
Submitted:
02 July 2024
Posted:
03 July 2024
You are already at the latest version
Abstract
Soil organic matter (SOM) is essential for nutrient cycling and soil carbon (C) accumu-lation, both of which are heavily influenced by the quality and quantity of plant litter. Since SOM dynamics in relation to plant diversity are poorly understood we investigated the effects of willow variety and mixture, and site on the soil C stocks, SOM chemical composition and ther-mal stability. Using pyrolysis-field ionization mass spectrometry we analysed the top 10 cm of soil from two 7-year-old experimental sites in Germany and Sweden, with monocultures and mixtures of two willow varieties (Salix spp.) belonging to different species. Overall, site had the strongest effect on SOM quality. Results showed significant variability across sites for willow identity and mixture effects on C accumulation and SOM chemistry. In the German site (Ros-tock), yearly soil C accumulation was higher (p < 0.05) for variety ‘Loden’ (1.0 Mg C ha-1 year-1) compared to ‘Tora’ (0.5 Mg C ha-1 year-1), whilst in the Swedish site (Uppsala), both varieties ex-hibited similar soil C accumulation rates of around 0.6 Mg C ha-1 year-1. Willow variety identity significantly affected SOM quality at both sites, while mixing had minor effects. Our findings emphasize the significance of site-specific context and variety or species identity in shaping soil C accumulation in willow plantations.
Keywords:
variety or species mixing
; Soil organic matter
; carbon sequestration
; chemical composition
; thermal stability
1. Introduction
Soils store more carbon (C) than both the atmosphere and all vegetation combined [1], thereby playing a critical role in terrestrial ecosystem CO2 exchange [2], and in balancing atmospheric CO2 emissions. Whether soils act as sinks or sources of CO2 depends on the balance between photosynthesis, respiration and stabilization of C belowground, and changes in soil C stocks have the potential to impact atmospheric CO2 levels and the global C budget [3]. Soil organic matter (SOM) comprises the largest active reservoir of terrestrial organic carbon [4], and plays a direct role in climate change mitigation [5,6,7]. However, the dynamics of SOM are complex and not fully understood [8], and improved understanding of its formation and stabilization is crucial for developing sustainable approaches to enhance soil C sequestration.
The SOM consists of decomposing plant material and microbial-derived compounds varying in size, extent of degradation, accessibility, and residence time [9]. The chemical composition and stability of SOM controls the rate at which microbial communities process organic matter, thus governing the accumulation of soil C [10,11]. Since soil respiration is influenced by the soil microbial activity a higher soil respiration rate could therefore reflect increased decomposition rates. In the present study we use chemistry indicating SOM stability as a proxy of SOM quality. The stability of SOM is partly determined by its constituent molecules, some of which are more resistant to decomposition (e.g., lignin, phenols and suberin; [12,13]) than others (e.g., free fatty acids and peptides; [14,15]). Higher proportions of recalcitrant molecules contribute to more stable SOM, which is more likely to accumulate over time [16]. Climatic and edaphic factors further complicate SOM dynamics, as temperature and precipitation influence microbial activity and soil type, which can be instrumental for SOM stabilization [17,18,19].
There are many wet-chemical, spectrometric and spectroscopic analytical methods available to assess the quality of SOM, e.g., [20,21]. Pyrolysis-field ionization mass spectrometry (Py-FIMS) is a powerful analytical tool, with which biogenic marker substances can be associated with molecular compound classes derived from soil samples, thus providing a means of characterizing SOM chemical composition while also providing the possibility to assess the thermal stability of these same biogenic markers [11,22]. It reveals the strength of chemical bonds within molecules or between reactive mineral surfaces and organic molecules in response to thermal stress, thus providing a measure of resistance to microbial decomposition [11,13]. This data-rich method provides a high-resolution overview of the SOM chemical structure with the potential to elucidate responses of belowground processes to factors such as plant species identity and plant species mixing. SOM chemical composition and thermal stability are important aspects to consider when examining how aboveground vegetation influences the stability of SOM in different environments.
The quantity and quality of plant litter input can vary greatly between species and vegetation types [23,24]. Multiple studies have shown plant diversity to enhance soil C sequestration [25,26], often through increased aboveground biomass production through altered resource competition or facilitative plant-plant interactions, thus increasing the amount of organic matter supplied to the soil and therefore the accumulation of C [27,28,29]. However, effects of species diversity on plant productivity are often highly context dependent [30,31]. Further, an increase in C storage with plant diversity can also result from longer persistence of plant litter due to slower decomposition [32]. Plant diversity, mainly studied in grasslands, but with evidence emerging from forest ecosystems, has been shown to affect SOM chemical diversity [33,34,35], microbial composition, activity and biomass [36,37], soil nutrient retention [38], and soil gas emissions [39]. Increased soil microbial biomass and consequently later necromass can make up more than half of SOC [40]. Plant diversity should therefore likely influence both SOM chemical composition and thermal stability, which could have major implications for SOM decomposition and ultimately ecosystem functioning [41,42,43,44]. Yet, the effects of plant diversity are not thoroughly explored in forest ecosystems, and especially the responses in SOM chemical composition and stability to tree species mixing remain poorly understood.
The cultivation of Salix spp. (willows) as short rotation coppice (SRC) has emerged as a promising approach to sustainably produce renewable biomass [45,46], with the largest cultivated areas found in China and Argentina, followed by Europe [47]. These production systems are characterized by short growth cycles of 2–5 years, after which the stems are harvested and shoots regrow rapidly from the stumps left in the soil [47]. In addition, willow SRC can serve as a model system for investigating plant diversity-productivity relationships [48], due to the fast growth and high phenotypic variability of willow species along with moderate to small phylogenetic contrasts. This enables the evaluation of subtle phenotypic differences on plant-plant interactions [49,50]. Different willow species and varieties can have differential impact on belowground C dynamics and soil microbiota [51,52,53], and willow species or varieties grown in mixtures can differ from their monoculture counterparts in terms of nitrogen economy and productivity [50,54]. In the present study, we used two phenotypically distinct willow varieties belonging to different species grown both as monoculture stands and as mixtures. The aim of the study was to evaluate the effects of willow variety identity, mixture and site on soil C accumulation, SOM chemical composition and thermal stability using Py-FIMS methodology. We hypothesized that: (i) willow variety identity affects soil C accumulation, SOM chemical composition and stability, (ii) willow variety mixtures accumulate similar amounts of soil C but differ in SOM chemical composition and stability compared to the equivalent monocultures at the same site, (iii) site conditions modulate the effects of variety identity and mixture on soil C accumulation, SOM chemical composition and stability.
2. Materials and Methods
2.1. Experimental Design and Plant Material
Two experimental sites, one in Rostock, Germany (54°02′ N 12°05′ E) and one in Uppsala, Sweden (59°49′ N 17°39′ E) were planted with willow in a short rotation coppice (SRC) system on former arable land in 2014. The two sites are part of the ECOLINK-Salix research trials as well as members of the global tree diversity network (TreeDivNet; [55]). Climatic conditions and soil properties differ between the two sites, notably clay content is roughly 10 times higher in the Uppsala site compared to the Rostock site (Table 1).
Two phenotypically distinct willow varieties belonging to different Salix species were used as stand components: ‘Loden’ (L; S. dasyclados Wimm.) and ‘Tora’ (T; S. schwerinii x S. viminalis). Generally, higher shoot biomass production is reported for ‘Tora’ than for ‘Loden’ [50,54], while leaf and fine root biomass production has been found to be higher for ‘Loden’ than for ‘Tora’ [57,58]. ‘Loden’ and ‘Tora’ also differ in terms of leaf chemistry and element stoichiometry [50,59], with [60] reporting weak effects of site, strong effects of variety and interaction effects between site and variety, on leaf litter decomposability as measured by fraction of remaining biomass after incubation. Willows can form both ecto- and arbuscular mycorrhizal associations [61], but are generally more associated with ectomycorrhizal fungi [62]. In addition, [63] reported on greater ectomycorrhizal colonization of root tips belonging to S. dasyclados Wimm. than S. viminalis L., which are similar to ‘Loden’ (a S. dasyclados variety) and ‘Tora’ (a S. schwerinii x S. viminalis variety), respectively. In the present study we use ‘Loden’ and ‘Tora’ in monoculture and the 2-component mixture. The willow varieties were planted in a randomized block design with 3 replicates for each monoculture (‘Loden’ and ‘Tora’) and mixture (‘Loden’:‘Tora’) for each site (i.e., 9 plots per site). All 9.6 × 9.6 m plots were planted in a hexagonal pattern with 12 rows, each row containing 12 plants at a distance of 0.8 m between neighboring plants, and further details can be found in [64].
2.2. Soil Sampling, Analyses and Measurements
Since the majority of Salix fine roots are found within the top 10 cm of soil [65,66], we focused our sampling efforts on this layer, assuming the most significant impact of Salix growth occurs here due to the turnover of fine roots and leaf litter. We collected nine soil samples per plot per site at experiment establishment in April 2014 and during the third cutting cycle in April 2021, using a soil auger (3 cm diameter). Subsamples were pooled per plot, dried at 40 degrees for 48 h and sieved (< 2 mm). The concentrations of C and N were determined using a CN analyzer (Vario EL, Elementar Analysensysteme, Hanau, Germany). Soil pH was determined in 0.01 M CaCl2 at a soil:solution ratio of 1:2.5 (w/v). The C stocks (Mg ha-1) were calculated for 2014 and 2021 by the following formula:
where Corg is the organic C concentration (g 100 g-1) in the soil, ρd is the dry bulk density (g cm-3), and depth is the soil sampling depth (10 cm). Yearly soil C accumulation (Mg C ha-1 year-1) was calculated as the difference between 2021 and 2014 C stocks averaged per year. The determination of the dry bulk density (ρd) was based on the use of 250 cm3 cores in 2014 and 2021 (ρd = dry weight [g] / volume [250 cm3]). The soil cores were dried at 105 °C until a constant weight was achieved.
Cstocks = Corg * ρd * depth
Double lactate-extractable phosphorus (Pdl), potassium (Kdl) and magnesium (Mgdl) were determined after extraction of 0.6 g soil with 30 ml lactate solution [67]. The element concentrations in the extract were measured with inductively coupled plasma-optical emission spectroscopy (ICP-OES, Optima 8300, Perkin Elmer, Waltham, Massachusetts, USA). The lactate-extractable concentrations have been considered to represent the plant-available fraction in soils [68].
CO2 respiration was measured at 4-week intervals (8 times) during the growing season (April to October) for year 2021 at the Uppsala site as a proxy for decomposition. Measurements were made using a portable infrared gas analyzer coupled to a 1296 cm3 dark chamber in closed air circuit (EgM-4 with SRC-1 probe type; PP Systems, Amesbury, Massachusetts, USA). The chamber was pressed firmly to the ground (avoiding weeds) for measurements in at least four different places per plot at each sampling occasion, evenly spread out with the intention to reflect the plot-level treatments. Due to cracking clay soil surfaces during dry summer months the sampling place varied between sampling occasions, ensuring a tight seal and avoiding cracks in the soil where large amounts of CO2 was released. Respiration was typically monitored between 9:00 and 14:00 CET on cloudy to half-cloudy days after a recent rainfall if possible for ca 120 s per measurement and calculated as a function of the linear increase of CO2 concentration in the chamber. Measurement quality was ensured by automatic baseline calibration by the ‘auto-zero’ option at least every 20 min and by keeping the start CO2 concentration in the chamber at ambient level [69].
2.4. Pyrolysis-Field Ionization Mass Spectrometry (Py-FIMS)
SOM chemical composition and thermal stability were analysed by pyrolysis-field ionization mass spectrometry (Py-FIMS) for samples from 2021 in three plots each for ‘Loden’, ‘Tora’ and ‘Loden’:‘Tora’ plots from the Rostock site and from two plots each from the Uppsala site. About 3 mg of the air-dried, ground and homogenized samples were thermally degraded by pyrolysis in the ion source (emitter: 4.7 kV, counter electrode –5.5 kV) of a double-focusing Finnigan MAT 95. The samples were heated in a vacuum of 10–4 Pa from 50 °C to 700 °C, in temperature steps of 10 °C over a time period of 18 min. Between magnetic scans the emitter was flash heated to avoid residues of pyrolysis products. About 65 spectra were recorded for the mass range m/z 15 to 900. Biogenic marker signals (m/z) were assigned to nine relevant compound classes according to [21] (Table 2). All recorded marker signals (m/z) were combined to obtain the total ion intensity (TII) for each measurement. The difference in sample weight before and after pyrolysis provides a measure of “volatile matter” (VM) and is used to normalize sample ion intensities per mg sample weight. The hexoses:pentoses ratio is a measure of microbial- to plant-derived sugars [70] and is used as a measure of microbial contribution to SOM. Additionally, the ion intensities at each temperature step during pyrolysis was calculated separately for each of the about 65 single scans. The ion intensities of each compound class, plotted against the volatilization temperature, provided distinct thermograms that could be evaluated in terms of the thermal stability of compound classes.
