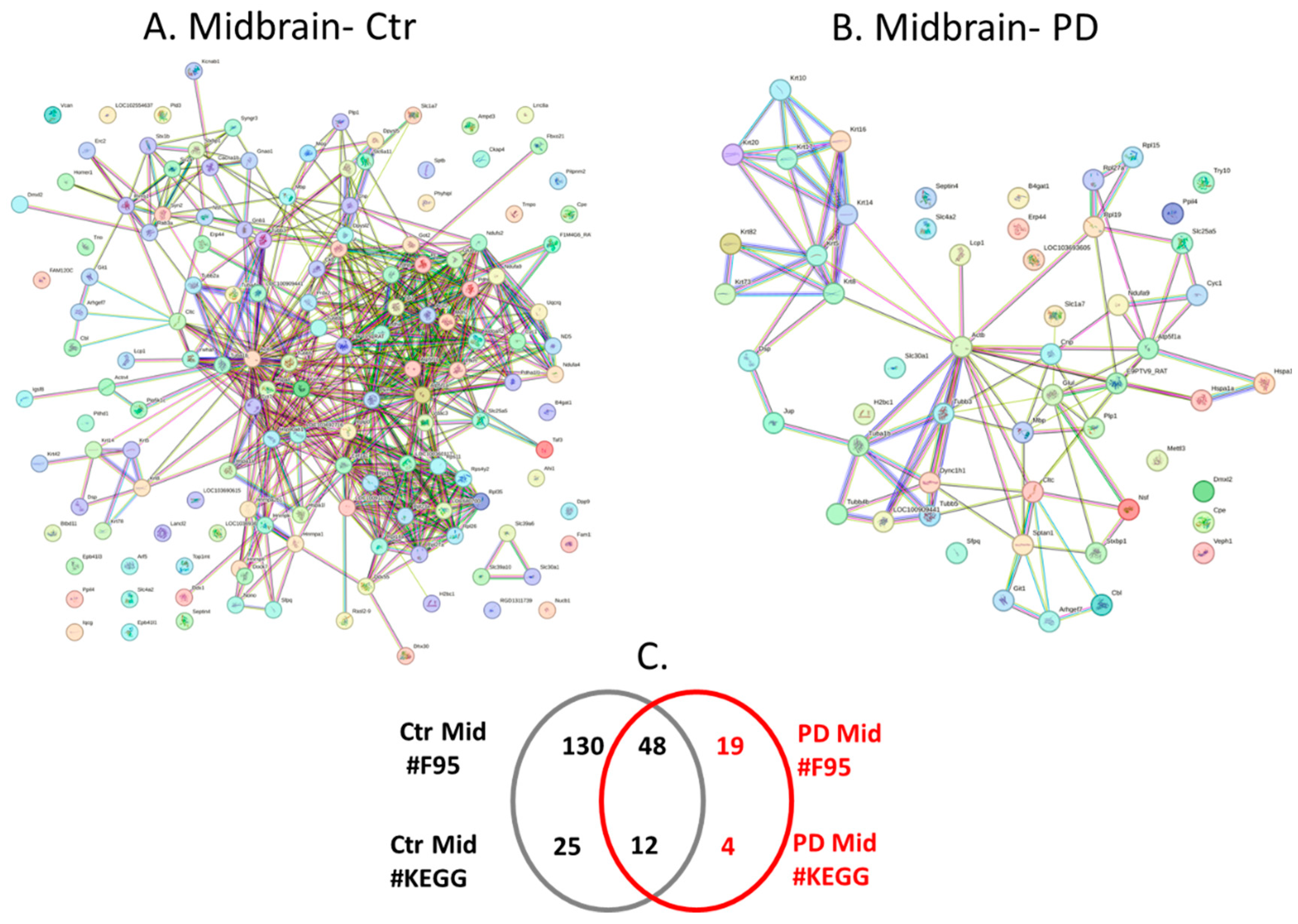
Figure 9.
