
Submitted:
01 July 2024
Posted:
02 July 2024
You are already at the latest version
Abstract
Neurobrucellosis in cetaceans, caused by Brucella ceti, is a relevant cause of death in striped dolphins (Stenella coeruleoalba) from the Mediterranean Sea. Serological tests are not used as a routinary technique for the diagnosis of this infection. We briefly describe the pathological findings of 10 free-ranging stranded cetaceans diagnosed with Brucella infection in our veterinary necropsy service from 2012 to 2023. Findings included focal diskospondylitis, non-suppurative meningitis, choroiditis and radiculitis. Additionally, an exploratory serological study was conducted in 66 frozen sera collected in the period 2012-2022 from 57 striped dolphins, 5 Risso’s dolphins (Grampus griseus), 2 common bottlenose dolphins (Tursiops truncatus), 1 common dolphin (Delphinus delphis) and 1 pilot whale (Globicephala melas), to compare antibody levels in Brucella-infected (n=8) and non-infected (n=58) animals, classified by cause of death, sex, age class and cetacean morbillivirus (CeMV) infection status. We performed a commercial competition ELISA (cELISA) using serial serum dilutions for each sample, considering a percentage of inhibition (PI) of ≥40% as positive. A titer of 1:160 was arbitrarily determined as the seropositivity threshold. Seropositive species included striped dolphins and Risso’s dolphins. Seroprevalence was higher in animals with neurobrucellosis (87.5%) compared to the overall prevalence (31.8%) and to other causes of death, indicating a high sensitivity but low specificity for neurobrucellosis. Animals with chronic CeMV seemed to have higher seroprevalences, as well as juveniles, which also had more disease prevalence. These results suggest that high antibody titers are not always protective against clinical brucellosis, although they may indicate a carrier state, and that CeMV may potentially condition Brucella epidemiology. More research is required to elucidate the epidemiology and pathogenesis and to disentangle the complicated host-pathogen interaction in Brucella species.
Keywords:
brucellosis
; serology
; antibodies
; ELISA
; morbillivirus
; dolphin
; stranding
; necropsy
1. Introduction
Brucellosis in cetaceans was first described in 1994 [1] and is caused by Brucella ceti [2]. Since then, infection and disease associated to B. ceti have been increasingly recognized in many cetacean species worldwide [3,4], and brucellosis is considered an emerging threat for odontocetes and mysticetes [5]. In the Mediterranean Sea, cetacean brucellosis was first diagnosed in 2013, and more cases have been reported since then [6,7,8,9,10], but serological evidence of infection was already detected in dolphins stranded along the Mediterranean coast of Spain in 1997-1999 [11], suggesting that Mediterranean dolphins were already exposed to the pathogen at that time. Disease due to B. ceti in cetaceans can occur as neurobrucellosis, reproductive disease, spinal diskospondylitis or abscesses (frequently cutaneous and subcutaneous) [4]. However, in many other instances, serological evidence of infection has been found in asymptomatic cetaceans, and B. ceti has been isolated from tissues of apparently healthy cetaceans, suggesting that Brucella-infected cetaceans may overcome initial infection and survive or become Brucella carriers (see for review [3,4]). Health assessment of wild cetacean populations is done usually through necropsy of stranded dolphins and laboratory investigation. Since the first description of disease associated to B. ceti in the Catalan Mediterranean coast [8], we have intensified efforts to detect Brucella in stranded cetaceans. In this paper we describe the cases of brucellosis detected in the period 2012 to 2023 in the North-western Mediterranean coast. Moreover, we perform an exploratory serological analysis using a commercial cELISA to compare antibody levels between infected and non-infected animals, age groups, sex, and cetacean morbillivirus (CeMV) status, with a special emphasis on striped dolphins. The general aims were to provide information about the epidemiology and pathogenesis of B. ceti infection, and to evaluate its impact on cetacean species in the area.
2. Materials and Methods
2.1. Cases Investigated and Necropsy Procedure
A total of 131 cetaceans stranded along the Catalan coast during the period 2012-2023 were necropsied at the Universitat Autònoma de Barcelona, according to standard procedures [12]. Necropsied species included striped dolphins (Stenella coeruleoalba) (n=103), common bottlenose dolphins (Tursiops truncatus) (n=13), Risso’s dolphins (Grampus griseus) (n=10), common dolphins (Delphinus delphis) (n=2), a fin whale (Balaenoptera physalus), a pilot whale (Globicephala melas) and a Cuvier’s beaked whale (Ziphius cavirostris). Conservation code ranged from 1 to 3 (animals in overt autolysis were not transported for necropsy) following the scale of Kuiken & García-Hartmann 1991 [13]. Retrieval of cerebrospinal fluid (CSF) was routinely attempted from the atlanto-occipital joint, or directly from the lateral ventricle after extraction of the brain. Frozen samples and swabs from multiple organs were collected for bacteriological and molecular investigations. A complete set of organs was sampled for histopathology, fixed in 10% neutral buffered formalin and routinely processed. Routine immunohistochemistry and RT-PCR for surveillance of CeMV were performed on lung, diaphragmatic lymph node and brain [14]. After the determination of the cause of death, animals were classified in different groups: bycatch, neurobrucellosis, CeMV, PEM (polioencephalomalacia of unknown origin), mother-calf separation, infectious causes, sinusitis by Crassicauda grampicola, erysipelas, other and unknown. “Infectious” cause of death comprised peritonitis, septicemia, protozoal infections, severe parasitism, mucormycosis, bacterial meningoencephalitis (negative Brucella culture), and necrotizing enteritis. “Other” included animals dying from tension pneumothorax, meningoencephalitis of unknown origin, abortion, pulmonary angiomatosis and muscular degeneration.
2.2. Serology for Brucella
Cardiac blood retrieval was attempted by cardiac puncture at reception of each cadaver with a 20 ml syringe and a 16G, 1.7 x 133 mm catheter needle (AngiocathTM, BD, REF 382259) and portioned in vacutainer plain tubes. Blood was centrifuged at 2,500 x g, and serum was aliquoted in 1 ml cryotubes and frozen at -80 ºC until use.
For the present serological study, available serum samples from 2012 to 2022, excluding those from 2023, (n=66; 57 striped dolphins, 5 Risso’s dolphins, 2 common bottlenose dolphins, 1 common dolphin and 1 pilot whale), were defrosted (see Supplementary File 1 for all cetacean samples). A commercial blocking ELISA (INgezim Brucella Compac, INGENASA, Madrid, Spain) was used following the manufacturer’s instructions. This competitive ELISA test (Brucella cELISA) is a multispecies kit developed for detection of blocking antibodies against LPS of Brucella abortus in domestic ruminants and swine serum samples, using a peroxidase-conjugated secondary monoclonal antibody directed to the LPS of B. abortus. The dilution of the serum sample for cetaceans is not established for the test and, therefore, sera were diluted as recommended by the manufacturer for ovine and caprine sera (1:5), bovine and porcine sera (1:10), and additionally at 1:20, 1:40, 1:80, and 1:160. Available serum of culture-confirmed Brucella-infected cases (n=8) were further diluted to 1:320, 1:640, 1:1280, 1:2560, 1:5120, and 1:10240. Optical density (OD) values were measured at 450 nm within 5 minutes after the addition of stop solution using a spectrophotometer (FLx800, Bio-Tek Instruments, Winooski, VT). OD values were used to calculate the final results, read as a percentage of inhibition (PI) in comparison to positive and negative control sera included in the kit, with the formula PI=100x [1-(OD test sample/OD negative control), where OD= optical density. Following the procedures of the test, a sample was classified as positive if the PI in the well was ≥40%.