2.5. Statistical Analyses
All statistical analyses were conducted in R (version 4.3.0; [75]). SOM chemical composition for the two willow varieties in monoculture and mixture per study site was visualized using principal component analysis (PCA; function rda in package vegan; [76]; Figure 2). The PCA was performed on the relative ion intensities for the 9 Py-FIMS compound classes (Table 2). We used a principal component approach for the compound class data due to significant covariance between several of the compound classes. Principal component scores (PC1 and PC2; function scores in package vegan; [76]) were used as response variables in mixed effects principal component regression models [77] to assess multivariate differences in SOM chemical composition between treatments.
The Shannon diversity index (H’), which accounts for both presence and relative abundance, was applied to our Py-FIMS biogenic marker signals (m/z) to quantify the chemical diversity of SOM (function diversity in package vegan; [76]):
where pi is the proportion of relative ion intensity for marker signal (m/z) i, and n is the number of measured marker signals in a given sample.
The SOM thermal stability, here used as an indicator of resistance to microbial decomposition [21], was calculated for total ion intensity (TII) as well as for each compound class separately (Table 2). This was done following [78] by dividing the sum of ion intensities volatilized at high temperature (> 400 °C) by ion intensities volatilized over the whole temperature range (50-650 °C).
Net diversity effects (NDE) of selected SOM properties were evaluated following [27]:
in which, the observed values under variety mixtures are compared to the expected mixture values, calculated as the average values under their monoculture counterparts. Mixture effects are characterized as non-additive if significantly different from zero (NDE ≠ 0) or additive if they are not (NDE = 0). To avoid any mixture effects being occluded by potential dominance effects, we applied weights to the expected mixture values based on their proportional basal area in the mixtures [29] (Supplementary Material Table S1).
Responses in soil C accumulation, nutrient concentrations, Py-FIMS compound classes and SOM thermal stability were modeled using mixed effects multiple linear regression models with the fixed effects of variety composition (‘Loden’, ‘Loden’:‘Tora’ and ‘Tora’), site (Rostock and Uppsala) and their interaction (function lme in package nlme; [79]). Block nested in site was added to the model as a random effect to account for site-specific block effects, which were especially noticeable at the Rostock site. Due to different variance structures at our sites, site-specific variance weights were included in the model. Soil CO2 respiration was sampled only in the Uppsala site and was modeled using a mixed effects multiple linear regression model in response to the fixed effects of variety composition and undergrowth ground cover (%). Block and plot nested in block were added as random effects to account for block effects and repeated measurements respectively. Temporal autocorrelation between months was accounted for using an autoregressive correlation structure. For each response variable in the papaer, the statistical significance of site, variety composition and their interaction was evaluated using analyses of variance (ANOVA; type = III, Kenward-Roger’s method, function anova in base R) and the results of these analyses are presented in Supplementary Material Tables S2–S7. This was followed by pairwise comparisons of estimated marginal means using a Tukey adjustment (function emmeans in package emmeans; [80]). The function contrast from the emmeans package was used to examine specific variations across sites, varieties within sites, varieties across sites, as well as the disparities between expected and observed mixture values within sites and observed mixture values across sites Estimated marginal means were used to calculate mean values due to the partially unbalanced nature of our dataset.
3. Results
3.1. Site Characteristics
The contents of soil C (p = 0.004), N (p = 0.001), Kdl (p = 0.002) and Mgdl (p = 0.011) were significantly lower at the Rostock than Uppsala site (Table 3), while C:N ratio and Pdl were similar between sites. No differences in the above characteristics were found between willow varieties grown in monoculture or mixture at either site.
The total thermal stability of bulk SOM was similar across sites, but total ion intensity (TII; p = 0.004, hexoses:pentoses ratio (p = 0.004) and chemical diversity (H’; p < 0.001) were all significantly higher in Rostock than Uppsala (Table 4). Volatile matter was significantly higher in Uppsala than Rostock (VM; p = 0.003). In terms of compound classes, the Uppsala site had significantly higher relative abundances of carbohydrates (CHYDR; 5.0% TII, p < 0.001), phenols and lignin monomers (PHLM; 5.9% TII, p < 0.001), alkylaromatics (ALKYL; 3.5% TII, p < 0.001), N-compounds (NCOMP; 2.2% TII, p < 0.001) and peptides (PEPTI; 2.3% TII, p = 0.001); while lower relative abundances of lipids (LIPID; -1.7% TII, p = 0.003), suberin (SUBER; -0.2% TII, p < 0.001) and free fatty acids (FATTY; -0.5% TII, p = 0.015) compared to the Rostock site.
3.2. Effects of Willow Variety
Soil C stocks measured at the establishment of the experiment in 2014 were similar across all plots within each site and averaged at 10.32 and 20.54 Mg C ha-1 for sites Rostock and Uppsala, respectively. Seven years after planting, yearly soil C accumulation rates measured at the Rostock site under ‘Loden’ (1.02 Mg C ha-1 year-1) were significantly higher (p = 0.004) than under ‘Tora’ (0.54 Mg C ha-1 year-1). Soil C accumulation rates measured at the Uppsala site were similar between ‘Loden’ (0.55 Mg C ha-1 year-1) and ‘Tora’ (0.65 Mg C ha-1 year-1). Furthermore, neither Loden nor Tora differ in C accumulation rates between sites (Figure 1).
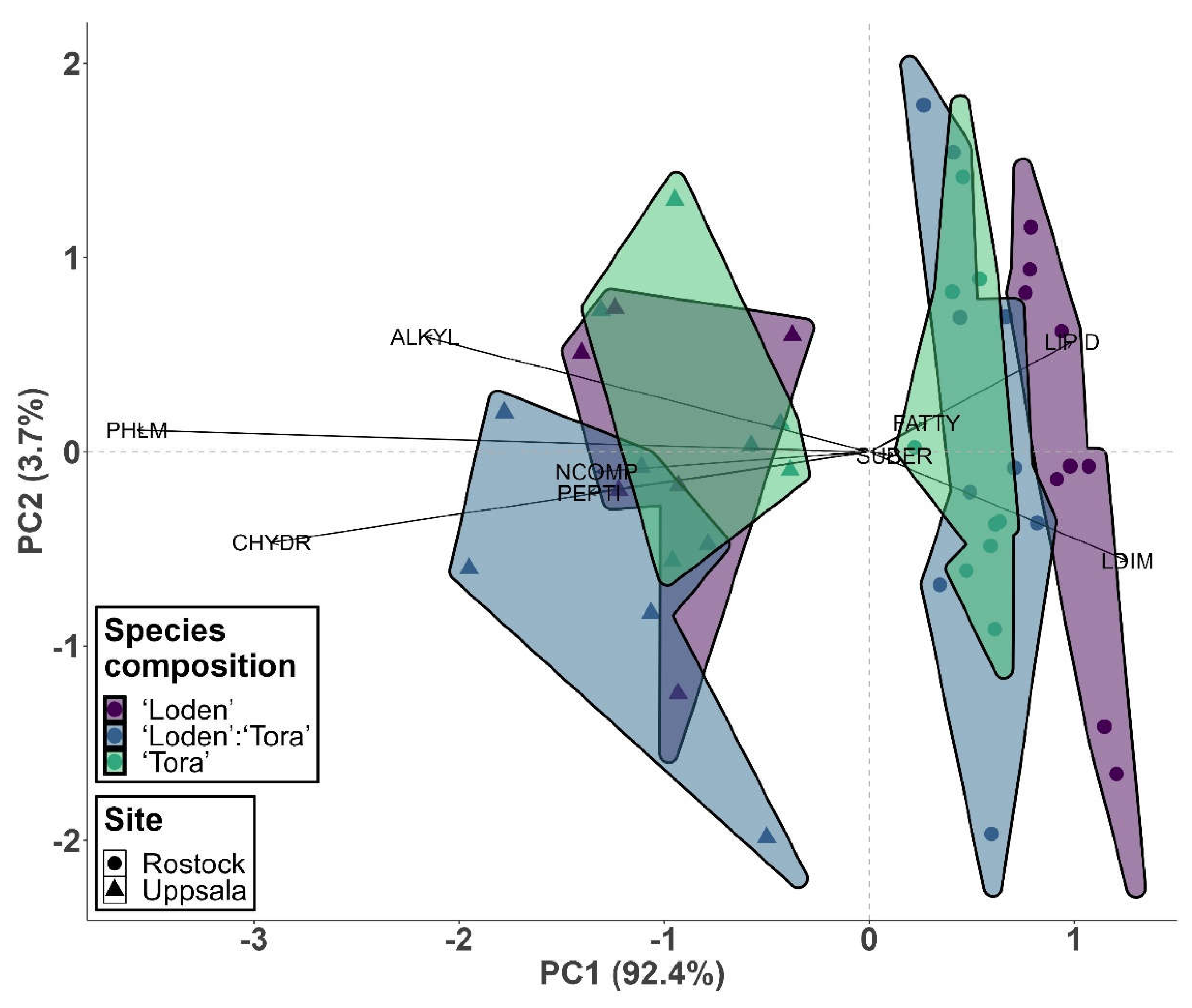
The SOM chemical composition differed between ‘Loden’ and ‘Tora’ (p = 0.036) and between the mixture and ‘Tora’ (p = 0.043) along PC1 at the Rostock site (Figure 2). No significant differences between ‘Loden’, ‘Loden’:‘Tora’ or ‘Tora’ were found along PC1 at the Uppsala site, or along PC2 at either site. Principal component 1 (PC1) explained 92.4% of the variation in the 9 compound classes from Py-FI mass spectra (Table 2) of soil samples under the willow varieties in monoculture and mixture and was strongly associated (p < 0.001) with all nine compound classes. Principal component 2 (PC2) explained 3.7% of the variation and correlated with compound classes lignin dimers (LDIM), lipids (LIPID) and free fatty acids (FATTY) (p < 0.05).
Figure 2.
SOM chemical composition: principal components plot of the relative ion intensities (% TII) of nine compound classes; CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER, FATTY in Py-FI mass spectra (for abbreviations, see Table 2) in topsoil (0-10 cm depth) for the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture and mixture at sites Rostock and Uppsala.
Figure 2.
SOM chemical composition: principal components plot of the relative ion intensities (% TII) of nine compound classes; CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER, FATTY in Py-FI mass spectra (for abbreviations, see Table 2) in topsoil (0-10 cm depth) for the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture and mixture at sites Rostock and Uppsala.
The SOM under ‘Tora’ exhibited significantly greater abundances of carbohydrates (CHYDR; 1.0% TII, p = 0.015), phenols and lignin monomers (PHLM; 1.8% TII, p = 0.018), N-compounds (NCOMP; 0.5% TII, p = 0.017), and peptides (PEPTI; 0.6% TII, p = 0.026), but significantly lower abundances of suberin (SUBER; -0.2% TII, p = 0.014) and free fatty acids (FATTY; -0.6% TII, p = 0.014), compared to SOM under ‘Loden’ at the Rostock site (Table 5). Conversely, at Uppsala, the abundance of lignin dimers (LDIM; 1.7% TII, p < 0.001) was significantly higher in SOM under ‘Tora’ than ‘Loden’, while peptides (PEPTI; -0.8% TII, p < 0.001) and free fatty acids (FATTY; -0.1% TII, p = 0.004) were significantly lower under ‘Tora’ compared to ‘Loden’. Additionally, comparing SOM chemical composition under each willow variety between sites, ‘Loden’ cultivated in Uppsala showed significantly higher abundances of carbohydrates (CHYDR; 5.5% TII, p = 0.003), phenols and lignin monomers (PHLM; 7.3% TII, p = 0.001), alkylaromatics (ALKYL; 4.5% TII, p = 0.002), N-compounds (NCOMP; 2.6% TII, p = 0.002), and peptides (PEPTI; 2.8% TII, p < 0.001) than ‘Loden’ cultivated in Rostock. In contrast, compound classes lignin dimers (LDIM; -2.6% TII, p = 0.040), lipids (LIPID; -2.0 % TII, p = 0.006), suberin (SUBER; -0.31% TII, p = 0.001), and free fatty acids (FATTY; -0.71% TII, p = 0.008) were significantly lower under ‘Loden’ when cultivated in Uppsala compared to Rostock. For ‘Tora’, significant differences between sites included higher abundances of carbohydrates (CHYDR; 3.8% TII, p = 0.010), phenols and lignin monomers (PHLM; 4.5% TII, p = 0.008), alkylaromatics (ALKYL; 2.9% TII, p = 0.009), N-compounds (NCOMP; 1.5% TII, p = 0.016) and peptides (PEPTI; 1.4% TII, p = 0.012), in Uppsala compared to Rostock. Conversely, abundances of lipids (LIPID; -1.2% TII, p = 0.037) and suberin (SUBER; -0.11% TII, p = 0.038) were significantly lower in Uppsala compared to Rostock under ‘Tora’.
