
Submitted:
01 July 2024
Posted:
02 July 2024
You are already at the latest version
Abstract
A total of 27 microsatellites markers , scattered all over the genom of barley, were used for characterization,and diversity analysis in 33 barley (Hordeum vulgare L.) accessions and cultivars of Algeria.Over the 27 genetic loci studied, 110 alleles were identified with a mean of 4.1 alleles per locus and the average polymorphism information content (PIC) of 0.46,the marker Ebma0806 showed the greatest value of PIC with 0.83.Genetic diversity at 27 microsatellite loci varied from 0 to 0.84 (Mean=0.50) for 33 Algerian cultivars. The genetic distance among the cultivars ranged from 0.2 to 0.85. Cluster analysis and principal coordinate analysis (PCoA) based on the SSR data clearly differentiate the all genotypes studied.
Keywords:
Genetic diversity
; SSR markers
; barley
; Algeria
1. Introduction
In genetic terms, characterization refers to the detection of variation in either DNA sequences or specific genes. Molecular characterization also helps determine the breeding behaviour of species, individual reproductive success and the existence of gene flow within and between populations, and its consequences [32]. Information about the genetic make-up of accessions helps decision making for conservation activities, which range from collecting and managing through identifying genes to adding value to genetic resources [8].
The knowledge of the extent and pattern of diversity in the crop species is a prerequisite for any crop improvement as it helps breeders in deciding suitable breeding strategies for their future improvement [41]. The systematic evaluation of the molecular diversity encompassed in barley genetic resources is a prerequisite for its efficient exploitation in breeding as well as for development of the strategies for optimal conservation of genetic diversity [28].
Molecular genetic marker analysis has clarified the structure of genetic diversity in a broad range of crops. Recent technological developments have made whole-genome sequence polymorphism and gene-targeted surveys possible, casting light on population dynamics and the impact of selection during domestication. Germplasm description has hence conferred analytical power for resolution of the genetic basis of trait variation and adaptation in major crops such as cereals, chickpea, grapevine, cacao, or banana [11]
Barley(HordeumVulgareL) has been the object of a very long history of mutation induction, both by chemicals and irradiation, producing one of the best collections the barley of mutants among plants and creating much diversity for breeding and genetic studies. The genetic variation which exists in barley germplasm worldwide is a consequence of many evolutionary pressures which have modified gene pool [22]. Thus Barley has the advantage, apart from being an important agricultural crop for food and feed, that it has also been used virtually worldwide as a model species for biological research.
Barley has been an important model species in the fields of genetics and mutagenesis, in particular for cereal species. This was because of its diploid self-pollinating crop species with a low chromosome number (2n = 14) and a relatively short life cycle, which can be cultivated In a wide range of climatic conditions and is easy to use in cross-breeding [38]
New genomics and molecular breeding technologies are allowing us to better understand and mine genetic variation. Cumulatively, the new resources empower the current generation of barley breeders and geneticists to continue the tradition of barley making simultaneous contributions to agriculture and to science. The great challenges of climate change and human population growth will require innovative, insightful, and productive germplasm enhancement and variety development [30].
Microsatellites markers are defined as simple sequence repeats (SSRs) which are based on unique DNA sequences that are flanking short repetitive traits of simple sequence motifs, for example – di or tri nucleotides. They are randomly distributed within the eukaryotic genomes [42]. SSRs are codominant, abundant, informative and their detection is very simple [27]. In many studies, SSR markers have been used to characterize barley from different countries [21,23,26,37,49]. No studies evaluating the molecular diversity of Algerian barley using this kind of markers have been conducted. The objective of the study was to explore the polymorphism of microsatellite markers on 33 Barley accessions originated from Algeria and analyse their genetic diversity.
2. Materials and Methods
2.1. Plant Materials
A total of 33 accessions were selected from diffferent regions of Algeria. Geographical details on the germplasm characterized in this study are provided in Table 1
2.2. Genomic DNA Isolation
Three seeds of each accession were sown, and the seedlings were grown in a greenhouse. Three weeks later, Young leaves of each accession were collected, and total genomic DNA was extracted using CTAB (cetyltrimethylammonium bromide) following the protocol described by [37] with minor modifications [45]. DNA quality was examined using electrophoresis with 1.0 % agarose-gel concentration (100 V during 1hour).
2.3. Microsatellite Markers
In total, 27 SSR markers representing the chromosomes 1H, 2H, 3H, 4H, 5H, 6H and 7H were selected for genotyping the 33 Algerian accessions of barley. They were HVM14, HVM22, HVM65, Bmag0206[47],HVLEU [4],Bmac0093,Bmac0316,Bmac067,Bmac0113,Bmac0040,Bmac0134,Bmac0096,Bmac0018,Bmac0209,Bmac0273,Bmac0032,Bmac0156,Ebmac0806,Ebmac0705,Bmag0013,Bmag0378,Bmag0009, Bmag0120, [36] ,Bmac0213, Bmac0181, HVM74([14].(Table 2)) .
PCR reaction was performed in a reaction volume of 10 μL containing 5x PCR buffer, 0.2mM of each dNTPs, 1pmol/μL of each primer, 0.025 μL of Taq DNA polymerase (Promega) and approximately 50 ng of genomic DNA. Primer sequences for each marker are given in Table 2.The reaction mixture was incubated in a thermal cycler at 94 °C for 5 min, followed by 30 cycles of 94 °C for 30 s, 55 -58°C for 40 s and 72 °C for 40 s, and finally 72 °C for 5 min. The PCR products were separated on 8% polyacrylamide gels, prepared on an OWL vertical electrophoresis system using 0.5 TBE buffer,which were then subjected to silver staining after running with ethidium bromide .Gel visualization was carried out, after 2 h (Molecular Imager® Gel Doc™ XR+ system, BIO-Rad).Size of each band was estimated simultaneously by means of a 100-bp DNA Ladder.
2.4. Analysis of Molecular Data
Power Marker software version 3.25 [25] was used to evaluate summary statistics, such as the number of alleles (Na), allele frequency, values of genetic diversity, Polymorphism information content (PIC) [7] of each locus and genetic distances between each pair of cultivars were measured by calculating the shared allele frequencies [17]. A dendrogram was constructed based on the genetic distance by using Neighbor-joining (NJ) method [39]. Using software GenAlEx 6 [33] an analyses of molecular variance (AMOVA ; [10] (, Genetic structure (effective number of alleles(Ne), mean expected heterozygosity (He), Wright’s F-statistics, *Wright’s statistics according to Weir and Cockerham1984 (Fst), number of private alleles (NPA), Hardy-Weinberg Equilibrium * P < .05, ** P < .01, *** P < .001 (HWE)and gene flow estimated(Nm) (Nm=0.25(1- FST) /FST)) and Principal Coordinates Analysis (PCoA) were carried.