The possible association between the level of hemolysis and the Brucella cELISA result was investigated in a non-published preliminary study. Briefly, the level of hemolysis was visually graded in the first dilution of samples when dispensed onto the ELISA plate into group 1 (non-hemolyzed or slightly hemolyzed serum), and group 2 (hemolyzed serum or hemolyzed blood). A comparison of positive and negative results in Brucella cELISA (for 1:20 dilutions and 1:40 dilutions) with the hemolysis score was performed with the Wilcoxon Two-Sample Test (Epi Info Package, www.cdc.gov/epiinfo/index.html).
2.3. Brucella Isolation
Brucella isolation, gold standard for brucellosis diagnosis, was attempted in dolphins with compatible lesions, a positive reaction to Bengal Rose Test (data not shown), or with CeMV infection. Samples used for Brucella isolation were frozen swabs from the lateral cerebral ventricle, cerebral tissue around lateral ventricle, cerebrospinal fluid (CSF), spleen, mesenteric lymph node, or swabs from spondylytic lesions. Bacterial culture was performed as previously described [8]. Briefly, tissue samples were superficially sterilized, homogenized in saline buffer and cultured in plates of both Farrell and CITA selective media. Isolates were identified as marine Brucella using a Bruce-ladder PCR [15]. Confirmation of Brucella ceti as the isolated species was done by a multiplex PCR adapted from Bruce-ladder [15].
2.4. Age Determination
The age of 57 striped dolphins for which serum was available was estimated using a Gompertz formula established for this species in the North Western Mediterranean Sea [16]. Also, animals were divided in three age ranges (fetus, calves, juveniles and adults), considering adulthood above 6 years [17]. The formula is not appliable to small animals under 120 cm (males) and 110 cm (females). For three calves of a few weeks of age (presence of fetal folds) age was estimated as 0.1 years. Age could not be estimated for dolphin N-604/17, which was found without a tail.
2.5. Data Analysis
Differences between antibody titers in striped dolphins, grouped by their cause of death and age, were compared in two-way contingency tables and using a Fisher Test. R software (version 4.4.0) was used to introduce data and elaborate the tables and StatCalc tool, from EpiInfo (version 7.2.6.0), to retrieve significance of the results. Significance was considered with p-value<0.05.
3. Results
3.1. Pathological Investigation
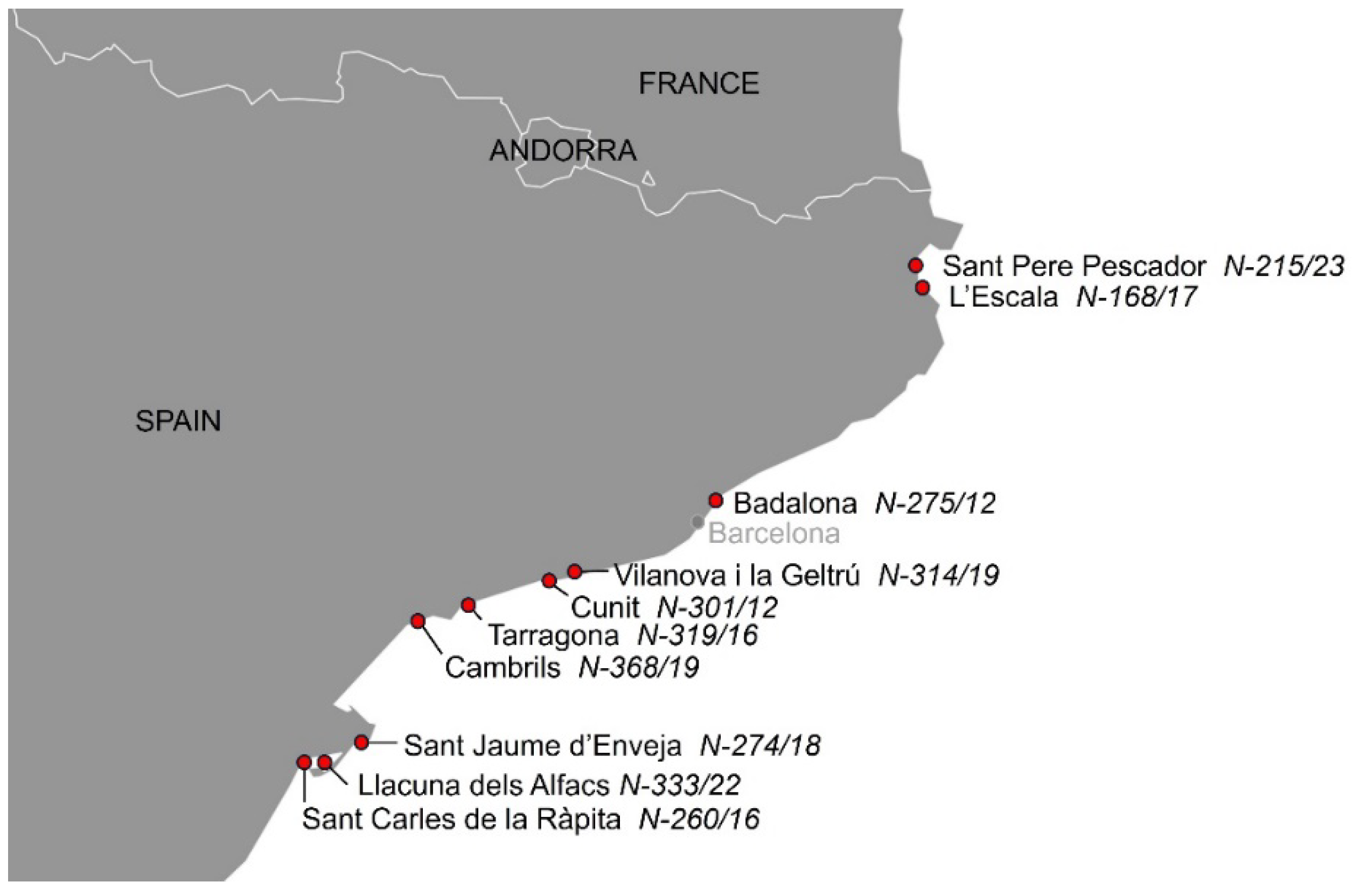
See Supplementary File 2 for biometrical data, cause of death, and ancillary test results in the 131 cetaceans. Ten cases of infection by Brucella ceti were detected among the 131 (7.6%) necropsied cetaceans, 9 striped dolphins and one common bottlenose dolphin. Eight of the 10 dolphins stranded alive and died shortly thereafter (n=5) or were euthanized due to bad prognosis (n=3). Six of them showed neurological signs, disorientation or abnormal swimming before death (301/12, 319/16, 368/19, 314/19, 333/22 and 215/23). Biometric and stranding data from the 10 cetaceans with brucellosis are shown in Table 1. A map with the geographic location of strandings is shown in Figure 1. Eight striped dolphins showed neurobrucellosis, characterized by a severe diffuse or multifocal non-suppurative meningitis or meningoencephalitis, in some cases more intense in the cerebellum and brainstem, and including choroiditis and radiculoneuritis. The other striped dolphin showed a subacute systemic CeMV infection, with an intense encephalitis and less meningeal involvement. Morbilliviral encephalitis was considered the cause of death in this dolphin. The common bottlenose dolphin had ankylosing spondylitis. Macroscopic and microscopic finding of the Brucella-infected cases are summarized in Table 2.