CO2 respiration, which was measured only at the Uppsala site, was similar across all variety compositions (Table 3). Chemical diversity (H’) was significantly higher under ‘Loden’ compared to ‘Tora’ at the Rostock site (p = 0.028), with no difference at the Uppsala site (Table 4). The ratio of microbial- to plant-derived sugars (hexoses:pentoses), an indicator of microbial contribution to SOM, was significantly higher in SOM under ‘Tora’ compared to ‘Loden’ and the ‘Loden’:‘Tora’ mixture at the Uppsala site (p = 0.021 and p = 0.026, respectively). Total ion intensity (TII) was similar within sites, while total thermal stability was lower under ‘Loden’:‘Tora’ compared to ‘Loden’ (p = 0.018) only at the Uppsala site. Volatile matter (VM) was lower under ‘Loden’:‘Tora’ compared to both ‘Loden’ and ‘Tora’ at the Uppsala site (p < 0.001 and p = 0.003, respectively).
The thermal stability of compound classes lignin dimers (LDIM; p = 0.047), lipids (LIPID; p = 0.030), alkylaromatics (ALKYL; p = 0.032), suberin (SUBER; p = 0.015) and free fatty acids (FATTY; p = 0.033) were all significantly higher under ‘Tora’ than ‘Loden’ at the Rostock site (Table 6 and Figure 3). In contrast, at the Uppsala site, only suberin had a greater thermal stability under ‘Tora’ (p < 0.001), representing the sole compound class that consistently responded to variety identity across sites. Compound classes carbohydrates (CHYDR; p = 0.004), phenols and lignin monomers (PHLM; p = 0.006), alkylaromatics (p = 0.022), N-compounds (NCOMP; p = 0.009), and peptides (PEPTI; p < 0.001) were all significantly more stable under ‘Loden’ than ‘Tora’ at the Uppsala site (Table 6 and Figure 4). In terms of comparisons of SOM thermal stability under each willow variety between sites, ‘Loden’ cultivated in Uppsala showed significantly higher thermal stability of compound classes phenols and lignin monomers (PHLM; p = 0.033), lipids (LIPID; p = 0.014), alkylaromatics (ALKYL; p = 0.038) and N-compounds (NCOMP; p = 0.022) compared to ‘Loden’ cultivated in Rostock. For ‘Tora’, no significant differences in SOM thermal stability were found between the sites.
At the Rostock site, the mean thermal stability of each compound class was as follows: lignin dimers > suberin > alkylaromatics > lipids > phenols and lignin monomers > peptides > N-compounds > carbohydrates > free fatty acids (Figure 3).
At the Uppsala site, the mean thermal stability of each compound class was as follows: lignin dimers > lipids > suberin > alkylaromatics > phenols and lignin monomers > N-compounds > peptides > carbohydrates > free fatty acids (Figure 4).
3.3. Effects of Variety Mixing
Expected values of yearly soil C accumulation in mixtures, as based on basal area weighted and averaged monoculture values, were similar to observed mixture values at either site (Figure 5). Expected and observed soil C accumulation in mixtures were 0.75 and 0.98 Mg C ha-1 year-1, respectively, at the Rostock site, and 0.61 and 0.16 Mg C ha-1 year-1, respectively, at the Uppsala site. At the Rostock site, observed mixture values were found to be significantly higher than at the Uppsala site (p = 0.046).
Expected SOM chemical composition in mixtures, as based on basal area weighted and averaged monoculture values, were mostly similar to the observed mixture values (Table 7). Significant non-additive diversity effects were found for lignin dimers (LDIM) and peptides (PEPTI) at the Uppsala site, where lignin dimer abundance was reduced under mixtures (NDE = -18.3%, p < 0.001) compared to monocultures and peptide abundance was elevated under mixtures compared to monocultures (NDE = 18.2%, p < 0.001).
Comparable to SOM chemical composition, SOM thermal stability was mostly similar between expected and observed mixture values for most compound classes, though some significant non-additive mixture effects were found, but only at the Uppsala site (Table 8 and Figure 4). There, the stability of compound classes ALKYL (NDE = -0.9%, p = 0.036) and suberin (SUBER; NDE = -22.7%, p < 0.001) were both lower under mixtures compared to monocultures.
4. Discussion
4.1. Effects of Site-Specific Environmental Conditions
Our study used two experimental sites with distinct climatic and edaphic circumstances, which provided a platform to investigate how local conditions modulate the influence of willows on SOM quality. Our results showed that the interaction between plants and the investigated soil properties is strongly influenced by local site conditions, where soil C accumulation, SOM chemical composition and thermal stability responded differently to both variety identity and mixture across sites (confirming hypothesis 3). Temperature, moisture and especially clay content, which differed between the two study sites, are widely documented as strong modifiers of decomposition rates [23,81], and microbial dynamics [82,83,84]. Therefore, we expected the lower temperatures and precipitation levels as well as higher clay content in Uppsala to contribute to lower SOM decomposition rates compared to Rostock. In the present study, this is partly supported by the lower ratio of microbial- to plant-derived sugars (hexoses:pentoses) found in Uppsala compared to Rostock, indicating lower microbial contribution to SOM [70,85]. Additional supporting evidence for lower decomposition rates in Uppsala compared to Rostock comes from a previous study on the same willow SRCs [60], showing slower leaf litter decomposition rates for monocultures and higher fractions of remaining N (immobilization of N) in mixtures in Uppsala compared to Rostock, though with only minor effects of climate. However, the highest accumulation of soil C was found under ‘Loden’ in Rostock, where we expected a higher decomposition rate compared to Uppsala. This discrepancy between likely outcomes based on literature evaluations and results may be explained by the generally greater biomass production at the Rostock site [64], leading to a greater litter input to the soil.
4.2. Effects of Willow Variety
In the present study, we demonstrated that the influence of willow SRC on soil C accumulation and SOM chemistry strongly depended on willow variety (confirming hypothesis 1). After seven years of growth, the net average annual C accumulation in the topsoil (0-10 cm depth) was consistently positive across all experimental plots, with C accumulation rates ranging from 0.06 to 1.29 Mg C ha-1 year-1. These values were comparable to mean soil C accumulation values documented under willow SRC for ‘Loden’ (0.73 Mg C ha-1 year-1) and ‘Tora’ (0.65 Mg C ha-1 year-1) near the Uppsala site over a 17 year period in [51]. In the site with climatically more favorable growth conditions (Rostock), ‘Loden’ had a higher C accumulation rate compared to ‘Tora’, whilst both varieties accumulated similar soil C amounts at the site with climatically less favorable growth conditions (Uppsala). A greater C accumulation under ‘Loden’ could be due to a higher fine root biomass production compared to ‘Tora’, as reported by [57] at the Rostock site. Furthermore, at the Rostock site, ‘Loden’ also had higher abundances of suberin compared to ‘Tora’, a compound primarily derived from roots [86]. This suggests a higher fine root biomass production under ‘Loden’ at the Rostock site, thus in line with the findings of [57]. Interestingly, the difference in C accumulation between ‘Loden’ and ‘Tora’ in Rostock was similar to what [51] reported in a site near Uppsala after 17 years of growth. It is therefore plausible that we in the future (e.g., after an additional 10 years) will see a significantly greater soil C accumulation under ‘Loden’ than ‘Tora’ also in the site with less favorable growth conditions (Uppsala). The difference in C accumulation between ‘Loden’ and ‘Tora’ in Rostock could also depend on differences in SOM chemistry, or results in different stability of the accumulated SOM. At the Rostock site, we found more chemically diverse SOM under ‘Loden’ compared to ‘Tora’, which may have contributed to a decreased decomposition and a subsequently promoted C accumulation for ‘Loden’ [4,87]. The relative enrichment of monomeric lignin building blocks under ‘Tora’ at the Rostock site, which can be explained by an advanced lignin decomposition [15], also supports enhanced decomposition in ‘Tora’ plots. Further, ‘Tora’ plots at the Rostock site displayed higher overall thermal stabilities of compound classes, suggesting higher decomposition rates as the more labile fractions of plant litter SOM would be decomposed first [11], leaving more recalcitrant molecules behind. A final potential explanation to the differences in C accumulation between ‘Loden’ and ‘Tora’, in addition to differential biomass production and decomposition rates, can depend on the interaction between mycorrhizal and saprotrophic fungi. Since ‘Loden’ is more dependent on the ectomycorrhizal association compared to ‘Tora’ [57,63] the saprotrophic activity under ‘Loden’ may have been suppressed as per the Gadgil effect [88]. The Gadgil effect implies suppression of saprotrophic activity in the presence of increased ectomycorrhizal colonization due to competition between the two functional groups and assimilate transfer from the host plant.
4.3. Effects of Variety Mixing
In the present study, we found no effect of variety mixing on soil C accumulation (confirming part of hypothesis 2), which is consistent with the findings of [89,90,91]. Since previous investigations of mixed-species willow plantations with ‘Loden’ and ‘Tora’ have found no indication of aboveground over-yielding [64,92], which is considered one of the main mechanisms of increased soil C sequestration in mixtures [25,93], this may explain the lack of response. However, even without increased plant productivity, ‘Loden’:‘Tora’ mixtures have been shown to affect other belowground processes such as soil phosphorus (P) cycling [57] e.g., through promoting alkaline phosphatase activity in P-deficient soils in mixtures compared to monocultures [92], and to increase endophytic root colonization [57], when compared to their monoculture counterparts. We detected some significant effects of variety mixing on SOM chemical composition and thermal stability (confirming the second part of hypothesis 2). At the Uppsala site, the detected effects of variety mixing on SOM chemical composition were found in the compound classes of lignin dimers and peptides, which revealed significantly higher and lower abundances, respectively, in variety mixtures compared to equivalent monocultures. Elevated levels of (lignin) decomposition in mixtures as a response to litter diversity is a possible explanation for these findings [94], which might be based on the higher general decomposition activity as revealed for the P cycling [57]. Since lignin is a rather stable fraction of SOM, lower lignin levels in mixtures could reduce SOM stability and decrease the long-term C sequestration in mixtures. Even so, the absence of effects of variety mixing on soil C accumulation and only some effects on SOM chemistry suggests that mixed willow cultivation does not greatly impair SOM quality or climate change mitigation potential compared to equivalent monoculture plantations at the same location. However, an increased decomposition of lignin in the mixtures in Uppsala could also indicate a decreased SOM formation. Finally, soil respiration, which is largely based on microbial activity in the soil, can reflect increased decomposition rates when respiration rates are higher. However, we did not find any difference in soil respiration between the treatments in the present study.
5. Conclusions
Our results highlight the complexity of plant-soil interactions and the challenges they present during interpretation. However, despite the complex nature of our results, our study revealed that willow variety or species identity could have a significant impact on soil C accumulation, SOM chemical composition and thermal stability. We found no evidence for non-additive effects of willow variety mixtures on soil C accumulation, but some non-additive effects on SOM chemical composition and thermal stability. Significant interaction between willow variety and site conditions have important implications for the management of willow SRC plantations. First, variety or species mixtures can be used in multifunctional plantations without greatly compromising soil C accumulation or SOM quality, as they mainly exhibit values similar to their monoculture counterparts. Second, the selection of willow varieties or species for willow SRC plantations need to consider the context-dependency of their expression under specific site conditions.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: PCA using Py-FI mass signals (m/z) instead of compound classes; Table S1: Basal area weights used for calculating net diversity effects (NDE); Table S2: Soil chemical characteristics: results of two-way ANOVAs for soil C:N ratios nutrient concentrations and soil CO2 respiration; Table S3: Py-FIMS parameters: results of two-way ANOVAs for various SOM properties; Table S4: Soil C accumulation: results of a two-way ANOVA of topsoil C accumulation rates; Table S5: Principal component regression: results of two-way ANOVAs for principal components (PC1 and PC2) derived from the relative ion intensity (% TII) of nine compound classes in topsoil; Table S6: SOM chemical composition: results of two-way ANOVAs for the relative ion intensity (% TII) of nine compound classes in Py-FI mass spectra in topsoil; Table S7: SOM thermal stability: results of two-way ANOVAs for the thermal stability of nine compound classes in Py-FI mass spectra in topsoil; Table S8: Py-FI mass spectral data of soil samples from Uppsala and Rostock; Table S9: C stocks and accumulation rates as well as P, Mg and K concentrations in topsoil.
Author Contributions
Conceptualization: Joel Jensen, Petra Fransson, Christel Baum, Peter Leinweber and Martin Weih; methodology, Joel Jensen; formal analysis, Joel Jensen; investigation, Joel Jensen and Kai-Uwe Eckhardt; resources, Christel Baum and Peter Leinweber; data curation, Kai-Uwe Eckhardt; writing-original draft preparation, Joel Jensen; writing-review and editing, Petra Fransson, Christel Baum, Peter Leinweber and Martin Weih; visualization, Joel Jensen; supervision, Petra Fransson, Christel Baum and Martin Weih. All authors have read and agreed to the published version of the manuscript.
Funding
This research received funding from FORMAS (The Swedish Research Council for Environment, Agricultural sciences and Spatial Planning) for the MixForChange project [grant number 2020-02339]. The establishment and management of the field trial was funded by grants from the Swedish Energy Agency [project numbers 36654-1, 36654-2]. Part of the soil analyses was funded by Deutsche Forschungsgemeinschaft (DFG) [project number BA 1494/9-1]. Petra Fransson was supported by the Swedish Research Council FORMAS [grant number 2016-01107].