The citrullinomes of sham/control and pre-motor PD rat olfactory bulb. A) Protein interaction networks of the protein citrullinome of control rat olfactory bulb. B) Protein interaction network of the protein citrullinome of PD rat olfactory bulb. C) A Venn diagram representing the number (#) of citrullinated protein hits (F95) and associated KEGG pathways in control and pre-motor PD olfactory bulb.
Figure 9.
The citrullinomes of sham/control and pre-motor PD rat olfactory bulb. A) Protein interaction networks of the protein citrullinome of control rat olfactory bulb. B) Protein interaction network of the protein citrullinome of PD rat olfactory bulb. C) A Venn diagram representing the number (#) of citrullinated protein hits (F95) and associated KEGG pathways in control and pre-motor PD olfactory bulb.
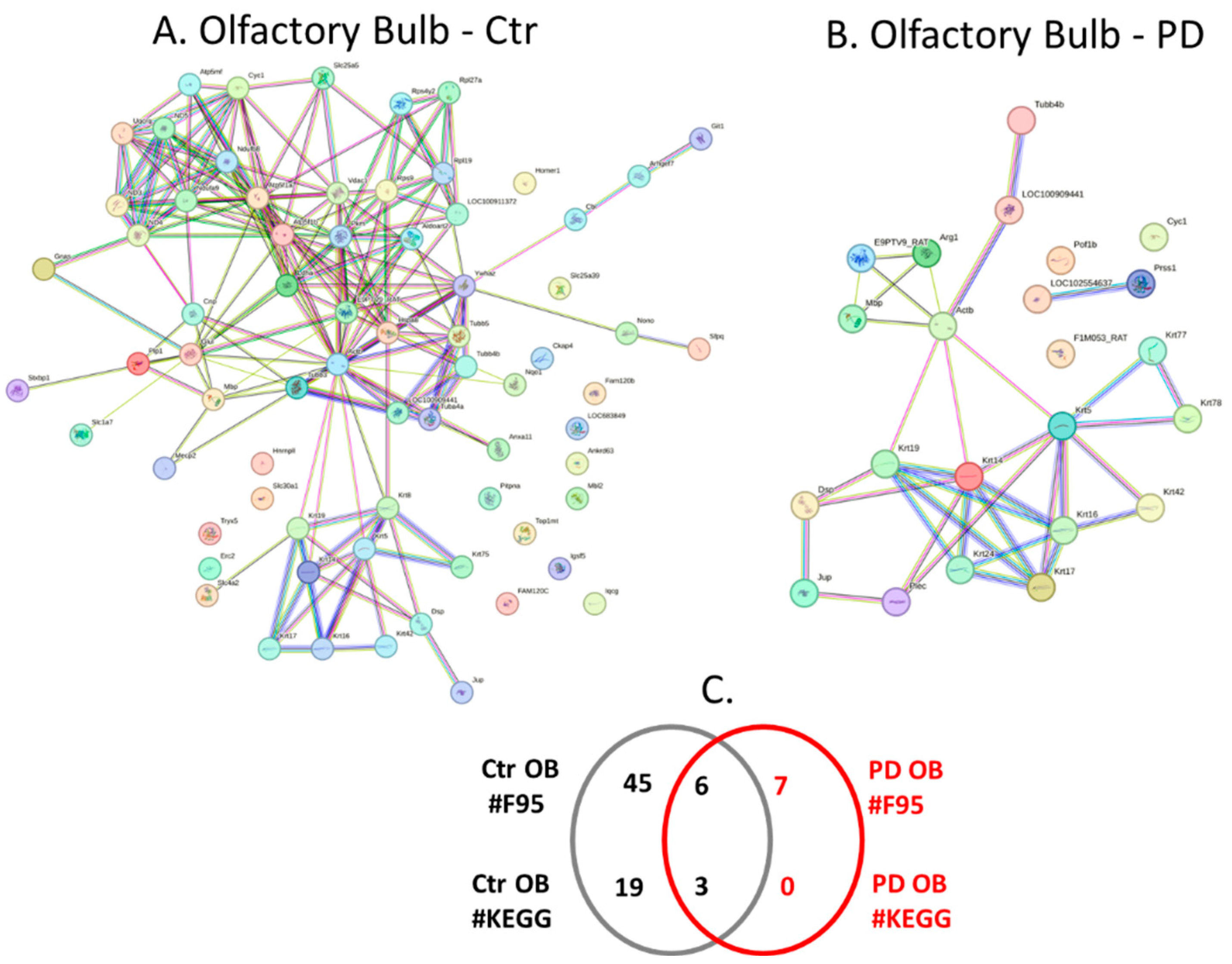
Figure 10.
