
Submitted:
01 July 2024
Posted:
02 July 2024
You are already at the latest version
Abstract
Obesity has increasingly become a worldwide epidemic, as demonstrated by epidemiological and clinical studies. Obesity may lead to development of a broad spectrum of cardiovascular diseases (CVDs), such as coronary heart disease (CHD), hypertension, heart failure (HF), cerebrovascular disease, atrial fibrillation (AF), ventricular arrhythmias, and sudden cardiac death (SCD). In addition to hypertension, there are other cardiometabolic risk factors (CRFs) such as visceral adiposity, dyslipidemia, insulin resistance, diabetes, elevated levels of fibrinogen, and C-reactive protein and others, all of which increase the risk of CVD events. Mechanisms involved between obesity and CVD mainly include insulin resistance, oxidative stress, inflammation, and adipokines dysregulation, which cause maladaptive structural and functional alterations of the heart, particularly left ventricular (LV) remodeling and diastolic dysfunction. Natural products of plants, provide a diversity of nutrients and different bioactive compounds, including phenolics, flavonoids, terpenoids, carotenoids, anthocyanins, vitamins, minerals, fibers, and others, which possess a wide range of biological activities including antihypertensive, antilipidemic, antidiabetic, and other activities, thus conferring cardiometabolic benefits. In this review, we discussed the main therapeutic interventions using extracts from herbs and plants in preclinical and clinical trials with protective properties targeting CRFs. Molecular mechanisms and therapeutic targets of herbs and plants extracts for the prevention and treatment of CRFs are also reviewed.
Keywords:
obesity
; cardiometabolic risk factors
; herbs
; plants
; extracts
; molecular mechanisms
; therapeutic targets
1. Introduction
Obesity is a worldwide public health problem and is a chronic non-transmissible disease whose incidence has been dramatically increasing around the world. This chronic metabolic disease affects distinct age, ethnic and social classes, and has a tremendous impact on the economy and quality of life. Obesity and being overweight are the fifth main causes of deaths globally [1,2]. According to the World Health Organization (WHO), overweight people are defined as having a body mass index (BMI) > 25, while people with a BMI > 30 are considered obese [1]. The pathophysiology of obesity implicates a combination of genetic/epigenetic, nutritional, and environmental factors that promote a chronic positive energy balance and expansion of body fat mass, specially of white adipose tissue (WAT) in visceral fat depots [1,2]. In obese people, WAT plays a key role in secreting lipolysis or lipid synthesis hormones, including inflammatory cytokines that regulate lipid metabolism and promotes a low-grade of chronic inflammation which has the potential to activate insulin resistance and endothelial dysfunction [1,3,4,5]. An excessive accumulation of fat in WAT, in addition to obesity, can also cause metabolic disorders such as dyslipidemia, adipocyte dysfunction, metabolic syndrome, hypertension, type 2 diabetes (T2D), metabolic dysfunction-associated steatotic liver disease (MASLD), cardiovascular disease, and even certain cancers [1,6,7]. In people obese it is presently accepted that oxidative stress is induced by obesity-related diseases such as hyperglycemia, insulin resistance, diabetes, dyslipidemia, atherosclerosis, and inflammation [1,4,8]. Furthermore, adipose tissue inflammation and oxidative stress cause a dysregulation of adipokines secretion, with a reduction in adiponectin and an increase in the secretion of resistin, leptin, and pro-inflammatory adipokines and cytokines, which contribute to cardiovascular stiffness, an impaired vascular relaxation, and finally to cardiac diastolic dysfunction [3]. In addition, the cardiac phenotype in obesity includes concentric left ventricular hypertrophy (LVH), myocardial fibrosis, microvascular dysfunction, arrhythmia, heart failure (HF)-particularly HF with preserved ejection fraction (HFpEF), atrial fibrillation (AF), cardiac remodeling, myocardial infarction, and left ventricular systolic dysfunction, causing deterioration in myocardial function and HF [3,9].
The complex interaction between several cardiometabolic dysfunctions and pathological processes plays an essential role in the pathogenesis of obesity, which is associated with the morbidity and mortality of diabetes and cardiovascular disease [1,4,8,10]. Moreover, cardiovascular diseases are closely associated with high cholesterol, obesity, smoking, diabetes, and a lack of physical activity. Therefore, a convenient lifestyle accompanied by healthy nutrition, reduction of energy-dense food consumption, physical activity and activities that reduce stress oxidative, are the most common therapeutic strategies to reduce cardiometabolic risk factors closely linked to obesity, such as hypertension, dyslipidemia, insulin resistance, diabetes, elevated levels of fibrinogen and C-reactive protein (CRP), including the low-grade chronic inflammation (Figure 1) [1,11].
Although exists several anti-obesity medications approved by the United States (US) Food and Drug Administration (FDA), which include liraglutide (Saxenda), naltrexone-bupropion (Contrave), orlistat (Xenical), phentermine-topiramate (Qsymia), semaglutide (Wegovy), setmelanotide (IMCIVREE), and tirzepatide (Mounjaro) [12,13]; however, the use of these drugs remains controversial as they are associated with a number of adverse side effects and weight regain when the medication is stopped [12,13]. Therefore, the World Health Organization (Committee, 1980) recommended the use of indigenous medicinal plants to treat obesity because of their easy availability, low costs, and relatively fewer side effects. Moreover, herbs and plants contain an unlimited source of phytochemicals, macronutrients, micronutrients, and antioxidants such as polyphenols which are known to prevent diseases associated with oxidative stress such as obesity and its related complications.
In this review article, we discussed the beneficial properties of several herbs and plant extracts (in total 21), as well as its active components against obesity, cardiometabolic risk factors, and associated pathophysiological processes to treat and prevent different cardiovascular diseases in preclinical and clinical trials, considering its molecular mechanisms underlying for their medicinal uses.
2. Phytochemical Constituents and Pharmacological Activities of Herbs and Plants with Cardiovascular Protective Effects
2.1. Allium sativum, Family Alliaceae
Garlic is one of the most well-known herbal medicines in the world and has been used as a spice or medicinal herb for many centuries. The major bioactive compounds of bulbs include sulfur compounds such as alliin, allicin, ajoene, vinyldithiins, diallyl disulfide, allyl methanethiosulfinate, diallyltrisulfide, dimethylmonotohexasulfide, and S- allylcysteine [14]. Garlic administered either in liquid form or capsules, has different antioxidant, anti-diabetic, anti-hypertensive, anti-atherosclerotic, anti-inflammation, endothelial-protecting, lipid-lowering, plasma fibrinogen-lowering, platelet aggregation-inhibiting, fibrinolytic activity-increasing, and other cardiovascular-protective effects [14,15,16]. In addition, aged garlic extract (AGE) has been used in previous human trails and has been shown to be safe [17].
2.2. Andrographis paniculata (Burm. F.) Wall. Ex Nees (Family: Acanthaceae)
Andrographis paniculata (Burm.f.) Nees is considered as a potent plant medicinal in most parts of Asia for the treatment of endocrine disorders, inflammation, and hypertension. Based on phytochemical tests, flavonoids, alkaloids, tannins, triterpenoids, and polyphenols have been isolated from Andrographis paniculata [18]. In addition, andrographolide is a natural diterpenoid lactone extracted from Andrographis paniculata (Burm.f.) Nees, and scientific studies revealed that andrographolide is the main phytoconstituent for its medicinal properties, such as anti-neoplasm anti-bacterial, anti-inflammatory, anti-malaria, antithrombotic, hepato-protective, anti-hypertensive, anti-diabetic, antioxidant, anti-apoptosis, anti-fibrosis, and cardioprotection activities [19].
2.3. Aronia melanocarpa (Michx.) Elliott. (Family: Rosaceae)
Black chokeberry, Aronia melanocarpa (Michx.) Elliot is a deciduous shrub native to eastern North America, and berries Aronia melanocarpa (chokeberry) contain a rich source of biologically active polyphenols such as anthocyanins, proanthocyanidins, and phenolic acid, which have strong antioxidant effects and cardioprotective benefits [20,21]. Other bioactive compounds have been identified present in the fruits and other parts of the plant as neochlorogenic and chlorogenic acids, cyanidin-3-galactoside, cyanidin-3-arabinoside, and (−)-epicatechin [21,22]. Aronia melanocarpa or black chokeberry has been found in multiple clinical trials to combat hyperglycemia-induced oxidative stress, blood pressure (BP), cholesterol and the macrovascular complications of diabetes including cardiovascular disease [21,22] The berries of Aronia melanocarpa also possess therapeutic benefits such as gastroprotective, hepatoprotective, antiproliferative or anti-inflammatory activities [22].
2.4. Camellia sinensis (Family: Theaceae)
Green tea derived from Camellia sinensis leaves, is one of the most popular beverages consumed worldwide. The plant is native to East Asia, possibly originating in southern China, including border areas of Myanmar and India [23]. Green tea extract (GTE) contains several bioactive components, including polyphenols, catechins, theobromine, caffeine, and flavonoids. The major catechins in green tea are (-)-epigallocatechin-3-gallate (EGCG), (-)-epigallocatechin (EGC), (-)-epicatechin-3-gallate (ECG), (-)-epicatechin (EC), and (þ)-catechin (C). Among them, EGCG represents approximately 50-70% of the total catechins from green tea leaves and is primarily responsible for the beneficial effect of green tea [23,24]. The polyphenolic compounds in green tea possess antioxidant properties preventing oxidative stress-caused diseases such as cancer, cardiovascular (e.g., stroke, coronary heart disease, and coronary atherosclerosis) and neurodegenerative diseases. In addition, green tea has beneficial effects on cardiovascular risk factors such as hypertension, lipid disorders, diabetes, endothelial dysfunction, and inflammation. Other beneficial effects include antibacterial, antiviral, antimicrobial, anti-obesity, anti-angiogenic, and anti-metabolic syndrome activities [23,24].
2.5. Caralluma fimbriata (Family: Apocynaceae)
Caralluma fimbriata, an edible succulent and wild medicinal plant growing in dry places, is found throughout Asia (Afghanistan, India, Iran, Pakistan, and Sri Lanka), Africa, Arabian Peninsula, Canary Islands, and Southeast Europe. The key phytochemical constituents of the herb are pregnane glycosides, flavone glycosides, megastigmane glycosides and saponins, including bitter princiaples, triterpenoids, and other flavonoids [25]. The pregnane glycosides, particularly rich in this plant, are known to suppress hunger and increase endurance. In addition, the extract of Caralluma fimbriata has hypoglycaemic, antioxidant, anti-adipogenic, antihypertensive properties [25,26,27]. The herb is also used to treat pain, fever, inflammation, and is commonly consumed by ethnic populations of Central India to manage obesity [25].
2.6. Cinnamomum zeylanicum (Ceylon cinnamon), Family: Lauraceae
Ceylon cinnamon is scientifically known as Cinnamomum zeylanicum Blume. Cinnamon is native to Sri Lanka and is one of the most important spices used daily by people around the world. The most important compounds of cinnamon are cinnamaldehyde and trans-cinnamaldehyde (Cin), which also are found in the essential oil, and both contribute to fragrance and to the various biological activities observed with cinnamon. In addition, this plant contains a variety of resinous compounds, such as cinnamate, cinnamic acid, and numerous essential oils [28]. Cinnamon has many health benefits, including anti-inflammatory, antioxidant, blood-glucose regulation, insulin sensitivity improving, anti-diabetic, lipid-lowering, antimicrobial, anticancer, and anti-cardiovascular properties; cinnamon has also been reported to have benefits against neurological disorders, such as Parkinson’s and Alzheimer’s diseases [28,29,30].
2.7. Citrullus colocynthis (Family: Cucurbitacea)
The Citrullus colocynthis (L.) Schrad is widely distributed in desert areas around the world, including Sudan, Morocco, Arabian desert, Jordan, Tunisia, Iran, India, and Pakistan. Citrullus colocynthis contains several compounds, mainly cucurbitacins and others such as alkaloids, flavonoids, coumarins, steroids, and phenolic acids [31]. The parts of this plant have been used in traditional medicine, and is widely used to treat constipation, mastitis, joint pain, diabetes, hypertension, inflammation, leukaemia, epilepsy, asthma, bronchitis, jaundice, leprosy, rheumatism, common cold, cough, toothache, wounds, and bacterial infection [31,32]. Moreover, in diabetic and nondiabetic animal models, the aqueous extract of Citrullus colocynthis have the hypoglycemic, anti-diabetic, hypolipidemic, and antihyperlipidemic effects, including antiplatelets and profibrinolytic activity [31,32,33]. However, some of the side effects caused by Citrullus colocynthis have been reported, such as nausea, vomiting, colic, diarrhea, hematochezia, and nephrosis [34].
2.8. Cacao (Theobroma cacao L.), Family: Malvaceae
Cacao is native to the jungles of South America and then extended to Mexico. Cocoa beans are the seeds, which are used mainly to produce chocolate, cocoa, and fat. Cocoa is one of the richest sources of polyphenols (about 6-8% by dry weight), which include mainly flavonoids, flavanols, flavanones, isoflavones, and nonflavonoids, as well as catechins, anthocyanidins/anthocyanins, flavonol glycosides, and procynanidins [35]. Black chocolate is considered one of the major sources of antioxidants. Several therapeutic effects have been attributed to cocoa-derived polyphenols, such as improvement of lipid peroxidation, insulin resistance, lipid profile, endothelial dysfunction, postprandial systolic blood pressure (SBP), oxidative stress, and inflammation, including lipid metabolism, and glucose metabolism [35,36,37].
2.9. Corni Fructus (Cornus ofcinalis Sieb. et Zucc.), Family: Cornaceae
Cornus officinalis Siebold et Zuccarini, usually known as Corni Fructus, is a herb and food plant in east Asia and is used in traditional Chinese medicine. Several chemical constituents have been identified in Corni Fructus, which are terpenoids, favonoids, tannins, polysaccharides, phenylpropanoids, sterols, carboxylic acids, furans, saponins, phenolic acid (gallic acid, tannic acid), loganin, and mineral substances. In addition, other phytochemicals are reported in Corni Fructus extracts, such as morroniside, 1,6-α-glucans, loganin, ursolic acid, oleanolic acid, cornuside, polymeric proanthocyanidins, 1,2,3-tri-O-galloyl-beta-D-glucose, 1,2,3,6-tetra-O-galloyl-beta-D-glucose, among others [38,39]. The components in Corni Fructus, such as iridoid glycoside, morroniside, loganin, and polyphenols, exhibit protective effects against hyperglycemia, oxidative stress, and cancer. In vivo and in vitro experimental studies indicate that Corni Fructus has several biological activities, including hypoglycemic, antioxidant, anti-inflammatory, antineoplastic, antimicrobial, anticancer, anti-apoptosis, anti-inflammation, anti-osteoporosis, immunoregulation, neuroprotective, hepatoprotective, nephroprotective, and cardiovascular protection [38,39,40,41]. However, clinical studies are still needed to confirm the reported pharmacological activities.
On the other hand, Corni Fructus has been frequently used for the treatment of asthenia diseases, liver, and kidney diseases, including reproductive system diseases since ancient times. Moreover, it is commonly used for the treatment of several conditions such as diabetes, frequent urination, impotence, and collapse with profuse sweating [38,39].
2.10. Cydonia oblonga Miller (Family: Rosaceae)
Cydonia oblonga Miller (COM) is a plant known by various names, including quince, aiva, bier, and marmelo. The fruit of COM contains various polyphenolic compounds, organic acids, ionone glycosides, and tetracyclic sesterterpenes, including chlorogenic acid, cryptochlorogenic acid, neochlorogenic acid, isochlorogenic acid, quercetin 3-rutinoside, quercetin 3-galactoside, quercetin 3-glucoside, kaempferol 3-glucoside, kaempferol 3-glycoside, and kaempferol 3-rutinoside. In the pulp, leaves, peel, seeds, and complete fruits of COM several citric, ascorbic, malic, oxalic, quinic, fumaric, and shikimic acids have been discovered [42,43]. The fruit of COM is commonly used in the Mediterranean region to prevent or treat obesity. In addition, the fruit of COM has been used for the treatment of hypertension, diabetes, cancer, cardiovascular diseases, respiratory disorders, hemolysis, and ulcers [42,44,45,46]. Several studies have reported the beneficial effects of COM extracts, such as antioxidant, anti-inflammatory, antiallergic, anti-depressant, and antistress, including positive effects on cardiovascular-associated factors such as BP, glucose metabolism, lipid profile, liver dysfunction, and thrombosis [42,45,46]. Moreover, the plant’s seeds have been used to treat diarrhea, dysentery, constipation, cough, sore throat, and bronchitis [42].
2.11. Ginkgo biloba (Family: Ginkgoaceae)
For centuries, the herb Ginkgo biloba has had a long history in traditional Chinese medicine to treat various medical conditions. The extracts of Ginkgo biloba (EGb) leaves have a wide variety of bioactive compounds, such as flavonoid heterosides (between 22% and 27%), represented by flavonol glycosides kaempferol, quercetin, myricetin, apigenin, isorhamnetin, luteolin and tamarixetin), diterpenes, sesquiterpenes, between 5% and 7% of terpene trilactones (ginkgolide A, ginkgolide B, ginkgolide C, ginkgolide J, ginkgolide M, ginkgolide K, ginkgolide L, and bilobalide), 2.8-3.4% correspond to ginkgolides A, B, and C, and 2.6-3.2% consists of bilobalide, phenolic acids, polysaccharides, steroids, and content of less than 5 mg/kg of ginkgolic acids, of which flavonoids and terpene lactones are usually considered to be the responsible for the pharmacological activity associated with this plant [47,48]. For instance, the flavonols of Ginkgo biloba have cardioprotective, antioxidant, antibacterial, and neuroprotective properties. Current pharmacological studies have shown that the flavonoids from Ginkgo biloba have prominent cardioprotective activities, such as regulating blood lipids, lowering blood sugar, inhibiting cardiomyocyte apoptosis, dilating blood vessels, antagonize platelet activating factor, and preventing myocardial ischemic injury, and vascular rupture [47,49,50]. It is important to note that many types of preparations based on Ginkgo biloba extract have been developed for the treatment of cardiovascular diseases. Ginkgo biloba is also used for the prevention and treatment of hypertension, atherosclerosis, peripheral arterial disease, peripheral venous disease, Raynaud’s phenomenon, and erectile dysfunction. The plant has also been used for diseases such as cognitive decline, dementia, and tinnitus [47,49,51,52].
2.12. Coffea (genus Coffea), Family: Rubiaceae
Coffee is widely consumed in the world and has a variety of phytochemicals. The main coffee polyphenol includes the glycosylated derivate forms of the polyphenol and chlorogenic acids (CGAs), such as esters of caffeic acid and quinic acid. Green coffee is raw coffee beans that have not been roasted, and it is rich in bioactive phytochemical compounds, mainly CGAs, caffeine, and soluble fiber (mostly galactomannans and arabinogalactan) [53,54]. Green coffee bean extract (GCBE) has antioxidant properties and neutralizes reactive oxygen species. In addition, studies have found that the CGA from GCBE, regulates vasoreactivity and glucose metabolism, including properties such as anti-cancer, anti-inflammatory, anti-lipidemic, anti-hypertensive, and anti-diabetic [55,56,57]. With respect to hypolipidemic effects, GCBE and its CGA reduce triglyceride (TG) and total cholesterol (TC) levels; however, the effects on high- and low-density lipoprotein cholesterol (LDL-C) levels are inconsistent. Some studies reported an increase in serum high-density lipoprotein cholesterol (HDL-C) after GCBE intake, while others reported non-significant results [58].
2.13. Hibiscus sabdariffa (Roselle), Family: Malvaceae
Hibiscus sabdariffa Linn is commonly known as roselle, which probably originated in West Africa and grows in the tropical and subtropical regions. Roselle contains several bioactive compounds, which has medicinal properties, such as phenolic acids (protocatechuic, chlorogenic caffeic acid , and gallic acids), flavonoids (quercetin-3-glucoside, methyl epigallocatechin, myricetin, quercetin, rutin, and kaempferol), anthocyanins (delphinidin-3-sambubioside and cyanidin-3-sambubioside), and organic acids (hibiscus acid, citric acid, hydroxycitric acid, malic acid, and tartaric acid), which are responsible for many biological activities [59,60]. This plant is commonly used as a traditional drink material and folk medicine against hypertension, pyrexia, liver disease, fever, inflammation, kidney and urinary bladder stones, and obesity. Roselle, mainly its calyx, has phytochemicals with various health benefits, such as anti-hyperglycemic, anti-hyperlipidemic, anti-hypertensive, antioxidative, anti-inflammatory, and anti-fibrosis effects [59,60,61,62]. Roselle water extracts also show anti-cancer, antibacterial, nephro- and hepato-protective, renal/diuretic effect, anti-cholesterol, and anti-diabetic effects among others; this might be related to inhibition of α-glucosidase and α-amylase, inhibition of angiotensin-converting enzymes (ACE), including the direct vasorelaxant effect or calcium channel modulation [59,60,63,64]. Besides, Hibiscus sabdariffa relaxes other smooth muscles, including the intestine, uterus, and bladder [60].
2.14. Ilex paraguariensis A.St.-Hil. (Mate), Family: Aquifoliaceae
Ilex paraguariensis, commonly known as yerba mate, is one of the most widely consumed plants in subtropical regions of South America (Brazil, Paraguay, Uruguay, and Argentina). This tree or shrub contains polyphenols derived from caffeoyl, manly monocaffeoyl quinic isomers (3-O-caffeoyl quinic or neochlorogenic acid, 5-O-caffeoyl quinic or chlorogenic acid and 4-O-caffeoyl quinic or cryptochlorogenic acid), caffeic acid, and dicaffeoyl quinic isomers (3,4-dicaffeoylquinic acid, 3,5-dicaffeoylquinic acid, and 4,5-dicaffeoylquinic acid), methylxanthines (caffeine, theophylline, and theobromine), flavonoids (quercetin, kaempferol, and rutin), tannins, and numerous triterpenic saponins that are derived from ursolic acid and are named as metesaponins [65,66]. Yerba mate exhibits various biological activities such as antioxidant, anti-inflammatory, anti-obesity, anticancer, immunomodulatory, improvement of glycemic and lipid metabolism, reversion of insulin resistance, inhibition of glycation and atherosclerosis, thermogenic and vasodilatation effects, a protective effect against induced DNA damage, and reduction of cardiovascular risk [65,66,67,68,69,70]. Moreover, yerba mate facilitates recovery from physical and mental fatigue, reduces the feeling of hunger, and works as a diuretic; and aqueous extract of this medicinal plant protects the myocardium against ischemia-reperfusion injury and decreases oxidative damage, which can be attributed to the potent antioxidant properties of the extract [65,71].
2.15. Moringa oleifera Lam., Family: Moringaceae
Moringa oleifera Lam. is native to the sub-Himalayan northern parts of India and commonly cultivated throughout tropical and sub-tropical countries. Moringa leaves are rich in many nutritious and bioactive compounds, including carotenoids, polyphenols, glucosinolates (the most abundant of them is 4-O-(α-l-rhamnopyranosyl-oxy)-benzylglucosinolate or also named glucomoringin), tannins, among others. With respect to polyphenolic compounds, are represented by flavonoids (mostly quercetin and kaempferol, in their as 3′-O-glycoside forms) and phenolic acids such as gallic, chlorogenic, which is an ester of dihydrocinnamic acid (caffeic acid), ellagic, quinic, and ferulic acids) [72,73]. The following compounds have hypotensive properties, such as sothiocyanates, thiocyanates, and nitriles, which are formed by enzymatic hydrolysis of the glucosinolates; and niaziminin also hypotensive, is a mustard oil glycoside isolated along with other glycosides (niazinin and niazimicin) from ethanolic extracts of Moringa oleifera leaves. The flavonol quercetin is a potent antioxidant and is found at concentrations as high as 100 mg/100 g of dried Moringa oleifera leaves [72,73]. The bioactive compounds of moringa oleifera are accountable for many medicinal properties such as, cholesterol-lowering, anti-obesity, antihyperlipidemic, anti-diabetic, antihypertensive, neuroprotective, anti-asthmatic, antitumor, anti-inflammatory, antioxidant, antipyretic, antiepileptic, antiulcer, antispasmodic, diuretic, hepatoprotective, antiviral, antimicrobial, antifungal, cardioprotective activity, protection against signs of aging, typhoid fever, malaria, diarrhea, and dysentery [72,73,74,75,76,77].