3. Results
3.1. Genetic Diversity
The genetic diversity of these Algerian genotypes is estimated on the basis of the number of alleles, the frequency of the alleles and the PIC values. We detected 110 alleles in Algerian accessions. The number of alleles per locus varied from one for markers Bmac 0032, Hvm65 and Hvm14 (monomorphic loci) to eight for the marker Bmag0009 and Ebmac0.806 with an average of 4.1 alleles per locus (Table 3). The effective number of alleles (Ne) ranged from 1.00 (bmac0032, hvm65 and hvm22) to 6.43 (ebmac0806) with an average of 2.69. Genetic diversity for 27 microsatellites loci at 33 Algerian Barley accessions varied from 0,00 (Bmac 0032, Hvm65 and Hvm14) to 0.84 (Bmag0009 and Ebmac0.806) with an average of 0.50 (Table 3). These results reveal that the markers chosen were sufficiently effective to detect the diversity in the Algerian accessions studied. Furthermore, indicators of locus diversity (polymorphism information content - PIC) were calculated (Table3), the PIC values of markers can provide an estimate of discrimination power in a set of accessions by taking not only the number of alleles, but also the relative frequencies of each allele [43]. Large variation was observed among the polymorphic loci ; PIC values ranged from 0.06 (Bmag0378) to 0.83(Ebma0806) with an average of 0.46.
Concerning the observed heterozygosity parameter, all loci showed(Ho) values close to zero, whereas the expected heterozygosity(He) ranged from 0.00 (bmac0032, hvm65 and hvm22) to 0.84 (Ebma0806) with an average of 0.50. For the gene flow estimated (Nm) ranged from 0.00 (bmac0032, hvm65 and hvm22) to 0.04 (bmag0009), with an average of 0.02. For Fst parameter, all loci showed higher values with an average of 0.81. No Hardy-Weinberg Equilibrium was performed for loci bmac0032, hvm65 and hvm22 because there were monomorphic, whereas all other loci deviated very significantly from HWE.
3.2. Accessions Genetic Structure
By structure analysis on the whole dataset of 33 accessions, and using all of the27 SSR loci, We performed the number of alleles (Na), effective number of alleles(Ne), mean expected heterozygosity (He) and number of private alleles(NPA) (Table 4).
The higher number of alleles per locus (Na=7) was observed for four accessions (chater ad, Saida SL, bourabaa AD, Saida Adf) whereas the effective number of alleles (Ne) ranged from 2.51 (Saida SL) to 4.51 (Chir B) with an average of 3.69 .The mean expected heterozygosity (He) was 0.72 and which ranged from 0.60 for Saida SL to 0.78 for O.byad BA2.
Although a total of 693 private alleles have been identified in 33 accessions studied, 401of them have a frequency greater than 5%. The highest number of private alleles having a frequency greater than 5%, was observed in SaidaR1and Faouar SBA1 accessions.
The Dendrogram constructed (Figure 1) classified the evaluated accessions into three major groups (Group-1, Group -2 and Group-3). In each group, the southern and northern accessions were mainly separated into five minor groups ;the first group consists of Safra BA,,O.beldi Ad ,Bourabaa Ad,Safra Ad ,Bourabaa Ker ,Safra BA,O,Byad BA2 and Obyad BA, the second group consists of Chater Ad ,Chater Ker Salt Ad Tichderett M d and Tichederett SBA and the third group consists of Ras elmouch1oa ,Raselmouch Ad , A zrir Ad and Ghir B. The fourth group is formed by the accessions Saida sl ,Saida Md ,Saida Ad ,Saida G3 and Saida G4.The fiftieth group consists of Rihane Adf , RihaneMd , Rihane SBA , Saida R1, Saida BA ,Siada B, ,Faouara SBA2, Faouara SBA1 ,Siada SBA,Faouara SBA and Askad SAB.
The lowest genetic distance (0.2) was observed between accession saidaG4 (Tlemcen) and Saida(Bechar) and the highest genetic distance of 0.85 is recorded between population Ras elmouch
(Oueld ali) and SaidaG3(Tlemcen) (Appendix A). These populations are the most genetically distant. Genetic relationships among barley varieties revealed by genetic similarity at SSR levels were in agreement with their roles in agricultural production and breeding [35].
3.3. Genetic Relationships and PCoA Analysis
The genetic structure was established by performing a principal coordinate analysis (PCoA) using SSR data. The PCoA of genetic distance between genotypes, based on gene frequencies revealed differentiation between accessions. The three axes explained 15, 8.48 and 7.29 % of the total variance, and separated the accessions into two clusters, northern accessions in one cluster and southren in another cluster (Figure 2). PCoA also confirmed the results obtained from Nei’s GD estimates
3.4. Analysis of Molecular Variance (AMOVA)
The AMOVA analysis (Table 6) revealded that the proportion of variance between accessions in the regions was very high, accounting for 97% of the total variation while the differnce between regions recorded only 3%. These results suggest that the genetic flow between region it’s very low.
4. Discussion
This study is the first report on genetic characterization of barley accessions of Algeria. All of the accessions were genotyped using 27 SSR markers assigned to seven chromosomes in barley. [34] reported that the information of polymorphism would be sufficient if more than 70 alleles were detected. [40] suggested that more than two SSRs from each of seven linkage groups should be selected to ensure the efficiency and representation of the genetic information among accessions.
In general, average number of alleles detected in this study in Algerian barley accessions (4.2) was higher than Tunisian Barley accessions reported by [12] (3.6), Indian varieties reported by [16] for Eritrean barley accessions (7.6). Molecular diversity in barley accessions from various geographic regions worldwide differs with respect to allelic richness, frequency of unique alleles, and extent of heterogeneity [28].For the effective number of alleles, we found an average (Ne=2.69) .This value was close to that reported by [5]for Italian barley landrace populations with an average value (Ne=2.75).The difference between average number of observed alleles and effective number of alleles was due to the uneven frequency of each allele [24].
Concerning the mean PIC value we found (0.46) which was close to that reported by several workers for SSR markers ([27,37,44]).For exemple, [27]reported a mean PIC value of 0.45 in a set of 96 elite breeding lines of barley from a single barley breeding program. . This value was lower than those reported by [18] for Tunisian barley where mean PIC value was 0.53.and [47].
On barley lines from six different countries using a set of 28 SSRs where mean PIC value was 0.58. The highest PIC value (0.83) was displayed by SSR marker (Ebma0806) having highest number of alleles. Thus, six of the loci were moderately polymorphic (0.25≤PIC≤ 0.5) and fifteen loci were highly polymorphic (0.5˃PIC), while six of the loci showed low polymorphism (0.25˂PIC). PIC provides an estimate of discriminatory power of a marker to differentiate genotypes based on both the number of alleles expressed and their relative frequencies [31] . The observed heterozygote (Ho=0.00) was lower than the exeted heterozygosity (He=0.5) same result was reported by [5] with (Ho=0.01) and (He=0.51).
The results indicated that all loci deviated very significantly from HWE except for three loci (bmac0032, hvm65 and hvm22) No Hardy-Weinberg Equilibrium was performed. Possible explanations for deviations of loci from HWE are heterozygote deficiency in loci, population size and the reproduction mode [24].
By genetic structure of 33 accessions Algerian, we identified that the accessions (chater ad, SaidaSL, bourabaaAD, SaidaAdf) contained the higher number of alleles per locus. The effective number of alleles and the expected heterozygosity are (Ne=3.69) and (He=0.72) respectively, these values were higher than reported by [5] with (Ne=2.09) and (He=0.42) respectively.
