
Submitted:
01 July 2024
Posted:
01 July 2024
You are already at the latest version
Abstract
Aquaporins (AQPs) are a family of integral membrane proteins that selectively transport water and glycerol across the cell membrane. Because AQPs are involved in a wide range of physiological functions and pathophysiological conditions, AQP-based therapeutics may have broad potential for clinical utility, including disorders of water and energy balance. However, AQP modulators have not yet been developed as suitable candidates for clinical applications. In this study, to identify potential modulators of AQPs, we screened 32 natural products by measuring the water and glycerol permeability of mouse erythrocyte membranes using a stopped-flow light scattering method. None of the tested natural compounds substantially affected osmotic water permeability. However, several compounds considerably affected glycerol permeability. Stichoposide C increased the glycerol permeability of mouse erythrocyte membranes, whereas rhizochalin decreased it at nanomolar concentrations. Immunohistochemistry revealed that AQP7 was the main aquaglyceroporin in mouse erythrocyte membranes. We further verified the effects of stichoposide C and rhizochalin on aquaglyceroporins using human AQP3 expressing keratinocyte cells. Stichoposide C, but not stichoposide D, increased AQP3-mediated transepithelial glycerol transport, whereas the peracetyl aglycon of rhizochalin was the most potent inhibitor of glycerol transport among the tested rhizochalin derivatives. Collectively, stichoposide C and the peracetyl aglycon of rhizochalin might function as modulators of AQP3 and AQP7, and suggest the possibility of these natural products as potential drug candidates for aquaglyceroporin modulators
Keywords:
Aquaglyceroporin
; stichoposide C
; peracetyl aglycon of rhizochalin
; stopped-flow light scattering
; glycerol permeability.
1. Introduction
Marine natural products are a rich source of potential drugs because of their high biodiversity, low toxicity, suitability for oral applications, and a variety of biological actions [1,2,3]. According to the global marine pharmaceutical pipeline website (www.marinepharmacology.org), 15 marine-derived drugs have been approved by the United States Food and Drug Administration (US FDA), and 32 marine-derived compounds are currently in various phases of clinical trials for drug development. Most have been developed or are under development as anticancer drugs, whereas a few are targeted for diseases other than cancer, such as pain, hypertriglyceridemia, Alzheimer’s disease, and viral infections. Considerable scientific and technological advances in analytical methods and functional assays in the past decade have opened up new opportunities for the exploration of marine natural products as novel potential therapeutic agents for more diverse disease entities [4,5,6].
Membrane transport proteins are important targets for drug discovery and delivery [7,8]. Transport proteins are essential for translocating solutes across the plasma or intracellular membranes, thus maintaining homeostasis. Transport proteins include channels and membrane transporters that are divided into three distinct classes: solute carriers (SLC), ATP-binding cassette (ABC) transporters, and ATPase ion pumps [9,10]. According to the data from The HUGO Gene Nomenclature Committee (https://genenames.org/), transport proteins account for approximately 6% of all human protein-coding genes: 427 genes of SLC superfamily, 312 genes of ABC transporters, 125 genes of ATPase, and 291 genes of channels. The loss or alteration of the function of membrane transporters and channels causes a wide variety of human diseases, including channelopathies [11,12,13]. Membrane transporters are rapidly emerging as potential drug targets but have remained understudied until recently [14,15]. Moreover, since membrane transporters function as drug transporters, they play key roles in the absorption, distribution, and elimination of drugs and determine the therapeutic efficacy and adverse reactions of drugs in the process of drug discovery and development [16,17,18].
Aquaporins (AQPs) are a family of small integral membrane proteins that primarily transport water across cell membranes along osmotic gradients. To date, 13 AQP subtypes have been found in mammals (AQP0-12), some of which permit transcellular passage of glycerol and urea as well as water (AQP3, 7, 9, and 10), and are thus called aquaglyceroporins [19]. AQPs are key players in maintaining normal water balance and energy homeostasis in the body [20,21]. AQPs have been implicated in multiple disorders including diabetes insipidus, brain edema, cancer, obesity, cataracts, and neuromyelitis optica [22,23,24,25,26]. In addition to AQP-based diseases, AQP modulators have potential therapeutic utility in various pathological conditions for correcting abnormal transepithelial fluid and glycerol transport [20,21,27]. Notwithstanding the vigorous efforts to identify aquaporin modulators, limited progress has been made in the development of aquaporin-based therapeutics [28]. The relatively low hit rate, poor druggability due to toxicity, and lack of specificity makes the exploration of new AQP-targeted drugs challenging [29,30]. Because marine natural products not only possess greater structural and chemical diversity but also less toxicity than synthetic chemical libraries, the difficulty in finding promising lead compounds as AQP modulators using synthetic compound libraries encouraged us to conduct the present study to screen marine natural products for novel drugs targeting AQPs.
2. Results
2.1. Effects of marine natural compounds on the osmotic water permeability of the erythrocyte membrane
We first investigated whether marine natural compounds could alter the osmotic water permeability of mouse erythrocyte membranes. To measure membrane water permeability, the kinetics of cell volume changes were examined using light scattering (Figure 1a). In the presence of an inwardly directed gradient of an impermeable solute, water moves out of the cells according to the osmotic gradient through AQPs. Consequently, the cell volume decreases, which causes an increase in light scattering. As shown in Figure 1b, in response to an inward gradient of 300 mM sucrose, the scattered light intensity rapidly increased and reached a steady state. HgCl2 is a well-known inhibitor of various AQPs and prevents cell shrinkage in response to osmotic gradients by inhibiting water efflux. DMSO also slows osmotic water movement owing to an osmotic clamping effect rather than direct inhibition of water efflux. The value of the osmotic water permeability coefficient (Pf), which was calculated from the time courses of the increase in light scattering upon exposure to 300 mM sucrose, was 0.033 cm/sec (Figure 1c). In the presence of DMSO or HgCl2, the Pf values decreased 30% and 88%, respectively. Figures 1, 3a and 3d show the representative light scattering traces for 32 marine natural compounds (100 nM) in response to 300 mM sucrose. The calculated Pf values in the presence of marine natural compounds were not changed when compared with that of the control (data not shown), suggesting no significant effects on osmotic water permeability.
2.2. Effects of marine natural compounds on glycerol permeability of erythrocyte membrane
We next investigated whether any marine natural compounds might change the glycerol permeability of the moue erythrocyte membrane. To measure membrane glycerol permeability, changes in cell volume in response to an imposed glycerol gradient were measured by light scattering (Figure 2a). In the presence of an inwardly directed gradient of glycerol, a permeable solute, water efflux through AQPs occurs first according to the osmotic gradient, followed by water influx accompanying glycerol influx through aquaglyceroporins. As shown in Figure 2b, scattered light intensity rapidly increased for the first ~0.3 s in response to an inwardly directed gradient of 300 mM glycerol in the same manner as that of sucrose, but slowly decreased afterwards as the cell volume was recovered by the entry of glycerol and water. DMSO slowed the initial increase in light scattering but did not affect the late decreasing phase. However, HgCl2, an inhibitor of AQPs including aquaglyceroporins, reduced not only the initial rise but also the late decreasing phase of light scattering. Figure 2c shows the representative light-scattering traces for 24 marine natural compounds (100 nM) in response to 300 mM glycerol. The glycerol permeability coefficients (Pglycerol) calculated from the time courses of the late decreasing phase of light scattering upon exposure to 300 mM glycerol alone or in the presence of HgCl2 were 3.71×10-6 cm/sec and 0.78×10-6 cm/sec, respectively (Figure 4). As shown in Figures 2, 3b and 3d, several marine natural compounds increased or decreased the decrement rate of the late phase of light-scattering kinetics driven by glycerol influx through aquaglyceroporins. Among them, most notably, stichoposide C increased glycerol permeability, whereas rhizochalin decreased it.
