
Submitted:
03 July 2024
Posted:
03 July 2024
You are already at the latest version
Abstract
Abstract: Currently, antibodies are major therapeutic modalities in biopharmaceutical research and development. Because of required high dose for certain clinical applications, antibodies are formulated at high protein concentration for convenience of administration, shipping and storage. We will describe the inter-molecular interactions that can occur in such a crowded environment of high protein concentration. Two types of repulsive interactions, i.e., electrostatic and excluded volume effects, and three types of attractive interactions, i.e., hydrophobic, electrostatic and cross-linking, need to be considered in such a crowded environments. We then show consequences of such inter-molecular interactions, leading to high viscosity, opalescence and phase separation of antibody solutions. Co-solvents are used to control the stability, structure and molecular interactions and thereby to alleviate these solution problems. Among various co-solvents, arginine has been extensively used to suppress protein-protein interactions at high protein concentration. Arginine weakly but extensively interacts with aromatic/hydrophobic groups as well as charged groups on proteins, leading to effective suppression of viscosity, opalescence and phase separation of antibody solution.
Keywords:
arginine
; antibodies
; viscosity
; opalescence
; phase separation
1. Introduction
A subcutaneous injection of high dose protein therapeutics in small volume necessitates formulation of the proteins at high concentration, leading to undesirable self-association/aggregation of the proteins, in particular therapeutic monoclonal antibodies [1,2]. Such high protein concentration also often causes high viscosity, opalescence and phase separation [3]. Such high protein concentration properties may be enhanced for antibody therapeutics due to their multi-domain structures. These high protein concentration formulations also increase the probability of both reversible and irreversible aggregation [4]. When injected, the irreversible aggregates may cause undesirable reactions in patients [1,2]. The reversible aggregates will dissociate upon injection and resultant dilution and hence should not cause aggregate-related side effects. However, that may not be the case for therapeutic antibodies, as it has been shown that antibody aggregates dissociate so slowly that they may behave similarly to the irreversible aggregates [4]. Antibodies at high concentration in particular lead to high viscosity. It has been shown that those antibodies that result in high viscosity tends to aggregate more, indicating a relation between protein-protein interaction (leading to high viscosity) and aggregate formation [5,6,7]. Such a contribution of self-association also leads to opalescence, phase separation and precipitation at high protein concentrations [3].
There are several potential inter-molecular interactions that occur at high protein concentrations as described in the next section. Co-solvents are used to control inter-molecular interactions and thereby mitigate high concentration-related problems. Arginine has been shown to be particularly useful in suppressing inter-molecular interactions. We here review the mode of inter-molecular interactions for proteins, in particular antibodies, and the use of co-solvents and the mechanism of their effects on inter-molecular interactions. For example, a protein stabilizing trehalose was shown to suppress antibody self-association [1]. Co-solvents that function at such a high concentration and hence occupy a large volume fraction of solvent water, thus “called co-solvent”, have been used to alter the stability, solubility and structure of macromolecule. Such co-solvents encompass salts [8,9], sugars [10,11,12], polyols [13], amino acids [12,14] and denaturants [15]. They are typically classified as salting-in or salting-out agents and stabilizing or destabilizing/denaturing agents. Arginine belongs in neither class, as its effects on proteins are complex. It is generally known that arginine suppresses protein aggregation on the one hand [16], but on the other hand occasionally transforms insoluble aggregate into soluble aggregates [17]. Arginine also has marginal effects on protein stability [16]. We will here summarize how the co-solvents, in particular arginine, suppress protein-protein interactions.
2. Inter-molecular interactions
Figure 1 shows two main modes of repulsive inter-molecular interactions. The protein molecule is simply expressed as a globular shape having a net positive charge, although many proteins are not globular: the same argument also applies when the protein molecular has a net negative charge. It should be noted that the assumed globular shape in Figure 1 is oversimplified. At high protein concentration, the distance, d, between molecules becomes closer. Figure 1A depicts repulsive interaction between the charged protein molecules. The electrostatic repulsive force in Figure 1A is related to 1/d2 and hence increases as the inter-molecular distance becomes closer. Since each protein molecule is repelled from the surrounding proteins, their Brownian motion will be suppressed even under the repulsive electrostatic interactions.
Figure 1B shows another main mode of repulsive inter-molecular interactions. When a protein molecular approaches second molecule, there is a space around the second molecule at the distance (R) from the surface of the second molecular to the center of the first molecule, shown by the dotted line (see Figure 1B). This space is not available for any protein molecule, but is available for water. Namely, the protein is depleted in this space and thus this effect is called “depletion effect”[18] or “excluded volume effect”[19]. This depletion results in entropy loss and increased free energy. This effect also restrict the motion of the protein molecules. It is important to note that the excluded volume (depletion) effect is independent of the chemical nature of the protein molecules (for example, whether negatively or positively charged)
Figure 2 shows attractive inter-molecular interactions. Even when the protein has net charges, local charges between two molecules can be attractive (Figure 2A). They can also induce charges on the second molecule, which can result in dipole-dipole or electrostatic attractive interactions. Figure 2B also shows hydrophobic interaction, where two potential forces may operate. One force is unfavorable hydrophobic hydration, termed iceberg [20]. When two hydrophobic regions on the protein surface are combined, the unfavorable hydrophobic hydration forming iceberg is released, resulting in stabilization of the combined state and thereby hydrophobic interactions between two proteins. There is another force, called van der Waals interaction, which results from induced dipole interactions between non-polar groups. This induced dipole interaction dissipates rapidly with distance between two molecules, as it is inversely related to 6th power of the distance R. Namely, the van der Waals force is stronger as the distance gets closer and closer. However, when it gets too close, electron crowds of any atoms on the protein surface become too close, which causes electrostatic repulsion. Thus, there is an optimal distance between the electrostatic repulsion and van der Waals force, where hydrophobic interaction is maximal. In any case, hydrophobic interaction is strongest at close distance. Figure 2C and D show protein-protein interactions mediated by multivalent ions. Multivalent anions may cross-link the positively charged protein molecules, while mutivalent caions may cross-link the negatively charged protein molecules.