2.16. Nigella sativa, Family: Ranunculaceae
This medicinal plant is popularly known as black seed or black cumin, and is mainly distributed in North Africa, the Middle East, Europe, and Asia. The major phytochemical constituent of the seeds from Nigella sativa is thymoquinone (particularly the essential oil), also include phytosterols (β-sitosterol and stigmasterol), alkaloids (e.g., nigellamines), saponins, dithymoquinone, nigellin, terpenes and terpenoids (such as thymoquinone and its derivatives), tocopherols, polyphenols (such as quercitrin and kaempferol), and miscellaneous components [78,79]. These bioactive components of the seeds are responsible for the pleiotropic pharmacological properties, such as antioxidant, anti-inflammatory, anti-hypertensive, anti-hepatotoxic, anti-cancer, hypoglycemic, anti-microbial, antifungal, anti-nephrotoxic, anti-hepatotoxic, lipid-lowering properties, and immunostimulating activities. The seeds of Nigella sativa are also used for the treatment of cardiovascular diseases, respiratory diseases (asthma and bronchitis), cough, headache, rheumatic disorders, fever, influenza, obesity, epilepsy, back pain, and gastrointestinal disorders (indigestion and diarrhea) as well as in cases of amenorrhea, dysmenorrhea, and skin infections [78,79,80].
2.17. Opuntia ficus indica, Family: Cactaceae
The species of genus Opuntia (approximately 200), grow extensively in desert or semi-desert regions in Mexico, the United States, and Mediterranean countries, among other countries. This plant is native to Mexico and is known as nopal, prickly-pear cactus in the Southern United States, and Indian fig cactus in Europe. Cladodes of opuntia ficus indica provides dietary fiber and bioactive compounds such as carotenoids (lutein, β-carotene, and β-cryptoxanthin), flavonoids (isorhamnetin-3-O-glucoside, kaempferol, quercetin, isoquercetin, nicotiflorin, and rutin), and phenolic compounds (coumaric callic acid, and 3,4-dihydroxybenzoic 4-hydroxybenzoic, and ferulic acid). Moreover, cladodes are rich in pectin, mucilage, minerals, malic acid, vitamins, and antioxidants. Meanwhile, prickly pear fruits contain bioactive compounds such as pigments (betaxanthins, betacyanins, and betalains) and flavonoids (kaempferol, quercetin, and isorhamnetin) [81,82]. Opuntia ficus indica has actions against atherosclerotic cardiovascular diseases, diabetes, obesity, hypertension, asthma, burns, edema, and indigestion, as well as other pharmacological effects including the antioxidant, neuroprotective, anti-inflammatory, antihypercholesterolemic, anti-ulcer, antimicrobial, antiviral potential, wound healing, skin protective, hepatoprotective, anticancer, effects on human infertility, and chemopreventive capacity. Moreover, opuntia ficus indica has effects on the bone health, kidneys, and gastrointestinal tract, including gastroprotective, sedative, analgesic, anxiolytic, cognitive and memory effects [81,82,83,84,85].
2.18. Platycodon grandiflorus, Family: Campanulaceae
Platycodon grandiflorus, a common Chinese herb, is mainly distributed in Northeast Asia, including China, Korean Peninsula, Japan, and Siberia, where it has been used for decades as a traditional medicinal herb. A phytochemical investigation revealed that platycodon grandiflorus contains at least 100 compounds, including steroidal saponins, flavonoids, polyacetylenes, sterols (e.g., stigmasterol), phenolic acids, and other bioactive compounds; among which, the saponins are considered the main active compounds [86,87]. Platycodon grandiflorus exhibit diverse pharmacological activities, such as antioxidant, anti-inflammatory, anti-tumor, anti-diabetic, anti-obesity, antibacterial, anti-apoptosis, hypocholesterolemic, hypoglycemic, immune enhancement, liver protection, improve insulin resistance and the lipid profile, decrease BP, alleviation of atopic dermatitis, as well as has relieving cough and asthma activities, apophlegmatic, antitussive, and cardiovascular system activities [86,87,88,89,90,91]. Platycodon grandiflorus was also reported to be used for the treatments of chest congestion, chest distress, diphtheria, dyspnea, mastitis, measles, dermatitis, dysentery, suppuration, chronic rhinitis, chronic tonsillitis, bronchitis, asthma, pulmonary abscesses, pulmonary tuberculosis, faucitis, bronchial asthma, and other conditions [86,87].
2.19. Punica granatum L., Family: Lythraceae
Punica granatum Linn., commonly known as pomegranate, is a small shrub with tasty fruit native to the Middle East, growing in subtropical and temperate regions and having a variety of planting distributions around the world. More than 60 bioactive components have been identified in pomegranate, which are categorized as phenols, flavonoids, triterpenes, alkaloids, sterols, vitamins, and unsaturated fatty acids. In addition, pomegranates are rich in polyphenolic antioxidants, such as tannins, anthocyanin, and flavonoids; these active components being the most abundant in the pomegranate. The main compounds isolated from pomegranate flowers are polyphenols, flavonoids, terpenoids, and triterpenoids, such as leanolic acid and ursolic acid. The pomegranate fruit includes hydrolyzable tannins like gallotannins and ellagitannins, as well as ellagic acid and its derivatives, gallic acid, anthocyanins, proanthocyanidins, flavonoids, sterols, lignans, terpenes, and terpenoids. Pomegranate peel is abundant in a variety of phenolics, ellagitannins, proanthocyanidins, microelements, and flavonoids, including kaempferol-3-O-glucoside. Punica granatum bark is rich in tannins, proanthocyanidins, anthocyanins, and terpenoids. Pomegranate juice is rich in antioxidants, such as polyphenols, flavonoids, ellagitannins, tannins, and anthocyanins [92,93,94]. In vivo and in vitro studies have been shown that extracts of different pomegranate fractions (peels, flowers, seeds, and juice) improve lipid metabolism in diseases such as atherosclerosis, metabolic dysfunction-associated steatotic liver disease (MASLD), metabolic syndrome, and type 2 diabetes, including a wide range of diseases, such as inflammation, Alzheimer’s disease, ulcers, diarrhea, erectile dysfunction, obesity, cancer, brain ischaemia, fibrosis, fungal and microbial infections. Pomegranate flowers are used for the treatment of cardiovascular disorders, diabetes, obesity, and some microbial infections (Salmonella entteriditis and Kentucky). Pomegranate seeds are used to treat heart diseases, diabetes, cancer, obesity, urinary disorders, and to prevent miscarriage and to improve male fertility. Additionally, pomegranate seeds have antimicrobial and antioxidant properties. Pomegranate peel extracts are traditionally used to treat diarrhea and ulcers. Other pharmaceutical properties reported in the pomegranate peels include anti-proliferative, anti-inflammatory, antioxidant, and anti-cancerous effects. Punica granatum bark has been used traditionally for the treatment of inflammation, diarrhea, malaria, nose bleeding, sore throat, ulcer, and hoarseness. Pomegranate juice has important biological actions, including antioxidant activity and cardiovascular protection. Moreover, consumption of pomegranate can relieve dental infections and menopausal symptoms, as well as improve the intestinal microbiota, thus preventing obesity and diabetes [92,93,94,95,96,97].
2.20. Salvia miltiorrhiza Bunge, Family: Lamiaceae
Salvia miltiorrhiza Bunge, commonly called danshen, is a perennial herb used in traditional Chinese medicine. Salvia miltiorrhiza contains more than 100 compounds, including salvianolic acid A/B/C/D/E/F/G, lithospermic acid, danshensu, caffeic acid, and rosmarinic acid, tanshinone I/IIA/IIB/V/VI, tanshindiol A, cryptotanshinone, dihydrotanshinone І, miltirone, dehydro miltirone, and isotanshinone, among others. The bioactive compounds in salvia miltiorrhiza extract are classified into two major groups, water-soluble phenolics (salvianolic acid and comfrey acid) and liposoluble tanshinones (diterpenoids), which are responsible for the main pharmacological properties of salvia miltiorrhiza [98,99]. Tanshinone IIA and salvianolate have various cardiovascular and pharmacological effects, including antioxidative, anti-inflammatory, endothelial protective, anticoagulation, vasodilation, myocardial protective, anticoagulation, vasodilation, and anti-atherosclerosis, as well as effects on reducing the proliferation and migration of vascular smooth muscle cells. Besides, salvianolates are composed of salvianolic acid B, rosmarinic acid, and lithospermic acid, which are widely used in the treatment of coronary heart disease. Meanwhile, tanshinones are more effective against the treatment of cardiovascular diseases and cerebrovascular diseases, including atherosclerosis, myocardial infarction, cardiac hypertrophy, myocardial ischemia reperfusion (I/R), and chronic heart failure. Salvia miltiorrhiza has other effects such as anti-diabetic, anti-inflammation, antioxidant, anti-fibrosis, and anti-apoptosis. Salvia miltiorrhiza is also used to treat malignant tumors, neurological, lung diseases, inflammatory diseases, gynecological diseases, liver diseases, renal diseases, and metabolic disorders such as atherosclerosis, hyperlipidemia, obesity, and other dyslipidemia-related diseases [98,99,100,101].
2.21. Taraxacum officinale L. (Dandelion), Family: Asteraceae
Taraxacum officinale L., also known as dandelion, a perennial herb and commonly regarded as a weed, is native to Eurasia and grows in America, Africa, New Zealand, and Australia. Dandelion has phenolic acids (chlorogenic acid, chicoric acid), flavonoids (luteolin derivatives, quercetin), and terpenes (sesquiterpene lactones). The leaves containing bitter sesquiterpene lactones (taraxinic acid and triterpenoids such as cycloartenol), while the roots have phenolic acids, inulin, sesquiterpene lactones, triterpenes, and sterols (taraxasterol, taraxerol, cycloartenol, beta-sitosterol, stigmasterol), and the compounds already mentioned, which contribute to its therapeutic properties [102,103]. Dandelion has been used as a phytomedicine for its holeretic, antirhemetic, diuretic, antibacterial, hypolipidemic, hypoglycemic, antithrombotic, anti-inflammatory, anti-obesity, antioxidant, and antiplatelet effects, as well its use against cancer and cardiovascular ailments. Moreover, dandelion is used as a remedy in kidney diseases, liver, kidney, and spleen disorders. Dandelion has high levels of phenolic acids, with antioxidant effects; coumarins with anti-inflammatory, anticancer, antibacterial, and antithrombotic properties; sesquiterpene lactones with anti-inflammatory and antibacterial effects; and triterpenes or phytosterols, which possess anti-atherosclerotic effects. Dandelion leaves and flowers contain polyphenols, predominantly hydroxycinnamic acid derivatives and flavonoids (apigenin and luteolin derivatives), all of which have antioxidant and hypocholesterolemic properties. Dandelion roots are rich in inulin, which has a hypoglycemic, probiotic, and immune-boosting effect; meanwhile, its phytochemicals such as phenolic acids and sesquiterpene lactones are responsible for its anti-diabetic properties. In general, bioactive compounds from dandelion roots possess bifidogenic, anti-inflammatory, and antifibrotic activities [102,103,104,105,106,107].
3. Pathological Processes Involved in Obesity
Overweight and obesity are increasingly common conditions in the world, due to the intake of calorie-dense foods and relatively inactive lifestyles, which create long-term imbalances between energy uptake and expenditure, and these conditions promote the deposition of fat mass in the body’s WAT, leading to phenotypic changes in this tissue such as adipocyte hypertrophy (cell size increase) and subsequently hyperplasia (cell number increase) [1,3,108]. In obesity, the hypertrophied WAT visceral adipocytes show lipolysis activation, leading to high levels of circulating non-esterified fatty acids (NEFAs) [109]. NEFAs in normal conditions are catabolized by the β-oxidation to provide energy tissues such as liver and muscle; however, in high concentrations they contribute to the development of insulin resistance [110]. Furthermore, hypertrophic visceral adipocytes contribute to elevated circulating triacylglycerol (TAG) levels mainly from de novo lipogenesis, in which fatty acids (FAs) are synthetized from carbohydrates or FAs are provided from chylomicrons and very-low-density lipoproteins (VLDL) [111]. Several studies suggest that oxidative stress plays a fundamental role as a factor linking obesity and its related complications. Furthermore, oxidative stress can induce an increase in preadipocyte proliferation, adipocyte differentiation, and the size of mature adipocytes. Obesity per se can also induce systemic oxidative stress through superoxide generation from NADPH oxidases, oxidative phosphorylation, protein kinase C activation (PKC), glyceraldehyde auto-oxidation, and polyol and hexosamine pathways. Besides, elevated plasma free FAs promote the generation of superoxide radicals; and hyperleptinemia, low antioxidant defense, chronic inflammation, and postprandial reactive oxygen species generation are other factors that also contribute to oxidative stress in obesity. Obesity-associated oxidative stress induces various pathological events, including insulin resistance and diabetes, liver failure, cardiovascular complications, sleep disorders, and asthma, including reproductive, oncological, and rheumatological problems [1,3,112]. Adipose tissue produces several adipokines including cytokines and hormones, which regulate energy homeostasis, glucose and lipid metabolism and cardiovascular functions [113]. Because obesity is directly associated to chronic inflammation, during the hypertrophy and hyperplasia of adipose tissue leads to the organ’s dysfunction and development of a pro-inflammatory microenvironment, the signaling pathway of NF-κB is activated and increases the expression of interleukin-6 (IL-6), tumor necrosis factor-α (TNF-α), and IL-1β through the TLR4/MyD88 signaling pathway; other molecules increased with excessive adiposity are leptin, IL-8, CRP, PAI-1, heptoglobin, angiotensinogen, inducible nitric oxide synthase, platelet-activating factor (PAF) and chemokines, such as monocyte chemotactic protein 1 (MCP1), which promotes migration of macrophages into the adipose tissue and induces the release of cytokines. Furthermore, in obese subjects, the levels of interleukin 10 (IL-10) are decreased, which worsens the metabolic profile due to IL-10 inhibiting the synthesis of pro-inflammatory cytokines [3,19,112,114,115]. The activation of NF-κB, TLR4 and mTOR lead to the attenuation of insulin signaling and insulin resistance in several tissues, which contribute to the development of obesity-related complications, including diabetes and atherosclerosis [116,117]. Platelets are key players in the thrombotic processes, and various platelet markers have been reported elevated in obese and T2D individuals, including the mean platelet volume, circulating levels of platelet microparticles, oxidation products, platelet-derived soluble P-selectin, and CD40L. Therefore, these markers contribute to an intersection between obesity, inflammation, and thrombotic phenotype [118]. Obesity is an altered health condition with changes in gut microbiota due to the consumption of improper diet, which affects the health status of the host. Gut microbiota plays an important role in energy balance, intestinal integrity, and immunity against invading pathogens. Approximately 100 trillion microbes colonize the human gut, which are represented by microorganisms such as bacteria, archaea, fungi, protozoa, and viruses. Gut microbiota is represented by the phyla Bacteroidetes, Firmicutes, Proteobacteria, Actinobacteria, and Verrucomicrobia, and approximately 90% of the total bacterial species belong to Bacteroidetes and Firmicutes. Obese individuals exhibited reduced proportion of Bacteroidetes and elevated level of Actinobacteria with no significant difference in Firmicutes, thus an increased Firmicutes/Bacteroidetes ratio is a biomarker of obesity susceptibility [119,120].
3.1. Plant and Herb Extracts with Anti-Obesity Activity
Previous studies have shown the nutritional and beneficial effects on metabolism of plants and herbs consumption, showing a wide range of bioactive compounds, many of them with different properties for healthy humans. There are several papers in which the use of various medicinal plants and herbs to treat obesity has been investigated. For instance, green tea is associated with beneficial health effects due to its body fat-reducing and hypocholesterolemic properties [121,122], and green tea aqueous extract (GTAE, 1.1% and 2%) administered in rats fed with a high-fat diet, lowed atherogenic index, reduced body weight gain (only with 2% of GTAE), and prevented visceral fat accumulation [123]. The visceral weight loss and improvement of lipid profile with green tea in rats may be due to increased thermogenesis and fat oxidation [124]. The principles present in the consumed Indian food plant, caralluma fimbriata extract (CFE) suppress appetite, and provide antiobesogenic and metabolic effects. In a Diet-Induced Obesity (DIO) rat model, CFE showed dose-dependent appetite suppressant, prevented liver weight and fat pad mass, and protected against atherogenesis in rats fed a cafeteria diet [125]. In overweight adults, CFE preserved body weight, decreased waist circumference, and reduced daily caloric intake over a 16-week period in overweight patients compared to a placebo. The mechanism for appetite suppression by CFE includes the reduction of ghrelin synthesis in the stomach and neuropeptide Y in the hypothalamus [26]. Bioactive compounds derived from cacao, which are rich in polyphenols (flavonoids), are beneficial against overweight and lipid disorders. In an obesity model induced by high fat diet (HFD) and fructose ingestion in rats, the cocoa extracts made with outer pod husk and kernel husk decreased the weight gain, reduced the SBP, and improved the lipid profile [37]. Ginkgo biloba extract (GBE), which contains a mixture of polyphenols with antioxidant properties, has several potentially beneficial effects, such as reduces food and energy intake, reduces body adiposity, improvement in insulin sensitivity, enhances insulin receptor and AKT phosphorylation, and reduces NF-κB p65 phosphorylation in retroperitoneal adipose tissue of obese rats, and reduction of weight gain in models of obesity induced by diet and ovariectomy [126,127,128]. These findings were confirmed in HFD induced obese male rats, GBE supplementation reduced energy intake and epididymal adipocyte volume [129]. Green coffee beans contain phytochemicals with beneficial effects on cardiometabolic disorders. The 3-caffeoylquinic acid (3-CQA) in green coffee bean extract (GCBE) was evaluated in HFD-induced obese mice, and this extract decreased body weight gain, liver weight and WAT weights, regulating adipogenesis and lipid metabolism-linked genes and proteins in WAT and liver [130]. In addition, in male albino Wistar rats fed with HFD, and intervened with GCBE when obesity was established, the extract decreased the body and organ weights, reduced TC, TG, LDL-C, VLDL, glucose, and insulin levels. GCBE also exhibited an increase of adiponectin levels, and decreased the expression of RBP4, whereas an increase in GLUT4 expression was observed in the adipose tissue [131]. Moringa oleifera leaf extracts (MOLE) possess anti-obesity effects in experimental animal models and were tested in HFD-induced obesity in rats. Thirteen metabolites were identified in MOLE, including flavonols, flavones and a phenolic acid. MOLE reduced weight gain and adiposity index, including glucose, insulin and HOMA-IR, and Revised Quantitative Insulin Sensitivity Check Index (R-QUICKI) was significantly increased by MOLE. In visceral fat mass, MOLE significantly reduced the levels of leptin and vaspin; meanwhile, diponectin, omentin and GLUT-4 expression were increased. MOLE significantly inhibited FAS and HMG-CoA reductase and elevated mRNA expression of MC4R and PPAR-α. In obese patients, the administration of Moringa oleifera hard gelatin capsules showed significant reduction of the average BMI, TC and LDL-C [74]. Previous studies have been showed that Ilex paraguariensis (yerba mate) aqueous extracts inhibited the progression of atherosclerosis and decreased the body weight, visceral fat, serum lipids, glucose, leptin, and insulin levels in HFD-fed-rats [132,133]. The treatment with Ilex paraguariensis extracts (IPE) in C57BL/6J mice fed a HFD, also reduced the accumulation of lipids in adipocytes, body weight gain, and obesity. Besides, the extract reduced serum cholesterol, serum TG, and glucose levels [69]. These findings were confirmed in another study, in which Yerba mate extract was also evaluated in mice fed with a HFD, and obese mice treated with yerba mate exhibited marked attenuation of weight gain, adiposity, a reduce in epididymal fat pad weight, and restoration of the serum levels of cholesterol, TGs, LDL-C, and glucose [68]. In obese rats primed by early weaning, IPE reversed abdominal obesity, leptin resistance and hypertriglyceridemia [134]. The chlorogenic acid in yerba mate is mainly responsible for these effects [135]. In another study Ilex paraguariensis was investigated in Korean subjects with obesity. Individuals with obesity were given oral supplements of Yerba Mate capsules, and this supplementation decreased body fat mass, percent body fat, and waist-hip ratio, which suggest that Yerba Mate supplementation may be an alternative for treating obese patients [136]. Platycodon grandiflorus root extract (PGE) was analyzed in obese Korean adults and revealed a significant decrease in body fat mass and body fat percentage, suggesting anti-obesogenic effects in overweight or obese adult humans [137]. Pomegranate extract (PomE) is rich in punicalagins and increases markers related to browning and thermogenesis in human differentiated adipocytes [138]. In addition, in preclinical study of HFD-induced obesity, pomegranate extract (PomE) increases the systemic energy expenditure, thus contributing to a reduction in the low grade of chronic inflammation and insulin resistance associated with obesity [95].
Oxidative stress results from elevated production of free radicals along with reduced levels of antioxidants, and plays an important role in cardiovascular diseases, including atherosclerosis and coronary artery disease [112]. CFE improves HFD-induced cardiac damage through reducing cardiac lipids such as total lipids, TG, TC, and free fatty acids (FFAs). Furthermore, CFE improves the activities of antioxidant enzymes, such as glutathione peroxidase (GPx), glutathione reductase (GR), glutathione peroxidase (GPx), catalase (CAT), superoxide dismutase (SOD), and glutathione-s-transferase (GST) [27]. Moringa oleifera leaf extracts (MOLE) were evaluated in HFD-induced obesity and cardiac damage in rats. The activities of antioxidant enzymes were decreased in animals which received a HFD; however, these antioxidant enzymes were significantly, and dose-dependently, enhanced by administration with MOLE [139].
Several diseases associated with obesity such as dyslipidemia, T2D, and cardiovascular disease, are closely related to low-grade inflammation. In WAT from obese mice, TNF-α, IL-6, leptin, CCR2, CCL2, and PAI-1 genes are upregulated. However, yerba mate extract administration decreased the expression of these genes [68]. In addition, yerba mate extract intake blunted the proinflammatory effects of HFD-induced obesity in rats through the phosphorylation of hypothalamic IKK and NF-κB p65 expression and increasing the protein levels of IκBα, adiponectin receptor-1, and IRS-2 [140]. Andrographis paniculata extract (APE) and its bioactive constituent andrographolide are known to possess anti-inflammatory and anti-apoptotic effects. APE was analyzed in myocardial tissue from HFD-induced obese mice. The animals fed with HFD developed myocardial inflammation, which potentially contributed to cardiac hypertrophy and myocardial apoptosis, but APE showed significant inhibition of these effects in obese mice [141].
Aronia melanocarpa contains a high content of procyanidins and anthocyanins. It was found that aronia melanocarpa extract significantly inhibits the amidolytic activity of thrombin and plasmin, the latter being the main fibrinolytic enzyme [142]. Moreover, patients with metabolic syndrome showed a significant reduction in the levels of TC, LDL-C, and TG, as well as an improvement in platelet aggregation, clotting, and fibrinolysis after aronia melanocarpa extract supplementation [143]. The effect of Citrullus colocynthis was investigated on blood haemostasis in HFD-induced obese rats, and was found that Citrullus colocynthis reversed HFD induced increases in fibrinogen and von Willebrand factor, thus Citrullus colocynthis has antiplatelets and profibrinolytic properties due to its potent hypoglycaemic and hypolipidaemic effect, its ability to reduced levels of circulatory TNF-α and IL-6 and its ability to lower the prothrombic leptin levels and elevate the antithrombic adiponectin levels [33]. Cydonia oblonga is traditionally used in Uyghur medicine to prevent cardiovascular diseases. Cydonia oblonga extract (COE) was explored on models (mice and rats) and markers of thrombosis. COE dose-dependently prolonged bleeding and the clotting time. Besides, COE reduced pulmonary embolism mortality, and dose-dependently increased thrombolysis, and reduced TXB2. Therefore, COE has an anti-thrombotic effect, probably at least in part related to an antithromboxane activity [46]. Garlic also inhibits platelet aggregation, and aged garlic extract (AGE) blocks both the activation and aggregation of human platelets. The mechanism implicated by AGEs in the inhibition of platelet aggregation includes an increase in cAMP levels through inhibition of cAMP phosphodiesterase activity, resulting a reduction in calcium mobilization and, therefore, suppresses the binding of GPIIa/IIIb receptors to fibrinogen [144]. GBE was investigated on experimental cardiac remodeling in rats induced by acute myocardial infarction. The results suggest that GBE may inhibit experimental myocardial remodeling in rats after acute myocardial infarction by reducing the transcription of TGF-β1, MMP-2 and MMP-9 genes and attenuating the extracellular matrix deposition by decreasing the levels of proteins such as type I collagen, MMP-2 and MMP-9 [145].
