
Submitted:
02 July 2024
Posted:
03 July 2024
You are already at the latest version
Abstract
Rhodiola rosea, a perennial herb from the Crassulaceae family, contains the bioactive compound salidroside, recognized as an adaptogen with significant therapeutic potential for bone metabolism. Salidroside promotes osteoblast proliferation and differentiation by activating critical signaling pathways, including bone morphogenetic protein-2 and adenosine monophosphate-activated protein kinase, essential for bone formation and growth. It enhances osteogenic activity by increasing alkaline phosphatase activity and mineralization markers, while upregulating key transcription factors such as runt-related transcription factor 2 and osterix. Additionally, salidroside facilitates angiogenesis via the hypoxia-inducible factor 1-alpha and vascular endothelial growth factor pathway, crucial for coupling bone formation with vascular support. Its antioxidant properties offer protection against bone loss by reducing oxidative stress and promoting osteogenic differentiation through the nuclear factor erythroid 2–related factor 2 pathway. Salidroside's ability to counteract the negative effects of glucocorticoids on bone cells and prevent steroid-induced osteonecrosis, along with its anti-inflammatory properties, enhances its potential in treating conditions like osteoarthritis. These attributes underscore its multifaceted role in bone metabolism. This article reviews the literature, including in vitro and in vivo studies primarily on rodent models, demonstrating salidroside's efficacy in mitigating bone loss, preserving bone microarchitecture, and accelerating fracture healing. Future research directions to explore salidroside’s applications in bone-related diseases are also highlighted.
Keywords:
salidroside
; adaptogens
; bone metabolism
; osteoporosis
; Rhodiola rosea
; bone loss
; fracture healing
; osteoblasts
1. Introduction
Rhodiola rosea (R. rosea) from the Crassulaceae plant family is named after the Greek words “rodia” or “rodion”, which refer to its pink color when dried. It goes by various names such as “golden root” due to its shiny appearance after purification and “arctic root” because of its habitat in cold regions. This small perennial herb thrives in harsh environmental conditions and low temperatures. It can be found in polar regions across North America, Europe, and Asia, characterized by compact stem leaves up to 40 cm tall and roots with thick rhizomes that release a distinct rosy fragrance when crushed. These rhizomes contain extracts responsible for the plant's medicinal properties [1]. R. rosea is categorized as an adaptogen, meaning it contains active components that help the human body adjust to environmental stressors. Its properties have been utilized for centuries in cases such as chronic stress, fatigue, insomnia, and concentration problems [2,3]. R. rosea has been the subject of numerous studies aimed at understanding its mechanisms of action and establishing its potential role in medicine [4,5]. It was noted early on that the medicinal properties of this substance are largely attributed to salidroside (SAL), a compound with various beneficial effects [6]. SAL belongs to the group of Phenylethanoid glycosides and is distinguished by its high biological availability among compounds in this group due to its relatively simple structure [7]. Additional compounds such as rosarin, rosin, and rosavin have been uncovered in subsequent studies and contribute to the unique properties of R. rosea [8]. However, SAL continues to garner the most attention, with increasing research on its effects annually [9]. Extensive analysis of this compound involves studying its regenerative and anti-inflammatory effects [10]. Other studies have demonstrated SAL's beneficial effect on pulmonary fibrosis in animals, protection of cardiac muscle from exhaustive exercise-induced injury, inhibition of myocardial remodeling following a heart attack, neuroprotective effects in Parkinson's disease and Alzheimer's disease, while also providing protection against ischemic stroke [11]. SAL exhibits anti-inflammatory actions in liver fibrosis progression and attenuates nonalcoholic fatty liver disease induced by a high-fat diet, while significantly influencing metabolic processes in bone tissue (demonstrating multifaceted osteoprotective effects) [12]. This last property could be most valuable, considering the steadily rising number of patients with diminished bone mineral density (BMD). The number of osteoporotic fractures in European Union countries reached 3.5 million in 2010 and is predicted to rise to 4.5 million cases by 2025, leading to an increase in research dedicated to prevent and treat this condition [13,14]. The escalating numbers not only reduce the quality of life for each patient but also burden the healthcare system with enormous costs. In 2019, Great Britain, Switzerland, and EU countries combined allocated more than €56 billion for treating osteoporosis (OP) and osteoporotic fractures, sparking increased interest in substances that could offer new and more effective methods for treating bone tissue diseases [15]. This study aims to systematically summarize the current knowledge on SAL as a promising molecule for preventing and treating OP and other bone diseases. The authors will focus on presenting the critical mechanisms by which SAL influences cellular and molecular levels, considering both in vitro and in vivo studies, including an exploration of its potential applications in clinical settings [16].
2. Biochemical Structure of Salidroside
SAL is a chemical compound belonging to the group of phenylpropanoid glycosides with the molecular formula C14H20O7 [17]. Its full chemical name is 2-(4-hydroxyphenyl)ethyl O-β-D-glucopyranoside [18]. The biochemical structure of SAL consists of an aglycone (tyrosol), which features a phenyl skeleton with a hydroxyl group (-OH) at the para position (4) and an ethyl group (-CH2CH2OH) attached to the phenyl ring [19]. The aglycone is linked to a glucose molecule via a glycosidic bond, forming a β-D-glucopyranoside [20]. Additionally, SAL is water-soluble [5]. The chemical structure of SAL is depicted in Figure 1.
3. Influence of Salidroside on Bone Metabolism
3.1. In vitro studies
3.1.1. Proliferation and Differentiation of Osteoblast Precursors
Chen et al. examined the effects of SAL on the proliferation of C3H10T1/2 (mouse pluripotent mesenchymal stem cell-like fibroblasts) and MC3T3-E1 (osteoblast precursors from mouse calvariae) cells. Cells were seeded in 96-well microplates (3 × 103 cells in 0.2 ml per well) and treated with various concentrations of SAL for 48 hours. Proliferation was assessed using the MTT assay (colorimetric assay for assessing cell metabolic activity). In C3H10T1/2 cells, SAL induced a slight proliferation increase (up to 12% compared to control) at concentrations ranging from 0.5 µM to 50 µM, while in MC3T3-E1 cells, the highest observed proliferation increase was 138% [21]. It is worth noting that SAL also significantly increased the proliferation of other cells analogous to osteoblasts derived from different rodent species, such as rat bone marrow-derived mesenchymal stem cells (rBMSCs) [22,23,24].
3.1.2. Activation of Bone Morphogenetic Protein 2 Signaling Pathway
SAL was evaluated as a potential activator of bone morphogenetic protein 2 (BMP-2), a crucial factor in osteoblast differentiation and growth. BMP-2 activity was measured using a luciferase reporter gene assay. Various concentrations of SAL were added to cells for 48 hours, and luciferase activity was assessed using the Dual-Luciferase® Reporter Assay System. A clear increase in reporter gene expression was observed for SAL concentrations ranging from 0.25 µM to 10 µM [21].
3.1.3. Alkaline Phosphatase Activity and Mineralization
Alkaline phosphatase (ALP) is a marker of osteoblast differentiation. C3H10T1/2 cells treated with SAL (0.5–10 µM) for 12 and 18 days showed a significant increase in ALP activity, with the highest increase observed with 5 µM SAL at 18 days. Alizarin Red S staining demonstrated increased osteoblast mineralization after 21 days of SAL treatment [21]. Additionally, mRNA expression of osteoblast markers ALP and osteopontin, as well as osteogenic transcription factors like runt-related transcription factor 2 (Runx2) and osterix (OSX), were significantly elevated in C3H10T1/2 cells treated with 5 µM SAL for 18 days, as measured by reverse transcription-polymerase chain reaction (RT-PCR). Similar effects were observed in MC3T3-E1 cells treated with SAL (0.5 - 5 µM) for 12 days [21].
3.1.4. Bone Morphogenetic Protein 2, Bone Morphogenetic Protein 6, and Bone Morphogenetic Protein 7 Expression
To determine if SAL influences BMP mRNA expression during osteoblast differentiation, 5 µM SAL was administered to C3H10T1/2 cells for 18 days, resulting in increased expression of BMP-2, BMP-6, and BMP-7 [21]. In MC3T3-E1 cells, 1 µM SAL administered for 6 and 12 days significantly increased BMP-2 mRNA expression as detected by RT-PCR. SAL treatment in both cell lines also induced rapid phosphorylation of Smad1/5/8 and enhanced extracellular signal-regulated kinase 1/2 (ERK1/2) pathway activation, which is also confirmed by other in vitro studies [22]. Co-treatment with noggin, a BMP inhibitor, or dorsomorphin, an inhibitor of the type I BMP receptor, significantly reduced SAL-induced ALP activity and Smad1/5/8 phosphorylation [21].