The cluster analysis for 27 SSR markers used in the study showed a high level of diversity among all the barley accessions. All accessions represented mainly three major groups. In each group, the southern and northern accessions were mainly separated into minor groups. This indicated that accessions collected from one location was almost similar than accessions taken from different states [2]. However, northern accessions Tichedret SBA and Tichedrett Media were grouped with sourthern accessions. The variety Tichedrett, has a behavior of resistance to water deficit, as well as an efficient root development [20].
The PCoA showed that 33 accessions were well scattered. Data analysis, i.e., NJ dendrogram and principal coordinate analysis, also revealed differentiation between accessions in general.
The AMOVA analysis showed that the proportion of variance between accessions in the regions was very high (97%) while the difference between regions recorded only 3%. Similar results were recorded for barley in Eritria, [3] analyzed 39 SSR in 24 fields of barley and observed a highest proportion of variance within the landraces (97.5%). [1] also analyzed six agronomic traits in 18 barley landraces from Ethiopia and found a large degree of variation within the landraces (97.3%). Aside from barley, a similar genetic structure was obtained for wheat, studying 39 moroccan varietes with 14 SSR markers, [15] found 96 % of the genetic variation within the landraces.
5. Conclusion
This study is the first report on the genetic characterization of barley accessions in Algeria. In addition, the results of DNA profiles obtained was conducted to determine the levels of genetic variation in algerian barley materials and can still be used for the reliable identification of barley germplasm. This study give us a real genetic potentiel about barley in Algeria, this result can be very helpful in management of biodiversity and genetic improvement.
Acknowledgments
Authors are grateful to the International Treaty for Plant Genetic Resources for Food and Agriculture/FAO, the European Union, the CRP-Wheat and ICARDA/Morocco Collaborative Grants Program for the financial support
Appendix A. Dissimilarity Matrix

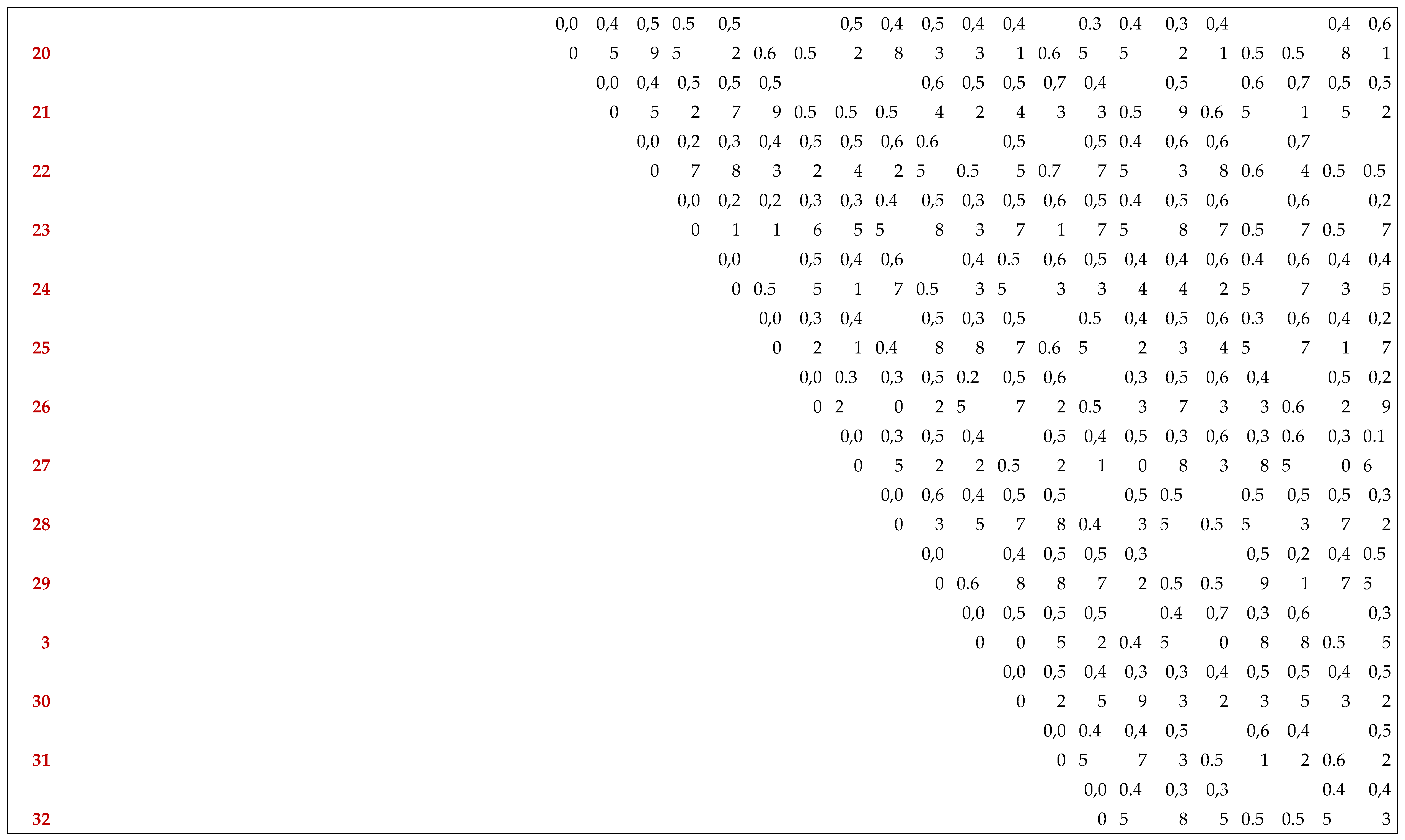

References
- Alemayehu F, Parlevliet JE. Variation between and within Ethiopian barley landraces.Euphytica ,1997 ;94:183-189.
- Babaei N,Abdullah N. A. P., Saleh G, Abdullah T. L. Isolation and characterization of microsatellite markers and analysis of genetic variability in Curculigo latifolia Dryand. Mol Biol Rep,2012 ; 39:9869–9877. [CrossRef]
- Backes, G. M., Orabi, J., Wolday, A., Yahyaoui, A., & Jahoor, A. High genetic diversity revealed in barley(Hordeum vulgare) collected from small-scale farmer’s fields in Eritrea. Genetic Resources and Crop Evolution,2009 ;56(1),85-97. https://doi.org/10.1007/s10722-008-9347-5.
- Becker J and Heun M .Barley microsatellites: allele variation and mapping Plant Molecular Biology,1995 ;27:835-845.
- Bellucci E, Bitocchi E, Rau D, Nanni L, Ferradini N, et al. Population Structure of Barley Landrace Populations and Gene-Flow with Modern Varieties,2013 ; PLoS ONE 8(12): e83891. [CrossRef]
- Benmohammed A.La production de l’orge et possibilités dedéveloppement en Algérie. ITGC, Alger. Céréaliculture,2004 ; 41, 34-38.
- Botstein, D., White, R.L., Skolnick, M., and Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet,1980 ; 32: 314- 331.
- De Vicente C.F.A., Guzmán J., Engels V., Ramanatha R.,2005. Genetic characterization and its use in decision making for the conservation of crop germplasm. the role of biotechnology -Villa Gualino, Turin, Italy – 5-7.
- Edwards, S. Tuvesson, M. Morgante, A. Massari, E. Maestri, N. Marmiroli, T. Sjakste, M. Ganal, W. Powell and R. Waugh. A simple sequence repeat-based linkage map of barley .Genetics,2000 ; 156:1997-2005.