Cardiometabolic parameters were evaluated in ApoE-/- mice fed an atherogenic diet and green coffee extract (GCE). Although GCE did not decrease atherosclerotic lesion progression or plasma lipid levels, it improved metabolic parameters, such as fasting glucose, insulin resistance, serum leptin, urinary catecholamines, and liver TGs. GCE also decreased weight gain, reduced adiposity, lower inflammatory infiltrate in adipose tissue, and protection against hepatic damage. Furthermore, the number of observed operational taxonomic untis (alpha diversity) diminished in ApoE-/- mice with an atherogenic diet, and it was recovered in the GCE-treated ApoE-/- mice [146]. Hibiscus sabdariffa extract (HSE) was evaluated in an experimental model of HFD-induced obesity in mice. HSE reduced weight in mice fed a HFD, and also improved glucose tolerance, insulin sensitivity and normalized LDL-C/HDL-C cholesterol ratio. HSE reduced the expression of different adipokines and pro-inflammatory mediators, and reinforced gut integrity by reducing the Firmicutes/Bacteroidetes ratio [147]. Salvia miltiorrhiza extract (SME) was investigated in rats with HFD-induced obesity. SME treatment markedly reduced weight, body fat index, lipid profile, glucose levels, and adipocyte vacuolation. The beneficial effects were accompanied with elevated concentrations of lipid factors such as cAMP, PKA, and HSL in the liver and adipose tissues, enhanced gut integrity, and ameliorated lipid metabolism. Furthermore, salvia miltiorrhiza extract reversed HFD-induced dysbacteriosis by promoting the abundance of Actinobacteriota and Proteobacteria and reducing the growth of Firmicutes and Desulfobacterita [101]. Therefore, the consumption of products derived from the plants and herbs previously analyzed should be considered as a new therapeutic strategy in the control of obesity and its associated disorders (Table 1).
4. Adipogenesis and Obesity
Accumulation and adipocyte differentiation are linked with the development of obesity. In the process of preadipocyte to adipocyte differentiation participates several transcription factors, being the most important cAMP response element-binding protein (CREB), CEBPβ and CCAAT/enhancer-binding protein beta and delta), CEBPα (CCAAT/enhancer-binding protein alpha (CEBPδ), and peroxisome proliferator activated receptor gamma (PPARγ), which control adipocyte differentiation. Adipocyte differentiation starts with CREB phosphorylation by PKA and ERK1/ERK2, and at the same time occurs the activation of CEBPβ and CEBPδ, which in turn activate CEBPα and PPARγ [148]. However, this process is much more complicated because it involves other biological signaling pathways. In addition to the adipocyte differentiation, PPARγ plays an important role in lipid storage and glucose homeostasis and is predominantly expressed in adipose tissue [149]. On the other hand, the PI3K/AKT pathway plays a critical role in transmitting insulin action in adipose tissue (increases glucose uptake by the glucose transporter 4 (GLUT4) membrane translocation) during adipogenesis of both WAT and brown adipose tissue (BAT). AKT is essential to induce PPARγ expression. The activation of PI3K/AKT signaling is determining for initiation of adipogenic transformation and adipocyte hyperplasia [150]. AMPK is a serine/threonine kinase which is expressed in several tissues (adipose, skeletal, liver, kidney, and hypothalamus), which regulates lipid/glucose homeostasis, autophagy, mitochondrial biogenesis, protein homeostasis, redox equilibrium, food intake, and insulin signaling. AMPK has a function as a cellular energy sensor. AMPK and adiponectin act in peripheral tissues and the central nervous system by regulating food intake. Consequently, the inhibition of hypothalamic AMPK activity along with an increase in adiponectin levels reduce food intake. In addition, AMPK inhibits de novo synthesis of cholesterol, FAs, and TGs, and activates FAs uptake and β-oxidation. It inhibits and phosphorylates proteins involved in the synthesis of FAs (FAS, ACC1, and SREBP-1c). AMPK inhibits the synthesis of cholesterol (phosphorylates and inhibits HMG-CoA reductase) AMPK inhibits the synthesis of cholesterol (phosphorylates and inhibits HMG-CoA reductase) and, through PGC-1α activation stimulates mitochondrial biogenesis and β-oxidation. AMPK inhibits adipogenesis via inhibition of the early mitotic clonal expansion (MCE) phase accompanied with reduction of early and late adipogenic factors including FAS, SREBP-1c and aP2 [151]. Therefore, the inhibition of differentiation into adipocytes by bioactive compounds from plant and herb extracts are beneficial for the loss of body fat and prevention of obesity (Table 1).
4.1. Adipogenesis as a Possible Target Against Obesity
The principal cause of obesity is energy overconsumption and/or insufficient energy expenditure, thereby, excessive food/energy intake leads to the expansion of WAT through de novo adipogenesis with the recruitment of new adipocytes (hyperplasia) and enlargement of existing adipocytes (hypertrophy). Chokeberry extract (Aronia melanocarpa) and its active polyphenols (seven anti-adipogenic polyphenolic phytochemicals) was investigated in HFD-induced obese mice. Amygdalin and prunasin were shown to inhibit 3T3-L1 adipocyte differentiation by suppressing the expressions of PPARγ, C/EBPα, SREBP-1c, FAS, and aP2. In addition, Chokeberry extract showed in obese mice significant decreases in body weight, serum TG, and LDL-C levels and improved insulin sensitivity [152]. The effects of cinnamon (Cinnamomum zeylanicum) extract were examined on the inhibition of adipocyte differentiation in 3T3-L1 fibroblast cells and in male mice fed a HFD. Cinnamon extract inhibited lipid accumulation and increased adiponectin and leptin genes in 3T3-L1 cells. In in vivo experiments, cinnamon extract elevated the expression of lipolysis-related proteins (AMPK, p-ACC, and CPT-1), and decreased the expression of lipid-synthesis-related proteins (SREBP-1c and FAS) in liver tissue [153]. Corni Fructus extract (CFE), which contain Corni Fructus, Dioscoreae Rhizoma, Aurantii Fructus Immaturus, and Platycodonis Radix, was shown to suppresses the differentiation of 3T3-L1 adipocytes by reducing the cellular induction of PPAR-γ, C/EBP-α, and lipin-1, including a significantly upregulation of AMPK-α phosphorylation. Moreover, CFE in obese mice fed HFD, induced weight loss in mice. Therefore, CFE has a potent anti-obesity activity due to the inhibition of adipocyte differentiation and adipogenesis [154]. Cydonia oblonga fruit extract (COFE) was tested on adipogenesis in 3T3-L1 preadipocytes. COFE inhibited intracellular TG deposition during adipogenesis. Furthermore, COFE treatment in 3T3-L1 cells induced upregulation of AMPK-α phosphorylation and downregulation of the adipogenic transcription factors (SREBP-1c, PPARγ, and C/EBPα). COFE also reduced the mRNA expression of FAS, ACLY, aP2, and lipoprotein lipase (LPL), including increased HSL and CPT-1 in 3T3-L1 cells [44]. Hibiscus sabdariffa extract (HSE) was examined on adipocyte differentiation in 3T3-L1 preadipocytes. HSE inhibited significantly the lipid droplet accumulation and attenuated adipogenic transcriptional factors, C/EBPα and PPARγ, during adipogenesis. HSE also reduced the expression of PI3-K/Akt and phosphorylation and expression of MEK-1/ERK during adipocyte differentiation. Taken together, HSE inhibits adipocyte differentiation through the regulation of PI3-K/Akt and ERK pathways, which play pivotal roles during adipogenesis [155]. Ilex paraguariensis extracts (IPE) was investigated in 3T3-L1 adipocytes and HFD-fed obese Sprague-Dawley (SD) rats. IPE inhibited intracellular lipid accumulation in 3T3-L1 adipocytes, increased AMPK-α, HSL, CaMKK, LKB1, PKA, C/EBPβ, IRβ, and IRS1(Tyr465), and decreased SREBP-1c, FAS, PPARγ, and IRS1 (Ser1101). Furthermore, an AMPK-α inhibitor abolished the effects exerted by IPE on intracellular lipid accumulation and HSL and FAS expression levels. In animals, IPE inhibited body weight gain and ameliorated serum cholesterol levels, and increased AMPK-α, PKA, Erk1/Erk2 (p44/p42), and UCP1 and and reduced those genes of mammalian target of rapamycin, S6 kinase, SREBP-1c, ap2, FAS, Il6, adiponectin, leptin, and Fabp4 in obese SD rats [156]. Moringa oleifera leaf petroleum ether extract (MOPEE) which has high levels of isoquercitrin, chrysin-7-glucoside, and quercitrin, was studied on lipid accumulation by in vitro and in vivo experiments. MOPEE suppressed adipogenesis in 3T3-L1 adipocytes by downregulated the expression of adipogenesis-associated proteins (PPARγ, C/EBPα and C/EBPβ, and FAS) and upregulated the expression of a lipolysis-associated protein (HSL). MOPEE also significantly increased the phosphorylation of AMPKα and ACC. In HFD-induced obese mice, MOPEE decreased body weight, epididymal, perirenal, mesenteric fat weight, and fat tissue size, including hepatic fat accumulation. Furthermore, MOPEE also decreased TC, LDL-C, and aspartate transferase (AST). Besides, MOPEE decreased the expression of adipogenesis-associated proteins (PPARγ and FAS) and upregulated the expression of a lipolysis-associated protein (ATGL) in liver and epididymal fat tissue. MOPEE also increased the phosphorylation of AMPKα and ACC in liver and epididymal fat tissue of obese mice. Therefore, MOPEE suppresses fat accumulation by inhibiting adipogenesis and promoting lipolysis [75]. A study explored the anti-adipogenic effects of lyophilised Opuntia cladode powders (OCP) in an in vitro and an in vivo HFD-induced obesity rat model. Two OCP were tested (O. streptacantha and O. ficus-indica). OCP impaired the differentiation in adipocytes (3T3 F442A) and decreased TG content and a low glucose uptake, thus suggesting an antiadipogenic effect. In SD rats, OCP slightly reduced body weight gain, liver and abdominal fat weights, improved some metabolic parameters and augmented TG excretion in the feces [157]. Platycodon grandiflorus extract (PGE) was investigated on the pre-adipocyte 3T3-L1 differentiation, pancreatic lipase activity, and HFD-induced obese rats. PGE inhibited 3T3-L1 pre-adipocyte differentiation and fat accumulation and reduced pancreatic lipase activity. In SD rats, PGE significantly reduced plasma TC and TG levels, body weight, and subcutaneous adipose tissue weight. PGE also reduced the size of subcutaneous adipocytes [158]. Taraxacum officinale (Dandelion) was investigated on adipocyte differentiation and lipogenesis in 3T3-L1 preadipocytes. Leaf and root extracts and a commercial root powder (caffeic and chlorogenic acids as the main phenolic constituents) were used in the study. All extract tested inhibited adipocyte differentiation and lipid accumulation in 3T3-L1 cells [105]. Therefore, the analyzed herbal and plant extracts play an important role during adipogenesis and lipid metabolism, supporting their therapeutic potential for the prevention and treatment of obesity (Table 1).

Table 1.
Extracts from different herbs and plants with anti-obesity properties.
| Name of herbs and plants and Method Extraction | Type of Study | Doses and Duration | Outcomes |
|---|---|---|---|
| Allium sativum (Galic) Aged Garlic Extract (15-20% aqueous ethanol) |
Isolated human platelets stimulated with ADP |
1.56 to 25% (v/v) | Inhibited platelet binding to fibrinogen by 40-70.4%, decreased PAC-1 binding to GPIIb/IIIa by 72%, and increased cAMP levels [144] |
| Andrographis paniculata (ethanolic extract) | 4-week-old male C57/BL6 mice with HFD (45% kcal from fat) |
2 g/kg/day, orally for a week | Attenuated cardiac hypertrophy and apoptosis, decreased ANP and BNP proteins, reduced cardiac collagen accumulation and fibrosis, inhibited COX-2, p-IκBα, and NF-κB proteins, reversed cardiac inflammation and myocardial apoptosis [141] |
| Aronia melanocar-pa (Chokeberry), methanol extract | 3T3-L1 adipo-cytes and 5-week-old male C57BL/6J mice with HFD (60% kcal from fat) | In vitro: 7 poly-phe-nols at 10 µM for 8 days. In vivo: 100 or 200mg/kg/day, oral-ly for 8 weeks | Inhibited 3T3-L1 adipocyte differentiation, de-creased body weight, serum TG, and LDL-C levels; and improved insulin sensitivity [152] |
| Aronia melanocarpa (Chokeberry) Polyphenol-rich extract (aqueous extract) |
Human platelets stimulated with ADP |
Platelet adhesion assay (range 0.5-100 µg/mL), thrombin activity (0.5-100 mg/mL), Plasmin activity (2.5, 5, 10, 20, 100 µg/mL) | Reduced ADP-activated platelet adhesion, increased overall potential of clotting and lysis, inhibited thrombin and plasmin amidolytic activity [142] |
| Aronia melanocarpa (Chokeberry), the extract was purchased from Agropharm SA (Poland) |
Patients with metabolic syndrome | 100 mg, three times daily for 2 months |
Reduced TC, LDL-C, and TG levels, inhibited platelet aggregation (less pronounced after 2 months), decreased potential for coagulation and clot formation, beneficial changes in coagulation and fibrinolysis parameters [143] |
| Camellia sinensis (Green tea aqueous extract, GTAE) | 12-week-old male Wistar rats with HFD (50% kcal from fat) |
1.1% and 2.0% GTAE for 8 weeks |
Reduced body weight gain (5.6% decrease at 2.0% GTAE), prevented visceral fat accumulation (17.8% reduction at 2.0% GTAE), lowered atherogenic index (14.3% reduction at both doses), reduced protein digestion (82.6% and 84.3% at 1.1% and 2.0% GTAE, respectively) [123] |
| Caralluma fimbriata (alcohol extract) |
Male Wistar rats (200-220 g) with cafeteria diet |
25, 50, 100 mg/kg/day for 90 days |
Inhibited food intake, prevention of body weight, liver weight, and fat pad mass gains, improved serum lipid and leptin profiles, and protection against atherogenesis [125] |
| Caralluma fimbriata (40% aqueous alcohol) |
Male Wistar rats (170–190 g) with HFD (60 kcal% from fat) |
200 mg/kg/day for 90 days |
Attenuated cardiac lipids and oxidative stress, and improved antioxidant enzyme activities [27] |
| Caralluma fimbriata (dry extract concentrate in gelatin capsules) |
Double-blind, randomized, placebo-controlled trial |
1 g/kg/day for 16 weeks | Reduced waist circumference, calorie intake, maintained body weight, reduced fat mass and BMI, and improved satiety markers [26] |
| Cinnamomum zeylanicum (70% ethylene alcohol) |
3T3-L1 cells and 7-week-old male C57BL/6J mice with a normal diet with 45% fat |
In vitro: 1, 3, 5, 7, 10 µg/mL for 3 days. In vivo: 1% cinnamon extract for 14 weeks |
In vitro: Inhibited lipid accumulation, increased adiponectin and leptin gene expression. In vivo: Reduced lipid synthesis, increased lipolysis, decreased VLDL-C, increased HDL-C, and lowered body fat and fatty tissue accumulation [153] |
| Citrullus colocynthis, hydro-alcoholic extract (80/20, v/v) |
9-week-old male Sprague-Dawley rats with HFD (45% kcal from fat) |
50 mg/kg/day, orally for 16 weeks |
Enhanced bleeding time and tPA levels, decreased PAI-1 and thromboxane B2, inhibited platelet aggregation, reversed HFD-induced increases in fibrinogen and von Willebrand factor, decreased food intake, pancreatic lipase activity, TNF-a, IL-6, and leptin, and increased adiponectin levels [33] |
| Coffea (Green coffee bean extract, GCBE from KPLC group: Montagne, France) |
5-week-old male C57BL/6J mice with HFD (60% Kcal from fat) |
Obesity induction for 4 weeks and then with extract (50, 100, 200 mg/kg/day) for 6 weeks |
Reduced body weight gain, liver weight, and white adipose tissue weights. Increased adiponectin and reduced leptin. GCBE upregulated mRNA levels of PPARα, ATGL, and HSL, and downregulated adipogenesis-related genes like C/EBPα, SREBP-1c, and PPARγ. GCBE increased pAMPK expression [130] |
| Coffea Arabica (aqueous extract) |
Male Wistar rats (160-180 g) with HFD (40% beef tallow) |
Obesity induction for 8 weeks and then with extract (200 mg/kg/day) for 8 weeks |
Decreased body and organ weights, reduced TC, TG, LDL-C, VLDL-C, glucose, and insulin levels, improved HOMA-IR, increased adiponectin, and reduced adipocyte hypertrophy [131] |
| Coffea canephora var. robusta beans (hot water extract) | 8-12-week-old male ApoE-/- mice with HFD (42% kcal from fat) |
At 2 weeks received 220 mg/kg/day for 14 weeks. At 4 weeks received HFD for 12 weeks |
Improved fasting glucose, insulin resistance, serum leptin, urinary catecholamines, and liver triglycerides. Reduced weight gain, adiposity, and inflammatory infiltrate in adipose tissue. Recovered operational taxonomic units (alpha diversity) [146] |
| Combination of Corni Fructus, Dioscoreae Rhizoma, Aurantii Fructus Immaturus, Platycodonis Radix (ethanol extract) |
3T3-L1 adipocytes and 5-week-old male C57BL/6J mice with HFD (60% kcal from fat) |
In vitro: 10, 50, 100 µg/mL for 48 hours. In vivo: Obesity induction for 4 weeks and then with extract (100 mg/kg/day) for 16 weeks |
Inhibited the differentiation of 3T3-L1 adipocytes and expressions of PPAR-γ, C/EBP-α, and lipin-1, increased phosphorylation of AMPK-α, and reduced weight gain in mice [154] |
| Cydonia oblonga (30% ethanol) |
3T3-L1 adipocytes | 0-600 µg/mL for 8 days |
Inhibited intracellular TG accumulation, induced AMPKα phosphorylation, downregulated adipogenic transcription factors (SREBP-1c, PPAR-γ, C/EBP-α), reduced mRNA expression of FAS, ACL, aP2, LPL, and increased mRNA expression of HSL and CPT-1 [44] |
| Cydonia oblonga (aqueous extract) | Male ICR mice (18-22 g) and male Wistar rats (300-350 g) | 20, 40, 80 mg/kg/day, orally for 14 days | Prolonged bleeding and clotting times, reduced pulmonary embolus mortality, increased thrombolysis, shortened ELT, reduced arterial and venous thrombus weights, decreased TXB2 and increased 6-keto-PGF1α levels [46] |
| Ginkgo biloba (extract obtained from Huacheng Biotech Inc. China) |
2-month-old male Wistar rats with HFD (57.3% from fat) |
Obesity induction for 2 months and then with extract (500 mg/kg/day), orally for 2 weeks |
Reduced energy intake, epididymal adipocyte volume, and lipid accumulation. It also reduced Plin 1 and Fasn mRNA and FAS protein levels [129] |
| Ginkgo biloba (unspecified extract) |
Male Sprague-Dawley rats (200-250 g) with acute myocardial infarction |
100 mg/kg/day, orally for 4 and 8 weeks |
Decreased TGF-β1, MMP-2, and MMP-9 mRNA transcription levels, reduced protein levels of type I collagen, MMP-2, and MMP-9, and inhibited myocardial remodeling after AMI [145] |
| Hibiscus sabdariffa (water extract) |
7-9 weeks old male C57BL/6J mice with HFD (60% kcal from fat) | 1, 10, 25 mg/kg/day for 42 days | Inhibited adipogenesis via PI3-K and MAPK pathways, reduced weight gain, improved glucose tolerance and insulin sensitivity, normalized LDL-C/HDL-C ratio, reduced inflammatory state in liver, reinforced gut integrity, and prebiotic effects on gut microbiota [147] |
| Hibiscus sabdariffa (hot water extract) |
3T3-L1 adipocytes | 2 mg/ml for 5 days | Inhibited adipocyte differentiation through PI3-K/Akt and ERK pathways, and decreased lipid droplet accumulation [155] |
| Ilex paraguariensis (Yerba mate), water extract |
6-week-old male Swiss strain mice with HFD |
Obesity induction for 8 weeks and then with extract (1 mg/kg) for 8 weeks |
Attenuation of weight gain, decreased adiposity and epididymal fat-pad weight, restored serum levels of cholesterol, TG, LDL-C, and glucose [68] |
| Ilex paraguariensis (Yerba mate), water extract |
6-week-old male C57BL/6J mice with HFD (60% kcal from fat) |
Obesity induction for 6 weeks and then with extract (0.5, 1, or 2 g/kg/day) for 4 weeks |
Reduced body weight gain, lower adipose tissue, decreased serum cholesterol, TG, and glucose levels [69] |
| Ilex paraguariensis (Yerba mate), 15% etanol extract |
6-week-old male Sprague-Dawley rats with HFD (40% kcal from fat) |
Daily supplementation of extract, 0.24% (w/w) for 60 days |
Reduced body weight, visceral fat, blood and hepatic lipid levels, improved glucose and insulin levels, enhanced AMPK phosphorylation, increased UCP2 and UCP3 expression [133] |
| Ilex paraguariensis (Yerba mate), water extract |
Early weaned Wistar rats |
1 g/kg BW/day, gavage for 30 days |
Reduced adipose mass (retroperitoneal and epididymal), total body fat, subcutaneous fat, visceral adipocyte area, TG, and hypothalamic NPY content; restored central leptin resistance, hyperphagia, and higher hypothalamic SOCS-3 content [134] |
| Ilex paraguariensis (Yerba mate), water extract (capsules) |
A randomized, double-blind, placebo-controlled clinical trial on obese Korean adults |
3 g/day for 12 weeks |
Decreased body fat mass, percent body fat, and WHR [136] |
| Ilex paraguariensis (Yerba mate), water extract |
8-week-old male Wistar rats with HFD (45% kcal from lard fat) |
100 mg/day in 3rd month of age and 200 mg/day in 4th month of age, daily for 2 months |
Reduced hypothalamic IKK phosphorylation and NF-κB p65 expression, increased IκBα and AdipoR1 expression, reduced IL-6 levels, increased IL-10/TNF-α ratio, and reduced low-grade inflammation [140] |
| Ilex paraguariensis (Yerba mate), water extract |
3T3-L1 adipocytes and 8-week-old male Sprague-Dawley rats with HFD (507.6 kcal/100 g) |
In vitro: 10, 50, 100 µg/mL for 7 days. In vivo: 500 mg/kg/day for 8 weeks |
In vitro: Suppressed lipid accumulation, increased AMPK, HSL, CaMKK, LKB1, PKA, C/EBPβ, IRβ, IRS1 (Tyr465), decreased SREBP-1c, FAS, PPARγ, and IRS1 (Ser1101). In vivo: suppressed body weight gain, improved serum cholesterol levels, increased AMPK, PKA, Erk1/Erk2, UCP1, reduced mTOR, S6K, SREBP-1c, ap2, FAS, IL-6, adiponectin, leptin, and Fabp4 [156] |
| Moringa oleifera (70% ethanol extract) |
Male albino rats (100 ± 20 g) with HFD (58% fat) and overweight/obese female patients |
In vivo: Obesity induction for 2 months and then with extract (200 and 400 mg/kg/day) for 1 month; patients: gelatine capsules (400 mg/day) for 8 weeks |
In rats, reduced final weight, adiposity index, glucose, insulin, and HOMA-IR. Increased R-QUICKI, adiponectin, omentin, GLUT-4, and PPAR-α expression. Reduced leptin and vaspin. Suppressed FAS and HMG-CoA reductase. In patients reduced BMI, TC, and LDL-C [74] |
| Moringa oleífera, Moringa oleifera leaf petroleum ether extract (MOPEE) |
3T3-L1 adipocytes and 7-week-old male C57BL/6J mice with HFD (60% kcal from fat) |
In vitro: 0, 50, 100, 200, and 400 µg/ml for 24 hours. In vivo: 0.125, 0.25, 0.5 g/kg/day for 14 weeks |
In vitro: Inhibited adipogenesis in a dose-dependent manner. Downregulated PPARγ, C/EBPα, C/EBPβ, FAS. Upregulated HSL, AMPKα, and ACC phosphorylation. In vivo: Decreased body weight, fat pad weight, and hepatic fat accumulation. Reduced TC, LDL-C, and AST levels. Downregulated PPARγ and FAS. Upregulated ATGL, AMPKα, and ACC phosphorylation [75] |
| Moringa oleifera (methanol extract from leaves) |
3-month-old male Wistar rats with HFD |
200 and 400 mg/kg/day for 12 weeks |
Alleviated serum biochemical abnormalities, balanced antioxidant status, and reestablished normal heart histology [139] |
| Opuntia streptacantha and Opuntia ficus-indica. Opuntia young cladode powders |
3T3-F442A adipocytes and 6-week-old male Sprague-Dawley rats with HFD (60% kcal from fat) |
In vitro: 1, 10, 100 μg/mL for 10 days. In vivo: 0.5% w/w for 8 weeks | In vitro: Impaired adipocyte differentiation and decreased TG, and reduced glucose uptake. In vivo: Slightly reduced body weight gain, liver and abdominal fat weights. Increased TG excretion in feces [157] |
| Platycodon grandiflorus (ethanol extract) |
Randomized, double-blind, placebo-controlled clinical trial on overweight or moderately obese adults |
571 mg, 1142 mg, 2855 mg (in tablets) per day for 12 weeks |
Decreased body fat mass and body fat percentage, reduced total abdominal and subcutaneous fat areas, increased muscle mass [137] |
| Platycodon grandifloras (water extract) | 3T3-L1 preadipocytes and 8-week-old male Sprague-Dawley rats with HFD (59.8% kcal from fat) | In vitro: various concentrations (10-50 mg/mL). In vivo: 150 mg/kg/day for 7 weeks |
Inhibited 3T3-L1 preadipocyte differentiation and fat accumulation. Decreased pancreatic lipase activity. In vivo: Reduced plasma TC) and TG levels, decreased body weight and subcutaneous adipose tissue weight, reduced size of subcutaneous adipocytes, repressed up-regulation of FABP mRNA in subcutaneous adipose tissue [158] |
| Punica granatum (Pomegranate), ethanol:water 70:30 | 6-week-old male C57BL/6 mice with HFD (45% of total fat) |
g/kg/3 days per week for 12-14 weeks |
Increased energy expenditure, reduced chronic inflammation and insulin resistance, promoted browning and thermogenesis in adipose tissue, reduced inflammatory markers, increased the reductive potential [95] |
| Salvia miltiorrhiza (75% etanol extract) |
8-9-week-old male Sprague-Dawley rats with HFD (45% kcal from fat) | 0.675, 1.35, 2.70 g/kg/day for 8 weeks | Reduced body weight, body fat index, serum lipid level, hepatic lipid accumulation, and adipocyte vacuolation. Improved gut integrity and lipid metabolism, altered gut microbiota composition [101] |
| Taraxacum officinale (95% ethanol extract) |
Porcine pancreatic lipase and 7-week-old male ICR mice |
In vitro: 50-250 µg/ml. In vivo: 400 mg/kg single dose for 240 min |
In vitro: inhibited pancreatic lipase activity. In vivo: decreased plasma TG levels and reduced AUC of plasma TG response curve [104] |
| Taraxacum officinale (leaf and root extracts in ethanol 60%) |
3T3-L1 adipocytes | 300-600 μg/μL for 6 days |
Inhibited adipocyte differentiation, reduced lipid and TG accumulation, regulated expression of genes and long non-coding RNAs involved in adipogenesis and lipid metabolism [105] |
| Theobroma cacao (aqueous extract) |
Wistar rats (250 ± 20 g) with HFD (45% kcal) and 20% fructose |
Obesity induction for 5 weeks and then with 100%, 10%, 1% pellet for 5 weeks |
Decreased body weight by 39%, systolic blood pressure by 27%, triglycerides by 55%, TC by 24%, LDL-C by 37%, and TG/HDL-C ratio by 54% [37] |
5. Plant and Herb Extracts Targeting Dyslipidemia and Adipokines in Obesity
Obesity-related dyslipidemia is considered as an atherogenic lipoprotein phenotype and one of the major risk factors for ischemic heart disease. The main manifestations of dyslipidemia include elevated plasma concentrations of TC, LDL-C and TGs, and low levels of HDL-C, which are important factors in hypertension and CVD [159]. Visceral obesity promotes insulin resistance in part mediated by high levels of FFAs and adipokines dysregulation. Adipokines such as leptin, resistin, and retinol-binding protein 4 increase insulin resistance, whereas adiponectin with anti-inflammatory and anti-lipogenic effects, increases insulin sensitivity. In addition, during obesity, pro-inflammatory mediators (leptin, resistin, IL-6, and TNF-α) may promote adipose tissue dysregulation and systemic insulin resistance [1].