The citrullinomes of sham/control and pre-motor PD rat striatum. A) Protein interaction networks of the protein citrullinome of control rat striatum. B) Protein interaction network of the protein citrullinome of PD rat striatum. C) A Venn diagram representing citrullinated protein hits (F95) and number of associated KEGG pathways in control and pre-motor PD striatum.
Figure 10.
The citrullinomes of sham/control and pre-motor PD rat striatum. A) Protein interaction networks of the protein citrullinome of control rat striatum. B) Protein interaction network of the protein citrullinome of PD rat striatum. C) A Venn diagram representing citrullinated protein hits (F95) and number of associated KEGG pathways in control and pre-motor PD striatum.
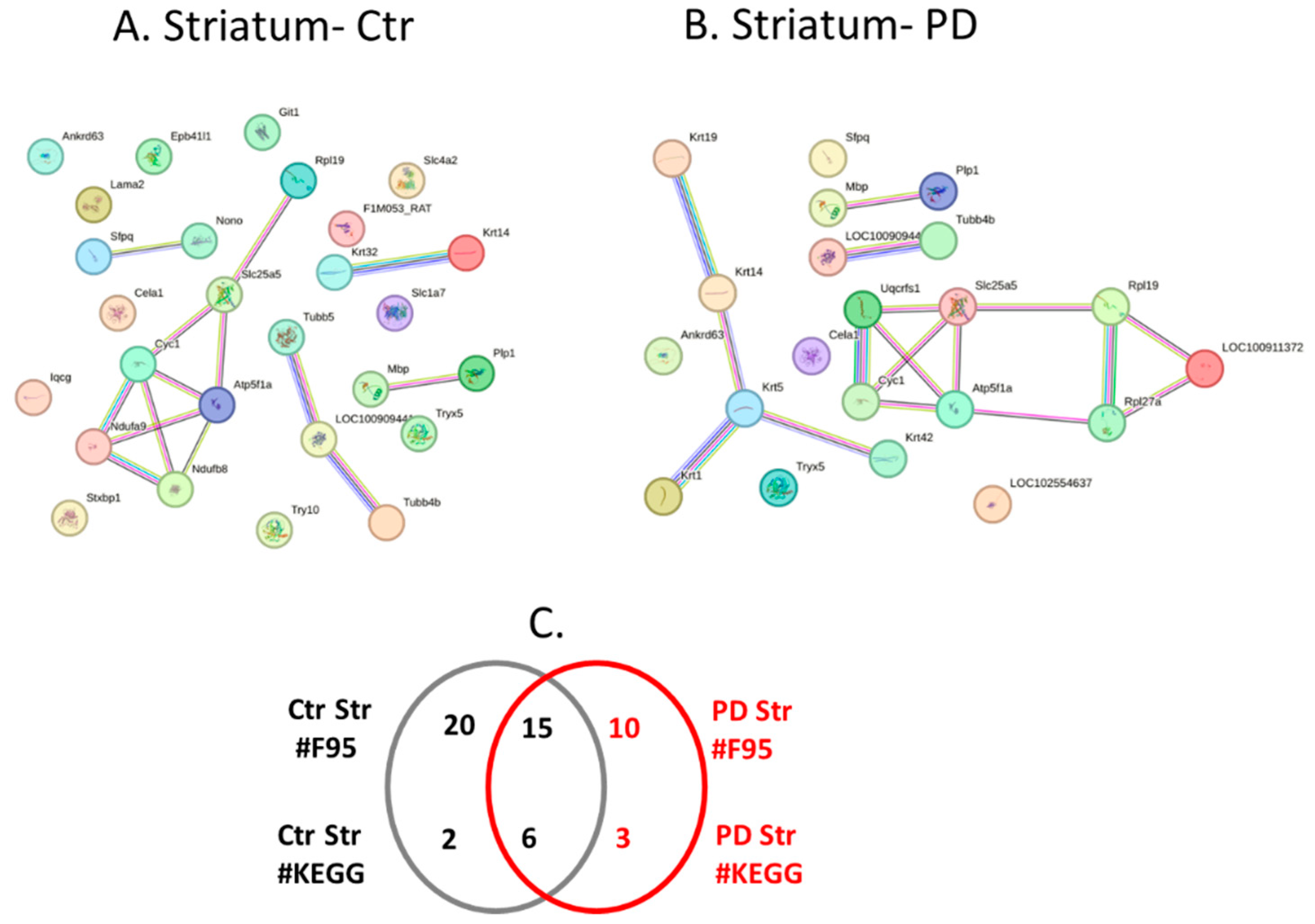
Figure 11.