- Excoffier L, Smouse PE and Quattro J .Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics,1992 ; 131:479-491.
- Glaszmann, J.; Kilian, B.; Upadhyaya, H.D.; Varshney, R.K.(2010). Accessing genetic diversity for crop improvement. Curr. Opin. Plant Biol. 13 :167–173.
- Hamza S, Ben Hamida W, Rebai A, Harrabi M (2004) SSR-based genetic diversity assessment among Tunisian winter barley and relationship with morphological traits. Euphytica.135:107-118.
- Harlan JR . Our Vanishing Genetic Resources. Science, 1975 ;188:618-621.
- Hearnden PR et al. A genetic map of 1,000 SSR and DArT markers in a wide barley cross. Theoretical and Applied ,2000 ; Genetics 115:383-391.
- Henkrar Fatima, Jamal El-Haddoury, Driss Iraqi, Najib Bendaou, Sripada M. Udupa Allelic variation at high-molecular weight and low-molecular weight glutenin subunit genes in Moroccan bread wheat and durum wheat cultivars. 3 Biotech ,2017 ; 7:287.
- Jaiswal s k, shree p.pandey,s. Sharma1,r. Prasad, l. c.Prasad1,r. p.s. Verma and arun k. joshi.Diversity in Indian barley (Hordeum vulgare) cultivars and identification of genotype-specific fingerprints using microsatellite markers. Indian Academy of Sciences,2010 ;89 : 46–54.
- Jin L, Chakraborty R .Estimation of genetic distance and coefficient of gene diversity from single-probe multilocus DNA fingerprinting Data. Mol Biol Evol,1993 ; 11:120–127.
- Kadri karim, Abdellawi RAWDA , Cheikh-Mhamed Hatem . Genetic diversity in barley genetic diversity in local Tunisian barley based on RAPD and SSR analysis. BioDiCon 2/1 ,2009 ; 27-35.
- Kaplan JK . Conserving the world’s plants. Agr Res ,1998 ; 46:4-9.
- Khaldoun A , Jacques Chery , P. Monneveux . Etude des caracteres d’enracinement et de leur role dans l’adaptation au deficit hydrique chez l’orge (Hordeum vulgare L.). Agronomie,10(5),1999,pp.369-379.
- Khodayari H, Saeidi H, Roofigar AA, Rahiminejad MR, PourkheirandishMand Komatsuda T . Genetic diversity of cultivated barley landraces in Iran measured using microsatellites. Int J Biosci Biochem Bioinform,2012 ; 2:278-290.
- Ladizinsky. G. Economic Botany ,1985 ;39 :191–199.
- Lamara M., Zhang Li Yi., Marchand S., Nicholas A., Tinker F.,Belzile.Comparative analysis of genetic diversity in Canadian barley assessed by SSR,DarT,and pedigree data.2013 ;Genome.Vol.56,N°6:351-358-. [CrossRef]
- Liao M, Wang Y, Rong X, Zhang Z, Li B, Wang L, Chen G . Development of new microsatellite DNA markers from Apostichopus japonicus and their cross-species application in Parastichopus parvimensis and Pathallus mollis. Int J Mol Sci ,2011 ;12:5862–5870.
- Liu, Muse .PowerMarker: an integrated analysis environment for genetic marker analysis”, Bioinformatics,2005 ; 21(9):2128-2129.
- Ledovskoy Y, Abugalieva S and Turuspekov Y . Comparative assessment of the genetic variation in wild and cultivated barley based on SSR markers. Asian Australas J Plant Sci Biotechnol ,2010 ;4:21-26.
- Matus IA, Hayes PM. Genetic diversity in three groups of barley germplasm assessed by simple sequence repeats. Genome ,2002 ;45:1095-1106.
- Malysheva-Otto L. V., Ganal M.W. and Roder M. S. Analysis of molecular diversity, population structure and linkage disequilibrium in a worldwide survey of cultivated barley germplasm (Hordeum vulgare L). BMC Genet.2006 ; 7- 6.
- Mebarek Lamara, Li Yi Zhang, Suzanne Marchand, Nicholas A. Tinker, François.Belzile-Comparative analysis of genetic diversity in Canadian barley assessed by SSR,DarT, and pedigree data.Genome.2013 ;Vol.56,N°6 :351-358. [CrossRef]
- Munoz-Amatrian Mara, Alfonso Cuesta-Marcos, Patrick M. Hayes and Gary J. Muehlbauer. Barley genetic variation: implications for crop improvement.Briefings in functional genomics,2014 ;Vol 13. NO 4. 341^350. [CrossRef]
- Nagl N, Taski-Ajdukovic K, Popovic A, Curcic A, Danojevic D, Kovacev L (2011) Estimation of genetic variation among related sugar beet genotypes by using RAPD. Genetika 43:575–582.
- Papa, R. & Gepts P. Asymmetry of gene flow and differential geographical structure of molecular diversity in wild and domesticated common bean (Phaseolus vulgaris L.) from Mesoamerica. Theor. Appl.Genet., 2003 ;106:239–250.
- Peakall R, Smouse PE. GenAlEx 6.5: genetic analysis in Excel.Population genetic software for teaching and research-an update.Bioinformatics 28:2537–2539,2012. [CrossRef]
- Pejic I, Ajmone-Marsan P, Morgante M, Kozumplick V, Castiglioni P, Taramino G, Motto M. Comparative analysis of genetic similarity among maize inbred lines detected by RFLPs, RAPDs, SSRs and AFLPs. Theor Appl Genet,1998 ;97(8):1248–1255. [CrossRef]
- QUIAN.
- Ramsay L , M. Macaulay, S. degli Ivanissevich, K. MacLean, L. Cardle, J. Fuller, K. J. Ramzi Chaabane ,Mouldi El Felah, Hammadi Ben Salah ,M’Barek Ben Naceur Chedly Abdelly, Dalila Ramla , Ahmad Nada ,Mahmoud Saker. Molecular Characterization of Tunisian Barley (Hordeum Vulgare L.) Genotypes using Microsatellites (SSRs) Markers. Eu ropean Journal of Scientific Research ISSN 1450-216X Vol.36 No.1 :pp.6-15,2009.
- Saghai Maroof M, Biyashev RM, Yang GP, Zhang Q and Allard RW .Extraordinarily polymorphic microsatelliteDNA in barley: species diversity, chromosomal locations, and population dynamics,1994 ;Proc Natl Acad Sci USA91:5466-5470.
- Saisho D, Takeda K . Barley: emergence as a new research material of crop science. Plant Cell,2011 ;Physiol 52:724-727.
- Saitou, N., Nei, M . The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution,1987 ; 4: 406-425.
- Shi YT, Bian HW, Han N, Pan JW, Tong WX, Zhu MY. Genetic Variation Analysis by RAPD of Some Barley Cultivars in China. Acta Agron Sin, 2004 ;30(3):258–265. (in Chinese).