In male SD rats fed with HFD were investigated the effects of a high hydrostatic pressure extract of garlic (HEG) on HDL-C level and hepatic apolipoprotein A-I (apoA-I) gene expression. In animals treated with HEG, plasma TC, TG and LDL-C levels were significantly decreased, while the plasma HDL-C level and mRNA level of hepatic apoA-I were significantly increased. Furthermore, HEG upregulated the gene expression of ATP-binding cassette transporter A1 (ABCA1) and lecithin-cholesterol acyl transferase (LCAT) in obese rats [160]. LVH is a risk factor for cardiovascular morbidity and mortality [161]. The effect of three Camellia sinensis teas (green, red, and white) were studied on LVH and insulin resistance in LDLr-/- mice fed a HFD. The teas partially prevented hyperlipidemia, increased HDL-C, reduced insulin resistance and CRP levels, and completely prevented LVH in LDLr-/- mice fed a HFD [162]. A systematic review and meta-analysis reported that green tea extract (GTE) significantly reduced TC, LDL-C, fasting blood sugar, hemoglobin A1c (HbA1c), and DBP, while increasing HDL-C [163]. Corni fructus extract (CFE) was administered in a rat model of diet-induced hypercholesterolemia, and the extract inhibited the elevation of both systolic and diastolic (BP), and lowered serum TC levels with a decrease in esterified cholesterol. Besides, the protein expression of SREBP-2 and PPARγ were elevated, indicating that CFE would activate FA oxidation [40]. Cydonia oblonga extract (COE) with flavonoids (> 60%) from leaves and fruit was analyzed on the blood lipid and antioxidant effects using hyperlipidaemic rat models. The flavonoids from COE significantly reduced serum TC, TG, LDL-C, ALT and AST, and increased HDL-C. Flavonoids improved the activity of SOD and GSH-Px in hepatic tissues, and reduced malondialdehyde acid (MDA) [45]. In rats fed with HFD, Ilex paraguariensis (yerba mate) extract reduced serum TG and TC and decreased the atherogenic index [164]. Ilex paraguariensis extract from leaves was investigated on hyperlipidemia induced in hamsters by a HFD. Yerba mate extract significantly reduced body-weight gain and lowered serum lipid levels, meanwhile, Yerba mate treatment increased antioxidant enzyme activity, ameliorated LPL and hepatic lipase activities in serum and liver, upregulated mRNA expression of PPARα, and downregulated mRNA expression of SREBP-1c and ACC in the liver. Therefore, Yerba mate extract regulates the expression of genes involved in lipid oxidation and lipogenesis [165]. Moreover, Yerba mate infusions were also studied in dyslipidemic individuals over 18 years of age (men and women). Yerba mate tea showed a significant increase in ferric reducing antioxidant potential and decreased glutathione concentrations but no significant changes in lipid hydroperoxide (LOOH), protein carbonyl, and paraoxonase-1 levels; thereby, Yerba mate tea increases plasma and blood antioxidant protection in patients with dyslipidemia [166]. However, a systematic review and meta-analysis found no differences in TC, LDL-C, HDL-C, and TG levels when comparing the yerba mate and control groups. The authors concluded that because the results are based on small inconclusive studies, more research is needed to confirm these findings [167]. Moringa oleifera leaves are used in India as a hypocholesterolemic agent in obese patients. A study reported that administration of the crude leaf extract of Moringa oleifera along with HFD reduced the HFD-induced increases in serum, liver, and kidney cholesterol levels. Furthermore, the crude extract increased serum albumin [76]. Nigella sativa has been used for the treatment and prevention of hyperlipidemia. A study analyzed different preparations reported of Nigella sativa including seed powder (100 mg-20 g daily), seed oil (20-800 mg daily), thymoquinone (3.5-20 mg daily), and seed extract (methanolic extract especially), and found that these preparations of Nigella sativa reduce plasma concentrations of TC, LDL-C, and TG, but the effect on HDL-C was not significant. The authors concluded that lipid-modifying properties of Nigella sativa could be attributed to the suppression of intestinal cholesterol absorption, reduced hepatic cholesterol synthesis, and up-regulation of LDL-C receptors [78]. Cholesterol reduction is critical for prevention of CVD. Opuntia ficus-indica extract (OFIE) was tested on inhibitory activity of pancreatic lipase enzyme (in vitro) and on hypercholesterolemia induced in mice by intraperitoneal administration of Triton WR-1339 (in vivo). The extracts significantly decreased blood cholesterol levels and inhibited pancreatic lipase activity. Therefore, OFIE prevents hypercholesterolemia by pancreatic lipase inhibition, partly attributed to its polyphenolic compounds [83]. Platycodon grandiflorus extract (PGE) was investigated in obese mice. The extract reduced body weight gain and improved plasma lipid profiles. Furthermore, leptin was significantly reduced whereas adiponectin was elevated. PGE also downregulated lipogenic genes (LPL, ACC, and FAS) expression and increased lipolysis genes (CPT-1, HSL, and UCP2) in WAT and liver. Moreover, PGE inhibited adipogenic transcriptional factors, such as PPARγ, C/EBPα, and SREBP-1c [88]. Another study related to Platycodon grandiflorus root extract in HFD-induced obese mice, reported that the extract exhibited antioxidant activity, meanwhile, in calf pulmonary arterial endothelial cells inhibited both oxLDL-C-induced cell death and lactate dehydrogenase release. In obese mice treated with Platycodon grandiflorus root extract increased antioxidant proteins and reduced plasma and hepatic lipid levels, thus demonstrating its beneficial effects on hyperlipidemia [89]. The representative adipokines secreted from adipose tissues with an increased plasma leptin, resistin, and TNF-α, and a reduced plasma adiponectin are related with systemic insulin resistance [1]. PGE in male ICR mice fed an HFD, markedly attenuated food intake, epididymal fat weight, body weight, adipocyte size, and blood glucose levels, meanwhile, maintained serum levels of adiponectin, resistin, leptin, fructosamine, and TGs. PGE also up-regulates adiponectin, and down-regulates TNF-α, and leptin in fat tissue. In L6 muscle cells, the extract elevated insulin-stimulated glucose uptake [90]. Taken together, the evidence from experimental and clinical studies suggest that plant extracts have lipid-lowering effects and adipokines regulation, which may be suitable for the prevention and treatment of obesity (Table 2).
6. Plant and Herb Extracts Against to Insulin Resistance, Hyperglycemia, and Diabetes
Several factors contribute to the growth of obesity and T2D such as unhealthy eating habits, physical inactivity, sedentary lifestyle, increased stress, and environmental factors, which are typical to global urbanization and modern society in the world [1]. Increased BMI and excessive visceral adipose tissue are known to be associated with insulin resistance and beta cell dysfunction, which can lead to glucose intolerance and T2D. Obesity plays an important role in the elevated prevalence of T2D, which is characterized by insulin resistance in several tissues, including muscle, liver, and adipose tissue. T2D is manifested by low insulin secretions from β-cells and peripheral insulin resistance, including high levels of fatty acids, which promote systemic inflammation. Obesity course with a chronic inflammatory state with high levels of cytokines such as TNF-α, IL-1β, monocyte chemoattractant protein-1 (MCP-1) and IL-6, which in part contributes to the pathogenesis of insulin resistance, promoting a reduction in glucose uptake in insulin-dependent tissues, which increases blood glucose levels, β-cell dysfunction in pancreas and results in endocrine dysfunction of adipose tissue. One of the signaling pathways involved in the inflammatory mechanisms is the activation of JNK1 by TNF-α, which results in serine phosphorylation of insulin receptor substrate 1 (IRS1) and impairs insulin signaling and subsequent insulin resistance [1,168].
In addition to obesity and T2D hyperglycemia is a known risk factor for the development of several health disorders such as oxidative stress, and cardiovascular diseases. Caralluma fimbriata extract (CFE) was investigated on the effects of HFD feeding on insulin resistance and oxidative stress in Wistar rats. Administration of CFE in obese animals resulted in a significant improvement in plasma glucose, insulin, leptin, and TGs. CFE aso prevented high levels of lipid peroxidation, protein oxidation and low GSH levels and low activities of enzymatic antioxidants [169]. Cinnamon extracts have been used to treat blood glucose. Ceylon cinnamon extract (CCE) was tested on carbohydrate digestion and post-meal blood glucose reduction in vitro (enzymatic assays) and in vivo (starch tolerance tests in rats), including 18 healthy female and male volunteers. In vitro study, CCE inhibited pancreatic α-amylase activity, and reduced the glycemic response to starch in a dose-dependent manner in rats. In healthy volunteers, CCE lowered the area under the curve of glycemia with no changes in the insulin secretion [30]. Citrullus colocynthis have been used against diabetes. Colocynth fruit extract was investigated on insulin action using 3T3-L1 adipocytes. Extracts of seed and pulp enhanced insulin-induced GLUT4 translocation and increased insulin-stimulated cellular glucose uptake. Moreover, extracts of pulp enhanced insulin-induced PKB phosphorylation without affecting phosphorylation of the insulin receptor [170]. However, a systematic review and meta-analysis in diabetic patients reported that Citrullus colocynthis does not have a significant effect on fasting blood sugar, HbA1c, LDL-C, TC, and TG indices, but increases HDL-C. These findings could be related to the relatively low quality of articles and the small number of included studies [32]. Corni fructus extract (CFE) was analyzed on blood glucose and insulin resistance in db/db mice. CFE suppressed an increase in blood glucose level during the oral glucose tolerance test. In addition, CFE lowered final fasting serum glucose and TG in diabetic mice. The mRNA expression of adiponectin, GLUT 4 and PPAR-γ in adipose tissue were higher in diabetic mice treated with CFE [41]. In another study Corni fructus extract was evaluated on kidneys of diabetic db/db mice. The activities of XO and SOD were significantly higher in diabetic mice with CFE, meanwhile the activities of CAT and GST were lowered in the diabetic mice with CFE. Besides, the mRNA expression of eNOS in kidneys was reduced in the diabetic mice with the extract. Therefore, Corni fructus extract has antioxidative actions which contribute to its renoprotective effects on diabetic nephropathy [171]. Ginkgo biloba extract (GBE) leaves (water and 12% ethanol extracts) from four different trees (1 and 2-males and 3 and 4-females) were analyzed on phenolic profile, antioxidant activity, and the potential in vitro inhibitory properties on α-amylase, α-glucosidase, and ACE enzymes, which are related to the diabetes and hypertension. Aqueous extracts had higher phenolic content than the ethanolic extracts. ACE activity was only inhibited by ethanolic extracts. The results showed a strong correlation between total phenolics and α-glucosidase inhibitory activity, and to a lesser degree positive correlation between total phenolics and α-amylase inhibitory activity [50]. EGb761, a Ginkgo biloba extract, has antioxidant and antiplatelet aggregation effects. It was analyzed the effect of EGb761 and its major bioactive compounds (bilobalide, kaempferol, and quercetin) on preventing atherosclerosis in vitro and in a rat model of T2D. EGb761 dose-dependently reduced intima-media ratio, proliferation of VSMCs and promoted greater apoptosis in obese rats with T2D. In vitro model, EGb761 also decreased proliferation and migration of VSMCs. Glucose and circulating adiponectin levels were ameliorated and plasma hsCRP concentrations were reduced in obesogenic and diabetic rats. Moreover, caspase-3 activity and DNA fragmentation were increased while monocyte adhesion and ICAM-1/VCAM-1 levels were decreased with EGb761 in obese rats with T2D. The bioactive compounds of EGb761, kaempferol and quercetin decreased VSMC migration and elevated caspase activity [51]. Green coffee extract (GCE) was evaluated by 10 weeks on glycemic indices, inflammation, and oxidative stress in individuals with T2D and overweight/obesity. GCE supplementation reduced body weight and BMI. In addition, patients with GCE had a lower fasting blood glucose (FBG) concentration, but there was no effect on insulin levels and HOMA-IR. However, there were significant improvements in SBP, TG level, HDL-C, and TG-to-HDL-C ratio. GCE supplementation had no effect on DBP, LDL-C, or TC, including MDA levels. GCE significantly decreased the hs-CRP levels in patients with T2D and overweight/obesity. Therefore, GCE possesses beneficial effects on lipid profile and inflammation in individuals with T2D and overweight/obesity [172]. Hibiscus sabdariffa polyphenolic extract (HPE) was analyzed on the T2D rat model. HPE decreased hyperglycemia and hyperinsulinemia, including serum TG, cholesterol, and the ratio of LDL-C/ HDL-C. Moreover, HPE significantly reduced the plasma advanced glycation end product (AGE) formation and lipid peroxidation in T2D rats. Furthermore, HPE inhibits the expression of connective tissue growth factor (CTGF) and receptor of AGE (RAGE). HPE retrieved the weight loss found in T2D rats [173]. Extracts of two varieties (red and white) of Hibiscus sabdariffa (Roselle) calyces were evaluated on carbohydrate hydrolyzing enzymes (α-amylase and α-glucosidase). The extracts inhibited the α-amylase and α-glucosidase activities in vitro, but the red variety exhibited higher α-glucosidase inhibitory activity than the white variety, while the white variety showed higher α-amylase inhibitory activity than the red variety. Besides, the red variety possesses higher antioxidant capacity, which appears to be more potent compared to white variety [63]. Hibiscus sabdariffa extract (HSE) was evaluated on the mechanism of adipogenesis and complications of the obesity-related insulin resistance in HFD-induced obese SD rats. HSE reduced food intake, body weight, lipid profiles, lipid peroxidation, inflammatory cytokines, serum leptin, insulin and duodenal glucose absorption, while significantly elevated glucose uptake of adipose tissue and muscle. Moreover, HSE prevents lipid accumulation by inhibiting differentiation of 3T3-L1 adipocytes through downregulation of genes involved in adipogenesis [62]. Roasted mate tea consumption (Ilex paraguariensis) was evaluated on the glycemic and lipid profiles of patients with T2D or pre-diabetes. Mate tea consumption reduced the levels of fasting glucose, HbA1c, and LDL-C of T2D patients. However, the consumption of mate tea did not change the intake of total energy, carbohydrate, protein, cholesterol, and fiber. In addition, mate tea consumption together with nutritional counseling reduced significantly the levels of LDL-C, HDL-C, and TG. Therefore, Mate tea consumption ameliorated the glucose levels and lipid profile of T2D patients [70]. Moringa oleifera has beneficial properties to reduce the risk of chronic metabolic diseases such as T2D. One study investigated capsules of Moringa oleifera dry leaf powder in subjects with prediabetes for 12 weeks. Moringa oleifera improved FBG and HbA1c. There were no significant changes in microbiota, hepatic and renal function markers or the appetite-controlling hormones, such as glucagon-like peptide 1 (GLP-1), ghrelin, and peptide YY (PYY) [77]. Opuntia ficus-indica var. saboten (OFS) dried powder extract was investigated using in vitro and in vivo models. OSF inhibited α-glucosidase activity in vitro and intestinal glucose absorption in db/db mice. In L6 muscle cells, OFS elevated dose-dependent glucose uptake, stimulated AMPK and p38 MAPK phosphorylations, and increased GLUT4 translocation to the cell membrane. OFS treatment in db/db mice dose-dependently prevented hyperinsulinemia, hyperglycemia, and glucose tolerance, including insulin resistance and quantitative insulin sensitivity check index. OFS ameliorated pancreatic function through elevated β-cell mass in db/db mice [174]. Another study of Opuntia ficus-indica extract (OFIE) prepared from the cladodes and a proprietary stem/fruit skin-blend was tested on blood glucose and plasma insulin in normal rats. OFIE significantly lowered blood glucose levels and significantly elevated basal plasma insulin levels, suggesting a direct action on pancreatic β-cells [85]. The flowering part of Punica granatum has been recommended in Unani literature for the treatment of diabetes. Punica granatum flower extract (PGFE) was analyzed on hyperglycemia in vivo and in vitro. PGF extract markedly reduced plasma glucose levels (postprandial hyperglycemia) in non-fasted Zucker diabetic fatty rats (a genetic model of obesity and T2D). In vitro, PGFE had a potent inhibitory effect on α-glucosidase activity [96]. Salvia miltiorrhiza extract (SME) was investigated on the expression of VEGF induced by high concentration of glucose in HMEC-1 cells, in which mitochondrial uncoupling protein 2 (UCP2) was knockdown by using UCP2 siRNA. HMEC-1 cells with 30 mM glucose resulted in a significant increase in the expression of VEGF mRNA, and high levels of ROS. SME significantly decreased VEGF mRNA and ROS formation in HMEC-1 cells with 30 mM glucose. Interestingly, the knockdown of UCP-2 abolished the reduction of VEGF expression and ROS formation by SME. Therefore, SME has antioxidant effects and can be used for the treatment of diabetic chronic vascular complications [100]. The anti-diabetic potential of fresh and shade-dried leaves of Taraxacum officinale was investigated. The extract of shade-dried Taraxacum officinale leaves demonstrated potent anti-diabetic activity in a dose-dependent manner by targeting α-amylase and α-glucosidase, having great potential to suppress post-prandial glucose rise and for better management of diabetes [107]. Plant extracts described in this section have anti-diabetic effects because they improve the glycemic control and lipid profile, postprandial hyperglycemia, insulin resistance, inflammatory cytokines, and oxidative stress in diabetic models. Therefore, its consumption combined with nutritional intervention could be a good strategy to decrease plasma glucose levels and lipid parameters in individuals with pre-diabetes and diabetes, which may reduce their risk of developing metabolic disorders and coronary artery disease (Table 3).
7. Plant and Herb Extracts with Anti-Hypertensive Effects
Obesity-associated hypertension is well documented in children and adults and in both sexes. Excess weight gain (especially increased visceral adiposity) is a major cause of hypertension and accounts for 65% to 75% of the risk for human primary hypertension and causes a cascade of associated cardiorenal and metabolic disorders. The mechanisms involved in obesity-associated hypertension are complex and include [1] physical compression of the kidneys from excess fat in and around the kidneys, [2] sympathetic nervous system (SNS) overactivation, [3] activation of the renin-angiotensin-aldosterone system (RAAS), [4] dysregulation in adipose tissue-secreted cytokines, such as leptin, insulin, resistance, TNF-α, and IL-6), [5] systemic insulin resistance, [6] endothelial dysfunction, and [7] structural and functional renal changes. In addition, SNS overactivation promotes elevations in heart rate, cardiac output, and renal tubular sodium reabsorption, which occur due to α-adrenergic and β-adrenergic receptor stimulation and indirectly through activation of other systems (e.g., RAAS) [175,176]. Weight loss is the main goal to reduce obesity-related hypertension, and current therapeutic approaches address the metabolic consequences of obesity, including dyslipidemia, inflammation, and diabetes.