3.1.5. Adenosine Monophosphate-Activated Protein Kinase Activation
Fu et al. explored the role of adenosine monophosphate-activated protein kinase (AMPK) in SAL-induced osteoblast proliferation and differentiation using MC3T3-E1 cells [25]. Cells were divided into six groups and cultured in osteogenic medium for 14 days. Three groups were treated with different concentrations of SAL (1 µM, 5 µM, 10 µM) for 48 hours. Another group was treated with 1 mM 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR), an AMPK activator, for 48 hours. A fifth group was treated with 10 µM SAL and 40 µM Compound C, an AMPK inhibitor. The control group was maintained in osteogenic medium without additional treatments. Cell proliferation was assessed using a Cell Counting Kit-8 (CCK8) assay and the optical density (OD) was measured. Results showed that SAL stimulated MC3T3-E1 cell proliferation in a dose- and time-dependent manner. After 24 hours, cells treated with 5 µM and 10 µM SAL showed significantly increased OD values compared to the control and 1 µM SAL groups. After 36 hours, all SAL-treated groups demonstrated significantly increased proliferation. The AICAR group also showed a significant increase in OD values at 24 and 36 hours. Co-treatment with Compound C and 10 µM SAL significantly reduced the pro-proliferative effect of SAL compared to 10 µM SAL alone [25]. ALP activity, a marker of osteoblast differentiation, was significantly increased in all SAL-treated groups compared to the control, with the highest increase observed in the 10 µM SAL group. The lowest increase was seen in the group treated with SAL and Compound C. Alizarin Red assay results indicated that SAL increased the number of calcium nodules, with the highest number in the AICAR group and the lowest in the SAL and Compound C group. Western blot analysis revealed that the expression of osteogenic proteins collagen type I alpha 1 (COL1A1), osteocalcin (OCN), and RUNX2 was significantly increased in the AICAR and SAL-treated groups, particularly in the 10 µM SAL group. However, the SAL and Compound C group showed a reduced effect. Furthermore, phosphorylated AMPK (p-AMPK) expression was significantly increased in the AICAR and SAL-treated groups compared to the control, whereas the SAL and Compound C group showed a decreased p-AMPK/AMPK ratio [25].
3.1.6. Cellular and Molecular Impact of Salidroside on Endothelial Cells
Guo et al. conducted an extensive study on the cellular and molecular impacts of SAL on vascular endothelial cells and the hypoxia-inducible factor 1-alpha (HIF-1α)/vascular endothelial growth factor (VEGF) signaling pathway, which plays a pivotal role in angiogenesis-osteogenesis coupling [25]. The primary objective of this research was to evaluate whether SAL could prevent bone loss and/or expedite the bone healing process. The HIF-1α pathway is essential for bone growth and blood vessel formation as it enhances the expression of VEGF in osteoblasts under hypoxic conditions. The researchers used the human endothelial cell line EA.hy926 and human umbilical vein endothelial cells (HUVEC) cultured with conditioned medium (CM) from osteoblasts (MG-63 cells) [26]. To assess the effects of SAL, the researchers performed a series of in vitro experiments. Initially, the 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay was employed to determine the viability and proliferation of EA.hy926 and HUVEC cells treated with CM and varying concentrations of SAL. The results revealed that CM supplemented with SAL significantly boosted cell proliferation and viability by up to 126% compared to the control group. Notably, there were no significant differences observed between the effects of 10 nM and 100 nM concentrations of SAL. These findings indicate that SAL-treated CM has the potential to markedly enhance the proliferation and viability of endothelial cells [26]. In another study, Guo et al. aimed to determine the cellular and molecular effects of salidroside on bone metabolism and the HIF-1α pathway in osteoblasts, thus potentially influencing fracture healing [27]. The research involved both in vitro and in vivo studies, with in vitro studies focusing on cellular models. The human osteoblast-like cell line MG-63 was purchased from ATCC, and rat calvarial osteoblast (ROB) cells were isolated enzymatically from the calvariae of newborn Sprague Dawley (SD) rats. These cells were treated with varying concentrations (1, 10, 100, or 1000 nM) of salidroside dissolved in Dimethyl Sulfoxide (DMSO) over 2, 4, and 6 days. Cell viability following salidroside treatment was assessed using MTS assay supplied by Shanghai Beibo Technologies, China. The findings from the in vitro studies demonstrated that salidroside significantly promoted cell proliferation by altering the cell cycle distribution of osteoblastic cells. Additionally, salidroside stimulated bone formation in osteoblasts by enhancing their differentiation and mineralization [27]. Specifically, salidroside significantly stimulated osteoblast differentiation and mineralization by inducing the expression of Runx2 and OSX. It was also observed that salidroside could activate the HIF-1α/VEGF signaling pathway, promoting both angiogenesis and osteogenesis through a non-autonomous mechanism. This was evidenced by a significant increase in HIF-1α expression at both mRNA and protein levels. Furthermore, salidroside promoted VEGF production by inducing HIF-1α expression and enhancing HIF-1α transactivation. The mechanism of action of SAL on the HIF-1α/VEGF signaling pathway is illustrated in Figure 2. It is additionally worth nothing that the mitogen-activated protein kinase (MAPK)/ERK and PI3K/Akt pathways were found to be essential for salidroside-mediated osteoblast proliferation and HIF-1α expression in MG-63 cells, indicating that salidroside exerts its effects on osteoblastic cells by triggering the activation of MAPK/ERK and PI3K/Akt pathways [27].
3.1.7. Proliferation and Viability Assays
The proliferation and viability of EA.hy926 and HUVEC cells were also analyzed using MTS assays [26]. The CM prepared from osteoblasts treated with SAL showed a substantial increase in the proliferation and viability of both EA.hy926 and HUVEC cells. The increase in cell viability reached up to 126%, indicating a significant enhancement compared to control cells not treated with SAL. The assays confirmed that CM with SAL significantly boosts the proliferation and viability of endothelial cells, a crucial factor for angiogenesis [26]. In these assays, the researchers observed no notable difference in the effects between the 10 nM and 100 nM concentrations of SAL, suggesting a plateau in the dose-response relationship at these concentrations. This implies that even at lower concentrations, SAL effectively promotes endothelial cell proliferation and viability, making it a potent agent for enhancing angiogenic processes [26].
3.1.8. Migration and Capillary Tube Formation
Cell migration is a vital process in angiogenesis where endothelial cells move to form new blood vessels. The researchers evaluated the impact of SAL on endothelial cell migration using the CM treated with different concentrations of SAL [26]. The results showed that CM alone could promote the migration of EA.hy926 and HUVEC cells. However, when SAL was added to the CM at concentrations of 10 nM and 100 nM, there was a significant increase in cell migration. Specifically, the migration of EA.hy926 cells increased by approximately 50%, while HUVEC cells showed an increase of about 30% compared to cells treated with CM alone [26]. To further understand the role of VEGF in this process, anti-VEGF antibodies were introduced to the culture. The addition of anti-VEGF antibodies significantly reversed the migration induced by CM with SAL, while the use of isotype control antibodies had no such effect. This reversal suggests that the angiogenic activity of SAL is heavily mediated by VEGF expression [26]. The study also examined the effect of SAL on capillary tube formation in vitro using HUVEC cells as a model. The ability of endothelial cells to form capillary-like structures is a hallmark of angiogenesis. The results indicated that both 10 nM and 100 nM concentrations of SAL in the CM significantly enhanced the formation of capillary tubes compared to CM alone. The length of the capillary tubes in the presence of SAL was 46% greater than in the CM-only group. Similar to the migration assays, the application of anti-VEGF antibodies significantly diminished the tube formation induced by SAL, confirming the critical role of VEGF in this process [26].
3.1.9. Expression of Hypoxia-Inducible Factor 1-Alpha and Vascular Endothelial Growth Factor
To delve deeper into the molecular mechanisms, the study of Guo et al. analyzed the expression levels of HIF-1α and VEGF in endothelial cells treated with SAL [26]. Western blot and RT-PCR analyses were employed to measure the mRNA and protein levels of these factors. The data revealed that SAL treatment significantly elevated the levels of HIF-1α and VEGF mRNA and proteins in EA.hy926 cells, with the most notable effects observed at the 10 nM concentration. This suggests that SAL can effectively enhance the expression of these key angiogenic factors at relatively low concentrations [26]. Further experiments were conducted to test the specificity of this response using 3-(5'-hydroxymethyl-2'-furyl)-1-benzylindazole (YC-1), a known inhibitor of HIF-1α. EA.hy926 cells were pre-treated with YC-1 before being exposed to SAL. The results demonstrated that YC-1 significantly inhibited the SAL-induced expression of HIF-1α and VEGF. However, SAL was able to partially counteract the inhibitory effects of YC-1, indicating that SAL mitigates the suppression of HIF-1α and VEGF expression [24]. Additionally, luciferase reporter assays were conducted to evaluate the transcriptional activity of HIF-1α in cells treated with various concentrations of SAL. The assays showed that SAL at 10 nM substantially increased HIF-1α transcriptional activity under hypoxic conditions, further supporting the role of SAL in promoting angiogenesis through the activation of the HIF-1α/VEGF signaling pathway [26].