Data Availability Statement
Data is available in the supplementary material, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Carvalhais, N.; Forkel, M.; Khomik, M.; Bellarby, J.; Jung, M.; Migliavacca, M.; Saatchi, S.; Santoro, M.; Thurner, M.; Weber, U. Global Covariation of Carbon Turnover Times with Climate in Terrestrial Ecosystems. Nature 2014, 514, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.A.; Exbrayat, J.-F.; van der Velde, I.R.; Feng, L.; Williams, M. The Decadal State of the Terrestrial Carbon Cycle: Global Retrievals of Terrestrial Carbon Allocation, Pools, and Residence Times. Proceedings of the National Academy of Sciences 2016, 113, 1285–1290. [Google Scholar] [CrossRef]
- Bellamy, P.H.; Loveland, P.J.; Bradley, R.I.; Lark, R.M.; Kirk, G.J. Carbon Losses from All Soils across England and Wales 1978–2003. Nature 2005, 437, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.; Hansel, C.M.; Kaiser, C.; Kleber, M.; Maher, K.; Manzoni, S.; Nunan, N.; Reichstein, M.; Schimel, J.P.; Torn, M.S.; et al. Persistence of Soil Organic Carbon Caused by Functional Complexity. Nat. Geosci. 2020, 13, 529–534. [Google Scholar] [CrossRef]
- Huguenin-Elie, O.; Delaby, L.; Klumpp, K.; Lemauviel-Lavenant, S. The Role of Grasslands in Biogeochemical Cycles and Biodiversity Conservation. In Improving grassland and pasture management in temperate agriculture; Burleigh Dodds Science Publishing, 2019; pp. 23–50 ISBN 1-351-11456-5.
- Lal, R. Soil Carbon Sequestration to Mitigate Climate Change. Geoderma 2004, 123, 1–22. [Google Scholar] [CrossRef]
- Lehman, R.M.; Cambardella, C.A.; Stott, D.E.; Acosta-Martinez, V.; Manter, D.K.; Buyer, J.S.; Maul, J.E.; Smith, J.L.; Collins, H.P.; Halvorson, J.J.; et al. Understanding and Enhancing Soil Biological Health: The Solution for Reversing Soil Degradation. Sustainability 2015, 7, 988–1027. [Google Scholar] [CrossRef]
- Ramesh, T.; Bolan, N.S.; Kirkham, M.B.; Wijesekara, H.; Kanchikerimath, M.; Srinivasa Rao, C.; Sandeep, S.; Rinklebe, J.; Ok, Y.S.; Choudhury, B.U.; et al. Chapter One - Soil Organic Carbon Dynamics: Impact of Land Use Changes and Management Practices: A Review. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press, 2019; Vol. 156, pp. 1–107.
- Lehmann, J.; Kleber, M. The Contentious Nature of Soil Organic Matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M. Thermal Adaptation of Decomposer Communities in Warming Soils. Frontiers in Microbiology 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Leinweber, P.; Jandl, G.; Baum, C.; Eckhardt, K.-U.; Kandeler, E. Stability and Composition of Soil Organic Matter Control Respiration and Soil Enzyme Activities. Soil Biology and Biochemistry 2008, 40, 1496–1505. [Google Scholar] [CrossRef]
- Gleixner, G. Soil Organic Matter Dynamics: A Biological Perspective Derived from the Use of Compound-Specific Isotopes Studies. Ecol Res 2013, 28, 683–695. [Google Scholar] [CrossRef]
- von Lützow, M.; Kögel-Knabner, I.; Ludwig, B.; Matzner, E.; Flessa, H.; Ekschmitt, K.; Guggenberger, G.; Marschner, B.; Kalbitz, K. Stabilization Mechanisms of Organic Matter in Four Temperate Soils: Development and Application of a Conceptual Model. Journal of Plant Nutrition and Soil Science 2008, 171, 111–124. [Google Scholar] [CrossRef]
- Jandl, G.; Leinweber, P.; Schulten, H.-R.; Ekschmitt, K. Contribution of Primary Organic Matter to the Fatty Acid Pool in Agricultural Soils. Soil Biology and Biochemistry 2005, 37, 1033–1041. [Google Scholar] [CrossRef]
- Schulten, H.-R.; Leinweber, P. Thermal Stability and Composition of Mineral-Bound Organic Matter in Density Fractions of Soil. European Journal of Soil Science 1999, 50, 237–248. [Google Scholar] [CrossRef]
- Hensgens, G.; Lechtenfeld, O.J.; Guillemette, F.; Laudon, H.; Berggren, M. Impacts of Litter Decay on Organic Leachate Composition and Reactivity. Biogeochemistry 2021, 154, 99–117. [Google Scholar] [CrossRef]
- Gabarrón-Galeote, M.A.; Trigalet, S.; Wesemael, B. van Soil Organic Carbon Evolution after Land Abandonment along a Precipitation Gradient in Southern Spain. Agriculture, Ecosystems & Environment 2015, 199, 114–123. [Google Scholar] [CrossRef]
- González-Domínguez, B.; Niklaus, P.A.; Studer, M.S.; Hagedorn, F.; Wacker, L.; Haghipour, N.; Zimmermann, S.; Walthert, L.; McIntyre, C.; Abiven, S. Temperature and Moisture Are Minor Drivers of Regional-Scale Soil Organic Carbon Dynamics. Sci Rep 2019, 9, 6422. [Google Scholar] [CrossRef] [PubMed]
- Le Roux, X.; Schmid, B.; Poly, F.; Barnard, R.L.; Niklaus, P.A.; Guillaumaud, N.; Habekost, M.; Oelmann, Y.; Philippot, L.; Salles, J.F. Soil Environmental Conditions and Microbial Build-up Mediate the Effect of Plant Diversity on Soil Nitrifying and Denitrifying Enzyme Activities in Temperate Grasslands. PLoS One 2013, 8, e61069. [Google Scholar] [CrossRef] [PubMed]
- Frimmel, F.H.; Abbt-Braun, G.; Heumann, K.G.; Hock, B.; Lüdemann, H.-D.; Spiteller, M. Refractory Organic Substances in the Environment; John Wiley & Sons, 2008; ISBN 978-3-527-61444-8.
- Leinweber, P.; Jandl, G.; Eckhardt, K.-U.; Schulten, H.-R.; Schlichting, A.; HofMann, D. Analytical Pyrolysis and Soft-Ionization Mass Spectrometry. In BIOPHYSICO-CHEMICAL PROCESSES INVOLVING NATURAL NONLIVING ORGANIC MATTER IN ENVIRONMENTAL SYSTEMS.; Senesi, N., Xing, B., Huang, P.M., Eds.; John Wiley & Sons, 2009; pp. 539–588.
- Schmidt, L.; Warnstorff, K.; Dörfel, H.; Leinweber, P.; Lange, H.; Merbach, W. The Influence of Fertilization and Rotation on Soil Organic Matter and Plant Yields in the Long-Term Eternal Rye Trial in Halle (Saale), Germany. Journal of Plant Nutrition and Soil Science 2000, 163, 639–648. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration; Springer Nature, 2020; ISBN 978-3-030-59631-6.
- Warembourg, F.R.; Roumet, C.; Lafont, F. Differences in Rhizosphere Carbon-Partitioning among Plant Species of Different Families. Plant and Soil 2003, 256, 347–357. [Google Scholar] [CrossRef]
- Augusto, L.; Boča, A. Tree Functional Traits, Forest Biomass, and Tree Species Diversity Interact with Site Properties to Drive Forest Soil Carbon. Nat Commun 2022, 13, 1097. [Google Scholar] [CrossRef]
- Chen, S.; Wang, W.; Xu, W.; Wang, Y.; Wan, H.; Chen, D.; Tang, Z.; Tang, X.; Zhou, G.; Xie, Z.; et al. Plant Diversity Enhances Productivity and Soil Carbon Storage. Proceedings of the National Academy of Sciences 2018, 115, 4027–4032. [Google Scholar] [CrossRef] [PubMed]
- Dawud, S.M.; Raulund-Rasmussen, K.; Domisch, T.; Finér, L.; Jaroszewicz, B.; Vesterdal, L. Is Tree Species Diversity or Species Identity the More Important Driver of Soil Carbon Stocks, C/N Ratio, and pH? Ecosystems 2016, 19, 645–660. [Google Scholar] [CrossRef]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.-D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive Biodiversity-Productivity Relationship Predominant in Global Forests. Science 2016, 354. [Google Scholar] [CrossRef] [PubMed]
- Scherer-Lorenzen, M.; Schulze, E.-D.; Don, A.; Schumacher, J.; Weller, E. Exploring the Functional Significance of Forest Diversity: A New Long-Term Experiment with Temperate Tree Species (BIOTREE). Perspectives in Plant Ecology, Evolution and Systematics 2007, 9, 53–70. [Google Scholar] [CrossRef]
- Jucker, T.; Avăcăriței, D.; Bărnoaiea, I.; Duduman, G.; Bouriaud, O.; Coomes, D.A. Climate Modulates the Effects of Tree Diversity on Forest Productivity. Journal of Ecology 2016, 104, 388–398. [Google Scholar] [CrossRef]
- Ratcliffe, S.; Wirth, C.; Jucker, T.; van der Plas, F.; Scherer-Lorenzen, M.; Verheyen, K.; Allan, E.; Benavides, R.; Bruelheide, H.; Ohse, B.; et al. Biodiversity and Ecosystem Functioning Relations in European Forests Depend on Environmental Context. Ecology Letters 2017, 20, 1414–1426. [Google Scholar] [CrossRef]
- Jastrow, J.D.; Amonette, J.E.; Bailey, V.L. Mechanisms Controlling Soil Carbon Turnover and Their Potential Application for Enhancing Carbon Sequestration. Climatic Change 2007, 80, 5–23. [Google Scholar] [CrossRef]
- Moujahid, L.E.; Roux, X.L.; Michalet, S.; Bellvert, F.; Weigelt, A.; Poly, F. Effect of Plant Diversity on the Diversity of Soil Organic Compounds. PLOS ONE 2017, 12, e0170494. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ding, Y.; Zhang, Y.; Wang, J.; Freedman, Z.B.; Liu, P.; Cong, W.; Wang, J.; Zang, R.; Liu, S. Evenness of Soil Organic Carbon Chemical Components Changes with Tree Species Richness, Composition and Functional Diversity across Forests in China. Global Change Biology 2023. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Song, Z.; Yang, Y.; Wang, J.; You, Y.; Zhang, X.; Shi, Z.; Nong, Y.; Ming, A.; et al. Introducing Nitrogen-Fixing Tree Species and Mixing with Pinus Massoniana Alters and Evenly Distributes Various Chemical Compositions of Soil Organic Carbon in a Planted Forest in Southern China. Forest Ecology and Management 2019, 449, 117477. [Google Scholar] [CrossRef]
- Beugnon, R.; Eisenhauer, N.; Bruelheide, H.; Davrinche, A.; Du, J.; Haider, S.; Hähn, G.; Saadani, M.; Singavarapu, B.; Sünnemann, M. Tree Diversity Effects on Litter Decomposition Are Mediated by Litterfall and Microbial Processes. Oikos 2023, e09751. [Google Scholar] [CrossRef]
- Wardle, D.A.; Nicholson, K.S. Synergistic Effects of Grassland Plant Spcies on Soil Microbial Biomass and Activity: Implications for Ecosystem-Level Effects of Enriched Plant Diversity. Functional Ecology 1996, 410–416. [Google Scholar] [CrossRef]
- Gubsch, M.; Roscher, C.; Gleixner, G.; Habekost, M.; Lipowsky, A.; Schmid, B.; SCHULZE, E.; Steinbeiss, S.; Buchmann, N. Foliar and Soil δ15N Values Reveal Increased Nitrogen Partitioning among Species in Diverse Grassland Communities. Plant, Cell & Environment 2011, 34, 895–908. [Google Scholar]
- Niklaus, P.A.; Le Roux, X.; Poly, F.; Buchmann, N.; Scherer-Lorenzen, M.; Weigelt, A.; Barnard, R.L. Plant Species Diversity Affects Soil–Atmosphere Fluxes of Methane and Nitrous Oxide. Oecologia 2016, 181, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Amelung, W.; Lehmann, J.; Kästner, M. Quantitative Assessment of Microbial Necromass Contribution to Soil Organic Matter. Global Change Biology 2019, 25, 3578–3590. [Google Scholar] [CrossRef]
- Banerjee, S.; van der Heijden, M.G.A. Soil Microbiomes and One Health. Nat Rev Microbiol 2023, 21, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of Biodiversity on Ecosystem Functioning: A Consensus of Current Knowledge. Ecological Monographs 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Isbell, F.; Calcagno, V.; Hector, A.; Connolly, J.; Harpole, W.S.; Reich, P.B.; Scherer-Lorenzen, M.; Schmid, B.; Tilman, D.; van Ruijven, J.; et al. High Plant Diversity Is Needed to Maintain Ecosystem Services. Nature 2011, 477, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Souto, X.C.; Chiapusio, G.; Pellissier, F. Relationships Between Phenolics and Soil Microorganisms in Spruce Forests: Significance for Natural Regeneration. J Chem Ecol 2000, 26, 2025–2034. [Google Scholar] [CrossRef]
- Karp, A.; Shield, I. Bioenergy from Plants and the Sustainable Yield Challenge. New Phytologist 2008, 179, 15–32. [Google Scholar] [CrossRef]
- Weih, M.; Hansson, P.-A.; Ohlsson, J.; Sandgren, M.; Schnürer, A.; Rönnberg-Wästljung, A.-C. Sustainable Production of Willow for Biofuel Use. In Achieving carbon negative bioenergy systems from plant materials; Burleigh Dodds Science Publishing, 2020; pp. 305–340.