The consumption of aged black garlic (ABG) is associated with improvements in several CVD risk factors. ABG extract along dietary recommendations was analyzed on CVD risk factors in subjects with moderate hypercholesterolemia. ABG extract for 6 weeks reduced DBP, particularly in men with a DBP > 75 mm Hg [177]. Andrographis paniculata extract (APE) was investigated using chronic intraperitoneal infusions by osmotic pumps in spontaneously hypertensive rats (SHRs) and Wistar-Kyoto (WKY) rats. APE significantly reduced the SBP of both SHRs and WKY rats. Plasma ACE activity and thiobarbituric acid (TBA) were significantly reduced in SHRs treated with APE [178]. In a meta-analysis of controlled clinical trials, berry extract from Aronia melanocarpa (chokeberry) was tested for an average of 6-8 weeks on TC and BP. Daily supplementation with berry extract significantly reduced SBP and TC, mainly in adults over the age of 50 years [179]. According to epidemiological studies, green tea (Camellia sinensis) consumption has protective effects against CVD. Green tea extract (GTE) with Ang II (induces endothelial dysfunction) were investigated for 13 days on arterial hypertension with high oxidative stress in male SD rats. GTE blunted the increased BP, LV mass index, media-to-lumen ratio, and hydroperoxide radicals, including HO-1, p22phox, and SOD-1 mRNA in aorta caused by Ang II [180]. In a systematic review and meta-analysis of randomized clinical trials, GTE was analyzed in 20 human randomized clinical trials comprising 1536 participants. GTE significantly reduced SBP, TC, and LDL-C. Adverse events reported were elevated BP, rash, and abdominal discomfort [181]. In a crossover randomized clinical trial, green tea was analyzed on BP, endothelial function, inflammatory activity, and metabolic profile in obese prehypertensive women. Participants received daily 3 capsules containing 500 mg of green tea extract (GTE) for 4 weeks, with a washout period of 2 weeks between treatments. Each GTE capsule had 260 mg of polyphenols. After 4 weeks of GTE supplementation, there was a significant reduction in SBP at 24 hours, daytime, and nighttime [182]. The intake of cocoa extract, which consisted of 1.4 g (415 mg flavanols) before and after 4 weeks of daily intake, was evaluated on postprandial cardiometabolic effects. The consumption of cocoa extract within an energy-restricted diet for 4 weeks showed a greater reduction in postprandial AUC of SBP compared to the control group and independently of body weight loss [36]. Leaf extract of ginkgo biloba (EGb761) was investigated on hypertension with hypercholesterolemia-induced renal injury in rats. Hypertension was caused by L-N(G)-nitroarginine methyl ester (L-NAME) and hypercholesterolemia was induced by a diet with 1% cholesterol. EGb761 exhibited a progressive reduction in the SBP, diastolic blood pressure (SBP) and mean arterial BP. Moreover, EGb761 decreased the excess of MDA and nitrite levels and recovered the low levels of intracellular reduced glutathione (GSH) caused by hypertension with hypercholesterolemia in the renal tissue. Furthermore, hypertension with hypercholesterolemia increased the expression of TNF-α, IL-6, and IL-1β levels in renal tissues and was inhibited by treatment with EGb761. Chronic hypertension with hypercholesterolemia induced inhibition of endothelial nitric oxide synthase (eNOS) and activation of inducible NO synthase (iNOS), but EGb761 activated eNOS and inhibited iNOS in the kidney tissues. Therefore, these findings suggest that EGb761 protects against hypertension with hypercholesterolemia-induced renal injury [52]. A new component group of Ginkgo biloba leaves (GBLCG), mainly composed of quercetin, kaempferol, and isorhamnetin, was investigated on reducing BP and ameliorating myocardial hypertrophy in SHRs. Total terpenoid lactones of GBLCG might be a novel cocrystal composed of Ginkgolide (A, B, C, J) and bilobalide. GBLCG had hypotensive activity and improved the myocardial hypertrophy. These effects could be due to the promoting of NO synthesis and release in endothelial cells, reducing oxidative stress, and inhibiting platelet aggregation [183]. Green coffee bean extract (GCE) has protective effects against hypertension in both SHRs and humans. A study investigated the dose-response relationship of GCE in 117 male subjects with mild hypertension. After 28 days of using GCE, the decrease in SBP and DBP were statistically significant compared with the placebo group. Therefore, GCE has antihypertensive effects in patients with mild hypertension [57]. It has been reported that Hibiscus sabdariffa can reduce BP in human and animal studies. The extract of the dried calyx of Hibiscus sabdariffa (HS) and Hibiscus anthocyanins (HAs) were investigated on left ventricular myocardial capillary length and surface area in SHRs. HS consumption significantly decreased SBP, DBP and LV mass in a dose-dependent fashion, but it had no effect on the heart rate. HS also significantly increased surface area and length density of myocardial capillaries and length density. Myocyte nuclear volume was reduced in rats with HS. There was an insignificant decrease in SBP and DBP with HA ingestion. This study showed that HS ingestion improves myocardial capillarization in SHRs through structural alterations linked to a reduction of myocardial mass and the promotion of new vessel formation [184]. Hibiscus sabdariffa extract (HSE) was investigated on RAAS in mild to moderate essential hypertensive Nigerians. After 4 weeks of treatment with HSE (150 mg/kg/day), the extract significantly (P < 0.001) reduced plasma aldosterone; meanwhile, serum ACE and plasma renin activity did not change significantly. The effects observed could be related to the presence of anthocyanins in the extract [185]. Hibiscus sabdariffa calyces (HSC) extract was analyzed on BP, vascular function and other cardiometabolic risk factors in men with 1% to 10% CVD risk. The consumption of aqueous extract of HSC significantly increases in % flow mediated dilatation of the branchial artery, and there was no significant decrease in SBP and DBP, non-significant increase in urinary and plasma nitric oxide (NOx) and reduced levels of plasma insulin and serum glucose, including TG and CRP. There was a significant improvement in the area under systemic antioxidant response curve and consumption of the HSC extract showed no significant changes in arterial stiffness [186]. HS extract was studied on isolated mesenteric arteries from normotensive (Wistar and WKY) and SHRs. HS extract caused a concentration-dependent relaxant effect on mesenteric arteries rings of SHRs (EC50 = 0.83 ± 0.08 mg/mL), WKY (EC50 = 0.46 ± 0.04 mg/mL), and Wistar rats (EC50 = 0.44 ± 0.08 mg/mL) pre-contracted with phenylephrine (10 µM). HS extract 2 mg/mL significantly reduced the peak of the L-type calcium current seen in cardiac myocytes by 24%. HS extract did not promote a membrane hyperpolarization of smooth muscle cells, which could suggest an absence of a direct effect on background potassium current. The authors concluded that HS extract probably implicates a vasorelaxant effect on small resistance arteries, which does not depend on the endothelium; and the reduction of L-type calcium current is part of this effect [64]. Nigella sativa seed extract (100 and 200 mg twice a day) supplement was evaluated in patients with mild hypertension. After 8 weeks, SBP and DBP values were statistically significantly reduced in a dose-dependent manner. The extract also caused a significant decline in the level of TC and LDL-C [80]. Platycodon grandiflorus (PG) is used to reduce inflammation and lower BP in the Chinese population. Platycodon grandiflorus root was tested for inhibiting Ang II-induced IGF-IIR activation and apoptosis pathway in H9c2 cells and SHRs. The crude extract of PG significantly inhibited the Ang II-induced IGFIIR signaling to avoid H9c2 cells apoptosis. PG extract suppressed Ang II-dependent JNK activation and SIRT1 degradation to decrease IGF-IIR activity. Besides, PG maintained SIRT1 stability to improve HSF1-mediated IGF-IIR suppression, which prevents H9c2 cells apoptosis. In SHRs, PG markedly decreased this apoptotic pathway in the heart tissues, thus PG could be considered for the treatment of heart diseases in hypertensive patients [91]. According to the antioxidant properties of pomegranate, its peel extract was analyzed on damage related to hypertension and aging in a SHR model. Pomegranate peel extract showed significant reduction in SBP and coronary ACE activity. The extract also reduced superoxide anion levels and vascular wall areas in the coronary of SHRs treated with peel extract. Therefore, this study suggests that pomegranate peel extract may have beneficial effects on coronary heart disease [97]. Antioxidant properties related to Leaf and root extracts of Taraxacum officinale were investigated in vitro and in vivo. For the in vivo model, experiments were performed on organ homogenate samples from L-NAME-induced Wistar rats. Leaf extract of Taraxacum officinale possessed significantly higher polyphenol and flavonoid, including free radical scavenging activity (EC50 0.37 compared to 1.34 mg/mL) and total antioxidant capacities (82.56% compared to 61.54% 3-ethylbenzothiazoline-6-sulfonic acid: ABTS, and 156 ± 5.28 compared to 40 ± 0.31 ferric reducing antioxidant power: FRAP). Both extracts significantly increased total antioxidant capacities (kidney and brain tissues) and reduced MDA levels (heart tissue) [106]. Taken together, there are several plants and herbs that their extracts have blood pressure-lowering properties in patients and animal models with hypertension. Therefore, these extracts could be used as a therapeutic strategy to prevent and treat hypertension-associated to obesity (Table 4).
8. Conclusions and Perspectives
Obesity and associated cardiovascular diseases have been recognized as a public health concern, mainly in countries where its prevalence is alarmingly high. According to the World Obesity Atlas 2023 estimation, 38% of the global population is currently overweight or obese. In Mexico, the projected trends in obesity prevalence (BMI ≥30kg/m2) will be very high by 2035, 47% in adults (https://www.worldobesity.org/resources/resource-library/world-obesity-atlas-2023, accessed on 28 June 2024). Currently, various agents are used to prevent or treat obesity and associated metabolic disorders; for instance, for lowering lipid (e.g., statins, inhibitors of enzyme HMG-CoA reductase) and body weight (e.g., orlistat, an inhibitor of pancreatic lipase), and common treatment strategies employed for CVDs include a combination of anticoagulant and antithrombotic therapy such as aspirin, clopidogrel (tienopyrin), apixaban, dabigatran, rivaroxban and warfarin. Unfortunately, the use of these medications causes potentially serious side effects, including nausea, vomiting, flatulence, diarrhea, insomnia, headache, hemorrhagic, and ischemic complications. For this reason, the World Health Organization (Committee, 1980) recommended the use of herbal and plant-based medicines. Compared with pharmaceutical agents, herbs and plants extract offer similar benefits against cardiometabolic risk factors associated with obesity without the side effects. Herbs and plants extract are rich source of various nutrients and medicinal phytochemicals, including vitamins, minerals, carotenoids, fatty acids and esters, oils, polysaccharides, proteins, polyphenols, fibers, catechins, flavonoids, terpenes, and other compounds. These bioactive phytochemicals show various beneficial properties against various human diseases. For example, polyphenols have antioxidant, anti-inflammatory, anti-hypertensive, and atherogenic effects, and they can inhibit platelet aggregation and activation [187], thus having potential protective role in several diseases such as obesity, T2D, and cardiovascular disease. Although most herb and plant extracts analyzed in this review were investigated in vitro and animals’ models, further future research studies in clinical trials are required to confirm the beneficial properties of these herbs and plants against cardiometabolic risk factors associated with obesity. It is well-known that obesity is associated with processed foods and high-calorie diets, including a sedentary lifestyle. Therefore, health care systems and governments in countries with a current high prevalence of obesity must encourage people to consume healthy nutrition (vegetables and fruits), physical activity, and maintain a healthy weight to avoid obesity and CVDs, which are the main cause of mortality globally. In 2021, CVDs accounted for 20.5 million deaths, of which around 80% occurred in low- and middle-income countries [188]. Therefore, overall, the consumption of herbs and plant teas should be recommended as a possible approach to reduce cardiovascular diseases associated with obesity.
Author Contributions
J.G.-C. and J.A.-B. conceived, designed, and revised the review. J.G.-C., D.L.-C., A.S.-R. and J.B.-G. wrote the draft, reviewed, and edited it. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by CONAHCYT, Mexico, under grant CF-2023-I-473 to J.G.-C.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gutiérrez-Cuevas, J.; Santos, A.; Armendariz-Borunda, J. Pathophysiological Molecular Mechanisms of Obesity: A Link between MAFLD and NASH with Cardiovascular Diseases. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Karri, S.; Sharma, S.; Hatware, K.; Patil, K. Natural Anti-Obesity Agents and Their Therapeutic Role in Management of Obesity: A Future Trend Perspective. Biomedicine and Pharmacotherapy 2019, 110. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-cuevas, J.; Sandoval-rodriguez, A.; Meza-rios, A.; Monroy-ramírez, H.C.; Galicia-moreno, M.; García-bañuelos, J.; Santos, A.; Armendariz-borunda, J. Molecular Mechanisms of Obesity-linked Cardiac Dysfunction: An Up-date on Current Knowledge. Cells 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Schuster, D. Obesity and the Development of Type 2 Diabetes: The Effects of Fatty Tissue Inflammation. Diabetes Metab Syndr Obes 2010. [Google Scholar] [CrossRef]
- McLaughlin, T.; Lamendola, C.; Liu, A.; Abbasi, F. Preferential Fat Deposition in Subcutaneous versus Visceral Depots Is Associated with Insulin Sensitivity. Journal of Clinical Endocrinology and Metabolism 2011, 96. [Google Scholar] [CrossRef]
- Konige, M.; Wang, H.; Sztalryd, C. Role of Adipose Specific Lipid Droplet Proteins in Maintaining Whole Body Energy Homeostasis. Biochim Biophys Acta Mol Basis Dis 2014, 1842. [Google Scholar] [CrossRef] [PubMed]
- Garin-Shkolnik, T.; Rudich, A.; Hotamisligil, G.S.; Rubinstein, M. FABP4 Attenuates PPARγ and Adipogenesis and Is Inversely Correlated with PPARγ in Adipose Tissues. Diabetes 2014, 63. [Google Scholar] [CrossRef]
- Vincent, H.K.; Taylor, A.G. Biomarkers and Potential Mechanisms of Obesity-Induced Oxidant Stress in Humans. Int J Obes 2006, 30. [Google Scholar] [CrossRef]
- Csige, I.; Ujvárosy, D.; Szabó, Z.; Lorincz, I.; Paragh, G.; Harangi, M.; Somodi, S.; Santulli, G. The Impact of Obesity on the Cardiovascular System. J Diabetes Res 2018, 2018. [Google Scholar] [CrossRef]
- Pi-Sunyer, X. The Medical Risks of Obesity. Postgrad Med 2009, 121. [Google Scholar] [CrossRef]
- Hossain, P.; Kawar, B.; El Nahas, M. Obesity and Diabetes in the Developing World — A Growing Challenge. New England Journal of Medicine 2007, 356. [Google Scholar] [CrossRef] [PubMed]
- Abdi Beshir, S.; Ahmed Elnour, A.; Soorya, A.; Parveen Mohamed, A.; Sir Loon Goh, S.; Hussain, N.; Al Haddad, A.H.I.; Hussain, F.; Yousif Khidir, I.; Abdelnassir, Z. A Narrative Review of Approved and Emerging Anti-Obesity Medications. Saudi Pharmaceutical Journal 2023, 31. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.G.; Park, C.Y. Anti-Obesity Drugs: A Review about Their Effects and Safety. Diabetes Metab J 2012, 36. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.S.; Ahmed, N. Antiglycation Properties of Aged Garlic Extract: Possible Role in Prevention of Diabetic Complications. In Proceedings of the Journal of Nutrition; 2006; Vol. 136.
- Imaizumi, V.M.; Laurindo, L.F.; Manzan, B.; Guiguer, E.L.; Oshiiwa, M.; Otoboni, A.M.M.B.; Araujo, A.C.; Tofano, R.J.; Barbalho, S.M. Garlic: A Systematic Review of the Effects on Cardiovascular Diseases. Crit Rev Food Sci Nutr 2023, 63. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.J.; Wang, P.Q.; Li, S.J.; Li, X.K.; Zhang, Y.Q.; Wang, J. Garlic for Hypertension: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Phytomedicine 2015, 22. [Google Scholar] [CrossRef]
- Nakagawa, S.; Masamoto, K.; Sumiyoshi, H.; Harada, H. Acute Toxicity Test of Garlic Extract. Journal of Toxicological Sciences 1984, 9. [Google Scholar] [CrossRef] [PubMed]
- Fardiyah, Q.; Ersam, T.; Suyanta; Slamet, A.; Suprapto; Kurniawan, F. New Potential and Characterization of Andrographis Paniculata L. Ness Plant Extracts as Photoprotective Agent. Arabian Journal of Chemistry 2020, 13. [CrossRef]
- Wong, S.K.; Chin, K.Y.; Ima-Nirwana, S. A Review on the Molecular Basis Underlying the Protective Effects of Andrographis Paniculata and Andrographolide against Myocardial Injury. Drug Des Devel Ther 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Kulling, S.E.; Rawel, H.M. Chokeberry (Aronia Melanocarpa) - A Review on the Characteristic Components and Potential Health Effects. Planta Med 2008, 74. [Google Scholar] [CrossRef]
- Banjari, I.; Misir, A.; Šavikin, K.; Jokić, S.; Molnar, M.; De Zoysa, H.K.S.; Waisundara, V.Y. Antidiabetic Effects of Aronia Melanocarpa and Its Other Therapeutic Properties. Front Nutr 2017, 4. [Google Scholar] [CrossRef]
- Jurikova, T.; Mlcek, J.; Skrovankova, S.; Sumczynski, D.; Sochor, J.; Hlavacova, I.; Snopek, L.; Orsavova, J. Fruits of Black Chokeberry Aronia Melanocarpa in the Prevention of Chronic Diseases. Molecules 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Brimson, J.M.; Prasanth, M.I.; Kumaree, K.K.; Thitilertdecha, P.; Malar, D.S.; Tencomnao, T.; Prasansuklab, A. Tea Plant (Camellia Sinensis): A Current Update on Use in Diabetes, Obesity, and Cardiovascular Disease. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Dinh, T.C.; Thi Phuong, T.N.; Minh, L.B.; Minh Thuc, V.T.; Bac, N.D.; Van Tien, N.; Pham, V.H.; Show, P.L.; Tao, Y.; Nhu Ngoc, V.T.; et al. The Effects of Green Tea on Lipid Metabolism and Its Potential Applications for Obesity and Related Metabolic Disorders - An Existing Update. Diabetes and Metabolic Syndrome: Clinical Research and Reviews 2019, 13. [Google Scholar] [CrossRef] [PubMed]
- Anwar, R.; Rabail, R.; Rakha, A.; Bryla, M.; Roszko, M.; Aadil, R.M.; Kieliszek, M. Delving the Role of Caralluma Fimbriata: An Edible Wild Plant to Mitigate the Biomarkers of Metabolic Syndrome. Oxid Med Cell Longev 2022, 2022. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Briskey, D.; dos Reis, C.; Mallard, A.R. The Effect of an Orally-Dosed Caralluma Fimbriata Extract on Appetite Control and Body Composition in Overweight Adults. Sci Rep 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Gujjala, S.; Putakala, M.; Bongu, S.B.R.; Ramaswamy, R.; Desireddy, S. Preventive Effect of Caralluma Fimbriata against High-Fat Diet Induced Injury to Heart by Modulation of Tissue Lipids, Oxidative Stress and Histological Changes in Wistar Rats. Arch Physiol Biochem 2022, 128. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.V.; Gan, S.H. Cinnamon: A Multifaceted Medicinal Plant. Evidence-based Complementary and Alternative Medicine 2014, 2014. [Google Scholar] [CrossRef]
- Roussel, A.M.; Hininger, I.; Benaraba, R.; Ziegenfuss, T.N.; Anderson, R.A. Antioxidant Effects of a Cinnamon Extract in People with Impaired Fasting Glucose That Are Overweight or Obese. J Am Coll Nutr 2009, 28. [Google Scholar] [CrossRef]
- Beejmohun, V.; Peytavy-Izard, M.; Mignon, C.; Muscente-Paque, D.; Deplanque, X.; Ripoll, C.; Chapal, N. Acute Effect of Ceylon Cinnamon Extract on Postprandial Glycemia: Alpha-Amylase Inhibition, Starch Tolerance Test in Rats, and Randomized Crossover Clinical Trial in Healthy Volunteers. BMC Complement Altern Med 2014, 14. [Google Scholar] [CrossRef]
- Cheng, X.; Qin, M.; Chen, R.; Jia, Y.; Zhu, Q.; Chen, G.; Wang, A.; Ling, B.; Rong, W. Citrullus Colocynthis (L.) Schrad.: A Promising Pharmaceutical Resource for Multiple Diseases. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Jafarizadeh, A.; Raeisi, S.A.; Ghassab-Abdollahi, N.; Yarani, R.; Araj-Khodaei, M.; Mirghafourvand, M. Effect of Citrullus Colocynthis on Glycemic Factors and Lipid Profile in Type II Diabetic Patients: A Systematic Review and Meta-Analysis. J Diabetes Metab Disord 2022, 21. [Google Scholar] [CrossRef] [PubMed]
- Alhawiti, N.M. Antiplatelets and Profibrinolytic Activity of Citrullus Colocynthis in Control and High-Fat Diet-Induced Obese Rats: Mechanisms of Action. Arch Physiol Biochem 2018, 124. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, R.; Amin, G.; Ardekani, M.R.S. A Review on Citrullus Colocynthis Schrad.: From Traditional Iranian Medicine to Modern Phytotherapy. Journal of Alternative and Complementary Medicine 2012, 18. [Google Scholar] [CrossRef] [PubMed]
- Davinelli, S.; Corbi, G.; Righetti, S.; Sears, B.; Olarte, H.H.; Grassi, D.; Scapagnini, G. Cardioprotection by Cocoa Polyphenols and ω-3 Fatty Acids: A Disease-Prevention Perspective on Aging-Associated Cardiovascular Risk. J Med Food 2018, 21. [Google Scholar] [CrossRef] [PubMed]