3.1.10. Effects of Salidroside on Glucocorticoid-Induced Osteoporosis
Xie et al. investigated the protective effects of SAL on glucocorticoid-induced OP, specifically focusing on dexamethasone-induced bone loss [28]. Dexamethasone, a synthetic glucocorticoid, is known for its anti-inflammatory and immunosuppressive properties, but it also inhibits osteoblast activity, reduces osteoprotegerin (OPG) production, decreases collagen synthesis, and impairs calcium absorption. These effects make glucocorticoid-induced OP the third most common type of OP after postmenopausal and senile OP [28]. The researchers utilized the MC3T3-E1 cell line, consisting of pre-osteoblasts isolated from the skulls of Mus musculus. The toxicity of SAL in these cells was assessed by incubating them with various concentrations of SAL (10, 20, 40, 60, 80, and 100 μM) for 24 hours. The impact of SAL on cell viability in the presence of dexamethasone was also evaluated. For the experiments, 5 x 104 MC3T3-E1 cells were seeded and treated with osteogenic induction medium (OIM) containing 50 μg/mL ascorbic acid, 10 mM β-glycerophosphate, and 0.1 μM dexamethasone. The cells were then treated with SAL and OIM once they reached 80% confluency for seven days. It was found that MC3T3-E1 cells exposed to SAL (1 to 100 μM) for three days did not show a reduction in viability. However, SAL concentrations of 10 to 100 μM did not significantly alleviate the reduction in cell viability caused by dexamethasone [28]. In another set of experiments in this study, MC3T3-E1 cells were cultured in OIM and exposed to dexamethasone (10 μM) and SAL (10 and 20 μM) for three weeks. Alizarin Red S staining, which indicates mineralized nodules, showed that dexamethasone significantly inhibited mineralization. However, this inhibitory effect was markedly reduced by the addition of SAL. Furthermore, SAL activated the transforming growth factor-beta (TGF-β)/Smad2/3 signaling pathway, essential for maintaining tissue homeostasis and regulating cellular responses to tissue injury and repair. SAL also mitigated the dexamethasone-induced suppression of protein expression levels of OSX, a transcription factor crucial for the differentiation of pre-osteoblasts into mature osteoblasts, and significantly enhanced ALP activity and mineralization, promoting osteogenic differentiation. The protective effects of SAL against dexamethasone-induced inhibition of osteogenic differentiation could be blocked by LY2109761, an inhibitor of TGF-β type I/II receptors [28]. Xue et al. conducted studies to investigate the effects of SAL on steroid-induced avascular necrosis of the femoral head (SANFH) using the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) signaling pathway [29]. These studies included both in vitro and in vivo experiments. In the in vitro experiments, primary rat osteoblasts were treated with SAL and dexamethasone to induce osteoblast apoptosis. The results demonstrated that SAL at concentrations of 10 nM and 100 nM significantly protected osteoblasts from dexamethasone-induced cell death and apoptosis. This protective effect was confirmed through TUNEL staining, which detects apoptotic cells, and Western blot analyses, which showed decreased levels of apoptotic markers such as Bcl-2 Associated X-protein (BAX), caspase-3 (CASP3), and caspase-9 (CASP9). These findings suggest that SAL mitigates mitochondrial apoptosis pathways, thereby protecting osteoblasts [29].
3.1.11. Salidroside's Role in Mitigating Osteoporosis Through Antioxidant Activity
Wang et al. aimed to demonstrate the protective effects of SAL against OP by inhibiting oxidative stress and promoting osteogenesis in a mouse model of ovariectomy(OVX)-induced estrogen deficiency [30]. OP is characterized by low bone mass, deterioration of bone microarchitecture, and increased bone fragility. Contributing factors include aging, reduced levels of estrogen, vitamin D, and calcium, as well as elevated levels of reactive oxygen species (ROS). Evidence suggests that the balance between ROS and antioxidants plays a significant role in the development of OP [30]. In vitro, the researchers used a tert-butyl hydroperoxide (t-BHP)-induced oxidative stress model in rat osteoblasts (ROBs) to assess the protective capabilities of SAL against cell apoptosis, osteogenic differentiation, antioxidant capacity, and nuclear factor erythroid 2-related factor 2 (Nrf2) expression. ROBs were isolated from the skulls of 4-week-old rats and cultured. The cells were exposed to various concentrations of t-BHP (50, 62.5, 75, 87.5, 100 μM) for six hours, and different concentrations of SAL (0.01, 0.1, 1, 10, 100 μM) were administered for 24 or 48 hours. Following pretreatment with SAL, the medium was replaced with t-BHP for six hours. Cell viability was assessed using the CCK-8 assay, and flow cytometry and Western blotting were used to analyze apoptosis and protein expression. The results showed that SAL pretreatment significantly protected ROBs from t-BHP-induced oxidative damage and apoptosis. SAL promoted osteogenic differentiation, enhanced antioxidant capacity, and increased Nrf2 expression while reducing Kelch-like ECH-associated protein 1 (Keap1) expression, which is an inhibitor of Nrf2. These findings indicate that SAL exerts its protective effects by activating the Nrf2 pathway and mitigating oxidative stress [30].
3.2. In Vivo Studies
3.2.1. Protective Effects Against Oxidative Stress
SAL's potential to protect against oxidative stress-induced bone loss was assessed using hydrogen peroxide (H2O2)-induced oxidative stress in MC3T3-E1 cells and an OVX-induced OP mouse model [31]. H2O2 significantly reduced cell viability, with a 300 µM concentration causing approximately 50% viability reduction after 24 hours. Pre-treatment with SAL (0.1 - 10 µM) for 24 hours significantly improved cell survival against subsequent H2O2 exposure. SAL also protected osteoblast function, as evidenced by increased ALP activity and enhanced expression of differentiation markers ALP, COL1A1, and OCN [29], which has also been observed in other in vivo studies [22,23,24]. Micro-computed tomography (micro-CT) scans of the femoral distal metaphysis and lumbar vertebrae from treated mice showed that SAL (20 mg/kg) significantly preserved bone microstructure parameters such as bone volume/total volume (BV/TV%), trabecular number (Tb.N), trabecular separation (Tb.Sp), trabecular thickness (Tb.Th), connectivity density (Conn.D), structure model index (SMI), and BMD. Additionally, SAL treatment increased glutathione (GSH) levels and decreased malondialdehyde (MDA) levels in the blood, indicating reduced oxidative stress. Histological analyses confirmed that SAL treatment mitigated bone trabecular thinning and increased calcium deposition [31].
3.2.2. Effects on Knee Osteoarthritis in Mice
Further in vivo studies were conducted on a knee osteoarthritis (KOA) model in mice [25]. Fifty 6-week-old male mice were divided into five groups: normal, KOA, SAL, AICAR, and SAL with Compound C. KOA was induced by intra-articular injection of monoiodoacetate. After three weeks, treatments commenced with SAL (50 mg/kg/day) administered intragastrically, AICAR (100 mg/kg/day), or SAL (50 mg/kg/day) with Compound C (20 mg/kg/day) administered intraperitoneally. The normal and KOA groups did not receive treatment. Mice were weighed weekly, and arthritis index (AI) was assessed. Body weight increased across all groups, with the highest increase in the normal group and the lowest in the KOA group. AI significantly increased in the KOA model groups compared to the normal group. SAL and AICAR treatments reduced AI after 21 days, while the combination of SAL and Compound C resulted in higher AI values [25]. Enzyme-linked immunosorbent assays (ELISA) on plasma samples showed elevated levels of inflammatory markers tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6), and reduced interleukin-10 (IL-10) in the KOA group compared to the normal group. SAL and AICAR treatments significantly decreased TNF-α and IL-6 levels and increased IL-10 levels. The combination of SAL and Compound C nullified these anti-inflammatory effects [25]. Western blot analysis of knee joint tissues revealed increased expression of osteogenic proteins RUNX2, ALP, COL1A1, and OCN in the SAL and AICAR groups compared to the KOA group. The combination of SAL and Compound C reduced this effect. Additionally, p-AMPK expression was higher in the SAL and AICAR groups compared to the KOA group, but lower in the SAL and Compound C group [25]. Histological analysis showed preserved joint structure with minimal inflammatory cell infiltration in the normal group. The KOA group exhibited bone erosion and cartilage degradation, while SAL and AICAR treatments reduced inflammatory cell presence and preserved joint structure. Tartrate-resistant acid phosphatase (TRAP) staining indicated higher osteoclast activity in the KOA group, which was reduced in the SAL and AICAR groups. The combination of SAL and Compound C increased TRAP expression. Nuclear factor of activated T cells, cytoplasmic 1 (NFATc1) expression related to osteoclast activity was also reduced by SAL and AICAR treatments but increased with the combination of SAL and Compound C [25].
3.2.3. Angiogenesis in Mouse Embryonic Metatarsals
In another in vivo study, mouse embryonic metatarsal bones were isolated to evaluate endothelial sprouting, a crucial aspect of angiogenesis [26]. The experimental groups included control, SAL, VEGF, SAL + anti-VEGF, and SAL + isotype control immunoglobulin G (IgG). SAL treatment significantly increased the area of endothelial sprouting compared to the control group. The inclusion of anti-VEGF antibodies markedly inhibited this sprouting, highlighting the critical role of VEGF in mediating the pro-angiogenic effects of SAL. This experiment provided compelling evidence that SAL promotes angiogenesis through VEGF-dependent pathways in an in vivo context [26].