- Singh Nivedita, Debjani Roy Choudhury, Gunjan Tiwari, Amit Kumar Singh, Sundeep Kumar, Kalyani Srinivasan, R. K. Tyagi, A. D. Sharma, N. K. Singh and Rakesh Singh.Genetic diversity trend in Indian rice varieties: an analysis using SSR markers-BMC,2016 ;Genetics-17:127-1 :13-. [CrossRef]
- Smith, D. N., and Devey, M. E. Occurrence and inheritance of microsatellites in Pinus radia.1994 ; Genome 37 :977–983. [CrossRef]
- Smith JSC, Kresovich S, Hopkins MS, Mitchell SE, Dean RE, Woodman WL, Lee M, Porter K . Genetic diversity among elite sorghum inbred lines assessed with simple sequence repeats. Aust. J. Crop Sci,2000 ; 40:226-232.
- Struss D, Plieske J (1998). The use of microsatellite markers for detection of genetic diversity in barley populations. Theor. Appl. Genet. 97:308-315.
- Udupa SM, Robertson LD, Weigand F, Baum M, Kahl G. Allelic variation at (TAA)n microsatellite loci in a world collection of chickpea (Cicer arietinum L.) Germplasm. Mol Genet Genom,1999 ; 261:354–363. [CrossRef]
- Van de Ven, M.; Powell, W.; Ramsay, G.; Waugh, R. Restriction fragment length polymorphisms as genetic markers in Vicia. Heredity . 1990 ;65 :329–342.
- Varshney RK et al. A high density barley microsatellite consensus map with 775 SSR loci. Theoretical and Applied Genetics ,2007 ;114:1091.C. de Vicente1 , F.A. Guzmán , J. Engels and V. Ramanatha Rao . genetic characterization and its use in decision making for the conservation of crop germplasm. the role of biotechnology -Villa Gualino, Turin, Italy ,2005 ;5-7.
- Xie W. & Nevo E.Wild emmer: genetic resources, gene mapping and transfer for wheat improvement. Euphytica ,2008 ;164 : 603–614.
- Yahiaoui S, Igartua E, Moralejo M, Ramsay L, Molina-Cano JL,Lasa JM, Gracia MP and Casas AM . Patterns of geneticand eco-geographical diversity in Spanish barleys.Theor Appl Genet ,2008 ;116:271-282.
Figure 1.
Dendrogram showing relationships among the 33 Algerian accessions of barley as revealed by Neighbour-joining method based on shared allele genetic distance. All the accessions were clustered into three major groups (Group-1, Group-2 and Group-3).
Figure 1.
Dendrogram showing relationships among the 33 Algerian accessions of barley as revealed by Neighbour-joining method based on shared allele genetic distance. All the accessions were clustered into three major groups (Group-1, Group-2 and Group-3).
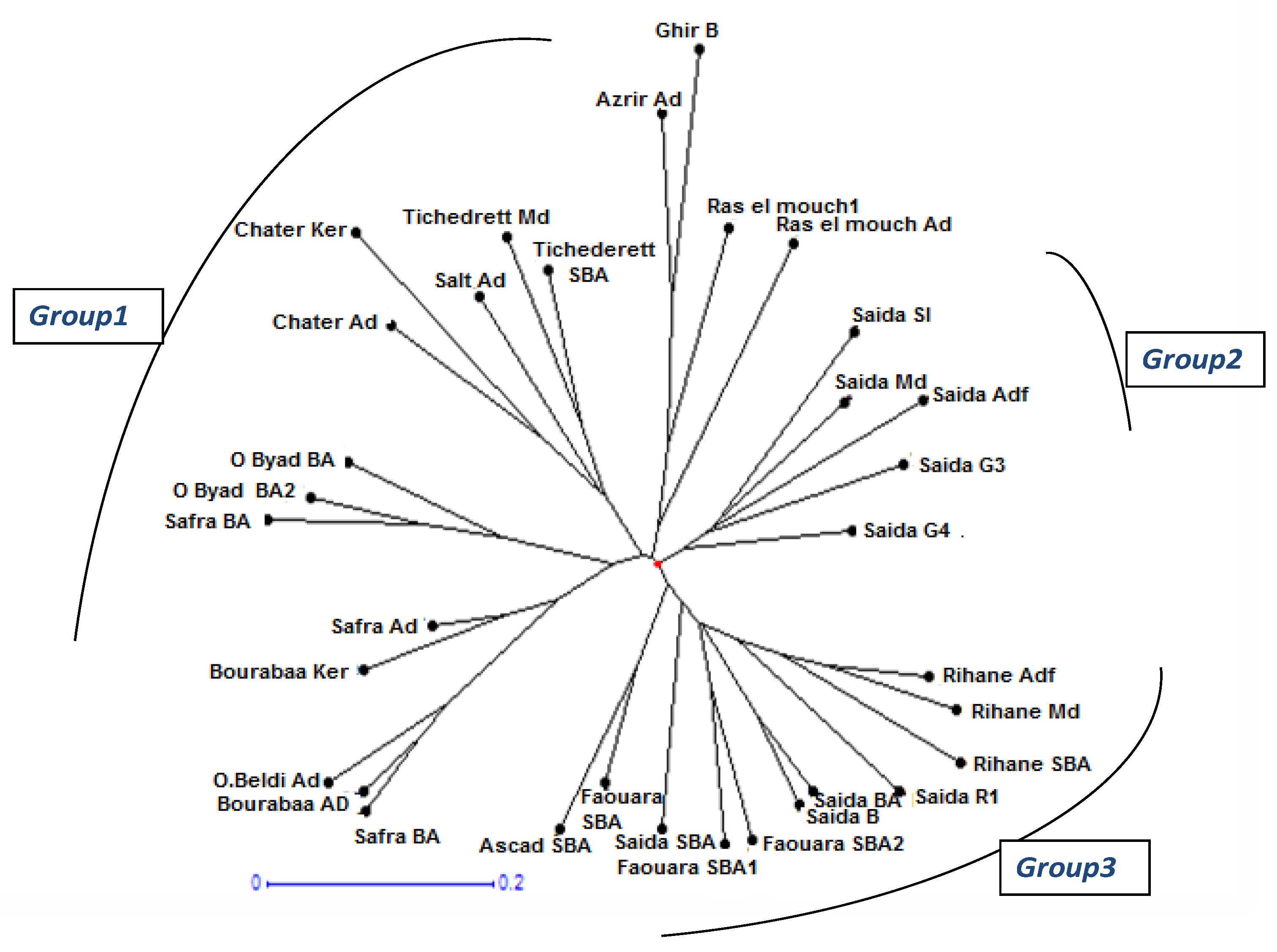
Figure 2.
Principal coordinate analysis (PCoA) plot generated from genetic distance calculations using the GENALEX package.
Figure 2.
Principal coordinate analysis (PCoA) plot generated from genetic distance calculations using the GENALEX package.
