
Submitted:
03 July 2024
Posted:
03 July 2024
You are already at the latest version
Abstract
Lanthanides and actinides are emerging contaminants, but little is known about their uptake and distribution by plants and their interactions in the rhizosphere. To better understand the fate of these metals in plants, we assessed the bioassociation of 2, 20 and 200 µM Eu(III) by five hydroponically grown crops endemic to Europe. The metal’s concentration and its speciation was monitored by inductively coupled plasma mass spectrometry and laser spectroscopy, whereas root exudation was investigated by chromatographic methods. It has been shown, that Eu(III) bioassociation is a two-stage process, involving rapid biosorption followed by accumulation in root tissue and distribution to the stem and leaves. Within 96 h of exposure time, the plant induces a change of Eu(III) speciation in the liquid medium, from a predominant Eu(III) aquo species, as calculated by thermodynamic modelling, to a species with longer luminescence lifetime. Root exudates such as citric, malic, and fumaric acid were identified in the cultivation medium and affect Eu(III) speciation in solution, as was shown by a change in the thermodynamic model. These results contribute to a comprehensive understanding of the fate of lanthanides in the biosphere and provide a basis for further investigations with the chemical analogues Cm(III) and Am(III).
Keywords:
europium
; speciation
; phytoremediation
; bioassociation
; laser spectroscopy
; lanthanides
; hydroponics
; plant uptake
; root exudates
; thermodynamic modelling
1. Introduction
Soil and water contaminated with actinides (An) and lanthanides (Ln) poses a risk for all living organisms. Though most of them occur naturally on earth, human activities are increasingly introducing these metals into the environment. Beside nuclear explosions, accidents and emissions from the nuclear fuel cycle, the mining and further processing of ores, dumping and the excessive use of phosphate fertilizers is primarily responsible for increased levels of An and Ln in the environment. The risk of exposure, e.g. by inhalation of dust, holds particularly true for workers, farmers and the public in the vicinity of such production, emission and dissemination sites [1]. However, these metals might also be absorbed by a larger number of people through the consumption of vegetables, fruits, food grains, fish, meat and milk [2,3,4]. In this regard, remediation of contaminated areas and ecosystems is extremely important to maintain food safety and compliance with environmental regulations. Conventional physico-chemical remediation techniques employ soil replacement, dumping, thermal desorption, soil washing with chelators and acids or immobilization by vitrification, solidification and electrokinetic remediation. These methods are often accompanied by ecologically detrimental side effects such as changes in edaphic conditions, creation of secondary contaminations, accelerated soil erosion and, due to energy consumption and high costs, economic inefficiency [5]. In contrast, biological remediation approaches are a cost-effective, environmentally friendly and viable alternative.
Phytoremediation uses green plants to remove contaminants from soils, surface waters and atmosphere or to mitigate their adverse effects in the environment. Additionally, phytoremediation contributes to carbon sequestration, maintains biodiversity, minimizes soil erosion and provides the feedstock for biofuel production [6]. The mechanisms involved in phytoremediation are phytoextraction, phytostabilization, rhizofiltration, phytovolatilization and phytodegradation. The absorption of contaminants from soils into the biomass, their concentration and following translocation into harvestable plant parts is denoted as phytoextraction, which is particularly suitable for diffusely contaminated sites with superficial pollutants [7]. Plants for phytoextraction should – in the best case – tolerate and accumulate certain levels of metals in their aboveground parts and gain a high biomass in relatively short time [8]. The effectiveness of bioassociation in natural environments strongly depends on edaphic conditions, especially on soil parameters such as pH, redox potential, organic matter content, soil mineralogy and the presence of nutrients, which ultimately all have an influence on the speciation, i.e. the bioavailability of the metal contamination [9]. Only free metal ions, soluble complexes and colloids in the soil solution are readily available for plant bioassociation, partially also ions adsorbed to inorganic soil components, whereas organically bound An and Ln, their oxides, carbonates and hydroxides are less bioavailable [8]. However, plants can influence the bioavailability of metals through root exudation, i.e. releasing organic compounds into the rhizosphere to solubilize or mobilize nutrients, thereby enhancing nutrient and metal uptake, respectively [10].
Furthermore, bioassociation of non-essential metals is plant type and species specific. This specificity is based on genetic and molecular factors, such as the presence of specialized metal transport proteins, metal chelators and the plant’s homeostasis mechanisms [11,12]. In this regard, plants can be categorized into three types depending on their ability to tolerate and accumulate metals present in the habitat they grow within:
- Indicator plants serve as sensitive biological indicators of environmental contamination and accumulate metals in their tissues at levels proportionate to the bioavailable metal concentration in the growth substrate [13,14]. Helophytes such as Rorippa amphibian can therefore be used to biomonitor the Fe, Mn, Cu and Cd load in rivers and sediments [15].
- Excluder plants are able to tolerate high concentrations of metals in their growth medium up to a threshold concentration by restricting the accumulation in the cells by binding of metals in the root zone, blocking uptake in the root tissue or by energy dependent efflux pumps [14,16,17]. Oenothera biennis, for example, is considered as Cd and Cu excluder plant, showing very low metal concentrations in the aboveground plant parts [18].
- Hyperaccumulators are plants capable to tolerate and actively accumulate specific metals to several percent of their dry mass in the shoots. First introduced by Brooks et al. for plants accumulating more than 1000 µg Ni g−1 dry weight (DW), hyperaccumulators are found among many plant groups and metals [13,19]. For Ln, the greatest number of hyperaccumulators are species within the group of ferns, with Dicranopteris dichotoma being a well-studied representative [20].
However, it is important to highlight that the processes responsible for the accumulation of metals in hyperaccumulators are specific for one, or at most, a small number of elements showing similar chemical properties, such as REE. Beside the type of contamination, various other factors must be taken into account when choosing candidates for phytoremediation. Bioaccumulation capacity, biomass production, ecological suitability with regard to climate and soil, maintenance requirements such as watering and fertilization and legal regulatory factors have to be assessed to select appropriate plant species [8].
In our present study, we selected five crops endemic to Europe that are both drought-resistant and robust. In addition, they have a low nutritional demand for their growth substrate and form branched root systems. We chose representative plants belonging to the family of grasses (Poaceae), namely Avena strigosa, plants of the Fabaceae family, namely Lotus corniculatus, Trifolium incarnatum and Medicago sativa, as well as a plant from the family of Linaceae, namely Linum usitatissimum. Beside their inherent botanical suitability for a potential application in phytoremediation in Europe, those plants are assigned an economic value as green manure, fodder and fiber raw material. In this regard, knowledge about bioassociation and translocation behavior of non-essential elements in the plant’s tissue is crucial to assess the risk of a transfer into the human food chain.
This model system study aimed to explore the potential of the aforementioned plant species for bioassociation of Eu(III) as chemical and non-radioactive analogue for Ln and the trivalent An Cm(III) and Am(III). Therefore, plants were grown in a hydroponic medium and exposed to variable concentrations of Eu(III). The liquid medium was analyzed by inductively coupled plasma mass spectrometry (ICP-MS) and time resolved laser induced fluorescence spectroscopy (TRLFS) to obtain information on the metal’s concentration and speciation. The amount of bioassociated Eu(III) was quantified in individual plant parts to draw conclusions about their potential for phytoremediation. Furthermore, root exudates were scrutinized by high performance liquid chromatography (HPLC) to unveil plant-related organic compounds that might have an influence on speciation and thus bioavailability of metals. Thermodynamic modelling was used to support the interpretation of TRLFS results and assess the influence of root exudates on the Eu(III) speciation throughout the experiment. Overall, we want to understand the process of bioassociation from both sides: from bioavailability, to rate and amount of uptake, to the interaction of the plant roots with the metal and the medium.
2. Materials and Methods
2.1. Plant Cultivation
Seeds of A. strigosa, L. usitatissimum and L. corniculatus were purchased from Sativa Rheinau AG (Switzerland) and seeds of M. sativa and T. incarnatum from Hof Jeebel GmbH & Co. KG (Germany). Seed were stored at 5 °C impervious to light for stratification. For germination and plant growth, a hydroponic system (Araponics, Belgium) was used. First, 2.5 % (w/v) sterilized plant agar in MilliQ water was pipetted into seed holders and plant seeds were placed some millimeters deep in the solidified agar. Seed holders prepared in this way were hung into a tap water filled hydroponic system in a phytochamber with a 16/8 h day/night cycle at 23.5/18.5 °C and 55 % relative humidity. After germination and formation of primary roots, the seedlings were transferred to polyurethane sponges (25 × 25 x 25 mm3) and from then on cultivated in sterilized modified Hoagland medium (pH = 5.5 ± 0.1, medium composition is listed in Table S1 in the Supporting Information), which was replaced weekly [21]. Aeration of plant roots and circulation of the medium was provided by aquarium pumps with connected bubble air stones.