3.2.4. Osteogenesis and Bone Healing
The in vivo aspect of Xue et al. study involved adult male rats to evaluate the protective effects of SAL against SANFH [29]. The rats were divided into control, model, and SAL treatment groups. The model and SAL groups received lipopolysaccharide (LPS) followed by methylprednisolone (MPS) to induce SANFH, while the SAL group additionally received SAL injections. Histological analysis of the femoral heads showed that the SAL-treated group had significantly fewer empty lacunae compared to the model group, indicating a protective effect of SAL against bone necrosis [29]. For another in vivo study, BALB/c mice were used, sourced from the Experimental Animal Center of the Academy of Military Medical Sciences in Beijing, China [26]. An isolated fracture was induced in the proximal third of the tibia of these mice, and they were randomly divided into four groups. Seven days post-surgery, the groups were administered different treatments: a vehicle (fracture model group, FM), 200 μM salidroside (low-dose salidroside group, SAL-L), 800 μM salidroside (high-dose salidroside group, SAL-H), and 200 μM desferrioxamine (DFO group). These treatments were administered to the fracture sites every other day. Seventeen days after the procedure, radiographic and histological analyses were conducted to evaluate the fracture healing status. The results indicated that salidroside significantly enhanced fracture healing. This enhancement was evidenced by improved bone formation and mineralization, increased angiogenesis within the callus, and accelerated overall fracture healing [26].
3.2.5. Effects on Osteoporosis Model in Rats
Ling Li et al. utilized an OVX-induced OP model in rats to study the protective effects of SAL on bone loss [32]. This model mimics postmenopausal OP in humans. The rats were divided into five groups: control, OVX, OVX + low-dose SAL (4 mg/kg), OVX + high-dose SAL (20 mg/kg), and OVX + raloxifene (RLX), a selective estrogen receptor modulator known to protect against OP. The results showed that SAL significantly increased bone mass and mineral apposition rates (MAR), which represent the rate of new bone formation. Serum levels of ALP, OCN, and prostaglandin E2 (PGE2) were also measured. The OVX group exhibited significantly elevated levels of ALP and OCN, indicative of increased bone turnover. However, treatment with high-dose SAL or RLX significantly reduced these levels, suggesting that SAL mitigates the adverse effects of estrogen deficiency on bone turnover [32]. Micro-CT analysis further revealed that SAL improved the microarchitecture of trabecular bone. Parameters such as BMD, trabecular area (Tb.Ar), Tb.Th, Tb.N, and Tb.Sp were all significantly improved in the SAL-treated groups compared to the OVX group. The histological examination confirmed these findings, showing that SAL partially reversed the negative impact of OVX on bone microarchitecture [30], which is also supported by other in vivo studies [22,23,24]. Additionally, the effects of SAL on osteoid parameters, including osteoid surface (OS), osteoid width (O.Wi), and osteoid volume (OV), were assessed. The OVX group showed increased osteoid parameters, indicative of impaired bone formation, while SAL treatment significantly reduced these parameters, further supporting its protective role [32]. Immunohistochemical analysis was used to assess the expression of HIF-1α and VEGF in bone tissue. The results showed increased expression of HIF-1α and VEGF in the SAL-treated groups, suggesting that SAL enhances bone healing and regeneration through the HIF-1α/VEGF pathway [32]. Another study conducted by Zheng et al. on rats provided detailed insights into the significant impact of SAL on the OPG/the receptor activator of nuclear factor kappa-Β ligand (RANKL) ratio [33]. The study used female Sprague-Dawley rats, which were OVX to induce a postmenopausal OP model and subsequently rendered diabetic through the administration of streptozotocin (STZ). SAL was administered orally at doses of 20, 40, and 80 mg/kg per day for a period of 12 weeks. The treatment significantly upregulated the expression of OPG while simultaneously downregulating RANKL across all dosage groups, with the most pronounced effects observed at the highest dose of 80 mg/kg. Corresponding to these changes, there was a significant increase in BMD in the SAL-treated groups compared to the control group, alongside notable improvements in overall bone health. These results underscore the potential of SAL as a therapeutic agent in preventing bone loss through the modulation of the OPG/RANKL signaling pathway, particularly in diabetic conditions exacerbated by estrogen deficiency [33]. The protective effects of SAL on OVX-induced bone loss were examined in 3-month-old female rats also [31]. Four weeks post-OVX, the rats were divided into groups receiving vehicle treatment, 4 mg/kg/day SAL, or 20 mg/kg/day SAL for 90 days via intragastric injection. Post-treatment histomorphometry of the left tibia revealed that SAL significantly mitigated trabecular bone loss, with higher doses providing better protection. SAL treatment also increased trabecular bone volume percentage (TBV%) and reduced trabecular absorption surface percentage (TRS%), indicative of decreased osteoclast activity [31].
4. Discussion
The research on SAL presented in this review paper elucidates its multifaceted role in promoting bone health through osteogenesis and angiogenesis. This comprehensive analysis of both in vitro (Table 1) and in vivo (Table 2) studies highlights SAL’s potential as a therapeutic agent in the treatment of OP and the enhancement of fracture healing. SAL has been shown to stimulate the proliferation and differentiation of osteoblast precursors, as evidenced by the increased activity of ALP and mineralization markers in studies conducted by Chen et al. [21]. This is achieved primarily through the activation of the BMP-2 signaling pathway, which is critical for osteoblast differentiation and bone growth. Furthermore, the upregulation of key osteogenic transcription factors such as Runx2 and OSX under SAL treatment reinforces its role in enhancing osteoblast activity and bone formation. These findings can be particularly useful in the field of orthopedics and bone regenerative medicine. Fu et al. demonstrated that SAL activates AMPK, a key regulator of cellular energy homeostasis, to promote osteoblast proliferation and differentiation while inhibiting bone resorption [25]. This activation of AMPK is crucial, as it not only supports osteoblast function but also mitigates the effects of bone resorptive conditions such as osteoarthritis. This discovery has significant implications for the treatment of metabolic bone diseases and inflammatory conditions affecting the joints, such as rheumatoid arthritis and osteoarthritis. SAL’s role in endothelial cell function and angiogenesis, as investigated by Guo et al., further underscores its therapeutic potential. By enhancing the HIF-1α and VEGF signaling pathways, SAL promotes angiogenesis, which is essential for bone healing and regeneration [26]. The coupling of osteogenesis and angiogenesis facilitated by SAL ensures adequate vascular support for newly forming bone tissues, enhancing the overall efficacy of bone repair processes. These insights are valuable for developing advanced therapies in vascularized tissue engineering and regenerative medicine, particularly for conditions such as peripheral artery disease and diabetic foot ulcers. Xie et al. explored the protective effects of SAL against glucocorticoid-induced OP [28]. Their findings revealed that SAL mitigates the inhibitory effects of dexamethasone on osteoblast viability and differentiation by activating the TGF-β/Smad2/3 signaling pathway [28]. This pathway is crucial for maintaining tissue homeostasis and promoting the differentiation of pre-osteoblasts into mature osteoblasts, highlighting SAL’s potential in counteracting the adverse effects of glucocorticoids on bone health. This research is particularly relevant for endocrinology and pharmacology, where managing side effects of long-term steroid use is critical, especially in conditions like Cushing's syndrome and chronic inflammatory diseases. Xue et al. provided evidence that SAL protects against SANFH through the PI3K/Akt signaling pathway [29]. SAL’s ability to prevent osteoblast apoptosis and promote cell survival under dexamethasone treatment indicates its potential to mitigate steroid-induced bone damage [29]. These findings have significant applications in sports medicine and orthopedic surgery, particularly in preventing and treating bone necrosis often seen in patients receiving high-dose corticosteroid therapy. The antioxidant properties of SAL, as detailed by Wang et al., play a significant role in protecting against OP by inhibiting oxidative stress and promoting osteogenesis through the Nrf2 pathway [30]. The activation of Nrf2 enhances the cells' antioxidant capacity, reducing oxidative damage and promoting bone health, especially in conditions of estrogen deficiency, as modeled in OVX rats. This aspect of SAL is crucial for geriatric medicine and preventive healthcare, focusing on mitigating age-related bone density loss and conditions such as postmenopausal OP. The consolidated graphical summary illustrating the impact of salidroside on the signaling pathways, as mentioned in the cited studies, can be found in Figure 3. In vivo studies further validate SAL’s efficacy in improving bone density, preserving bone microarchitecture, and enhancing bone formation [22,23,24]. For instance, the studies by Li et al. demonstrated that SAL significantly increased bone mass and mineral apposition rates in an OVX-induced OP model, indicating its potential as a therapeutic agent for postmenopausal OP [32]. Additionally, SAL’s ability to enhance angiogenesis and accelerate fracture healing through both cell-autonomous and non-autonomous mechanisms was evidenced in mouse models, supporting its application in clinical settings for improving skeletal regeneration and repair [27]. These findings are highly relevant for orthopedic surgery and rehabilitation medicine, particularly in the context of fracture healing and recovery from bone surgeries. The therapeutic potential of SAL in bone health and regenerative medicine is promising, warranting further investigation and potential clinical applications. Its multifaceted mechanisms of action, including osteogenic, angiogenic, anti-inflammatory, and antioxidant properties, position SAL as a valuable compound in the prevention and treatment of various bone-related conditions. The therapeutic potential of SAL extends to its anti-inflammatory properties in conditions like KOA, as shown by its ability to reduce inflammatory markers and improve joint structure in animal models [25]. Osteoarthritis is often associated with the formation of subchondral cysts, which can weaken the structural integrity of bones and increase the risk of fractures. This highlights the broader applicability of SAL in managing inflammatory bone diseases and potentially mitigating fracture risks associated with KOA. This application is pertinent to rheumatology and pain management specialties, addressing common issues such as joint degeneration and chronic pain. Despite the extensive research on SAL, none of the studies to date have considered its combined effects with rosavin, another bioactive compound that naturally occurs with SAL in R. rosea. Rosavin has been demonstrated to possess osteogenic and anti-osteoporotic properties [34,35]. Investigating the synergistic effects of SAL and rosavin could provide a more comprehensive understanding of their potential in bone health. This unexplored combination may reveal enhanced or complementary mechanisms of action, potentially leading to more effective therapeutic strategies for OP and other bone-related disorders. Future studies should aim to elucidate the interactions between these compounds to fully leverage the therapeutic potential of R. rosea [32,33]. SAL has been used for years in the form of extracts from R. rosea to treat a variety of conditions, including fatigue, depression, anxiety, and cardiovascular disorders. Its potential osteoprotective effects and the convenience of oral administration present a significant advantage. However, studies on its bioavailability in the human gastrointestinal tract are essential. Research should focus on determining how effectively SAL is absorbed and utilized when taken orally. If oral bioavailability proves inadequate, alternative delivery methods, such as transdermal or intravenous administration, may need to be explored to ensure its therapeutic efficacy in bone-related conditions. This is particularly important for pharmacology and drug delivery systems [36,37]. Future research should focus on clinical trials to evaluate the efficacy and safety of SAL in human subjects, particularly in the treatment of OP, glucocorticoid-induced bone loss, and fracture healing. It is essential to conduct studies on human bone tissue cells under both in vitro conditions and in vivo using tissues obtained from patients after orthopedic procedures. Additionally, further elucidation of the signaling pathways involved in SAL’s action will provide deeper insights into its mechanisms and potential combinatory effects with other therapeutic agents.
