
Submitted:
30 June 2024
Posted:
01 July 2024
You are already at the latest version
Abstract
Direct-acting antivirals (DAAs) revolutionized therapeutics of chronic hepatitis C. The emergence and transmission of HCV variants with resistance-associated substitutions (RASs) can undermine HCV treatment. This study aimed to assess the prevalence and temporal trends of RASs in HCV with a particular focus on the clinically-relevant RASs (cr-RASs). Near-complete HCV GenBank sequences archived in the Los Alamos HCV Database were analyzed. The study period was divided into two phases; before 2011 and 2011 onwards. Identification of RASs across three DAA classes; NS3, NS5A, and NS5B inhibitors was based on the 2020 EASL guidelines. The AASLD-IDSA recommendations were used to identify cr-RASs for three HCV genotypes/subtypes (1a, 1b, and 3) for four DAA regimens; ledipasvir/sofosbuvir; elbasvir/grazoprevir; sofosbuvir/velpatasvir; and glecaprevir/pibrentasvir. The final HCV dataset comprised 3443 sequences and the prevalences of RASs were 50.4%, 60.2%, and 25.3% in NS3, NS5A, and NS5B, respectively. In subtype 1a, resistance to ledipasvir/sofosbuvir was 32.8%, while resistance to elbasvir/grazoprevir was 33.0%. For genotype 3, resistance to sofosbuvir/velpatasvir and glecaprevir/pibrentasvir was 4.2% and 24.9%, respectively. Significant increase in cr-RASs was observed across the two study phases as follows: for ledipasvir/sofosbuvir in subtype 1a, cr-RASs increased from 30.2% to 35.8% (p=0.019); for elbasvir/grazoprevir in subtype 1a, cr-RASs increased from 30.4% to 36.1% (p=0.018); In subtype 1b, neither ledipasvir/sofosbuvir nor elbasvir/grazoprevir showed any cr-RASs in the first phase, but both were present at a prevalence of 6.5% in the second phase (p<0.001); for sofosbuvir/velpatasvir in genotype 3, cr-RASs increased from 0.9% to 5.2% (p=0.006); and for glecaprevir/pibrentasvir, cr-RASs increased from 12.0% to 29.1% (p<0.001). Rising prevalence of HCV RASs and cr-RASs was discernible. This highlights the necessity for ongoing surveillance and adaptation of novel therapeutics to manage resistance effectively. Updating of the clinical guidelines and treatment regimens are recommended to counteract the evolving resistance to DAAs.
Keywords:
Resistance
; therapy
; treatment response
; treatment as prevention
; HCV elimination
1. Introduction
Hepatitis C virus (HCV) infection continues to represent a formidable public health challenge, with an estimated 50 million people having the chronic HCV globally [1,2,3]. Additionally, about one million new HCV infections are estimated to occur each year, with an estimated mortality of slightly less than quarter a million in 2022 [1]. The chronic nature of HCV infection in a majority of infected individuals can lead to grave outcomes including cirrhosis and hepatocellular carcinoma (HCC), contributing significantly to global morbidity and mortality [4,5,6,7].
In 2011 onwards, the approval and widespread use of different direct-acting antivirals (DAAs) represented a critical breakthrough for treatment of HCV [8,9]. The DAAs were reported to offer cure rates exceeding 90% across various HCV genotypes (e.g., genotype 1 and 4) [10,11]. Nevertheless, lower cure rates (66–80%) were reported for genotype 3 especially among cirrhotic patients, justifying its description as the possible “last stand” for HCV or alternatively “the new HCV villain” [12,13,14,15]. In addition to the high cure rates, the use of DAAs reduced the duration of treatment needed to achieve sustained virologic response (SVR) into 12 weeks [16,17]. Importantly, the introduction and use of DAAs enhanced the implementation of the treatment as prevention (TasP) strategy, significantly reducing HCV transmission (e.g., among injection drug users (IDUs) in Europe) [18,19]. This was highlighted by van Santen et al. in a multinational cohort study which showed a decrease in pooled incidence of HCV from 0.91 per 100 person-years in 2015 to 0.41 per 100 person-years in 2019 [20]. The TasP strategy is critical to achieve the World Health Organization (WHO) goal of HCV elimination as a public health threat by 2030 considering the current lack of an effective HCV vaccine and the challenges to achieve such a vaccine in the near future [21,22,23].
The current therapeutic strategies for HCV employ a combination of DAAs that target specific viral proteins, namely the NS3 (protease), NS5A (critical component of HCV replication complex), and NS5B (the HCV RNA-dependent RNA polymerase (RdRp)) [3,24]. The HCV treatment regimens are tailored based on various factors such as the HCV genotype and viral load, prior treatment, co-infection with human immunodeficiency virus (HIV), and the presence of liver cirrhosis among other factors [25,26,27].
Despite the remarkable success of DAAs to reduce the HCV burden, the emergence of resistance-associated substitutions (RASs) represents an increasing challenge [28,29,30]. The rapid evolution of HCV driven by error-prone RdRp facilitates the emergence of RASs with subsequent selective advantage under drug pressure [31,32]. The RASs are defined as the amino acid substitutions that reduce the potency of a DAA in vitro or in vivo, with the possibility of treatment failure [30,33,34]. Importantly, the RASs vary based on the specific HCV genotypes/subtypes due to the high genetic diversity of HCV, which leads to different reference sequences (wild types) for different HCV genotypes/subtypes [3,35].
The characterization of RASs initially emerged from studies involving patients treated with DAAs [34,36]. It is important to emphasize that the detection of a RAS per se is not necessarily an indication of resistance to DAAs. Instead, the likelihood of a DAA to select for a RAS conferring resistance is influenced by the genetic barrier to resistance, the level of drug exposure, and the replicative fitness of the HCV variant [37]. It is also important to highlight that the genetic barriers to resistance vary across different DAAs and are further influenced by variations in HCV genotypes/subtypes [38,39,40].
The evaluation of real-world clinical impact of RASs have been limited and showed variable results [41,42,43,44,45]. In these studies, the presence of RASs was infrequently associated with treatment failure; nevertheless, RASs were associated treatment failure more commonly in patients with cirrhosis [46]. Notably, the presence of baseline NS5A RASs is among the factors to be considered for therapy with NS5A inhibitors —at least for some regimens— due to the relatively low genetic barrier for resistance [27,38,47].
An important concept should be clarified at this point in relation to the distinction between RASs versus the clinically relevant RASs (cr-RASs) [3,26]. The cr-RASs can be defined as the mutations that have been empirically shown to reduce the effectiveness of DAAs manifested in high fold change decrease in potency [26,47,48]. Thus, the characterization of cr-RAS can be crucial for optimizing HCV treatment as their presence can necessitate adjustments in therapeutic approaches, such as the selection of more potent DAA combinations or the inclusion of additional therapeutic agents to overcome resistance [46,47].
Among the available DAAs in formulation, four regimens—ledipasvir/sofosbuvir (Harvoni), elbasvir/grazoprevir (Zepatier), sofosbuvir/velpatasvir (Epclusa), and glecaprevir/pibrentasvir (Mavyret)—have been distinguished for their broad genotypic effectiveness and high barrier to resistance [3,49,50]. These regimens offer potent therapeutic options to accommodate a wide range of patient demographics and virological profiles. Therefore, monitoring the prevalence and trends of cr-RASs among these regimens is essential to guide clinical and public health strategies aimed at HCV management, prevention, and possible elimination. This surveillance can enable the prediction of potential treatment outcomes and proactively adapt new therapeutic approaches. Moreover, understanding these trends helps in anticipating future challenges in the fight against HCV, which would help to ensure that HCV therapeutic regimens remain effective and that the goals of reducing HCV disease burden and achieving viral elimination by 2030 are met. Therefore, this study aimed to assess the prevalence of RASs, cr-RASs, and its temporal trend using publicly available HCV GenBank sequences across the commonly found genotypes/subtypes.
2. Materials and Methods
2.1. Study Design
This study was based on detailed analysis of near-complete HCV genomes, which were obtained from the GenBank sequences archived in the Los Alamos HCV Database (LAHCVDB) [51,52,53]. The selection of near-complete genomes aimed to ensure highly accurate HCV genotype/subtype assignment, which was essential for subsequent analysis of RASs prevalence per HCV genotype/subtype.
2.2. Sequence Retrieval, Selection, Alignment, and Subtyping
The initial phase of sequence retrieval started by downloading full HCV genomes available at LAHCVDB. A total of 5,004 HCV sequences in FASTA format were retrieved. The metadata for each sequence was downloaded as well including data on genotype/subtype, country of sequence collection (if available), year of sequence collection, and accession number. These metadata were standardized in the FASTA headers using FaBox version 1.61, an online fasta sequence toolbox [54]. Sequence visualization and translation into corresponding amino acids (aa) sequence was conducted in MEGA6 software [55].
To accommodate subsequent temporal analysis, the retrieved HCV sequences which lacked complete metadata regarding the timing of collection were excluded. The remaining sequences underwent alignment against the reference HCV sequence (H77, GenBank accession number: AF009606) using the MAFFT multiple sequence alignment tool.[56,57]
Then, the aligned HCV sequences were manually inspected for alignment accuracy (by the first and senior authors independently) and then the alignment was subdivided into three HCV genomic regions based on their positions relative to the H77 reference: NS3 (positions: 3420–5312), NS5A (positions: 6258–7601), and NS5B (positions: 7602–9374). Afterwards, the sequences in each dataset were screened to exclude any that contained stop codons or extensive regions of missing nucleotides (greater than 30 bases).
The final dataset included 3,488 sequences for NS3, 3,487 for NS5A, and 3,488 for NS5B, excluding the reference sequence H77. Additional filtration steps were applied to exclude genotype 5 sequences due to their limited numbers (n=5) and any recombinant forms (n=4), and non-1a, non-1b genotype 1 (n=36) due to their limited numbers which would preclude reliable assessment of RASs prevalence in these genetic variants of HCV. The final list of 3443 HCV sequences analyzed in this study are available in (Supplementary Table S1).
Finally, HCV subtype assignment was confirmed for the final HCV nucleotide sequences using the COMET HCV tool, which rapidly classifies HCV sequences into subtypes [58].
2.3. Assessment of RASs in the Final HCV Dataset for the Three DAA Classes
To identify RAS indicating resistance to DAAs across three key classes; NS3 inhibitors, NS5A inhibitors, and NS5B inhibitors, we opted to adhere to the 2020 European Association for the Study of the Liver (EASL) Guidelines for the Treatment of Hepatitis C [25]. This involved analyzing specific aa positions known to influence drug response within the NS3, NS5A, and NS5B protein regions [25]. The positions analyzed were 36, 41, 43, 54, 55, 56, 80, 122, 155, 156, 158, 166, 168, 170, and 175 for NS3; 24, 26, 28, 29, 30, 31, 32, 38, 58, 62, 92, and 93 for NS5A; 150, 159, 206, 282, 316, 320, and 321 for NS5B NIs, and 314, 316, 368, 395, 411, 414, 445, 446, 448, 553, 554, 555, 556, 557, 558, 559, 561, and 565 for NNIs NS5B [25].
For the prevalence analysis, we quantified the occurrence of genotype-specific RAS at these positions and assessed their prevalence across the whole HCV genotype/subtype dataset. The presence of ambiguous aa, denoted by “?” in the alignment, which could result from low sequencing quality, or the presence of multiple viral strains within the sample, a conservative approach was adopted; ambiguous amino acids were not classified as resistant. This methodological caution was chosen to avoid overestimation of RASs prevalence.
2.4. Assessment of Clinically-Relevant RASs by DAA and HCV Genotypes/Subtypes
As stated in the Introduction, it was crucial to distinguish between mere RAS and (cr-RAS). Therefore, we put a special focus on the cr-RASs relying on the “HCV Guidance” by AASLD and IDSA criteria for identification of cr-RAS as follows [26].
The AASLD-IDSA recommendations for testing, managing, and treating hepatitis C were based on the use data for three genotypes/subtypes (1a, 1b, and 3) and concerning four DAA regimens ledipasvir/sofosbuvir (Harvoni); elbasvir/grazoprevir (Zepatier); sofosbuvir/velpatasvir (Epclusa); and glecaprevir/pibrentasvir (Mavyret) [26]. For Harvoni, the following RASs were deemed cr-RAS for subtype 1a: Q30H/R, L31M/V, or Y93C/H/N; and for subtype 1b: L31V or Y93H. For Zepatier, the following RASs were deemed cr-RAS for subtype 1a: M28A/T, Q30H/R, L31M/V, or Y93C/H/N and for subtype 1b: Y93H. For Epclusa, the following RAS was deemed cr-RAS for genotype 3: Y93H. For Mavyret, the following RAS was deemed cr-RAS for genotype 3: A30K [26].