These effects may be compounded for antibodies, which are made of multiple domains and have an asymmetric shape, as described above (Figure 3). The domain-domain interactions are known to be dynamic as well. Such asymmetric shape results in stronger concentration dependence of viscosity, meaning that more protein-protein interactions may occur at lower protein concentration (leading to greater viscosity increase) [21]. It has been shown that antibodies ensemble different orientations of domains. Unlike globular proteins, antibodies may have different conformers with different domain-domain interactions, as depicted inFigure 3
Further compounding factor by having multiple domains is each domain contributing to inter-molecular interaction via. Fab-Fab interaction, Fab-Fc interaction and, to lesser extent, Fc-Fc interaction [22,23,24]. With anti-glucagon receptor IgG1, it was shown that hydrophobic interaction dominates inter-molecular interaction leading to the high viscosity at high antibody concentration. This is based on the observation that addition of NaCl increased the viscosity and addition of arginine decreased the viscosity. Mutation analysis confirmed that those mutations that reduce hydrophobic patches decreased solution viscosity [25].
3. High antibody concentration
High protein concentration formulation, in particular for antibody therapeutics, is preferred for cost-effective applications of antibodies. This strategy is however hampered by high viscosity, appearance of opalescence and phase separation, most likely caused by different mode and level of protein-protein interactions and aggregation/self-association of the proteins. As depicted inFigure 4, the distance between protein molecules becomes less and less, which increase the probability of either repulsive or attractive molecular interactions. These molecular interactions may be transient, but sufficient to restrict molecular motions, leading to either high viscosity or opalescence. Local fluctuations of such transient interactions can cause fluctuation of protein concentration and hence refractive index and thereby opalescence or turbidity. To mitigate such high concentration-mediated viscosity or opalescence, various approaches are taken to suppress protein-protein interactions and arginine has been shown to be effective in suppressing viscosity, opalescence and phase separation and stated as a “winner” among various approaches for reducing these phenomena: note that 17 monoclonal antibody drugs out of total 20 protein therapeutics use arginine-containing formulations [26].
3.1. Viscosity
More specifically, high viscosity occurs due to protein-protein interaction, especially with highly interactive multi-domain antibodies. This may be intuitively understood as depicted in Figure 1 and 2, where movement of a protein molecule is controlled by surrounding (same) protein molecules through different mechanisms [27]. High concentration of macromolecules inherently causes molecular crowding due to excluded volume effect (Figure 1B), which interferes with the molecular motion of the solute by the surrounding macromolecules [28]. The excluded volume effect is unrelated to the chemical nature of the macromolecules and simply occurs due to repulsive steric hindrance, which increases with the macromolecular concentration and size and therefore cannot be readily controlled by arginine or any other solution conditions. There are of course molecular interactions due to their chemical properties, i.e., attractive and repulsive interactions, as shown in Figure 1 and Figure 2. It can be easily speculated that a protein molecule would be difficult to move when there is attractive interaction between the protein molecules, which should lead to an increases in viscosity. Such attractive interaction is most likely mediated by electrostatic interactions, although hydrophobic interaction (Figure 2B) or cross-linking (Figure 2C and D) cannot be excluded. Since the proteins involved in attractive interactions are within the same molecules, they have an identical net charge. For example, if one molecule is negatively charged, then all other protein molecules are also negatively charged, which would cause repulsive interactions. The electrostatic attractive interactions thus occur by uneven charge distribution and thereby resultant local negative or positive charges, dipole moment or charge fluctuation. In either case, charge shielding by ions can suppress these electrostatic interactions. When hydrophobic interactions, in addition to electrostatic interactions, are involved, arginine (which is ionic) is effective in suppressing both hydrophobic and electrostatic interactions.
There is second possible macromolecular interaction, which is repulsive. This is exactly the case when the protein molecules possess net charges that are not sufficiently screened by ions. In the presence of such a repulsive interactions, the molecular motion of a protein molecules is also restricted, resulting in increased viscosity. In such a case as well, ionic arginine can suppress the solution viscosity, although such repulsive interaction and resultant increased viscosity may be suppressed by salt, as involvement of hydrophobic interaction in repulsive interaction is unlikely [29,30].
Viscosity of solution is expressed in a simple manner by Einstein equation: η = η0 (1 + 2.5 θ), where η is the viscosity of the protein solution, η0 is the viscosity of water and θ is the volume fraction of the protein in water. The equation indicates that the viscosity of water increases with the added particles, in proportion to the volume fraction of the particle at dilute particle concentration, as the solution viscosity is related to friction between water molecules and particles and Brownian motion of the particles. Both the friction and Brownian motion are related to the size and shape of the particle. The constant 2.5 does not always apply for any particles. In Figure 5 are depicted examples of three protein molecules to exist in dilute solution, where, the volume fraction is identical in three cases. However, the viscosity increases in the order of the particle size and shape (A < B < C). The constant is greater than 2.5 for aggregated particles. The Einstein equation only applies dilute dissociated particle solution. When the particles are electrically charged, however, the solution viscosity also deviates from Einstein relation due to the effects of charges on the surrounding ions and water molecules even at dilute particle concentration. This is not due to the electrostatic protein-protein interactions that dominate at high protein concentrations as depicted in Figure 1 and Figure 2. The charged particles bind water and ions, forming electrical double layer. When the solution moves, there is deformation or dragging of the electrical double layer, which causes increased viscosity [31]. This effect of electric charges on solution viscosity is different from the molecular interactions at high protein concentrations, as shown in Figure 1 and Figure 2, which also lead to high viscosity. When the particle concentration increases, there is inter-particle interactions, which cause deviation from the linear relation to particle concentration. The viscosity increases much sharper than the linear relation predicted by Einstein relation or the modified relations. Such non-linear relation was expressed as a polynomial function of volume fraction or even more drastically by Mooney equation [31]. Namely, as the concentration and particle number increase, there will be increased transient molecular interactions, leading to non-linear increase in solution viscosity.
Co-solvents and sequence modification have been used to reduce molecular interactions and thereby high viscosity [32]. For example, mutation analysis showed that reduced hydrophobicity leads to lower viscosity. Such a lower viscosity was ascribed to reduced attractive protein-protein interaction and thereby faster diffusion coefficient of the mutant antibody than the wild-type antibody, whose diffusion is slowed by increasing protein concentration [33]. More specifically, importance of Fv domains of the antibody was implicated for high viscosity [34]. It was shown that increased hydrophobic patches or decreased isoelectric point (pI) in Fv domains correlated with high viscosity. The decrease in pI was responsible for decreased net charges and hence lower electrostatic repulsion at the formulation pH of ~5. This suggests that high viscosity occurs due to reduced repulsive electrostatic repulsion and increased hydrophobic interaction. Retention on hydrophobic interaction chromatography has been shown to correlate with the tendency of antibodies to suffer high viscosity, as protein binding to hydrophobic resin in hydrophobic chromatography (HIC) is related the hydrophobicity of native proteins [35].