2.3. Effects of stichoposide C, rhizochalin, and their derivatives on glycerol permeability of erythrocyte membrane
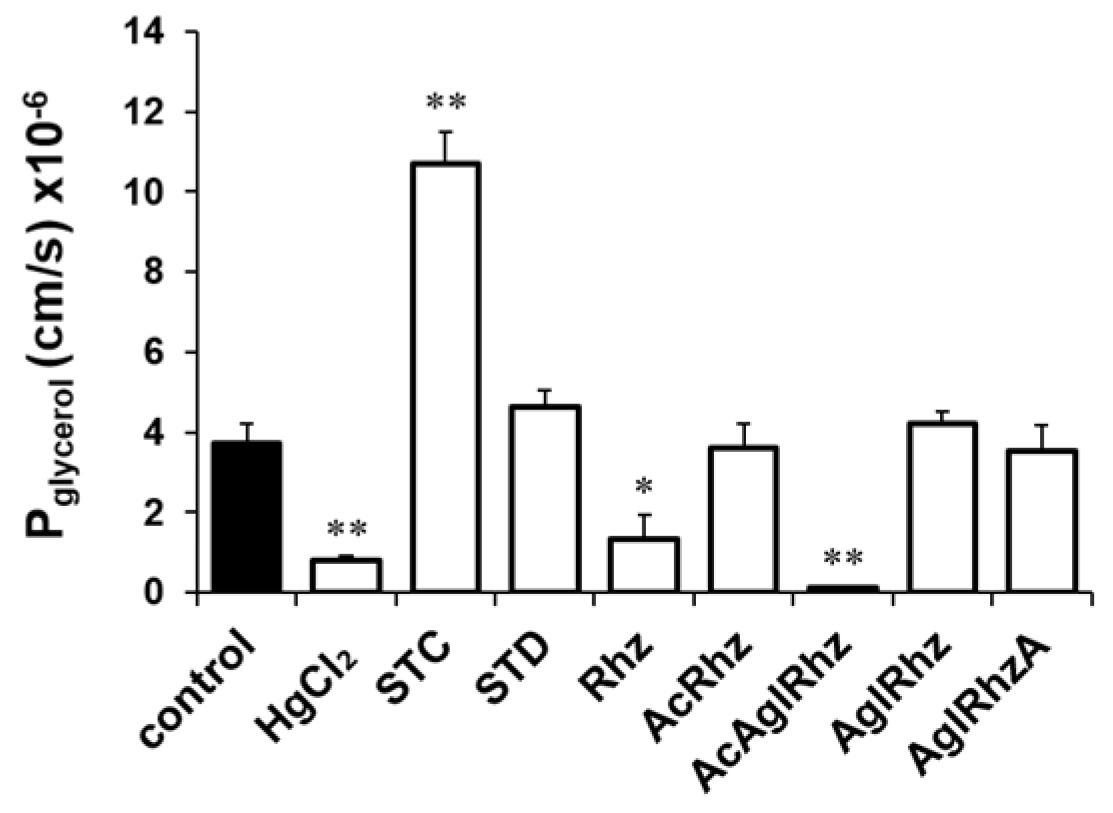
The effect of stichoposide C on the glycerol permeability of erythrocyte membranes was further investigated. The promoting effect of stichoposide C on glycerol permeability was concentration-dependent in the range from 100 nM to 1 μM (Figure 3c). In contrast to stichoposide C, stichoposide D, a structural analog of stichoposide C, did not exert any significant influence on the late phase of light-scattering kinetics up to a concentration of 1 μM (Figure 3b). The calculated Pglycerol values in the presence of stichoposide C or Stichoposide D were 10.71×10-6 cm/sec (2.88-fold increase, P < 0.01) and 4.64×10-6 cm/sec (1.25-fold increase), respectively (Figure 4).
The effects of rhizochalin and its derivatives on the glycerol permeability of erythrocyte membranes were also investigated (Figure 3e). The peracetyl aglycon of rhizochalin decreased glycerol permeability more profoundly than rhizochalin. However, aglycon of rhizochalin, aglycon of Rhizochalin A, and rhizochalin peracetate did not have any significant effect on glycerol permeability, in contrast to rhizochalin. Both rhizochalin and the peracetyl aglycon of rhizochalin exhibited concentration-dependent inhibition of glycerol permeability (Figure 3f). The calculated Pglycerol values in the presence of rhizochalin or the peracetyl aglycon of rhizochalin were 1.33×10-6 cm/sec (2.79-fold decrease, P<0.05) and 0.15×10-6 cm/sec (24.73-fold decrease, P < 0.01), respectively, indicating that the peracetyl aglycon of rhizochalin was a more potent inhibitor of glycerol permeability than that of rhizochalin (Figure 4).
2.4. Expression of AQP subtypes in mouse erythrocytes
To elucidate which AQP subtypes are responsible for water and glycerol permeability, we analyzed CD1 mouse erythrocytes by immunohistochemistry using nine AQP subtypes. As shown in Figure 5, AQP1 and AQP7 were strongly expressed in mouse erythrocyte membranes, and AQP7 was the main aquaglyceroporin that determined glycerol permeability in mouse erythrocytes.
Figure 4.
Glycerol permeability coefficients of mouse erythrocytes in the presence of stichoposide, rhizochalin, and their derivatives. Glycerol permeability coefficients (Pglycerol) were calculated from the time course of scattered light intensity in response to 300 mM glycerol gradient. Erythrocytes were incubated with 0.5 mM of HgCl2 or 100 nM of the compounds for 10 min before measurement. STC, stichoposide C; STD, stichoposide D; rhizochalin (Rhz); AglRhz, aglycon of rhizochalin; AcRhz, peracetyl rhizochalin; AcAglRhz, peracetyl aglycon of rhizochalin; AglRhzA, aglycon of rhizochalin A. Values are mean ± SD. *P<0.05, **P<0.01.
Figure 4.
Glycerol permeability coefficients of mouse erythrocytes in the presence of stichoposide, rhizochalin, and their derivatives. Glycerol permeability coefficients (Pglycerol) were calculated from the time course of scattered light intensity in response to 300 mM glycerol gradient. Erythrocytes were incubated with 0.5 mM of HgCl2 or 100 nM of the compounds for 10 min before measurement. STC, stichoposide C; STD, stichoposide D; rhizochalin (Rhz); AglRhz, aglycon of rhizochalin; AcRhz, peracetyl rhizochalin; AcAglRhz, peracetyl aglycon of rhizochalin; AglRhzA, aglycon of rhizochalin A. Values are mean ± SD. *P<0.05, **P<0.01.
2.5. Effects of stichoposide C, rhizochalin, and their derivatives on AQP3-mediated transepithelial glycerol transport
The effects of stichoposide C and rhizochalin on aquaglyceroporins were further explored using AQP3-expressing HaCaT cell monolayers to directly measure transepithelial glycerol transport (Figure 6a). When cells grown on permeable supports were exposed to 20 mM glycerol in the lower basolateral chamber, the glycerol concentrations in the upper apical chambers increased with time up to 30 min in both YFP-hAQP1 and YFP-hAQP3 HaCaT cells (Figure 6b). However, glycerol concentrations in YFP-hAQP3 HaCaT cells were over two-fold higher than those in YFP-hAQP1 HaCaT cells at every measured time point, that validated our method to measure AQP3-mediated transepithelial glycerol transport using YFP-hAQP3 HaCaT cells. Stichoposide C significantly stimulated AQP3-mediated basolateral glycerol transport in YFP-hAQP3 HaCaT cell monolayers (Figure 6c and 6d). The glycerol concentrations in the upper apical chambers at 10 min and inside the cells at 30 min were 2.30-fold (P < 0.01) and 1.97-fold (P<0.05) increased, respectively, by Stichoposide C. The peracetyl aglycon of rhizochalin significantly decreased the glycerol concentrations in the upper apical chambers at 10 min (1.63-fold, P<0.05) and inside the cells at 30 min (1.52-fold, P<0.05). Stichoposide D increased, and rhizochalin decreased AQP3-mediated transepithelial glycerol transport to some extent, but there were no statistically significant differences.
3. Discussion
In this study, we aimed to identify potential AQP modulators in marine natural products. Although none of the marine natural compounds tested had significant effects on osmotic water permeability, we found several compounds that significantly affected glycerol permeability in mouse erythrocytes. We confirmed the modulatory effects of stichoposide C and rhizochalin on aquaglyceroporins through transepithelial glycerol transport assays using AQP3-expressing human keratinocytes.
Natural products, including plants, animals, and minerals, continue to be the best sources of novel drugs owing to their structural diversity and biodiversity [5,6]. In 2023, 10 new natural products or their direct derivatives were drug approved by the US FDA, which corresponds to 18% of total 55 new drugs on the market [32]. Because the total number of species and biochemical diversity in oceans are higher than those on land, marine natural products are more invaluable sources for potential drug discovery than terrestrial products [3,4]. Moreover, natural products released into water are rapidly diluted and, therefore, need to be highly potent to have any effect. These advantages of marine natural products, along with the growing appreciation for functional assays and phenotypic screening, may further contribute to the revival of interest in natural products for drug discovery and development [33,34].