2.5. Data and Statistical Analysis
Statistical analysis was conducted using IBM SPSS Statistics for Windows, Version 27.0 (IBM Corp., Armonk, NY). To explore the associations between categorical variables, we employed the chi-squared test (χ2). To evaluate temporal trends, we utilized the linear-by-linear association test (LBL). The threshold for statistical significance was set at p < 0.050. To compute the prevalences, we applied the adjusted Wald method to obtain 95% confidence intervals (CIs). For effect size in differences in the prevalence of RASs and cr-RASs over the two study phases, we calculated the Cohen’s d using the following formula= (M2 − M1) ⁄ SD pooled, where SD pooled = √ ((SD12 + SD22) ⁄ 2) and M is the mean calculated based on assigning the presence of RAS as “1” and its absence as “zero”.
Geographically, the analysis categorized countries into six regions: (1) North America, Western Europe, and Australia; (2) Eastern Europe, Russia, and Former Soviet Union countries; (3) Asia; (4) the Middle East and North Africa (MENA); (5) Africa; and (6) Latin America and the Caribbean. The study timeline was bifurcated into two phases for temporal analysis: the first phase spanning from 1983 to 2010, before the approval of the first DAA in 2011, and the second phase from 2011 to 2022. This division was based on the attempt to examine the RASs trends before and after the widespread introduction of DAAs.
3. Results
3.1. HCV Dataset Characteristics
The final HCV dataset comprised a total of 3443 sequences distributed as follows: subtype 1a (n=1534, 44.6%), subtype 1b (n=617, 17.9%), genotype 2 (n=244, 7.1%), genotype 3 (n=851, 24.7%), genotype 4 (n=116, 3.4%), and genotype 6 (n=81, 2.4%). In the first phase of the study (1983–2010) the number of included HCV sequences was 1650 (47.9%), while in the second phase of the study (2011–2022) the number of HCV sequences was 1793 (52.1%). The number of HCV sequences belonging to genotype 3 increased significantly over the study period rising from 13.1% in the first phase to 35.4% in the second phase (P<0.001, LBL), while subtypes 1a and 1b decreased significantly from 50.7% and 21.6% in the first phase to 38.9% and 14.6%, respectively.
The majority of included sequences were collected in North America/Western Europe, or Australia with 2393 sequences (69.5%). The second most common region was Asia with 413 sequences (12.0%), followed by Africa (n=107, 3.1%), the MENA (n=23, 0.7%), Latin America/the Caribbean (n=20, 0.6%), and only eight sequences were collected in Eastern Europe/Russia (0.2%). A total of 479 HCV sequences lacked information regarding country/region of collection. The overall prevalence of any RASs in the study sample was 85.8% (95% CI: 84.6%–87.0%), with an increase in the RASs prevalence from 78.9% (95% CI: 76.9%–80.9%) in the first phase to 92.1% (95% CI: 90.8%–93.3%, P<0.001, LBL) in the second phase.
3.2. The Prevalence and Trends of NS3 RASs
The overall prevalence of RASs in the NS3 for the included sequences was 50.4% (95% CI: 48.7%–52.1%). The majority of these NS3 RASs were observed as a single RAS within the sequences (n=1182, 68.1%), followed by three NS3 RASs (n=342, 19.7%), two RASs (n=202, 11.6%), and only nine sequences harbored four NS3 RASs (0.5%). The full description of the NS3 RASs stratified per HCV GT/SGT is shown in (Table 1).
Stratified per genotype/subtype, the highest prevalence of NS3 RASs was found in genotype 6 (80/81, 98.8%), while the lowest prevalence was found in genotype 4 (1/116, 0.9%, p<0.001, χ2= 1078.375, Figure 1A).
Over the study period, the prevalence of NS3 RASs increased from 45.6% (752/1650) during the first phase to 54.8% (983/1793) during the second phase (p<0.001, χ2=29.401, Cohen’s d=0.181). The increasing trend was statistically significant for subtype 1a, subtype b, genotype 3 (p<0.001), and genotype 2 (p=0.014, Figure 2A).
Per region, the highest percentage of NS3 RASs was observed in Africa (n=64/107, 59.8%), followed by Asia (n=239/413, 57.9%), while the lowest percentage was reported in Eastern Europe/Russia (n=2/8, 25.0%, p<0.001, χ2=22.651, Figure 3A).
3.3. The Prevalence and Trends of NS5A RASs
The overall prevalence of RASs in the NS5A for the included sequences was 60.2% (95% CI: 58.6%–61.9%). The vast majority of HCV sequences harboring NS5A RASs contained either three RASs (n=745, 35.9%), a single RAS (n=660, 31.8%), or two RASs (n=633, 30.5%), while 33 sequences contained four RASs (1.6%), and only two sequences contained five RASs (0.1%). The full description of the NS5A RASs stratified per HCV genotype/subtype is shown in (Table 2).
At least one NS5A RAS was found in all genotype 4 and genotype 6 sequences yielding a prevalence rate of 100% in these two genotypes. Apart from these two genotypes, the highest prevalence of NS5A RASs was observed in subtype 1b (n=551/617, 89.3%) while the lowest prevalence of NS5A RASs was observed in genotype 3 (n=396/851, 46.5%) and subtype 1a (n=721/1533, 47.0%, Figure 1B).
The prevalence of NS5A RASs increased from 57.0% (940/1649) during the first phase to 63.2% (1133/1793) during the second phase (p<0.001, χ2=13.722, Cohen’s d=0.123). The increasing trend was statistically significant for subtype 1a, genotype 2, and genotype 3 (p<0.001, Figure 2B).
Per region, the highest percentage of NS5A RASs was observed in the MENA and Africa (n=22/23, 95.7% and n=98/107, 91.6%, respectively), while the lowest percentage was reported in Western Europe/North America/Australia (n=1355/2392, 56.6%, p<0.001, χ2=122.158, Figure 3B).
3.4. The Prevalence and Trends of NS5B RASs
The overall prevalence of RASs in the NS5B for the included sequences was 25.3% (95% CI: 23.9%–26.8%). The majority of sequences identified with RASs had a single RAS (n=502, 57.6%), followed by sequences with three RASs (n=216, 24.8%), or two RASs (n=149, 17.1%), while only five HCV sequences had four RASs (0.6%). The full description of the NS5B RASs stratified per HCV genotype/subtype is shown in (Table 3).
The NS5B RASs were totally absent in all genotype 4 and genotype 6 sequences. Apart from these two genotypes, the highest prevalence of NS5B RASs was observed in subtype 1b (n=215/617, 34.8%) and subtype 1a (n=521/1533, 34.0%), while the lowest prevalence of NS5B RASs was observed in genotype 2 (n=1/244, 0.4%, Figure 1C).
The prevalence of NS5B RASs significantly increased from 17.5% (289/1650) during the first phase to 32.5% (583/1793) during the second phase (p<0.001, χ2=102.229, Cohen’s d=0.351). The increasing trend was statistically significant only for subtype 1b and genotype 3 (p<0.001, Figure 2C).
Per region, the highest percentage of NS5B RASs was observed in Eastern Europe/Russia (n=5/8, 62.5%) and Africa (n=58/107, 54.2%), while the lowest percentage was reported in Asia (n=66/413, 16.0%, p<0.001, χ2=108.898, Figure 3C).
3.5. The Prevalence of cr-RASs in Four Different DAA Regimens
We proceeded to testing the prevalence of cr-RASs as defined in AASLD-IDSA for four currently used DAA regimens namely Harvoni, Zepatier, Epclusa, and Mavyret.
A total of 502/1533 of subtype 1a had any cr-RAS in relation to Harvoni. A total of 506/1533 of subtype 1a had any cr-RAS in relation to Zepatier. The prevalence of resistance to Harvoni in subtype 1a was 32.8% (95% CI: 30.4%–35.1%). The prevalence of resistance to Harvoni and Zepatier in subtype 1b was 2.9% (95% CI: 1.7%–4.4%). The prevalence of resistance to Zepatier in subtype 1a was 33.0% (95% CI: 30.7%–35.4%), while the prevalence of resistance to Epclusa in genotype 3 was 4.2% (95% CI: 3.0%–5.7%). For Mavyret in genotype 3, the prevalence of resistance was 24.9% (95% CI: 22.0%–27.8%). The full details of the cr-RASs for the four DAA regimens are presented in (Table 4).
For the trends of cr-RAS, the prevalence of cr-RAS for Harvoni in subtype 1a increased from 30.2% during the first phase into 35.8% in the second phase (p=0.019, LBL, Cohen’s d=0.128). For Zepatier in subtype 1a, the prevalence of cr-RAS increased from 30.4% during the first phase into 36.1% in the second phase (p=0.018, LBL, Cohen’s d=0.106). In subtype 1b, for both Harvoni and Zepatier, the cr-RAS was totally absent in the first phase and was detected at a prevalence of 6.5% in the second phase (p<0.001, LBL). In genotype 3 and for Epclusa, the prevalence of cr-RAS increased from 0.9% in the first phase into 5.2% in the second phase (p=0.006, LBL, Cohen’s d=0.620). For Mavyret in genotype 3, the prevalence of cr-RAS increased from 12.0% in the first phase into 29.1% in the second phase (p<0.001, Cohen’s d=0.463, LBL, Figure 4).
Finally, the highest prevalence of cr-RASs was reported in Asia for SGT1a for both Harvoni and Zepatier, in Africa for SGT1b for Harvoni, Zepatier, and Epclusa for GT3, while the highest prevalence of Mavyret cr-RAS for GT3 was reported in Latin America/the Caribbean (Figure 5).
4. Discussion
The findings of this study demonstrated an increasing trend in the prevalence of RASs, including cr-RASs, over the study period. This trend is aligned with the relatively recent widespread adoption of DAAs as a curative therapeutic strategy for chronic hepatitis C, with subsequent selection of drug-resistant HCV variants that is an unintended consequence of widespread DAA use. However, the detection of RASs during the first phase of the study, prior to the approval of DAAs in 2011, suggested the natural occurrence of RASs, an observation that is consistent with the known rapid evolutionary rate of the HCV [32]. The relatively lower prevalence of naturally occurring RASs in the first phase of the study may be attributed to the lower replicative fitness of variants harboring these substitutions [60,61].
Consistent with our finding of a relatively high prevalence of RASs even prior to widespread use of DAAs, previous studies documented the presence of RASs in DAA-naïve populations [62]. For example, an early study by Paolucci et al. in Italy found that among treatment-naïve individuals, the prevalence of RASs in NS5A and NS5B for subtype 1a was 12.5%, while for subtype 1b, it was significantly higher at 53.3% and 90% for NS5A and NS5B, respectively [63]. A study involving 108 Argentinean DAA-naïve patients reported NS5A and NS5B RASs in 25.8% and 6.3% of the HCV sequences analyzed, respectively [64]. A more recent study in Saint Petersburg found a 12% prevalence of RASs among 42 patients [65].
In this comprehensive study, despite inherent biases in the analyzed HCV sequence data, we utilized 3,443 HCV sequences to assess trends in RASs across various HCV genotypes/subtypes and geographic regions from 1983 to 2022. Our findings indicated a clear temporal increase in both RASs and cr-RASs. Specifically, the analysis revealed substantial RAS prevalence rates of 50.4% in NS3, 60.2% in NS5A, and 25.3% in NS5B, suggesting a widespread occurrence of RASs. This trend is supported by the results of an earlier study by Chen et al. in 2016, which reported an overall RASs prevalence of 58.7% across 1,459 sequences [66]. Additionally, common occurrence of NS5A RASs in particular was reported by Kitrinos et al. at a baseline rate of 9.4% in subtypes 1a or 1b patients [67]. The notable escalation in RASs prevalence over time, as demonstrated by the significant rise from the first phase of our study period to the second can explain the results of this study compared to the aforementioned study on the global prevalence of RASs [66]. This discrepancy can also be attributed to the possible inclusion of HCV sequences collected from DAA-experienced individuals who are more likely to harbor drug-resistant variants [47,68,69,70].
Further evidence of increasing RASs prevalence is provided by a recent study from the SHARED consortium, led by Howe et al.[68] This multinational effort involved collecting HCV sequence-linked metadata from 3,355 patients across 22 countries and reported that post-DAA exposure, the frequency of RASs surged from natural baseline levels to 60% in NS3, 80% in NS5A, and 37% in NS5B for specific DAAs, supporting the trends observed in our study. This incremental increase in RASs emphasizes the critical need for ongoing surveillance and novel management strategies to mitigate the impact of RASs on HCV treatment efficacy.