3.2. Culture of Brucella
Brucella culture was attempted in 26 cetaceans, including cases with compatible histological lesions, epididymitis, or a positive Bengal Rose test. Three additional dolphins with a positive result in a qPCR against Brucella in brain tissue were included (data not shown). Brucella ceti was isolated in 9/26 cetaceans (7 striped dolphins with neurobrucellosis, 1 striped dolphin with subacute morbilliviral encephalitis, and the common bottlenose dolphin with diskospondylitis). In the 8 striped dolphins, Brucella was cultured from CNS samples (brain, CSF, ventricle swabs, spinal cord), or mesenteric lymph node, and in the bottlenose dolphin, from the diskospondylitic lesion [8] (see Table 2). Neurobrucellosis diagnosis in an additional striped dolphin (N-368/19) was based on highly compatible lesions and a low Ct result in the Brucella qPCR in CSF (data not shown). In this case, Brucella culture was attempted but the sample was contaminated due to problems during storage. A positive Brucella PCR result has prompted the inclusion of this case in the neurobrucellosis group for the serological analysis.
The level of haemolysis did not condition the results of the cELISA, as tested in the preliminary study (data not shown).
Figure 1.
Map showing the stranding location of 10 cetaceans with Brucella infection.
3.3. Serological Investigation
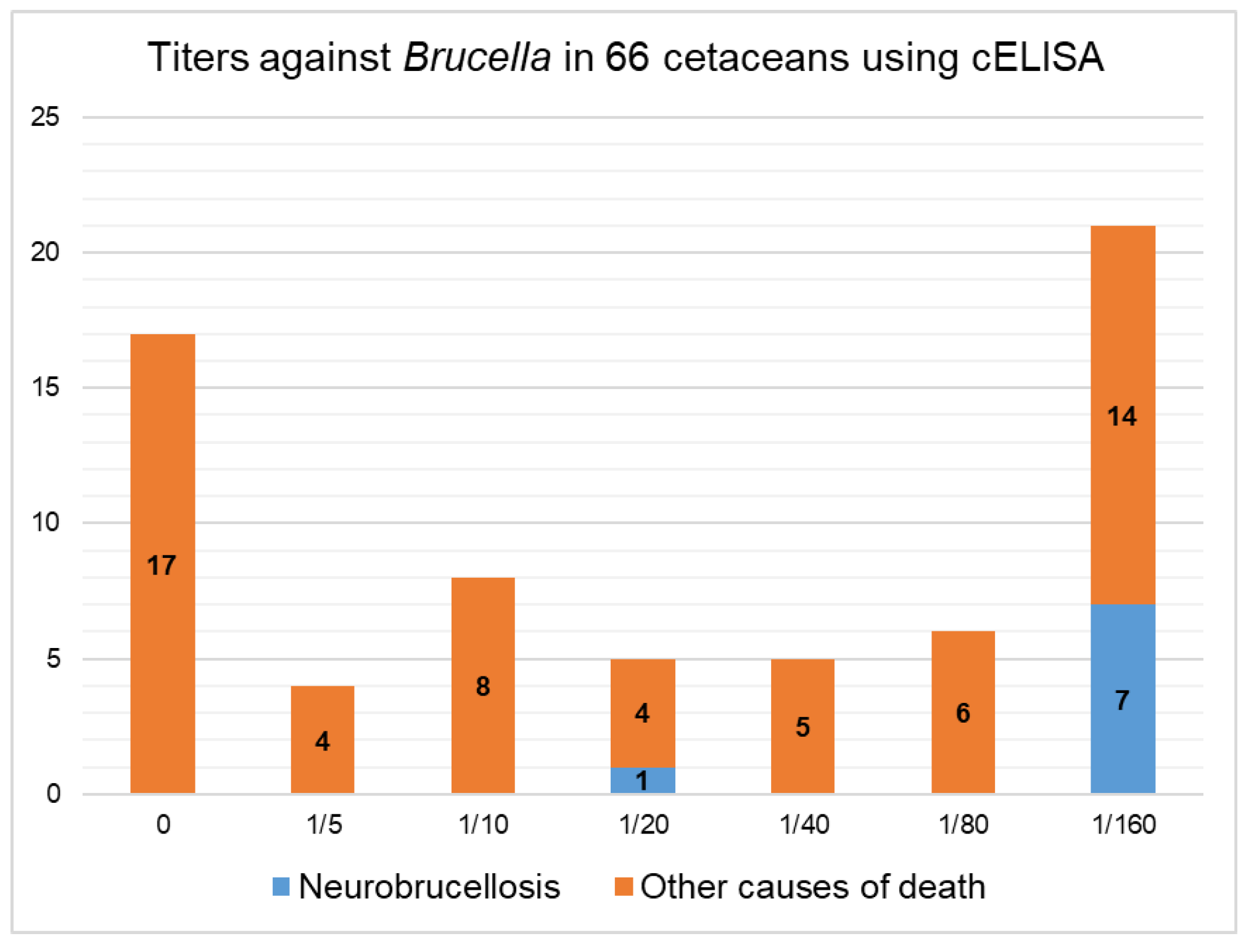
A multispecies cELISA was used in an exploratory study to detect antibodies against the LPS of Brucella sp. in 66 cetaceans, 8 of them infected with Brucella. This test has not been validated for cetaceans, and there is no cut-off established to consider one sample as positive or negative. Furthermore, a panel of reference sera of infected and non-infected cetaceans is not available, and will hardly be so in the future. As a consequence, both sensitivity and specificity of the test for cetacean serum are unknown. All dolphins were tested at serial dilutions from 1:5 to 1:160, and the last dilution with a PI ≥40% was arbitrarily considered the titer of that serum sample. The majority of cetaceans (7 of 8) with Brucella isolation were positive at the last dilution used (1:160). However, a high proportion of sera (21 of 66; 31.8%) were still positive (PI ≥40%) at the 1:160 dilution (see Figure 2). The overall Brucella antibody estimated prevalence was 33.3% for striped dolphins and 31.8% for all cetaceans using the dilution of 1:160 and the cut-off of PI ≥40%. Further serum dilutions performed with neurobrucellosis cases showed titers of 1:320 (n=1), 1:640 (n=2), 1:1280 (n=2), and 1:5120 (n=1). The PI for the different dilutions of each cetacean serum are shown in Supplementary File 1.
Figure 1.
Antibody titer against Brucella using a cELISA in 66 cetaceans highlighting Brucella-infected cases. Vertical axis shows the absolute number of animals.
Figure 1.
Antibody titer against Brucella using a cELISA in 66 cetaceans highlighting Brucella-infected cases. Vertical axis shows the absolute number of animals.