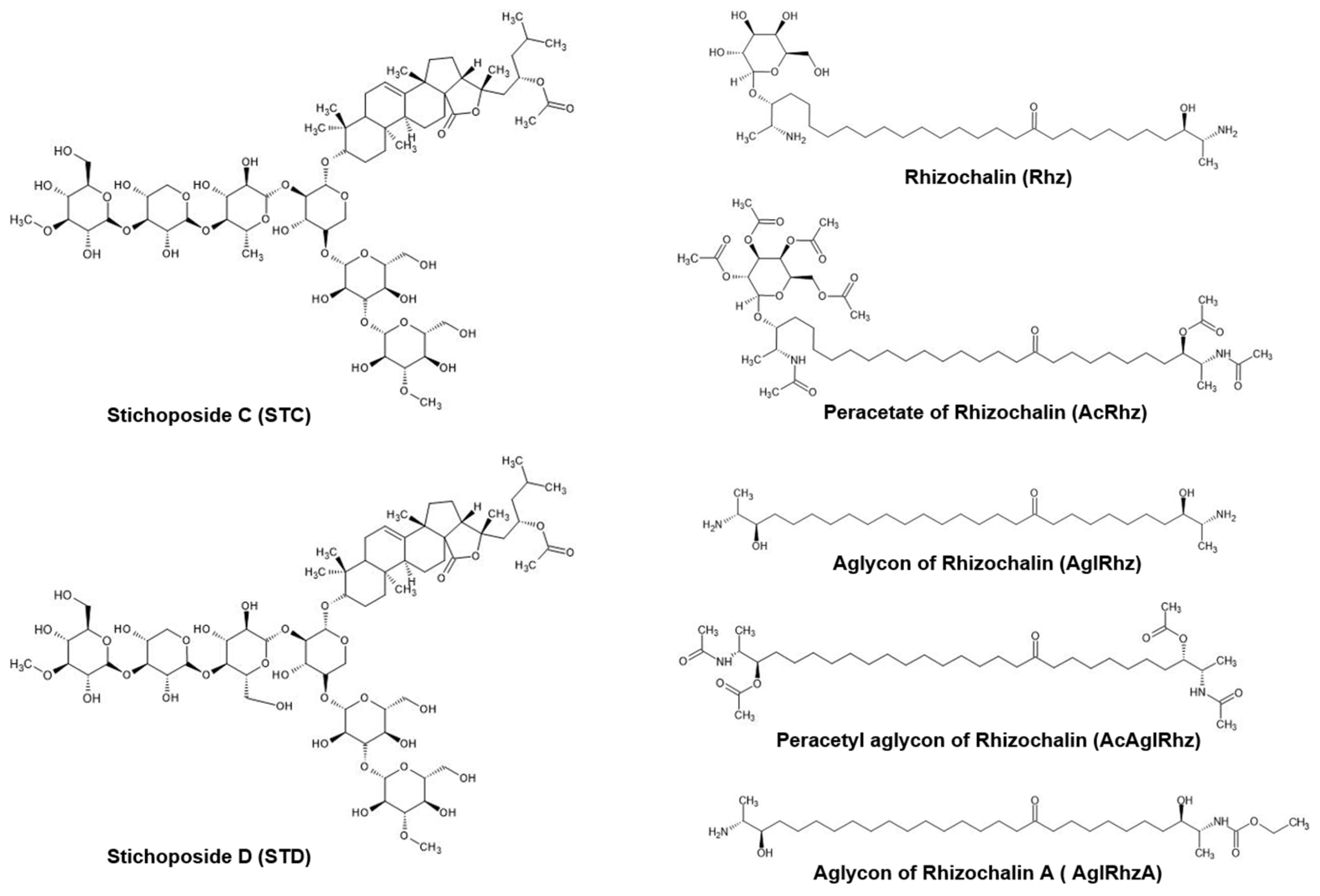
Triterpene glycosides are widely distributed not only in plants, but also in marine invertebrates, especially echinoderms, octocorals, and sponges [35,36]. Many investigators have attempted to develop marine triterpene glycosides as candidate anticancer agents based on in vitro and in vivo studies [37,38,39]. Stichoposides C and D are triterpene glycosides extracted from sea cucumbers of the family Stichopodidae [40,41]. Stichoposide C is a quinovose-containing hexoside, whereas stichoposide D is a glucose-containing hexoside (Figure 7). Stichoposide C is an active membrane-acting agent with anticancer activity, and is more potent than stichoposide D [42,43]. In the present study, stichoposide C strongly stimulated glycerol permeability in murine erythrocytes and AQP3-expressing human keratinocytes, whereas stichoposide D did not. Based on the notion that a linear tetrasaccharide fragment in triterpene glycosides is essential for the actions leading to the modification of the cellular membrane [37,44,45], having quinovose, rather than glucose, as a second monosaccharide unit in glycosides seems to be critical for interaction with 5(6)-unsaturated sterols of cellular membranes in contact with aquaglyceroporins or direct interaction with channel proteins. Otherwise, STC-specific activation of the sphingomyelinase-ceramide pathway might account for the different effects on aquaglyceroporins [38]. However, further studies on the structure–activity relationship of these molecules are needed to improve the efficacy and safety of these compounds in activating aquaglyceroporins.
Rhizochalin is a marine two-headed sphingolipid-like natural product isolated from the sponge Rhizochalina incrustata [46,47]. These compounds differ from classical sphingolipids in the presence of polar groups at α,ω-positions, which contain a terminal methyl group instead of hydroxymethyl group unlike sphingoid bases, thus representing a unique class of bipolar lipids [48,49]. Rhizochalin and its analogs have been reported to have antibacterial, antifungal, and cytotoxic activities [50,51,52,53]. In the present study, the peracetyl aglycon of rhizochalin exhibited the most potent inhibitory effect on glycerol permeability in murine erythrocytes and human AQP3-expressing epithelial cells. It might be assumed that the absence of a glucopyranosyl sugar moiety on one polar end, together with the presence of three acetyl moieties on the other polar end of two-headed bipolar lipids (Figure 7), favors an intimate interaction with aquaglyceroporin or neighboring membrane lipids and regulatory proteins. The molecular mechanism underlying the inhibitory effect of the peracetyl aglycon of rhizochalin on aquaglyceroporins and the structure–activity relationship of rhizochalin analogs require further investigation.
Aquaglyceroporins mediate various physiological and pathophysiological processes [54,55,56]. Water-selective AQPs function primarily in water and salt homeostasis, whereas water/glycerol-transporting aquaglyceroporins are primarily involved in energy metabolism and lubrication. Because aquaglyceroporin dysfunction is implicated in various human diseases and symptoms, including obesity, nonalcoholic fatty liver disease, psoriasis, cancer, polyuria, and glyceroluria, functional modulators of aquaglyceroporins are promising therapeutic targets for a wide range of clinical conditions [24,57,58,59]. A few dozen aquaglyceroporin modulators have been reported to date, but none have been developed or are under development. Metal compounds, such as HgCl2 and p-chloromercuribenzene sulfonate (pCMBS), were first identified as AQP inhibitors, including aquaglyceroporins, and are still the most commonly used inhibitors in functional assays [60]. The gold compound Auphen ([Au(phen)Cl2]Cl) and its copper analog Cuphen ([Cu(phen)Cl2]) inhibited AQP3 and AQP7 [61,62]. Recently, several aquaglyceroporin inhibitors were identified by screening a commercially available library of small molecules: DFP00173 for AQP3, Z433927330 for AQP7, and HTS13286 for AQP9 inhibitors [63,64]. Natural compounds were also reported to have modulating effects on aquaglyceroporins which are usually from plants, such as glycolic acid, 18β-glycyrrhetinic acid, curcumin, chrysin and phloretin [59]. However, the application of aquaglyceroporin modulators in clinical trials is still far from being implemented, possibly because of the lack of selective and potent modulators that can be administered in vivo.
In this study, we searched for potential AQP modulators among marine natural products and found that stichoposide C and the peracetyl aglycon of rhizochalin were potential aquaglyceroporin modulators. Since mouse erythrocytes express AQP7 and transepithelial glycerol transport was measured using human AQP3-expressing cells, it can be stated that stichoposide C and the peracetyl aglycon of rhizochalin function as modulators of AQP3 and AQP7. AQP3 activators may exert therapeutic effects on defects in the hydration, lubrication, and proliferation of the skin, bladder, vagina, and respiratory system, whereas AQP7 activators may be applicable as useful adjuvants for obesity treatment. Therefore, AQP3 and AQP7 inhibitors are worthy of development as anticancer drugs. Further studies are needed to verify the therapeutic effects of stichoposide C and the peracetyl aglycon of rhizochalin in animal models of human diseases or in 3D bioengineered disease models.
4. Materials and Methods
4.1. Marine natural products
The marine natural products were provided by Dr. V. A. Stonik at the Pacific Institute of Bioorganic Chemistry, Far Eastern Branch of the Russian Academy of Science, Vladivostok, Russian Federation. Thirty two compounds tested were extracted from marine natural products. The compounds were dissolved in phosphate-buffered saline (PBS) or dimethyl sulfoxide (DMSO) as 1–2 mM stock solutions, and diluted to working concentrations ranging from 100 nM to 1 μM.
4.2. Erythrocyte preparation
Thirty CD1 mice, 6–8 weeks old, were used in this experiment. The mice were anesthetized with avertin (2,2,2-tribromoethanol-tert-amyl alcohol, 250 mg/kg intraperitoneally). Blood was collected from the inferior vena cava in heparinized tubes. Freshly obtained whole blood was centrifuged at 120 × g for 10 min to remove the plasma and buffy coat. The erythrocytes were washed three times and suspended in PBS to adjust the final hematocrit to 2%. All animal protocols were approved by the Dong-A University Medical School Institutional Animal Care and Use Committee (DIACUC-10-6-1).
4.3. Stopped-flow light scattering measurements
Water and glycerol permeabilities were measured via light scattering using a stopped-flow apparatus (SFM-3, Biologic, France). The erythrocyte suspension was subjected to a 300 mM inwardly directed gradient of either sucrose or glycerol. The kinetics of decreasing cell volume were measured from the time course of 90° scattered light intensity at 530 nm. The osmotic water and glycerol permeability coefficients were calculated from the time course of light scattering using the following equations:
dV(t)/dt = Pf x SAV x MVW x (Cin - Cout)
where V(t) is the relative red cell volume as a function of time, SAV is the red cell surface area-to-initial volume ratio, MVW is the molar volume of water (18 cm3/mole), and Cin and Cout (mole/cm3) are the initial concentrations of the intracellular and extracellular solutes, respectively. The average erythrocyte surface area and volume were 7.2 × 10-7 cm2 and 3.1 × 10-11 cm3, respectively [31].