On the other hand, the role of these RASs in DAA treatment effectiveness remains to be fully elucidated and might be a contributory factor to consider in case of treatment failure depending on the drug regimen [30,43,71,72,73,74]. For example, a systematic review and meta-analysis by Singh et al. reported that baseline RASs, decrease the odds of achieving SVR12 in patients with genotype 3 infection [44]. To the contrary, Sarrazin et al. reported the lack of evidence to suggest that known RASs can affect the efficacy of rescue treatments involving multi-target therapies involving sofosbuvir, velpatasvir, and voxilaprevir [43,75].
In this study, the observed disparity in the prevalence of RASs across the NS5A, NS3, and NS5B regions, with NS5A RASs being notably higher, aligns with the biological characteristics of these variants. NS5A is a regulatory protein integral to the HCV replication cycle assisting in the formation of the replication complex and enhances the replication machinery activity through its interactions with both viral and host proteins [76]. This multifunctional role underlines the persistence of NS5A resistant variants, which demonstrate significant replicative fitness, thus allowing them to prevail longer than those in NS3 and NS5B [77,78,79,80]. This persistence is further complicated by the fact that NS5A inhibitors, despite their potent antiviral effects, inherently possess a low barrier to resistance [38,81]. This concept is supported by findings from Nitta et al., who documented that specific mutations, namely Y93H and R30Q/A92K in NS5A, were associated with enhanced viral propagation [82,83].
An important part of this study was the evaluation of the prevalence of cr-RASs for four major DAA regimens—Harvoni, Zepatier, Epclusa, and Mavyret—as defined by the AASLD and the IDSA [26,59]. In contrast to RASs which are mutations within HCV that has the potential to reduce the effectiveness of DAAs, these cr-RASs are the result of specific mutations that have been demonstrated through clinical studies to significantly affect the efficacy of DAAs, leading to reduced treatment response rates or treatment failures.
Out of 1,533 subtype 1a sequences analyzed, 502 showed cr-RASs that affect the efficacy of Harvoni, translating into a prevalence rate of 32.8%. In contrast, only a small fraction (2.9%) of the 617 subtype 1b sequences displayed resistance to both Harvoni and Zepatier. The lower level of resistance in subtype 1b compared to subtype 1a can be explained by previous evidence showing that subtype 1b strains have a higher barrier of resistance compared to subtype 1a strains, with subsequent higher rates of treatment failure [84,85].
In clinical trials, the post-treatment resistance of HCV to ledipasvir/sofosbuvir (Harvoni) has been primarily evaluated in patients infected by subtypes 1a or 1b. For example, Wyles et al. showed that virological failure was reported infrequently with Harvoni, occurring in 51 out of 2144 patients (2.4%), and among those who experienced treatment failure, 74.5% harbored detectable mutations associated with resistance to the NS5A inhibitor ledipasvir [69]. Common RASs observed included Q30R/H and/or Y93H/N in subtype 1a and Y93H in subtype 1b, with 35.3% of patients presenting two or more RASs leading to a significant reduction in ledipasvir susceptibility [69]. Additionally, in a phase 2 study, only one patient with pre-existing ledipasvir resistance developed the sofosbuvir RAS S282T at the point of treatment failure [69]. In this study, the cr-RASs Q30R/H and Y93C/H/N in subtype 1a were reported at rates of 27.8% and 1.9% respectively, while the cr-RAS Y93H in subtype 1b was observed at a rate of 2.8% of the analyzed sequences indicating its frequent detection and possible implication on treatment failure associated with Harvoni. On the other hand, the RAS S282T was totally absent in subtypes 1a and 1b sequences analyzed in this study indicating its infrequent occurrence. In earlier study, baseline RASs in NS5A and NS5B were reported at rate of 16% and 2.5%, with minimal impact on patient responses to ledipasvir/sofosbuvir therapy [86]. Additionally, in the aforementioned clinical trial, the impact of NS5A RASs when affecting the outcomes, can be overcome by extending the treatment duration to 24 weeks or intensifying therapy [86,87].
Despite being classified as cr-RAS, a Japanese clinical trial found that the Y93H in subtype 1b occurred at a baseline frequency of 17.9% [88]. The study concluded that the presence of baseline NS5A RASs did not affect treatment outcomes in subtype 1b Japanese patients treated with ledipasvir/sofosbuvir (Harvoni) for 12 weeks [88]. To the contrary, another Japanese study which involved 493 patients with subtype 1b infection showed the association between virologic failure and NS5A and NS5B cr-RASs [89]. This suggests the necessity for further research to draw reliable conclusions about the real-world impact of cr-RASs.
In real-world studies, the significant impact of cr-RAS was demonstrated in several studies as follows. In a cohort of Mongolian patients, Shih-Jer Hsu et al. showed that a higher proportion of the cr-RAS Y93H was the only independent factor associated with treatment failure in subtype 1b infection [90]. A Spanish study analyzed samples from five patients with subtype 1b or genotype 3 infections using deep sequencing before and after failing sofosbuvir/velpatasvir/voxilaprevir therapy [91]. Post-treatment, four patients exhibited the NS5A cr-RAS, Y93H, indicating the selection of HCV RASs that may complicate effective salvage therapies, hinting to the necessity of RAS testing in such a scenario [91].
From a wider perspective, and considering the indeterminate nature of RASs’ impact on DAA therapeutic efficacy, there are valid arguments to show why RASs need to be considered in HCV research and practice. The increasing prevalence of RASs in HCV as observed in this study can represent a significant concern since those designated as cr-RASs can lead to treatment failure [29,67,92,93]. Additionally, variants harboring RASs can persist especially in high-risk populations with lower adherence to therapeutic regimens (e.g., among IDUs) [94,95,96,97]. Consequently, there would be an increased probability of transmission of RASs with subsequent negative impact on DAA-naïve individuals [98]. Moreover, the emergence and spread of RASs can limit the effectiveness of existing drugs, which is particularly concerning for patients who have limited treatment options due to previous DAA failures or those with advanced liver disease [42,99,100]. Furthermore, the treatment failure negatively impacts patient health and results in increased healthcare costs. This is related to the fact that managing DAA failure involves more complex and prolonged treatment strategies, additional monitoring, and potentially second-line therapies, all of which contribute to higher healthcare expenditures [101]. This is of particular concern in low- and middle-income settings considering the high cost of DAAs even for generic drugs [102,103,104]. Thus, the emergence and spread of RASs could hinder the global efforts for hepatitis C elimination as a public health threat by complicating treatment outcomes and increasing the burden of chronic HCV infections [105].
One important observation of this study was the higher prevalence of RASs in Asia and Africa. This can be attributed to several factors as follows. In many parts of Asia and Africa, access to the latest DAAs can be limited, and when available are often expensive. This leads to inconsistent treatment adherence or the use of suboptimal treatment regimens, which can result in the emergence and persistence of RASs [75]. Additionally, older and less effective DAAs that require longer treatment durations may still be in use in some regions due to economic constraints. These therapies often have a higher risk of developing resistance compared to newer DAAs [68,106]. Furthermore, some regions in Asia and Africa have a higher prevalence of HCV genotypes/subtypes that are naturally more prone to develop certain types of RASs, influencing treatment outcomes [107]. Importantly, the systematic approach for surveillance and monitoring of HCV genetic variants in some regions of Asia and Africa is expected to be less developed. Subsequently, this may delay the detection of RASs and deciphering its possible role in treatment failure, allowing for RASs spread within the population. The lower prevalence of surveillance of RASs in these regions can be inferred by the lower number of available HCV sequences in GenBank which was illustrated in this study. Lower levels of healthcare quality and the availability of diagnostics for HCV resistance testing can lead to less optimal management of treated individuals, facilitating the persistence and spread of resistant strains.
Finally, is it important to highlight several inevitable caveats that should be taken into consideration upon interpreting the study results as follows. First, this study relied on HCV sequences available from the GenBank and Los Alamos HCV Database and this might have introduced biases related to the geographic and temporal representation of samples. These databases may not comprehensively reflect the current global diversity of HCV due to underrepresentation of certain regions or HCV genotype/subtypes, which was reflected by the dominance of samples from North America and Western Europe. Second, we opted to include near-complete genomes aiming to enhance the accuracy of genotype/subtype assignments; nevertheless, this approach may have resulted in inadvertent exclusion of potentially informative partial HCV sequences. Third, the exclusion of sequences without complete metadata, particularly regarding the timing of collection limited the ability to conduct a more comprehensive analysis by reducing the overall sample size of sequences available for analysis. Fourth, it is important to highlight that the reliance on sequences available from GenBank via different submitting laboratories over a long period of time with varying sequencing technologies and bioinformatics tools carries the inherent limitation of varying sequence quality particularly in relation to the detection of minor variants. Finally, it is important to stress out that the results obtained from the HCV dataset in this study may not be generalizable to all clinical scenarios, particularly in cases where novel or rare subtypes as well as recombinant forms are involved. The clinical relevance of the identified RASs needs to be further validated in prospective clinical studies to ensure their applicability in diverse real-world settings.
5. Conclusions
In this study, comprehensive analysis of 3,443 HCV sequences collected from 1983 to 2022 demonstrated significant increase in the prevalence of RASs and cr-RASs across different HCV genotypes/subtypes and regions. A notably high prevalence of RASs across the three analyzed genomic regions was observed at 50.4%, 60.2%, and 25.3% in NS3, NS5A, and NS5B, respectively. The increasing trend in RASs across genomic regions, genotypes/subtypes and geographic regions, could raise concerns about the ability of these RASs to compromise the efficacy of current therapeutic DAA regimens. This is of particular importance for the increasing prevalence of cr-RASs across four major DAA regimens.
Given these findings, it is crucial to maintain ongoing surveillance of RASs prevalence, and develop new antivirals including novel DAAs to overcome potential resistance. This approach is essential to preserve the efficacy of current DAAs, reduce HCV transmission, and ultimately achieving the goal of HCV elimination by 2030.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: The full list of GenBank accession numbers for the HCV sequences analyzed in this study.
Author Contributions
Conceptualization, M.S.; methodology, all authors; validation, M.S.; formal analysis, M.S.; investigation, all authors; data curation, all authors; visualization: M.S.; writing—original draft preparation, M.S.; writing—review and editing, all authors; visualization, M.S.; supervision, M.S.; project administration, M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was approved by the Institutional Review Board, Faculty of Pharmacy, Applied Science Private University (Approval Number 2024-PHA-19). Consent to participate is not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available in the supplementary material section of this article.
Acknowledgments
We sincerely thank all the originating and submitting laboratories who have sequenced and shared the genomic data for HCV in GenBank and the Los Alamos HCV database.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- World Health Organization (WHO). Hepatitis C - Key Facts. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-c (accessed on 19 April 2024).