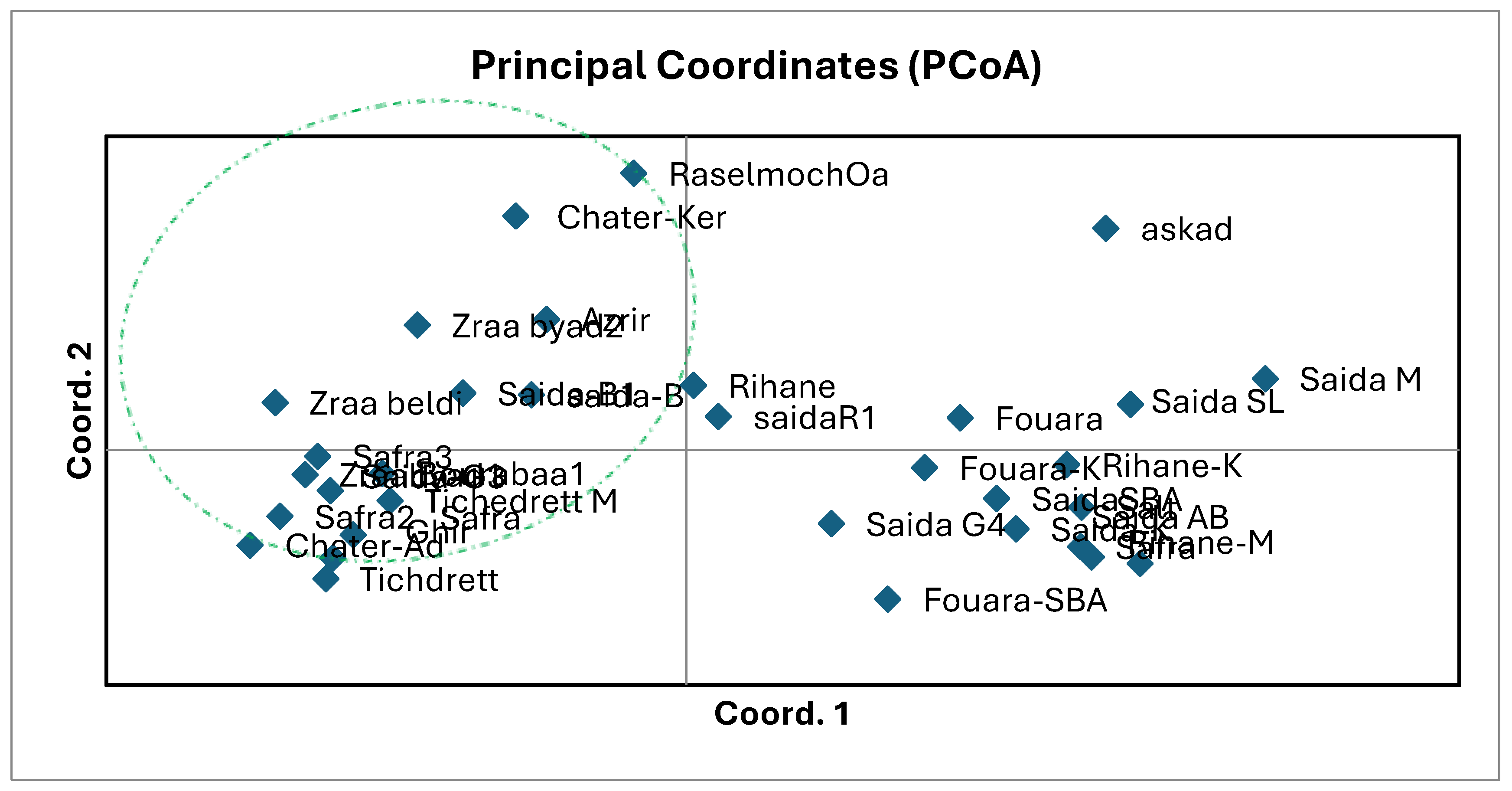

Table 1.
Names and origin of accessions of barley studied in Algeria.
| Accessions | region | locations | Longitude | Latitude | Altitude |
|---|---|---|---|---|---|
| Saida G3 | tlemcen | tlemcen | 1°19’4.82”O | 34°53’18.26”N | 745m |
| Saida G4 | tlemcen | 1°19’4.82”O | 34°53’18.26”N | 745m | |
| Saida R1 | Sid abdli | 1° 7’58.47”O | 35° 3’49.79”N | 465m | |
| Saida | Sidi belabbes Sidi belabbes 0°38’17.72”O 35°12’7.84”N |
Sidi belabbes | 0°38’17.72”O | 35°12’8.01”N | 478m |
| Saida2 | Sidi lahcene | 0°43’29.18”O | 35°12’6.11”N | 515m | |
| Ascad | Sidi belabbes | 0°38’17.72”O | 35°12’7.84”N | 478m | |
| Rihane | Sidi belabbes | 0°38’17.72”O | 35°12’7.84”N | 478m | |
| Faouara | Sidi belabbes | 0°38’17.72”O | 35°12’7.84”N | 478m | |
| Tichedrett | Sidi belabbes | 0°38’17.72”O | 35°12’7.84”N | 478m | |
| Faouara2 | Sidi belabbes | 0°38’17.72”O | 35°12’7.84”N | 478m | |
| Azrir | Adrar | oueled ali | 0° 5’52.95”O | 28°42’51.56”N | 251m |
| Safra | oueled ali | 0° 5’52.95”O | 28°42’51.56”N | 251m | |
| Bourabaa | adrar | 0°11’12.99”O | 27°58’18.55”N | 257m | |
| Salt | oueled ali | 0° 5’52.95”O | 28°42’51.56”N | 251m | |
| Ras el mouch | oueled ali | 0° 5’52.95”O | 28°42’51.56”N | 251m | |
| Chater | kerzaz | 1°26’19.08”O | 29°27’44.71”N | 387m | |
| Bourabaa | kerzaz | 1°26’19.08”O | 29°27’44.71”N | 387m | |
| Chater 2 | adrar | 0°11’12.99”O | 27°58’18.55”N | 257m | |
| Zraa beldi | zaouiet konta | 0°12’3.68”O | 27°13’34.73”N | 204m | |
| Orge ALG ad | oueled ali | 0° 5’52.95”O | 28°42’51.56”N | 251m | |
| Ghir | Bechar | bechar | 2°10’58.22”O | 31°33’28.58”N | 789m |
| Saida | bechar | 2°10’58.22”O | 31°33’28.58”N | 789m | |
| Safra3 | beni abbes | 2°10’1.66”O | 30° 7’56.89”N | 505m | |
| Saida | beni abbes | 2°10’1.66”O | 30° 7’56.89”N | 505m | |
| Safra2 | beni abbes | 2°10’1.66”O | 30° 7’56.89”N | 505m | |
| Zraa byad1 | beni abbes | 2°10’1.66”O | 30° 7’56.89”N | 505m | |
| Zraa byad2 | bechar | 2°10’58.22”O | 31°33’28.58”N | 789m | |
| Rihane | Ain defla | khemiss meliana | 2°12’49.78”E | 36°15’12.70”N | 281m |
| Saida | khemiss meliana | 2°12’49.78”E | 36°15’12.70”N | 281m | |
| Faouara | khemiss meliana | 2°12’49.78”E | 36°15’12.70”N | 281m | |
| Saida | Media | beni slimane | 2°56’15.27”E | 36° 3’12.55”N | 588m |
| Rihane | beni slimane | 2°56’15.27”E | 36° 3’12.55”N | 588m | |
| Tichedrett | beni slimane | 2°56’15.27”E | 36° 3’12.55”N | 588m |
Table 2.