4.4. Tissues preparation and immunohistochemistry
CD1 mouse kidneys were fixed in 10% neutral buffered formalin for 48 h, paraffin-embedded, and sectioned at 5 μm using a microtome (RM 2125RT, Leica, Wetzlar, Germany). Paraffin sections were deparaffinized with xylene, serially rehydrated in 100%, 95%, and 70% ethanol, and boiled for 15 min in citrate buffer (10 mM sodium citrate, 0.05% Tween 20, and pH 6.0) for antigen retrieval. After blocking with 5% bovine serum albumin for 2 h, the sections were incubated with the following primary antibodies overnight at 4°C at a dilution of 1:300: anti-AQP1, AQP2, AQP3, and AQP4 (Merck Millipore, Billerica, MA, USA); anti-AQP5, AQP6, and AQP7 (Alomone Labs, Jerusalem, Israel); anti-AQP8 (Mybiosource, CA, USA); and anti-AQP9 (Bioss, Woburn, MA, USA). Subsequently, the sections were incubated with secondary anti-rabbit horseradish peroxidase (HRP)-linked IgG (Dako, Glostrup, Denmark) for 40 min at 25°C, and HRP activity was determined by adding DAB+ chromogen (Dako). The sections were mounted using Marinol Mount (Muto Pure Chemicals, Tokyo, Japan) solution and xylene (Muto Pure Chemicals) (1:1), and then visualized using a digital slide scanner (Pannoramic Midi, 3DHistech, Budapest, Hungary) at the Neuroscience Translational Research Solution Center (Busan, South Korea).
4.5. Cell cultures
HaCaT cells, an immortalized human keratinocyte line, were stably transfected with a plasmid encoding yellow fluorescent protein (YFP) and either human AQP1 (YFP-hAQP1) or human AQP3 (YFP-hAQP3) (gifts from Dr. Alan Verkman at the University of California, San Francisco, California, USA) and selected using G418 (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA). Cells were grown in DMEM medium (Gibco, Thermo Fisher Scientific) supplemented with 10% fetal bovine serum (Hyclone™, Cytiva, Logan, UT, USA), penicillin G (100 U/mL), streptomycin (100 μg/mL) and maintained at 37°C in a humidified 5% CO2 incubator.
4.6. Measurement of transepithelial glycerol transport
YFP-hAQP1-HaCaT cells and YFP-hAQP3-HaCaT cells were seeded at a density of 2 × 104 cells/well onto 6.5 mm, 0.4 μm pore size Transwell permeable supports (Corning Life Sciences, Tewksbury, MA, USA), cultured for 3–5 days to a confluent state, and serum-deprived for 2–3 days before the experiments. Once the cells reached a confluent state, the integrity of the cellular barriers was assessed by transepithelial electrical resistance (TEER) using an epithelial voltohmmeter. When the TEER values were above 200 Ω.cm2, transepithelial flux experiments were performed.
Glycerol transport across HaCaT cell monolayers was determined by measuring the glycerol concentrations of the culture medium in the upper and lower compartments, as well as in the Transwell inserts. The cells were pretreated with marine natural products or HgCl2 for 10 min, and 20 mM glycerol was added to the bath solution on the basal side of the cells. Aliquots (10 µL) of samples were collected from the medium of the opposite side from glycerol addition at the indicated time points. Glycerol concentrations were measured using a glycerol assay kit (Sigma-Aldrich, Saint Louis, MO, USA) according to the manufacturer’s instructions.
4.7. Statistical analysis
All the data were expressed as mean ± standard deviation (SD). The statistical significance of differences between groups was analyzed using Student’s t-test or one-way ANOVA. P<0.05 was considered statistically significant.
Author Contributions
Conceptualization, J.Y.K. and H.R.B.; methodology, J.W.I., J.H.L. and H.R.B.; software, S.J. and M.S.; validation, J.W.I., S.J. and H.R.B.; formal analysis, J.W.I. and H.R.B.; investigation, J.W.I. and J.H.L.; resources, J.Y.K. and V.A.S.; data curation, J.W.I. and H.R.B.; writing—original draft preparation, J.W.I., J.H.L. and H.R.B.; writing—review and editing, M.S. and H.R.B.; visualization, J.W.I. and J.H.L.; supervision, , J.Y.K. and H.R.B; project administration, H.R.B.; funding acquisition, H.R.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Research Foundation of Korea (NRF) grant. funded by the Korean government (MSIT) (No. 2021R111A3057381).
Institutional Review Board Statement
Not applicable
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stonik, V.A. Marine natural products: A way to new drugs. Acta Naturae 2009, 1, 15-25.
- Blunt, J.W.; Copp, B.R.; Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2017, 34, 235-294. [CrossRef]
- Jiménez, C. Marine natural products in medicinal chemistry. ACS Med. Chem. Lett. 2018, 9, 959-961. https://doi: 10.1021/acsmedchemlett.8b00368.
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 69-85. [CrossRef]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111-129. [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200-216. [CrossRef]
- The International Transporter Consortium.; Giacomini, K.M.; Huang, S.M.; Tweedie, D.J.; Benet, L.Z.; Brouwer, K.L.R.; Chu, X.; Dahlin, A.; Evers, R.; Fischer, V.; Hillgren, K.M.; et al. Membrane transporters in drug development. Nat. Rev. Drug Discov. 2010, 9, 215-236. [CrossRef]
- Alam, S.; Doherty, E.; Ortega-Prieto, P.; Arizanova, J.; Fets, L. Membrane transporters in cell physiology, cancer metabolism and drug response. Dis. model. mech. 2023, 16. [CrossRef]
- Pizzagalli, M.D.; Bensimon, A.; Superti-Furga, G. A Guide to Plasma Membrane Solute Carrier Proteins. FEBS J. 2021, 288, 2784–2835. [CrossRef]
- Thomas, C.; Tampé, R. Structural and Mechanistic Principles of ABC Transporters. Annu. Rev. Biochem. 2020, 89, 605–636. [CrossRef]
- Kass, R.S. The Channelopathies: Novel Insights into Molecular and Genetic Mechanisms of Human Disease. J. Clin. Invest. 2005, 115, 1986–1989. [CrossRef]
- Hediger, M.A.; Clémençon, B.; Burrier, R.E.; Bruford, E.A. The ABCs of Membrane Transporters in Health and Disease (SLC Series): Introduction. Molecular Aspects of Medicine 2013, 34, 95–107. [CrossRef]
- Harraz, O.F.; Delpire, E. Recent Insights into Channelopathies. Physiol. Rev. 2024, 104, 23–31. [CrossRef]
- Lin, L.; Yee, S.W.; Kim, R.B.; Giacomini, K.M. SLC Transporters as Therapeutic Targets: Emerging Opportunities. Nat. Rev. Drug Discov. 2015, 14, 543–560. [CrossRef]
- César-Razquin, A.; Snijder, B.; Frappier-Brinton, T.; Isserlin, R.; Gyimesi, G.; Bai, X.; Reithmeier, R.A.; Hepworth, D.; Hediger, M.A.; Edwards, A.M.; et al. A Call for Systematic Research on Solute Carriers. Cell 2015, 162, 478–487. [CrossRef]
- Mizuno, N.; Niwa, T.; Yotsumoto, Y.; Sugiyama, Y. Impact of Drug Transporter Studies on Drug Discovery and Development. Pharmacol. Rev. 2003, 55, 425–461. [CrossRef]
- Shugarts, S.; Benet, L.Z. The Role of Transporters in the Pharmacokinetics of Orally Administered Drugs. Pharm. Res. 2009, 26, 2039–2054. [CrossRef]
- Galetin, A.; Brouwer, K.L.R.; Tweedie, D.; Yoshida, K.; Sjöstedt, N.; Aleksunes, L.; Chu, X.; Evers, R.; Hafey, M.J.; Lai, Y.; et al. Membrane Transporters in Drug Development and as Determinants of Precision Medicine. Nat. Rev. Drug Discov. 2024, 23, 255–280. [CrossRef]
- Ishibashi, K.; Morishita, Y.; Tanaka, Y. The Evolutionary Aspects of Aquaporin Family. Adv. Exp. Med. Biol. 2017, 969, 35–50. [CrossRef]
- Verkman, A.S. Aquaporins at a Glance. J. Cell Sci. 2011, 124, 2107–2112. [CrossRef]
- Verkman, A.S. Aquaporins. Curr. Biol. 2013, 23, R52–R55. [CrossRef]
- Moeller, H.B.; Rittig, S.; Fenton, R.A. Nephrogenic Diabetes Insipidus: Essential Insights into the Molecular Background and Potential Therapies for Treatment. Endocr. Rev. 2013, 34, 278–301. [CrossRef]
- Manley, G.T.; Fujimura, M.; Ma, T.; Noshita, N.; Filiz, F.; Bollen, A.W.; Chan, P.; Verkman, A.S. Aquaporin-4 Deletion in Mice Reduces Brain Edema after Acute Water Intoxication and Ischemic Stroke. Nat. Med. 2000, 6, 159–163. [CrossRef]
- Maeda, N.; Hibuse, T.; Funahashi, T. Role of Aquaporin-7 and Aquaporin-9 in Glycerol Metabolism; Involvement in Obesity. In Handb. Exp. Pharmacol.; Beitz, E., Ed.; Springer Naure: Berlin, Germany, 2009; Volume 190, pp. 233–249. [CrossRef]
- Berry, V.; Francis, P.; Kaushal, S.; Moore, A.; Bhattacharya, S. Missense Mutations in MIP Underlie Autosomal Dominant ‘polymorphic’ and Lamellar Cataracts Linked to 12q. Nat. Genet. 2000, 25, 15–17. [CrossRef]
- Papadopoulos, M.C.; Verkman, A.S. Aquaporin 4 and Neuromyelitis Optica. Lancet Neurol. 2012, 11, 535–544. [CrossRef]
- Verkman, A.S. Aquaporins in Clinical Medicine. Annu. Rev. Med. 2012, 63, 303–316. [CrossRef]
- Tradtrantip, L.; Jin, B.J.; Yao, X.; Anderson, M.O.; Verkman, A.S. Aquaporin-Targeted Therapeutics: State-of-the-Field. Adv. Exp. Med. Biol. 2017, 969, 239–250. [CrossRef]
- Verkman, A.S.; Anderson, M.O.; Papadopoulos, M.C. Aquaporins: Important but Elusive Drug Targets. Nat. Rev. Drug Discov. 2014, 13, 259–277. [CrossRef]
- Salman, M.M.; Kitchen, P.; Yool, A.J.; Bill, R.M. Recent Breakthroughs and Future Directions in Drugging Aquaporins. Trends Pharmacol. Sci. 2022, 43, 30–42. [CrossRef]
- Yang, B.; Fukuda, N.; van Hoek, A.; Matthay, M.A.; Ma, T.; Verkman, A.S. Carbon Dioxide Permeability of Aquaporin-1 Measured in Erythrocytes and Lung of Aquaporin-1 Null Mice and in Reconstituted Proteoliposomes. J. Biol. Chem. 2000, 275, 2686–2692. [CrossRef]
- de la Torre, B.G.; Albericio, F. The Pharmaceutical Industry in 2023: An Analysis of FDA Drug Approvals from the Perspective of Molecules. Molecules 2024, 29, 585. [CrossRef]
- Montaser, R.; Luesch, H. Marine Natural Products: A New Wave of Drugs? Future Med. Chem. 2011, 3, 1475–1489. [CrossRef]
- Romano, G.; Almeida, M.; Varela Coelho, A.; Cutignano, A.; Gonçalves, L.G.; Hansen, E.; Khnykin, D.; Mass, T.; Ramšak, A.; Rocha, M.S.; et al. Biomaterials and Bioactive Natural Products from Marine Invertebrates: From Basic Research to Innovative Applications. Mar. Drugs. 2022, 20, 219. [CrossRef]
- Stonik, V.A.; Kalinin, V.I.; Avilov, S.A. Toxins from Sea Cucumbers (Holothuroids): Chemical Structures, Properties, Taxonomic Distribution, Biosynthesis and Evolution. J. Nat. Toxins 1999, 8, 235–248.