- Polaris Observatory HCV Collaborators. Global change in hepatitis C virus prevalence and cascade of care between 2015 and 2020: a modelling study. Lancet Gastroenterol Hepatol 2022, 7, 396–415. [Google Scholar] [CrossRef] [PubMed]
- Sallam, M.; Khalil, R. Contemporary Insights into Hepatitis C Virus: A Comprehensive Review. Microorganisms 2024, 12, 1035. [Google Scholar] [CrossRef] [PubMed]
- GBD 2017 Cirrhosis Collaborators. The global, regional, and national burden of cirrhosis by cause in 195 countries and territories, 1990-2017: a systematic analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol Hepatol 2020, 5, 245–266. [Google Scholar] [CrossRef] [PubMed]
- Moon, A.M.; Singal, A.G.; Tapper, E.B. Contemporary Epidemiology of Chronic Liver Disease and Cirrhosis. Clin Gastroenterol Hepatol 2020, 18, 2650–2666. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.Q.; Terrault, N.A.; Tacke, F.; Gluud, L.L.; Arrese, M.; Bugianesi, E.; Loomba, R. Global epidemiology of cirrhosis - aetiology, trends and predictions. Nat Rev Gastroenterol Hepatol 2023, 20, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Kim, B.K.; Yon, D.K.; Lee, S.W.; Lee, H.G.; Chang, H.H.; Park, S.; Koyanagi, A.; Jacob, L.; Dragioti, E.; et al. Global burden of primary liver cancer and its association with underlying aetiologies, sociodemographic status, and sex differences from 1990-2019: A DALY-based analysis of the Global Burden of Disease 2019 study. Clin Mol Hepatol 2023, 29, 433–452. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.J.; Ghany, M.G. Current and future therapies for hepatitis C virus infection. N Engl J Med 2013, 368, 1907–1917. [Google Scholar] [CrossRef] [PubMed]
- Geddawy, A.; Ibrahim, Y.F.; Elbahie, N.M.; Ibrahim, M.A. Direct Acting Anti-hepatitis C Virus Drugs: Clinical Pharmacology and Future Direction. J Transl Int Med 2017, 5, 8–17. [Google Scholar] [CrossRef]
- Calleja, J.L.; Crespo, J.; Rincón, D.; Ruiz-Antorán, B.; Fernandez, I.; Perelló, C.; Gea, F.; Lens, S.; García-Samaniego, J.; Sacristán, B.; et al. Effectiveness, safety and clinical outcomes of direct-acting antiviral therapy in HCV genotype 1 infection: Results from a Spanish real-world cohort. J Hepatol 2017, 66, 1138–1148. [Google Scholar] [CrossRef] [PubMed]
- Crespo, J.; Calleja, J.L.; Fernández, I.; Sacristan, B.; Ruiz-Antorán, B.; Ampuero, J.; Hernández-Conde, M.; García-Samaniego, J.; Gea, F.; Buti, M.; et al. Real-World Effectiveness and Safety of Oral Combination Antiviral Therapy for Hepatitis C Virus Genotype 4 Infection. Clin Gastroenterol Hepatol 2017, 15, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.; Patel, K.; Naggie, S. Genotype 3 Infection: The Last Stand of Hepatitis C Virus. Drugs 2017, 77, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Goossens, N.; Negro, F. Is genotype 3 of the hepatitis C virus the new villain? Hepatology 2014, 59, 2403–2412. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Ji, F.; Yeo, Y.H.; Ogawa, E.; Stave, C.D.; Dang, S.; Li, Z.; Furusyo, N.; Cheung, R.C.; Nguyen, M.H. Systematic review and meta-analysis: real-world effectiveness of direct-acting antiviral therapies in chronic hepatitis C genotype 3 in Asia. BMJ Open Gastroenterol 2018, 5, e000209. [Google Scholar] [CrossRef]
- Isfordink, C.J.; van de Laar, T.J.W.; Rebers, S.P.H.; Wessels, E.; Molenkamp, R.; Knoester, M.; Baak, B.C.; van Nieuwkoop, C.; van Hoek, B.; Brakenhoff, S.M.; et al. Direct-Acting Antiviral Treatment for Hepatitis C Genotypes Uncommon in High-Income Countries: A Dutch Nationwide Cohort Study. Open Forum Infect Dis 2021, 8, ofab006. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.V.; Hussaini, T.; Yoshida, E.M. Concordance of sustained virologic response at weeks 4, 12 and 24 post-treatment of hepatitis c in the era of new oral direct-acting antivirals: A concise review. Ann Hepatol 2016, 15, 154–159. [Google Scholar] [PubMed]
- Chen, J.; Florian, J.; Carter, W.; Fleischer, R.D.; Hammerstrom, T.S.; Jadhav, P.R.; Zeng, W.; Murray, J.; Birnkrant, D. Earlier sustained virologic response end points for regulatory approval and dose selection of hepatitis C therapies. Gastroenterology 2013, 144, 1450–1455. [Google Scholar] [CrossRef] [PubMed]
- Fraser, H.; Martin, N.K.; Brummer-Korvenkontio, H.; Carrieri, P.; Dalgard, O.; Dillon, J.; Goldberg, D.; Hutchinson, S.; Jauffret-Roustide, M.; Kåberg, M.; et al. Model projections on the impact of HCV treatment in the prevention of HCV transmission among people who inject drugs in Europe. J Hepatol 2018, 68, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, H.H.; Mahmud, S.; Chemaitelly, H.; Abu-Raddad, L.J. Treatment as prevention for hepatitis C virus in the Middle East and North Africa: a modeling study. Front Public Health 2023, 11, 1187786. [Google Scholar] [CrossRef] [PubMed]
- van Santen, D.K.; Sacks-Davis, R.; Stewart, A.; Boyd, A.; Young, J.; van der Valk, M.; Smit, C.; Rauch, A.; Braun, D.L.; Jarrin, I.; et al. Treatment as prevention effect of direct-acting antivirals on primary hepatitis C virus incidence: Findings from a multinational cohort between 2010 and 2019. EClinicalMedicine 2023, 56, 101810. [Google Scholar] [CrossRef] [PubMed]
- Hellard, M.; Schroeder, S.E.; Pedrana, A.; Doyle, J.; Aitken, C. The Elimination of Hepatitis C as a Public Health Threat. Cold Spring Harb Perspect Med 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.L. Challenges and Promise of a Hepatitis C Virus Vaccine. Cold Spring Harb Perspect Med 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Duncan, J.D.; Urbanowicz, R.A.; Tarr, A.W.; Ball, J.K. Hepatitis C Virus Vaccine: Challenges and Prospects. Vaccines (Basel) 2020, 8, 90. [Google Scholar] [CrossRef] [PubMed]
- Izhari, M.A. Molecular Mechanisms of Resistance to Direct-Acting Antiviral (DAA) Drugs for the Treatment of Hepatitis C Virus Infections. Diagnostics (Basel) 2023, 13, 3102. [Google Scholar] [CrossRef] [PubMed]
- European Association for the Study of the Liver. EASL recommendations on treatment of hepatitis C: Final update of the series(☆). J Hepatol 2020, 73, 1170–1218. [Google Scholar] [CrossRef] [PubMed]
- AASLD-IDSA. Recommendations for testing, managing, and treating hepatitis C. Available online: https://www.hcvguidelines.org/evaluate/resistance (accessed on 3 May 2024).
- Bhattacharya, D.; Aronsohn, A.; Price, J.; Lo Re, V. Hepatitis C Guidance 2023 Update: AASLD-IDSA Recommendations for Testing, Managing, and Treating Hepatitis C Virus Infection. Clin Infect Dis 2023. [Google Scholar] [CrossRef] [PubMed]
- Bagaglio, S.; Uberti-Foppa, C.; Morsica, G. Resistance Mechanisms in Hepatitis C Virus: implications for Direct-Acting Antiviral Use. Drugs 2017, 77, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, S.; Hashmi, A.H.; Khan, A.; Asad Raza Kazmi, S.M.; Manzoor, S. Emergence and Persistence of Resistance-Associated Substitutions in HCV GT3 Patients Failing Direct-Acting Antivirals. Front Pharmacol 2022, 13, 894460. [Google Scholar] [CrossRef] [PubMed]
- Malandris, K.; Kalopitas, G.; Theocharidou, E.; Germanidis, G. The Role of RASs /RVs in the Current Management of HCV. Viruses 2021, 13, 2096. [Google Scholar] [CrossRef] [PubMed]
- Eltahla, A.A.; Leung, P.; Pirozyan, M.R.; Rodrigo, C.; Grebely, J.; Applegate, T.; Maher, L.; Luciani, F.; Lloyd, A.R.; Bull, R.A. Dynamic evolution of hepatitis C virus resistance-associated substitutions in the absence of antiviral treatment. Sci Rep 2017, 7, 41719. [Google Scholar] [CrossRef] [PubMed]
- Preciado, M.V.; Valva, P.; Escobar-Gutierrez, A.; Rahal, P.; Ruiz-Tovar, K.; Yamasaki, L.; Vazquez-Chacon, C.; Martinez-Guarneros, A.; Carpio-Pedroza, J.C.; Fonseca-Coronado, S.; et al. Hepatitis C virus molecular evolution: transmission, disease progression and antiviral therapy. World J Gastroenterol 2014, 20, 15992–16013. [Google Scholar] [CrossRef] [PubMed]
- Fourati, S.; Pawlotsky, J.M. Virologic Tools for HCV Drug Resistance Testing. Viruses 2015, 7, 6346–6359. [Google Scholar] [CrossRef] [PubMed]
- Soria, M.E.; García-Crespo, C.; Martínez-González, B.; Vázquez-Sirvent, L.; Lobo-Vega, R.; de Ávila, A.I.; Gallego, I.; Chen, Q.; García-Cehic, D.; Llorens-Revull, M.; et al. Amino Acid Substitutions Associated with Treatment Failure for Hepatitis C Virus Infection. J Clin Microbiol 2020, 58. [Google Scholar] [CrossRef] [PubMed]
- Kalaghatgi, P.; Sikorski, A.M.; Knops, E.; Rupp, D.; Sierra, S.; Heger, E.; Neumann-Fraune, M.; Beggel, B.; Walker, A.; Timm, J.; et al. Geno2pheno[HCV] - A Web-based Interpretation System to Support Hepatitis C Treatment Decisions in the Era of Direct-Acting Antiviral Agents. PLoS One 2016, 11, e0155869. [Google Scholar] [CrossRef] [PubMed]
- Lontok, E.; Harrington, P.; Howe, A.; Kieffer, T.; Lennerstrand, J.; Lenz, O.; McPhee, F.; Mo, H.; Parkin, N.; Pilot-Matias, T.; et al. Hepatitis C virus drug resistance-associated substitutions: State of the art summary. Hepatology 2015, 62, 1623–1632. [Google Scholar] [CrossRef] [PubMed]
- Feld, J.J. Resistance testing: Interpretation and incorporation into HCV treatment algorithms. Clin Liver Dis (Hoboken) 2017, 9, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Kliemann, D.A.; Tovo, C.V.; Gorini da Veiga, A.B.; Machado, A.L.; West, J. Genetic Barrier to Direct Acting Antivirals in HCV Sequences Deposited in the European Databank. PLoS One 2016, 11, e0159924. [Google Scholar] [CrossRef] [PubMed]