- Ibero-Baraibar, I.; Suárez, M.; Arola-Arnal, A.; Zulet, M.A.; Martinez, J.A. Cocoa Extract Intake for 4 Weeks Reduces Postprandial Systolic Blood Pressure Response of Obese Subjects, Even after Following an Energy-Restricted Diet. Food Nutr Res 2016, 60. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, I.; Ortiz, A.; Sanchez-Pardo, M.; Garduño-Siciliano, L.; Hernández-Ortega, M.; Villarreal, F.; Meaney, E.; Najera, N.; Ceballos, G.M. Obesity and Cardiovascular Risk Improvement Using Cacao By-Products in a Diet-Induced Obesity Murine Model. J Med Food 2019, 22. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Feng, Z.L.; Chen, H.B.; Wang, F.S.; Lu, J.H. Corni Fructus: A Review of Chemical Constituents and Pharmacological Activities. Chinese Medicine (United Kingdom) 2018, 13. [Google Scholar] [CrossRef]
- Gao, X.; Liu, Y.; An, Z.; Ni, J. Active Components and Pharmacological Effects of Cornus Officinalis: Literature Review. Front Pharmacol 2021, 12. [Google Scholar] [CrossRef]
- Park, C.H.; Cho, E.J.; Yokozawa, T. Protection against Hypercholesterolemia by Corni Fructus Extract and Its Related Protective Mechanism. J Med Food 2009, 12. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, K.S.; Lee, T.J.; Kim, Y.C. Antidiabetic Effects of Corni Fructus Extract on Blood Glucose and Insulin Resistance in Db/Db Mice. Toxicol Res 2009, 25. [Google Scholar] [CrossRef]
- Amerizadeh, A.; Vaseghi, G.; Esmaeilian, N.; Asgary, S. Cardiovascular Effects of Cydonia Oblonga Miller (Quince). Evidence-based Complementary and Alternative Medicine 2022, 2022. [Google Scholar] [CrossRef] [PubMed]
- Wojdyło, A.; Oszmiański, J.; Bielicki, P. Polyphenolic Composition, Antioxidant Activity, and Polyphenol Oxidase (PPO) Activity of Quince (Cydonia Oblonga Miller) Varieties. J Agric Food Chem 2013, 61. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Jung, J.I.; Hwang, J.S.; Hwang, M.O.; Kim, E.J. Cydonia Oblonga Miller Fruit Extract Exerts an Anti-Obesity Effect in 3T3-L1 Adipocytes by Activating the AMPK Signaling Pathway. Nutr Res Pract 2023, 17. [Google Scholar] [CrossRef] [PubMed]
- Umar, A.; Iskandar, G.; Aikemu, A.; Yiming, W.; Zhou, W.; Berké, B.; Begaud, B.; Moore, N. Effects of Cydonia Oblonga Miller Leaf and Fruit Flavonoids on Blood Lipids and Anti-Oxydant Potential in Hyperlipidemia Rats. J Ethnopharmacol 2015, 169. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Abdurahman, A.; Umar, A.; Iskander, G.; Abdusalam, E.; Berké, B.; Bégaud, B.; Moore, N. Effects of Cydonia Oblonga Miller Extracts on Blood Hemostasis, Coagulation and Fibrinolysis in Mice, and Experimental Thrombosis in Rats. J Ethnopharmacol 2014, 154. [Google Scholar] [CrossRef] [PubMed]
- Silva, H.; Martins, F.G. Cardiovascular Activity of Ginkgo Biloba—An Insight from Healthy Subjects. Biology (Basel) 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Mohanta, T.K.; Tamboli, Y.; Zubaidha, P.K. Phytochemical and Medicinal Importance of Ginkgo Biloba L. Nat Prod Res 2014, 28. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Zhu, F.; Pan, M.; Liu, Q.; Wang, P. Pharmacokinetic, Metabolism, and Metabolomic Strategies Provide Deep Insight Into the Underlying Mechanism of Ginkgo Biloba Flavonoids in the Treatment of Cardiovascular Disease. Front Nutr 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Pinto, M.; Kwon, Y.I.; Apostolidis, E.; Lajolo, F.M.; Genovese, M.I.; Shetty, K. Potential of Ginkgo Biloba L. Leaves in the Management of Hyperglycemia and Hypertension Using in Vitro Models. Bioresour Technol 2009, 100. [Google Scholar] [CrossRef]
- Lim, S.; Yoon, J.W.; Kang, S.M.; Choi, S.H.; Cho, B.J.; Kim, M.; Park, H.S.; Cho, H.J.; Shin, H.; Kim, Y.B.; et al. Egb761, a Ginkgo Biloba Extract, Is Effective against Atherosclerosis in Vitro, and in a Rat Model of Type 2 Diabetes. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Abdel-Zaher, A.O.; Farghaly, H.S.M.; El-Refaiy, A.E.M.; Abd-Eldayem, A.M. Protective Effect of the Standardized Extract of Ginkgo Biloba (EGb761) against Hypertension with Hypercholesterolemia-Induced Renal Injury in Rats: Insights in the Underlying Mechanisms. Biomedicine and Pharmacotherapy 2017, 95. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. American Journal of Clinical Nutrition 2004, 79. [Google Scholar] [CrossRef] [PubMed]
- Nunes, F.M.; Coimbra, M.A. Chemical Characterization of Galactomannans and Arabinogalactans from Two Arabica Coffee Infusions as Affected by the Degree of Roast. J Agric Food Chem 2002, 50. [Google Scholar] [CrossRef] [PubMed]
- Blum, J.; Lemaire, B.; Lafay, S. Effect of a Green Decaffeinated Coffee Extract on Glycaemia. Nutrafoods 2007, 6. [Google Scholar]
- Asbaghi, O.; Kashkooli, S.; Mardani, M.; Rezaei kelishadi, M.; Fry, H.; Kazemi, M.; Kaviani, M. Effect of Green Coffee Bean Extract Supplementation on Liver Function and Inflammatory Biomarkers: A Meta-Analysis of Randomized Clinical Trials. Complement Ther Clin Pract 2021, 43. [Google Scholar] [CrossRef] [PubMed]
- Kozuma, K.; Tsuchiya, S.; Kohori, J.; Hase, T.; Tokimitsu, I. Antihypertensive Effect of Green Coffee Bean Extract on Mildly Hypertensive Subjects. Hypertension Research 2005, 28. [Google Scholar] [CrossRef] [PubMed]
- Asbaghi, O.; Sadeghian, M.; Nasiri, M.; Khodadost, M.; Shokri, A.; Panahande, B.; Pirouzi, A.; Sadeghi, O. The Effects of Green Coffee Extract Supplementation on Glycemic Indices and Lipid Profile in Adults: A Systematic Review and Dose-Response Meta-Analysis of Clinical Trials. Nutr J 2020, 19. [Google Scholar] [CrossRef] [PubMed]
- Sapian, S.; Ibrahim Mze, A.A.; Jubaidi, F.F.; Mohd Nor, N.A.; Taib, I.S.; Abd Hamid, Z.; Zainalabidin, S.; Mohamad Anuar, N.N.; Katas, H.; Latip, J.; et al. Therapeutic Potential of Hibiscus Sabdariffa Linn. in Attenuating Cardiovascular Risk Factors. Pharmaceuticals 2023, 16. [Google Scholar] [CrossRef] [PubMed]
- Da-Costa-Rocha, I.; Bonnlaender, B.; Sievers, H.; Pischel, I.; Heinrich, M. Hibiscus Sabdariffa L. - A Phytochemical and Pharmacological Review. Food Chem 2014, 165. [Google Scholar] [CrossRef]
- Gurrola-Díaz, C.M.; García-López, P.M.; Sánchez-Enríquez, S.; Troyo-Sanromán, R.; Andrade-González, I.; Gómez-Leyva, J.F. Effects of Hibiscus Sabdariffa Extract Powder and Preventive Treatment (Diet) on the Lipid Profiles of Patients with Metabolic Syndrome (MeSy). Phytomedicine 2010, 17. [Google Scholar] [CrossRef]
- Janson, B.; Prasomthong, J.; Malakul, W.; Boonsong, T.; Tunsophon, S. Hibiscus Sabdariffa L. Calyx Extract Prevents the Adipogenesis of 3T3-L1 Adipocytes, and Obesity-Related Insulin Resistance in High-Fat Diet-Induced Obese Rats. Biomedicine and Pharmacotherapy 2021, 138. [Google Scholar] [CrossRef]
- Ademiluyi, A.O.; Oboh, G. Aqueous Extracts of Roselle (Hibiscus Sabdariffa Linn.) Varieties Inhibit α-Amylase and α-Glucosidase Activities in Vitro. J Med Food 2013, 16. [Google Scholar] [CrossRef]
- Alsayed, A.M.A.; Zhang, B.L.; Bredeloux, P.; Boudesocque-Delaye, L.; Yu, A.; Peineau, N.; Enguehard-Gueiffier, C.; Ahmed, E.M.; Pasqualin, C.; Maupoil, V. Aqueous Fraction from Hibiscus Sabdariffa Relaxes Mesenteric Arteries of Normotensive and Hypertensive Rats through Calcium Current Reduction and Possibly Potassium Channels Modulation. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- de Vasconcellos, A.C.; Frazzon, J.; Zapata Noreña, C.P. Phenolic Compounds Present in Yerba Mate Potentially Increase Human Health: A Critical Review. Plant Foods for Human Nutrition 2022, 77. [Google Scholar] [CrossRef] [PubMed]
- Paluch, E.; Okińczyc, P.; Zwyrzykowska-Wodzińska, A.; Szperlik, J.; Żarowska, B.; Duda-Madej, A.; Babelewski˛, P.; Włodarczyk, M.; Wojtasik, W.; Kupczyński, R.; et al. Composition and Antimicrobial Activity of Ilex Leaves Water Extracts. Molecules 2021, 26. [Google Scholar] [CrossRef]
- dos Santos, T.W.; Miranda, J.; Teixeira, L.; Aiastui, A.; Matheu, A.; Gambero, A.; Portillo, M.P.; Ribeiro, M.L. Yerba Mate Stimulates Mitochondrial Biogenesis and Thermogenesis in High-Fat-Diet-Induced Obese Mice. Mol Nutr Food Res 2018, 62. [Google Scholar] [CrossRef]
- Arçari, D.P.; Bartchewsky, W.; Dos Santos, T.W.; Oliveira, K.A.; Funck, A.; Pedrazzoli, J.; De Souza, M.F.F.; Saad, M.J.; Bastos, D.H.M.; Gambero, A.; et al. Antiobesity Effects of Yerba Maté Extract (Ilex Paraguariensis) in High-Fat Diet-Induced Obese Mice. Obesity 2009, 17. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-R.; Lee, H.-Y.; Kim, J.-H.; Moon, D.-I.; Seo, M.-Y.; Park, S.-H.; Choi, K.-H.; Kim, C.-R.; Kim, S.-H.; Oh, J.-H.; et al. Anti-Obesity and Anti-Diabetic Effects of Yerba Mate ( Ilex Paraguariensis ) in C57BL/6J Mice Fed a High-Fat Diet. Lab Anim Res 2012, 28. [Google Scholar] [CrossRef] [PubMed]
- Klein, G.A.; Stefanuto, A.; Boaventura, B.C.B.; De Morais, E.C.; Da Cavalcante, L.S.; De Andrade, F.; Wazlawik, E.; Di Pietro, P.F.; Maraschin, M.; Da Silva, E.L. Mate Tea (Ilex Paraguariensis) Improves Glycemic and Lipid Profiles of Type 2 Diabetes and Pre-Diabetes Individuals: A Pilot Study. J Am Coll Nutr 2011, 30. [Google Scholar] [CrossRef]
- González Arbeláez, L.F.; Fantinelli, J.C.; Ciocci Pardo, A.; Caldiz, C.I.; Ríos, J.L.; Schinella, G.R.; Mosca, S.M. Effect of an Ilex Paraguariensis (Yerba Mate) Extract on Infarct Size in Isolated Rat Hearts: The Mechanisms Involved. Food Funct 2016, 7. [Google Scholar] [CrossRef]
- Azlan, U.K.; Mediani, A.; Rohani, E.R.; Tong, X.; Han, R.; Misnan, N.M.; Jam, F.A.; Bunawan, H.; Sarian, M.N.; Hamezah, H.S. A Comprehensive Review with Updated Future Perspectives on the Ethnomedicinal and Pharmacological Aspects of Moringa Oleifera. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Mbikay, M. Therapeutic Potential of Moringa Oleifera Leaves in Chronic Hyperglycemia and Dyslipidemia: A Review. Front Pharmacol 2012, 3 MAR. [Google Scholar] [CrossRef]
- Ezzat, S.M.; El Bishbishy, M.H.; Aborehab, N.M.; Salama, M.M.; Hasheesh, A.; Motaal, A.A.; Rashad, H.; Metwally, F.M. Upregulation of MC4R and PPAR-α Expression Mediates the Anti-Obesity Activity of Moringa Oleifera Lam. in High-Fat Diet-Induced Obesity in Rats. J Ethnopharmacol 2020, 251. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wang, Y.; Jiang, W.W.; Luo, X.F.; Dai, T.Y.; Peng, L.; Song, S.; Li, L.F.; Tao, L.; Shi, C.Y.; et al. Moringa Oleifera Leaf Petroleum Ether Extract Inhibits Lipogenesis by Activating the AMPK Signaling Pathway. Front Pharmacol 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Ghasi, S.; Nwobodo, E.; Ofili, J.O. Hypocholesterolemic Effects of Crude Extract of Leaf of Moringa Oleifera Lam in High-Fat Diet Fed Wistar Rats. J Ethnopharmacol 2000, 69. [Google Scholar] [CrossRef]
- Gómez-Martínez, S.; Díaz-Prieto, L.E.; Castro, I.V.; Jurado, C.; Iturmendi, N.; Martín-Ridaura, M.C.; Calle, N.; Dueñas, M.; Picón, M.J.; Marcos, A.; et al. Moringa Oleifera Leaf Supplementation as a Glycemic Control Strategy in Subjects with Prediabetes. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Asgary, S.; Sahebkar, A.; Goli-Malekabadi, N. Ameliorative Effects of Nigella Sativa on Dyslipidemia. J Endocrinol Invest 2015, 38. [Google Scholar] [CrossRef]
- Derosa, G.; D’Angelo, A.; Maffioli, P.; Cucinella, L.; Nappi, R.E. The Use of Nigella Sativa in Cardiometabolic Diseases. Biomedicines 2024, 12. [Google Scholar] [CrossRef]
- Dehkordi, F.R.; Kamkhah, A.F. Antihypertensive Effect of Nigella Sativa Seed Extract in Patients with Mild Hypertension. Fundam Clin Pharmacol 2008, 22. [Google Scholar] [CrossRef]
- Madrigal-Santillán, E.; Portillo-Reyes, J.; Madrigal-Bujaidar, E.; Sánchez-Gutiérrez, M.; Izquierdo-Vega, J.A.; Izquierdo-Vega, J.; Delgado-Olivares, L.; Vargas-Mendoza, N.; Álvarez-González, I.; Morales-González, Á.; et al. Opuntia Spp. in Human Health: A Comprehensive Summary on Its Pharmacological, Therapeutic and Preventive Properties. Part 2. Plants 2022, 11. [Google Scholar] [CrossRef]
- Abbas, E.Y.; Ezzat, M.I.; El Hefnawy, H.M.; Abdel-Sattar, E. An Overview and Update on the Chemical Composition and Potential Health Benefits of Opuntia Ficus-Indica (L.) Miller. J Food Biochem 2022, 46. [Google Scholar] [CrossRef]
- Padilla-Camberos, E.; Flores-Fernandez, J.M.; Fernandez-Flores, O.; Gutierrez-Mercado, Y.; Carmona-De La Luz, J.; Sandoval-Salas, F.; Mendez-Carreto, C.; Allen, K. Hypocholesterolemic Effect and in Vitro Pancreatic Lipase Inhibitory Activity of an Opuntia Ficus-Indica Extract. Biomed Res Int 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- López-Romero, P.; Pichardo-Ontiveros, E.; Avila-Nava, A.; Vázquez-Manjarrez, N.; Tovar, A.R.; Pedraza-Chaverri, J.; Torres, N. The Effect of Nopal (Opuntia Ficus Indica) on Postprandial Blood Glucose, Incretins, and Antioxidant Activity in Mexican Patients with Type 2 Diabetes after Consumption of Two Different Composition Breakfasts. J Acad Nutr Diet 2014, 114. [Google Scholar] [CrossRef]
- Butterweck, V.; Semlin, L.; Feistel, B.; Pischel, I.; Bauer, K.; Verspohl, E.J. Comparative Evaluation of Two Different Opuntia Ficus-Indica Extracts for Blood Sugar Lowering Effects in Rats. Phytotherapy Research 2011, 25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, Y.; Yang, D.; Zhang, C.; Zhang, N.; Li, M.; Liu, Y. Platycodon Grandiflorus - An Ethnopharmacological, Phytochemical and Pharmacological Review. J Ethnopharmacol 2015, 164. [Google Scholar] [CrossRef]
- Ji, M.Y.; Bo, A.; Yang, M.; Xu, J.F.; Jiang, L.L.; Zhou, B.C.; Li, M.H. The Pharmacological Effects and Health Benefits of Platycodon Grandiflorus - A Medicine Food Homology Species. Foods 2020, 9. [Google Scholar] [CrossRef]
- Hwang, K.A.; Hwang, Y.J.; Im, P.R.; Hwang, H.J.; Song, J.; Kim, Y.J. Platycodon Grandiflorum Extract Reduces High-Fat Diet-Induced Obesity Through Regulation of Adipogenesis and Lipogenesis Pathways in Mice. J Med Food 2019, 22. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.J.; Kim, S.H.; Park, J.W.; Lee, Y.J.; Ham, S.S. Platycodon Grandiflorum Root Attenuates Vascular Endothelial Cell Injury by Oxidized Low-Density Lipoprotein and Prevents High-Fat Diet-Induced Dyslipidemia in Mice by up-Regulating Antioxidant Proteins. Nutrition Research 2012, 32. [Google Scholar] [CrossRef]
- Ahn, Y.M.; Kim, S.K.; Kang, J.S.; Lee, B.C. Platycodon Grandiflorum Modifies Adipokines and the Glucose Uptake in High-Fat Diet in Mice and L6 Muscle Cells. Journal of Pharmacy and Pharmacology 2012, 64. [Google Scholar] [CrossRef]
- Lin, Y.C.; Lin, C.H.; Yao, H.T.; Kuo, W.W.; Shen, C.Y.; Yeh, Y.L.; Ho, T.J.; Padma, V.V.; Lin, Y.C.; Huang, C.Y.; et al. Platycodon Grandiflorum (PG) Reverses Angiotensin II-Induced Apoptosis by Repressing IGF-IIR Expression. J Ethnopharmacol 2017, 205. [Google Scholar] [CrossRef]
- Hou, C.; Zhang, W.; Li, J.; Du, L.; Lv, O.; Zhao, S.; Li, J. Beneficial Effects of Pomegranate on Lipid Metabolism in Metabolic Disorders. Mol Nutr Food Res 2019, 63. [Google Scholar] [CrossRef] [PubMed]
- Maphetu, N.; Unuofin, J.O.; Masuku, N.P.; Olisah, C.; Lebelo, S.L. Medicinal Uses, Pharmacological Activities, Phytochemistry, and the Molecular Mechanisms of Punica Granatum L. (Pomegranate) Plant Extracts: A Review. Biomedicine and Pharmacotherapy 2022, 153. [Google Scholar] [CrossRef] [PubMed]
- Mokgalaboni, K.; Dlamini, S.; Phoswa, W.N.; Modjadji, P.; Lebelo, S.L. The Impact of Punica Granatum Linn and Its Derivatives on Oxidative Stress, Inflammation, and Endothelial Function in Diabetes Mellitus: Evidence from Preclinical and Clinical Studies. Antioxidants 2023, 12. [Google Scholar] [CrossRef]
- Reguero, M.; Gómez de Cedrón, M.; Sierra-Ramírez, A.; Fernández-Marcos, P.J.; Reglero, G.; Quintela, J.C.; Ramírez de Molina, A. Pomegranate Extract Augments Energy Expenditure Counteracting the Metabolic Stress Associated with High-Fat-Diet-Induced Obesity. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Li, Y.; Wen, S.; Kota, B.P.; Peng, G.; Li, G.Q.; Yamahara, J.; Roufogalis, B.D. Punica Granatum Flower Extract, a Potent α-Glucosidase Inhibitor, Improves Postprandial Hyperglycemia in Zucker Diabetic Fatty Rats. J Ethnopharmacol 2005, 99. [Google Scholar] [CrossRef]
- dos Santos, R.L.; Dellacqua, L.O.; Delgado, N.T.B.; Rouver, W.N.; Podratz, P.L.; Lima, L.C.F.; Piccin, M.P.C.; Meyrelles, S.S.; Mauad, H.; Graceli, J.B.; et al. Pomegranate Peel Extract Attenuates Oxidative Stress by Decreasing Coronary Angiotensin-Converting Enzyme (ACE) Activity in Hypertensive Female Rats. Journal of Toxicology and Environmental Health - Part A: Current Issues 2016, 79. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Zhu, R.; Tian, Y.; Chen, B.; Li, R.; Li, L.; Wang, L.; Che, Y.; Zhao, D.; Mo, F.; et al. Salvia Miltiorrhiza in Diabetes: A Review of Its Pharmacology, Phytochemistry, and Safety. Phytomedicine 2019, 58. [Google Scholar] [CrossRef]
- Ren, J.; Fu, L.; Nile, S.H.; Zhang, J.; Kai, G. Salvia Miltiorrhiza in Treating Cardiovascular Diseases: A Review on Its Pharmacological and Clinical Applications. Front Pharmacol 2019, 10. [Google Scholar] [CrossRef]
- Qian, S.; Huo, D.; Wang, S.; Qian, Q. Inhibition of Glucose-Induced Vascular Endothelial Growth Factor Expression by Salvia Miltiorrhiza Hydrophilic Extract in Human Microvascular Endothelial Cells: Evidence for Mitochondrial Oxidative Stress. J Ethnopharmacol 2011, 137. [Google Scholar] [CrossRef]
- Ai, Z.L.; Zhang, X.; Ge, W.; Zhong, Y.B.; Wang, H.Y.; Zuo, Z.Y.; Liu, D.Y. Salvia Miltiorrhiza Extract May Exert an Anti-Obesity Effect in Rats with High-Fat Diet-Induced Obesity by Modulating Gut Microbiome and Lipid Metabolism. World J Gastroenterol 2022, 28. [Google Scholar] [CrossRef]
- Kania-Dobrowolska, M.; Baraniak, J. Dandelion (Taraxacum Officinale L.) as a Source of Biologically Active Compounds Supporting the Therapy of Co-Existing Diseases in Metabolic Syndrome. Foods 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Olas, B. New Perspectives on the Effect of Dandelion, Its Food Products and Other Preparations on the Cardiovascular System and Its Diseases. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kang, M.-J.; Kim, M.-J.; Kim, M.-E.; Song, J.-H.; Lee, Y.-M.; Kim, J.-I. Pancreatic Lipase Inhibitory Activity of Taraxacum Officinale in Vitro and in Vivo. Nutr Res Pract 2008, 2. [Google Scholar] [CrossRef] [PubMed]
- González-Castejón, M.; García-Carrasco, B.; Fernández-Dacosta, R.; Dávalos, A.; Rodriguez-Casado, A. Reduction of Adipogenesis and Lipid Accumulation by Taraxacum Officinale (Dandelion) Extracts in 3T3L1 Adipocytes: An in Vitro Study. Phytotherapy Research 2014, 28. [Google Scholar] [CrossRef] [PubMed]
- Aremu, O.O.; Oyedeji, A.O.; Oyedeji, O.O.; Nkeh-Chungag, B.N.; Sewani Rusike, C.R. In Vitro and in Vivo Antioxidant Properties of Taraxacum Officinale in Nω-Nitro-L-Arginine Methyl Ester (L-NAME)-Induced Hypertensive Rats. Antioxidants 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Murtaza, I.; Laila, O.; Drabu, I.; Ahmad, A.; Charifi, W.; Popescu, S.M.; Mansoor, S. Nutritional Profiling, Phytochemical Composition and Antidiabetic Potential of Taraxacum Officinale, an Underutilized Herb. Molecules 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or Hyperplasia: Dynamics of Adipose Tissue Growth. PLoS Comput Biol 2009, 5. [Google Scholar] [CrossRef] [PubMed]
- Stich, V.; Berlan, M. Physiological Regulation of NEFA Availability: Lipolysis Pathway. Proceedings of the Nutrition Society 2004, 63. [Google Scholar] [CrossRef] [PubMed]
- Frühbeck, G.; Méndez-Giménez, L.; Fernández-Formoso, J.A.; Fernández, S.; Rodríguez, A. Regulation of Adipocyte Lipolysis. Nutr Res Rev 2014, 27. [Google Scholar] [CrossRef]
- Lafontan, M.; Langin, D. Lipolysis and Lipid Mobilization in Human Adipose Tissue. Prog Lipid Res 2009, 48. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab Syndr Relat Disord 2015, 13. [Google Scholar] [CrossRef] [PubMed]
- Ahima, R.S.; Flier, J.S. Adipose Tissue as an Endocrine Organ. Trends Endocrinol Metab 2000, 11, 327–332. [Google Scholar] [CrossRef]
- Macdougall, C.E.; Wood, E.G.; Loschko, J.; Scagliotti, V.; Cassidy, F.C.; Robinson, M.E.; Feldhahn, N.; Castellano, L.; Voisin, M.B.; Marelli-Berg, F.; et al. Visceral Adipose Tissue Immune Homeostasis Is Regulated by the Crosstalk between Adipocytes and Dendritic Cell Subsets. Cell Metab 2018, 27. [Google Scholar] [CrossRef]
- Gotoh, K.; Inoue, M.; Masaki, T.; Chiba, S.; Shimasaki, T.; Ando, H.; Fujiwara, K.; Katsuragi, I.; Kakuma, T.; Seike, M.; et al. A Novel Anti-Inflammatory Role for Spleen-Derived Interleukin-10 in Obesity-Induced Inflammation in White Adipose Tissue and Liver. Diabetes 2012, 61. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, G.; Zhang, H.; Karin, M.; Bai, H.; Cai, D. Hypothalamic IKKbeta/NF-KappaB and ER Stress Link Overnutrition to Energy Imbalance and Obesity. Cell 2008, 135, 61–73. [Google Scholar] [CrossRef]
- Milanski, M.; Degasperi, G.; Coope, A.; Morari, J.; Denis, R.; Cintra, D.E.; Tsukumo, D.M.L.; Anhe, G.; Amaral, M.E.; Takahashi, H.K.; et al. Saturated Fatty Acids Produce an Inflammatory Response Predominantly through the Activation of TLR4 Signaling in Hypothalamus: Implications for the Pathogenesis of Obesity. Journal of Neuroscience 2009, 29. [Google Scholar] [CrossRef] [PubMed]
- Barale, C.; Russo, I. Influence of Cardiometabolic Risk Factors on Platelet Function. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Kesika, P.; Suganthy, N.; Chaiyasut, C. A Review on Role of Microbiome in Obesity and Antiobesity Properties of Probiotic Supplements. Biomed Res Int 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Chakraborti, C.K. New-Found Link between Microbiota and Obesity. World J Gastrointest Pathophysiol 2015, 6. [Google Scholar] [CrossRef]
- Crespy, V.; Williamson, G. A Review of the Health Effects of Green Tea Catechins in in Vivo Animal Models. In Proceedings of the Journal of Nutrition; 2004; Vol. 134.