Poly-glutamine was found effective in reducing high viscosity of a mAb, perhaps masking the positively charged patches that may be responsible for electrostatic interactions [36].
3.2. Opalescence
Another issue at a high antibody concentration is appearance of the solution. When antibody solu tion is concentrated at low temperature (e.g., in cold box), a gel-like structure is often observed. Such changes in high antibody concentration solution are observed without conformational changes and hence most likely due to self-association of antibody molecules. Upon visual inspection, a solution of antibody at high concentration often exhibits bluish color, which is called opalescence [37]. Is this comparable to the phenomenon of a blue sky, where small particles in air effectively reflect shorter-wavelength light? The mechanism of opalescence has been investigated using light microscopy [38]. Bovine γ-globulin solution showed transparent appearance below 50 mg/ml and white suspension at 50 mg/ml when stored at room temperature. This 50 mg/ml suspension was opaque, indicating that particulates were formed and caused light scattering of all visible wavelengths, resulting in a white appearance. Interestingly, further increase to 120 mg/ml resulted in opalescence with bluish appearance. It may be possible that increasing antibody concentration from 50 to 120 mg/ml resulted in conversion from large to small particles and hence wavelength dependence of light scattering. This conversion appears to be opposite to what is expected from increasing protein concentration. As expected, further increase to 230 mg/ml resulted in whitish suspension and resulted in opaque appearance again. Transmission electron microscopic observation of the 230 mg/ml γ-globulin solution showed net-work formation of the proteins due to extensive protein-protein interactions. This opalescence appearance can be suppressed by salts and more effectively by arginine [39,40].
Opalescence is a consequence of gradient of refractive index in solution, which causes reflection of low wavelength light (for blueish color appearance). Such differential light reflection may arise from small particles or fluctuation of refractive index but due to network formation of antibody molecules.
3.3. Liquid-liquid phase separation
These high viscosity and opalescence are mediated by weak protein-protein interactions that prop agate to form a transient net-work throughout the entire solution. In cold temperature and high antibody concentration, liquid droplets are sometimes observed [39]. This may be ascribed to reversible stronger attractive protein-protein interactions, which generate a protein cluster or a condensate. This is depicted in Figure 6, where an unstable intermediate structure (A) with perhaps a single-bonded dimer structure may be formed. This unstable intermediate may further reversibly associate to form condensates. While some condensates (B) with less bonds may be unstable and converted to more stable condensates (C) with greater number of bonds. The solution surrounding the condensate would have higher refractive index than the bulk phase and be observable as different phase, i.e., liquid droplet. The condensates and hence liquid droplet may become round, as an addition of a protein molecule (dark blue in D or E) would be more stable with multiple bonds as in E than D, which would make the condensate more spherical. This condensate formation and hence droplet formation can be suppressed by arginine.
3.4. Aggregation and amyloid
Another issue with antibodies is amyloid fibril formation, although it is not related to a whole antibody structure [41]. It occurs in vivo when immunoglobulin light chain is secreted into blood circulation. The light chains readily undergo conformational changes and deposit as fibrils on various tissues [42]. Fibril formation was also observed in vitro with various immunoglobulin variable domains [43]. To our knowledge, arginine has never been shown to disrupt aggregated immunoglobulin light chain fibrils. Arginine is highly unlikely to disaggregate pre-formed fibrils due to no denaturing capacity of arginine and the observed resistance of these fibrils to strong protein denaturant, sodium dodecyl sulfate. However, it has been shown in vitro to suppress amyloid formation and often lead to formation of different types of aggregates [44,45].
3.5. Reversible aggregation
Reversible antibody aggregation at high concentration may not be a problem, as injection should lead to dilution and thereby dissociation of reversible aggregates. However, it may not be the case for antibodies. It has been shown that the aggregated antibody dissociate so slowly that the aggregated species may persist long enough in circulation to cause aggregate-related pharmacological problems [46].
4. The effects of arginine
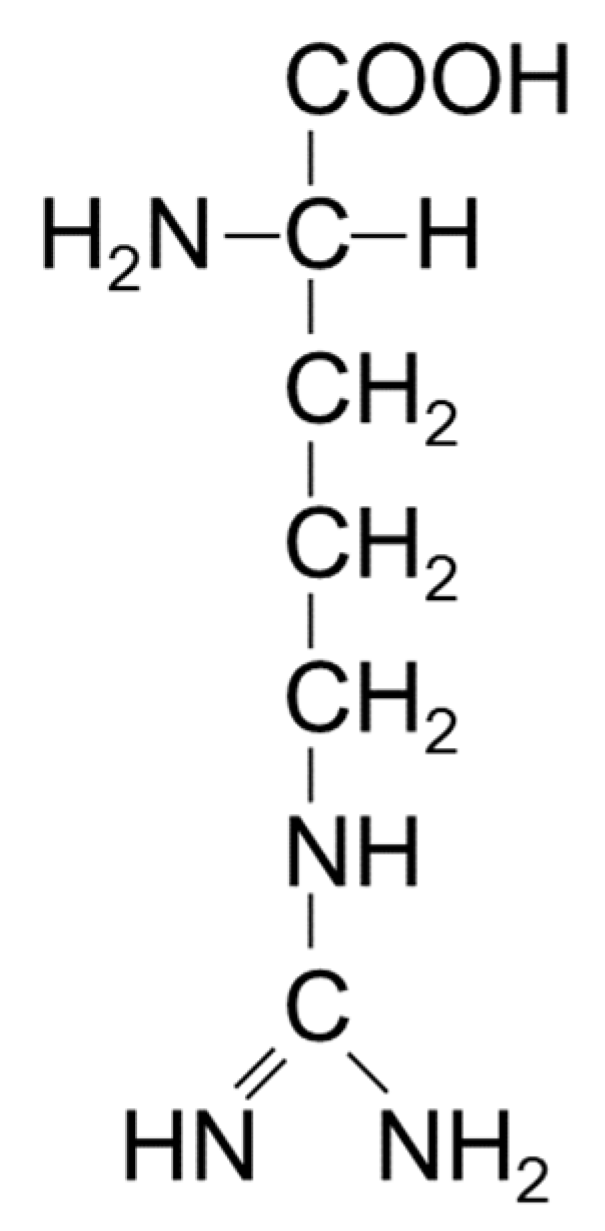
Among various co-solvents, arginine is unique as a co-solvent in controlling the solubil ity and sta bility of proteins and nucleic acids [16,47]. It is noted here that arginine normally refers to arginine hydrochloride, as arginine is generally used as a chloride form around neutral pH. However, any acid can be used to titrate basic arginine to neutral to acidic pH range, as in, for example, arginine sulfate that has been used for protein chromatography and formulation [48,49]. Arginine (mostly arginine hydrochloride) is used at moderate to high concentration (e.g., even above 1 M) to be effective as a co-solvent. Requirement of high concentration to be effective indicates weak, but extensive interactions with the target macromolecules, here antibodies. What are the cause of weak interactions? Figure 7 shows the chemical structure of arginine. It was described that arginine can interact with proteins through multiple mechanisms, including electrostatic force, π-cation interactions and aromatic/hydrophobic interactions. The guanidinium group has an aromatic character, which can participate in aromatic interactions with aromatic side chains, i.e., tryptophan, tyrosine and phenyl alanine. The planar nature of the guanidinium group has been shown to bind to aromatic groups through π-π stacking between guanidinium group of arginine and aromatic side chains of protein [50]. It can also confer hydrogen bonding to the peptide groups of the proteins. The positive charges of the amino and guanidinium group offer π-cation interactions with the above aromatic side chains. Since it is ionic, arginine interacts with the charged groups of the proteins. As seen in Figure 7, arginine may also show affinity for non-polar groups through three methylene groups. With regard to the interaction of arginine hydrochloride with proteins, the electrostatic force can be substituted for by NaCl or any ionic solute. At moderate to high concentration, however, the π-cation or interactions and aromatic/hydrophobic interactions, which cannot be replaced by NaCl, dominate arginine-protein interactions. However, such binding interaction with proteins is too weak or too limited to denature proteins, different from protein binding denaturants such as urea and guanidine hydrochloride [16].