3.1.1. Serological Results Comparison by Cause of Death
Cetaceans investigated serologically are classified by cause of death, species and cELISA results in Table 3. Seropositivity (at dilution 1:160) was only detected in striped dolphins and Risso’s dolphins. Striped dolphins with neurobrucellosis (n=8) had the highest proportion of seropositives (87.5%). High antibody titers (1:160), however, were also observed in striped dolphins dying from bycatch, CeMV infection, PEM, and with cause of death classified as “other”. Animals with neurobrucellosis showed a statistically significant higher proportion of seropositives compared to the groups of bycatch (4/15; 26.6%), striped dolphins not dying from neurobrucellosis (12/49; 24.5%), and the rest of all cetaceans dying from different causes (14/66; 21.2%). Regarding other species, only 2/5 Risso’s dolphins were seropositive, a calf dying from maternal separation and a bycaught adult (Table 2). None of the Rissos’ dolphins investigated serologically showed lesions compatible with brucellosis and in the two cases where culture was attempted, results were negative.
3.1.1. Serology by Age Class
The proportion of seropositive striped dolphins within the different age classes is shown in Table 4. Juveniles, compared to adults, presented a higher proportion of seropositivity (53.9 vs 29.3%) and they comprised the majority of neurobrucellosis cases (20.8 vs 6.3%), although differences were not statistically significant. Seropositivity was 33% for both male and female striped dolphins (data not shown).
3.1.1. Brucella Serology in CeMV-Infected Cetaceans
A CeMV outbreak in the Northwestern Mediterranean Sea accounted for the death of 17 cetaceans from 2016 to 2021, including systemic and chronic CNS cases [14] (see Table 5). High titers against Brucella within this group were frequent in dolphins diagnosed with CeMV-chronic CNS forms. Only one of four CeMV cases where Brucella isolation was attempted yielded a positive result (N-319/16, stranded in 2016, with CNS lesions attributed to CeMV). In 4 of these cases Brucella culture was attempted and was negative.
4. Discussion
The results show that brucellosis is regularly detected in small cetaceans in the western Mediterranean Sea, and it is the most relevant single bacterial cause of death in striped dolphins in the area, confirming previous similar findings from Mediterranean regions [14,18]. B. ceti was isolated in 10 out of 131 (7.6%) necropsied cetaceans, with this figure being the first estimated prevalence of the disease in dolphins from any Mediterranean coastal region. In 8 striped dolphins neurobrucellosis was considered the primary cause of death, whereas B. ceti was considered a comorbidity in one striped dolphin with systemic CeMV infection and in one common bottlenose dolphin with B. ceti diskospondylitis and a CNS mucormycosis [8].
Understanding the pathogenesis of cetacean neurobrucellosis remains elusive, mostly due to the lack of knowledge about the types of immune responses elicited by B. ceti in healthy and in sick cetaceans. It is generally accepted that Th1 responses are relevant in controlling replication of intracellular bacteria, and that a strong humoral (Th2-mediated) response seems to be unprotective, being antibodies a more useful indicator of infection than a proper defense mechanisms (for review, see [19,20]. Accordingly, a study with B. melitensis in mice suggested that antibodies are not decisive in the control of infection [21], although antibodies against LPS have proven to confer certain protection in B. abortus [19,22]. As in other mammals, it may be hypothesized that cetaceans with a predominant Th1 response to B. ceti survive the infection, and a shift to a Th2 response constitutes the hallmark of disease progression. However, to our knowledge, specific T-cell response against Brucella sp. has never been measured in dolphins, and the role of serum antibodies against the bacteria is unknown. Several serologic tests have been used in cetacean species, either developed for terrestrial mammals or adapted to marine mammals [11,23,24,25,26,27,28,29,30]. These studies have shown a high seroprevalence of Brucella-infection ranging from 7.6% to 60%, both in cetaceans and pinnipeds, in many places of the world. However, the sensitivity (Se) and specificity (Sp) of these tests have not been established for cetaceans, and this knowledge is hindered by the lack of reference panel sera for cetaceans. Bearing in mind these limitations we applied a commercial cELISA for Brucella diagnosis in cetaceans, not to establish Se and Sp values for this test, but rather to obtain a more defined picture of the prevalence of B. ceti infection and disease, and to understand the practical utility of serologic results when applied to stranded cetaceans.
We observed a higher seropositivity in striped dolphins with neurobrucellosis than in the total of analyzed cetaceans and than in striped dolphins dying from other causes, considering seropositivity when PI≥40% at a serum dilution of 1:160. Causes of death in seropositive animals, however, were varied and included infectious and non-infectious causes, indicating a low specificity for the cELISA to predict neurobrucellosis. If these cetaceans with high titers represent subclinical forms of latent Brucella infections inducing humoral immunity remains to be studied. In humans, whose pathogenesis of neurobrucellosis is often compared to cetaceans, neurobrucellosis is not the most common manifestation of disease [31,32] and occurs in subacute to chronic infections [33,34]. Thus, it is likely that the diagnosed cases presented in this work represent just the tip of the iceberg of an endemicity of Brucella infection in the Mediterranean, as suggested previously [14] and supported by the high seroprevalence reported in other studies around the world in many cetacean species (see [4] for review).
The lack of specificity of serological tests is often attributed to cross-reactions with other LPS of Gram-negative bacteria. In terrestrial animals, Campylobacter spp., Salmonella spp., Pasteurella spp., Yersinia enterocolytica [11], Francisella tularensis and Vibrio cholerae [35] have been associated to cross-reactions. Terrestrial bacteria, however, can not always be found in marine environments. Salmonella spp. have been isolated from free-ranging cetaceans [36]. In the seropositive dolphins of this paper, Psychrobacter phenylpyruvicus (N-28/21), Photobacterium damselae (N-557/17, N-232/18, N-42/18), Pseudomonas spp. (N-557/17), Vibrio sp. (N-232/18, N-329/18), and Escherichia coli (N-329/18) have been isolated. The pathogenicity of some of these species is unclear and their role in cross-reaction is speculative.
The presence of high seropositivity in the cases of neurobrucellosis probably indicates that the humoral response is not effective against the pathogen. In our cases, Th1 response cytokines (IL-1 and TNFα) have been detected in Brucella-induced meningitis by immunohistochemistry, which could suggest a role of this type of response in brucellosis [37]. As speculated by the authors, it may be that in these cases, for unknown reasons, cellular response is ineffective. Polymorphisms in some molecules of the immune system may also play a role in individual susceptibility [38,39].
After analyzing the serological results, the authors hypothesized that the rise of antibodies, instead of conferring protection, contributes to disease progression in animals with neurobrucellosis. Nonetheless, the authors failed to find literature supporting that. Antibodies in humans with brucellosis can persist several months after the remission of symptoms, clinical relapse [31,32] and, in areas with endemicity, repeated infections [31]. In rats inoculated with B. abortus antigens, antibodies are present a minimum of 120 days [40]. There is a report of a captive bottlenose dolphin with Brucella osteomyelitis with sustained high antibody titers against Brucella [41]. Considering this, it is possible that antibodies would be indicative of chronicity rather than accountable for disease development.
Interestingly, the authors observed that juvenile striped dolphins were the age group with higher seroprevalence and proportion of neurobrucellosis. In the Italian coast, similar results have been reported, with 6/8 striped dolphins with brucellosis being juveniles [18]. In our cases, this may be due to a larger contribution of anthropogenic interactions as cause of death for adults (21/64; 32.8%), diminishing the relative importance of neurobrucellosis. Immune immaturity as a predisposing cause for neurologic form of brucellosis in juveniles, however, can not be discarded. The results from this work contrast with some serologic in human patients, where they found a positive correlation between the age and seroprevalence of Brucella antibodies [42,43].