2.2. Bioassociation Experiment
After four to eight weeks of plant growth – depending on plant species – hydroponic bioassociation experiments were conducted with 250 mL modified Hoagland medium with reduced phosphate concentration (1 µM KH2PO4) including the respective amount of a EuCl3 ∙ 6 H2O stock solution to obtain concentrations of 2, 20 and 200 µM. After initial sampling of the medium, the experiment was started by immersing the roots of a single plant per flask. The liquid in the flask was continuously aerated via a tube and an aquarium pump. Samples of the liquid medium were taken 20 min, 2.5 h, 24 h, 48 h, 72 h and 96 h after start of the experiment. As the liquid volume decreased during the experiment due to metabolism and transpiration of the plant as well as evaporation from the Erlenmeyer flask, the initial volume was topped up with phosphate-reduced medium without Eu(III) before each sampling. The experiment was terminated after 96 h by removing the roots from the Eu(III)-containing solution. Roots were washed three times for 5 min each with MilliQ water to remove residues of the medium and weakly sorbed Eu(III) from the root surface. Roots were blotted dry, samples of root tissue were taken for TRLFS and then roots and shoots (A. strigosa, T. incarnatum) and roots, stem and leaves (L. corniculatus, M. sativa, L. usitatissimum), respectively, were separated.
2.3. Digestion of Plant Material and Determination of Eu(Iii) Concentration in Plant Material
Upon drying of the individual plant parts at 80 °C for at least three days, dry masses were determined and the dry plant material was incinerated in a muffle furnace over night at 480 °C in quartz glass crucibles. The ash was digested in 2 mL concentrated HNO3, transferred to a volumetric flask and topped up with MilliQ water. Eu(III) concentration was measured by ICP-MS as triplicate by a NexION 350x (Perkin Elmer, USA) in an Ar plasma with Sc, Rh and Lu as internal standards. The Eu(III) content in the plant parts was then presented as the mean value of four independent biological replicates in relation to the dry mass.
2.4. Speciation Analysis by TRLFS and Parallel Factor Analysis (PARAFAC)
Liquid samples of the medium were pipetted in a quartz semi-microcuvette, while samples of plant tissue were placed in transparent 1.5 mL Eppendorf vials in the laser beam and analyzed at 25 °C. A diode-pumped Nd:YAG laser with an optical parametric oscillator (Ekspla NT230-50-SH/SF-SCU, Lithuania) and energy pulses of 1 - 2 mJ at an operating frequency of 50 Hz were used. The excitation wavelength was 394 nm. A Shamrock SR-303i-A spectrograph (Andor Technology, UK) coupled with an ICCD camera iStar DH320T-18U-63 cooled to -35 °C was controlled using Andor Solis software. An optical grating with 300 l/mm was used and the spectra were recorded with an initial delay of 12 μs in relation to the laser pulse in a spectral range of 500 to 780 nm over 200 accumulations. The time interval between the laser pulse and the start of spectrum acquisition was varied dynamically between 0 and 700 μs. A series of time-resolved spectra consisted of 21 individual spectra.
Afterwards, a PARAFAC analysis was performed to extract single component spectra, species distribution and the respective luminescence lifetimes as reported in Drobot et al. [22].
2.5. Chromatographic Analysis of Cultivation Media by HPLC
In order to investigate the root exudates from A. strigosa, the liquid medium was sampled after 96 h of root contact time. Due to very low intensities in the chromatograms, a separate experiment was conducted as follow. Three plant roots of A. strigosa, cultivated like described in section 2.1., were washed under running deionized water to remove residual medium from the roots and subsequently hung into 100 mL MilliQ water for 2 h to induce the secretion of root exudates, as reported in the literature [23]. Irrespective of the sampling method, the liquids were filtered through a 0.22 µm syringe filter and immediately analyzed by HPLC or stored at -20 °C until measurement. A negative control sample consisted of pure MilliQ filtered through a 0.22 µm syringe filter. Standards of fumaric, citric and malic acid were prepared as 10 mM stock solutions in MilliQ water and root exudate samples were spiked in such a way that final concentrations of 10, 250 and 250 µM, respectively, were obtained. Chromatography was performed on an HPLC system (1200 series, Agilent Technologies, USA) equipped with a sulphonated polystyrol/divinylbenzol resin column (Nucleogel ION 300 OA, Macherey-Nagel, Germany) and a diode array detector. The column was kept at 70 °C and the flow rate of 5 mM H2SO4 was set to 0.4 mL/min. Injection volume of the samples was varied between 50 and 100 µL.
2.6. Thermodynamic Modelling of Eu(III) Speciation in Hydroponic Solution
To further verify the experimental results with modeling, speciation calculations were carried out with PHREEQC Interactive version 3.7.3-15968. The presence of carbon dioxide in a free atmosphere was simulated and taken into account to reproduce the experimental conditions. The complexation constants were taken from Paul Scherrer Institute (PSI)/Nagra Chemical Thermodynamic Database/TDB 2020, Friedrich et al. [24], Heller et al. [25] and Taube et al. [26].
3. Results and Discussion
3.1. Eu(III) Concentration in Hydroponic Solution
Analysis of the Eu(III) concentration in the medium throughout the experiment was performed to obtain insights into the progress of Eu(III) removal from the solution. Different bioassociation kinetics were observed depending on the initial metal concentration in the medium, as shown in Figure 1. All curves demonstrate an instantaneous decrease in Eu(III) concentration upon immersing the roots in the metal-containing solution. This was especially pronounced for the low concentration flasks, in which 32 % (L. usitatissimum) up to 74 % (L. corniculatus) of the initial 2 µM Eu(III) were withdrawn from the medium within 20 min plant contact time. For the 200 µM flasks, between 4 % (L. usitatissimum) up to 38 % (L. corniculatus) of initial Eu(III) was removed within 20 min. This observation suggests the metabolism-independent process of biosorption, in which the Eu(III) is adsorbed physico-chemically to the root surface. Biosorption of metal ions on functional groups, such as carboxylate and phosphate groups, of living and dead biomass is well known for plant cells and higher plants and usually involves ion exchange mechanisms [27,28,29,30,31].
This initial concentration drop in the medium was followed by a further universal decrease of Eu(III) concentration. This trend can be attributed to Eu(III) being taken up by the root tissue. However, it is unclear whether this uptake occurred actively through ion channels into the symplast or passively through the apoplast, i.e. the intercellular space outside the plasma membrane. After 24 hours, the majority of the 2 µM Eu(III) was already bioassociated. Almost quantitative bioassociation was achieved in the 20 µM flasks after 24 h for L. corniculatus, after 48 h for T. incarnatum, and after 96 h for A. strigosa and M. sativa as well. In the case of L. usitatissimum, 7 ± 3 µM Eu(III) still remained in solution after 96 h. 200 µM Eu(III), however, were not entirely removed from the medium, irrespective of the plant species. Differences in the extent of Eu(III) removal by these were particularly evident in the high concentration approaches. L. usitatissimum removed the least Eu(III) from the solution, leaving 146 ± 15 µM in the flask, whereas L. corniculatus and T. incarnatum removed the most, leaving only 23 ± 16 µM and 13 ± 8 µM Eu(III) in the flask, respectively.
In general, plants with a higher biomass and more branched root system also showed a higher removal of Eu(III) from the medium. For the same growth period, different biomasses were obtained due to diverse plant morphologies. This means that L. usitatissimum, which had a taproot with few lateral roots, was able to bioassociate less Eu(III) than L. corniculatus, which had a strong taproot with a broad root crown (see Figure S2 in the Supporting Information). To account for these differences, the amount of bioassociated Eu(III) is related to the dry weight of the plant parts in the following section. Furthermore, the superficial loosely biosorbed fraction of Eu(III) will be washed off with MilliQ water to solely take into account the Eu(III) bound to or taken up into the root tissue.
3.2. Bioassociated Eu(III) in Plants
Roots were separated from shoots for incineration and subsequent acid digestion of the individual plant parts. In the case of M. sativa, L. corniculatus and L. usitatissimum, a further partitioning of shoots into stem and leaves was possible. For reasons of comparability, Figure 2 solely depicts data from roots and shoots. For detailed stem and leaves data, please refer to Figure S1 in the Supporting Information.
Quantification by ICP-MS revealed, that a higher initial Eu(III) concentration in the medium resulted in a higher mass of bioassociated Eu(III) in the respective plant parts. These findings agree with the soil-to-root/shoot relationships proposed by Kalis et al., in which the metal content in shoots is related to the metal content in roots, which in turn is related to the metal adsorption to the root surface, depending on the concentration in the soil solution [32]. Since we were working in a hydroponic model system without soil, our medium is considered equivalent to the soil solution in the proposed concept.