The above general discussion applies to the interactions between arginine and antibodies. However, it should be noted that antibody is a multi-domain protein and may be considered as an aggregate of multiple globular domains. These globular domains are packed by inter-domain interactions into an overall antibody structure, as depicted in Figure 3 [51]. Thus, the suppressive effects of arginine on protein-protein interactions may cause alteration in domain-domain interactions of antibodies. Arginine has been shown to increase the local flexibility in antibody structure [52,53]. In fact, arginine significantly contributes to modulation of reversible protein-protein interactions, as has recently been reviewed [54]. There are many advantages of using arginine in antibody purification [55,56]. Arginine was also found to suppress antibody aggregation during thermal stress [32]. However, adverse effects of arginine on antibodies have also been observed. Arginine resulted in increased aggregation and formation of subvisible particles of immunoglobulin solution against thermal or benzyl alcohol treatments [57,58]. Arginine was also found to reduce monomer recovery of a monoclonal antibody upon refolding from urea denaturation [59] and increased formation of subvisible particles of human intravenous immunoglobulin G solution against thermal or agitation stress [60]. Arginine residues in antibodies, not arginine co-solvent, may also contribute to poly-reactivity (non-specific antigen binding) [61]. These results suggest that arginine-antibody interaction can interfere with the domain-domain interactions and thereby lead to incorrect domain packing or antigen binding. However, arginine clearly shows suppression of antibody-antibody interactions, as described below.
5. Conclusions
Protein-protein interaction at high protein concentration leads to high viscosity, opalescence and phase separation as well as protein precipitation. This protein-protein interaction is further compounded for antibodies due to their conformation dynamics, which may augment inter-molecular interactions. We have speculated that such conformational dynamics of antibodies can lead to poly-reactivity of antibodeis, which may be mediated by altered self-association of antibodies [61]. We then suggested that acid used in Protein A chromatography may cause conformational heterogeneity and an importance of non-affinity purication [56]. We have described how arginine interacts with protein and nucleic acid. Regarding the interaction with proteins, we noted some detail of high concentration antibody formulation that generate high viscosity, opalescence and droplet formation and suppression of these phenomena by arginine. With regard to nucleic acid, we have reviewed available data on the interaction of arginine with the phosphate backbones and nucleobases in nucleic acids, showing their favorable interactions with nucleobases and backbone phosphate groups. Arginine being ionic, it can interact electrostatically with negative charges on the nucleic acids but also interact with nucleobases through multiple mechanisms, such as π-cation and hydrophobic/aromatic interactions. Such interactions can affect the interactions between proteins and nucleic acids, possibly leading to higher quality products in various nucleic acid applications.
Funding
We received no external funding.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank Dr. Daisuke Ejima for his insightful suggestions, which helped us strengthen the quality of the manuscript.
Conflicts of Interest
Tsutomu Arakawa was formerly affiliated with the for-profit company Alliance Protein Laboratories but currently has no conflicts of interest. C.S. and Teruo Akuta are employees of the for-profit company Kyokuto Pharmaceuticals. It is important to note that they do not have any conflicts of interest. S.N. and K.S. are staff members at University of Tsukuba and have no conflicts of interest.