- Kalinin, V.I.; Ivanchina, N.V.; Krasokhin, V.B.; Makarieva, T.N.; Stonik, V.A. Glycosides from Marine Sponges (Porifera, Demospongiae): Structures, Taxonomical Distribution, Biological Activities and Biological Roles. Mar. Drugs. 2012, 10, 1671–1710. [CrossRef]
- Aminin, D.L.; Menchinskaya, E.S.; Pisliagin, E.A.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I. Anticancer Activity of Sea Cucumber Triterpene Glycosides. Mar. Drugs. 2015, 13, 1202–1223. [CrossRef]
- Yun, S.H.; Sim, E.H.; Han, S.H.; Kim, T.R.; Ju, M.H.; Han, J.Y.; Jeong, J.S.; Kim, S.H.; Silchenko, A.S.; Stonik, V.A.; et al. In Vitro and in Vivo Anti-Leukemic Effects of Cladoloside C2 Are Mediated by Activation of Fas/Ceramide Synthase 6/P38 Kinase/c-Jun NH2-Terminal Kinase/Caspase-8. Oncotarget 2017, 9, 495–511. [CrossRef]
- Menchinskaya, E.S.; Dyshlovoy, S.A.; Venz, S.; Jacobsen, C.; Hauschild, J.; Rohlfing, T.; Silchenko, A.S.; Avilov, S.A.; Balabanov, S.; Bokemeyer, C.; et al. Anticancer Activity of the Marine Triterpene Glycoside Cucumarioside A2-2 in Human Prostate Cancer Cells. Mar. Drugs. 2024, 22, 20. [CrossRef]
- Kitagawa, I.; Kobayashi, M.; Inamoto, T.; Yasuzawa, T.; Kyogoku, Y. The Structures of Six Antifungal Oligoglycosides, Stichlorosides A1, A2, B1, B2, C1, and C2, from the Sea Cucumber Stichopus Chloronotus (BRANDT). Chem. Pharm. Bull. 1981, 29, 2387–2391. [CrossRef]
- Stonik, V.A.; Mal’tsev, I.I.; Kalinovskii, A.I.; Conde, C.; Elyakov, G.B. Glycosides of Marine Invertegrates. XI. Two New Triterpene Glycosides from Holothurians of the family Stichopadidae. Chem. Nat. Compd. 1982, 18, 177–182. [CrossRef]
- Yun, S.H.; Park, E.S.; Shin, S.W.; Na, Y.W.; Han, J.Y.; Jeong, J.S.; Shastina, V.V.; Stonik, V.A.; Park, J.I.; Kwak, J.Y. Stichoposide C Induces Apoptosis through the Generation of Ceramide in Leukemia and Colorectal Cancer Cells and Shows in Vivo Antitumor Activity. Clin. Cancer Res. 2012, 18, 5934–5948. [CrossRef]
- Fedorov, S.N.; Dyshlovoy, S.A.; Kuzmich, A.S.; Shubina, L.; Avilov, S.A.; Silchenko, A.S.; Bode, A.M.; Dong, Z.; Stonik, V.A. In Vitro Anticancer Activities of Some Triterpene Glycosides from Holothurians of Cucumariidae, Stichopodidae, Psolidae, Holothuriidae and Synaptidae Families. Nat. Prod. Commun. 2016, 11, 1239–1242.
- Park J.I.; Bae H.R.; Kim C.G.; Stonik V.A.; Kwak J.Y. Relationships between chemical structures and functions of triterpene glycosides isolated from sea cucumbers. Front Chem. 2014, 9, 2:77. doi: 10.3389/fchem.2014.00077.
- Bahrami, Y.; Franco, C.M.M. Acetylated Triterpene Glycosides and Their Biological Activity from Holothuroidea Reported in the Past Six Decades. Mar. Drugs. 2016, 14, 147. [CrossRef]
- Makarieva, T.N.; Denisenko, V.A.; Stonik, V.A.; Milgrom, Yu.M.; Rashkes, Ya.V. Rhizochalin, a Novel Secondary Metabolite of Mixed Biosynthesis from the Sponge Rhizochalina Incrustata. Tetrahedron Lett. 1989, 30, 6581–6584. [CrossRef]
- Makarieva, T.N.; Guzii, A.G.; Denisenko, V.A.; Dmitrenok, P.S.; Santalova, E.A.; Pokanevich, E.V.; Molinski, T.F.; Stonik, V.A. Rhizochalin A, a Novel Two-Headed Sphingolipid from the Sponge Rhizochalina Incrustata. J. Nat. Prod. 2005, 68, 255–257. [CrossRef]
- Molinski, T.F.; Makarieva, T.N.; Stonik, V.A. (-)-Rhizochalin Is a Dimeric Enantiomorphic (2R)-Sphingolipid: Absolute Configuration of Pseudo-C(2v)-Symmetric Bis-2-Amino-3-Alkanols by CD. Angew. Chem., Int. Ed. Engl. 2000, 39, 4076–4079.
- Pruett, S.T.; Bushnev, A.; Hagedorn, K.; Adiga, M.; Haynes, C.A.; Sullards, M.C.; Liotta, D.C.; Merrill, A.H. Biodiversity of Sphingoid Bases (“sphingosines”) and Related Amino Alcohols. J. Lipid Res. 2008, 49, 1621–1639. [CrossRef]
- Nicholas, G.M.; Hong, T.W.; Molinski, T.F.; Lerch, M.L.; Cancilla, M.T.; Lebrilla, C.B. Oceanapiside, an Antifungal Bis-Alpha,Omega-Amino Alcohol Glycoside from the Marine Sponge Oceanapia Phillipensis. J. Nat. Prod. 1999, 62, 1678–1681. [CrossRef]
- Jin, J.O.; Shastina, V.; Park, J.I.; Han, J.Y.; Makarieva, T.; Fedorov, S.; Rasskazov, V.; Stonik, V.A.; Kwak, J.Y. Differential Induction of Apoptosis of Leukemic Cells by Rhizochalin, Two Headed Sphingolipids from Sponge and Its Derivatives. Biol. Pharm. Bull. 2009, 32, 955–962. [CrossRef]
- Fedorov, S.N.; Makarieva, T.N.; Guzii, A.G.; Shubina, L.K.; Kwak, J.Y.; Stonik, V.A. Marine Two-Headed Sphingolipid-like Compound Rhizochalin Inhibits EGF-Induced Transformation of JB6 P+ Cl41 Cells. Lipids 2009, 44, 777–785. [CrossRef]
- Dyshlovoy, S.A.; Hauschild, J.; Venz, S.; Krisp, C.; Kolbe, K.; Zapf, S.; Heinemann, S.; Fita, K.D.; Shubina, L.K.; Makarieva, T.N.; et al. Rhizochalinin Exhibits Anticancer Activity and Synergizes with EGFR Inhibitors in Glioblastoma In Vitro Models. Mol. Pharm. 2023, 20, 4994–5005. [CrossRef]
- Hara-Chikuma, M.; Verkman, A.S. Physiological Roles of Glycerol-Transporting Aquaporins: The Aquaglyceroporins. Cell. Mol. Life Sci. 2006, 63, 1386–1392. [CrossRef]
- Rojek, A.; Praetorius, J.; Frøkiaer, J.; Nielsen, S.; Fenton, R.A. A Current View of the Mammalian Aquaglyceroporins. Annu. Rev. Physiol. 2008, 70, 301–327. [CrossRef]
- Calamita, G.; Delporte, C. Involvement of Aquaglyceroporins in Energy Metabolism in Health and Disease. Biochim. 2021, 188, 20–34. [CrossRef]
- Sohara, E.; Rai, T.; Miyazaki, J.; Verkman, A.S.; Sasaki, S.; Uchida, S. Defective Water and Glycerol Transport in the Proximal Tubules of AQP7 Knockout Mice. Am. J. Physiol. Renal Physiol. 2005, 289, F1195-1200. [CrossRef]
- Calamita, G.; Perret, J.; Delporte, C. Aquaglyceroporins: Drug Targets for Metabolic Diseases? Front. Physiol. 2018, 9, 851. [CrossRef]
- Pimpão, C.; Wragg, D.; da Silva, I.V.; Casini, A.; Soveral, G. Aquaglyceroporin Modulators as Emergent Pharmacological Molecules for Human Diseases. Front. Mol. Biosci. 2022, 9, 845237. [CrossRef]
- Yang, B.; Kim, J.K.; Verkman, A.S. Comparative Efficacy of HgCl2 with Candidate Aquaporin-1 Inhibitors DMSO, Gold, TEA+ and Acetazolamide. FEBS Lett. 2006, 580, 6679–6684. [CrossRef]
- Martins, A.P.; Marrone, A.; Ciancetta, A.; Galán Cobo, A.; Echevarría, M.; Moura, T.F.; Re, N.; Casini, A.; Soveral, G. Targeting Aquaporin Function: Potent Inhibition of Aquaglyceroporin-3 by a Gold-Based Compound. PLoS One 2012, 7, e37435. [CrossRef]
- Nave, M.; Castro, R.E.; Rodrigues, C.M.; Casini, A.; Soveral, G.; Gaspar, M.M. Nanoformulations of a Potent Copper-Based Aquaporin Inhibitor with Cytotoxic Effect against Cancer Cells. Nanomedicine (Lond.) 2016, 11, 1817–1830. [CrossRef]
- Jelen, S.; Wacker, S.; Aponte-Santamaría, C.; Skott, M.; Rojek, A.; Johanson, U.; Kjellbom, P.; Nielsen, S.; de Groot, B.L.; Rützler, M. Aquaporin-9 Protein Is the Primary Route of Hepatocyte Glycerol Uptake for Glycerol Gluconeogenesis in Mice. J. Biol. Chem. 2011, 286, 44319–44325. [CrossRef]
- Sonntag, Y.; Gena, P.; Maggio, A.; Singh, T.; Artner, I.; Oklinski, M.K.; Johanson, U.; Kjellbom, P.; Nieland, J.D.; Nielsen, S.; et al. Identification and Characterization of Potent and Selective Aquaporin-3 and Aquaporin-7 Inhibitors. J. Biol. Chem. 2019, 294, 7377–7387. [CrossRef]
Figure 1.