Although the initially applied Eu(III) concentrations in the medium were varied by order of magnitudes, the sum of bioassociated Eu(III) did not always reflect this proportionality. As an example, exposing M. sativa to 2 µM Eu(III) resulted in bioassociation values for roots and shoots of 219 ± 111 and 7 ± 4 µgEu/gDW, respectively. When applying ten times as much, i.e. 20 µM Eu(III), the bioassociated Eu(III) in the root (1692 ± 276 µgEu/gDW) almost scales by this factor, but not in shoots (10 ± 8 µgEu/gDW). This peculiarity becomes even more apparent when comparing with the result of the highest applied Eu(III) concentration, in which 5734 ± 591 and 45 ± 29 µgEu/gDW were found in roots and shoots, respectively. These data suggest the presence of selective uptake and distribution pathways which limit the bioassociation of non-essential elements like Eu(III) [33]. This can be achieved by deposition of the metal into vacuoles, chelation or precipitation at the cell wall or actively pumping the metal out of the cells again [34]. Furthermore, the Casparian strip prevents the translocation of non-essential metals and restricts the inward movement and further distribution of solutes and water through the xylem. Therefore, the majority of Eu(III), namely 95.1 % (L. usitatissimum, 200 µM) up to 99.8 % (T. incarnatum, 200 µM) remained in the roots, as emphasized in the bar charts in Figure 2. Nevertheless, a certain amount of Eu(III) was traced in green plant parts. This is due to the fact that the Casparian strip is not fully developed at roots tips and disrupted where lateral roots protrude from the parental roots. Hence, Eu(III) can enter the xylem without passing a cell membrane and is translocated upwards [35,36].
For plants that allowed separation of stem and leaves, Eu(III) was distributed differently between these tissues depending on the initial Eu(III) concentration, as illustrated in Figure S1 in the Supporting Information. M. sativa and L. usitatissimum accumulated more Eu(III) in the stem (58 ± 25 and 216 ± 136 µgEu/gDW, respectively) than in the leaves (10 ± 1 and 92 ± 38 µgEu/gDW, respectively) when exposed to 200 µM Eu(III). The cation exchange capacity of pectic substances in the cell walls of the xylem vessels is well known [37]. An association of Eu(III) with carboxyl and hydroxyl groups of cell walls of the vascular cylinder can therefore be a reason for accumulation in the stem of the examined plants.
For lower concentrations and in the case of L. corniculatus, Eu(III) was either equally distributed between stem and leaves or accumulated more in the leaves. When comparing the amount of bioassociated Eu(III) in aboveground tissue, L. usitatissimum accumulated the most Eu(III), irrespective of the initial concentration, in numbers 309 ± 174, 86 ± 32 and 24 ± 10 µgEu/gDW at 200, 20 and 2 µM Eu(III) initial concentration. When only roots are considered, L. usitatissimum accumulated the most Eu(III) at 20 and 2 µM initial Eu(III) concentration (3183 ± 1219 and 563 ± 145 µgEu/gDW, respectively) and in the 200 µM approach A. strigosa (14243 ± 4659 µgEu/gDW) and T. incarnatum (12345 ± 2810 µgEu/gDW) showed the highest Eu(III) bioassociation.
A quantitative insight into the bioassociation of Eu(III) on contact with plant roots has just been obtained, revealing its distribution in different tissues. This raises the question of the fate of bioassociated and residual Eu(III) in the medium in terms of its speciation. To answer this question, the metal’s speciation during the experiment was analyzed using TRLFS.
3.3. Speciation of Eu(III) in Liquid Medium
Knowledge of the speciation of Eu(III) in the medium as a function of exposure time provides insight into the bioavailability of the metal on the one hand, and on the other hand allows to observe plant-induced alterations of the Eu(III)-containing medium during the experiment. For this purpose, the liquid medium was analyzed after each sampling by TRLFS and the measured sum spectra were later deconvoluted by the means of PARAFAC. The interpretation of the results was further supported with thermodynamic modelling of Eu(III) speciation in the liquid medium.
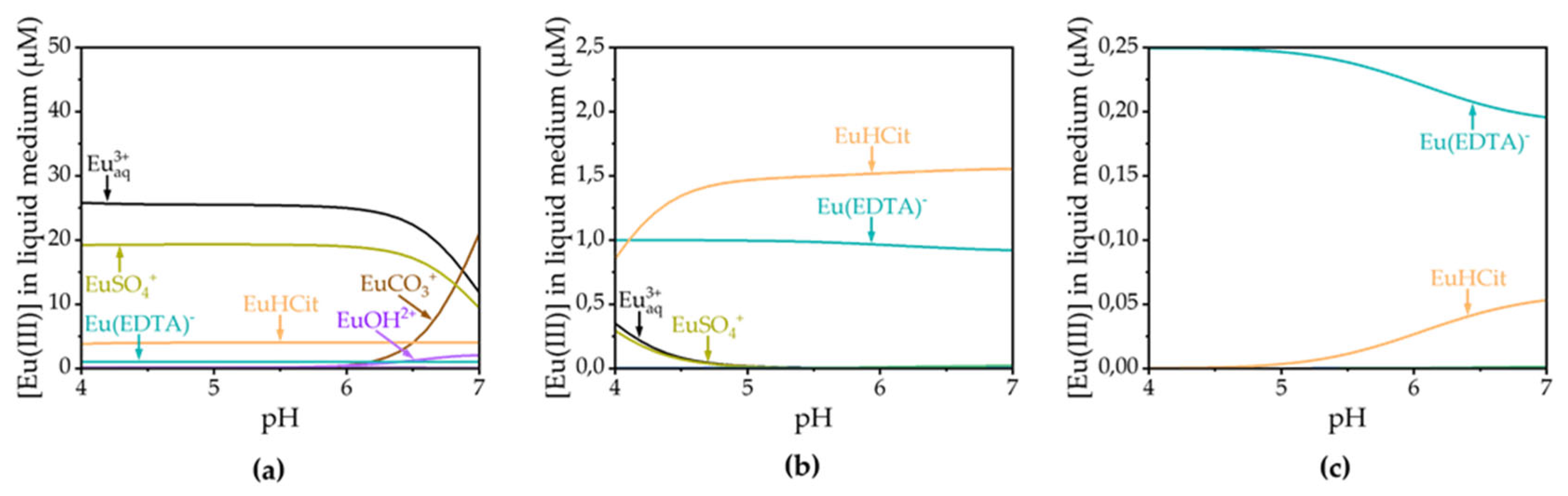
A comparable alteration in the Eu(III) luminescence spectra was observed over time in the liquid medium across various plant species, as can be seen in Figure S3 in the Supporting Information. The spectra are all characterized by an intensity increase of the 5D0 → 7F0 and the 5D0 → 7F2 transitions, a change of spectral characteristics, especially the 5D0 → 7F4 transition at low concentrations, and variance of luminescence lifetime as experiment time progresses. The lower the initial Eu(III) concentration, the faster the speciation change occurred. Luminescence spectra of the medium in contact with M. sativa are depicted in Figure 3, representatively. For the 200 and 20 µM approach, the Eu(III)-aquo ion dominates the speciation before plant contact according to thermodynamic modelling. Given a F2/F1 ratio around 0.4 and a luminescence lifetime of τ0 = 110 µs, this is in good agreement with the recorded spectra and values from the literature [38]. Though Eu(EDTA)- is the predominant Eu(III) species in the 2 µM approach according to thermodynamic modelling, the initial Eu(III) luminescence spectra are still dominated by the Eu(III)-aquo species. This is due to the fact that EDTA is initially bound to Fe(III) and hence not available for Eu(III) complexation, which is neglected in the thermodynamic database. Contributions by other complexing medium components are not observable in the spectra of the 200 µM approach, although e.g. EuSO4+ plays a role in the modelled speciation. The contribution of EuSO4+ and Eu(EDTA)- to the modelled speciation becomes even more pronounced in the 20 and 2 µM experiments and simultaneously becomes apparent by a slightly increased luminescence lifetime and F2/F1 ratio for 20 and 2 µM samples in comparison.
As a result of the contact of the medium with plant roots, a speciation change becomes recognizable. In order to clearly separate the species contributing to the sum spectra shown in Figure 4, a deconvolution by the means of PARAFAC was done with the spectral data.
In Figure 4, PARAFAC evaluation results of the 200 µM Eu(III) liquid medium in experiments with M. sativa are shown, representatively. Based on characteristic features in peak splitting, shape and intensity as well as different luminescence decay curves, sum spectra were deconvoluted into single components. As already stated before, the short-lived Eu(III)-aquo species is the dominating species at the start of the experiment. A second, longer-lived intermediate species (Species 2) arises within the first hours of the experiment. While the fraction of the Eu(III)-aquo species decreases with time, species 2 gains intensity until 72 h. At the end of the bioassociation time, a third species with a notably longer luminescence lifetime dominates the spectrum. In control flasks without plant material, no such speciation change was observable and the Eu(III)-aquo species persisted until the end of the experiment. This allows the occurrence of species 2 and 3 to be considered as a plant-related complexation of the Eu(III) ion. Various low molecular weight organic compounds are continuously and passively released into the rhizosphere [39]. Among these root exudates, organic and amino acids show great potential to induce a speciation change of Eu(III) due to their affinity to trivalent REE ions [40,41]. In section 3.5, this issue will further be evaluated.