- Weih, M. Willow. In Biofuel crops: production, physiology and genetics; CABI, 2013; pp. 415-426 ISBN 978-1- 84593-885-7.
- Weih, M.; Glynn, C.; Baum, C. Willow Short-Rotation Coppice as Model System for Exploring Ecological Theory on Biodiversity–Ecosystem Function. Diversity 2019, 11, 125. [Google Scholar] [CrossRef]
- Lindh, M.; Hoeber, S.; Weih, M.; Manzoni, S. Interactions of Nutrient and Water Availability Control Growth and Diversity Effects in a Salix Two-Species Mixture. Ecohydrology 2022, 15, e2401. [Google Scholar] [CrossRef]
- Weih, M.; Nordh, N.-E.; Manzoni, S.; Hoeber, S. Functional Traits of Individual Varieties as Determinants of Growth and Nitrogen Use Patterns in Mixed Stands of Willow (Salix Spp.). Forest Ecology and Management 2021, 479, 118605. [Google Scholar] [CrossRef]
- Baum, C.; Amm, T.; Kahle, P.; Weih, M. Fertilization Effects on Soil Ecology Strongly Depend on the Genotype in a Willow (Salix Spp.) Plantation. Forest Ecol Manag 2020, 466. [Google Scholar] [CrossRef]
- Baum, C.; Leinweber, P.; Weih, M.; Lamersdorf, N.; Dimitriou, I. Effects of Short Rotation Coppice with Willows and Poplar on Soil Ecology. Agriculture and Forestry Research 2009, 3, 183–196. [Google Scholar]
- Kalita, S.; Potter, H.K.; Weih, M.; Baum, C.; Nordberg, Å.; Hansson, P.-A. Soil Carbon Modelling in Salix Biomass Plantations: Variety Determines Carbon Sequestration and Climate Impacts. Forests 2021, 12, 1529. [Google Scholar] [CrossRef]
- Hoeber, S.; Fransson, P.; Prieto-Ruiz, I.; Manzoni, S.; Weih, M. Two Salix Genotypes Differ in Productivity and Nitrogen Economy When Grown in Monoculture and Mixture. Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Verheyen, K.; Vanhellemont, M.; Auge, H.; Baeten, L.; Baraloto, C.; Barsoum, N.; Bilodeau-Gauthier, S.; Bruelheide, H.; Castagneyrol, B.; Godbold, D.; et al. Contributions of a Global Network of Tree Diversity Experiments to Sustainable Forest Plantations. Ambio 2016, 45, 29–41. [Google Scholar] [CrossRef] [PubMed]
- ISRIC, F. World Reference Base for Soil Resources. World soil resources reports 1998, 84. [Google Scholar]
- Baum, C.; Hrynkiewicz, K.; Szymańska, S.; Vitow, N.; Hoeber, S.; Fransson, P.M.A.; Weih, M. Mixture of Salix Genotypes Promotes Root Colonization With Dark Septate Endophytes and Changes P Cycling in the Mycorrhizosphere. Frontiers in Microbiology 2018, 9. [Google Scholar] [CrossRef]
- Weih, M.; Nordh, N.-E. Characterising Willows for Biomass and Phytoremediation: Growth, Nitrogen and Water Use of 14 Willow Clones under Different Irrigation and Fertilisation Regimes. Biomass and Bioenergy 2002, 23, 397–413. [Google Scholar] [CrossRef]
- Ågren, G.I.; Weih, M. Plant Stoichiometry at Different Scales: Element Concentration Patterns Reflect Environment More than Genotype. New Phytologist 2012, 194, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Hoeber, S.; Fransson, P.; Weih, M.; Manzoni, S. Leaf Litter Quality Coupled to Salix Variety Drives Litter Decomposition More than Stand Diversity or Climate. Plant Soil 2020, 453, 313–328. [Google Scholar] [CrossRef]
- An, H.; Liu, Y.; Zhao, X.; Huang, Q.; Yuan, S.; Yang, X.; Dong, J. Characterization of Cadmium-Resistant Endophytic Fungi from Salix Variegata Franch. in Three Gorges Reservoir Region, China. Microbiological Research 2015, 176, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Hrynkiewicz, K.; Ciesielska, A.; Haug, I.; Baum, C. Ectomycorrhiza Formation and Willow Growth Promotion as Affected by Associated Bacteria: Role of Microbial Metabolites and Use of C Sources. Biology and Fertility of Soils 2010, 46, 139–150. [Google Scholar] [CrossRef]
- Püttsepp, Ü.; Rosling, A.; Taylor, A.F.S. Ectomycorrhizal Fungal Communities Associated with Salix Viminalis L. and S. Dasyclados Wimm. Clones in a Short-Rotation Forestry Plantation. Forest Ecology and Management 2004, 196, 413–424. [Google Scholar] [CrossRef]
- Hoeber, S.; Arranz, C.; Nordh, N.E.; Baum, C.; Low, M.; Nock, C.; Scherer-Lorenzen, M.; Weih, M. Genotype Identity Has a More Important Influence than Genotype Diversity on Shoot Biomass Productivity in Willow Short-Rotation Coppices. Gcb Bioenergy 2018, 10, 534–547. [Google Scholar] [CrossRef]
- Cunniff, J.; Purdy, S.J.; Barraclough, T.J.P.; Castle, M.; Maddison, A.L.; Jones, L.E.; Shield, I.F.; Gregory, A.S.; Karp, A. High Yielding Biomass Genotypes of Willow (Salix Spp.) Show Differences in below Ground Biomass Allocation. Biomass and Bioenergy 2015, 80, 114–127. [Google Scholar] [CrossRef] [PubMed]
- Rytter, R.-M.; Hansson, A.-C. Seasonal Amount, Growth and Depth Distribution of Fine Roots in an Irrigated and Fertilized Salix Viminalis L. Plantation. Biomass and Bioenergy 1996, 11, 129–137. [Google Scholar] [CrossRef]
- VDLUFA Methode A 6.2.1.2 - Bestimmung von Phosphor Und Kalium Im Doppellactat (DL)-Auszug. In VDLUFA-Methodenbuch, Band I, Die Untersuchung von Boden; Verband Deutscher Landwirtschaftlicher Untersuchungs- und, Forschungsanstalten, Ed.; VDLUFA-Verlag: Darmstadt, Germany, 2002. [Google Scholar]
- Hylander, L.D.; Svensson, H.; Simán, G. Different Methods for Determination of Plant Available Soil Phosphorus. Communications in Soil Science and Plant Analysis 1996, 27, 1501–1512. [Google Scholar] [CrossRef]
- Parker, T.C.; Clemmensen, K.E.; Friggens, N.L.; Hartley, I.P.; Johnson, D.; Lindahl, B.D.; Olofsson, J.; Siewert, M.B.; Street, L.E.; Subke, J.-A.; et al. Rhizosphere Allocation by Canopy-Forming Species Dominates Soil CO2 Efflux in a Subarctic Landscape. New Phytologist 2020, 227, 1818–1830. [Google Scholar] [CrossRef] [PubMed]
- Oades, J.M. Soil Organic Matter and Structural Stability: Mechanisms and Implications for Management. Plant Soil 1984, 76, 319–337. [Google Scholar] [CrossRef]
- Hempfling, R.; Zech, W.; Schulten, H.-R. CHEMICAL COMPOSITION OF THE ORGANIC MATTER IN FOREST SOILS: 2. MODER PROFILE. Soil Science 1988, 146, 262–276. [Google Scholar] [CrossRef]
- Leinweber, P.; Kruse, J.; Baum, C.; Arcand, M.; Knight, J.D.; Farrell, R.; Eckhardt, K.-U.; Kiersch, K.; Jandl, G. Chapter Two - Advances in Understanding Organic Nitrogen Chemistry in Soils Using State-of-the-Art Analytical Techniques. In Advances in Agronomy; Sparks, D.L., Ed.; Advances in Agronomy; Academic Press, 2013; Vol. 119, pp. 83–151.
- Leinweber, P.; Blumenstein, O.; Schulten, H.-R. Organic Matter Composition in Sewage Farm Soils: Investigations by 13C-NMR and Pyrolysis-Field Ionization Mass Spectrometry. European Journal of Soil Science 1996, 47, 71–80. [Google Scholar] [CrossRef]
- Schnitzer, M.; Schulten, H.R. The Analysis of Soil Organic Matter by Pyrolysis-Field Ionization Mass Spectrometry. Soil Science Society of America Journal 1992, 56, 1811–1817. [Google Scholar] [CrossRef]
- Core, R. TEAM, 2017. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Online: https://www. r-project. org, 2022. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘Vegan. ’ Community ecology package, version 2013, 2, 1–295. [Google Scholar]
- Keithley, R.B.; Mark Wightman, R.; Heien, M.L. Multivariate Concentration Determination Using Principal Component Regression with Residual Analysis. TrAC Trends in Analytical Chemistry 2009, 28, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Baumann, K.; Eckhardt, K.-U.; Schöning, I.; Schrumpf, M.; Leinweber, P. Clay Fraction Properties and Grassland Management Imprint on Soil Organic Matter Composition and Stability at Molecular Level. Soil Use and Management 2022, 38, 1578–1596. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. Nonlinear Mixed-Effects Models. R package version 2012, 3, 1–89. [Google Scholar]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R package version 2018, 1, 3. [Google Scholar]
- Castellano, M.J.; Mueller, K.E.; Olk, D.C.; Sawyer, J.E.; Six, J. Integrating Plant Litter Quality, Soil Organic Matter Stabilization, and the Carbon Saturation Concept. Global change biology 2015, 21, 3200–3209. [Google Scholar] [CrossRef] [PubMed]
- Baumann, K.; Dignac, M.-F.; Rumpel, C.; Bardoux, G.; Sarr, A.; Steffens, M.; Maron, P.-A. Soil Microbial Diversity Affects Soil Organic Matter Decomposition in a Silty Grassland Soil. Biogeochemistry 2013, 114, 201–212. [Google Scholar] [CrossRef]
- Carney, K.M.; Matson, P.A. The Influence of Tropical Plant Diversity and Composition on Soil Microbial Communities. Microb Ecol 2006, 52, 226–238. [Google Scholar] [CrossRef]
- Stockmann, U.; Adams, M.A.; Crawford, J.W.; Field, D.J.; Henakaarchchi, N.; Jenkins, M.; Minasny, B.; McBratney, A.B.; Courcelles, V. de R. de; Singh, K.; et al. The Knowns, Known Unknowns and Unknowns of Sequestration of Soil Organic Carbon. Agriculture, Ecosystems & Environment 2013, 164, 80–99. [Google Scholar] [CrossRef]
- Cheshire, M.V.; Christensen, B.T.; Sørensen, L.H. Labelled and Native Sugars in Particle-Size Fractions from Soils Incubated with 14C Straw for 6 to 18 Years. Journal of Soil Science 1990, 41, 29–39. [Google Scholar] [CrossRef]
- Angst, G.; Heinrich, L.; Kögel-Knabner, I.; Mueller, C.W. The Fate of Cutin and Suberin of Decaying Leaves, Needles and Roots – Inferences from the Initial Decomposition of Bound Fatty Acids. Organic Geochemistry 2016, 95, 81–92. [Google Scholar] [CrossRef]
- Yang, S.; Jansen, B.; Absalah, S.; Kalbitz, K.; Chunga Castro, F.O.; Cammeraat, E.L.H. Soil Organic Carbon Content and Mineralization Controlled by the Composition, Origin and Molecular Diversity of Organic Matter: A Study in Tropical Alpine Grasslands. Soil and Tillage Research 2022, 215, 105203. [Google Scholar] [CrossRef]
- Gadgil, R.L.; Gadgil, P. Mycorrhiza and Litter Decomposition. Nature 1971, 233, 133–133. [Google Scholar] [CrossRef] [PubMed]
- Cremer, M.; Kern, N.V.; Prietzel, J. Soil Organic Carbon and Nitrogen Stocks under Pure and Mixed Stands of European Beech, Douglas Fir and Norway Spruce. Forest Ecology and Management 2016, 367, 30–40. [Google Scholar] [CrossRef]
- Waring, B.G.; Smith, K.R.; Belluau, M.; Khlifa, R.; Messier, C.; Munson, A.; Paquette, A. Soil Carbon Pools Are Affected by Species Identity and Productivity in a Tree Common Garden Experiment. Frontiers in Forests and Global Change 2022, 5. [Google Scholar] [CrossRef]
- Wiesmeier, M.; Prietzel, J.; Barthold, F.; Spörlein, P.; Geuß, U.; Hangen, E.; Reischl, A.; Schilling, B.; von Lützow, M.; Kögel-Knabner, I. Storage and Drivers of Organic Carbon in Forest Soils of Southeast Germany (Bavaria) – Implications for Carbon Sequestration. Forest Ecology and Management 2013, 295, 162–172. [Google Scholar] [CrossRef]
- Koczorski, P.; Furtado, B.U.; Gołębiewski, M.; Hulisz, P.; Baum, C.; Weih, M.; Hrynkiewicz, K. The Effects of Host Plant Genotype and Environmental Conditions on Fungal Community Composition and Phosphorus Solubilization in Willow Short Rotation Coppice. Frontiers in Plant Science 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Barry, K.E.; Weigelt, A.; van Ruijven, J.; de Kroon, H.; Ebeling, A.; Eisenhauer, N.; Gessler, A.; Ravenek, J.M.; Scherer-Lorenzen, M.; Oram, N.J.; et al. Chapter Two - Above- and Belowground Overyielding Are Related at the Community and Species Level in a Grassland Biodiversity Experiment. In Advances in Ecological Research; Eisenhauer, N., Bohan, D.A., Dumbrell, A.J., Eds.; Mechanisms underlying the relationship between biodiversity and ecosystem function; Academic Press, 2019; Vol. 61, pp. 55–89.