Caracteristic of the 27 SSR markers used for molecular characterization.
| Name | Chr | Repeat | Size | Primer sequense |
|---|---|---|---|---|
| Bmag0013 | 3H | (CT)21 | 151 | F:AAGGGGAATCAAAATGGGAG R: TCGAATAGGTCTCCGAAGAAA |
| Bmac0067 | 3H | (AC)18 | 171 | F:AACGTACGAGCTCTTTTTCTA R :ATGCCAACTGCTTGTTTAG |
| Bmac0093 | 2H | (AC)24 | 151 | F:CGTTTGGGACGTATCAAT R : GGGAGTCTTGAGCCTACTG |
| Bmac0213 | 1H | (AC)23 | 168 | F : ATGGATGCAAGACCAAAC R : CTATGAGAGGTAGAGCAGCC |
| Bmac0040 | 6H | (AC)20 | 236 | F :AGCCCGATCAGATTTACG R : TTCTCCCTTTGGTCCTTG |
| Bmac0181 | 4H | (AC)20 | 177 | F:ATAGATCACCAAGTGAACCAC R: GGTTATCACTGAGGCAAATAC |
| Bmac0113 | 5H | (AT)7(AC)18 | 187 | F:TCAAAAGCCGGTCTAATGCT R:GTGCAAAGAAAATGCACAGATAG |
| Bmac0134 | 2H | (AC)28 | 148 | F:CCAACTGAGTCGATCTCG R :CTTCGTTGCTTCTCTACCTT |
| Bmac0096 | 5H | (AT)6(AC)16 | 173 | F:GCTATGGCGTACTATGTATGGTTG R :TCACGATGAGGTATGATCAAAGA |
| Bmac0316 | 6H | (AC)19 | 135 | F:ATGGTAGAGGTCCCAACTG R :ATCACTGCTGTGCCTAGC |
| Bmac0018 | 6H | (AC)11 | 138 | F:GTCCTTTACGCATGAACCGT R : ACATACGCCAGACTCGTGTG |
| Bmac0209 | 3H | (AC)13 | 176 | F:CTAGCAACTTCCCAACCGAC R :ATGCCTGTGTGTGGACCAT |
| Bmac0273 | 7H | (AC)2(AG)20 | 186 | F:ACAAAGCTCGTGGTACGT R: AGGGAGTATTTCACCCTTG |
| Bmac0032 | 1H | (AC)7T(CA)15(AT)9 | 215 | F:CCATCAAAGTCCGGCTAG R GTCGGGCCTCATACTGAC |
| Bmac0156 | 6H | (GA)13 | 0.162 | F:AGGAAGTCATTGCGTGAG R:TGATCAAGAATGATAACATGG |
| Bmag0378 | 2H | (AG)14 | 147 | F : CTTTTGTTTCCGTAGCATCTA R ATCCAACTATAGTAGCAAAGCC |
| Bmag0009 | 6H | (AG)13 | 172 | F:AAGTGAAGCAAGCAAACAAACA R :ATCCTTCCATATTTTGATTAGGCA |
| Bmag0120 | 7H | (AG)15 | 230 | F:ATTTCATCCCAAAGGAGAC R GTCACATAGACAGTTGTCTTCC |
| Bmag0206 | 7H | (GT)5(AG)14 | 239 | F:TTTTCCCCTATTATAGTGACG R:TAGAACTGGGTATTTCCTTGA |
| Ebmac0705 | 3H | (AC)16 | 150 | F:GTGGAAAACTGAGTGAAACTC R TTGAGGAGAAGTAATGACGAT |
| Ebmac0806 | 6H | (CA)4(GA)(CA)8,(CA)5 | 168 | F:ACTAAGTCCTTTCACGAGGA R :GTGTGTAGTAGGTGGGTACTTG |
| HVM22 | 6H | (AC)13 | 0.167 | F:TTTTGGGGGATGCCTACATA R TTTCAAATGGTTGGATTGGA |
| HVM65 | 6H | (GA)10 | 0.129 | F:AGACATCCAAAAAATGAACCA R:TGGTAACTTGTCCCCCAAAG |
| HVM14 | 6H | (CA)11 | 0.158 | F:CGATCAAGGACATTTGGGTAAT R :AACTCTTCGGGTTCAACCAATA |
| HVM74 | 6H | (GA)13 | 0.162 | F:AGGAAGTCATTGCGTGAG R:TGATCAAGAATGATAACATGG |
| Hvleu | 5H | (ATTT)4 | 0.166 | F:TTGGAAGTGTACAGCAATGGAG R TGAAAGGCCCCACAAGATAG |
| Ryd2 | 3H |
Table 3.
Genetic diersity parameters of the 27 loci investigated loci in Barley accessions.
| Locus. | Ss | NA | MAF | G.div | PIC | Ne | He | Ho | Fst | Nm | HWE |
|---|---|---|---|---|---|---|---|---|---|---|---|
| bmac0213 | 33 | 5 | 0.48 | 0.68 | 0.64 | 3,39 | 0,70 | 0.00 | 0,88 | 0,034 | *** |
| bma0181 | 33 | 3 | 0.61 | 0.55 | 0.49 | 2,24 | 0,55 | 0.00 | 0,91 | 0,026 | *** |
| bmac0040 | 33 | 3 | 0.84 | 0.28 | 0.26 | 1,38 | 0,27 | 0.00 | 0,95 | 0,012 | *** |
| bmac0093 | 33 | 5 | 0.60 | 0.59 | 0.55 | 2,43 | 0,59 | 0.00 | 0,90 | 0,028 | *** |
| bmac0018 | 33 | 6 | 0.48 | 0.64 | 0.59 | 2,81 | 0,64 | 0.00 | 0,89 | 0,030 | *** |
| bmac0156 | 33 | 3 | 0.90 | 0.19 | 0.18 | 1,23 | 0,19 | 0.00 | 0,97 | 0,008 | *** |
| bmac0032 | 33 | 1 | 1.00 | 0.00 | 0.00 | 1,00 | 0,00 | 0,00 | 1,00 | 0,000 | - |
| bmac0067 | 33 | 6 | 0.35 | 0.76 | 0.73 | 4,2 | 0,76 | 0.00 | 0,87 | 0,037 | *** |
| bmac0113 | 33 | 5 | 0.36 | 0.70 | 0.64 | 3,33 | 0,70 | 0.00 | 0,88 | 0,033 | *** |
| bmac0273 | 33 | 3 | 0.90 | 0.18 | 0.17 | 1,22 | 0,18 | 0.00 | 0,96 | 0,008 | *** |
| bmac0134 | 33 | 7 | 0.37 | 0.75 | 0.71 | 3,98 | 0,75 | 0.00 | 0,87 | 0,036 | *** |
| bmac0096 | 33 | 4 | 0.63 | 0.56 | 0.52 | 2,29 | 0,56 | 0.00 | 0,90 | 0,026 | *** |
| bmac0209 | 33 | 4 | 0.53 | 0.57 | 0.49 | 2,39 | 0,57 | 0.00 | 0,90 | 0,027 | *** |
| bmac0316 | 33 | 4 | 0.47 | 0.60 | 0.52 | 2,51 | 0,60 | 0.00 | 0,89 | 0,028 | *** |
| ebmac0806 | 33 | 8 | 0.21 | 0.84 | 0.83 | 6,43 | 0,84 | 0.00 | 0,85 | 0,041 | *** |
| ebmac0705 | 33 | 3 | 0.50 | 0.57 | 0.48 | 2,35 | 0,57 | 0.00 | 0,90 | 0,027 | *** |
| bmag0378 | 33 | 2 | 0.97 | 0.06 | 0.06 | 1,06 | 0,06 | 0.00 | 0,99 | 0,003 | *** |