- Lin, J.K.; Lin-Shiau, S.Y. Mechanisms of Hypolipidemic and Anti-Obesity Effects of Tea and Tea Polyphenols. Mol Nutr Food Res 2006, 50. [Google Scholar] [CrossRef]
- Bajerska, J.; Wozniewicz, M.; Jeszka, J.; Drzymala-Czyz, S.; Walkowiak, J. Green Tea Aqueous Extract Reduces Visceral Fat and Decreases Protein Availability in Rats Fed with a High-Fat Diet. Nutrition Research 2011, 31. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Sanchez, K.; Leyva, M.J.; Wu, M.; Betts, N.M.; Aston, C.E.; Lyons, T.J. Green Tea Supplementation Affects Body Weight, Lipids, and Lipid Peroxidation in Obese Subjects with Metabolic Syndrome. J Am Coll Nutr 2010, 29. [Google Scholar] [CrossRef] [PubMed]
- Kamalakkannan, S.; Rajendran, R.; Venkatesh, R. V.; Clayton, P.; Akbarsha, M.A. Antiobesogenic and Antiatherosclerotic Properties of Caralluma Fimbriata Extract. J Nutr Metab 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- Banin, R.M.; Hirata, B.K.S.; Andrade, I.S.; Zemdegs, J.C.S.; Clemente, A.P.G.; Dornellas, A.P.S.; Boldarine, V.T.; Estadella, D.; Albuquerque, K.T.; Oyama, L.M.; et al. Beneficial Effects of Ginkgo Biloba Extract on Insulin Signaling Cascade, Dyslipidemia, and Body Adiposity of Diet-Induced Obese Rats. Brazilian Journal of Medical and Biological Research 2014, 47. [Google Scholar] [CrossRef] [PubMed]
- Hirata, B.K.S.; Banin, R.M.; Dornellas, A.P.S.; De Andrade, I.S.; Zemdegs, J.C.S.; Caperuto, L.C.; Oyama, L.M.; Ribeiro, E.B.; Telles, M.M. Ginkgo Biloba Extract Improves Insulin Signaling and Attenuates Inflammation in Retroperitoneal Adipose Tissue Depot of Obese Rats. Mediators Inflamm 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Banin, R.M.; de Andrade, I.S.; Cerutti, S.M.; Oyama, L.M.; Telles, M.M.; Ribeiro, E.B. Ginkgo Biloba Extract (GbE) Stimulates the Hypothalamic Serotonergic System and Attenuates Obesity in Ovariectomized Rats. Front Pharmacol 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Hirata, B.K.S.; Cruz, M.M.; De Sá, R.D.C.C.; Farias, T.S.M.; Machado, M.M.F.; Bueno, A.A.; Alonso-Vale, M.I.C.; Telles, M.M. Potential Anti-Obesogenic Effects of Ginkgo Biloba Observed in Epididymal White Adipose Tissue of Obese Rats. Front Endocrinol (Lausanne) 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.K.; Park, S.B.; Lee, D.R.; Lee, H.J.; Jin, Y.Y.; Yang, S.H.; Suh, J.W. Green Coffee Bean Extract Improves Obesity by Decreasing Body Fat in High-Fat Diet-Induced Obese Mice. Asian Pac J Trop Med 2016, 9. [Google Scholar] [CrossRef]
- Seliem, E.M.; Azab, M.E.; Ismail, R.S.; Nafeaa, A.A.; Alotaibi, B.S.; Negm, W.A. Green Coffee Bean Extract Normalize Obesity-Induced Alterations of Metabolic Parameters in Rats by Upregulating Adiponectin and GLUT4 Levels and Reducing RBP-4 and HOMA-IR. Life 2022, 12. [Google Scholar] [CrossRef]
- Pamplona Mosimann, A.L.; Wilhelm-Filho, D.; Da Silva, E.L. Aqueous Extract of Ilex Paraguariensis Attenuates the Progression of Atherosclerosis in Cholesterol-Fed Rabbits. BioFactors 2006, 26. [Google Scholar] [CrossRef]
- Pang, J.; Choi, Y.; Park, T. Ilex Paraguariensis Extract Ameliorates Obesity Induced by High-Fat Diet: Potential Role of AMPK in the Visceral Adipose Tissue. Arch Biochem Biophys 2008, 476. [Google Scholar] [CrossRef]
- Lima, N.D.S.; Franco, J.G.; Peixoto-Silva, N.; Maia, L.A.; Kaezer, A.; Felzenszwalb, I.; De Oliveira, E.; De Moura, E.G.; Lisboa, P.C. Ilex Paraguariensis (Yerba Mate) Improves Endocrine and Metabolic Disorders in Obese Rats Primed by Early Weaning. Eur J Nutr 2014, 53. [Google Scholar] [CrossRef] [PubMed]
- Gugliucci, A.; Bastos, D.H.M.; Schulze, J.; Souza, M.F.F. Caffeic and Chlorogenic Acids in Ilex Paraguariensis Extracts Are the Main Inhibitors of AGE Generation by Methylglyoxal in Model Proteins. Fitoterapia 2009, 80. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Oh, M.R.; Kim, M.G.; Chae, H.J.; Chae, S.W. Anti-Obesity Effects of Yerba Mate (Ilex Paraguariensis): A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. BMC Complement Altern Med 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Kwon, E.Y.; Kim, J.W.; Lee, Y.; Ryu, R.; Yun, J.; Kim, M.; Choi, M.S. Intervention Study on the Efficacy and Safety of Platycodon Grandiflorus Ethanol Extract in Overweight or Moderately Obese Adults: A Single-Center, Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Reguero, M.; de Cedrón, M.G.; Reglero, G.; Quintela, J.C.; de Molina, A.R. Natural Extracts to Augment Energy Expenditure as a Complementary Approach to Tackle Obesity and Associated Metabolic Alterations. Biomolecules 2021, 11. [Google Scholar] [CrossRef]
- Mabrouki, L.; Rjeibi, I.; Taleb, J.; Zourgui, L. Cardiac Ameliorative Effect of Moringa Oleifera Leaf Extract in High-Fat Diet-Induced Obesity in Rat Model. Biomed Res Int 2020, 2020. [Google Scholar] [CrossRef]
- Pimentel, G.D.; Lira, F.S.; Rosa, J.C.; Caris, A. V.; Pinheiro, F.; Ribeiro, E.B.; Oller do Nascimento, C.M.; Oyama, L.M. Yerba Mate Extract (Ilex Paraguariensis) Attenuates Both Central and Peripheral Inflammatory Effects of Diet-Induced Obesity in Rats. Journal of Nutritional Biochemistry 2013, 24. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.L.; Shibu, M.A.; Lii, C.K.; Viswanadha, V.P.; Lin, Y.L.; Lai, C.H.; Chen, Y.F.; Lin, K.H.; Kuo, W.W.; Huang, C.Y. Andrographis Paniculata Extract Attenuates Pathological Cardiac Hypertrophy and Apoptosis in High-Fat Diet Fed Mice. J Ethnopharmacol 2016, 192. [Google Scholar] [CrossRef]
- Sikora, J.; Markowicz-Piasecka, M.; Broncel, M.; Mikiciuk-Olasik, E. Extract of Aronia Melanocarpa-Modified Hemostasis: In Vitro Studies. Eur J Nutr 2014, 53. [Google Scholar] [CrossRef]
- Sikora, J.; Broncel, M.; Markowicz, M.; Chałubiński, M.; Wojdan, K.; Mikiciuk-Olasik, E. Short-Term Supplementation with Aronia Melanocarpa Extract Improves Platelet Aggregation, Clotting, and Fibrinolysis in Patients with Metabolic Syndrome. Eur J Nutr 2012, 51. [Google Scholar] [CrossRef] [PubMed]
- Allison, G.L.; Lowe, G.M.; Rahman, K. Aged Garlic Extract Inhibits Platelet Activation by Increasing Intracellular CAMP and Reducing the Interaction of GPIIb/IIIa Receptor with Fibrinogen. Life Sci 2012, 91. [Google Scholar] [CrossRef]
- Li, W.; Luo, Z.; Liu, X.; Fu, L.; Xu, Y.; Wu, L.; Shen, X. Effect of Ginkgo Biloba Extract on Experimental Cardiac Remodeling. BMC Complement Altern Med 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Caro-Gómez, E.; Sierra, J.A.; Escobar, J.S.; Álvarez-Quintero, R.; Naranjo, M.; Medina, S.; Velásquez-Mejía, E.P.; Tabares-Guevara, J.H.; Jaramillo, J.C.; León-Varela, Y.M.; et al. Green Coffee Extract Improves Cardiometabolic Parameters and Modulates Gut Microbiota in High-Fat-Diet-Fed ApoE -/- Mice. Nutrients 2019, 11. [Google Scholar] [CrossRef]
- Diez-Echave, P.; Vezza, T.; Rodríguez-Nogales, A.; Ruiz-Malagón, A.J.; Hidalgo-García, L.; Garrido-Mesa, J.; Molina-Tijeras, J.A.; Romero, M.; Robles-Vera, I.; Pimentel-Moral, S.; et al. The Prebiotic Properties of Hibiscus Sabdariffa Extract Contribute to the Beneficial Effects in Diet-Induced Obesity in Mice. Food Research International 2020, 127. [Google Scholar] [CrossRef] [PubMed]
- Evseeva, M.N.; Balashova, M.S.; Kulebyakin, K.Y.; Rubtsov, Y.P. Adipocyte Biology from the Perspective of in Vivo Research: Review of Key Transcription Factors. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Sun, C.; Mao, S.; Chen, S.; Zhang, W.; Liu, C. Ppars-Orchestrated Metabolic Homeostasis in the Adipose Tissue. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Savova, M.S.; Mihaylova, L. V.; Tews, D.; Wabitsch, M.; Georgiev, M.I. Targeting PI3K/AKT Signaling Pathway in Obesity. Biomedicine and Pharmacotherapy 2023, 159. [Google Scholar] [CrossRef]
- Ahmad, B.; Serpell, C.J.; Fong, I.L.; Wong, E.H. Molecular Mechanisms of Adipogenesis: The Anti-Adipogenic Role of AMP-Activated Protein Kinase. Front Mol Biosci 2020, 7. [Google Scholar] [CrossRef]
- Kim, N.H.; Jegal, J.; Kim, Y.N.; Heo, J.D.; Rho, J.R.; Yang, M.H.; Jeong, E.J. Chokeberry Extract and Its Active Polyphenols Suppress Adipogenesis in 3T3-L1 Adipocytes and Modulates Fat Accumulation and Insulin Resistance in Diet-Induced Obese Mice. Nutrients 2018, 10. [Google Scholar] [CrossRef]
- Oh, J.; Ahn, S.H.; Zhou, X.; Lim, Y.J.; Hong, S.; Kim, H.S. Effects of Cinnamon (Cinnamomum Zeylanicum) Extract on Adipocyte Differentiation in 3T3-L1 Cells and Lipid Accumulation in Mice Fed a High-Fat Diet. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.L.; Jeon, Y.D.; Park, J.; Rim, H.K.; Jeong, M.Y.; Lim, H.; Ko, S.G.; Jang, H.J.; Lee, B.C.; Lee, K.T.; et al. Corni Fructus Containing Formulation Attenuates Weight Gain in Mice with Diet-Induced Obesity and Regulates Adipogenesis through AMPK. Evidence-based Complementary and Alternative Medicine 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; So, H.; Youn, M.J.; Kim, H.J.; Kim, Y.; Park, C.; Kim, S.J.; Ha, Y.A.; Chai, K.Y.; Kim, S.M.; et al. Hibiscus Sabdariffa L. Water Extract Inhibits the Adipocyte Differentiation through the PI3-K and MAPK Pathway. J Ethnopharmacol 2007, 114. [Google Scholar] [CrossRef]
- Kudo, M.; Gao, M.; Hayashi, M.; Kobayashi, Y.; Yang, J.; Liu, T. Ilex Paraguariensis A.St.-Hil. Improves Lipid Metabolism in High-Fat Diet-Fed Obese Rats and Suppresses Intracellular Lipid Accumulation in 3T3-L1 Adipocytes via the AMPK-Dependent and Insulin Signaling Pathways. Food Nutr Res 2024, 68. [Google Scholar] [CrossRef] [PubMed]
- Héliès-Toussaint, C.; Fouché, E.; Naud, N.; Blas-Y-Estrada, F.; del Socorro Santos-Diaz, M.; Nègre-Salvayre, A.; Barba de la Rosa, A.P.; Guéraud, F. Opuntia Cladode Powders Inhibit Adipogenesis in 3 T3-F442A Adipocytes and a High-Fat-Diet Rat Model by Modifying Metabolic Parameters and Favouring Faecal Fat Excretion. BMC Complement Med Ther 2020, 20. [Google Scholar] [CrossRef]
- Park, Y.S.; Yoon, Y.; Ahn, H.S. Platycodon Grandiflorum Extract Represses Up-Regulated Adipocyte Fatty Acid Binding Protein Triggered by a High Fat Feeding in Obese Rats. World J Gastroenterol 2007, 13. [Google Scholar] [CrossRef] [PubMed]
- Ezenwaka, C.E.; Okoye, O.; Esonwune, C.; Onuoha, P.; Dioka, C.; Osuji, C.; Oguejiofor, C.; Meludu, S. High Prevalence of Abdominal Obesity Increases the Risk of the Metabolic Syndrome in Nigerian Type 2 Diabetes Patients: Using the International Diabetes Federation Worldwide Definition. Metab Syndr Relat Disord 2014, 12. [Google Scholar] [CrossRef]
- Lee, S.; Joo, H.; Kim, C.T.; Kim, I.H.; Kim, Y. High Hydrostatic Pressure Extract of Garlic Increases the HDL Cholesterol Level via Up-Regulation of Apolipoprotein A-I Gene Expression in Rats Fed a High-Fat Diet. Lipids Health Dis 2012, 11. [Google Scholar] [CrossRef]
- Kannel, W.B. Prevalence and Natural History of Electrocardiographic Left Ventricular Hypertrophy. Am J Med 1983, 75. [Google Scholar] [CrossRef]
- Ferreira, M.C.L.; Lima, L.N.; Cota, L.H.T.; Costa, M.B.; Orsi, P.M.E.; Espíndola, R.P.; Albanez, A. V.; Rosa, B.B.; Carvalho, M.G.S.; Garcia, J.A.D. Effect of Camellia Sinensis Teas on Left Ventricular Hypertrophy and Insulin Resistance in Dyslipidemic Mice. Brazilian Journal of Medical and Biological Research 2020, 53. [Google Scholar] [CrossRef]
- Zamani, M.; Kelishadi, M.R.; Ashtary-Larky, D.; Amirani, N.; Goudarzi, K.; Torki, I.A.; Bagheri, R.; Ghanavati, M.; Asbaghi, O. The Effects of Green Tea Supplementation on Cardiovascular Risk Factors: A Systematic Review and Meta-Analysis. Front Nutr 2023, 9. [Google Scholar] [CrossRef] [PubMed]
- Balzan, S.; Hernandes, A.; Reichert, C.L.; Donaduzzi, C.; Pires, V.A.; Gasparotto, A.; Cardozo, E.L. Lipid-Lowering Effects of Standardized Extracts of Ilex Paraguariensis in High-Fat-Diet Rats. Fitoterapia 2013, 86. [Google Scholar] [CrossRef]
- Gao, H.; Long, Y.; Jiang, X.; Liu, Z.; Wang, D.; Zhao, Y.; Li, D.; Sun, B. liang Beneficial Effects of Yerba Mate Tea (Ilex Paraguariensis) on Hyperlipidemia in High-Fat-Fed Hamsters. Exp Gerontol 2013, 48. [Google Scholar] [CrossRef]
- Boaventura, B.C.B.; Di Pietro, P.F.; Stefanuto, A.; Klein, G.A.; de Morais, E.C.; de Andrade, F.; Wazlawik, E.; da Silva, E.L. Association of Mate Tea (Ilex Paraguariensis) Intake and Dietary Intervention and Effects on Oxidative Stress Biomarkers of Dyslipidemic Subjects. Nutrition 2012, 28. [Google Scholar] [CrossRef] [PubMed]
- Masson, W.; Barbagelata, L.; Lobo, M.; Nogueira, J.P.; Corral, P.; Lavalle-Cobo, A. Effect of Yerba Mate (Ilex Paraguariensis) on Lipid Levels: A Systematic Review and Meta-Analysis. Plant Foods for Human Nutrition 2022. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, P.; Weiskirchen, R. The Role of Obesity in Type 2 Diabetes Mellitus—An Overview. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Sudhakara, G.; Mallaiah, P.; Sreenivasulu, N.; Sasi Bhusana Rao, B.; Rajendran, R.; Saralakumari, D. Beneficial Effects of Hydro-Alcoholic Extract of Caralluma Fimbriata against High-Fat Diet-Induced Insulin Resistance and Oxidative Stress in Wistar Male Rats. J Physiol Biochem 2014, 70. [Google Scholar] [CrossRef] [PubMed]
- Drissi, F.; Lahfa, F.; Gonzalez, T.; Peiretti, F.; Tanti, J.F.; Haddad, M.; Fabre, N.; Govers, R. A Citrullus Colocynthis Fruit Extract Acutely Enhances Insulin-Induced GLUT4 Translocation and Glucose Uptake in Adipocytes by Increasing PKB Phosphorylation. J Ethnopharmacol 2021, 270. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, B.H.; Kim, Y.C. Antioxidative Action of Corni Fructus Aqueous Extract on Kidneys of Diabetic Mice. Toxicol Res 2011, 27. [Google Scholar] [CrossRef]
- Khalili-Moghadam, S.; Hedayati, M.; Golzarand, M.; Mirmiran, P. Effects of Green Coffee Aqueous Extract Supplementation on Glycemic Indices, Lipid Profile, CRP, and Malondialdehyde in Patients with Type 2 Diabetes: A Randomized, Double-Blind, Placebo-Controlled Trial. Front Nutr 2023, 10. [Google Scholar] [CrossRef]
- Peng, C.H.; Chyau, C.C.; Chan, K.C.; Chan, T.H.; Wang, C.J.; Huang, C.N. Hibiscus Sabdariffa Polyphenolic Extract Inhibits Hyperglycemia, Hyperlipidemia, and Glycation-Oxidative Stress While Improving Insulin Resistance. J Agric Food Chem 2011, 59. [Google Scholar] [CrossRef]
- Leem, K.H.; Kim, M.G.; Hahm, Y.T.; Kim, H.K. Hypoglycemic Effect of Opuntia Ficus-Indica Var. Saboten Is Due to Enhanced Peripheral Glucose Uptake through Activation of AMPK/P38 MAPK Pathway. Nutrients 2016, 8. [Google Scholar] [CrossRef]
- Hall, J.E.; Do Carmo, J.M.; Da Silva, A.A.; Wang, Z.; Hall, M.E. Obesity-Induced Hypertension: Interaction of Neurohumoral and Renal Mechanisms. Circ Res 2015, 116. [Google Scholar] [CrossRef]
- Shariq, O.A.; Mckenzie, T.J. Obesity-Related Hypertension: A Review of Pathophysiology, Management, and the Role of Metabolic Surgery. Gland Surg 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Valls, R.M.; Companys, J.; Calderón-Pérez, L.; Salamanca, P.; Pla-Pagà, L.; Sandoval-Ramírez, B.A.; Bueno, A.; Puzo, J.; Crescenti, A.; Del Bas, J.M.; et al. Effects of an Optimized Aged Garlic Extract on Cardiovascular Disease Risk Factors in Moderate Hypercholesterolemic Subjects: A Randomized, Crossover, Double-Blind, Sustainedand Controlled Study. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Tan, B. Hypotensive Activity of Aqueous Extract of Andrographis Paniculata in Rats. Clin Exp Pharmacol Physiol 1996, 23. [Google Scholar] [CrossRef]
- Hawkins, J.; Hires, C.; Baker, C.; Keenan, L.; Bush, M. Daily Supplementation with Aronia Melanocarpa (Chokeberry) Reduces Blood Pressure and Cholesterol: A Meta Analysis of Controlled Clinical Trials. J Diet Suppl 2021, 18. [Google Scholar] [CrossRef] [PubMed]
- Antonello, M.; Montemurro, D.; Bolognesi, M.; Di Pascoli, M.; Piva, A.; Grego, F.; Sticchi, D.; Giuliani, L.; Garbisa, S.; Rossi, G.P. Prevention of Hypertension, Cardiovascular Damage and Endothelial Dysfunction with Green Tea Extracts. Am J Hypertens 2007, 20. [Google Scholar] [CrossRef]
- Onakpoya, I.; Spencer, E.; Heneghan, C.; Thompson, M. The Effect of Green Tea on Blood Pressure and Lipid Profile: A Systematic Review and Meta-Analysis of Randomized Clinical Trials. Nutrition, Metabolism and Cardiovascular Diseases 2014, 24. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, L. de P.; Nogueira Neto, J.F.; Klein, M.R.S.T.; Sanjuliani, A.F. Short-Term Effects of Green Tea on Blood Pressure, Endothelial Function, and Metabolic Profile in Obese Prehypertensive Women: A Crossover Randomized Clinical Trial. J Am Coll Nutr 2017, 36. [Google Scholar] [CrossRef]
- Liang, H.; Yuan, X.; Sun, C.; Sun, Y.; Yang, M.; Feng, S.; Yao, J.; Liu, Z.; Zhang, G.; Li, F. Preparation of a New Component Group of Ginkgo Biloba Leaves and Investigation of the Antihypertensive Effects in Spontaneously Hypertensive Rats. Biomedicine and Pharmacotherapy 2022, 149. [Google Scholar] [CrossRef] [PubMed]
- Inuwa, I.; Ali, B.H.; Al-Lawati, I.; Beegam, S.; Ziada, A.; Blunden, G. Long-Term Ingestion of Hibiscus Sabdariffa Calyx Extract Enhances Myocardial Capillarization in the Spontaneously Hypertensive Rat. Exp Biol Med 2012, 237. [Google Scholar] [CrossRef] [PubMed]
- Nwachukwu, D.C.; Aneke, E.I.; Obika, L.F.; Nwachukwu, N.Z. Effects of Aqueous Extract of Hibiscus Sabdariffa on the Renin-Angiotensin-Aldosterone System of Nigerians with Mild to Moderate Essential Hypertension: A Comparative Study with Lisinopril. Indian J Pharmacol 2015, 47. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, S.M.; Ukeyima, M.T.; Spencer, J.P.E.; Lovegrove, J.A. Acute Effects of Hibiscus Sabdariffa Calyces on Postprandial Blood Pressure, Vascular Function, Blood Lipids, Biomarkers of Insulin Resistance and Inflammation in Humans. Nutrients 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Ignat, M.V.; Coldea, T.E.; Salanță, L.C.; Mudura, E. Plants of the Spontaneous Flora with Beneficial Action in the Management of Diabetes, Hepatic Disorders, and Cardiovascular Disease. Plants 2021, 10. [Google Scholar] [CrossRef]
- Di Cesare, M.; McGhie, D.V.; Perel, P.; Mwangi, J.; Taylor, S.; Pervan, B.; Kabudula, C.; Narula, J.; Bixby, H.; Pineiro, D.; et al. The Heart of the World. Glob Heart 2024, 19. [Google Scholar] [CrossRef]
Figure 1.
Overview of obesity. Obesity is associated with the development of cardiometabolic risk factors and cardiovascular diseases. However, healthy lifestyle and intake of plant extracts may prevent theses pathological conditions.
Figure 1.
Overview of obesity. Obesity is associated with the development of cardiometabolic risk factors and cardiovascular diseases. However, healthy lifestyle and intake of plant extracts may prevent theses pathological conditions.