- Hall, S.J.; Huang, W.; Timokhin, V.I.; Hammel, K.E. Lignin Lags, Leads, or Limits the Decomposition of Litter and Soil Organic Carbon. Ecology 2020, 101, e03113. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Soil C accumulation: estimated marginal means of topsoil (0-10 cm depth) C accumulation rates for the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture at sites Rostock and Uppsala. Site-specific bulk densities 1.3 and 1.4 g cm3 used in calculations of C stocks in Rostock and Uppsala respectively. Different uppercase letters indicate significant pairwise differences between willow varieties within each site (A-B; p < 0.05). Different lowercase letters indicate significant pairwise differences for each willow variety between sites (a-b; p < 0.05). Standard errors are shown as error bars.
Figure 1.
Soil C accumulation: estimated marginal means of topsoil (0-10 cm depth) C accumulation rates for the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture at sites Rostock and Uppsala. Site-specific bulk densities 1.3 and 1.4 g cm3 used in calculations of C stocks in Rostock and Uppsala respectively. Different uppercase letters indicate significant pairwise differences between willow varieties within each site (A-B; p < 0.05). Different lowercase letters indicate significant pairwise differences for each willow variety between sites (a-b; p < 0.05). Standard errors are shown as error bars.
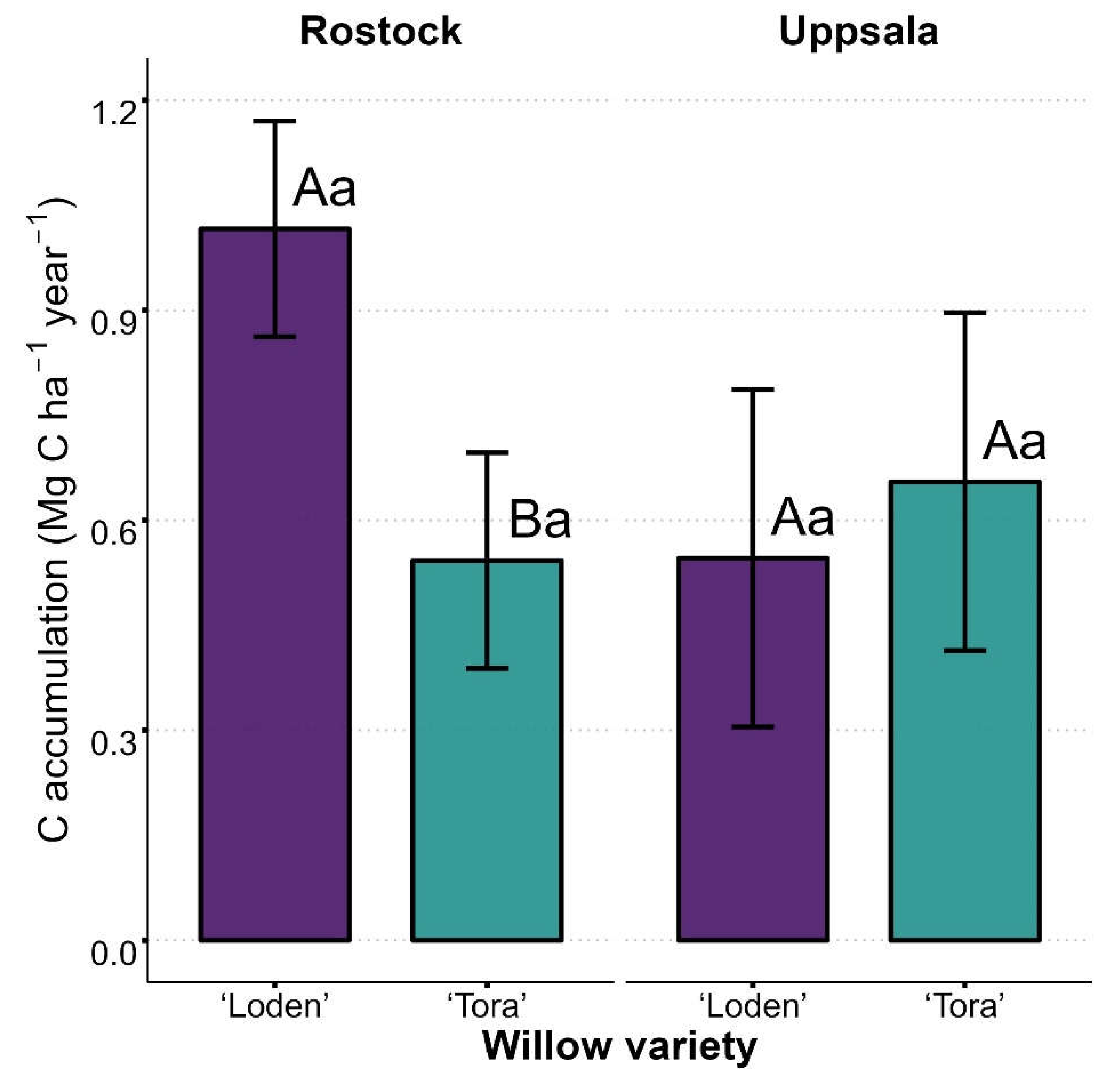
Figure 3.
SOM thermal stability: thermograms showing ions volatilized under pyrolysis (0-650 °C) of nine compound classes; CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER, FATTY in Py-FI mass spectra (for abbreviations, see Table 2) in topsoil (0-10 cm depth) for the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture and mixture at the Rostock site. Higher values of relative ion intensity indicate a greater abundance of specific compounds in the pyrolyzed SOM. The temperature at which compounds volatilize reflects their thermal stability, with ions volatilized over 400 °C being regarded as stable and ions volatilized under 400 °C being regarded as labile.
Figure 3.
SOM thermal stability: thermograms showing ions volatilized under pyrolysis (0-650 °C) of nine compound classes; CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER, FATTY in Py-FI mass spectra (for abbreviations, see Table 2) in topsoil (0-10 cm depth) for the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture and mixture at the Rostock site. Higher values of relative ion intensity indicate a greater abundance of specific compounds in the pyrolyzed SOM. The temperature at which compounds volatilize reflects their thermal stability, with ions volatilized over 400 °C being regarded as stable and ions volatilized under 400 °C being regarded as labile.
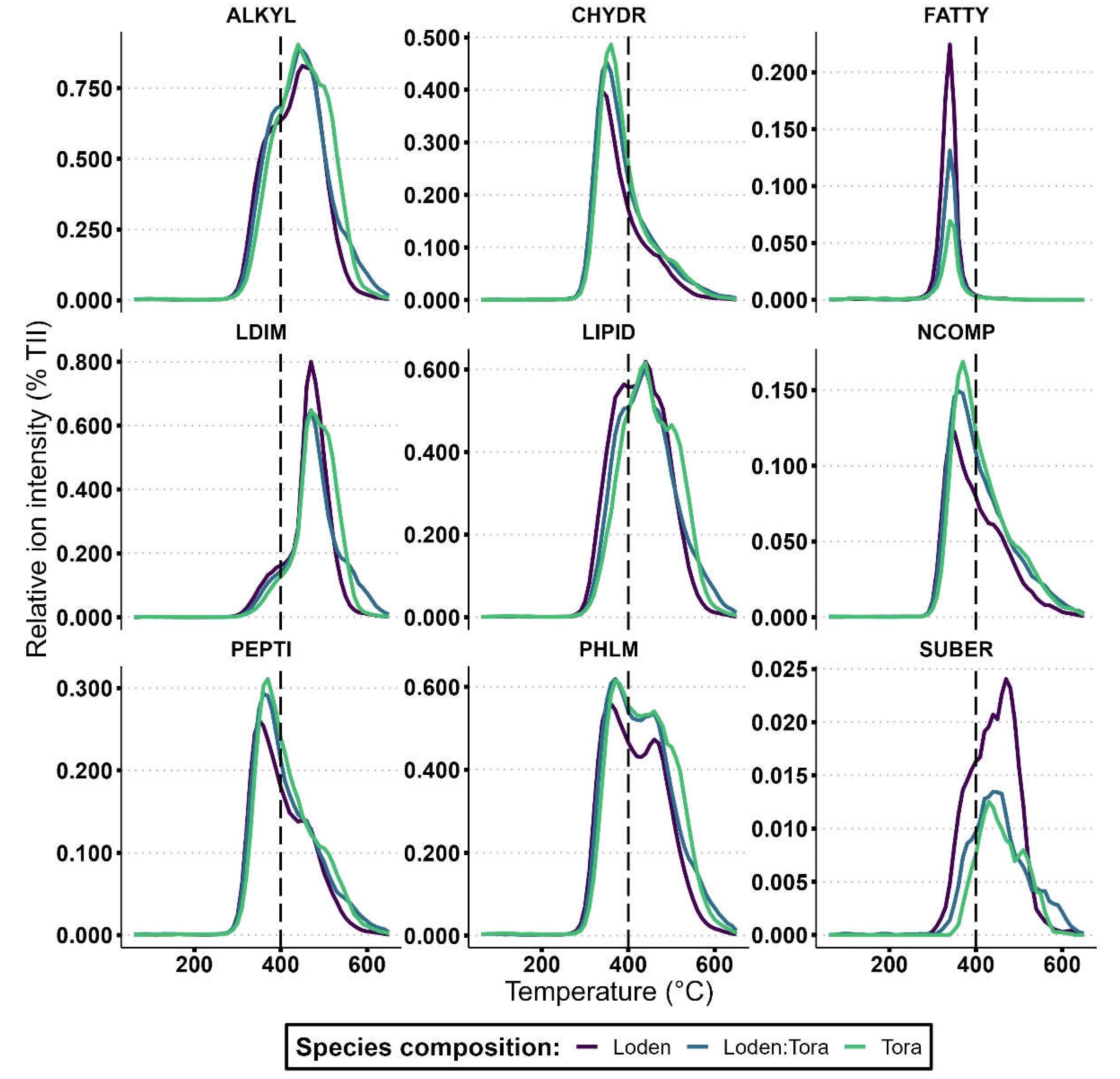
Figure 4.
SOM thermal stability: thermograms showing ions volatilized under pyrolysis (0-650 °C) of nine compound classes; CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER, FATTY in Py-FI mass spectra (for abbreviations, see Table 2) in topsoil (0-10 cm depth) for the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture and mixture at the Uppsala site. Higher values of relative ion intensity indicate a greater abundance of specific compounds in the pyrolyzed SOM. The temperature at which compounds volatilize reflects their thermal stability, with ions volatilized over 400 °C being regarded as stable and ions volatilized under 400 °C being regarded as labile.
Figure 4.
SOM thermal stability: thermograms showing ions volatilized under pyrolysis (0-650 °C) of nine compound classes; CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER, FATTY in Py-FI mass spectra (for abbreviations, see Table 2) in topsoil (0-10 cm depth) for the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture and mixture at the Uppsala site. Higher values of relative ion intensity indicate a greater abundance of specific compounds in the pyrolyzed SOM. The temperature at which compounds volatilize reflects their thermal stability, with ions volatilized over 400 °C being regarded as stable and ions volatilized under 400 °C being regarded as labile.