References
- Lui, L.H.; van der Walle, C.F.; Brochchini, S.; Velayuolhan, A. Discovering novel small molecule compound for prevention of monoclonal self-association. Antibodies 2022, 11, 40. [Google Scholar] [CrossRef]
- Li, W.; Probakaran, P.; Chen, W.; Zhu, Z.; Feng, Y.; Dimitrov, D.S. Antibody aggregation: insights from sequence and structure. Antibodies 2016, 5, 19. [Google Scholar] [CrossRef] [PubMed]
- Sibanda, N.; Shanmugam, R.K.; Curtis, R. The relationship between protein-protein interactions and liquid-liquid phase separa tion for monoclonal antibodies, Mol. Pharm. 2023, 20, 2662–2674. [Google Scholar] [CrossRef]
- Moore, J.M.R.; Patapoff, T.W.; Cromwell, M.E.M. Kinetics and thermodynamics of dimer formation and dissociation for a re combinant humanized monoclonal antibody to vascular endothelial growth factor. Biochemistry 1999, 38, 13960–13967. [Google Scholar] [CrossRef]
- Kingsbury, J.S.; Saini, A.; Auclair, S.M.; Fu, L.; Lantz, M.M.; Halloran, K.T.; Calero-Rubio, C.; Schwenger, W.; Airiau, C.Y.; Zhang, J.; Gokarn, Y.R. A single molecular descriptor to predict solution behavior of therapeutic antibodies. Sci Adv 2020, 5, eabb0372. [Google Scholar] [CrossRef] [PubMed]
- Chaudhri, A. The role of amino acid sequence in the self-association of therapeutic monoclonal antibodies: Insights frpm co arce-grained modeling. J. Phys. Chem. 2013, B117, 1269–1279. [Google Scholar] [CrossRef]
- Yadav, S.; Shire, S.J.; Kalonia, D.S. Factors affecting the viscosity in high concentration solutions of different monoclonal anti bodies. J. Pharm. Sci. 2010, 99, 4812–4829. [Google Scholar] [CrossRef]
- Hofmeister, F. Zur Lehre von der wirkung der salze. Arch. Expt. Pathl. Pharmakol. 1888, 24, S12–S23. [Google Scholar] [CrossRef]
- Traube, J. The theory of attraction pressure. J. Phys. Chem. 1910, 14, 471–475. [Google Scholar] [CrossRef]
- Traube, J. The attraction pressure. J. Phys. Chem. 1910, 14, 452–470. [Google Scholar] [CrossRef]
- Liu, L.; Qi, W.; Schwartz, D.K.; Randolph, T.W.; Carpenter, J.H. The effects of excipients on protein aggregation during agi tation: an interfacial shear rheology study. J. Pharm. Sci. 2013, 102, 2460–2470. [Google Scholar] [CrossRef]
- Yancey, P.H.; Clark, M.E.; Hand, S.C.; Bowlus, R.D.; Somero, G.N. Living with water stress: evolution of osmolyte systems. Science 1982, 217, 1214–1222. [Google Scholar] [CrossRef]
- Gerlsma, S.Y. Reversible denaturation of ribonuclease in aqueous solutions as influenced by polyhydric alcohols and some other additives. J. Biol. Chem. 1968, 243, 857–961. [Google Scholar] [CrossRef]
- Arakawa, T.; Timasheff, S.N. The stabilization of proteins by osmolytes. Biophys. J. 1985, 47, 411–414. [Google Scholar] [CrossRef]
- Tanford, C. Protein Denaturation: Part C. Theoretical models for the mechanism of denaturation. Adv. Protein Chem. 1968, 23, 121–282. [Google Scholar] [CrossRef]
- Tsumoto, K.; Umetsu, M.; Kumagai, I.; Ejima, D.; Philo, J.S.; Arakawa, T. Role of arginine in protein refolding, solubilization, and purification. Biotechnol. Prog. 2004, 20, 1301–1308. [Google Scholar] [CrossRef]
- Shibata, C.; Iwashita, K.; Shiraki, K. Selective separation method of aggregates from IgG solution by aqueous two-phase system. Protein Expr. Purif. 2019, 161, 57–62. [Google Scholar] [CrossRef]
- Asakura, S.; Oosawa, F. On interaction between two bodies immersed in a solution of macromolecules. J. Chem. Phys. 1954, 22, 1255. [Google Scholar] [CrossRef]
- Minton, A.P. Excluded volume as a determinant of protein structure and stability. Biophys. J. 1980, 32, 77–79. [Google Scholar] [CrossRef]
- Kauzmann, W. Structural factors in protein denaturation. J. Cell Physiol. Suppl. 1956, 47, 113–131. [Google Scholar] [CrossRef]
- Tian, Z.; Jiang, X.; Chen, Z.; Huang, C.; Qian, F. Quantifyinh protein shape to elucidate its inf;uence on solution viscosity in high-concentration electrolyte solutions. Mol. Pharm. 2024, 21, 1719–1728. [Google Scholar] [CrossRef]
- Lilyestrom, W.G.; Yadav, S.; Shire, S.J.; Scherer, T.M. Monoclonal antibody self-association, cluster formation, and rheology at high concentrations. J. Phys. Chem. B. 2013, 117, 6373–6384. [Google Scholar] [CrossRef]
- Arora, J.; Hu, Y.; Esfandiary, R.; Sathish, H.A.; Bishop, S.M.; Joshi, S.B.; Middaugh, C.R.; Volkin, D.B.; Weis, D.D. Charge-mediated Fab-Fc interactions in an IgG1 antibody induce reversible self-association, cluster formation, and elevated viscosity. MABs 2016, 8, 1561–1574. [Google Scholar] [CrossRef]