5. Conclusions
SAL presents a promising therapeutic avenue for enhancing bone health and treating various bone-related disorders. Its ability to promote osteogenesis and angiogenesis, coupled with its protective effects against oxidative stress and inflammation, positions it as a valuable candidate for future clinical applications in bone regeneration and OP treatment.
Author Contributions
Conceptualization, P.W.; Methodology, P.W., E.C., S.B., P.T., Ł.A.P., W.Ł., M.O., E.U.S., A.L-G. and B.Ż.; Software, P.W., P.T., M.O. and D.S.; Validation, P.W., Ł.A.P., E.W., A.M., E.U.S. and D.S.; Formal Analysis, P.W., E.C., S.B., P.T., W.Ł., B.Ż., K.R.-P. and D.S.; Investigation, P.W., E.C., S.B., P.T., W.Ł., M.O., E.U.S. and D.S.; Resources, P.W., E.W., A.M., M.O. and B.Ż.; Data Curation, P.W., P.T., E.W., A.M., A.L-G., B.Ż. and D.S.; Writing – Original Draft Preparation, P.W., E.C., S.B., P.T., Ł.A.P. and W.Ł.; Writing – Review & Editing, P.W., P.T., Ł.A.P., E.W., A.M., K.R.-P. and D.S.; Visualization, P.W., P.T., Ł.A.P. and A.L-G.; Supervision, P.W., Ł.A.P., W.Ł., E.U.S., B.Ż., K.R.-P. and D.S.; Project Administration, P.W. and D.S.; Funding Acquisition, P.W., E.C., S.B., W.Ł., K.R.-P. and D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
For this type of study formal consent is not required.
Data Availability Statement
Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Panossian, A.; Wikman, G. Effects of Adaptogens on the Central Nervous System and the Molecular Mechanisms Associated with Their Stress-Protective Activity. Pharmaceuticals 2010, 3, 188–224. [Google Scholar] [CrossRef] [PubMed]
- Darbinyan, V.; Aslanyan, G.; Amroyan, E.; Gabrielyan, E.; Malmström, C.; Panossian, A. Clinical trial of Rhodiola rosea L. extract SHR-5 in the treatment of mild to moderate depression. Nordic Journal of Psychiatry 2000, 61, 343–348. [Google Scholar] [CrossRef]
- Olsson, E.M.; von Schéele, B.; Panossian, A.G. A randomised double-blind placebo-controlled parallel-group study of the standardised extract SHR-5 of the roots of Rhodiola rosea in the treatment of subjects with stress-related fatigue. Planta Medica 2009, 75, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Kelly, G.S. Rhodiola rosea: A possible plant adaptogen. Alternative Medicine Review 2001, 6, 293–302. [Google Scholar] [PubMed]
- Brown, R.P.; Gerbarg, P.L.; Ramazanov, Z. Rhodiola rosea: A phytomedicinal overview. HerbalGram 2002, 56, 40–52. [Google Scholar]
- Ishaque, S.; Shamseer, L.; Bukutu, C.; Vohra, S. Rhodiola rosea for physical and mental fatigue: A systematic review. BMC Complementary and Alternative Medicine 2012, 12, 70. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Li, Q.; Yao, L.; Jiang, Z.; Jin, G.; Zhao, X. Salidroside: Advances in pharmacological activities and molecular mechanisms of action. Journal of Ethnopharmacology 2020, 261, 113112. [Google Scholar] [CrossRef]
- Chan, Y.S.; Cheng, L.N.; Wu, J.H.; Chan, E.; Kwan, Y.W.; Lee, S.M.; Leung, G.P.; Yue, P.Y.; Wong, R.N. A review of the pharmacological effects of Rhodiola rosea L. and its bioactive compounds. Phytotherapy Research 2012, 26, 1727–1734. [Google Scholar] [CrossRef]
- Zhang, J.; Zeng, Y.; Li, Y.; Lu, D.; Wang, X.; Ji, X. Salidroside regulates cardiac dysfunction in heart failure by enhancing mitochondrial biogenesis and inhibiting oxidative stress. Biomedicine & Pharmacotherapy 2018, 105, 119–128. [Google Scholar] [CrossRef]
- Li, Y.; Pham, V.; Bui, M.; Song, L.; Wu, C.; Walia, A.; Uchio, E.; Smith-Liu, F.; Zi, X. Rhodiola rosea: An herb with anti-stress, anti-aging, and anti-cancer properties. Cancer Cell International 2019, 19, 202. [Google Scholar]
- Hernlund, E.; Svedbom, A.; Ivergård, M.; Compston, J.; Cooper, C.; Stenmark, J.; McCloskey, E.V.; Jönsson, B.; Kanis, J.A. Osteoporosis in the European Union: medical management, epidemiology, and economic burden. Archives of Osteoporosis 2013, 8, 136. [Google Scholar] [CrossRef] [PubMed]
- Kanis, J.A.; Norton, N.; Harvey, N.C.; Jacobson, T.; Johansson, H.; Lorentzon, M.; Liu, E.; Vandenput, L.; McCloskey, E.V. SCOPE 2021: a new scorecard for osteoporosis in Europe. Archives of Osteoporosis 2022, 17, 104. [Google Scholar] [CrossRef] [PubMed]
- Pisani, P.; Renna, M.D.; Conversano, F.; Casciaro, E.; Di Paola, M.; Quarta, E.; Muratore, M.; Casciaro, S. Major osteoporotic fragility fractures: Risk factor updates and societal impact. World Journal of Orthopedics 2016, 7, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Huang, K.; Xu, H. Protective effects of Rhodiola rosea on oxidative stress and liver injury in rats. Molecular and Cellular Biochemistry 2009, 331, 31–41. [Google Scholar]
- Fan, X.J.; Wang, Y.; Wang, L.; Zhu, M. Salidroside induces apoptosis and autophagy in human colorectal cancer cells through inhibition of PI3K/Akt/mTOR pathway. Oncology Reports 2016, 6, 3559–3567. [Google Scholar] [CrossRef] [PubMed]
- György, Z.; Hohtola, A. Production of cinnamyl glycosides in compact callus aggregate cultures of Rhodiola rosea through biotransformation of cinnamyl alcohol. In Protocols for in vitro cultures and secondary metabolite analysis of aromatic and medicinal plants; Jain, S.M., Saxena, P.K., Eds.; Humana Press: Totowa, NJ, USA, 2009; pp. 305–312. [Google Scholar]