The brain region specific citrullinomes of control and pre-motor PD rat brains. A) KEGG and Reactome pathways associated to the protein citrullinomes of the six brain regions assessed, in control and PD groups. B) Biological GO, Molecular Function GO and Cellular Component GO pathways associated to the protein citrullnomes of the six brain regions assessed, in control and PD groups.
Figure 11.
The brain region specific citrullinomes of control and pre-motor PD rat brains. A) KEGG and Reactome pathways associated to the protein citrullinomes of the six brain regions assessed, in control and PD groups. B) Biological GO, Molecular Function GO and Cellular Component GO pathways associated to the protein citrullnomes of the six brain regions assessed, in control and PD groups.
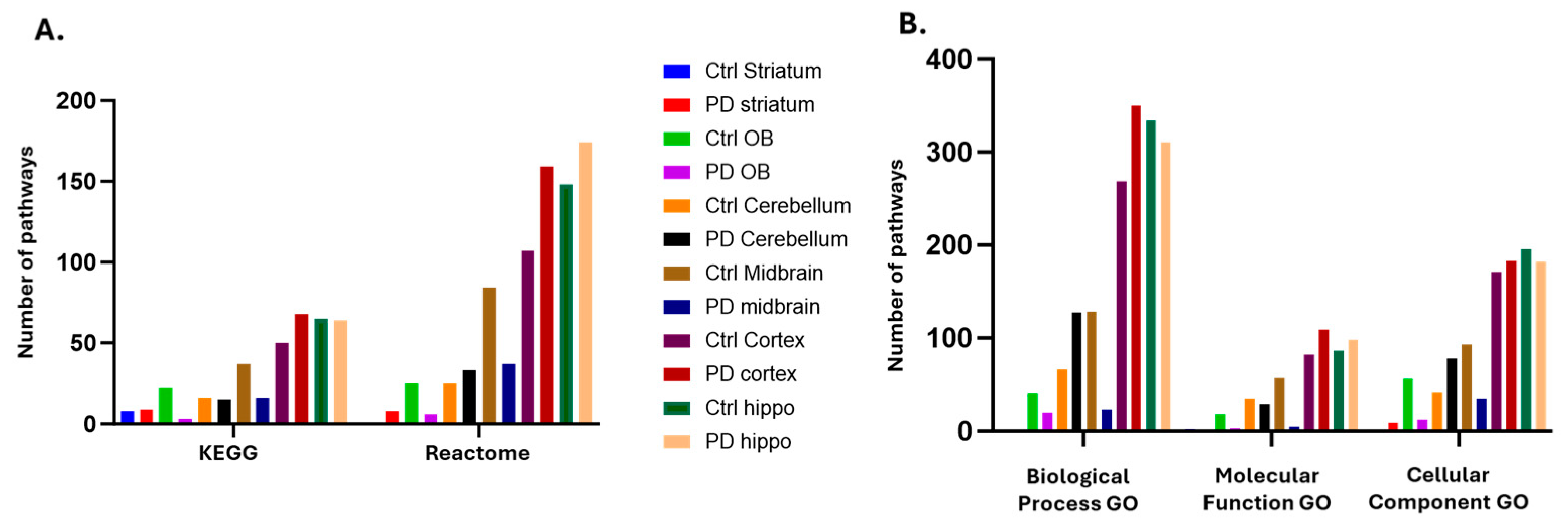
Figure 12.