A cluster of Brucella-seropositive dolphins occurred in the period 2018-2021, in CNS-localized forms of CeMV (n=5) (see Supplementary File 1). The authors speculate that the previous immune suppression of the systemic phase prompted the increased replication and transmission of subclinical Brucella, without causing death, and the persistence of Brucella antibodies until the development of the chronic form of CeMV. Another hypothesis is that the immunosuppressive acute phase transiently limited antibody production. At least in the present cases, there is no direct correlation between the massive immunesuppression in acute CeMV infection and the development of neurobrucellosis. In other publications the two coinfections are more frequent, with 4/8 striped dolphins testing positive for both pathogens [18].
5. Conclusions
A seroprevalence of 31.8% against Brucella LPS antigens in cetaceans from the Catalan coast of the Mediterranean has been detected. This percentage is significantly higher in cetaceans with Brucella infection (87.5%). Seropositive species include striped dolphins and Risso’s dolphins.
The cELISA designed for ruminants and pigs in serum samples of cetaceans is a potentially sensitive method to predict neurobrucellosis, but the specificity is low. This may reflect a proportion of subclinical carriers of the bacteria. Cross-reactions or subclinical forms of brucellosis are possible causes of high antibody titers. More research efforts are needed to dilucidate the epidemiology of the infection and the latency of bacteria in target organs. Cetaceans with chronic forms of CeMV seem to have higher antibody titers against Brucella. CeMV infection, in the acute immunosuppressive form, may enhance Brucella replication in carrier individuals and increase the horizontal transmission of the bacteria.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Table S1: Percentage of inhibition of all cELISA tested cetaceans; Table S2: Biometric data, ancillary tests and cause of death of the 131 studied cetaceans.
Author Contributions
Conceptualization, M.D.; methodology, M.D. and L.M.; validation, M.D.; formal analysis, A.A.; investigation, M.D., S.W., L.P., L.G., A.P., and L.M.; resources, M.D.; data curation, L.M.; writing—original draft preparation, M.D., L.M. and M.C.; writing—review and editing, A.P., S.W., and M.C.; visualization, L.M.; supervision, M.D.; project administration, M.D.; funding acquisition, M.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Servei de Fauna i Flora (Generalitat de Catalunya), and Servei de Diagnòstic de Patologia Veterinària (Universitat Autònoma de Barcelona), permit number AG-2022-1439.
Institutional Review Board Statement
Not applicable (animals included in this study stranded in the coast and died naturally or were euthanized under authorized veterinary services).
Data Availability Statement
data supporting reported results can be found in Supplementary Materials.
Acknowledgments
We thank Blanca Pérez and Aida Neira from the Servei de Diagnòstic de Patologia Veterinària, Universitat Autònoma de Barcelona, and Maite Martin and Marta Perez. We thank the brucellosis research group from CITA de Aragón (MJ de Miguel, S. Serrano, S. Andrés-Barranco and PM Muñoz), for culture and identification of Brucella isolates. Thanks to Silvio Špičić Laboratory for Bacterial Zoonoses, Croatian Veterinary Institute, Zagreb, Croatia ST 27 for kindly providing DNA of Brucella ceti CRO350.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ewalt, D.R.; Payeur, J.B.; Martin, B.M.; Cummins, D.R.; Miller, W.G. Characteristics of a Brucella Species from a Bottlenose Dolphin (Tursiops truncatus). J. Vet. Diagn. Invest. 1994, 6, 448–452. [Google Scholar] [CrossRef]
- Foster, G.; Osterman, B.S.; Godfroid, J.; Jacques, I.; Cloeckaert, A. Brucella ceti Sp. Nov. and Brucella pinnipedialis Sp. Nov. for Brucella Strains with Cetaceans and Seals as Their Preferred Hosts. Int. J. Syst. Evol. Microbiol. 2007, 57, 2688–2693. [Google Scholar] [CrossRef]
- Nymo, I.H.; Tryland, M.; Godfroid, J. A Review of Brucella Infection in Marine Mammals, with Special Emphasis on Brucella pinnipedialis in the Hooded Seal (Cystophora cristata). Vet. Res. 2011, 42, 93. [Google Scholar] [CrossRef]
- Guzmán-Verri, C.; González-Barrientos, R.; Hernández-Mora, G.; Morales, J.-A.; Baquero-Calvo, E.; Chaves-Olarte, E.; Moreno, E. Brucella ceti and Brucellosis in Cetaceans. Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef]
- Van Bressem, M.; Raga, A.; Di Guardo, G.; Jepson, P.; Duignan, P.; Siebert, U.; Barrett, T.; Santos, Mc.; Moreno, I.; Siciliano, S.; et al. Emerging Infectious Diseases in Cetaceans Worldwide and the Possible Role of Environmental Stressors. Dis. Aquat. Organ. 2009, 86, 143–157. [Google Scholar] [CrossRef]
- Alba, P.; Terracciano, G.; Franco, A.; Lorenzetti, S.; Cocumelli, C.; Fichi, G.; Eleni, C.; Zygmunt, M.S.; Cloeckaert, A.; Battisti, A. The Presence of Brucella ceti ST26 in a Striped Dolphin (Stenella coeruleoalba) with Meningoencephalitis from the Mediterranean Sea. Vet. Microbiol. 2013, 164, 158–163. [Google Scholar] [CrossRef]
- Garofolo, G.; Zilli, K.; Troiano, P.; Petrella, A.; Marotta, F.; Di Serafino, G.; Ancora, M.; Di Giannatale, E. Brucella ceti from Two Striped Dolphins Stranded on the Apulia Coastline, Italy. J. Med. Microbiol. 2014, 63, 325–329. [Google Scholar] [CrossRef]
- Isidoro-Ayza, M.; Ruiz-Villalobos, N.; Pérez, L.; Guzmán-Verri, C.; Muñoz, P.M.; Alegre, F.; Barberán, M.; Chacón-Díaz, C.; Chaves-Olarte, E.; González-Barrientos, R.; et al. Brucella ceti Infection in Dolphins from the Western Mediterranean Sea. BMC Vet. Res. 2014, 10, 206. [Google Scholar] [CrossRef]
- Cvetnić, Ž.; Duvnjak, S.; Đuras, M.; Gomerčić, T.; Reil, I.; Zdelar-Tuk, M.; Špičić, S. Evidence of Brucella Strain ST27 in Bottlenose Dolphin (Tursiops truncatus) in Europe. Vet. Microbiol. 2016, 196, 93–97. [Google Scholar] [CrossRef]
- Grattarola, C.; Petrella, A.; Lucifora, G.; Di Francesco, G.; Di Nocera, F.; Pintore, A.; Cocumelli, C.; Terracciano, G.; Battisti, A.; Di Renzo, L.; et al. Brucella ceti Infection in Striped Dolphins from Italian Seas: Associated Lesions and Epidemiological Data. Pathogens 2023, 12, 1034. [Google Scholar] [CrossRef] [PubMed]
- Van Bressem, M.; Van Waerebeek, K.; Raga, J.A.; Godfroid, J.; Brew, S.D.; MacMillan, A.P. Serological Evidence of Brucella Species Infection in Odontocetes from the South Pacific and the Mediterranean. Vet. Rec. 2001, 148, 657–661. [Google Scholar] [CrossRef]
- Pugliares, K.R.; Bogomolni, A.; Touhey, K.M.; Herzig, S.M.; Harry, C.T.; Moore, M.J. Marine Mammal Necropsy : An Introductory Guide for Stranding Responders and Field Biologists; Woods Hole Oceanographic Institution: Woods Hole, MA, 2007. [Google Scholar]
- Kuiken, Thijs; García-Hartmann, Manuel Proceedings of the First European Cetacean Society Workshop on Cetacean Pathology: Dissection Techniques and Tissue Sampling.; Leiden, The Netherlands, 1991; p. 39.