- Kanai, S.; Liu, J.; Patapoff, T.W.; Shire, S.J. Revsersible self-association of a concentrated monoclonal antibody solution mediated by Fab-Fab interaction that impacts solution viscosity. J. Pharm. Sci. 2008, 97, 4219–4227. [Google Scholar] [CrossRef]
- Dai, J.; Izadi, S.; Zarzar, J.; Wu, P.; Oh, A.; Carter, P.J. Variable domain mutational analysis to probe the molecular mechanisms of high viscosit of an IgG1 antibody. MABS 2024, 16, 2304282. [Google Scholar] [CrossRef]
- Ren, S. Effects of arginine in therapeutic protein formulations: a decade review and perspectives. Antib. Ther. 2023, 6, 265–276. [Google Scholar] [CrossRef]
- Hong, T.; Iwashita, K.; Shiraki, K. Viscosity control of protein solution by small solutes: a review. Curr. Protein Pept. Sci. 2018, 19, 746–758. [Google Scholar] [CrossRef]
- Xu, A.Y.; Castellanos, M.M.; Mattison, K.; Kruger, S.; Curtis, J.E. Studying excipient modulated physical stabilioty and viscosity of monoclonal antibody formulations using small angle-light scattering. Mol. Pharm. 2019, 16, 4319–4338. [Google Scholar] [CrossRef]
- Inoue, N.; Takai, E.; Arakawa, T.; Shiraki, K. Arginine and lysine reduce the high viscosity of serum albumin solutions for pharmaceutical injection. J. Biosci. Bioeng. 2014, 117, 539–543. [Google Scholar] [CrossRef]
- Inoue, N.; Takai, E.; m Arakawa, T.; Shiraki, K. Specific decrease in solution viscosity of antibodies by arginine for therapeutic formulations. Mol. Pharm. 2014, 11, 1889–1896. [Google Scholar] [CrossRef]
- Pindrus, M.A.; Shire, S.J.; Yadav, S.; Kalonia, D.S. Challenges in determining intrinsic viscosity under low ionic strength. Phar Res. 2017, 34, 836–846. [Google Scholar] [CrossRef]
- Nishinami, S.; Yoshizawa, S.; Arakawa, T.; Shiraki, K. Allantoin and hydantoin as new protein aggregation suppressors. Int. J. Biol. Macromol. 2018, 114, 497–503. [Google Scholar] [CrossRef]
- Armstrong, G.B.; Shah, V.; Sanches, P.; Patel, M.; Casey. R.; Jamieson, C.; Burley, G.A.; Lewis, W.J.; Rattray, Z. Enhancing viscosity control in antibody formulations: A framework for the biophysical screening of mutations targeting solvent-accessible hydrophobic and electrostatic patches. Molecules 2004, 23, 2345–2357. [Google Scholar] [CrossRef]
- Malkowski, E.K.; Chen, H.T.; Wang, T.; Wu, L.; Huang, J.; Mock, M.; Underhill, P.; Pelegri-O’Day, E.; Maglalang, E.; Winters, D.; Tessier, P.M. Reduction of monoclonal antibody viscosity using interpretable machine learning. MABS 2024, 16, 2303781. [Google Scholar] [CrossRef]
- Arakawa, T.; Tomioka, Y.; Kurosawa, Y.; Akuta, T. Elucidating the mechanism of additive effects at high concentrations on hydrophobic interaction chromatography. J. Chromatogr. A. 2023, 1702, 464091. [Google Scholar] [CrossRef]
- Lapenna, A.; Dagallier, C.; Huille, S.; Tribet, C. Poly(glutamic acid)-based viscosity reducers for concentrated formulations of a monclonal IgG antibody. Mol. Pharm. 2024, 21, 982–991. [Google Scholar] [CrossRef]
- Salinas, B.A.; Satishi, H.A.; Bishop, S.M.; Hann, N.; Carpenter, J.H.; Randolph, T.W. Understanding and modulating opalescence and viscosity in a monoclonal antibody formulation. J. Pharm. Sci. 2010, 99, 82–93. [Google Scholar] [CrossRef]
- Nakauchi, Y.; Nishinami, S.; Murakami, Y.; Ogura, T.; Kano, H.; Shiraki, K. Opalescence arising from network assembly in antibody solution. Mol. Pharm. 2022, 19, 1160–1167. [Google Scholar] [CrossRef]
- Oki, S.; Nishinami, S.; Shiraki, K. Arginine suppresses opalescence and liquid-liquid phase separation in IgG solutions. Int. J. Biol. Macromol. 2018, 118, 1708–1712. [Google Scholar] [CrossRef]
- Oki, S.; Nishinami, S.; Nakauchi, Y.; Ogura, T.; Shiraki, K. Arginine and its derivatives suppress the opalescence of an antibody solution. J. Pharm. Sci. 2022, 111, 1126–1132. [Google Scholar] [CrossRef]
- Stevens, F.J.; Kisilevsky, R. Immunoglobulin light chains, glycosaminoglycans, and amyloid. Cell Mol. Life Sci. 2000, 57, 441–449. [Google Scholar] [CrossRef]
- Ramirez-Alvarado, M. Amyloid formation in light chain amyloidosis. Curr. Top Med. Chem. 2012, 12, 2523–2533. [Google Scholar] [CrossRef]
- Stevens, P.W.; Raffen, R.; Hanson, D.K.; Deng, Y.L.; Berrios-Hammond, M.; Westholm, F.A.; Murphy, C.; Eulitz, M.; Wetzel, R.; Solomon, A.; et al. Recombinant immunglobulin variable domains generated from synthetic genes provide a system for in vitro characterization of light-chain amyloid proteins. Protein Sci. 1995, 4, 421–432. [Google Scholar] [CrossRef]
- Minakata, E.N.; Popiel, H.A.; Tada, M.; Takahashi, T.; Yamane, H.; Saitoh, Y.; Takahashi, Y.; Ozawa, D.; Takeda, A.; Takeuchi, T.; et al. Arginine is a disease modifier for polyQ disease models that stabilize polyQ protein conformation. Brain 2020, 143, 1811–1825. [Google Scholar] [CrossRef]