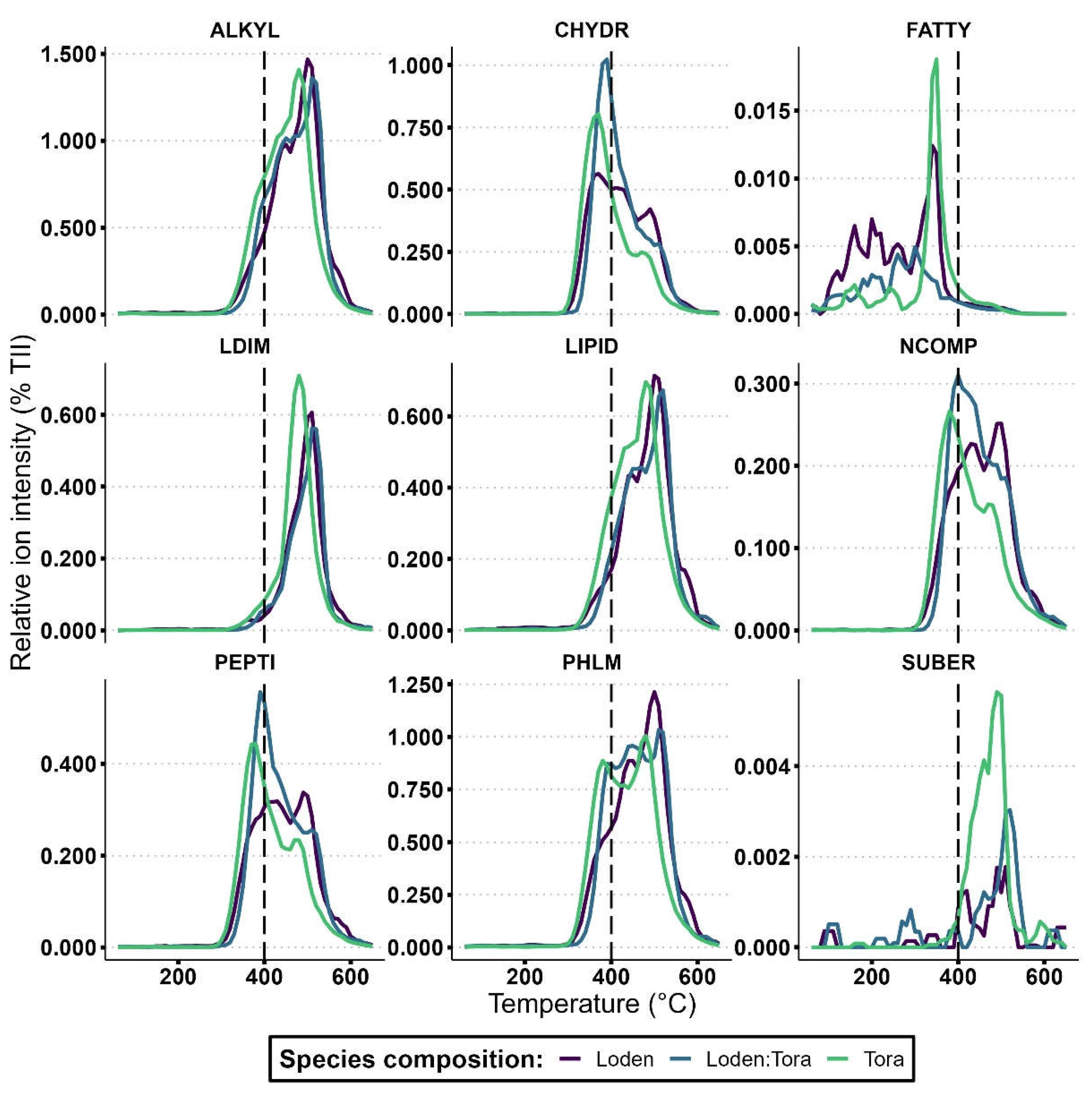
Figure 5.
Soil C accumulation: estimated marginal means of expected and observed topsoil (0-10 cm depth) C accumulation rates at sites Rostock and Uppsala. Site-specific bulk densities 1.3 and 1.4 g cm3 used in calculations of C stocks in Rostock and Uppsala, respectively. Expected values represent the basal area weighted and averaged mean values of the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture and observed values represent the mean values of ‘Loden’ and ‘Tora’ grown in mixture (see Equation (3)). Different uppercase letters indicate significant pairwise differences between expected and observed mixture values within each site (A-B; p < 0.05). Different lowercase letters indicate significant pairwise differences for observed mixture values between sites (a-b; p < 0.05). Standard errors are shown as error bars.
Figure 5.
Soil C accumulation: estimated marginal means of expected and observed topsoil (0-10 cm depth) C accumulation rates at sites Rostock and Uppsala. Site-specific bulk densities 1.3 and 1.4 g cm3 used in calculations of C stocks in Rostock and Uppsala, respectively. Expected values represent the basal area weighted and averaged mean values of the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture and observed values represent the mean values of ‘Loden’ and ‘Tora’ grown in mixture (see Equation (3)). Different uppercase letters indicate significant pairwise differences between expected and observed mixture values within each site (A-B; p < 0.05). Different lowercase letters indicate significant pairwise differences for observed mixture values between sites (a-b; p < 0.05). Standard errors are shown as error bars.
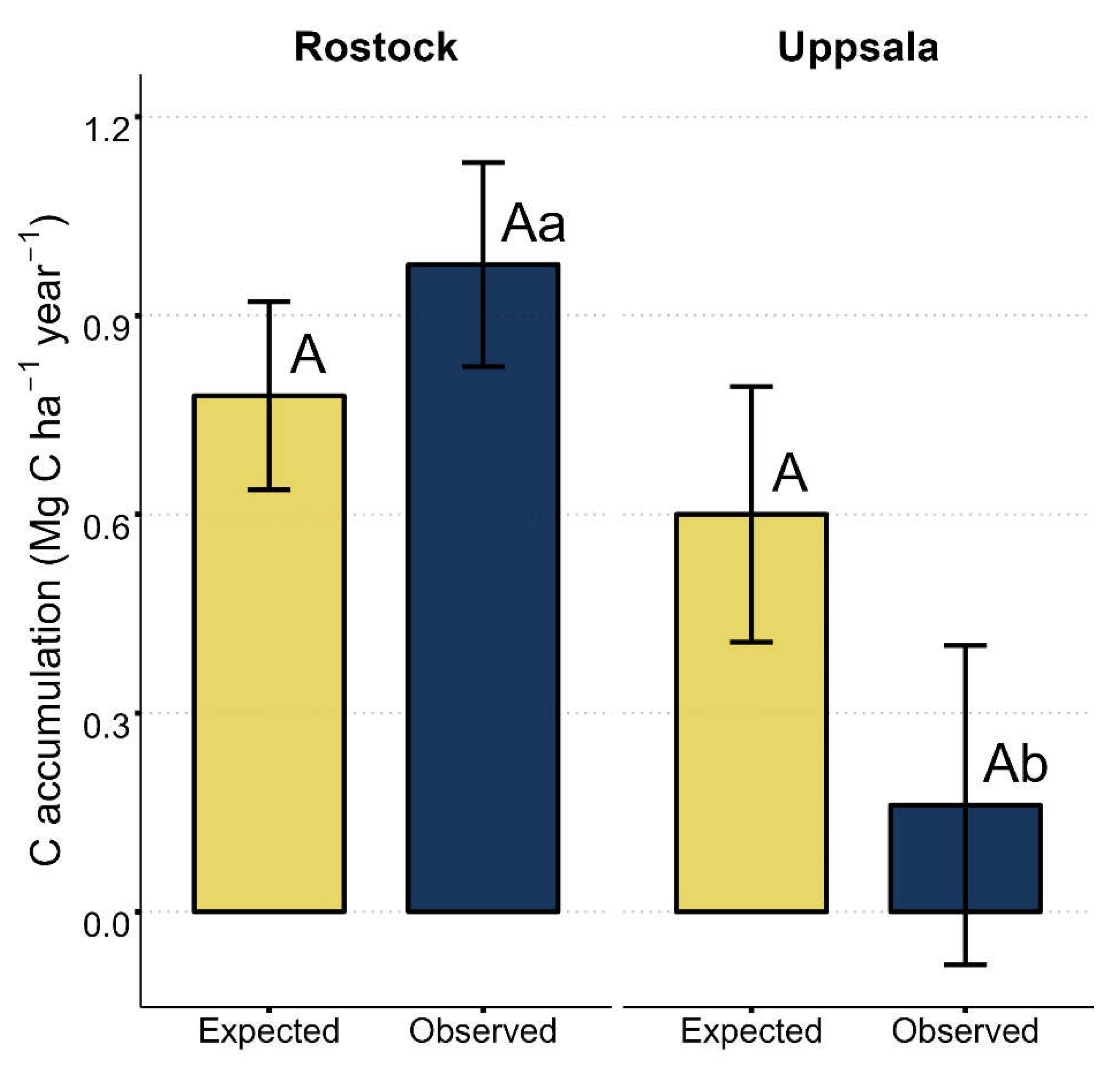

Table 1.
Site characteristics: soil group [56], topsoil (0-10 cm depth) properties measured in 2021 and climatic conditions during time since establishment at sites Rostock and Uppsala.
Table 1.
Site characteristics: soil group [56], topsoil (0-10 cm depth) properties measured in 2021 and climatic conditions during time since establishment at sites Rostock and Uppsala.
| Site | Soil group | pH | Bulk density [g cm-3] |
Clay content [%] |
MAT [° C] |
MAP [mg g-1] |
|---|---|---|---|---|---|---|
| Uppsala | Vertic Cambisol | 5.2 | 1.4 | 52 | 7.53 | 500 |
| Rostock | Stagnic Cambisol | 6.2 | 1.3 | 5 | 10.35 | 730 |
Table 2.
Pyrolysis-field ionization mass spectrometry (Py-FIMS) parameters and compound classes with associated biomolecules, as assigned by; [21,71,72,73,74].
| Py-FIMS parameters | Explanations |
|---|---|
| Hexoses:pentoses | Ratio of microbial- to plant-derived sugars |
| TII | Total ion intensity (106 counts mg-1) |
| VM | Volatile matter in % (weightbefore pyrolysis/weightafter pyrolysis) |
| CHYDR | Carbohydrates with pentose and hexose subunits |
| PHLM | Phenols and lignin monomers |
| LDIM | Lignin dimers |
| LIPID | Lipids, alkanes, alkenes, bound fatty acids, and alkylmonoesters |
| ALKYL | Alkylaromatics |
| NCOMP | Mainly heterocyclic N-containing compounds |
| PEPTI | Peptides (amino acids, peptides and aminosugars) |
| SUBER | Suberin |
| FATTY | Free fatty acids C16-C34 |
Table 3.
Soil chemical characteristics and soil respiration: estimated marginal means of topsoil (0-10 cm depth) soil C:N ratios, nutrient concentrations of C, N, Kdl, Mgdl and Pdl as well as soil CO2 respiration for varieties ‘Loden’ and ‘Tora’ grown in monoculture and mixture at sites Rostock and Uppsala. Different letters indicate significant differences across sites (x-y) and within each site (a-b; p < 0.05, Tukey’s post hoc test).
Table 3.
Soil chemical characteristics and soil respiration: estimated marginal means of topsoil (0-10 cm depth) soil C:N ratios, nutrient concentrations of C, N, Kdl, Mgdl and Pdl as well as soil CO2 respiration for varieties ‘Loden’ and ‘Tora’ grown in monoculture and mixture at sites Rostock and Uppsala. Different letters indicate significant differences across sites (x-y) and within each site (a-b; p < 0.05, Tukey’s post hoc test).
| Site | Variety composition |
C:N | C [%] |
N [%] |
Kdl~~·[mg g-1] | Mgdl [mg g-1] |
Pdl [mg g-1] |
CO2 [g C m-2 h-1] |
|---|---|---|---|---|---|---|---|---|
| Rostock | ‘Loden’ | 10.80 xa | 1.26 ya | 0.12 ya | 9.99 ya | 20.42 ya | 4.40 xa | - |
| ‘Loden’:‘Tora’ | 10.63 xa | 1.22 ya | 0.11 ya | 10.22 ya | 21.06 ya | 4.40 xa | - | |
| ‘Tora’ | 10.22 xa | 1.01 ya | 0.10 ya | 10.71 ya | 21.96 ya | 4.02 xa | - | |
| Uppsala | ‘Loden’ | 10.69 xa | 1.89 xa | 0.18 xa | 22.23 xa | 32.10 xa | 4.27 xa | 0.493 a |
| ‘Loden’:‘Tora’ | 10.58 xa | 1.58 xa | 0.16 xa | 23.62 xa | 27.94 xa | 5.04 xa | 0.453 a | |
| ‘Tora’ | 10.85 xa | 1.90 xa | 0.17 xa | 19.89 xa | 28.76 xa | 3.60 xa | 0.461 a |
Table 4.
Py-FIMS parameters: estimated marginal means of TII (total ion intensity; 106 counts mg-1) and total thermal stability of bulk SOM (ions volatilized > 400 °C / ions volatilized 50-650 °C), VM (volatile matter), H’ (SOM chemical diversity) and hexoses:pentoses (ratio of microbial- to plant-derived sugars) by site and by variety composition. Different letters indicate significant differences across sites (x-y) and within each site (a-b; p < 0.05, Tukey’s post hoc test).
Table 4.