3.4. Speciation of Bioassociated Eu(III)
In order to obtain not only a quantitative but also a qualitative insight into Eu(III) bioassociation, plant parts exposed to various Eu(III) concentrations were analyzed by laser spectroscopy at the end of the experiment (96 h). The luminescence spectra of the roots of each plants are visualized in Figure 5.
Although some Eu(III) luminescence signals were detected in the shoot, their intensity and signal-to-noise ratio (S/N) were too low to allow meaningful analysis due to the low concentration of the metal in this part of the plant and high background intensities.
Although different plant species from several plant families have been used experimentally, the luminescence spectra of root-associated Eu(III) are similar, especially at 200 and 20 µM initial concentration. The luminescence lifetimes range between 133 and 269 µs, depending on plant species and applied Eu(III) concentration in the medium. All spectra all characterized by a 5D0 → 7F2/5D0 → 7F1 ratio (F2/F1) > 1, which demonstrates that it is not the Eu(III)-aquo ion being present whose F2/F1 ratio is < 1. Notably, the intensity of the 5D0 → 7F2 band is higher for roots exposed to 2 µM. A possible explanation for this observation is, that the lower total amount of Eu(III) is strongly bound to specific sites rather than being unspecifically sorbed as in the presence of an excess of Eu(III).
Due to the large number of possible binding partners and subcellular processes to which Eu(III) is subjected to, no clear statement on speciation can be made on the basis of the available data. However, it is very likely that the recorded luminescence spectra are sum spectra of several individual Eu(III) species, as it has been shown in other studies with plant cells and entire plants [29,31,42]. To elucidate different binding sites and species, further investigations with chemical microscopy and electron microscopy in combination with energy-dispersive X-ray spectroscopy are necessary.
3.5. Identification of Root Exudates
As demonstrated in the species distribution in Figure 4, plants induced an alteration of Eu(III) speciation with time. Based on spectroscopic data, no unambiguous identification can be made. Therefore, HPLC of the liquid medium before and after plant contact was employed in search for possible complexing compounds. Using the HPLC setup mentioned in Section 2.5, no signals were detected in the Eu(III)-containing phosphate-reduced medium. After the end of the experiment, however, several signals at retention times between 9 and 31 min were clearly visible, though with low intensity (see Figure S4 in the Supporting Information). As only phosphate-reduced medium was added to refill the flask during the experiment, the signals detected can be attributed to metabolites exuded by plants, i.e. root exudates.
Root exudate composition can be complex, but usually contains sugars, amino acids, extracellular enzymes, phenolics and organic acids [10]. Especially intermediate products of the Krebs cycle, such as citric, malic, succinic and fumaric acid have been identified as root exudates for various plants [43,44,45,46]. In a natural system, they serve to solubilize essential metals in soil or enhance their bioavailability, but can be responsible for an involuntary uptake of An and Ln in the same way. Knowledge about their occurrence is therefore desirable to accurately implement these compounds in thermodynamic calculations.
Consequently, roots of three plants of A. strigosa were hung into MilliQ water in order to collect root exudates in a separate experiment. In contrast to the negative control, a number of peaks were observed in the MilliQ water loaded with exudate. By comparison of the chromatograms with references of multiple organic acids, we were able to match the signals in the sample with the retention times of citric, malic and fumaric acid, as shown in Figure 6. We then checked whether the limit of detection (LOD), which is defined in the European Pharmacopoeia as the concentration of a component providing a S/N of three, was exceeded [47]. For the peaks at 13.79, 16.66 and 23.45 min we determined S/N of 3.2, 5.8 and 4.8, respectively, which are all above the LOD. Based on the overlap of signals with known references, we can conclude that citrate, malate and fumarate are present in root exudates with statistical significance. Unfortunately, the signal intensity did not allow for a reasonable quantification of root exudates, since this would require S/N > 10. However, based on our experience and three-point calibrations with the above references, we hypothesize a low micromolar range for those organic acids.
In light of the evidence indicating the presence of root exudates and the observed decrease in Eu(III) concentration due to bioassociation processes, our objective was to ascertain whether these factors could be responsible for the speciation change as observed in TRLFS (see Figure 3). Therefore, we set up another thermodynamic modelling of Eu(III) speciation in the liquid medium after 96 h. Based on experience, we assumed fumaric, citric and malic acid concentrations to be 0.4, 4 and 40 µM and derived final Eu(III) concentrations of 0.25, 2.5 and 50 µM from data shown in Figure 1. The results of the modelling are shown in Figure 7.
It becomes evident, that root exudates have an influence on Eu(III) speciation in the liquid medium in the examined concentration ranges, especially citric acid. While the influence on the speciation in the experiment with the highest Eu(III) concentration is minor, the EuHCit species becomes the dominant one in the 20 µM approach. From the diagrams shown below one can additionally derive the importance of EDTA as a chelating agent in the medium, as already stated in Section 3.3. Interestingly, the luminescence lifetime of species 3 (τ3 = 162 µs) of the PARAFAC evaluation is in good agreement with the EuCitH luminescence lifetime reported in the literature (τEuCitH = 162 µs at pH = 5.5) [25]. The Eu(III) speciation change observed throughout the experiment can therefore be partly attributed to the release of root exudates.
5. Conclusions
In the context of phytoremediation, various agricultural plants were tested for their ability to bioassociate Eu(III) in hydroponic culture. Bioassociation began with an immediate biosorption upon root exposure to the Eu(III)-amended medium and continued with uptake into the root tissue thereafter. The concentration of Eu(III) in the plant tissues increased with the initial concentration of Eu(III). Over 95 % of the bioassociated Eu(III) was retained in the roots, although transport to aboveground tissues was observed in all plants. In relation to dry plant weight, A. strigosa and T. incarnatum accumulated the highest amount of Eu(III) at high concentrations (200 µM), while L. usitatissimum showed the highest accumulation of Eu(III) at lower concentrations in the medium (20, 2 µM). Currently, there is limited information on threshold concentrations for REE hyperaccumulators in the literature.
With regard to threshold concentrations for heavy metal hyperaccumulation (1000 up to 10,000 µg/g dry weight of the shoots, depending on the metal), the plants analyzed herein are neither hyperaccumulators nor excluders, though L. usitatissimum was found to accumulate up to 362 µgEu/gDW in the shoots [48]. Rather, the plants used can be considered indicator plants (group 1 in section 1) , as the metal content in their tissue is proportional to the concentration of bioavailable metal in the growth substrate [13,14].
We further assessed Eu(III) speciation in plant roots and the liquid medium to understand systemic and plant-induced metal complexation throughout the experiment. While Eu(III) speciation before plant contact is dictated by the Eu(III)-aquo ion and minor contributions from medium components such as SO42- and EDTA4-, a speciation change occurred when plant roots were introduced. This change was more pronounced for low Eu(III) concentration samples. PARAFAC deconvolution separated several distinct Eu(III) species which evolved with time and comprise different spectral characteristics and luminescence lifetimes. Analysis of the liquid medium by HPLC revealed the presence of root exudates. In a separate experiment, these organic compounds were identified as citrate, malate and fumarate, which are released by the roots. As substituted by thermodynamic modelling, especially citric acid is partially responsible for a complexation of the remaining Eu(III) in solution.
A quantitative description of Eu(III) distribution in plants was obtained, along with a qualitative insight into Eu(III) speciation in the medium. Root exudates relevant for thermodynamic models of metal biogeochemistry were also identified. It is evident that plant metabolites must be taken into account when predicting Ln and An mobility in natural systems. Further studies in model and natural systems are needed to understand the influence of rhizosphere processes on metal association. The data presented can be valuable in filling knowledge gaps in metal biogeochemistry. The results obtained with Eu(III) can be applied to model the behavior of Ln(III), Cm(III) and Am(III) in the environment.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, M.K., R.S., J.R.; methodology, M.K., R.S., M.V., B.D. ; software, B.D.; validation, M.K.; formal analysis, M.K. and V.D.; investigation, M.K.; resources, T.S.; data curation, M.K.; writing—original draft preparation, M.K.; writing—review and editing, R.S., V.D., M.V., T.S.; visualization, M.K.; supervision, R.S.; project administration, J.R.; funding acquisition, J.R.
Funding
The research was funded by the Federal Ministry of Education and Research as a part of the joint project ‘RENA’ (Biological remediation by natural association processes) under contract numbers 02NUK066A and 02NUK066D and by the Horizon 2020 Euroatom research and training program as a part of the joint project ‘RadoNorm’ (Towards effective radiation protection based on improved scientific evidence and social considerations – focus on Radon and NORM’) under grant agreement No. 900009.
Data Availability Statement
The data presented in this study are openly available in RODARE at 10.14278/rodare.3027, reference number 3027.
Acknowledgments
The authors thank S. Beutner for multiple ICP-MS and F. Lehmann for HPLC measurements.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rajačić, M.M.; Sarap, N.B.; Janković, M.M.; Nikolić, J.D.; Todorović, D.J.; Pantelić, G.K. Radioactivity in Chemical Fertilizers.