- Das, U.; Hariprasad, G.; Ethayathulla, A.S.; Manral, P.; Das, T.K.; Pasha, S.; Mann, A.; Gangull, M.; Verma, A.K.; Bhat, R.; et al. Inhibition of protein aggregation: supramolecular assemblies of arginine hold the key. PLoS One 2007, 11, e1176. [Google Scholar] [CrossRef]
- Moore, J.M.R.; Patapoff, T.W.; Cromwell, M.E.M. Kinetics and thermodynamics of dimer formation and dissociation for a recombinant humanized monoclonal antibody to vascular endothelial growth factor. Biochemistry 1999, 38, 13960–13967. [Google Scholar] [CrossRef]
- Hirano, A.; Tokunaga, H.; Tokunaga, M.; Arakawa, T.; Shiraki, K. The solubility of nucleobases in aqueous arginine solutions. Arch. Biochem. Biophys. 2010, 197, 90–96. [Google Scholar] [CrossRef]
- Mozgovics, M.; Fischer, A.; Brocard, C.; Jungbauer, A.; Lingg, N. L-Arginine sulfate reduces irreversible protein binding in immobilized metal affinity chromatography. J. Chromatogr. A. 2023, 1706, 464246. [Google Scholar] [CrossRef]
- Maity, H.; Karkaria, C.; Davagnino, J. Mapping of solution components, pH changes, protein stability and the elimination of protein precipitation during freeze-thawing of fibroblast growth factor 20. Int. J. Pharm. 2009, 378, 122–135. [Google Scholar] [CrossRef]
- Ferrari, L.; Stucchi, R.; Konstantoulea, K.; van de Kamp, G.; Kos, R.; Geerts, W.J.C.; van Bezouwen, L.S.; Förster, F.G.; Altelaar, M.; Hoogenraad, C.C.; et al. Arginine π-stacking drives binding to fibrils of the Alzheimer protein Tau. Nat. Commun. 2020, 11, 571. [Google Scholar] [CrossRef]
- Li, W.; Prabakaran, P.; Chen, W.; Zhu, Z.; Feng, Y.; Dimitrov, D.S. Antibody aggregation: insights from sequence and structure. Antibodies (Basel) 2016, 5, 19. [Google Scholar] [CrossRef]
- Manikwar, P.; Majumdar, R.; Hickey, J.M.; Thakkar, S.V.; Samra, H.S.; Sathish, H.A.; Bishop, S.M.; Middaugh, C.R.; Weiss, D.D.; Volkin, D.B. Correlating excipient effects on conformational and storage stability of an IgG monoclonal antibody with local dynamics as measured by hydrogen/deuterium-exchange mass spectrometry. J. Pharm. Sci. 2013, 102, 2136–2151. [Google Scholar] [CrossRef] [PubMed]
- Lilyestrom, W.G.; Shire, S.J.; Scherer, T.M. Influence of the cosolute environment on IgG solution structure analyzed by small-angle X-ray scattering. J. Phys. Chem. B. 2012, 116, 9611–9618. [Google Scholar] [CrossRef]
- Gupta, M.N.; Uversky, V.N. Biological importance of arginine: A comprehensive review of the roles in structure, disorder, and functionality of peptides and proteins. Int J Biol Macromol. 2024, 257, 128646. [Google Scholar] [CrossRef]
- Arakawa, T.; Philo, J.S.; Tsumoto, K.; Yumioka, R.; Ejima, D. Elution of antibodies from a Protein-A column by aqueous arginine solutions. Protein Expr. Purifi. 2004, 36, 244–248. [Google Scholar] [CrossRef]
- Arakawa, T.; Tomioka, Y.; Nakagawa, M.; Sakuma, C.; Kurosawa, Y.; Ejima, D.; Tsumoto, K.; Akuta, T. Non-affinity purification of antibodies. Antibodies (Basel) 2023, 12, 15. [Google Scholar] [CrossRef]
- Yoshizawa, S.; Arakawa, T.; Shiraki, K. Thermal aggregation of human immunoglobulin G in arginine solutions: Contrasting effects of stabilizers and destabilizers. Int. J. Biol. Macomol. 2017, 104, 650–655. [Google Scholar] [CrossRef]
- Yoshizawa, S.; Oki, S.; Arakawa, T.; Shiraki, K. Trimethylamine N-oxide (TMAO) is a counteracting solute of benzyl alcohol for multi-dose formulation of immunoglobulin. Int. J. Biol. Macromol. 2018, 107, 984–989. [Google Scholar] [CrossRef]
- Svilenov, H.L.; Kulakova, A.; Zalar, M.; Golovanov, A.P.; Harris, P.; Winter, G. Orthogonal techniques to study the effect of pH, sucrose, and arginine salts on monoclonal antibody physical stability and aggregation during long-term storage. J Pharm Sci. 2020, 109, 584–594. [Google Scholar] [CrossRef]
- Hada, S.; Burlakoti, U.; Kim, K.H.; Han, J.S.; Kim, M.J.; Kim, N.A.; Jeong, S.H. ; A comprehensive evaluation of arginine and its derivatives as protein formulation stabilizers. Int J Pharm. 2023, 647, 123545. [Google Scholar] [CrossRef]
- Arakawa, T.; Akuta, T. Mechanistic insight into poly-reactivity of immune antibodies upon acid denaturation or arginine mutation in antigen-binding regions. Antibodies (Basel) 2023, 12, 64. [Google Scholar] [CrossRef]
Figure 1.
Repulsive inter-molecular interaction.

Figure 2.
Attractive inter-molecular interaction.
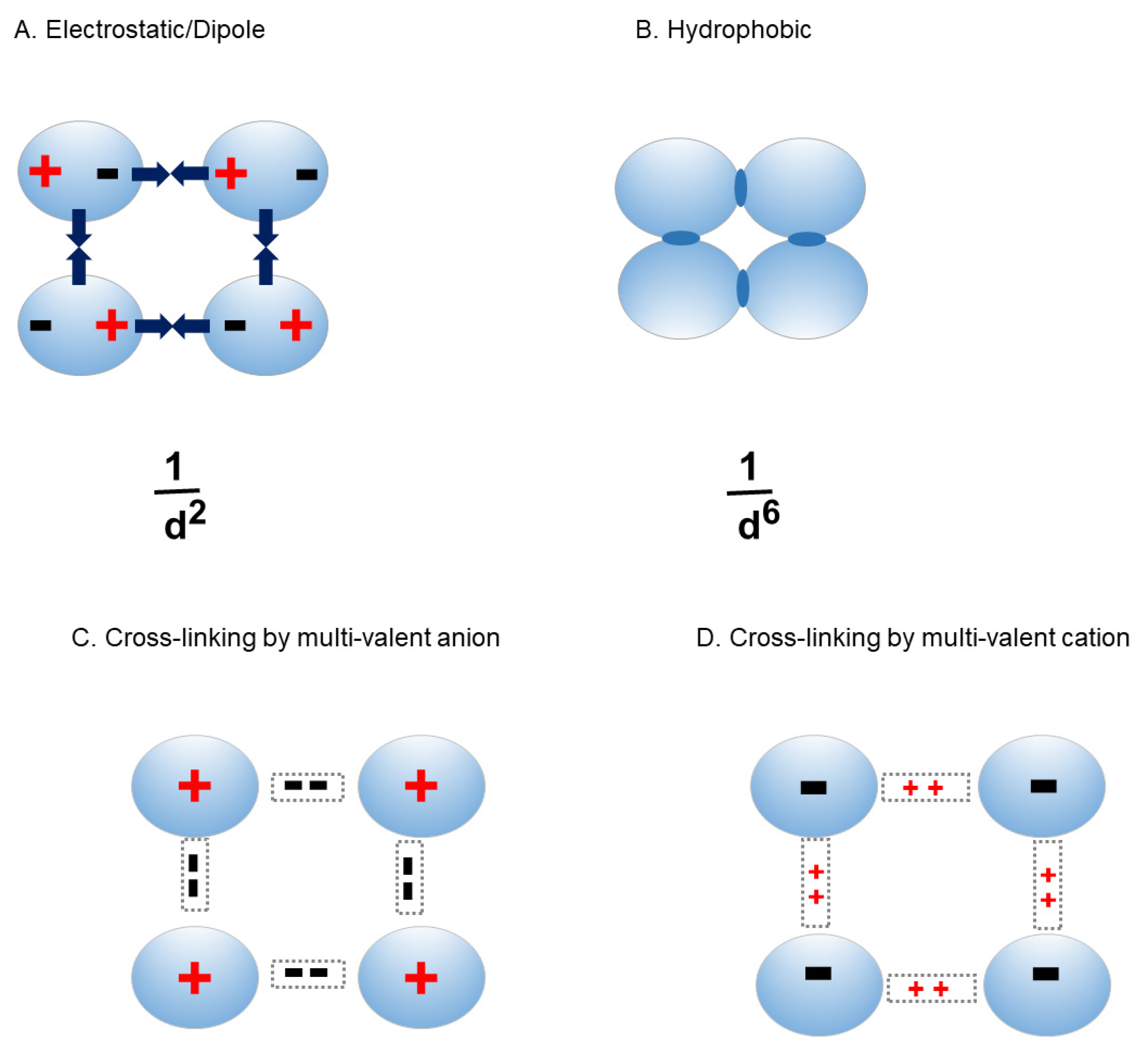
Figure 3.
Dynamic conformational ensemble of antibodies.