Effects of various marine natural products on osmotic water permeability in mouse erythrocytes. (a) Assay method. Mouse erythrocytes were subjected to a 300 mM sucrose gradient. A rapid decrease of the cell volume due to the water efflux through AQPs increased light scattering intensity. AQP inhibitors prevented water efflux and cell shrinkage. (b) Osmotic water permeability (Pf) was measured from the time course of scattered light intensity in response to 300 mM inwardly directed sucrose gradient (control). The initial slopes of scattered light intensity decreased in the presence of dimethylsulfoxide (DMSO, 0.1%) and HgCl2 (0.5 mM). (c) The Pf values calculated from traces in (b). Values are mean ± SD. *P<0.05, **P<0.01. (d) The representative light scattering traces for 24 marine natural compounds (100 nM) in response to 300 mM sucrose.
Figure 1.
Effects of various marine natural products on osmotic water permeability in mouse erythrocytes. (a) Assay method. Mouse erythrocytes were subjected to a 300 mM sucrose gradient. A rapid decrease of the cell volume due to the water efflux through AQPs increased light scattering intensity. AQP inhibitors prevented water efflux and cell shrinkage. (b) Osmotic water permeability (Pf) was measured from the time course of scattered light intensity in response to 300 mM inwardly directed sucrose gradient (control). The initial slopes of scattered light intensity decreased in the presence of dimethylsulfoxide (DMSO, 0.1%) and HgCl2 (0.5 mM). (c) The Pf values calculated from traces in (b). Values are mean ± SD. *P<0.05, **P<0.01. (d) The representative light scattering traces for 24 marine natural compounds (100 nM) in response to 300 mM sucrose.
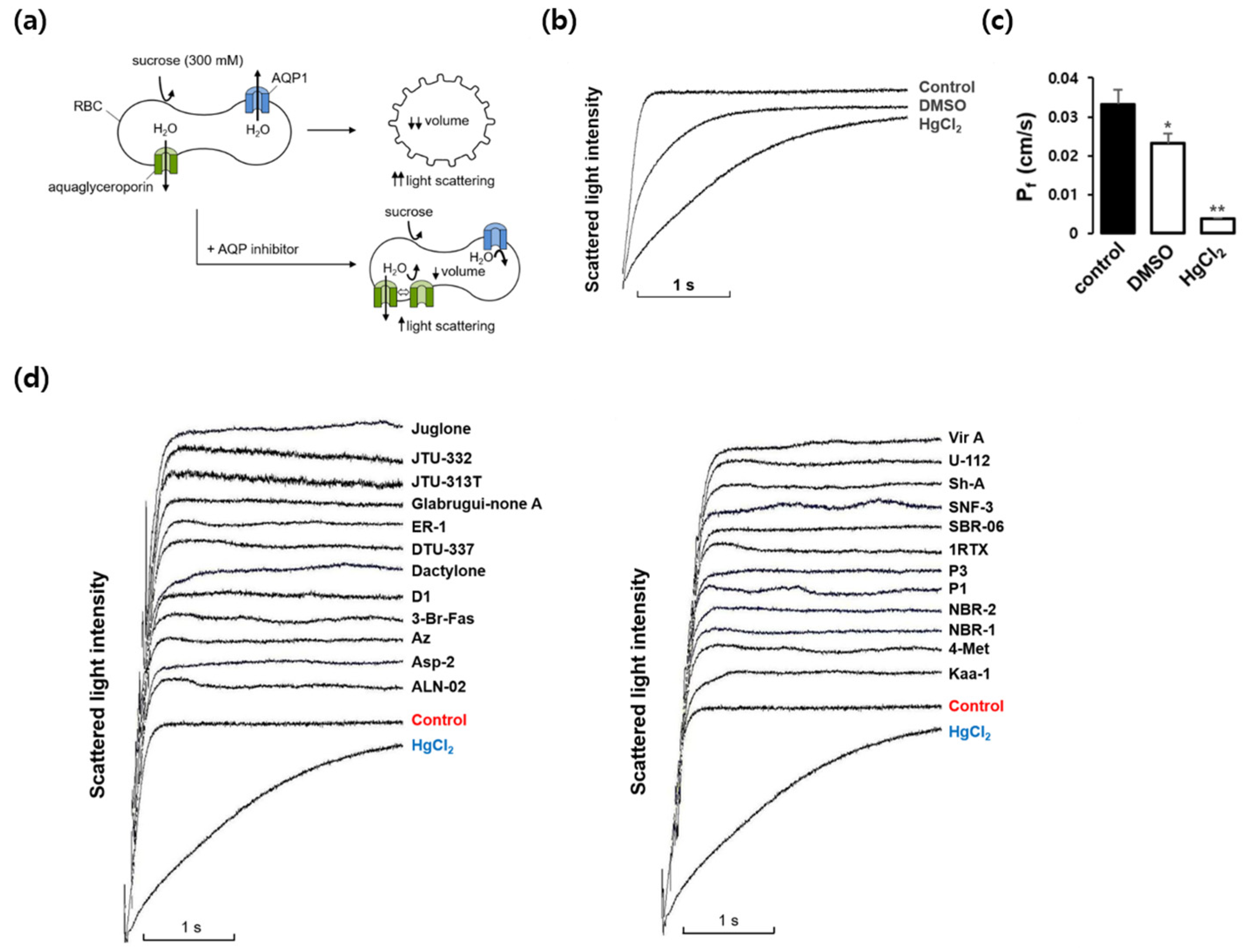
Figure 2.
Effects of various marine natural products on glycerol permeability in mouse erythrocytes. (a) Assay method. Mouse erythrocytes were subjected to a 300 mM glycerol gradient. A rapid decrease of the cell volume (increased light scattering) due to the water efflux through AQPs was followed by cell re-swelling (decreased light scattering) due to glycerol and water influx. Aquaglyceroporin inhibitors prevented the late re-swelling phase. (b) Glycerol permeability was measured from the time course of scattered light intensity in response to 300 mM inwardly directed glycerol gradient (control). HgCl2 (0.5 mM); DMSO, dimethylsulfoxide (0.1%). (c) The representative light scattering traces for 24 marine natural compounds (100 nM) in response to 300 mM glycerol.
Figure 2.
Effects of various marine natural products on glycerol permeability in mouse erythrocytes. (a) Assay method. Mouse erythrocytes were subjected to a 300 mM glycerol gradient. A rapid decrease of the cell volume (increased light scattering) due to the water efflux through AQPs was followed by cell re-swelling (decreased light scattering) due to glycerol and water influx. Aquaglyceroporin inhibitors prevented the late re-swelling phase. (b) Glycerol permeability was measured from the time course of scattered light intensity in response to 300 mM inwardly directed glycerol gradient (control). HgCl2 (0.5 mM); DMSO, dimethylsulfoxide (0.1%). (c) The representative light scattering traces for 24 marine natural compounds (100 nM) in response to 300 mM glycerol.