- Zifferero, M. A Post-Chernobyl View. In Radionuclides in the Food Chain; Harley, J.H., Schmidt, G.D., Silini, G., Eds.; Springer: London, 1988. [Google Scholar]
- Yemisi Olobatoke Manny Mathuthu, R.; Vet Anim Sci, T.J.; Yemisi OLOBATOKE, R.; Mathuthu, M. Radionuclide Exposure in Animals and the Public Health Implications. Turk J Vet Anim Sci 39. [CrossRef]
- Dinis, M.; Future, A.F.-M.S.A.C. for the; 2007, undefined Exposure Assessment to Radionuclides Transfer in Food Chain. Springer 2005.
- Luo, J.; Qi, S.; Gu, X.W.S.; Wang, J.; Xie, X. Evaluation of the Phytoremediation Effect and Environmental Risk in Remediation Processes under Different Cultivation Systems. J Clean Prod 2016, 119, 25–31. [Google Scholar] [CrossRef]
- Hu, C.; Ou, Y.; Zhang, D.; Zhang, H.; Yan, C.; Zhao, Y.; Zheng, Z. Phytoremediation of the Polluted Waigang River and General Survey on Variation of Phytoplankton Population. Environmental Science and Pollution Research 2012, 19, 4168–4175. [Google Scholar] [CrossRef]
- Rulkens, W.; Tichy, R.; Technology, J.G.-W.S. and; 1998, undefined Remediation of Polluted Soil and Sediment: Perspectives and Failures. Elsevier.
- Salt, D.E.; Blaylock, M.; Kumar, N.P.B.A.; Dushenkov, V.; Ensley, B.D.; Chet, I.; Raskin, I. Phytoremediation: A Novel Strategy for the Removal of Toxic Metals from the Environment Using Plants. Bio/Technology 1995 13:5 1995, 13, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Pinto, E.; Aguiar, A.A.R.M.; Ferreira, I.M.P.L.V.O. Influence of Soil Chemistry and Plant Physiology in the Phytoremediation of Cu, Mn, and Zn. CRC Crit Rev Plant Sci 2014, 33, 351–373. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic press, 1995.
- Colangelo, E.P.; Guerinot, M. Lou Put the Metal to the Petal: Metal Uptake and Transport throughout Plants. Curr Opin Plant Biol 2006, 9, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Corso, M.; García De La Torre, V.S. Biomolecular Approaches to Understanding Metal Tolerance and Hyperaccumulation in Plants. Metallomics 2020, 12, 840–859. [Google Scholar] [CrossRef] [PubMed]
- Leitenmaier, B.; Küpper, H. Compartmentation and Complexation of Metals in Hyperaccumulator Plants. Front Plant Sci 2013, 4, 55114. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.J.M. Accumulators and Excluders - Strategies in the Response of Plants to Heavy Metals. J Plant Nutr 1981, 3, 643–654. [Google Scholar] [CrossRef]
- Pajević, S.; Borišev, M.; Rončević, S.; Vukov, D.; Igić, R. Heavy Metal Accumulation of Danube River Aquatic Plants - Indication of Chemical Contamination. Cent Eur J Biol 2008, 3, 285–294. [Google Scholar] [CrossRef]
- Lux, A.; Martinka, M.; Vaculík, M.; White, P.J. Root Responses to Cadmium in the Rhizosphere: A Review. J Exp Bot 2011, 62, 21–37. [Google Scholar] [CrossRef]
- Van Hoof, N.A.L.M.; Koevoets, P.L.M.; Hakvoort, H.W.J.; Bookum, W.M. Ten; Schat, H.; Verkleij, J.A.C.; Ernst, W.H.O. Enhanced ATP-dependent Copper Efflux across the Root Cell Plasma Membrane in Copper-tolerant Silene Vulgaris. Wiley Online LibraryNALM Van Hoof, PLM Koevoets, HWJ Hakvoort, WM Ten Bookum, H Schat, JAC VerkleijPhysiologia Plantarum, 2001•Wiley Online Library 2001, 113, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Zhou, Q.; Wang, X. Identification of Weed Plants Excluding the Uptake of Heavy Metals. Environ Int 2005, 31, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Brooks, R.R. Geobotany and Hyperaccumulators. In Plants that Hyperaccumulate Heavy Metals; CAB International: Wallingford, UK, 1998; pp. 55–94. [Google Scholar]
- Wang, X.P.; Shan, X.Q.; Zhang, S.Z.; Wen, B. Distribution of Rare Earth Elements among Chloroplast Components of Hyperaccumulator Dicranopteris Dichotoma. Anal Bioanal Chem 2003, 376, 913–917. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. Circular. California Agricultural Experiment Station 1950, 347. [Google Scholar]
- Drobot, B.; Steudtner, R.; Raff, J.; Geipel, G.; Brendler, V.; Tsushima, S. Combining Luminescence Spectroscopy, Parallel Factor Analysis and Quantum Chemistry to Reveal Metal Speciation – a Case Study of Uranyl( vi ) Hydrolysis. Chem Sci 2015, 6, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Khorassani, R.; Hettwer, U.; Ratzinger, A.; Steingrobe, B.; Karlovsky, P.; Claassen, N. Citramalic Acid and Salicylic Acid in Sugar Beet Root Exudates Solubilize Soil Phosphorus. BMC Plant Biol 2011, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, S.; Sieber, C.; Drobot, B.; Tsushima, S.; Barkleit, A.; Schmeide, K.; Stumpf, T.; Kretzschmar, J. Eu(III) and Cm(III) Complexation by the Aminocarboxylates NTA, EDTA, and EGTA Studied with NMR, TRLFS, and ITC—An Improved Approach to More Robust Thermodynamics. Molecules 2023, 28, 4881. [Google Scholar] [CrossRef] [PubMed]
- Heller, A.; Barkleit, A.; Foerstendorf, H.; Tsushima, S.; Heim, K.; Bernhard, G. Curium( Iii ) Citrate Speciation in Biological Systems: A Europium( Iii ) Assisted Spectroscopic and Quantum Chemical Study. Dalton Transactions 2012, 41, 13969–13983. [Google Scholar] [CrossRef] [PubMed]
- Taube, F.; Drobot, B.; Rossberg, A.; Foerstendorf, H.; Acker, M.; Patzschke, M.; Trumm, M.; Taut, S.; Stumpf, T. Thermodynamic and Structural Studies on the Ln(III)/An(III) Malate Complexation. Inorg Chem 2019, 58, 368–381. [Google Scholar] [CrossRef]
- Diniz, V.; Volesky, B. Biosorption of La, Eu and Yb Using Sargassum Biomass. Water Res 2005, 39, 239–247. [Google Scholar] [CrossRef]
- Vogel, M.; Günther, A.; Rossberg, A.; Li, B.; Bernhard, G.; Raff, J. Biosorption of U(VI) by the Green Algae Chlorella Vulgaris in Dependence of PH Value and Cell Activity. 2010. [CrossRef]
- Klotzsche, M.; Vogel, M.; Sachs, S.; Raff, J.; Stumpf, T.; Drobot, B.; Steudtner, R. How Tobacco (Nicotiana Tabacum) BY-2 Cells Cope with Eu(III) – A Microspectroscopic Study. Analyst 2023. [Google Scholar] [CrossRef] [PubMed]
- Jessat, J.; Sachs, S.; Moll, H.; John, W.; Steudtner, R.; Hübner, R.; Bok, F.; Stumpf, T. Bioassociation of U(VI) and Eu(III) by Plant (Brassica Napus) Suspension Cell Cultures - A Spectroscopic Investigation. Environ Sci Technol 2021, 55, 6718–6728. [Google Scholar] [CrossRef] [PubMed]
- Jessat, J.; John, W.A.; Moll, H.; Vogel, M.; Steudtner, R.; Drobot, B.; Hübner, R.; Stumpf, T.; Sachs, S. Localization and Chemical Speciation of Europium(III) in Brassica Napus Plants. Ecotoxicol Environ Saf 2023, 254, 114741. [Google Scholar] [CrossRef] [PubMed]
- Kalis, E.J.J.; Temminghoff, E.J.M.; Visser, A.; Van Riemsdijk, W.H. Metal Uptake by Lolium Perenne in Contaminated Soils Using a Four-Step Approach. Environ Toxicol Chem 2007, 26, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Leavitt, S.W.; Dueser, R.D.; Goodell, H.G. Plant Regulation of Essential and Non-Essential Heavy Metals. J Appl Ecol 1979, 16, 203. [Google Scholar] [CrossRef]
- Antonovics, J.; Bradshaw, A.; Turner, R. Heavy Metal Tolerance in Plants. Adv Ecol Res 1971. [Google Scholar]
- Tanton, T.W.; Crowdy, S.H. The Distribution of Lead Chelate in the Transpiration Stream of Higher Plants. Pestic Sci 1971, 2, 211–213. [Google Scholar] [CrossRef]
- Haynes, R.J. Ion Exchange Properties of Roots and Ionic Interactions within the Root Apoplasm: Their Role in Ion Accumulation by Plants. The Botanical Review 1980, 46, 75–99. [Google Scholar] [CrossRef]
- Knight, A.; Crooke, W.; Inkson, R. Cation-Exchange Capacities of Tissues of Higher and Lower Plants and Their Related Uronic Acid Contents. Nature 1961. [Google Scholar] [CrossRef]
- Kimura, T.; Choppin, G.R.; Kato, Y.; Yoshida, Z. Determination of the Hydration Number of Cm(III) in Various Aqueous Solutions; 1996; Vol. 72;
- Nguyen, C. Rhizodeposition of Organic C by Plant: Mechanisms and Controls. In Sustainable Agriculture; Springer: Dordrecht, 2009; ISBN 978-90-481-2666-8. [Google Scholar]
- Wang, Z.; Zhang, S.; Shan, X.Q. Effects of Low-Molecular-Weight Organic Acids on Uptake of Lanthanum by Wheat Roots. Plant Soil 2004, 261, 163–170. [Google Scholar] [CrossRef]
- Liu, C.; Ding, T.X.; Liu, W.S.; Tang, Y.T.; Qiu, R.L. Phosphorus Mediated Rhizosphere Mobilization and Apoplast Precipitation Regulate Rare Earth Element Accumulation in Phytolacca Americana. Plant Soil 2023, 483, 697–709. [Google Scholar] [CrossRef]
- Stadler, J.; Vogel, M.; Steudtner, R.; Drobot, B.; Kogiomtzidis, A.L.; Weiss, M.; Walther, C. The Chemical Journey of Europium(III) through Winter Rye (Secale Cereale L.) – Understanding through Mass Spectrometry and Chemical Microscopy. Chemosphere 2023, 313, 137252. [Google Scholar] [CrossRef] [PubMed]
- Lipton, D.; Blanchar, R.; Physiology, D.B.-P.; 1987, undefined Citrate, Malate, and Succinate Concentration in Exudates from P-Sufficient and P-Stressed Medicago Sativa L. Seedlings. academic.oup.comDS Lipton, RW Blanchar, DG BlevinsPlant Physiology, 1987•academic.oup.com 1987, 85, 315–317.