- Cuvertoret-Sanz, M.; López-Figueroa, C.; O’Byrne, A.; Canturri, A.; Martí-Garcia, B.; Pintado, E.; Pérez, L.; Ganges, L.; Cobos, A.; Abarca, M.; et al. Causes of Cetacean Stranding and Death on the Catalonian Coast (Western Mediterranean Sea), 2012-2019. Dis. Aquat. Organ. 2020, 142, 239–253. [Google Scholar] [CrossRef]
- López-Goñi, I.; García-Yoldi, D.; Marín, C.M.; De Miguel, M.J.; Barquero-Calvo, E.; Guzmán-Verri, C.; Albert, D.; Garin-Bastuji, B. New Bruce-Ladder Multiplex PCR Assay for the Biovar Typing of Brucella suis and the Discrimination of Brucella suis and Brucella canis. Vet. Microbiol. 2011, 154, 152–155. [Google Scholar] [CrossRef]
- Calzada, Nuria; Aguilar, Alex; Lockyer, Christina; Grau, Esteve Patterns of Growth and Physical Maturity in the Western Mediterranean Striped Dolphin, Stenella coeruleoalba (Cetacea: Odontoceti). Can. J. Zool. 1997, 75, 632–637. [CrossRef]
- Calzada, N.; Aguilar, A. Flipper Development in the Mediterranean Striped Dolphin (Stenella coeruleoalba). Anat. Rec. 1996, 245, 708–714. [Google Scholar] [CrossRef]
- Garofolo, G.; Petrella, A.; Lucifora, G.; Di Francesco, G.; Di Guardo, G.; Pautasso, A.; Iulini, B.; Varello, K.; Giorda, F.; Goria, M.; et al. Occurrence of Brucella ceti in Striped Dolphins from Italian Seas. PLOS ONE 2020, 15, e0240178. [Google Scholar] [CrossRef]
- Dorneles, E.M.S.; Teixeira-Carvalho, A.; Araújo, M.S.S.; Sriranganathan, N.; Lage, A.P. Immune Response Triggered by Brucella abortus Following Infection or Vaccination. Vaccine 2015, 33, 3659–3666. [Google Scholar] [CrossRef]
- De Figueiredo, P.; Ficht, T.A.; Rice-Ficht, A.; Rossetti, C.A.; Adams, L.G. Pathogenesis and Immunobiology of Brucellosis. Am. J. Pathol. 2015, 185, 1505–1517. [Google Scholar] [CrossRef]
- Dadelahi, A.S.; Abushahba, M.F.N.; Ponzilacqua-Silva, B.; Chambers, C.A.; Moley, C.R.; Lacey, C.A.; Dent, A.L.; Skyberg, J.A. Interactions between B Cells and T Follicular Regulatory Cells Enhance Susceptibility to Brucella Infection Independent of the Anti-Brucella Humoral Response. PLOS Pathog. 2023, 19, e1011672. [Google Scholar] [CrossRef]
- Cloeckaert, A.; Jacques, I.; Bosseray, N.; Limet, J.N.; Bowden, R.; Dubray, G.; Plommet, M. Protection Conferred on Mice by Monoclonal Antibodies Directed against Outer-Membrane-Protein Antigens of Brucella. J. Med. Microbiol. 1991, 34, 175–180. [Google Scholar] [CrossRef]
- Hernández-Mora, G.; Palacios-Alfaro, J.D.; González-Barrientos, R. Wildlife Reservoirs of Brucellosis: Brucella in Aquatic Environments.
- Meegan, J.; Field, C.; Sidor, I.; Romano, T.; Casinghino, S.; Smith, C.R.; Kashinsky, L.; Fair, P.A.; Bossart, G.; Wells, R.; et al. Development, Validation, and Utilization of a Competitive Enzyme-Linked Immunosorbent Assay for the Detection of Antibodies against Brucella Species in Marine Mammals. J. Vet. Diagn. Invest. 2010, 22, 856–862. [Google Scholar] [CrossRef]
- Prenger-Berninghoff, E.; Siebert, U.; Stede, M.; König, A.; Weiß, R.; Baljer, G. Incidence of Brucella Species in Marine Mammals of the German North Sea. Dis. Aquat. Organ. 2008, 81, 65–71. [Google Scholar] [CrossRef]
- Lynch, M.; Duignan, P.J.; Taylor, T.; Nielsen, O.; Kirkwood, R.; Gibbens, J.; Arnould, J.P.Y. Epizootiology of Brucella Infection in Australian Fur Seals. J. Wildl. Dis. 2011, 47, 352–363. [Google Scholar] [CrossRef]
- Norman, S.A.; Delaney, M.A.; Haman, K.H.; Thomas, A.C.; Godfroid, J.; Larsen, A.K.; Nymo, I.H.; Robbe-Austerman, S.; Quance, C.; Rhyan, J.C.; et al. Application of Real-Time Quantitative PCR Assays for Detecting Marine Brucella spp. in Fish. J. Vet. Diagn. Invest. 2018, 30, 150–154. [Google Scholar] [CrossRef]
- Hoover-Miller, A.; Dunn, J.; Field, C.; Blundell, G.; Atkinson, S. Seroprevalence of Brucella Antibodies in Harbor Seals in Alaska, USA, with Age, Regional, and Reproductive Comparisons. Dis. Aquat. Organ. 2017, 126, 1–12. [Google Scholar] [CrossRef]
- Kershaw, J.; Stubberfield, E.; Foster, G.; Brownlow, A.; Hall, A.; Perrett, L. Exposure of Harbour Seals Phoca vitulina to Brucella in Declining Populations across Scotland. Dis. Aquat. Organ. 2017, 126, 13–23. [Google Scholar] [CrossRef]
- Kroese, M.V.; Beckers, L.; Bisselink, Y.J.W.M.; Brasseur, S.; Van Tulden, P.W.; Koene, M.G.J.; Roest, H.I.J.; Ruuls, R.C.; Backer, J.A.; IJzer, J.; et al. Brucella pinnipedialis in Grey Seals (Halichoerus gryphus) and Harbor Seals (Phoca vitulina) in the Netherlands. J. Wildl. Dis. 2018, 54, 439. [Google Scholar] [CrossRef]
- Ruiz-Mesa, J.D.; Sánchez-Gonzalez, J.; Reguera, J.M.; Martín, L.; Lopez-Palmero, S.; Colmenero, J.D. Rose Bengal Test: Diagnostic Yield and Use for the Rapid Diagnosis of Human Brucellosis in Emergency Departments in Endemic Areas. Clin. Microbiol. Infect. 2005, 11, 221–225. [Google Scholar] [CrossRef]