- Panossian, A.; Wikman, G.; Sarris, J. Rosenroot (Rhodiola rosea): Traditional use, chemical composition, pharmacology, and clinical efficacy. Phytomedicine 2010, 17, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wang, Y.; Guo, H.; Li, W.; Yan, S.; Zhao, S. Phytochemical analysis of traditional Chinese medicine using liquid chromatography coupled with mass spectrometry. Journal of Chromatography A 2015, 1428, 155–165. [Google Scholar] [CrossRef]
- Grech-Baran, M.; Sowa-Kasprzak, K. Salidroside as an adaptogen compound. Natural Product Communications 2013, 8, 1435–1440. [Google Scholar] [CrossRef]
- Panossian, A.; Wikman, G. Pharmacology of Schisandra chinensis Bail. : An overview of Russian research and uses in medicine. Journal of Ethnopharmacology 2009, 118, 183–212. [Google Scholar] [CrossRef]
- Chen, J.J.; Zhang, N.F.; Mao, G.X.; He, X.B.; Zhan, Y.C.; Deng, H.B.; Wang, Z. Salidroside stimulates osteoblast differentiation through BMP signaling pathway. Food and Chemical Toxicology 2013, 62, 499–505. [Google Scholar] [CrossRef]
- Pan, X.; Peng, X.; Jiang, C.; Yu, H.; Yang, Y.; Chen, H. Salidroside promotes the osteogenic differentiation of rat bone marrow mesenchymal stem cells through the BMP/Smad pathway. Drug Design Development and Therapy 2019, 13, 2501–2511. [Google Scholar]
- Li, J.; Guo, W. The effects of salidroside on bone marrow mesenchymal stem cells and their application in the treatment of osteoporosis. Life Sciences 2018, 207, 333–340. [Google Scholar] [CrossRef]
- Pan, X.; Liu, H.; Liu, J.; Shu, B.; Sun, L. Salidroside promotes osteogenic differentiation and bone formation in primary rat bone marrow stromal cells. Experimental and Therapeutic Medicine 2013, 5, 1265–1273. [Google Scholar] [CrossRef]
- Fu, S.; Yan, M.; Fan, Q.; Xu, J. Salidroside promotes osteoblast proliferation and differentiation via the activation of AMPK to inhibit bone resorption in knee osteoarthritis mice. Tissue Cell 2022, 79, 101917. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Yang, J.; Chen, Y.; Jin, X.; Li, Z.; Wen, X.; Xia, Q.; Wang, Y. Salidroside improves angiogenesis-osteogenesis coupling by regulating the HIF-1α/VEGF signalling pathway in the bone environment. European Journal of Pharmacology 2020, 884, 173394. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.Q.; Qi, L.; Yang, J.; Wang, Y.; Wang, C.; Li, Z.M.; Li, L.; Qu, Y.; Wang, D.; Han, Z.M. Salidroside accelerates fracture healing through cell-autonomous and non-autonomous effects on osteoblasts. Cell Tissue Research 2017, 367, 197–211. [Google Scholar] [CrossRef]
- Xie, B.; Zhou, H.; Liu, H.; Liao, S.; Zhou, C.; Xu, D. Salidroside alleviates dexamethasone-induced inhibition of bone formation via transforming growth factor-beta/Smad2/3 signaling pathway. Phytotherapy Research 2023, 37, 1938–1950. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Feng, Z.; Li, Z.; Pan, X. Salidroside inhibits steroid-induced avascular necrosis of the femoral head via the PI3K/Akt signaling pathway: In vitro and in vivo studies. Molecular Medicine Reports 2018, 17, 3751–3757. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.F.; Chang, Y.Y.; Zhang, X.M.; Gao, M.T.; Zhang, Q.L.; Li, X.; Zhang, L.; Yao, W.F. Salidroside protects against osteoporosis in ovariectomized rats by inhibiting oxidative stress and promoting osteogenesis via Nrf2 activation. Phytomedicine 2022, 99, 154020. [Google Scholar] [CrossRef]
- Zhang, J.K.; Yang, L.; Meng, G.L.; Yuan, Z.; Fan, J.; Li, D.; Liu, J. Protection by salidroside against bone loss via inhibition of oxidative stress and bone-resorbing mediators. PloS One 2013, 8, e57251. [Google Scholar] [CrossRef]
- Li, L.; Qu, Y.; Jin, X.; Guo, X.Q.; Wang, Y.; Qi, L.; Yang, J.; Zhang, P.; Li, L.Z. Protective effect of salidroside against bone loss via hypoxia-inducible factor-1α pathway-induced angiogenesis. Scientific Reports 2016, 6, 32131. [Google Scholar] [CrossRef]
- Zheng, H.; Qi, S.; Chen, C. Salidroside Improves Bone Histomorphology and Prevents Bone Loss in Ovariectomized Diabetic Rats by Upregulating the OPG/RANKL Ratio. Molecules 2018, 23, 2398. [Google Scholar] [CrossRef]
- Wang, S.; Feng, Y.; Zheng, L.; He, P.; Tan, J.; Cai, J.; Wu, M.; Ye, X. Rosavin: Research Advances in Extraction and Synthesis, Pharmacological Activities and Therapeutic Effects on Diseases of the Characteristic Active Ingredients of Rhodiola rosea L. Molecules 2023, 28, 7412. [Google Scholar] [CrossRef] [PubMed]
- Wojdasiewicz, P.; Turczyn, P.; Lach-Gruba, A.; Poniatowski, Ł.A.; Purrahman, D.; Mahmoudian-Sani, M.-R.; Szukiewicz, D. The Role of Rosavin in the Pathophysiology of Bone Metabolism. Int. J. Mol. Sci. 2024, 25, 2117. [Google Scholar] [CrossRef] [PubMed]
- Jamróz, W.; Arafat, B.; Gultekin, H.E. 3D-printed dosage forms for oral administration: a review. Drug Delivery and Translational Research 2023, 13, 210–223. [Google Scholar]
- Pan, X.; Liu, H.; Liu, J.; Shu, B.; Sun, L. Advances in Oral Drug Delivery Systems: Challenges and Opportunities. Pharmaceutics 2023, 15, 484. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The schematic representation of salidroside (C14H20O7) structure. The 2D and 3D structures were accustomed according to the PubChem database (PubChem CID: 159278).
Figure 1.
The schematic representation of salidroside (C14H20O7) structure. The 2D and 3D structures were accustomed according to the PubChem database (PubChem CID: 159278).
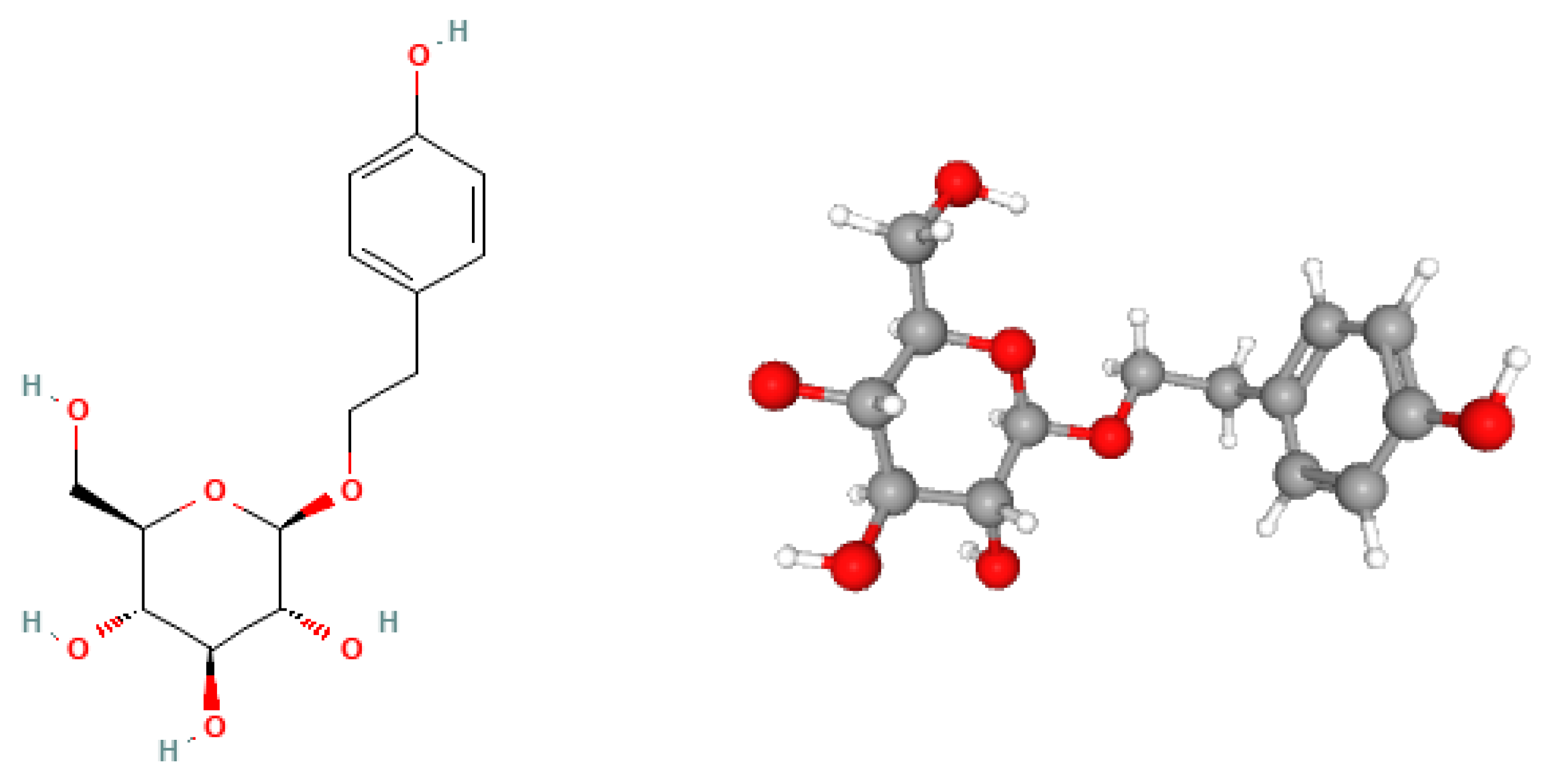
Figure 2.