- Cento, V.; Mirabelli, C.; Salpini, R.; Dimonte, S.; Artese, A.; Costa, G.; Mercurio, F.; Svicher, V.; Parrotta, L.; Bertoli, A.; et al. HCV genotypes are differently prone to the development of resistance to linear and macrocyclic protease inhibitors. PLoS One 2012, 7, e39652. [Google Scholar] [CrossRef]
- Vidal, L.L.; Soares, M.A.; Santos, A.F. NS3 protease polymorphisms and genetic barrier to drug resistance of distinct hepatitis C virus genotypes from worldwide treatment-naïve subjects. J Viral Hepat 2016, 23, 840–849. [Google Scholar] [CrossRef]
- de Salazar, A.; Dietz, J.; di Maio, V.C.; Vermehren, J.; Paolucci, S.; Müllhaupt, B.; Coppola, N.; Cabezas, J.; Stauber, R.E.; Puoti, M.; et al. Prevalence of resistance-associated substitutions and retreatment of patients failing a glecaprevir/pibrentasvir regimen. J Antimicrob Chemother 2020, 75, 3349–3358. [Google Scholar] [CrossRef] [PubMed]
- Pisaturo, M.; Starace, M.; Minichini, C.; De Pascalis, S.; Occhiello, L.; Fraia, A.D.; Messina, V.; Sangiovanni, V.; Claar, E.; Coppola, N. Virological patterns of hepatitis C virus patients with failure to the current-generation direct-acting antivirals. Int J Antimicrob Agents 2020, 56, 106067. [Google Scholar] [CrossRef]
- Sarrazin, C.; Cooper, C.L.; Manns, M.P.; Reddy, K.R.; Kowdley, K.V.; Roberts, S.K.; Dvory-Sobol, H.; Svarovskia, E.; Martin, R.; Camus, G.; et al. No impact of resistance-associated substitutions on the efficacy of sofosbuvir, velpatasvir, and voxilaprevir for 12 weeks in HCV DAA-experienced patients. J Hepatol 2018, 69, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.D.; Maitra, S.; Singh, N.; Tyagi, P.; Ashraf, A.; Kumar, R.; Shalimar. Systematic review with meta-analysis: impact of baseline resistance-associated substitutions on the efficacy of glecaprevir/pibrentasvir among chronic hepatitis C patients. Aliment Pharmacol Ther 2020, 51, 490–504. [Google Scholar] [CrossRef] [PubMed]
- Sayan, M.; Yıldırım, F.S.; Akhan, S.; Yıldırım, A.A.; Şirin, G.; Cabalak, M.; Demir, M.; Can, S.; Ersöz, G.; Altıntaş, E.; et al. NS5A resistance - associated substitutions in chronic hepatitis C patients with direct acting antiviral treatment failure in Turkey. Int J Infect Dis 2020, 95, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Ridruejo, E.; Pereson, M.J.; Flichman, D.M.; Di Lello, F.A. Hepatitis C virus treatment failure: Clinical utility for testing resistance-associated substitutions. World J Hepatol 2021, 13, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Wyles, D.L.; Luetkemeyer, A.F. Understanding Hepatitis C Virus Drug Resistance: Clinical Implications for Current and Future Regimens. Top Antivir Med 2017, 25, 103–109. [Google Scholar] [PubMed]
- Onorato, L.; Pisaturo, M.; Starace, M.; Minichini, C.; Di Fraia, A.; Astorri, R.; Coppola, N. Virological Factors Associated with Failure to the Latest Generation of Direct Acting Agents (DAA) and Re-Treatment Strategy: A Narrative Review. Viruses 2021, 13, 432. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, C.; Singh, M.; Das, D.; Chaudhuri, S.; Mukhopadhyay, A. Current therapeutics against HCV. Virusdisease 2021, 32, 228–243. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M. Era of direct acting anti-viral agents for the treatment of hepatitis C. World J Hepatol 2018, 10, 670–684. [Google Scholar] [CrossRef]
- Sayers, E.W.; Cavanaugh, M.; Clark, K.; Ostell, J.; Pruitt, K.D.; Karsch-Mizrachi, I. GenBank. Nucleic Acids Res 2020, 48, D84–d86. [Google Scholar] [CrossRef]
- Kuiken, C.; Hraber, P.; Thurmond, J.; Yusim, K. The hepatitis C sequence database in Los Alamos. Nucleic Acids Res 2008, 36, D512–516. [Google Scholar] [CrossRef]
- Yusim, K.; Richardson, R.; Tao, N.; Dalwani, A.; Agrawal, A.; Szinger, J.; Funkhouser, R.; Korber, B.; Kuiken, C. Los alamos hepatitis C immunology database. Appl Bioinformatics 2005, 4, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Villesen, P. FaBox: an online toolbox for fasta sequences. Molecular Ecology Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Struck, D.; Lawyer, G.; Ternes, A.M.; Schmit, J.C.; Bercoff, D.P. COMET: adaptive context-based modeling for ultrafast HIV-1 subtype identification. Nucleic Acids Res 2014, 42, e144. [Google Scholar] [CrossRef] [PubMed]
- AASLD-IDSA HCV Guidance Panel. Hepatitis C Guidance 2018 Update: AASLD-IDSA Recommendations for Testing, Managing, and Treating Hepatitis C Virus Infection. Clin Infect Dis 2018, 67, 1477–1492. [Google Scholar] [CrossRef] [PubMed]
- Svarovskaia, E.S.; Dvory-Sobol, H.; Parkin, N.; Hebner, C.; Gontcharova, V.; Martin, R.; Ouyang, W.; Han, B.; Xu, S.; Ku, K.; et al. Infrequent development of resistance in genotype 1-6 hepatitis C virus-infected subjects treated with sofosbuvir in phase 2 and 3 clinical trials. Clin Infect Dis 2014, 59, 1666–1674. [Google Scholar] [CrossRef] [PubMed]
- Eltahla, A.A.; Leung, P.; Pirozyan, M.R.; Rodrigo, C.; Grebely, J.; Applegate, T.; Maher, L.; Luciani, F.; Lloyd, A.R.; Bull, R.A. Dynamic evolution of hepatitis C virus resistance-associated substitutions in the absence of antiviral treatment. Scientific Reports 2017, 7, 41719. [Google Scholar] [CrossRef] [PubMed]
- Bertoli, A.; Sorbo, M.C.; Aragri, M.; Lenci, I.; Teti, E.; Polilli, E.; Di Maio, V.C.; Gianserra, L.; Biliotti, E.; Masetti, C.; et al. Prevalence of Single and Multiple Natural NS3, NS5A and NS5B Resistance-Associated Substitutions in Hepatitis C Virus Genotypes 1-4 in Italy. Sci Rep 2018, 8, 8988. [Google Scholar] [CrossRef] [PubMed]
- Paolucci, S.; Fiorina, L.; Mariani, B.; Gulminetti, R.; Novati, S.; Barbarini, G.; Bruno, R.; Baldanti, F. Naturally occurring resistance mutations to inhibitors of HCV NS5A region and NS5B polymerase in DAA treatment-naïve patients. Virol J 2013, 10, 355. [Google Scholar] [CrossRef] [PubMed]
- Martínez, A.P.; García, G.; Ridruejo, E.; Culasso, A.C.; Pérez, P.S.; Pereson, M.J.; Neukam, K.; Flichman, D.; Di Lello, F.A. Hepatitis C virus genotype 1 infection: Prevalence of NS5A and NS5B resistance-associated substitutions in naïve patients from Argentina. J Med Virol 2019, 91, 1970–1978. [Google Scholar] [CrossRef] [PubMed]
- Valutite, D.; Ostankova, Y.; Semenov, A.; Lyalina, L.; Totolian, A. Distribution of Primary Resistance Mutations in Saint Petersburg in Patients with Chronic Hepatitis C. Diagnostics (Basel) 2022, 12, 1054. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.W.; Li, H.; Ren, H.; Hu, P. Global prevalence of pre-existing HCV variants resistant to direct-acting antiviral agents (DAAs): mining the GenBank HCV genome data. Sci Rep 2016, 6, 20310. [Google Scholar] [CrossRef] [PubMed]
- Kitrinos, K.M.; Corsa, A.C.; Worth, A.; Hedskog, C.; Brainard, D.M.; Miller, M.D.; Mo, H. Nonstructural protein 5A resistance profile in patients with chronic hepatitis C treated with ledipasvir-containing regimens without sofosbuvir. J Viral Hepat 2018, 25, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Howe, A.Y.M.; Rodrigo, C.; Cunningham, E.B.; Douglas, M.W.; Dietz, J.; Grebely, J.; Popping, S.; Sfalcin, J.A.; Parczewski, M.; Sarrazin, C.; et al. Characteristics of hepatitis C virus resistance in an international cohort after a decade of direct-acting antivirals. JHEP Rep 2022, 4, 100462. [Google Scholar] [CrossRef] [PubMed]
- Wyles, D.; Dvory-Sobol, H.; Svarovskaia, E.S.; Doehle, B.P.; Martin, R.; Afdhal, N.H.; Kowdley, K.V.; Lawitz, E.; Brainard, D.M.; Miller, M.D.; et al. Post-treatment resistance analysis of hepatitis C virus from phase II and III clinical trials of ledipasvir/sofosbuvir. J Hepatol 2017, 66, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Lahser, F.; Galloway, A.; Hwang, P.; Palcza, J.; Brunhofer, J.; Wahl, J.; Robertson, M.; Barr, E.; Black, T.; Asante-Appiah, E.; et al. Interim analysis of a 3-year follow-up study of NS5A and NS3 resistance-associated substitutions after treatment with grazoprevir-containing regimens in participants with chronic HCV infection. Antivir Ther 2018, 23, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Kjellin, M.; Kileng, H.; Akaberi, D.; Palanisamy, N.; Duberg, A.S.; Danielsson, A.; Kristiansen, M.G.; Nöjd, J.; Aleman, S.; Gutteberg, T.; et al. Effect of the baseline Y93H resistance-associated substitution in HCV genotype 3 for direct-acting antiviral treatment: real-life experience from a multicenter study in Sweden and Norway. Scand J Gastroenterol 2019, 54, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Perales, C.; Quer, J.; Gregori, J.; Esteban, J.I.; Domingo, E. Resistance of Hepatitis C Virus to Inhibitors: Complexity and Clinical Implications. Viruses 2015, 7, 5746–5766. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M.; Gupta, E. Emerging resistance to directly-acting antiviral therapy in treatment of chronic Hepatitis C infection-A brief review of literature. J Family Med Prim Care 2020, 9, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Sharafi, H.; Alavian, S.M. Hepatitis C resistance to NS5A inhibitors: Is it going to be a problem? World J Hepatol 2018, 10, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, C. Treatment failure with DAA therapy: Importance of resistance. J Hepatol 2021, 74, 1472–1482. [Google Scholar] [CrossRef]
- Li, H.C.; Yang, C.H.; Lo, S.Y. Hepatitis C Viral Replication Complex. Viruses 2021, 13, 520. [Google Scholar] [CrossRef] [PubMed]