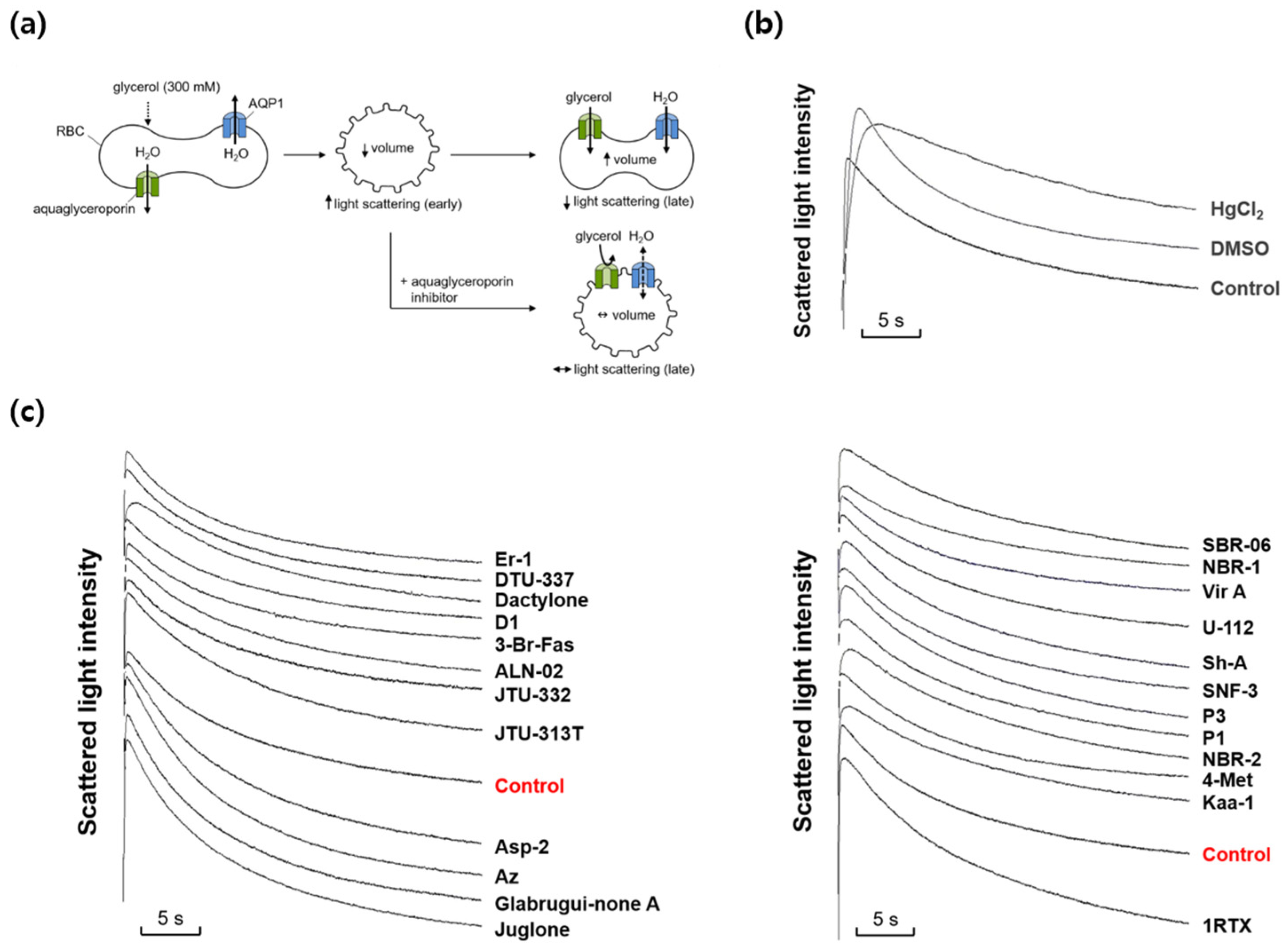
Figure 3.
Effects of stichoposide, rhizochalin, and their derivatives on glycerol permeability in mouse erythrocytes. (a) The representative light scattering traces for stichoposide C (STC) and stichoposide D (STD) in response to 300 mM sucrose. Erythrocytes were incubated with 1 μM of the compounds for 10 min before measurement. (b) The representative light scattering traces for STC and STD in response to 300 mM glycerol. Erythrocytes were incubated with 100 nM of the compounds for 10 min before measurement. (c) The representative light scattering traces for the absence (control) or presence of the indicated concentrations of STC in response to 300 mM glycerol. (d) The representative light scattering traces for rhizochalin (Rhz) and its derivatives in response to 300 mM sucrose. Erythrocytes were incubated with 1 μM of the compounds for 10 min before measurement. AglRhz, aglycon of rhizochalin; AcRhz, peracetyl rhizochalin; AcAglRhz, peracetyl aglycon of rhizochalin; AglRhzA, aglycon of rhizochalin A. (e) The representative light scattering traces for Rhz and its derivatives in response to 300 mM glycerol. Erythrocytes were incubated with 100 nM of the compounds for 10 min before measurement. (f) The representative light scattering traces for the absence (control) or presence of the indicated concentrations of Rhz in response to 300 mM glycerol.
Figure 3.
Effects of stichoposide, rhizochalin, and their derivatives on glycerol permeability in mouse erythrocytes. (a) The representative light scattering traces for stichoposide C (STC) and stichoposide D (STD) in response to 300 mM sucrose. Erythrocytes were incubated with 1 μM of the compounds for 10 min before measurement. (b) The representative light scattering traces for STC and STD in response to 300 mM glycerol. Erythrocytes were incubated with 100 nM of the compounds for 10 min before measurement. (c) The representative light scattering traces for the absence (control) or presence of the indicated concentrations of STC in response to 300 mM glycerol. (d) The representative light scattering traces for rhizochalin (Rhz) and its derivatives in response to 300 mM sucrose. Erythrocytes were incubated with 1 μM of the compounds for 10 min before measurement. AglRhz, aglycon of rhizochalin; AcRhz, peracetyl rhizochalin; AcAglRhz, peracetyl aglycon of rhizochalin; AglRhzA, aglycon of rhizochalin A. (e) The representative light scattering traces for Rhz and its derivatives in response to 300 mM glycerol. Erythrocytes were incubated with 100 nM of the compounds for 10 min before measurement. (f) The representative light scattering traces for the absence (control) or presence of the indicated concentrations of Rhz in response to 300 mM glycerol.
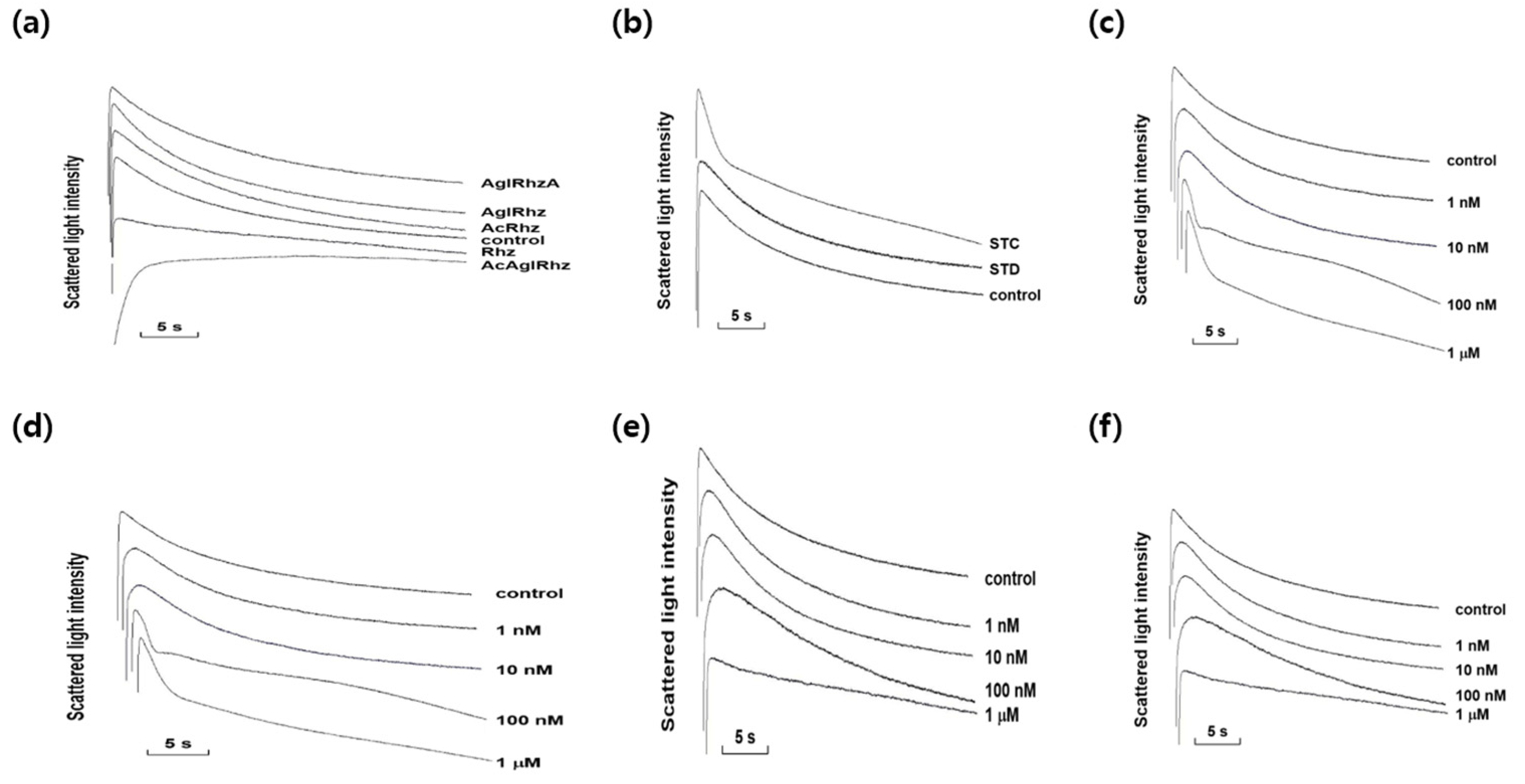
Figure 5.