| bmag0206 | 33 | 4 | 0.48 | 0.64 | 0.57 | 2,79 | 0,64 | 0.00 | 0,89 | 0,030 | *** |
| bmag0013 | 33 | 3 | 0.54 | 0.57 | 0.49 | 2,24 | 0,55 | 0,00 | 0,91 | 0,026 | *** |
| bmag0009 | 33 | 8 | 0.24 | 0.84 | 0.82 | 5,95 | 0,83 | 0.00 | 0,86 | 0,040 | *** |
| bmag0120 | 33 | 3 | 0.42 | 0.66 | 0.58 | 2,90 | 0,66 | 0.00 | 0,89 | 0,031 | *** |
| hvm65 | 33 | 1 | 1.00 | 0.00 | 0.00 | 1,00 | 0,00 | 0.00 | 1,00 | 0,000 | - |
| hvm22 | 33 | 1 | 1.00 | 0.00 | 0.00 | 1,00 | 0,00 | 0.00 | 1,00 | 0,000 | - |
| hvm74 | 33 | 4 | 0.58 | 0.58 | 0.52 | 2,37 | 0,58 | 0.00 | 0,90 | 0,027 | *** |
| hvm14 | 33 | 6 | 0.31 | 0.78 | 0.75 | 4,5 | 0,79 | 0.00 | 0,87 | 0,037 | *** |
| hvleu | 33 | 6 | 0.33 | 0.78 | 0.74 | 4,45 | 0,78 | 0.00 | 0,87 | 0,037 | *** |
| ryd2 | 33 | 2 | 0.90 | 0.17 | 0.16 | 1,21 | 0,17 | 0.00 | 0,97 | 0,008 | *** |
| Mean | 4.2 | 0.59 | 0.50 | 0.46 | 2,69 | 0,50 | 0,92 | 0,024 | |||
| SE | 0,381 | 0,28 | 0,05 | 0,009 | 0,003 |
Ss : sample size,Na: number of alleles, MAF : Major.Allele.Frquency ,G.div : Gene diversity ,Ne: effective number of alleles, PIC: polymorphic information content, FST: Wright’s F-statistics, *Wright’s statistics according to Weir and Cockerham.1984, He: expected heterozygosity, HWE: Hardy-Weinberg Equilibrium * P < .05, ** P < .01, *** P < .001 and Nm: gene flow estimated (Nm=0.25(1- FST)/FST.
Table 4.
Genetic diversity parameters acording to studied Barley accessions across 27 loci.
| Accessions | Na | Ne | He | Fst | NPA | ||
|---|---|---|---|---|---|---|---|
| Freq.˂5 | Freq.≥5 | Total | |||||
| saltad | 6 | 3,09 | 0,68 | 0,83 | 4 | 17 | 21 |
| Raselmoch1oa | 5 | 3,74 | 0,73 | 0,81 | 5 | 12 | 17 |
| azrirad | 6 | 3,74 | 0,73 | 0,81 | 10 | 11 | 21 |
| safraBA | 5 | 3,25 | 0,69 | 0,82 | 12 | 6 | 18 |
| Chir B | 6 | 4,51 | 0,78 | 0,80 | 8 | 15 | 23 |
| safra AD | 5 | 3,25 | 0,69 | 0,82 | 6 | 11 | 17 |
| obyad ba1 | 6 | 2,75 | 0,64 | 0,84 | 0 | 17 | 17 |
| bourabaa Ba | 6 | 3,79 | 0,74 | 0,81 | 14 | 8 | 22 |
| chater ad | 7 | 4,09 | 0,76 | 0,81 | 10 | 8 | 18 |
| chaterker | 6 | 3,70 | 0,73 | 0,81 | 17 | 11 | 28 |
| saida SBA | 4 | 3,20 | 0,69 | 0,83 | 7 | 15 | 22 |
| faouara SBA | 6 | 3,31 | 0,70 | 0,82 | 0 | 10 | 10 |
| saidaG3 | 6 | 3,70 | 0,73 | 0,81 | 19 | 6 | 25 |
| SaidaSL | 7 | 2,51 | 0,60 | 0,85 | 12 | 10 | 22 |
| rihane SBA | 6 | 3,89 | 0,74 | 0,81 | 22 | 6 | 28 |
| saida G4 | 6 | 3,27 | 0,69 | 0,82 | 12 | 10 | 22 |
| rihane Md. | 6 | 4,43 | 0,77 | 0,80 | 17 | 6 | 23 |
| saida r1 | 7 | 3,84 | 0,74 | 0,81 | 0 | 21 | 21 |
| FaouaraSBA | 5 | 3,57 | 0,72 | 0,82 | 12 | 16 | 28 |
| Saida BA | 6 | 4,30 | 0,77 | 0,81 | 6 | 10 | 16 |
| AscadSBA | 5 | 3,63 | 0,73 | 0,82 | 12 | 10 | 22 |
| bourabaaAD. | 7 | 4,30 | 0,77 | 0,81 | 4 | 15 | 19 |
| Faouara SBA1 | 6 | 4,30 | 0,77 | 0,81 | 8 | 21 | 29 |
| RaselmouchAD | 6 | 4,04 | 0,75 | 0,81 | 5 | 19 | 24 |
| obyadBA2 | 6 | 4,45 | 0,78 | 0,80 | 5 | 17 | 22 |
| RihaneAdf | 7 | 4,12 | 0,76 | 0,81 | 5 | 16 | 21 |
| SafraAd. | 5 | 3,76 | 0,73 | 0,81 | 14 | 15 | 29 |
| SaidaMd | 6 | 4,30 | 0,77 | 0,81 | 6 | 11 | 17 |
| SaidaAdf | 7 | 3,21 | 0,69 | 0,83 | 5 | 16 | 21 |
| TichedrettMd | 4 | 3,79 | 0,74 | 0,81 | 19 | 9 | 28 |
| oBeldiAd | 5 | 3,41 | 0,71 | 0,82 | 0 | 10 | 10 |
| TichedrettSBA | 5 | 3,89 | 0,74 | 0,81 | 11 | 6 | 17 |
| SaidaB | 5 | 2,77 | 0,64 | 0,84 | 5 | 10 | 15 |
| Mean | 3.69 | 0.72 | 0.82 | ||||
Na: number of alleles, Ne: effective number of alleles, He: mean expected heterozygosity, Fst : Wright’s F-statistics, *Wright’s statistics according to Weir and Cockerham.1984, NPA: number of private alleles.
Table 6.
Results of the Analysis of Molecular Variance (AMOVA).
| Source | degrees of freedom(Df) | Sum of squares (SS) | Means of squares (MS) | Est. Var. | % | Stat | value | prob |
|---|---|---|---|---|---|---|---|---|
| Among populations | 5 | 87,737 | 17,547 | 0,206 | 3% | |||
| Among individuals within populations | 27 | 415,657 | 15,395 | 7,697 | 97% | 0,026 | 0,138 | |
| Total | 65 | 503,394 | 7,903 | 100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