- Scheel, T.K.; Gottwein, J.M.; Mikkelsen, L.S.; Jensen, T.B.; Bukh, J. Recombinant HCV variants with NS5A from genotypes 1-7 have different sensitivities to an NS5A inhibitor but not interferon-α. Gastroenterology 2011, 140, 1032–1042. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, S.; Kanda, T.; Wu, S.; Shirasawa, H.; Yokosuka, O. Hepatitis C virus NS5A inhibitors and drug resistance mutations. World J Gastroenterol 2014, 20, 2902–2912. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Williford, S.E.; McGivern, D.R.; Burch, C.L.; Hu, F.; Benzine, T.; Ingravallo, P.; Asante-Appiah, E.; Howe, A.Y.M.; Swanstrom, R.; et al. Evolutionary pathways to NS5A inhibitor resistance in genotype 1 hepatitis C virus. Antiviral Res 2018, 158, 45–51. [Google Scholar] [CrossRef]
- Wyles, D.; Mangia, A.; Cheng, W.; Shafran, S.; Schwabe, C.; Ouyang, W.; Hedskog, C.; McNally, J.; Brainard, D.M.; Doehle, B.P.; et al. Long-term persistence of HCV NS5A resistance-associated substitutions after treatment with the HCV NS5A inhibitor, ledipasvir, without sofosbuvir. Antivir Ther 2018, 23, 229–238. [Google Scholar] [CrossRef]
- Kyuregyan, K.K.; Kichatova, V.S.; Karlsen, A.A.; Isaeva, O.V.; Solonin, S.A.; Petkov, S.; Nielsen, M.; Isaguliants, M.G.; Mikhailov, M.I. Factors Influencing the Prevalence of Resistance-Associated Substitutions in NS5A Protein in Treatment-Naive Patients with Chronic Hepatitis C. Biomedicines 2020, 8, 80. [Google Scholar] [CrossRef] [PubMed]
- Nitta, S.; Asahina, Y.; Matsuda, M.; Yamada, N.; Sugiyama, R.; Masaki, T.; Suzuki, R.; Kato, N.; Watanabe, M.; Wakita, T.; et al. Effects of Resistance-Associated NS5A Mutations in Hepatitis C Virus on Viral Production and Susceptibility to Antiviral Reagents. Sci Rep 2016, 6, 34652. [Google Scholar] [CrossRef] [PubMed]
- Nitta, S.; Asahina, Y.; Kato, T.; Tsuchiya, J.; Inoue-Shinomiya, E.; Sato, A.; Tsunoda, T.; Miyoshi, M.; Kawai-Kitahata, F.; Murakawa, M.; et al. Impact of novel NS5A resistance-associated substitutions of hepatitis C virus detected in treatment-experienced patients. Sci Rep 2019, 9, 5722. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, C.; Zeuzem, S. Resistance to direct antiviral agents in patients with hepatitis C virus infection. Gastroenterology 2010, 138, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Pellicelli, A.M.; Romano, M.; Stroffolini, T.; Mazzoni, E.; Mecenate, F.; Monarca, R.; Picardi, A.; Bonaventura, M.E.; Mastropietro, C.; Vignally, P.; et al. HCV genotype 1a shows a better virological response to antiviral therapy than HCV genotype 1b. BMC Gastroenterology 2012, 12, 162. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, C.; Dvory-Sobol, H.; Svarovskaia, E.S.; Doehle, B.P.; Pang, P.S.; Chuang, S.M.; Ma, J.; Ding, X.; Afdhal, N.H.; Kowdley, K.V.; et al. Prevalence of Resistance-Associated Substitutions in HCV NS5A, NS5B, or NS3 and Outcomes of Treatment With Ledipasvir and Sofosbuvir. Gastroenterology 2016, 151, 501–512. [Google Scholar] [CrossRef]
- Wang, G.P.; Terrault, N.; Reeves, J.D.; Liu, L.; Li, E.; Zhao, L.; Lim, J.K.; Morelli, G.; Kuo, A.; Levitsky, J.; et al. Prevalence and impact of baseline resistance-associated substitutions on the efficacy of ledipasvir/sofosbuvir or simeprevir/sofosbuvir against GT1 HCV infection. Sci Rep 2018, 8, 3199. [Google Scholar] [CrossRef] [PubMed]
- Mizokami, M.; Dvory-Sobol, H.; Izumi, N.; Nishiguchi, S.; Doehle, B.; Svarovskaia, E.S.; De-Oertel, S.; Knox, S.; Brainard, D.M.; Miller, M.D.; et al. Resistance Analyses of Japanese Hepatitis C-Infected Patients Receiving Sofosbuvir or Ledipasvir/Sofosbuvir Containing Regimens in Phase 3 Studies. J Viral Hepat 2016, 23, 780–788. [Google Scholar] [CrossRef]
- Mawatari, S.; Oda, K.; Tabu, K.; Ijuin, S.; Kumagai, K.; Fujisaki, K.; Hashiguchi, M.; Inada, Y.; Uto, H.; Hiramine, Y.; et al. The co-existence of NS5A and NS5B resistance-associated substitutions is associated with virologic failure in Hepatitis C Virus genotype 1 patients treated with sofosbuvir and ledipasvir. PLoS One 2018, 13, e0198642. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.J.; Enkhzaya, S.; Lin, Y.Y.; Tseng, T.C.; Khosbayar, T.; Tsai, C.H.; Wang, T.S.; Enkhtuya, D.; Ivshinkhorol, D.; Naranzul, N.; et al. Resistance-associated substitution and ledipasvir/sofosbuvir therapy in Mongolian chronic hepatitis C patients. J Formos Med Assoc 2020, 119, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Cehic, D.; Rando, A.; Rodriguez-Frias, F.; Gregori, J.; Costa, J.G.; Carrión, J.A.; Macenlle, R.; Pamplona, J.; Castro-Iglesias, A.; Cañizares, A.; et al. Resistance-associated substitutions after sofosbuvir/velpatasvir/voxilaprevir triple therapy failure. J Viral Hepat 2021, 28, 1319–1324. [Google Scholar] [CrossRef] [PubMed]
- Dietz, J.; Susser, S.; Vermehren, J.; Peiffer, K.H.; Grammatikos, G.; Berger, A.; Ferenci, P.; Buti, M.; Müllhaupt, B.; Hunyady, B.; et al. Patterns of Resistance-Associated Substitutions in Patients With Chronic HCV Infection Following Treatment With Direct-Acting Antivirals. Gastroenterology 2018, 154, 976–988. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Mao, X.; Yu, K.; Suo, C.; Jin, L.; Zhang, T.; Chen, X. Prevalence of HCV resistance-associated substitutions among treatment-failure patients receiving direct-acting antiviral agents. J Viral Hepat 2020, 27, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Dietz, J.; Müllhaupt, B.; Buggisch, P.; Graf, C.; Peiffer, K.H.; Matschenz, K.; Schattenberg, J.M.; Antoni, C.; Mauss, S.; Niederau, C.; et al. Long-term persistence of HCV resistance-associated substitutions after DAA treatment failure. J Hepatol 2023, 78, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Sagnelli, E.; Starace, M.; Minichini, C.; Pisaturo, M.; Macera, M.; Sagnelli, C.; Coppola, N. Resistance detection and re-treatment options in hepatitis C virus-related chronic liver diseases after DAA-treatment failure. Infection 2018, 46, 761–783. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.; Jin, B.; Lee, H.W.; Park, H.J.; Park, J.Y.; Kim, D.Y.; Han, K.H.; Ahn, S.H.; Kim, S. Evolution and persistence of resistance-associated substitutions of hepatitis C virus after direct-acting antiviral treatment failures. J Viral Hepat 2018, 25, 1251–1259. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, M.J.; Lipsey, D.; Ganova-Raeva, L.; Punkova, L.T.; Agyemang, L.; Sue, A.; Ramachandran, S.; Khudyakov, Y.; Litwin, A.H. A Phylogenetic Analysis of Hepatitis C Virus Transmission, Relapse, and Reinfection Among People Who Inject Drugs Receiving Opioid Agonist Therapy. J Infect Dis 2020, 222, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Esposito, I.; Marciano, S.; Haddad, L.; Galdame, O.; Franco, A.; Gadano, A.; Flichman, D.; Trinks, J. Prevalence and Factors Related to Natural Resistance-Associated Substitutions to Direct-Acting Antivirals in Patients with Genotype 1 Hepatitis C Virus Infection. Viruses 2018, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Di Maio, V.C.; Cento, V.; Lenci, I.; Aragri, M.; Rossi, P.; Barbaliscia, S.; Melis, M.; Verucchi, G.; Magni, C.F.; Teti, E.; et al. Multiclass HCV resistance to direct-acting antiviral failure in real-life patients advocates for tailored second-line therapies. Liver Int 2017, 37, 514–528. [Google Scholar] [CrossRef] [PubMed]
- Di Maio, V.C.; Barbaliscia, S.; Teti, E.; Fiorentino, G.; Milana, M.; Paolucci, S.; Pollicino, T.; Morsica, G.; Starace, M.; Bruzzone, B.; et al. Resistance analysis and treatment outcomes in hepatitis C virus genotype 3-infected patients within the Italian network VIRONET-C. Liver Int 2021, 41, 1802–1814. [Google Scholar] [CrossRef] [PubMed]
- Solitano, V.; Plaz Torres, M.C.; Pugliese, N.; Aghemo, A. Management and Treatment of Hepatitis C: Are There Still Unsolved Problems and Unique Populations? Viruses 2021, 13, 1048. [Google Scholar] [CrossRef] [PubMed]
- Graham, C.S.; Swan, T. A path to eradication of hepatitis C in low- and middle-income countries. Antiviral Res 2015, 119, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Ur-Rehman, T.; Ali, M.; Haque, S.; Rasheed, F.; Lougher, E.; Nawaz, M.S.; Paudyal, V. Improving access to the treatment of hepatitis C in low- and middle-income countries: evaluation of a patient assistance programme. International Journal of Clinical Pharmacy 2021, 43, 958–968. [Google Scholar] [CrossRef]
- Zahid, H.; Aslam, K.; Dahl, E.H.; Abbassi, W.; Adan, S.; Van den Bergh, R.; Balinska, M.A.; Luck, N.H. DAA treatment failures in a low-resource setting with a high burden of hepatitis C infections: a case series. Oxf Med Case Reports 2022, 2022, omac049. [Google Scholar] [CrossRef]
- Manns, M.P.; Maasoumy, B. Breakthroughs in hepatitis C research: from discovery to cure. Nature Reviews Gastroenterology & Hepatology 2022, 19, 533–550. [Google Scholar] [CrossRef]
- Jacobson, I.M. The HCV Treatment Revolution Continues: Resistance Considerations, Pangenotypic Efficacy, and Advances in Challenging Populations. Gastroenterol Hepatol (N Y) 2016, 12, 1–11. [Google Scholar] [PubMed]
- Shah, R.; Ahovegbe, L.; Niebel, M.; Shepherd, J.; Thomson, E.C. Non-epidemic HCV genotypes in low- and middle-income countries and the risk of resistance to current direct-acting antiviral regimens. J Hepatol 2021, 75, 462–473. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The overall prevalence of RASs per HCV genotype/subtype for NS3 (A); NS5A (B); and NS5B (C).
Figure 1.
The overall prevalence of RASs per HCV genotype/subtype for NS3 (A); NS5A (B); and NS5B (C).
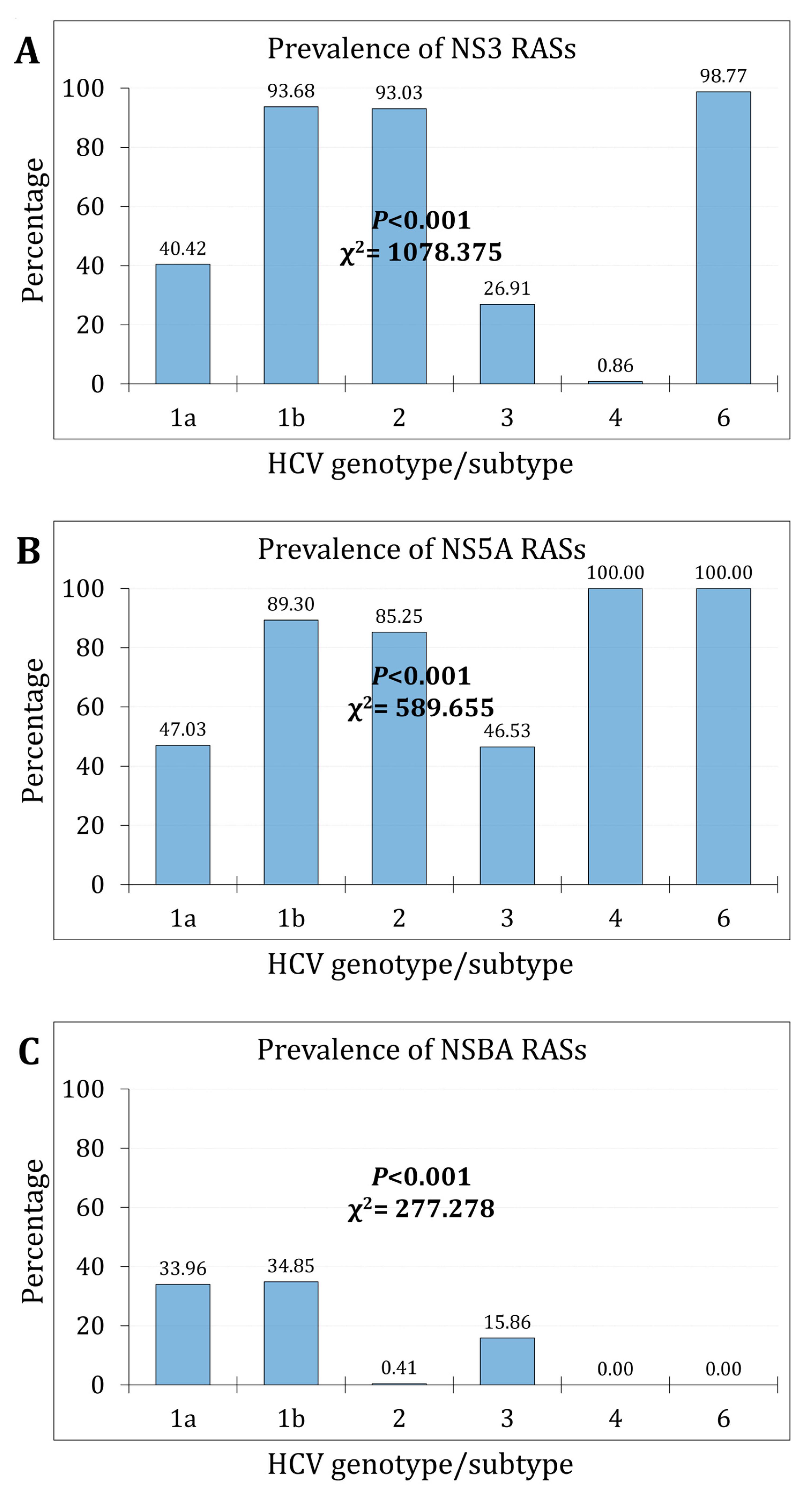
Figure 2.
The RASs trends over the study period per HCV genotype/subtype for NS3 (A); NS5A (B); and NS5B (C).
Figure 2.
The RASs trends over the study period per HCV genotype/subtype for NS3 (A); NS5A (B); and NS5B (C).
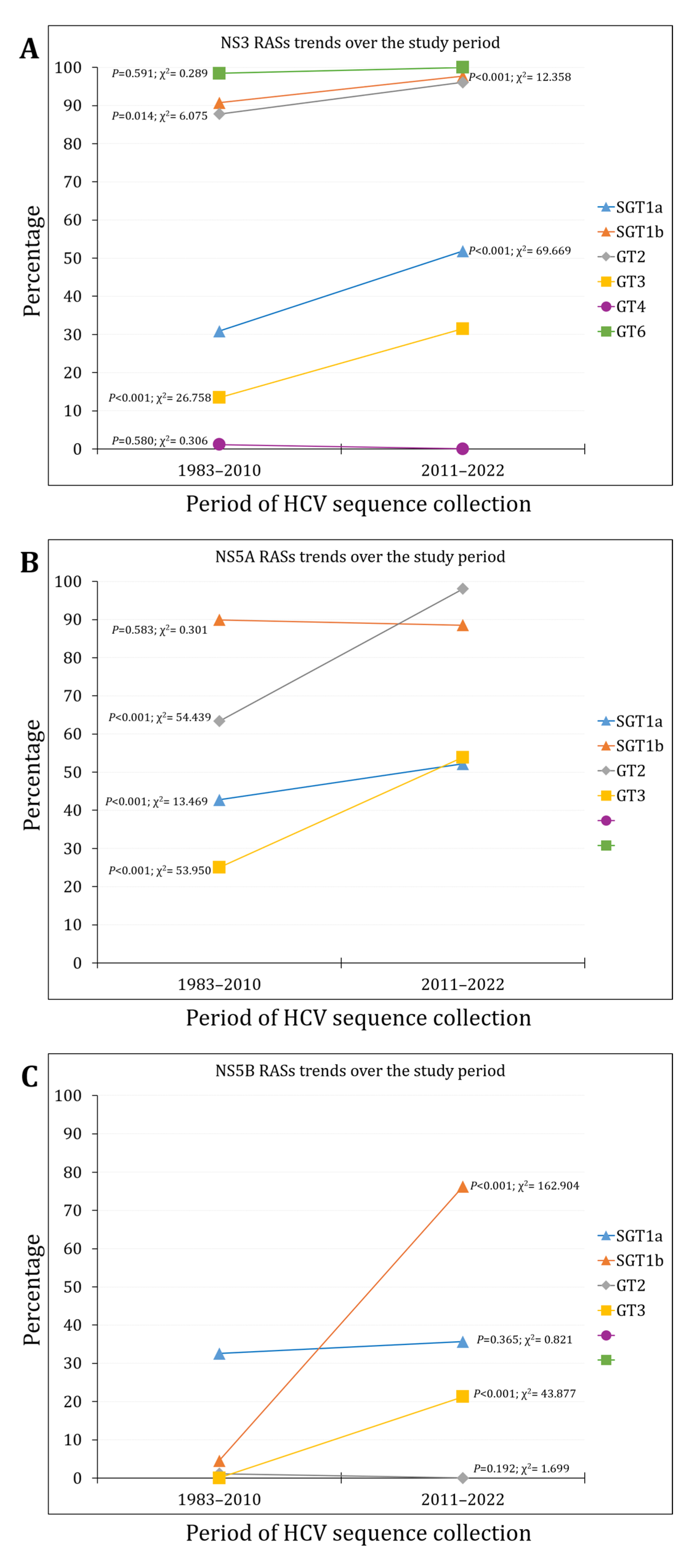
Figure 3.