Experimental setup. Three animals per group were used for sham/control and the pre-motor PD model, respectively. Brains were extracted at the end of the treatment and the six brain regions under study were excised and analysed for PADs by western blotting and for brain-region specific citrullinomes by proteomics analysis (LC-MS/MS).
Figure 12.
Experimental setup. Three animals per group were used for sham/control and the pre-motor PD model, respectively. Brains were extracted at the end of the treatment and the six brain regions under study were excised and analysed for PADs by western blotting and for brain-region specific citrullinomes by proteomics analysis (LC-MS/MS).
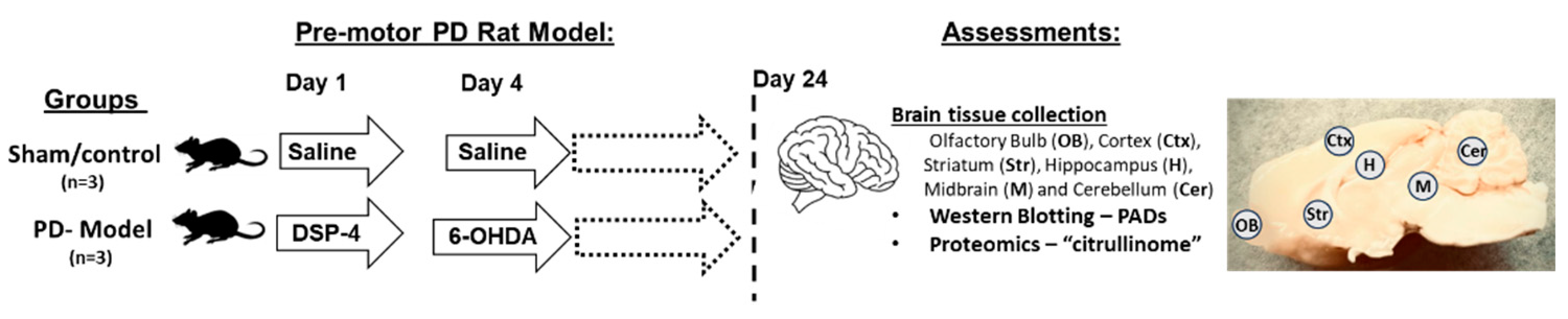

Table 1.
Summary of PAD isozymes (PAD1,2,3,4 and 6) detection, as assessed by western blotting in sham/control (CTR) and pre-motor PD (PD) cortex, hippocampus, cerebellum, midbrain, olfactory bulb and striatum. Scoring, following normalization with beta-actin, is shown as + low positive, ++ medium positive, +++ high positive. Brackets () indicate that the full score is not reached; statistically significant differences (p < 0.05) are indicated by a red star (*).
Table 1.
Summary of PAD isozymes (PAD1,2,3,4 and 6) detection, as assessed by western blotting in sham/control (CTR) and pre-motor PD (PD) cortex, hippocampus, cerebellum, midbrain, olfactory bulb and striatum. Scoring, following normalization with beta-actin, is shown as + low positive, ++ medium positive, +++ high positive. Brackets () indicate that the full score is not reached; statistically significant differences (p < 0.05) are indicated by a red star (*).
| Brain region | Exp group | PAD1 | PAD2 | PAD3 | PAD4 | PAD6 |
| Cortex | CTR | (+) | ++ | + | +++ | ++ |
| Cortex | PD | (+) | +++ * | ++ * | +++ | +++ |
| Hippocampus | CTR | (+) | + | ++ | ++ | ++ |
| Hippocampus | PD | (+) | (+) | +(+) | +(+) | ++(+) |
| Cerebellum | CTR | + | ++ | + | + | + |
| Cerebellum | PD | + | ++ | ++ * | +(+) * | +(+) * |
| Midbrain | CTR | (+) | ++ | +(+) | +(+) | ++ |
| Midbrain | PD | + | ++ | +(+) | + | ++ |
| Olfactory bulb | CTR | (+) | + | + | +(+) | + |
| Olfactory bulb | PD | + | + | +(+) * | ++ | ++ |
| Striatum | CTR | ++ | +++ | ++ | +++ | + |
| Striatum | PD | + * | + * | (+) * | + * | (+) * |
Table 2.