Py-FIMS parameters: estimated marginal means of TII (total ion intensity; 106 counts mg-1) and total thermal stability of bulk SOM (ions volatilized > 400 °C / ions volatilized 50-650 °C), VM (volatile matter), H’ (SOM chemical diversity) and hexoses:pentoses (ratio of microbial- to plant-derived sugars) by site and by variety composition. Different letters indicate significant differences across sites (x-y) and within each site (a-b; p < 0.05, Tukey’s post hoc test).
| Site | Variety composition |
TII [106 counts mg-1] |
Total thermal stability |
Chemical diversity [H’] |
Hexoses: pentoses |
Volatile matter |
|---|---|---|---|---|---|---|
| Rostock | ‘Loden’ | 53.3 xa | 0.68 xa | 6.71 ya | 5.79 xa | 2.83 xa |
| ‘Loden’:‘Tora’ | 39.1 xa | 0.72 xa | 6.30 ya | 5.73 xab | 2.92 xa | |
| ‘Tora’ | 34.8 xa | 0.77 xa | 5.82 ya | 5.67 xb | 2.75 xa | |
| Uppsala | ‘Loden’ | 14.8 ya | 0.84 xa | 14.66 xa | 5.47 ya | 1.89 yb |
| ‘Loden’:‘Tora’ | 14.2 ya | 0.82 xb | 9.69 xb | 5.45 ya | 1.91 yb | |
| ‘Tora’ | 24.7 ya | 0.83 xab | 13.36 xa | 5.53 ya | 2.28 ya |
Table 5.
SOM chemical composition: estimated marginal means (± SE) of relative ion intensities (% TII) of nine compound classes; CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER, FATTY in Py-FI mass spectra (for abbreviations, see Table 2) in topsoil (0-10 cm depth) for the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture at sites Rostock and Uppsala. Different uppercase letters indicate significant pairwise differences between willow varieties within each site (A-B; p < 0.05). Different lowercase letters indicate significant pairwise differences for each willow variety between sites (a-b; p < 0.05).
Table 5.
SOM chemical composition: estimated marginal means (± SE) of relative ion intensities (% TII) of nine compound classes; CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER, FATTY in Py-FI mass spectra (for abbreviations, see Table 2) in topsoil (0-10 cm depth) for the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture at sites Rostock and Uppsala. Different uppercase letters indicate significant pairwise differences between willow varieties within each site (A-B; p < 0.05). Different lowercase letters indicate significant pairwise differences for each willow variety between sites (a-b; p < 0.05).
| Compound classes | Rostock | Uppsala | |||
|---|---|---|---|---|---|
| ‘Loden’ | ‘Tora’ | ‘Loden’ | ‘Tora’ | ||
| CHYDR | 4.0 (0.19) Bb | 5.0 (0. 19) Ab | 9.5 (0.82) Aa | 8.8 (0.82) Aa | |
| PHLM | 9.9 (0.36) Bb | 11.7 (0.36) Ab | 17.2 (0.82) Aa | 16.2 (0.82) Aa | |
| LDIM | 6.8 (0.64) Aa | 7.1 (0.64) Aa | 4.2 (0.59) Bb | 5.9 (0.59) Aa | |
| LIPID | 10.3 (0.29) Aa | 9.9 (0.29) Aa | 8.4 (0.24) Ab | 8.8 (0.24) Ab | |
| ALKYL | 13.3 (0.49) Ab | 14.9 (0.49) Ab | 17.8 (0.38) Aa | 17.9 (0.38) Aa | |
| NCOMP | 1.6 (0. 10) Bb | 2.1 (0.10) Ab | 4.1 (0.36) Aa | 3.6 (0. 36) Aa | |
| PEPTI | 3.6 (0.24) Bb | 4.1 (0.24) Ab | 6.4 (0.21) Aa | 5.5 (0.21) Ba | |
| SUBER | 0.33 (0.03) Aa | 0.16 (0.03) Ba | 0.02 (0.01) Ab | 0.05 (0.01) Ab | |
| FATTY | 0.87 (0.13) Aa | 0.31 (0.13) Ba | 0.17 (0.07) Ab | 0.04 (0.07) Ba | |
Table 6.
SOM thermal stability: estimated marginal means (± SE) of thermal stability (ions volatilized > 400 °C / ions volatilized 50-650 °C) of nine compound classes; CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER, FATTY in Py-FI mass spectra (for abbreviations, see Table 2) in topsoil (0-10 cm depth) for the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture at sites Rostock and Uppsala. Different lowercase letters indicate significant pairwise differences for each willow variety between sites (a-b; p < 0.05).
Table 6.
SOM thermal stability: estimated marginal means (± SE) of thermal stability (ions volatilized > 400 °C / ions volatilized 50-650 °C) of nine compound classes; CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER, FATTY in Py-FI mass spectra (for abbreviations, see Table 2) in topsoil (0-10 cm depth) for the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture at sites Rostock and Uppsala. Different lowercase letters indicate significant pairwise differences for each willow variety between sites (a-b; p < 0.05).
| Compound classes | Rostock | Uppsala | |||
|---|---|---|---|---|---|
| ‘Loden’ | ‘Tora’ | ‘Loden’ | ‘Tora’ | ||
| CHYDR | 0.32 (0.08) Aa | 0.36 (0.08) Aa | 0.60 (0.08) Aa | 0.51 (0.08) Ba | |
| PHLM | 0.58 (0.06) Ab | 0.67 (0.06) Aa | 0.82 (0.05) Aa | 0.77 (0.05) Ba | |
| LDIM | 0.88 (0.02) Ba | 0.93 (0.02) Aa | 0.95 (0.02) Aa | 0.97 (0.02) Aa | |
| LIPID | 0.67 (0.04) Bb | 0.80 (0.04) Aa | 0.90 (0.03) Aa | 0.90 (0.03) Aa | |
| ALKYL | 0.70 (0.04) Bb | 0.80 (0.04) Aa | 0.88 (0.04) Aa | 0.87 (0.04) Ba | |
| NCOMP | 0.46 (0.07) Ab | 0.53 (0.07) Aa | 0.80 (0.06) Aa | 0.68 (0.06) Ba | |
| PEPTI | 0.48 (0.07) Aa | 0.54 (0.07) Aa | 0.72 (0.06) Aa | 0.64 (0.06) Ba | |
| SUBER | 0.76 (0.06) Ba | 0.90 (0.06) Aa | 0.88 (0.05) Ba | 0.97 (0.05) Aa | |
| FATTY | 0.02 (0.01) Ba | 0.07 (0.01) Aa | 0.10 (0.07) Aa | 0.12 (0.07) Aa | |
Table 7.
SOM chemical composition: estimated marginal means (± SE) of expected and observed relative ion intensities (% TII) of nine compound classes: CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER and FATTY in Py-FI mass spectra (for abbreviations, see Table 2), as well as net diversity effect (NDE; Equation (3)) in topsoil (0-10 cm depth) at sites Rostock and Uppsala. Expected values represent the basal area weighted and averaged mean values of the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture and observed values represent the mean values of ‘Loden’ and ‘Tora’ grown in mixture (see Equation (3)). Asterisks indicate significant pairwise differences between expected and observed values within each site (*** = p < 0.001, ** = p < 0.01, * = p < 0.05, n.s. = not significant).
Table 7.
SOM chemical composition: estimated marginal means (± SE) of expected and observed relative ion intensities (% TII) of nine compound classes: CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER and FATTY in Py-FI mass spectra (for abbreviations, see Table 2), as well as net diversity effect (NDE; Equation (3)) in topsoil (0-10 cm depth) at sites Rostock and Uppsala. Expected values represent the basal area weighted and averaged mean values of the Salix varieties ‘Loden’ and ‘Tora’ grown in monoculture and observed values represent the mean values of ‘Loden’ and ‘Tora’ grown in mixture (see Equation (3)). Asterisks indicate significant pairwise differences between expected and observed values within each site (*** = p < 0.001, ** = p < 0.01, * = p < 0.05, n.s. = not significant).
| Compound classes |
Rostock | Uppsala | |||||
|---|---|---|---|---|---|---|---|
| Expected | Observed | NDE | Expected | Observed | NDE | ||
| CHYDR | 4.6 (0.14) | 5.0 (0.19) | 9.2% n.s. | 9.0 (0.62) | 10.6 (0.82) | 17.1% n.s. | |
| PHLM | 11.0 (0.26) | 11.6 (0.36) | 5.8% n.s. | 16.5 (0.63) | 17.4 (0.82) | 5.7% n.s. | |
| LDIM | 7.0 (0.62) | 6.6 (0.64) | -5.6% n.s. | 5.4 (0.59) | 4.4 (0.59) | -18.3% *** | |
| LIPID | 10.1 (0.23) | 9.9 (0.29) | -1.9% n.s. | 8.6 (0.20) | 8.0 (0.24) | -7.2% n.s. | |
| ALKYL | 14.2 (0.35) | 14.6 (0.49) | 2.8% n.s. | 17.8 (0.29) | 17.6 (0.38) | -1.0% n.s. | |
| NCOMP | 1.9 (0.07) | 2.1 (0.25) | 10.2% n.s. | 3.8 (0.27) | 4.5 (0.23) | 20.1% n.s. | |
| PEPTI | 3.9 (0.22) | 4.1 (0.24) | 4.0% n.s. | 5.8 (0.20) | 6.8 (0.21) | 18.2% *** | |
| SUBER | 0.23 (0.03) | 0.20 (0.03) | -14.8% n.s. | 0.04 (0.01) | 0.03 (0.01) | -18.0% n.s. | |
| FATTY | 0.55 (0.10) | 0.52 (0.13) | -6.5% n.s. | 0.08 (0.07) | 0.13 (0.07) | 59.4% n.s. | |
Table 8.
SOM thermal stability: estimated marginal means (± SE) of expected and observed thermal stability (ions volatilized > 400 °C / > 0 °C) of nine compound classes: CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER and FATTY in Py-FI mass spectra (for abbreviations, see Table 2), as well as the net diversity effect (NDE; Equation (3)) in topsoil (0-10 cm depth) at sites Rostock and Uppsala. Expected values represent the basal area weighted and averaged mean values of varieties ‘Loden’ and ‘Tora’ grown in monoculture and observed values represent the mean values of ‘Loden’ and ‘Tora’ grown in mixture (see Equation (3)). Asterisks indicate significant pairwise differences between expected and observed values within each site (*** = p < 0.001, ** = p < 0.01, * = p < 0.05, n.s. = not significant).
Table 8.
SOM thermal stability: estimated marginal means (± SE) of expected and observed thermal stability (ions volatilized > 400 °C / > 0 °C) of nine compound classes: CHYDR, PHLM, LDIM, LIPID, ALKYL, NCOMP, PEPTI, SUBER and FATTY in Py-FI mass spectra (for abbreviations, see Table 2), as well as the net diversity effect (NDE; Equation (3)) in topsoil (0-10 cm depth) at sites Rostock and Uppsala. Expected values represent the basal area weighted and averaged mean values of varieties ‘Loden’ and ‘Tora’ grown in monoculture and observed values represent the mean values of ‘Loden’ and ‘Tora’ grown in mixture (see Equation (3)). Asterisks indicate significant pairwise differences between expected and observed values within each site (*** = p < 0.001, ** = p < 0.01, * = p < 0.05, n.s. = not significant).
| Compound classes |
Rostock | Uppsala | |||||
|---|---|---|---|---|---|---|---|
| Expected | Observed | NDE | Expected | Observed | NDE | ||
| CHYDR | 0.34 (0.08) | 0.36 (0.08) | 3.3% n.s. | 0.54 (0.08) | 0.53 (0.08) | -1.0% n.s. | |
| PHLM | 0.63 (0.05) | 0.63 (0.06) | -1.0% n.s. | 0.78 (0.05) | 0.78 (0.05) | 0.3% n.s. | |
| LDIM | 0.91 (0.02) | 0.91 (0.02) | -0.6% n.s. | 0.96 (0.02) | 0.95 (0.02) | -1.5% n.s. | |
| LIPID | 0.74 (0.04) | 0.74 (0.04) | -0.7% n.s. | 0.90 (0.03) | 0.90 (0.03) | 0.4% n.s. | |
| ALKYL | 0.76 (0.04) | 0.74 (0.04) | -2.0% n.s. | 0.87 (0.04) | 0.86 (0.04) | -0.9% * | |
| NCOMP | 0.48 (0.06) | 0.50 (0.07) | -0.2% n.s. | 0.71 (0.06) | 0.74 (0.06) | 3.55% n.s. | |
| PEPTI | 0.52 (0.07) | 0.50 (0.07) | -2.5% n.s. | 0.67 (0.07) | 0.67 (0.06) | 0.4% n.s. | |
| SUBER | 0.84 (0.05) | 0.83 (0.06) | -1.8% n.s. | 0.99 (0.05) | 0.76 (0.05) | -22.7% *** | |
| FATTY | 0.05 (0.01) | 0.04 (0.01) | -19.8% n.s. | 0.12 (0.05) | 0.15 (0.07) | 30.8% n.s. | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