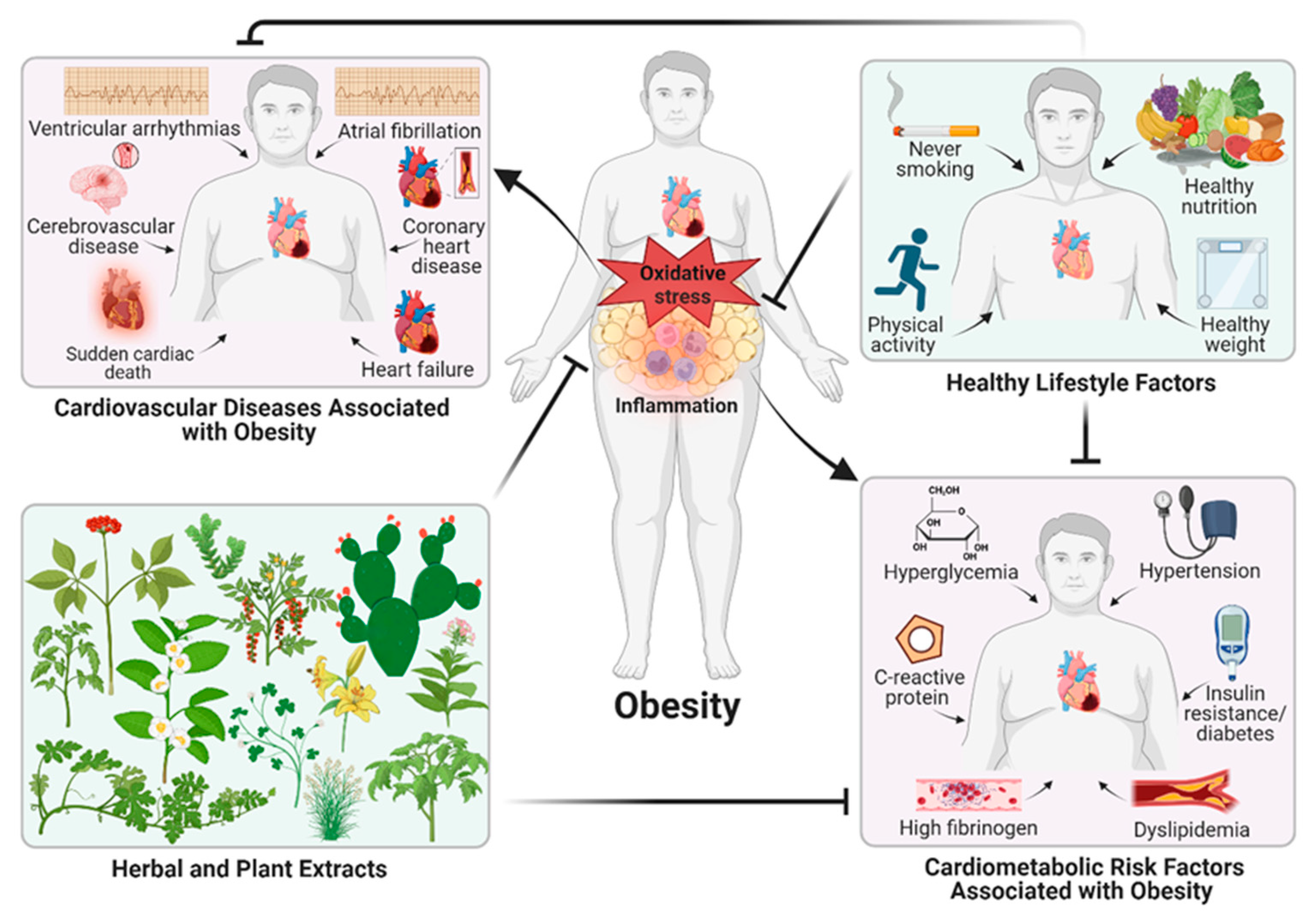
Table 2.
Extracts from different herbs and plants targeting dyslipidemia and adipokines in obesity.
| Name of herbs and plants and Method Extraction | Type of Study | Doses and Duration | Outcomes |
|---|---|---|---|
| Allium sativum (garlic), high hydrostatic pressure extract | 5-week-old male Sprague-Dawley rats with HFD (45% kcal from fat) |
2% (w/w) of extract for 5 weeks | Decreased in plasma TG and LDL-C levels, increased in HDL-C levels, reduced hepatic TG and TC levels, upregulated hepatic apoA-I, ABCA1, and LCAT gene expression [160] |
| Camellia sinensis, teas (green, red, and white) |
3-month-old male LDLr-/- mice with HFD (20% fat with 1.25% cholesterol, and 0.5% cholic acid) |
25 mg/kg body weight daily for 60 days | Prevented left ventricular hypertrophy, partially prevented hyperlipidemia and insulin resistance, and reduced CRP levels [162] |
| Camellia sinensis, green tea extract (GTE) |
Systematic review and meta-analysis of randomized clinical trials |
Varied dosages, some ≥1000 mg/day, others <1000 mg/day, and durations with subgroup analyses based on ≥12 weeks vs. <12 weeks |
Significant reduced total cholesterol (TC) and LDL-C. Decreased fasting blood sugar, and HbA1c. Small increased HDL-C. Reduced diastolic blood pressure [163] |
| Corni Fructus, extract produced by Tsumura Juntendo Inc. (Tokyo, Japan) |
5-week-old male Wistar rats with a high cholesterol diet (1% cholesterol and 0.5% cholic acid) |
50, 100, and 200 mg/kg/day for 10 days | Lowered blood pressure and serum cholesterol levels. Decreased atherogenic index, increased cholesterol and bile acid excretion. Reduced lipid peroxidation, up-regulated SREBP-2 and PPARα expression, and enhanced fatty acid oxidation [40] |
| Cydonia oblonga (ethanol extract) | Male Sprague Dawley rats (240 ± 20 g) induced with hyperlipidemia | Hyperlipidemia induction for 21 days and then with 40, 80, 160 mg/kg/day for 4 weeks |
Reduced serum TC, TG, LDL-C, ALT, AST, increased HDL-C, reduced MDA, improved SOD and GSH-Px activity in hepatic tissues [45] |
| Ilex paraguariensis (Yerba mate), hydroethanolic extract and n-butanolic fraction | 8-week-old male Wistar rats with HFD (60% kcal from fat) with cholesterol (2%) and cholic acid (0.2%) |
Hyperlipidemia induction for 30 days and then with 200, 400, 800 mg/kg/day for 30 days | Reduced serum TG, cholesterol, and atherogenic index [164] |
| Ilex paraguariensis (Yerba mate), aqueous extract | A systematic review and meta-analysis |
Various doses in included studies | No significant change in TC, LDL-C, HDL-C, and TG levels [167] |
| Ilex paraguariensis (Yerba mate), aqueous extract |
8-week-old male Syrian golden hamsters with HFD (15% lard and 0.2% cholesterol) |
Hyperlipidemia induction for 4 weeks and then with 1%, 2%, and 4% w/v for 4 weeks |
Decreased body weight gain, lowered serum lipid levels, increased antioxidant enzyme activity, improved lipoprotein lipase (LPL) and hepatic lipase (HL) activities, and upregulated PPARα and LDL-C receptor mRNA expression. Reduced SREBP-1c and acetyl CoA carboxylase mRNA expression [165] |
| Ilex paraguariensis (Yerba mate), aqueous extract |
Randomized clinical trial with dyslipidemic individuals | 1 L/day (20 mg/mL) for 90 days | Increased serum antioxidant capacity and GSH, and decreased LDL-C [166] |
| Moringa oleífera (aqueous extract) | Male Wistar rats with HFD (3% fat) | 1 mg/g for 30 days | Decreased cholesterol levels in serum, liver, and kidney. Increased serum albumin [76] |
| Nigella sativa, seed powder, seed oil, and seed (methanolic extract) |
Systematic review of experimental and clinical studies |
Variable treatment time of seed powder (100 mg-20 g daily), seed oil (20-800 mg daily), and seed extract (6, 9, 14, and 21 g/kg) | Reduced TC, LDL-C, and TG. No significant effect on HDL-C [78] |
| Opuntia ficus-indica (aqueous extract) |
Triton-induced hypercholesterolemia in male Balb-c mice |
500 mg/kg in a single administration for 16 hours plus fasting for 8 hours | Significantly decreased cholesterol levels. Inhibited pancreatic lipase with IC50 = 588.5 μg/mL [83] |
| Platycodon grandiflorus (water extract) |
9-week-old male C57BL/6J mice with HFD |
1 g/kg/day for 8 weeks | Reduced body weight gain by 7.5%, improved plasma lipid profiles, decreased leptin, increased adiponectin, downregulated lipogenic gene expression, increased lipolysis gene expression, and inhibited adipogenic transcription factors [88] |
| Platycodon grandiflorus (70% ethanol extract) |
5-week-old male C57BL/6J mice with HFD (40% of fat) |
Dyslipidemia induction for 5 weeks and then with 25 and 75 mg/kg/day for 4 weeks |
Reduced plasma and hepatic lipid levels, upregulated antioxidant proteins, inhibited oxLDL-C-induced cell death and lactate dehydrogenase release, exhibited antioxidant activity in vitro and in vivo [89] |
| Platycodon grandiflorus, extract (water, 50% ethanol, and 80% ethanol) |
L6 muscle cells and 9-weeks-old male ICR mice with HFD (60% kcal from fat) |
1% and 5% extract in diet for 6 weeks | Reduced food intake, body weight, epididymal fat weight, adipocyte size,and blood glucose levels. Maintained serum adiponectin, resistin, leptin, fructosamine, and triglycerides. Upregulated adiponectin mRNA, downregulated TNF-a and leptin mRNA in WAT. In L6 muscle cells increased insulin-stimulated glucose uptake [90] |
Table 3.
Extracts from different herbs and plants targeting insulin resistance, hyperglycemia, and diabetes.
Table 3.
Extracts from different herbs and plants targeting insulin resistance, hyperglycemia, and diabetes.
| Name of herbs and plants and Method Extraction | Type of Study | Doses and Duration | Outcomes |
|---|---|---|---|
| Caralluma fimbriata (hydro-alcoholic extract) |
Male Wistar rats with HFD (60% of fat) |
200 mg/kg/day for 90 days |
Reduced hyperglycemia, hyperinsulinemia, hyperleptinemia, hypertriglyceridemia, oxidative stress, and improved insulin sensitivity [169] |
| Ceylon Cinnamon (hydro-alcoholic extract) |
Pancreatic alpha-amylase activity. 7-week-old male Wistar Han IGS rats. A randomized, placebo-controlled, cross-over clinical trial in healthy subjects |
In vitro: 0-100 µg/ml. In vivo: 6.25, 12.5, 25, 50, 100 mg/kg for 5 weeks. Humans: 1 g of extract (two 500 mg capsules), single dose post-meal |
In vitro: Inhibited pancreatic alpha-amylase (IC50 = 25 μg/mL). In vivo: Reduced glycemic response to starch. Human: Lowered postprandial glycemia by 14.8% (0-120 min) and 21.2% (0-60 min) without increasing insulin secretion [30] |
| Citrullus colocynthis (petroleum ether, water or 80% methanol, ethyl acetate and n-butanol, crude aqueous extracts) | 3T3-L1 adipocytes | 4, 20, or 100 μg/ml for 24, 48, and 96 hours |
Enhanced insulin-induced GLUT4 translocation and glucose uptake, and increased insulin-induced PKB phosphorylation [170]. |
| Citrullus colocynthis (Tablets, Capsules, or Oral Drops) |
Randomized Controlled Clinical Trials | Different doses for 30 to 60 days |
No significant effect on FBS, HbA1c, LDL-C, TC, and TG. Increased HDL-C levels [32] |
| Corni fructus (water extract) | 7-week-old male C57BUKsJ-db/db mice and C57BL/6 mice | 500 mg/kg/day for 8 weeks | Reduced blood glucose levels, improved insulin resistance, and increased glucose utilization [41] |
| Corni fructus (aqueous extract) |
7-week-old male C57BL/KsJ-db/db mice | 500 mg/kg/day for 8 weeks |
Reduced oxidative stress, increased SOD activity, decreased XO, CAT, and GST activities. Lower mRNA expression of eNOS in kidneys [171] |
| Ginkgo biloba (aqueous and 12% ethanol extracts) |
α-amylase and α-glucosidase activities |
10, 25 and 50 mg/ml of Ginkgo leaf extract |
Aqueous extracts had higher total phenolic content but only ethanolic extracts inhibited ACE, strong correlation between total phenolics and α-glucosidase inhibitory activity, and to a lesser degree positive correlation between total phenolics and α-amylase inhibitory activity [50] |
| Ginkgo biloba (EGb761) | RAoSMCs and HUVECs. 5-week-old male Otsuka Long-Evans Tokushima Fatty rats and 5-week-old male ApoE-/- mice |
Obesity and insulin resistance induction for 24 weeks (rats). 2 months all mice with HFD (42% fat, 1.25% cholesterol). All animals with 100 mg/kg and 200 mg/kg for 6 weeks (rats), and for 2 months (mice) |
Reduced intima-media ratio. Induced greater apoptosis in rats, improved glucose homeostasis and increased circulating adiponectin levels, decreased plasma hsCRP concentrations. In vitro: Decreased VSMC proliferation and migration, Increased caspase-3 activity and DNA fragmentation, decreased monocyte adhesion and ICAM-1/VCAM-1 levels. Kaempferol and quercetin: Reduced VSMC migration and increased caspase activity, and protected against atherosclerosis [51] |
| Green Coffee (Coffea), aqueous extract) |
A randomized, double-blind, placebo-controlled trial |
400 mg (capsules) twice per day for 10 weeks |
Decreased SBP, TG, hs-CRP, increased HDL-C, and marginally significant reduction in FBG. No significant changes in DBP, LDL-C, TC, insulin levels, HOMA-IR, and MDA [172] |
| Hibiscus sabdariffa, (polyphenolic extract by methanol) | 8-week-old male Sprague-Dawley rats with HFD and STZ |
Type 2 diabetes induction: HFD for 7 weeks and then HFD and STZ for 2 weeks. Doses 100 mg/kg and 200 mg/kg for 7 weeks | Reduced hyperglycemia, hyperinsulinemia, serum TG, cholesterol, and LDL-C/HDL-C ratio. Decreased plasma AGE formation and lipid peroxidation. Inhibited CTGF and RAGE expression in aortic regions. Improved weight loss in diabetic rats [173] |
| Hibiscus sabdariffa (aqueous extract) | α-amylase and α-glucosidase activities | Red and white varieties; IC50 values: 25.2 µg/mL (red) and 47.4 µg/mL (white) for α-glucosidase inhibition; 90.5 µg/mL (white) and 187.9 µg/mL (red) for α-amylase inhibition |
Both varieties inhibited α-amylase and α-glucosidase activities, red variety exhibited higher α-glucosidase inhibitory activity, while the white variety showed higher α-amylase inhibitory activity, strong antioxidant properties, particularly in the red variety [63] |
| Hibiscus sabdariffa (aqueous extract) | 3T3-L1 cells and male Sprague Dawley rats (100-120 g) with HFD | In vitro: 0.1, 0.5, 1 mg/ml. In vivo: 250 and 500 mg/kg/day for 8 weeks | Reduced body weight, food intake, lipid profiles, inflammatory cytokines, lipid peroxidation, serum leptin, insulin, and duodenal glucose absorption. Increased glucose uptake in adipose tissue and muscle, downregulated adipogenic gene expression [62] |
| Ilex paraguariensis (Yerba mate), aqueous extract |
T2DM and pre-diabetes subjects | 330 mL of roasted mate tea 3 times a day for 60 days | T2DM: Significant decrease in fasting glucose, HbA1c, and LDL-C. Pre-diabetes: Significant decrease in LDL-C, non-HDL-C, and TG. Improved glycemic control and lipid profile, reduced risk of coronary disease [70] |
| Moringa oleifera (dry leaf powder) | A double-blind, randomized, placebo controlled, parallel group clinical trial |
2400 mg/day (6 capsules/day) for 12 weeks |
Significant decrease FBG and HbA1c. No significant changes in microbiota, hepatic and renal function markers, or appetite-controlling hormones [77] |
| Opuntia ficus-indica var. saboten (hot water extract) | α-Glucosidase activity. L6 muscle cells. 5-week-old male C57BL/6J db/db mice and their non-diabetic heterozygous littermates (db/-), and 9-week-old male ICR mice |
α-Glucosidase activity (1, 5, 10 mg/ml). L6 muscle cells (1-200 µg/ml). db/db mice (1 and 2 g/kg BW) and db/- mice (1 g/kg BW) for 4 weeks | Inhibited α-glucosidase activity and intestinal glucose absorption. In L6 muscle cells, increased glucose uptake, stimulated AMPK and p38 MAPK phosphorylation, and increased GLUT4. In db/db mice, improved hyperglycemia, hyperinsulinemia, glucose tolerance, and regenerated β-cells [174] |
| Opuntia ficus-indica (cladodes and stem/fruit skin-blend ratio 75/25) hot water extract | Wistar rats either sex weighing 250–350 g |
0.176-176 mg/kg for 180 min and glucose (i.p., 2 g/kg in 5 mL) 30 min after extracts administration | Both extracts lowered blood glucose levels (in doses as low as 6 mg/kg). The blend increased basal plasma insulin levels [85] |
| Punica granatum (methanolic extract) | α-glucosidase activity assay. Zucker diabetic fatty (ZDF) rats and Zucker lean (ZL) rats (14-15 weeks old) |
α-glucosidase activity (200 µl of extract for 5 min). 500 mg/kg body weight, oral in 5% acacia once daily for 2 weeks | Lowered plasma glucose levels in non-fasted ZDF rats, inhibited postprandial hyperglycemia, potent inhibitory effect on α-glucosidase activity (IC50: 1.8 µg/ml) [96] |
| Salvia miltiorrhiza (water extract) | HMEC-1 cells | 10 µg/ml of extract in 30 mM glucose condition for 48 hours | Decreased VEGF mRNA and ROS formation induced by high glucose, and UCP-2 siRNA abolished these effects [100] |
| Salvia miltiorrhiza (different extracts) | Review of preclinical and clinical studies on diabetes and complication |
Not applicable | SM exhibits anti-diabetic activities, including anti-inflammation, anti-oxidation, anti-fibrosis, and anti-apoptosis. Key pathways involved are Wnt/β-catenin, TSP-1/TGF-β1/STAT3, JNK/PI3K/Akt, and others. Main compounds include salvianolic acids and diterpenoid tanshinones [98] |
| Taraxacum officinale (aqueous extract) | α-amylase and α-glucosidase activities |
1, 10, 20, 30 mg/mL | Shade-dried leaves demonstrated potent antidiabetic activity via inhibiting α-amylase and α-glucosidase in a dose-dependent manner [107] |
Table 4.
Extracts from different herbs and plants with anti-hypertensive effects.
| Name of herbs and plants and Method Extraction | Type of Study | Doses and Duration | Outcomes |
|---|---|---|---|
| Allium sativum (aged black garlic extract) |
Randomized, crossover, double-blind, sustained, and controlled study; individuals with moderate hypercholesterolemia |
250 mg (1.25 mg SAC)/tablet/day ABG for 6 weeks, with 3 weeks of washout |
Significantly decreased DBP, particularly in men with a baseline DBP higher than 75 mm Hg, and improved cardiovascular risk factors [177] |
| Andrographis paniculata (aqueous extract) |
Male SHR and WKY rats, aged 14-15 weeks |
2.8, 1.4, 0.7 g/kg for 13 days |
Lowered SBP in SHR and WKY rats, reduced plasma ACE activity and kidney TBA level in SHR. No significant effect on lung ACE activity [178] |
| Aronia melanocarpa (chokeberry), berry extracts | Meta-analysis of controlled clinical trials; included randomized, placebo-controlled trials |
Daily supplementation for an average of 6-8 weeks | Significantly reduces systolic blood pressure and TC, with stronger effects in adults over the age of 50 years [179] |
| Camellia sinensis (Green tea), green tea extract (GTE) |
Crossover, randomized, double-blind, placebo-controlled clinical trial |
3 capsules daily, each containing 500 mg of GTE (260 mg polyphenols per capsule), for 4 weeks with a 2-week washout period between treatments | Significant decrease SBP at 24 hours, daytime, and nighttime in obese prehypertensive women. No significant changes in DBP or other metabolic parameters [182] |
| Camellia sinensis (Green Tea), green tea extract (GTE) | 13-week-old male Sprague-Dawley rats | High dose (700 g/kg/day) or low dose (350 g/kg/day) Ang II dose for 13 days, 6 mg/mL GTE in drinking water | GTE prevented hypertension, left ventricular hypertrophy, vascular remodeling, and endothelial dysfunction induced by high Ang II dose. It blunted increases in oxidative stress markers [180] |
| Camellia sinensis (Green tea), green tea extract (GTE) |
A systematic review and meta-analysis of randomized clinical trials |
Various doses and durations across multiple studies | Green tea epigallocatechins have ACE inhibitor properties. Green tea lowers blood pressure by suppressing NADPH oxidase activity and reducing reactive oxygen species. Some meta-analyses reported beneficial effects on blood vessel dilation and lipid profile [181]. |
| Cocoa (flavanols-rich cocoa extract) | Clinical trial, crossover, randomized, double-blind |
1.4 g of cocoa extract (415 mg flavanols) daily for 4 weeks | Reduced postprandial SBP after daily cocoa extract intake within an energy-restricted diet [36] |
| Ginkgo biloba (Standardized leaf extract, EGb761) |
Male adult Wistar rats (120-160 g), hypertension inducedby L-NAME and hypercholesterolemia induced by 1% cholesterol diet |
100 mg/kg/day orally for 12 weeks | Reduced systolic, diastolic, and mean arterial BP. Improved serum lipid profile, protected against renal injury, reduced renal oxidative stress, nitrosative stress, and inflammation. Decreased renal TNF-α, IL-6, IL-1β, and iNOS protein expressions, and increased eNOS protein expression [52] |
| Ginkgo biloba (new component group of Ginkgo biloba leaves, GBLCG), 50% ethanol extract |
Male Wistar rats and spontaneously hypertensive rats (SHRs), 200 ± 20 g |
4.4, 2.2, and 1.1 mg/kg for 120 days | Reduced blood pressure and improved myocardial hypertrophy by promoting NO synthesis and release in endothelial cells, reducing oxidative stress, inhibiting platelet aggregation, and promoting lesion circulation. The hypotensive activity of GBLCG (4.4 mg/kg) was better than Ginkgo biloba extract [183] |
| Coffea (green coffee bean extract, GCE) hot-water extract | Healthy male volunteers (aged 30 to 50 years), with mild hypertension |
46 mg, 93 mg, or 185 mg of GCE daily for 28 days |
Dose-dependent reduction in SBP. Reduction in DBP was also observed [57] |
| Hibiscus sabdariffa (dried calyx and hibiscus anthocyanins), water extract |
12-week-old male SHR | 10%, 15%, and 20% Hibiscus sabdariffa for 10 weeks. 50, 100, and 200 mg/kg red anthocyanin by oral gavage for 5 days |
Hibiscus sabdariffa reduced SBP, DBP, and LV mass; increased myocardial capillary surface area and length density. Red anthocyanin did not significantly reduce the SBP and DBP [184] |
| Hibiscus sabdariffa (aqueous extract) | Wistar, Wistar-Kyoto (WKY), and SHR of about 16 weeks old |
SHR (EC50 = 0.83 ± 0.08 mg/mL), WKY (EC50 = 0.46 ± 0.04 mg/mL), and Wistar rats (EC50 = 0.44 ± 0.08 mg/mL) | Concentration-dependent relaxant effect on mesenteric arteries and reduced L-type calcium current [64] |
| Hibiscus sabdariffa calyces (HSC), aqueous extract of calyces | A randomized, controlled, single-blinded, acute, cross-over trial | 7.5 g HSC in 250 mL Buxton water, at time 0 min followed by a medium fat lunch at 120 min in a random order separated by a two-week washout period | Significant increase in % flow mediated dilatation, non-significant decrease in SBP and DBP, non-significant increase in urinary and plasma NOx, reduced response of serum glucose, plasma insulin, serum triacylglycerol, and CRP levels. Significant improvement in systemic antioxidant response. No significant changes in arterial stiffness [186] |
| Hibiscus sabdariffa (aqueous extract) | Double-blind randomized controlled trial |
150 mg/kg daily for 4 weeks |
Reduced plasma aldosterone, serum ACE, and increased plasma renin activity [185] |
| Nigella sativa (seed), boiled water extract |
Randomized, double-blind, placebo-controlled trial. Healthy male volunteers with mild hypertension | 100 mg and 200 mg twice a day for 8 weeks | Significant reduction in SBP and DBP in a dose-dependent manner. Reduced TC and LDL-C levels [80] |
| Platycodon grandifloras (roots) (aqueous extract) | H9c2 myoblasts. SHRs and WKYs rats (about 300 g) |
1.25, 2.5, 5 µg/µL for in vitro. 100 and 200 mg/kg/day for 50 days for in vivo | Suppressed Ang II-induced IGF-IIR signaling, reduced cardiomyocyte apoptosis, decreased SBP and DBP in SHRs [91] |
| Punica granatum (pomegranate peel), ethanol (95°GL) extract |
Female SHRs (4 and 28 weeks old) | 25 mg/100 g rat for 30 days |
Reduced SBP, coronary ACE activity, oxidative stress, and vascular remodeling in hypertensive female rats [97] |
| Taraxacum officinale (leaves and roots), 70% ethanol extract | ABTS and FRAP. L-NAME-induced hypertensive Wistar rats (150 g to 200 g), both sexes | 500 mg/kg/day for 21 days | Leaves possessed higher polyphenol and flavonoid, free radical scavenging activity, and total antioxidant capacities. Leaves and roots extract significantly increased total antioxidant capacities (kidney and brain tissues) and reduced MDA levels (heart tissue) [106] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