- Hoffland, E.; Findenegg, G.R.; Nelemans, J.A. Solubilization of Rock Phosphate by Rape - II. Local Root Exudation of Organic Acids as a Response to P-Starvation. Plant Soil 1989, 113, 161–165. [Google Scholar] [CrossRef]
- Ohwaki, Y.; Hirata, H.; Ohwaki, Y. Differences in Carboxylic Acid Exudation among P-Starved Leguminous Crops in Relation to Carboxylic Acid Contents in Plant Tissues and Phospholipid Level in Roots. Soil Sci Plant Nutr 1992, 38, 235–243. [Google Scholar] [CrossRef]
- Ae, N.; Arihara, J.; Okada, K.; Yoshihara, T.; Johansen, C.; Link, R.F. Phosphorous Uptake by Pigeon Pea and Its Role in Cropping Systems of the Indian Subcontinent. J. Fac. Sci. Hokkaido Univ. Ser 1990, 34, 3. [Google Scholar]
- Keitel, S. Pharmacopoeial Standards: European Pharmacopoeia. In Encyclopedia of Pharmaceutical Science and Technology, Fourth Edition; CRC Press, 2013; pp. 2691–2703 ISBN 9781351124874.
- Baker, A.; phytologist, S.W.-N. ; 2002, undefined In Search of the Holy Grail: A Further Step in Understanding Metal Hyperaccumulation? JSTORAJM Baker, SN WhitingNew phytologist, 2002•JSTOR 2002, 155, 1–4. [Google Scholar] [CrossRef]
Figure 1.
Eu(III) concentration in the liquid medium during bioassociation experiments for different plant species. The color code in the legend applies for all graphs. (a) 2 µM initial Eu(III) concentration; (b) 20 µM initial Eu(III) concentration; (c) 200 µM initial Eu(III) concentration.
Figure 1.
Eu(III) concentration in the liquid medium during bioassociation experiments for different plant species. The color code in the legend applies for all graphs. (a) 2 µM initial Eu(III) concentration; (b) 20 µM initial Eu(III) concentration; (c) 200 µM initial Eu(III) concentration.
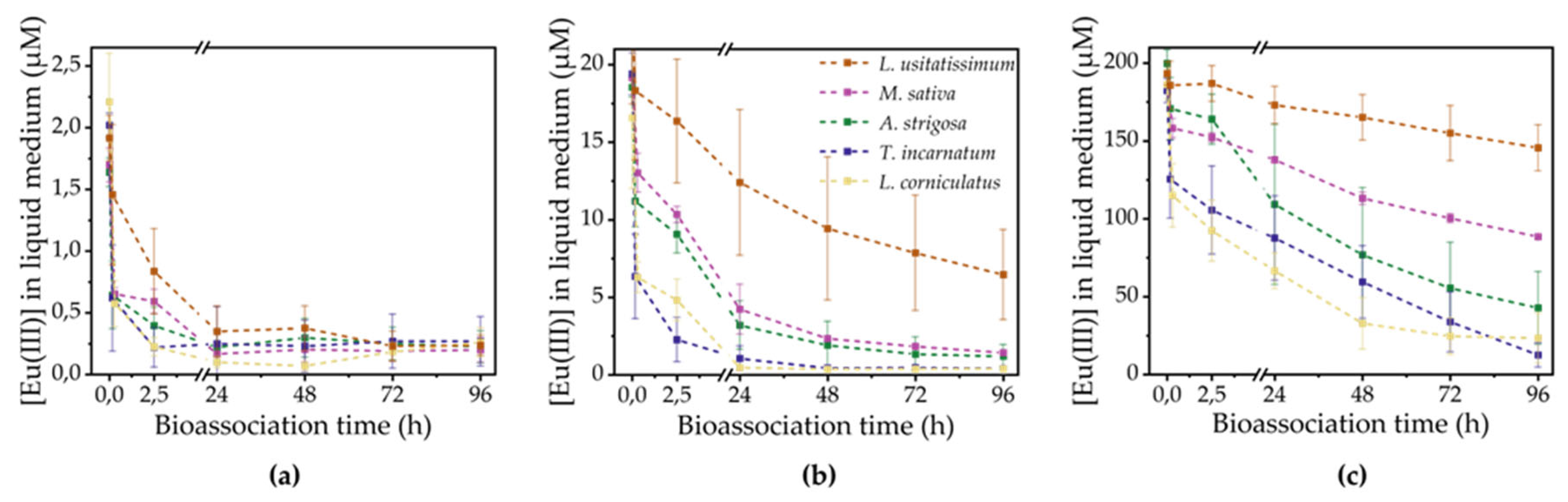
Figure 2.
Quantification of bioassociated Eu(III) in roots and shoots after 96 h exposure of plant roots with initial concentrations of 200, 20 and 2 µM Eu(III) in the liquid medium. Data show Eu(III) in roots and shoots of (a) A. strigosa, (b) T. incarnatum, (c) M. sativa, (d) L. corniculatus and (e) L. usitatissimum, respectively. For (c), (d) and (e), further data on the partitioning of stem from leaves is available in the Supporting Information. Bars represent the mean and error bars the standard deviation of four biological replicates. .
Figure 2.
Quantification of bioassociated Eu(III) in roots and shoots after 96 h exposure of plant roots with initial concentrations of 200, 20 and 2 µM Eu(III) in the liquid medium. Data show Eu(III) in roots and shoots of (a) A. strigosa, (b) T. incarnatum, (c) M. sativa, (d) L. corniculatus and (e) L. usitatissimum, respectively. For (c), (d) and (e), further data on the partitioning of stem from leaves is available in the Supporting Information. Bars represent the mean and error bars the standard deviation of four biological replicates. .