The schematic illustration of the impact of salidroside (SAL) and hypoxia on the hypoxia-inducible factor 1-alpha (HIF-1α)/vascular endothelial growth factor (VEGF) signaling pathway. SAL and hypoxia inhibit the degradation of HIF-1α (red designation), leading to the accumulation of HIF-1α (green designation) in the cytoplasm. Subsequently, HIF-1α translocates to the nucleus, where it dimerizes with hypoxia-inducible factor 1-beta (HIF-1β) and binds to the hypoxia-responsive element (HRE). This interaction initiates the transcription of the VEGF gene, which in turn stimulates the synthesis of VEGF in the cytoplasm. The newly synthesized VEGF is then secreted out of the cell, where it binds to VEGF receptors (VEGF-R) on the surface of endothelial cells, stimulating their proliferation and leading to the formation of new blood vessels. It is noteworthy that SAL promotes angiogenesis in a manner similar to hypoxia, but potentially offers a more favorable anabolic mechanism, as it does not induce biological stress in the cells.
Figure 2.
The schematic illustration of the impact of salidroside (SAL) and hypoxia on the hypoxia-inducible factor 1-alpha (HIF-1α)/vascular endothelial growth factor (VEGF) signaling pathway. SAL and hypoxia inhibit the degradation of HIF-1α (red designation), leading to the accumulation of HIF-1α (green designation) in the cytoplasm. Subsequently, HIF-1α translocates to the nucleus, where it dimerizes with hypoxia-inducible factor 1-beta (HIF-1β) and binds to the hypoxia-responsive element (HRE). This interaction initiates the transcription of the VEGF gene, which in turn stimulates the synthesis of VEGF in the cytoplasm. The newly synthesized VEGF is then secreted out of the cell, where it binds to VEGF receptors (VEGF-R) on the surface of endothelial cells, stimulating their proliferation and leading to the formation of new blood vessels. It is noteworthy that SAL promotes angiogenesis in a manner similar to hypoxia, but potentially offers a more favorable anabolic mechanism, as it does not induce biological stress in the cells.
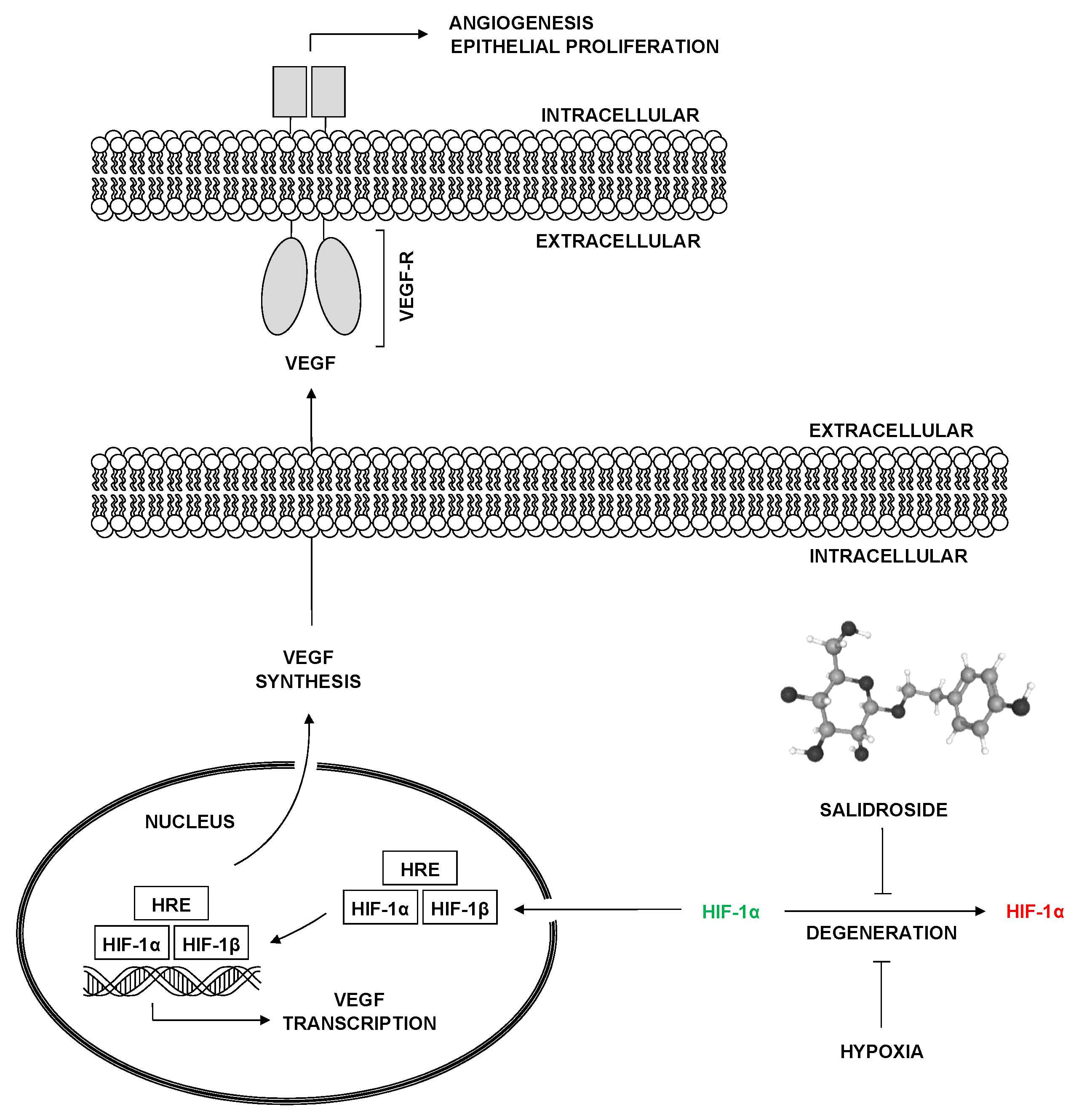
Figure 3.
Diagram summarizing the effects of salidroside (SAL) on cellular signaling pathways. Stimulatory effects are highlighted in green, whereas inhibitory effect is marked in red. Nrf2 - nuclear factor erythroid 2–related factor 2, BMP-2 - bone morphogenetic protein 2, BMP-6 - bone morphogenetic protein 6, BMP-7 - bone morphogenetic protein 7, PI3K/Akt - phosphoinositide 3-kinase/protein kinase B, AMPK - adenosine monophosphate-activated protein kinase, MAPK/ERK - mitogen-activated protein kinase/extracellular signal-regulated kinase, HIF-1α/VEGF - hypoxia-inducible factor 1-alpha/vascular endothelial growth factor, TGF-β/Smad2/3 - transforming growth factor-beta/smad family member 2/3. .
Figure 3.
Diagram summarizing the effects of salidroside (SAL) on cellular signaling pathways. Stimulatory effects are highlighted in green, whereas inhibitory effect is marked in red. Nrf2 - nuclear factor erythroid 2–related factor 2, BMP-2 - bone morphogenetic protein 2, BMP-6 - bone morphogenetic protein 6, BMP-7 - bone morphogenetic protein 7, PI3K/Akt - phosphoinositide 3-kinase/protein kinase B, AMPK - adenosine monophosphate-activated protein kinase, MAPK/ERK - mitogen-activated protein kinase/extracellular signal-regulated kinase, HIF-1α/VEGF - hypoxia-inducible factor 1-alpha/vascular endothelial growth factor, TGF-β/Smad2/3 - transforming growth factor-beta/smad family member 2/3. .

Table 1.
Effects of SAL on various cell lines in in vitro studies focusing on bone metabolism. rBMSC - rat bone marrow-derived mesenchymal stem cells, ALP - alkaline phosphatase, COL1A1 - collagen type I alpha 1, MG-63 - human osteoblast-like cell line, ROB - rat osteoblasts, MAPK/ERK - mitogen-activated protein kinase/extracellular signal-regulated kinase, PI3K/Akt - phosphoinositide 3-kinase/protein kinase B, C3H10T1/2 - mouse pluripotent mesenchymal stem cell-like fibroblasts, MC3T3-E1 - a mouse clonal, osteoblast-like cell line, BMP - bone morphogenetic protein, Smad - family of proteins that transduce extracellular signals from transforming growth factor beta (TGF-β) ligands, Runx2 - runt-related transcription factor 2, OSX - osterix, BAX - B-cell lymphoma (Bcl)-2-associated X protein, CASP3 - Caspase-3, CASP9 - Caspase-9, EA.hy926 - human endothelial cell line, HUVEC - human umbilical vein endothelial cells, HIF-1α - hypoxia-inducible factor 1-alpha, VEGF - vascular endothelial growth factor, AMPK - adenosine monophosphate-activated protein kinase, OCN - osteocalcin, Nrf2 - nuclear factor erythroid 2-related factor 2, Keap1 - Kelch-like ECH-associated protein 1, TGF-β - transforming growth factor-beta.