The RASs prevalence per region for NS3 (A); NS5A (B); and NS5B (C). WE/NA/AUS: Western Europe, North America, or Australia; EU/RUS: Eastern Europe, Russia; MENA: Middle East/North Africa; LA/Car: Latin America, the Caribbean.
Figure 3.
The RASs prevalence per region for NS3 (A); NS5A (B); and NS5B (C). WE/NA/AUS: Western Europe, North America, or Australia; EU/RUS: Eastern Europe, Russia; MENA: Middle East/North Africa; LA/Car: Latin America, the Caribbean.
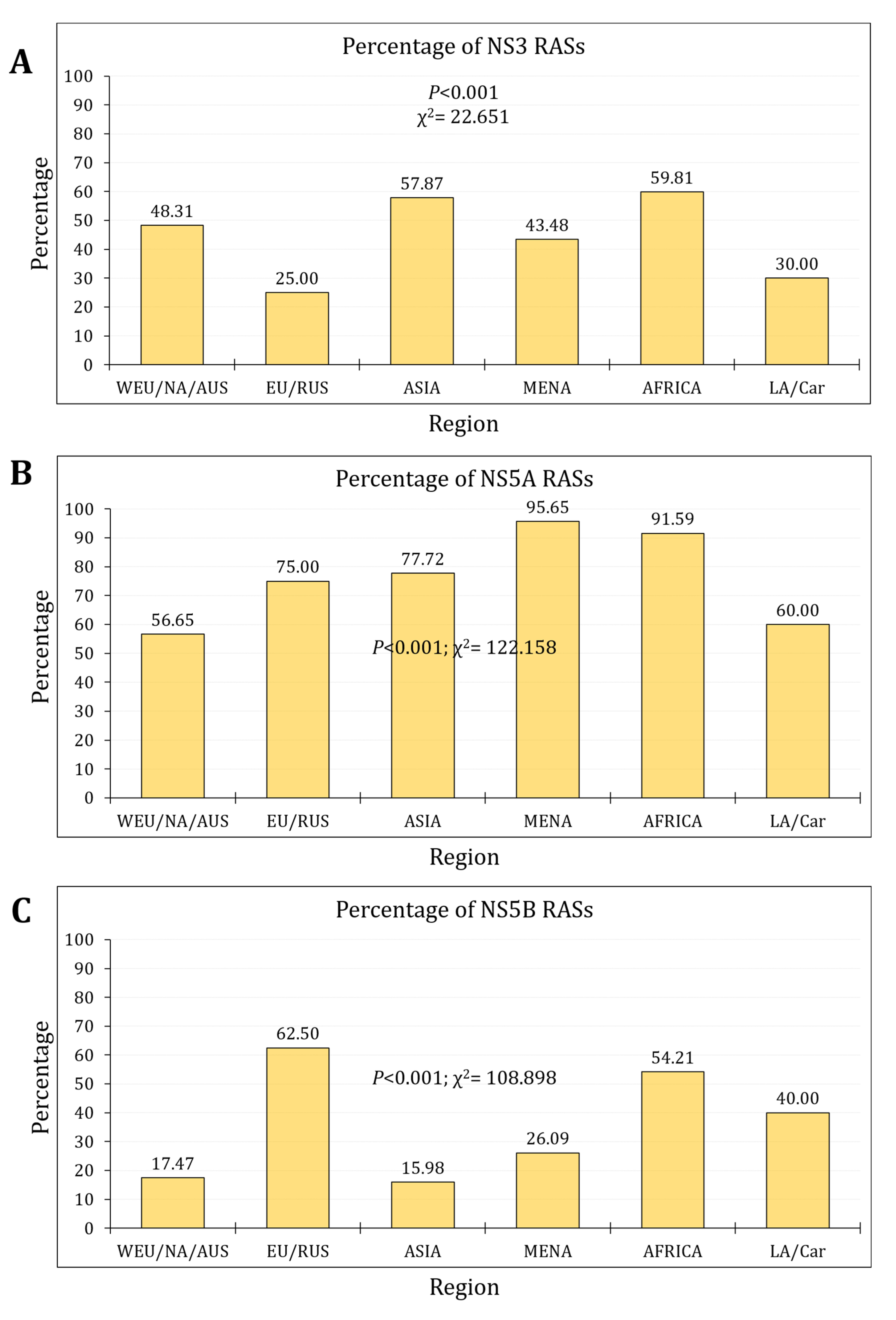
Figure 4.
The temporal trends of clinically-relevant resistance-associated substitutions (cr-RASs) in four direct-acting antiviral (DAA) regimens.
Figure 4.
The temporal trends of clinically-relevant resistance-associated substitutions (cr-RASs) in four direct-acting antiviral (DAA) regimens.
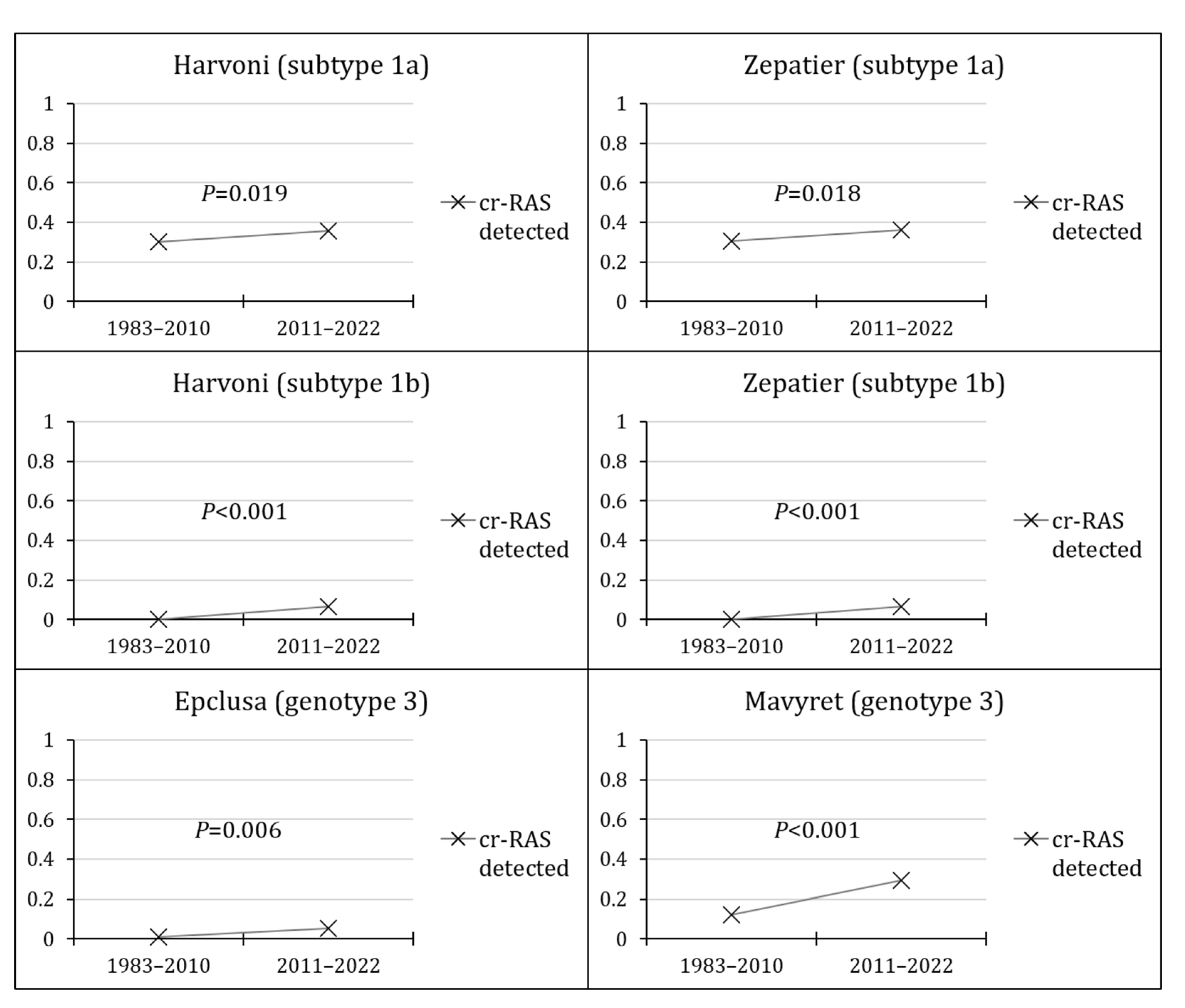
Figure 5.
The prevalence of clinically-relevant resistance-associated substitutions (cr-RASs) in four direct-acting antiviral (DAA) regimens per region.
Figure 5.
The prevalence of clinically-relevant resistance-associated substitutions (cr-RASs) in four direct-acting antiviral (DAA) regimens per region.
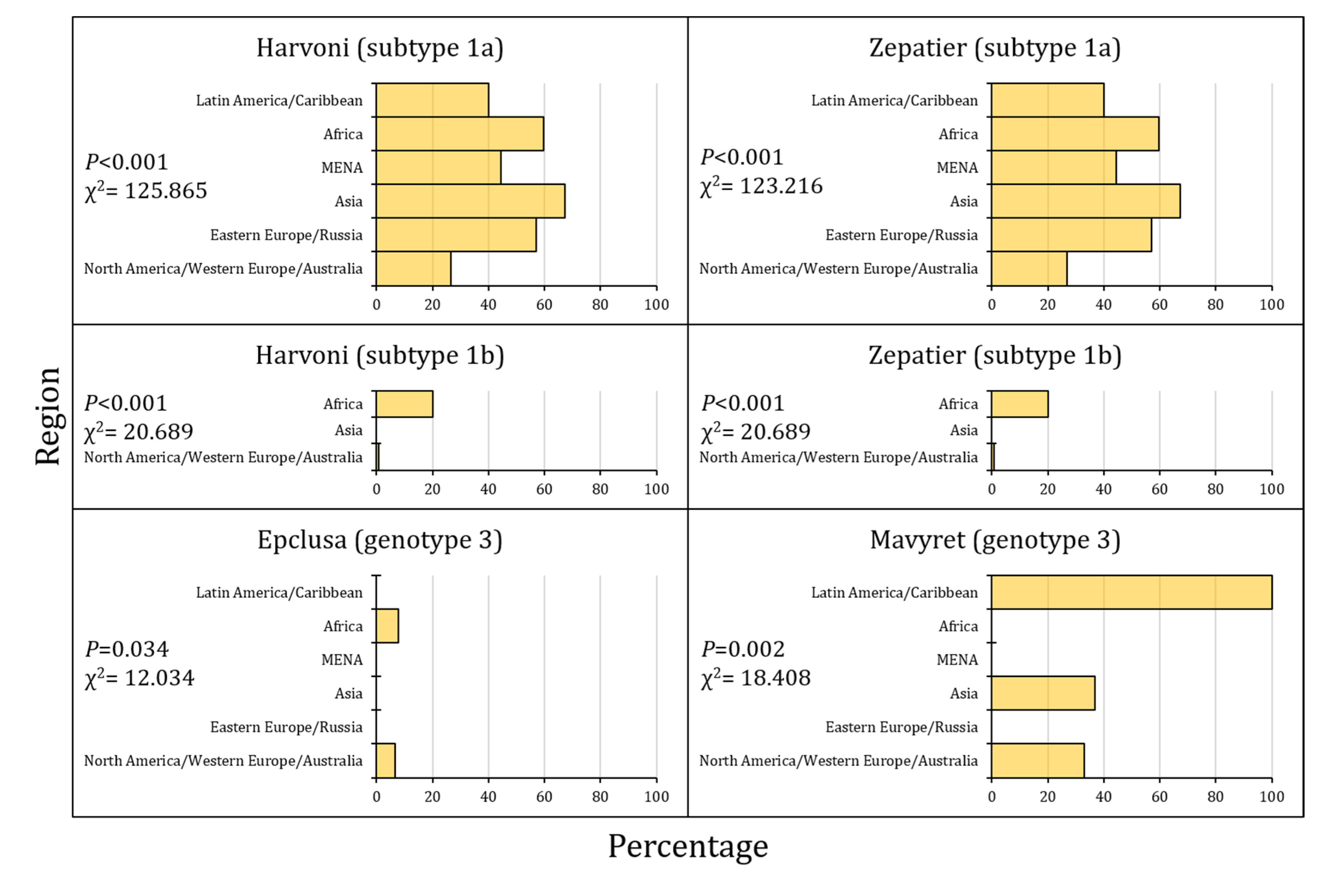

Table 1.