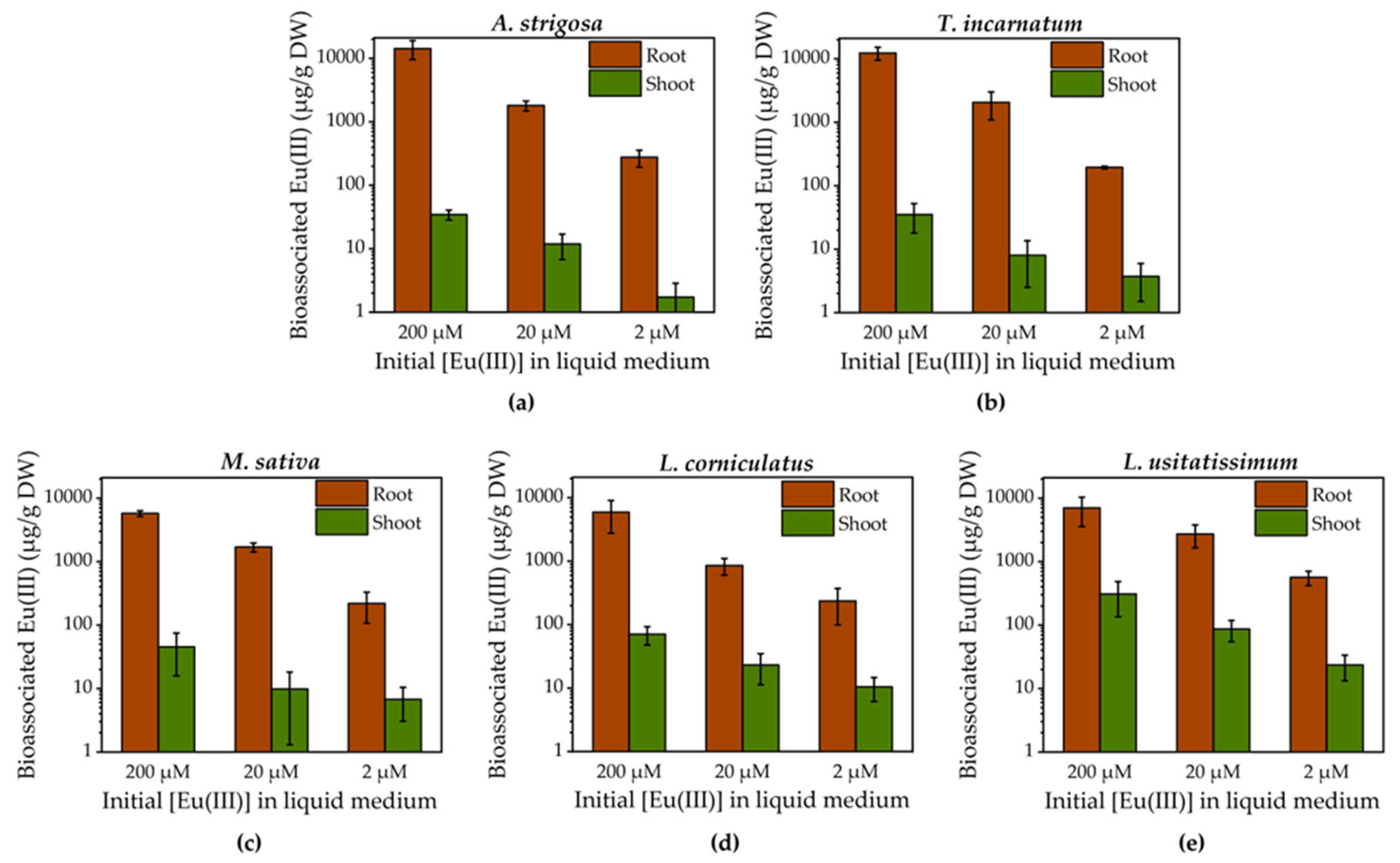
Figure 3.
Normalized luminescence spectra of liquid medium throughout the bioassociation experiment with M. sativa and initial Eu(III) concentrations of (a) 2 µM, (b) 20 µM and (c) 200 µM.
Figure 3.
Normalized luminescence spectra of liquid medium throughout the bioassociation experiment with M. sativa and initial Eu(III) concentrations of (a) 2 µM, (b) 20 µM and (c) 200 µM.
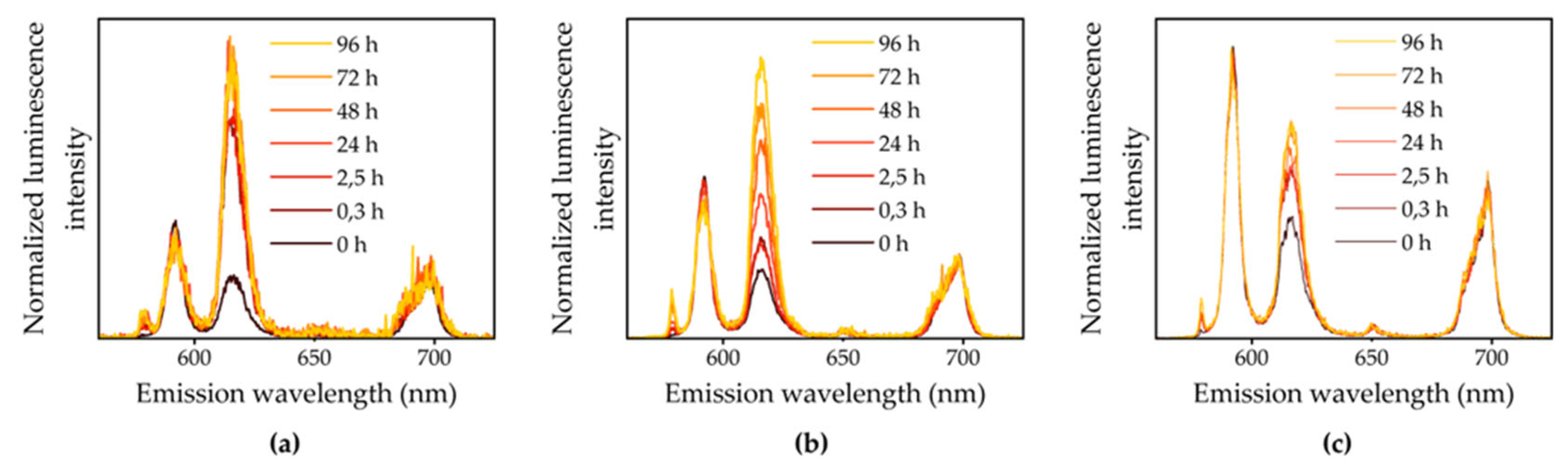
Figure 4.
Peak deconvolution results from TRLFS data of the liquid medium (200 µM Eu(III)) collected throughout a bioassociation experiment over 96 h with M. sativa. (a) Normalized single component spectra of Eu(III) species present in the liquid medium; (b) calculated species distribution over time based on luminescence intensities and total Eu(III) medium concentration (semi-transparent, see also Fig. 1); (c) Luminescence decay curves of single components with respective luminescence lifetimes.
Figure 4.
Peak deconvolution results from TRLFS data of the liquid medium (200 µM Eu(III)) collected throughout a bioassociation experiment over 96 h with M. sativa. (a) Normalized single component spectra of Eu(III) species present in the liquid medium; (b) calculated species distribution over time based on luminescence intensities and total Eu(III) medium concentration (semi-transparent, see also Fig. 1); (c) Luminescence decay curves of single components with respective luminescence lifetimes.
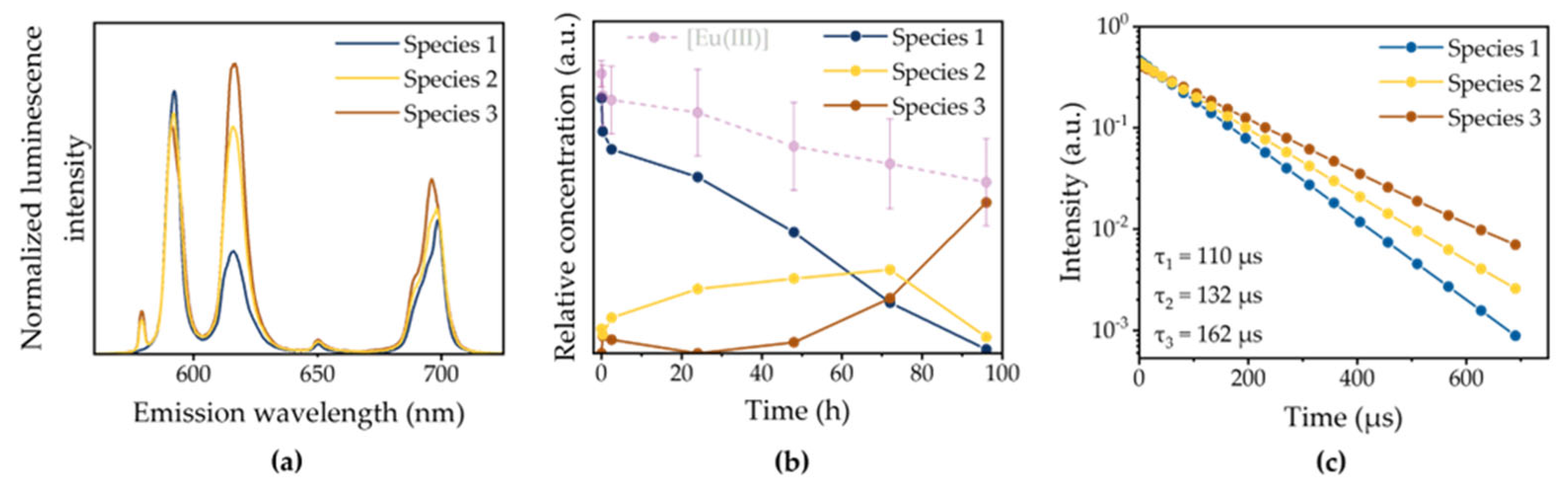
Figure 5.
Normalized luminescence spectra of plant roots after exposure to 200, 20 and 2 µM Eu(III) in the liquid medium for 96 h. Excitation wavelength giving rise to Eu(III) luminescence was 394 nm. Spectra were background corrected, if necessary, and normalized to the area of the 5D0 → 7F1 transition.
Figure 5.