KEGG pathways associated with the brain region specific citrullinomes. Protein interaction networks were built based on citrullinated protein hits names, per brain region (cortex CTX, hippocampus HIP, striatum STR, olfactory bulb OB, midbrain MID, cerebellum CER), in sham/control (ctr) and pre-motor PD (PD) brains. KEGG pathways were identified in STRING. All KEGG pathways identified are listed below and a tick (V) indicates the KEGG pathway associated with each brain-region specific citrullinome. The columns for the PD brain regions are highlighted in grey.
Table 2.
KEGG pathways associated with the brain region specific citrullinomes. Protein interaction networks were built based on citrullinated protein hits names, per brain region (cortex CTX, hippocampus HIP, striatum STR, olfactory bulb OB, midbrain MID, cerebellum CER), in sham/control (ctr) and pre-motor PD (PD) brains. KEGG pathways were identified in STRING. All KEGG pathways identified are listed below and a tick (V) indicates the KEGG pathway associated with each brain-region specific citrullinome. The columns for the PD brain regions are highlighted in grey.
| KEGG pathway | Ctr CTX | PD CTX | CtrlHIP | PD HIP | Ctr STR |
PD STR | Ctr OB | PD OB | Ctr MB | PD MB | Ctr CER | PD CER |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Oxidative phosphorylation | V | V | V | V | V | V | V | V | V | |||
| Synaptic vesicle cycle | V | V | V | V | V | V | V | |||||
| Ribosome | V | V | V | V | V | V | V | V | ||||
| Parkinson’s disease | V | V | V | V | V | V | V | V | V | V | V | |
| Retrograde endocannabinoid signalling | V | V | V | V | V | V | ||||||
| Huntington disease | V | V | V | V | V | V | V | V | V | V | V | |
| Prion disease | V | V | V | V | V | V | V | V | V | V | V | |
| Gap junction | V | V | V | V | V | V | V | V | ||||
| Non-alcoholic fatty liver disease | V | V | V | V | V | V | V | |||||
| Thermogenesis | V | V | V | V | V | V | V | V | V | V | ||
| Alzheimer’s disease | V | V | V | V | V | V | V | V | V | V | ||
| GABAergic synapse | V | V | V | V | V | |||||||
| Amyotrophic lateral sclerosis | V | V | V | V | V | V | V | V | V | V | ||
| Endocrine and other factor-regulated calcium reabsorption | V | V | ||||||||||
| Glutamatergic synapse | V | V | V | V | V | V | V | |||||
| Phagosome | V | V | V | V | V | V | V | V | ||||
| Cardiac muscle contraction | V | V | V | V | ||||||||
| Biosynthesis of amino acids | V | V | V | V | V | V | V | |||||
| Carbon metabolism | V | V | V | V | V | |||||||
| Oestrogen signalling pathway | V | V | V | V | V | V | V | V | V | V | V | |
| Necroptosis | V | V | V | V | V | |||||||
| Endocytosis | V | V | V | V | V | V | V | |||||
| Metabolic pathways | V | V | V | V | V | V | ||||||
| Cocaine addiction | V | V | ||||||||||
| Insulin secretion | V | V | V | V | ||||||||
| Apoptosis | V | V | V | V | V | V | ||||||
| cGMP-PKG signalling pathway | V | V | V | |||||||||
| Alcoholism | V | V | V | |||||||||
| Pyruvate metabolism | V | V | V | V | V | V | ||||||
| Inositol phosphate metabolism | V | |||||||||||
| Phosphatidylinositol signalling system | V | V | ||||||||||
| Progesterone mediated oocyte maturation | V | |||||||||||
| Axon guidance | V | |||||||||||