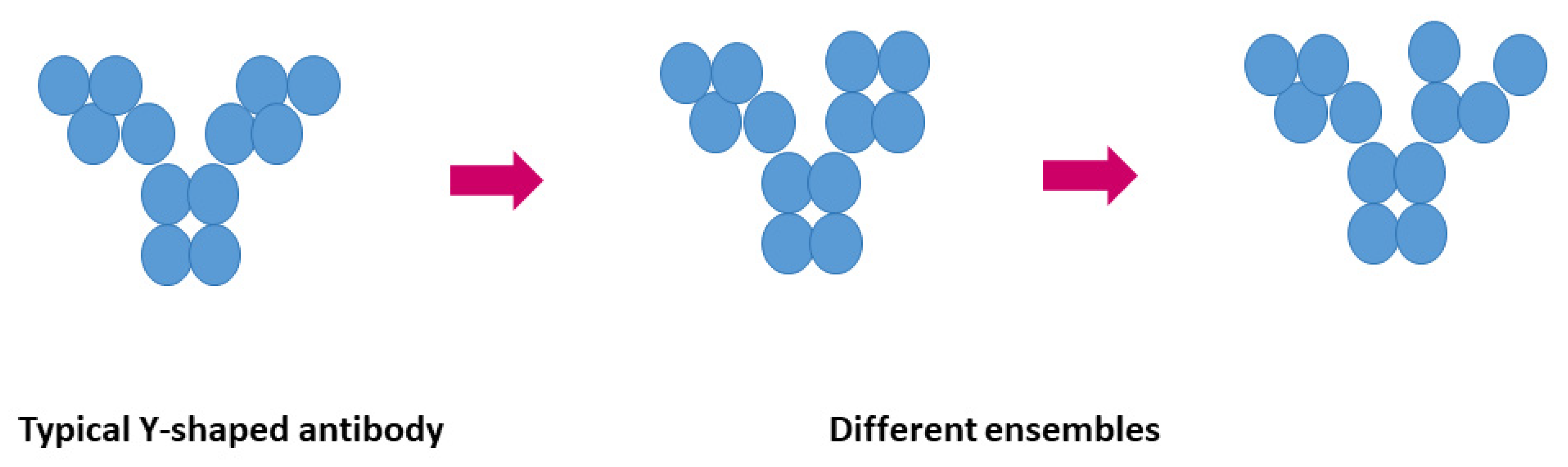
Figure 4.
Molecular interactions leading to high viscosity and opalescence of antibody solution.
Figure 5.
The effects of protein-protein interactions on solution viscosity.

Figure 6.
The mode of protein-protein interactions leading to condensate and phase separation.
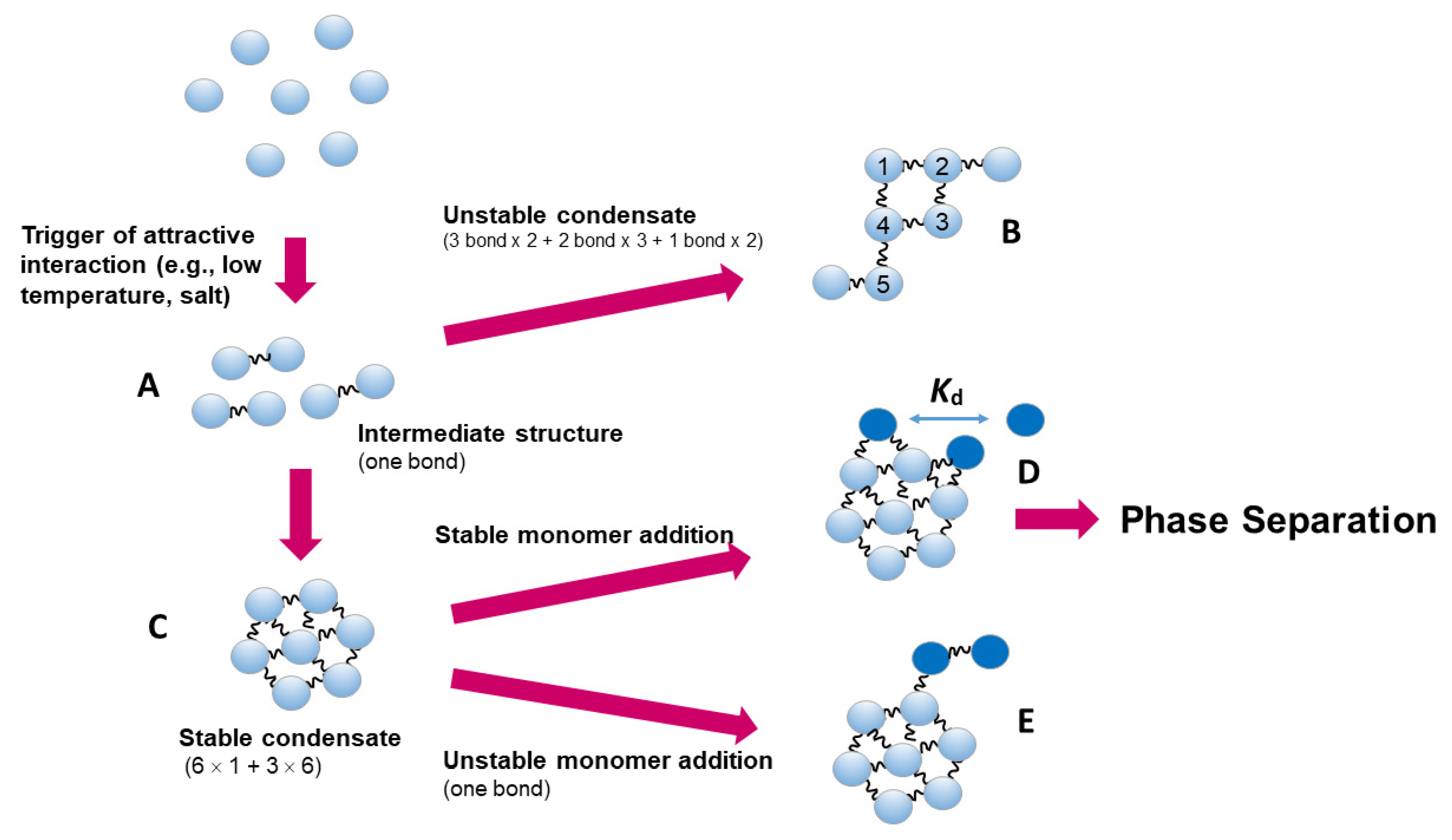
Figure 7.
The chemical structure of arginine.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