- Casanova, A.; Ariza, J.; Rubio, M.; Masuet, C.; Díaz, R. BrucellaCapt versus Classical Tests in the Serological Diagnosis and Management of Human Brucellosis. Clin. Vaccine Immunol. 2009, 16, 844–851. [Google Scholar] [CrossRef]
- Zhao, S.; Cheng, Y.; Liao, Y.; Zhang, Z.; Yin, X.; Shi, S. Treatment Efficacy and Risk Factors of Neurobrucellosis. Med. Sci. Monit. 2016, 22, 1005–1012. [Google Scholar] [CrossRef]
- Soares, C.N.; Angelim, A.I.M.; Brandão, C.O.; Santos, R.Q.; Mehta, R.; Silva, M.T.T.D. Neurobrucellosis: The Great Mimicker. Rev. Soc. Bras. Med. Trop. 2022, 55, e0567–2021. [Google Scholar] [CrossRef]
- Solís García Del Pozo, J.; Lorente Ortuño, S.; Navarro, E.; Solera, J. Detection of IgM Antibrucella Antibody in the Absence of IgGs: A Challenge for the Clinical Interpretation of Brucella Serology. PLoS Negl. Trop. Dis. 2014, 8, e3390. [Google Scholar] [CrossRef]
- Grattarola, C.; Gallina, S.; Giorda, F.; Pautasso, A.; Ballardini, M.; Iulini, B.; Varello, K.; Goria, M.; Peletto, S.; Masoero, L.; et al. First Report of Salmonella 1,4,[5],12:I:- In Free-Ranging Striped Dolphins (Stenella coeruleoalba), Italy. Sci. Rep. 2019, 9, 6061. [Google Scholar] [CrossRef] [PubMed]
- Rebollada-Merino, A.; Giorda, F.; Pumarola, M.; Martino, L.; Gomez-Buendia, A.; Romani-Cremaschi, U.; Casalone, C.; Mattioda, V.; Di Nocera, F.; Lucifora, G.; et al. Neurobrucellosis (Brucella ceti) in Striped Dolphins (Stenella coeruleoalba): Immunohistochemical Studies on Immune Response and Neuroinflammation. Vet. Pathol. 2024, 03009858241250336. [Google Scholar] [CrossRef]
- Bravo, M.J.; Colmenero, J.D.; Martín, J.; Alonso, A.; Caballero, A. Polymorphism of the Transmembrane Region of the MICA Gene and Human Brucellosis. Tissue Antigens 2007, 69, 358–360. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; He, H.-Y.; Ojha, S.C.; Shi, H.; Sun, C.-F.; Deng, C.-L.; Sheng, Y.-J. Association of IL-6, IL-10 and TGF-Β1 Gene Polymorphisms with Brucellosis: A Systematic Review with Meta-Analysis. Microb. Pathog. 2019, 135, 103640. [Google Scholar] [CrossRef]
- Khatun, M.; Islam, A.; Baek, B.K. Comparative Analysis of Humoral Immune Response and Cognate Antigen Detection in Experimentally Infected Sprague Dawley Rats with Brucella abortus Biotype 1. Vector-Borne Zoonotic Dis. 2024, 24, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Goertz, C.E.C.; Frasca, S.; Bohach, G.A.; Cowan, D.F.; Buck, J.D.; French, R.A.; De Guise, S.; Maratea, J.; Hinckley, L.; Ewalt, D.; et al. Brucella Sp. Vertebral Osteomyelitis with Intercurrent Fatal Staphylococcus aureus Toxigenic Enteritis in a Bottlenose Dolphin (Tursiops truncatus). J. Vet. Diagn. Invest. 2011, 23, 845–851. [Google Scholar] [CrossRef]
- Islam, Md.A.; Khatun, Mst. M.; Werre, S.R.; Sriranganathan, N.; Boyle, S.M. A Review of Brucella Seroprevalence among Humans and Animals in Bangladesh with Special Emphasis on Epidemiology, Risk Factors and Control Opportunities. Vet. Microbiol. 2013, 166, 317–326. [Google Scholar] [CrossRef]
- Abo-Shehada, M.N.; Odeh, J.S.; Abu-Essud, M.; Abuharfeil, N. Seroprevalence of Brucellosis among High Risk People in Northern Jordan. Int. J. Epidemiol. 1996, 25, 450–454. [Google Scholar] [CrossRef]

Table 1.
Biometric data and stranding information fo the 10 dolphins with brucellosis. J = juvenile. Ad = Adult. D = found dead. A = stranded alive. All animals are striped dolphins except N-275/12, which is a common bottlenose dolphin.
Table 1.
Biometric data and stranding information fo the 10 dolphins with brucellosis. J = juvenile. Ad = Adult. D = found dead. A = stranded alive. All animals are striped dolphins except N-275/12, which is a common bottlenose dolphin.
| ID | Date | Place of stranding | Age class | Age estimation | Length (cm) | Weight (Kg) | Sex | A/D |
|---|---|---|---|---|---|---|---|---|
|
N-275/12 |
2012/05/23 |
Badalona | Ad | - | 300 | Unknown | M | D |
|
N-301/12 |
2012/06/03 |
Cunit | Ad | 6.25 | 184 | 54.5 | F | A |
| N-260/16 | 2016/06/19 |
Sant Carles de la Ràpita |
J | 3.88 | 170 | 39.5 | F | D |
|
N-319/16 |
2016/09/08 |
Tarragona | J | 5.35 | 180 | 47 | F | A |
| N-168/17 | 2017/04/18 |
L’Escala |
Ad | 10.39 | 193 | 79 | M | A |
|
N-274/18 |
2018/06/24 |
Sant Jaume d’Enveja | J | 2.43 | 152 | 38 | M |
A |
|
N-314/19 |
2019/09/01 |
Vilanova i la Geltrú |
J | 2.8 | 159 | 41 | F | A |
|
N-368/19 |
2019/10/12 |
Cambrils | J | 5.23 | 175 | 47 | M | A |
| N-333/22 | 2022/09/22 |
Llacuna dels Alfacs | Ad | 6.15 | 180 | 47.5 | M | A |
| N-215/23 | 2023/06/22 | Sant Pere Pescador | Ad | 13.79 | 197 | 59 | M | A |
Table 2.
Summary of the signalment and pathological findings in 9 striped dolphins and a bottlenose dolphin (N-275/12) (*, previously published in [8]) diagnosed with brucellosis or with Brucella infection. CSF = cerebrospinal fluid. Only lesions attributable to Brucella ceti or relevant to cause of death are included.
Table 2.