| Oocyte meiosis | V | V | ||||||||||
| Cholinergic synapse | V | V | V | V | ||||||||
| Spinocerebellar ataxia | V | |||||||||||
| Phospholipase D signalling pathway | V | |||||||||||
| Rap1 signalling pathway | V | |||||||||||
| 2-Oxocarboxylic acid metabolism | V | V | ||||||||||
| Nitrogen metabolism | V | V | V | V | ||||||||
| Pentose phosphate pathway | V | V | ||||||||||
| Fructose and mannose metabolism | V | V | V | V | ||||||||
| Bacterial invasion of epithelial cells | V | V | V | V | V | |||||||
| Leukocyte transendothelial migration | V | V | ||||||||||
| Morphine addiction | V | V | V | V | ||||||||
| Regulation of actin cytoskeleton | V | V | V | |||||||||
| Citrate cycle (TCA cycle) | V | V | V | V | V | |||||||
| Vasopressin-regulated water reabsorption | V | V | V | |||||||||
| Arginine biosynthesis | V | V | V | V | ||||||||
| Glycolysis/Gluconeogenesis | V | V | V | V | V | V | V | |||||
| Central carbon metabolism in cancer | V | V | V | V | V | |||||||
| SNARE interactions in vesicular transport | V | V | V | |||||||||
| Alanine, aspartate and glutamate metabolism | V | V | V | V | ||||||||
| Cysteine and methionine metabolism | V | V | V | V | ||||||||
| HIF-1 signalling pathway | V | V | V | V | V | |||||||
| Legionellosis | V | V | V | V | V | |||||||
| Salmonella infection | V | V | V | V | ||||||||
| Oxytocin signalling pathway | V | V | V | V | ||||||||
| Tight junction | V | V | V | |||||||||
| mTOR signalling pathway | V | |||||||||||
| Viral carcinogenesis | V | V | V | V | V | |||||||
| Collecting duct acid secretion | V | |||||||||||
| Glyoxylate and dicarboxylate metabolism | V | V | V | |||||||||
| Butanoate metabolism | V | |||||||||||
| Dopaminergic synapse | V | V | V | V | ||||||||
| Calcium signalling pathway | V | V | V | V | ||||||||
| Ferroptosis | V | V | ||||||||||
| Systemic lupus erythematosus | V | V | ||||||||||
| Long-term depression | V | V | ||||||||||
| Hippo signalling pathway | V | |||||||||||
| Staphylococcus aureus infection | V | V | V | V | V | |||||||
| Gastric acid secretion | V | V | V | V | ||||||||
| Arrhythmogenic right ventricular cardiomyopathy | V | V | V | |||||||||
| Influenza A | V | |||||||||||
| Glucagon signalling pathway | V | V | V | V | V | V | V | |||||
| GnRH secretion | V | |||||||||||
| Type II diabetes mellitus | V | V | V | V | ||||||||
| Apelin signalling pathway | V | |||||||||||
| Proteoglycans in cancer | V | V | V | |||||||||
| NOD-like receptor signalling pathway | V | |||||||||||
| Platelet activation | V | |||||||||||
| Cellular senescence | V | V | ||||||||||
| Yersinia infection | V | |||||||||||
| Human immunodeficiency virus 1 infection | V | V | V | V | ||||||||
| Propanoate metabolism | V | V | ||||||||||
| Serotonergic synapse | V | V | V | |||||||||
| Beta-Alanine metabolism | V | |||||||||||
| Thyroid hormone signalling pathway | V | |||||||||||
| Fc gamma R-mediated phagocytosis | V | |||||||||||
| Choline metabolism in cancer | V | |||||||||||
| Spliceosome | V | |||||||||||
| Antigen processing and presentation | V | V | ||||||||||
| Protein processing in endoplasmic reticulum | V | |||||||||||
| Human cytomegalovirus infection | V |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