The full list of NS3 RASs detected in the included sequences stratified per HCV genotype/subtype.
Table 1.
The full list of NS3 RASs detected in the included sequences stratified per HCV genotype/subtype.
| HCV genotype/subtype: RASs | N (%) | 1a (n=1534) | 1b (n=617) | 2 (n=244) | 3 (n=851) | 4 (n=116) | 6 (n=81) |
|---|---|---|---|---|---|---|---|
| 1a: V36A/C/F/G/L/M 1b: V36A/C/G/L/M 6: V36I |
330 (14.8) | 121 (7.9) | 209 (33.9) | - | - | - | 0 |
| 1a: Q41R 1b: Q41R 3: Q41K 4: Q41R 6: Q41K/R |
0 | 0 | 0 | - | 0 | 0 | 0 |
| 1a: F43I/L/S/V 1b: F43I/S/V 2: F43V |
0 | 0 | 0 | 0 | - | - | - |
| 1a: T54A/S 1b: T54A/C/G/S |
165 (7.7) | 33 (2.2) | 132 (21.4) | - | - | - | - |
| 1a: V55I 1b: V55A 2: V55A/I |
38 (1.6) | 14 (0.9) | 0 | 24 (9.8) | - | - | - |
| 1a: Y56H 1b: Y56H/L/F 2: Y56H/F 3: Y56H 4: Y56H 6: Y56H |
23 (0.7) | 1 (0.1) | 9 (1.5) | 13 (5.3) | 0 | 0 | 0 |
| 1a: Q80K/L/R 1b: Q80H/K/L/R 3: Q80K/R 4: Q80R 6: L80K/Q |
656 (20.5) | 377 (24.6) | 200 (32.4) | - | 0 | 0 | 79 (97.5) |
| 1a: S122G/N/R 1b: S122A/D/G/I/N/R/T 6: S122T |
216 (9.7) | 162 (10.6) | 50 (8.1) | - | - | - | 4 (4.9) |
| 1a: R155G/I/K/M/Q/S/T/V/W 1b: R155C/G/I/K/L/Q/M/S/T/W 3: R155K 4: R155C/K |
11 (0.4) | 8 (0.5) | 2 (0.3) | - | 1 (0.1) | 0 | - |
| 1a: A156G/P/S/T/V 1b: A156G/P/S/T/V 2: A156L/M/T/V 3: A156G/P/T/V 4: A156G/H/K/L/S/T/V 6: A156T/V |
0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 1a: V158I 1b: V158I |
2 (0.1) | 1 (0.1) | 1 (0.2) | - | - | - | - |
| 3: A166S/T/Y | 228 (26.8) | - | - | - | 228 (26.8) | - | - |
| 1a: D168A/C/E/F/G/H/I/K/L/N/Q/R/T/V/Y 1b: D168A/C/E/F/G/H/I/K/L/N/Q/T/V/Y 2: D168A/E/F/G/H/N/S/T/V/Y 3: Q168H/K/L/R 4: D168A/E/G/H/T/V 6: D168A/E/G/H/V/Y |
421 (12.2) | 43 (2.8) | 186 (30.1) | 190 (77.9) | 1 (0.1) | 1 (0.9) | 0 |
| 1a: I/V170T 1b: I/V170A/L/T 6: I170V |
7 (0.3) | 0 | 0 | - | - | - | 7 (8.6) |
| M175L | 551 (89.3) | - | 551 (89.3) | - | - | - | - |
| Sequences with any NS3 RASs | 1735 (50.4) | 620 (40.4) | 578 (93.7) | 227 (93.0) | 229 (26.9) | 1 (0.9) | 80 (98.8) |
RAS: Resistance-associated substitution; HCV: Hepatitis C virus. The source of these RASs is the 2020 EASL recommendations on treatment of hepatitis C [25].
Table 2.
The full list of NS5A RASs in the included sequences stratified per HCV genotype/subtype.
| HCV genotype/subtype: RASs | N (%) | 1a (n=1533) | 1b (n=617) | 2 (n=244) | 3 (n=851) | 4 (n=116) | 6 (n=81) |
|---|---|---|---|---|---|---|---|
| 1a: K24E/Q/R/T 1b: Q24K 2: T24A/S 3: S24F 6: Q24H |
1051 (31.6) | 502 (32.7) | 344 (55.8) | 205 (84.0) | 0 | - | 0 |
| 1a: K26E | 0 | 0 | - | - | - | - | - |
| 1a: M28A/G/S/T/V 1b: L28A/M/T 2: L/F28C/S 3: M28T/K 4: L28M/S/T/V 6: F/L28A/I/L/M/T/V |
757 (22.7) | 67 (4.4) | 522 (84.6) | 0 | 0 | 84 (72.4) | 81 (100.0) |
| 1a: P29R 1b: P29S, del29 2: P29S |
0 | 0 | 0 | 0 | - | - | - |
| 1a: Q30C/D/E/G/H/K/L/N/R/T/Y, del30 1b: R30G/H/P/Q/S 2: L30H/S 3: A30D/E/K/S 4: L30F/G/H/R/S 6: R30E/H/N/S |
1105 (33.1) | 491 (32.0) | 351 (56.9) | 3 (1.2) | 230 (27.0) | 27 (23.3) | 3 (3.7) |
| 1a: L31I/F/M/P/V 1b: L31F/I/M/V/W 2: L31I/M/V 3: L31F/I/M/P/V 4: M/L31I/V 6: L31I/M/V |
251 (7.5) | 6 (0.4) | 21 (3.4) | 15 (6.1) | 190 (22.3) | 0 | 19 (23.5) |
| 1a: P32L/S, del32 1b: P32F/L/S, del32 6: P32A/L/Q/R/S |
0 | 0 | 0 | - | - | - | 0 |
| 1a: S38F | 0 | 0 | - | - | - | - | - |
| 1a: H58C/D/L/P/R 1b: P58A/D/L/S/R/T 4: T58A/P/S 6: T58A/G/H/N/S |
750 (32.0) | 637 (41.6) | 3 (0.5) | - | - | 76 (65.5) | 34 (42.0) |
| 1b: Q/E62D 3: S62L |
23 (1.6) | - | 6 (1.0) | - | 17 (2.0) | - | - |
| 1a: A92K/T 1b: A92E/K/T/V 2: C92R/S/T/W 3: E92K 6: E92T |
200 (6.0) | 9 (0.6) | 189 (30.6) | 2 (0.8) | 0 | - | 0 |
| 1a: Y93C/F/H/L/N/R/S/T/W 1b: Y93C/H/N/R/S/T 2: Y93F/N/H 3: Y93H/N/S 4: Y93C/H/N/S/R/W 6: T93A/H/N/S |
166 (5.0) | 41 (2.7) | 17 (2.8) | 48 (19.7) | 52 (6.1) | 4 (3.4) | 4 (4.9) |
| Sequences with any NS5A RASs | 2073 (62.0) | 721 (47.0) | 551 (89.3) | 208 (85.2) | 396 (46.5) | 116 (100.0) | 81 (100.0) |
RAS: Resistance-associated substitution; HCV: Hepatitis C virus. The source of these RASs is the 2020 EASL recommendations on treatment of hepatitis C [25].
Table 3.
The full list of NS5B RASs in the included sequences stratified per HCV genotype/subtype.
| HCV genotype/subtype: RASs | N (%) | 1a (n=1534) | 1b (n=617) | 2 (n=244) | 3 (n=851) | 4 (n=116) | 6 (n=81) |
|---|---|---|---|---|---|---|---|
| NIs | |||||||
| 3: A150V | 100 (11.8) | - | - | - | 100 (11.8) | - | - |
| 1a: L159F 1b: L159F 2: L159F 3: L159F |
45 (1.4) | 40 (2.6) | 3 (0.5) | 0 | 2 (0.2) | - | - |
| 3: K206E | 43 (5.1) | - | - | - | 43 (5.1) | - | - |
| 1a: S282G/R/T 1b: S282G/R/T 2: S282G/R/T 3: S282G/R/T 4: S282C/G/R/T 6: S282G/R/T |
1/3443 (0.03) | 0 | 0 | 1 (0.4) | 0 | 0 | 0 |
| 1a: C316H/R 1b: C316F/H/N |
9 (0.4) | 5 (0.3) | 4 (0.6) | - | - | - | - |
| 1a: L320I/F/V | 0 | 0 | - | - | - | - | - |
| 1a: V321A 1b: V321I 3: V321A 4: V321A |
0 | 0 | 0 | - | 0 | 0 | - |
| NNI | |||||||
| 1a: L314H | 0 | 0 | - | - | - | - | - |
| 1a: C316Y 1b: C316H/N/Y/W |
5 (0.2) | 0 | 5 (0.8) | - | - | - | - |
| 1b: S368T | 0 | - | 0 | - | - | - | - |
| 1a: A395G | 0 | 0 | - | - | - | - | - |
| 1b: N411S | 0 | - | 0 | - | - | - | - |
| 1a: M414I/T/V 1b: M414I/T/V |
8 (0.4) | 7 (0.5) | 1 (0.2) | - | - | - | - |
| C445F/Y | 203 (32.9) | - | 203 (32.9) | - | - | - | - |
| E446K/Q | 418 (27.2) | 418 (27.2) | - | - | - | - | - |
| 1a: Y448C/H 1b: Y448C/H |
6 (0.3) | 4 (0.3) | 2 (0.3) | - | - | - | - |
| 1a: A553T/V 1b: A553V |
280 (13.0) | 85 (5.5) | 195 (31.6) | - | - | - | - |
| 1a: G554S 1b: G554S |
4 (0.2) | 3 (0.2) | 1 (0.2) | - | - | - | - |
| 1a: Y555H | 0 | 0 | - | - | - | - | - |
| 1a: S556G/R 1b: S556G/R |
343 (15.9) | 142 (9.3) | 201 (32.6) | - | - | - | - |
| 1a: G557R | 0 | 0 | - | - | - | - | - |
| 1a: G558R 1b: G558R |
0 | 0 | 0 | - | - | - | - |
| 1a: D559G/N 1b: D559G/N |
2 (0.09) | 2 (0.1) | 0 | - | - | - | - |
| 1a: Y561H/N | 1 (0.07) | 1 (0.07) | - | - | - | - | - |
| 1a: S565F | 0 | 0 | - | - | - | - | - |
| Sequences with any NS5B RASs | 872 (25.3) | 521 (34.0) | 215 (34.8) | 1 (0.4) | 135 (15.9) | 0 | 0 |
RAS: Resistance-associated substitution; HCV: Hepatitis C virus; NI: Nucleoside inhibitor; NNI: non-nucleoside inhibitor. The source of these RASs is the 2020 EASL recommendations on treatment of hepatitis C [25].
Table 4.
The clinically-relevant RASs identified for four DAAs.
| aa Position in NS5A | 28 | 30 | 31 | 93 |
|---|---|---|---|---|
| Regimen | ||||
| Harvoni (subtype 1a) | - | Q30H/R | L31M/V | Y93C/H/N |
| N (%) | 426 (27.8) | 83 (5.4) | 29 (1.9) | |
| Harvoni (subtype 1b) | - | - | L31V | Y93H |
| N (%) | 0 | 17 (2.8) | ||
| Zepatier (subtype 1a) | M28A/T | Q30H/R | L31M/V | Y93C/H/N |
| N (%) | 6 (0.4) | 426 (27.8) | 83 (5.4) | 29 (1.9) |
| Zepatier (subtype 1b) | - | - | - | Y93H |
| N (%) | 17 (2.8) | |||
| Epclusa (genotype 3) | - | - | - | Y93H |
| N (%) | 35 (4.1) | |||
| Mavyret (genotype 3) | - | A30K | - | - |
| N (%) | 211 (24.8) |
N: Number; aa: amino acid. The source of these cr-RASs is the AASLD-IDSA HCV Guidance [59].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