Summary of the signalment and pathological findings in 9 striped dolphins and a bottlenose dolphin (N-275/12) (*, previously published in [8]) diagnosed with brucellosis or with Brucella infection. CSF = cerebrospinal fluid. Only lesions attributable to Brucella ceti or relevant to cause of death are included.
| ID | Macroscopic lesions | Histopathology | Brucella culture | Others | Cause of death | Available serum |
|---|---|---|---|---|---|---|
| N-275/12* | Chronic suppurative diskospondylitis Multifocal brain malacia |
Mycotic pyogranulomatous-necrotizing meningoencephalomyelitis |
Positive (vertebral abscess) | Mycotic encephalitis | Mycotic encephalitis | No |
| N-301/12* | Not relevant | Non-suppurative meningoencephalitis, diffuse, more intense in cerebellum, brainstem and spinal cord; choroiditis; radiculoneuritis | Positive (brain) | - | Neurobrucellosis | Yes |
| N-260/16 | Emaciation; cerebral edema; absence of recent ingesta | Non-suppurative meningoencephalitis, diffuse, more intense in brainstem; choroiditis; radiculoneuritis, multifocal and milder | Positive (brain) | - | Neurobrucellosis | Yes |
| N-319/16 | Low body condition; absence of recent ingesta; meningeal congestion | Non-suppurative meningoencephalitis | Positive (brain, ventricle swab) | CeMV intensely positive (RT-PCR and IHC) | CeMV encephalitis | Yes |
| N-168/17 | Absence of recent ingesta; meningeal and CSF turbidity |
Non-suppurative meningitis, more intense in brainstem and cerebellum | Positive (lateral ventricle swab, spinal cord, mesenteric LN, brain, CSF) | - | Neurobrucellosis | Yes |
| N-274/18 | Loss of body condition; brain edema and meningeal hemorrhage |
Non-suppurative meningitis, diffuse, with multifocal encephalitis; choroiditis | Positive (lateral ventricle swab) | Aeromonas sobria and P. damselae in lung, liver, CNS | Neurobrucellosis | Yes |
| N-314/19 | CSF turbidity; brain edema and meningeal hemorrhage |
Non-suppurative meningoencephalitis, more intense in brainstem, cerebellum and thalamus; choroiditis; radiculoneuritis, multifocal and milder | Positive (CSF) | - | Neurobrucellosis | Yes |
| N-368/19 | Emaciation; absence of recent ingesta; CSF turbidity | Non-suppurative meningitis, localized in brainstem and, with less intensity, cerebellum |
Negative (contaminated sample) PCR intensely positive in CSF. |
- | Neurobrucellosis | Yes |
| N-333/22 | Slight loss of body condition; absence of regent ingesta; brain edema; meningeal and CSF turbidity | Non-suppurative meningitis, localized in brainstem and cerebellum, choroiditis | Positive (brain, CSF) | - | Neurobrucellosis | Yes |
| N-215/23 | Absence of regent ingesta; CSF turbidity; hydrocephalus |
Non-suppurative meningitis, especially in brainstem and, with lesser intensity, cerebellum; choroiditis; radiculoneuritis, multifocal and milder |
Positive (CSF, ventricular swab) | - | Neurobrucellosis | No |
Table 3.
Antibody titers in the cELISA against Brucella LPS in serum samples of 66 cetaceans. The column on the right (%Pos) shows the percentage of seropositives (PI ≥40% at dilution 1:160) in each group. *In one dolphin, the cause of death was encephalitis by CeMV. ♦Chronic forms of CeMV stranded in the period 2018-2021. Sc = Stenella coeruleoalba; Gg = Grampus griseus; Tt = Tursiops truncatus; Dd = Delphinus delphis; Gm = Globicephala melas.
Table 3.
Antibody titers in the cELISA against Brucella LPS in serum samples of 66 cetaceans. The column on the right (%Pos) shows the percentage of seropositives (PI ≥40% at dilution 1:160) in each group. *In one dolphin, the cause of death was encephalitis by CeMV. ♦Chronic forms of CeMV stranded in the period 2018-2021. Sc = Stenella coeruleoalba; Gg = Grampus griseus; Tt = Tursiops truncatus; Dd = Delphinus delphis; Gm = Globicephala melas.
| Species | Cause of death | n | Antibody titer | |||||||
| >1:5 | 1:5 | 1:10 | 1:20 | 1:40 | 1:80 | 1:160 | %Pos | |||
| Sc | Neurobrucellosis* | 8 | 1 | 7 | 87.5% | |||||
| Bycatch | 15 | 4 | 1 | 3 | 1 | 2 | 4 | 26.6% | ||
| CeMV | 12 | 5 | 1 | 1 | 1 | 4♦ | 33.3% | |||
| Infectious** | 6 | 1 | 1 | 2 | 2 | 0% | ||||
| PEM | 4 | 1 | 1 | 2 | 50% | |||||
| Mother-calf separation | 2 | 1 | 1 | 0% | ||||||
| Other*** | 7 | 1 | 2 | 2 | 2 | 28.6% | ||||
| Unknown | 3 | 1 | 2 | 0% | ||||||
| TOTAL | 57 | 13 | 4 | 7 | 5 | 4 | 5 | 19 | 33.3% | |
| Gg | Sinusitis Crassicauda | 3 | 2 | 1 | 0% | |||||
| Bycatch | 1 | 1 | 100% | |||||||
| Mother-calf separation | 1 | 1 | 100% | |||||||
| TOTAL | 5 | 2 | 1 | 2 | 40% | |||||
| Tt | Mother-calf separation | 1 | 1 | 0% | ||||||
| Erysipelas | 1 | 1 | 0% | |||||||
| TOTAL | 2 | 1 | 1 | 0% | ||||||
| Dd | Unknown | 1 | 1 | 0% | ||||||
| Gm | CeMV | 1 | 1 | 0% | ||||||
| TOTAL | 66 | 45 | 21 | 31.8% | ||||||
Table 4.
Seropositivity for Brucella (left column) and neurobrucellosis cases (right column) by age class. Serum was available in n=57 striped dolphins. Right column includes all striped dolphins, with or without serological analysis, with isolation of Brucella ceti.
Table 4.
Seropositivity for Brucella (left column) and neurobrucellosis cases (right column) by age class. Serum was available in n=57 striped dolphins. Right column includes all striped dolphins, with or without serological analysis, with isolation of Brucella ceti.
| Seropositives (%) | Neurobrucellosis (%) | |
| Adults | 12/41 (29.3%) | 4/64 (6.3%) |
| Juveniles | 7/13 (53.9%) | 5/24 (20.8%) |
| Calves | 0/3 (0%) | 0/14 (0%) |
| Fetus | 0 | 0/1 (0%) |
| Total | 19/57 (33.3%) | 9/88 (14%) |
Table 5.
Antibody titers against Brucella spp. in cetaceans infected with CeMV. *Brucella coinfection. Sc = Stenella coeruleoalba; Gm = Globicephala melas; J = juvenile; Ad = Adult. Bold indicates seropositivity.
Table 5.
Antibody titers against Brucella spp. in cetaceans infected with CeMV. *Brucella coinfection. Sc = Stenella coeruleoalba; Gm = Globicephala melas; J = juvenile; Ad = Adult. Bold indicates seropositivity.
| ID | Species | Age group | CeMV form | Titer |
| N-319/16 | Sc | J | Systemic* | 1/160 |
| N-044/17 | Sc | J | Systemic | >1:5 |
| N-045/17 | Sc | J | Systemic | >1:5 |
| N-077/17 | Sc | J | Systemic | >1:5 |
| N-454/17 | Sc | J | Systemic | >1:5 |
| N-488/17 | Sc | Ad | Systemic | 1/5 |
| N-497/17 | Sc | Ad | Systemic | 1/10 |
| N-604/17 | Sc | Ad | Chronic | >1:5 |
| N-001/18 | Sc | Ad | Chronic | 1/40 |
| N-232/18 | Sc | J | Chronic | 1/160 |
| N-293/18 | Sc | Ad | Chronic | 1/160 |
| N-362/18 | Sc | Ad | Chronic | 1/160 |
| N-361/19 | Sc | J | Chronic | 1/160 |
| N-334/21 | Sc | Ad | Chronic | 1/80 |
| N-023/21 | Gm | J | Chronic | >1:5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