Table 1.
Effects of SAL on various cell lines in in vitro studies focusing on bone metabolism. rBMSC - rat bone marrow-derived mesenchymal stem cells, ALP - alkaline phosphatase, COL1A1 - collagen type I alpha 1, MG-63 - human osteoblast-like cell line, ROB - rat osteoblasts, MAPK/ERK - mitogen-activated protein kinase/extracellular signal-regulated kinase, PI3K/Akt - phosphoinositide 3-kinase/protein kinase B, C3H10T1/2 - mouse pluripotent mesenchymal stem cell-like fibroblasts, MC3T3-E1 - a mouse clonal, osteoblast-like cell line, BMP - bone morphogenetic protein, Smad - family of proteins that transduce extracellular signals from transforming growth factor beta (TGF-β) ligands, Runx2 - runt-related transcription factor 2, OSX - osterix, BAX - B-cell lymphoma (Bcl)-2-associated X protein, CASP3 - Caspase-3, CASP9 - Caspase-9, EA.hy926 - human endothelial cell line, HUVEC - human umbilical vein endothelial cells, HIF-1α - hypoxia-inducible factor 1-alpha, VEGF - vascular endothelial growth factor, AMPK - adenosine monophosphate-activated protein kinase, OCN - osteocalcin, Nrf2 - nuclear factor erythroid 2-related factor 2, Keap1 - Kelch-like ECH-associated protein 1, TGF-β - transforming growth factor-beta.
| First Author, Year (Reference) | Cell Lines | Biological manifestation |
|---|---|---|
| Pan et al., 2013 [24] | rBMSC | Promotion of osteoblast differentiation and bone formation, upregulation of osteogenic markers ALP and COL1A1. |
| Guo et al., 2017 [27] | MG-63, ROB | Enhanced proliferation and differentiation of osteoblasts, activation of MAPK/ERK and PI3K/Akt pathways, increased Runx2 and OSX expression. |
| Li et al., 2018 [23] | rBMSC | Increased osteogenic differentiation, enhanced ALP activity, and mineralization, promotion of bone formation, increased expression of Runx2 and ALP. |
| Xue et al., 2018 [29] | ROB | Protection against apoptosis, activation of PI3K/Akt pathway, reduction of apoptotic markers such as BAX, CASP3, and CASP9, increased expression of Runx2 and OSX. |
| Chen et al., 2019 [21] | C3H10T1/2, MC3T3-E1, rBMSC | Increased proliferation and differentiation of osteoblasts, enhanced ALP activity, and mineralization via BMP/Smad pathway activation, increased expression of Runx2 and OSX. |
| Guo et al., 2020 [26] | EA.hy926, HUVEC, MG-63, ROB | Increased proliferation, migration, capillary formation, and mineralization through HIF-1α/VEGF pathway activation, increased expression of ALP, Runx2, OSX, and VEGF. |
| Fu et al., 2022 [25] | MC3T3-E1 | Enhanced proliferation and differentiation of osteoblasts, AMPK activation, inhibition of bone resorption, increased expression of ALP, COL1A1, OCN, and Runx2. |
| Wang et al., 2022 [30] | ROB | Protection against oxidative stress, increased Nrf2 activation, decreased Keap1 expression, promotion of osteogenesis, increased expression of ALP and Runx2. |
| Xie et al., 2023 [28] | MC3T3-E1 | Protection against dexamethasone-induced inhibition, increased ALP activity, activation of TGF-β/Smad2/3 pathway, increased expression of OSX and ALP. |
Table 2.
Effects of SAL on various animal models in in vivo studies focusing on bone metabolism. OVX – ovariectomy, RANKL - receptor activator of nuclear factor kappa-Β ligand, OPG - osteoprotegerin, HIF-1α - hypoxia-inducible factor 1-alpha, VEGF - vascular endothelial growth factor, OP – osteoporosis, SANFH - steroid-induced avascular necrosis of the femoral head, PI3K/Akt - phosphoinositide 3-kinase/protein kinase B, Bcl-2 - B-cell lymphoma 2, BAX - Bcl-2-associated X protein, CASP3 - Caspase-3, MDA - malondialdehyde, ROS - reactive oxygen species, SOD - superoxide dismutase, GSH-Px - glutathione peroxidase, CD31 - Cluster of Differentiation 31 (platelet endothelial cell adhesion molecule), KOA - knee osteoarthritis, AMPK - adenosine monophosphate-activated protein kinase, TNF-α - tumor necrosis factor alpha, IL-1β - interleukin 1 beta, IL-6 - interleukin 6, Nrf2 - nuclear factor erythroid 2-related factor 2, Runx2 - runt-related transcription factor 2, ALP - alkaline phosphatase, OCN – osteocalcin, TGF-β - transforming growth factor-beta, Smad - family of proteins that transduce extracellular signals from transforming growth factor beta (TGF-β) ligands, COL1A1 - collagen type I alpha 1.
Table 2.
Effects of SAL on various animal models in in vivo studies focusing on bone metabolism. OVX – ovariectomy, RANKL - receptor activator of nuclear factor kappa-Β ligand, OPG - osteoprotegerin, HIF-1α - hypoxia-inducible factor 1-alpha, VEGF - vascular endothelial growth factor, OP – osteoporosis, SANFH - steroid-induced avascular necrosis of the femoral head, PI3K/Akt - phosphoinositide 3-kinase/protein kinase B, Bcl-2 - B-cell lymphoma 2, BAX - Bcl-2-associated X protein, CASP3 - Caspase-3, MDA - malondialdehyde, ROS - reactive oxygen species, SOD - superoxide dismutase, GSH-Px - glutathione peroxidase, CD31 - Cluster of Differentiation 31 (platelet endothelial cell adhesion molecule), KOA - knee osteoarthritis, AMPK - adenosine monophosphate-activated protein kinase, TNF-α - tumor necrosis factor alpha, IL-1β - interleukin 1 beta, IL-6 - interleukin 6, Nrf2 - nuclear factor erythroid 2-related factor 2, Runx2 - runt-related transcription factor 2, ALP - alkaline phosphatase, OCN – osteocalcin, TGF-β - transforming growth factor-beta, Smad - family of proteins that transduce extracellular signals from transforming growth factor beta (TGF-β) ligands, COL1A1 - collagen type I alpha 1.
| First Author, Year (Reference) | Animal Model | Biological effect |
|---|---|---|
| Pan et al., 2013 [24] | Sprague Dawley rats (OVX-induced bone loss model) | Reduced oxidative stress, increased bone mass, reduced RANKL, increased OPG. |
| Guo et al., 2017 [27] | BALB/c mice (tibia fracture model) | Accelerated fracture healing, enhanced osteoblast proliferation and differentiation, increased HIF-1α and VEGF. |
| Li et al., 2018 [23] | Sprague Dawley rats (OVX-induced OP model) | Increased bone mass and mineral apposition rates, improved bone microarchitecture, increased HIF-1α and VEGF. |
| Xue et al., 2018 [29] | Sprague Dawley rats (SANFH model) | Reduced osteoblast apoptosis, increased osteogenic differentiation via PI3K/Akt, increased Bcl-2, decreased BAX and CASP3. |
| Zheng et al., 2018 [33] | Sprague Dawley rats (OVX-induced OP model with diabetes) | Improved bone histomorphology, prevention of bone loss, upregulation of the OPG/RANKL ratio |
| Chen et al., 2019 [21] | C57BL/6 mice (oxidative stress model) | Preserved bone microstructure, reduced MDA and ROS, increased osteoblast function and antioxidant enzymes such as SOD and GSH-Px. |
| Guo et al., 2020 [26] | C57BL/6 mice (angiogenesis model using mouse embryonic metatarsals) | Enhanced endothelial sprouting, increased VEGF, increased CD31-positive endothelial cells. |
| Fu et al., 2022 [25] | C57BL/6 mice (KOA model) | Reduced inflammation, increased osteogenic protein expression, enhanced AMPK activation, reduced TNF-α, IL-1β, and IL-6. |
| Wang et al., 2022 [30] | Sprague Dawley rats (OVX-induced OP model) | Reduced oxidative stress, increased osteogenesis via Nrf2, increased Runx2, ALP, and OCN. |
| Xie et al., 2023 [28] | C57BL/6 mice (dexamethasone-induced OP model) | Mitigated inhibitory effects of dexamethasone on osteogenesis, activated TGF-β/Smad2/3, increased ALP and COL1A1. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