Expression of AQP subtypes in mouse erythrocytes. Paraffin-embedded sections of formalin-fixed CD1 mouse kidney were prepared to observe erythrocytes in the blood vessels. Immunohistochemistry using anti-AQP1-AQP9 antibodies was performed.
Figure 5.
Expression of AQP subtypes in mouse erythrocytes. Paraffin-embedded sections of formalin-fixed CD1 mouse kidney were prepared to observe erythrocytes in the blood vessels. Immunohistochemistry using anti-AQP1-AQP9 antibodies was performed.
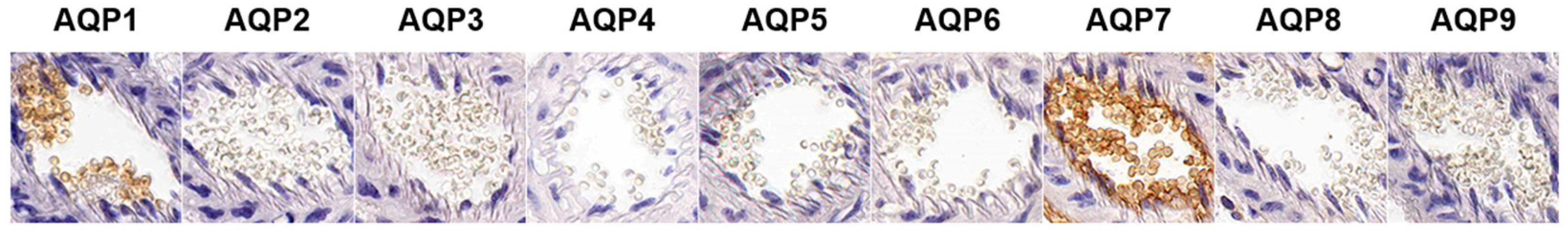
Figure 6.
AQP3-mediated transepithelial glycerol transport in HaCaT cell monolayers. (a) Assay method. YFP-hAQP3 HaCaT cells were grown to a confluence on permeable supports until the TEER values reached 200 Ω.cm2. Cells were exposed to 20 mM glycerol in the lower chamber of the Transwell. The basal-to-apical glycerol transport across the cell monolayer was assessed by measuring the glycerol concentration in the upper chamber as well as inside the cells. (b) Transepithelial glycerol transport across the monolayers of YFP-hAQP1 and YFP-hAQP3 HaCaT cells was assessed by measuring the glycerol concentrations of the upper chamber at the indicated time points after the addition of 20 mM glycerol in the lower chamber. Values are mean ± SD. *P<0.05, **P<0.01. (c) Transepithelial glycerol transport across YFP-hAQP3 HaCaT cell monolayer was assessed by measuring the glycerol concentrations of the upper chamber 10 min after the addition of 20 mM glycerol in the lower chamber. Cells were pretreated with 1 μM of stichoposide C (STC), stichoposide D (STD), rhizochalin (Rhz) or peracetyl aglycon of rhizochalin (AcAglRhz) for 10 min. Values are mean ± SD. *P<0.05, **P<0.01. (d) Transepithelial glycerol transport across YFP-hAQP3 HaCaT cell monolayer was assessed by measuring the glycerol concentrations inside the cells 30 min after the addition of 20 mM glycerol in the lower chamber. Values are mean ± SD. *P<0.05.
Figure 6.
AQP3-mediated transepithelial glycerol transport in HaCaT cell monolayers. (a) Assay method. YFP-hAQP3 HaCaT cells were grown to a confluence on permeable supports until the TEER values reached 200 Ω.cm2. Cells were exposed to 20 mM glycerol in the lower chamber of the Transwell. The basal-to-apical glycerol transport across the cell monolayer was assessed by measuring the glycerol concentration in the upper chamber as well as inside the cells. (b) Transepithelial glycerol transport across the monolayers of YFP-hAQP1 and YFP-hAQP3 HaCaT cells was assessed by measuring the glycerol concentrations of the upper chamber at the indicated time points after the addition of 20 mM glycerol in the lower chamber. Values are mean ± SD. *P<0.05, **P<0.01. (c) Transepithelial glycerol transport across YFP-hAQP3 HaCaT cell monolayer was assessed by measuring the glycerol concentrations of the upper chamber 10 min after the addition of 20 mM glycerol in the lower chamber. Cells were pretreated with 1 μM of stichoposide C (STC), stichoposide D (STD), rhizochalin (Rhz) or peracetyl aglycon of rhizochalin (AcAglRhz) for 10 min. Values are mean ± SD. *P<0.05, **P<0.01. (d) Transepithelial glycerol transport across YFP-hAQP3 HaCaT cell monolayer was assessed by measuring the glycerol concentrations inside the cells 30 min after the addition of 20 mM glycerol in the lower chamber. Values are mean ± SD. *P<0.05.
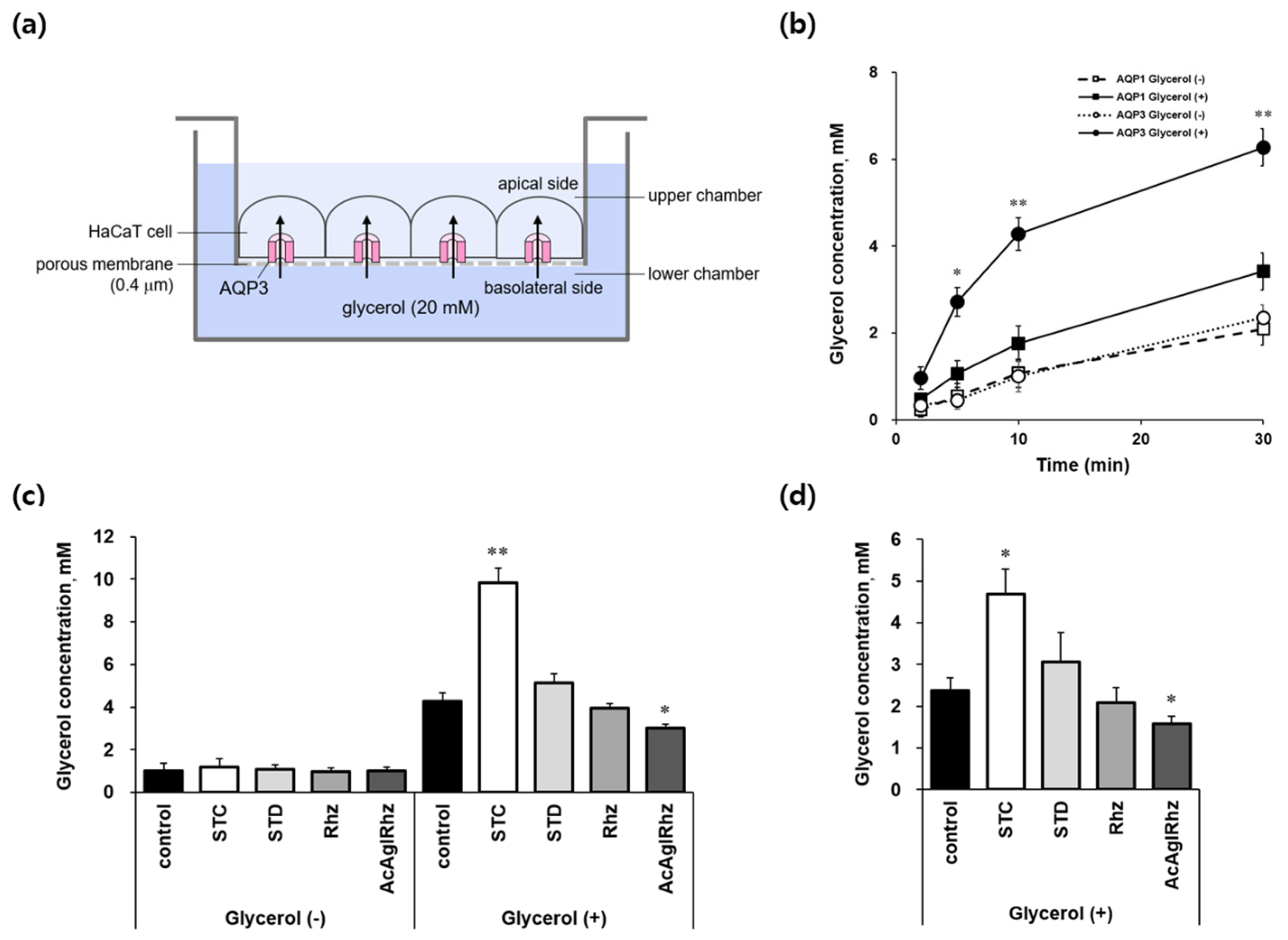
Figure 7.
Structures of Stichoposide, Rhizochalin and their derivatives.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