Normalized luminescence spectra of plant roots after exposure to 200, 20 and 2 µM Eu(III) in the liquid medium for 96 h. Excitation wavelength giving rise to Eu(III) luminescence was 394 nm. Spectra were background corrected, if necessary, and normalized to the area of the 5D0 → 7F1 transition.
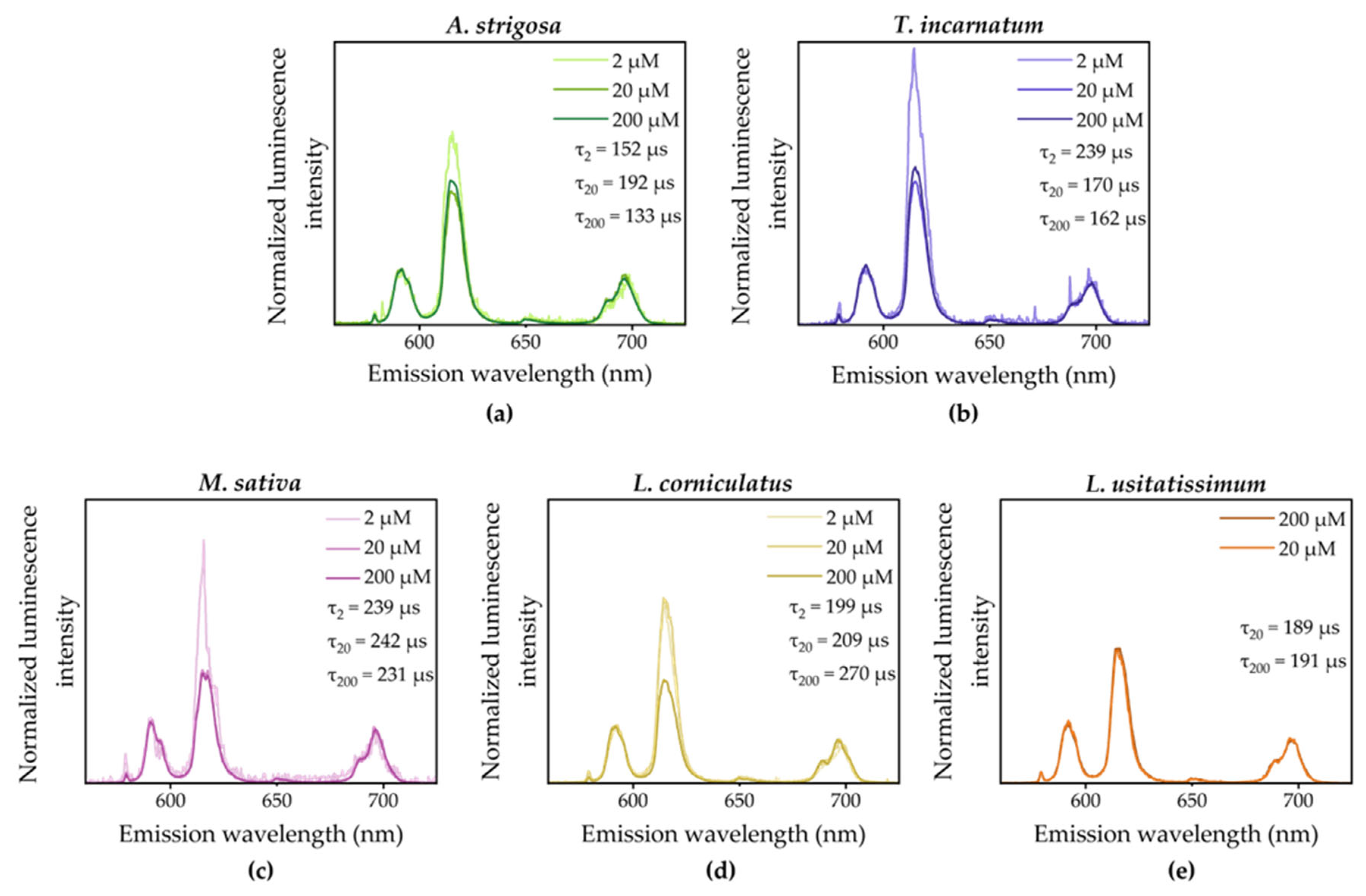
Figure 6.
Chromatograms of root exudate samples from A. strigosa and root exudate samples spiked with 250 µM citrate, 250 µM malate and 10 µM fumarate. The latter were normalized to 1 in the range considered. References of malate always include fumarate as a factory impurity, which become detectable even at very low concentrations due to its large absorption coefficient. Negative control and root exudate sample are shown in their original intensities. Chromatograms were cut at 8.5 min (after breakthrough peak) and after 35 min, because no relevant signals were detected outside that range.
Figure 6.
Chromatograms of root exudate samples from A. strigosa and root exudate samples spiked with 250 µM citrate, 250 µM malate and 10 µM fumarate. The latter were normalized to 1 in the range considered. References of malate always include fumarate as a factory impurity, which become detectable even at very low concentrations due to its large absorption coefficient. Negative control and root exudate sample are shown in their original intensities. Chromatograms were cut at 8.5 min (after breakthrough peak) and after 35 min, because no relevant signals were detected outside that range.
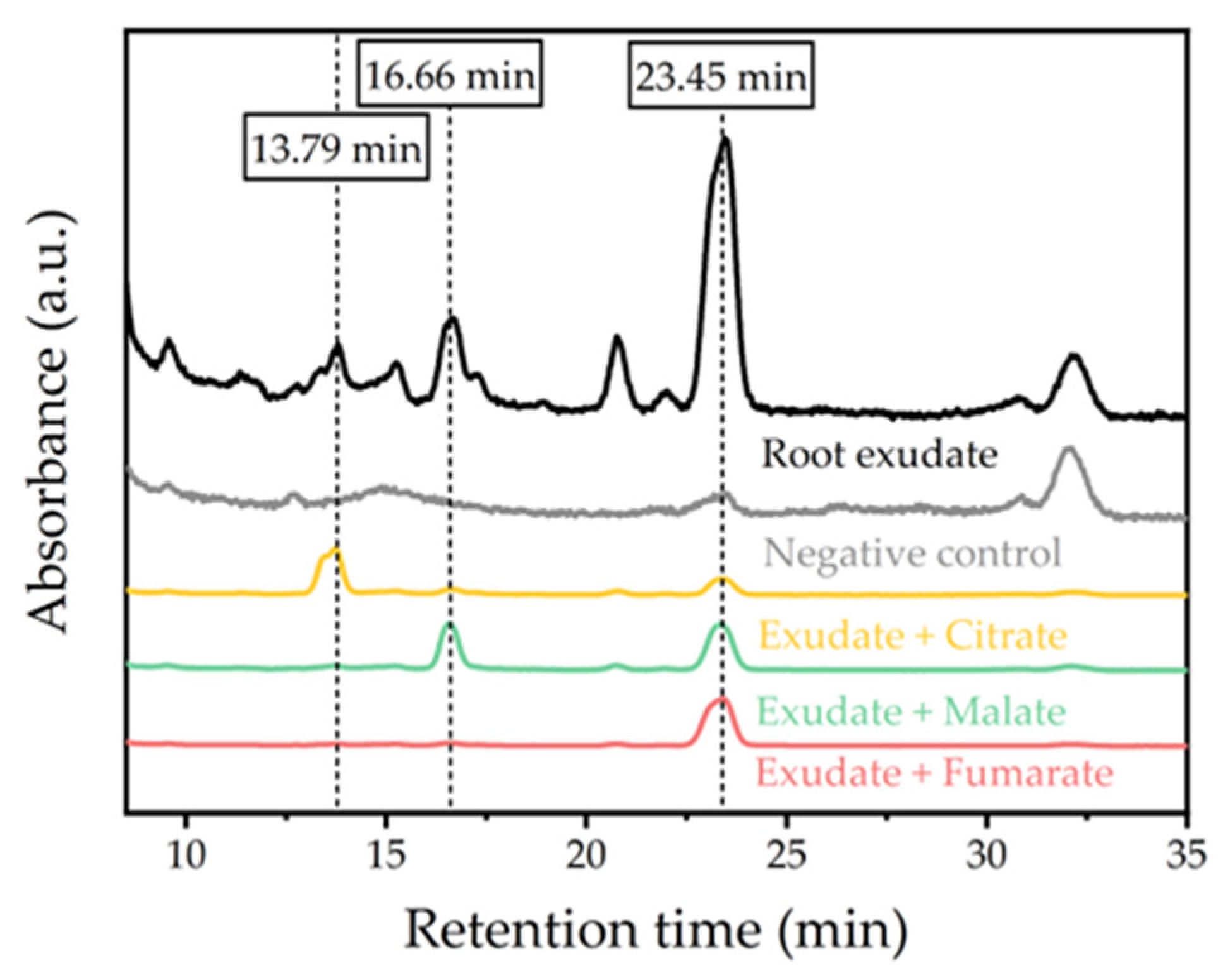
Figure 7.
Calculated speciation of the remaining Eu(III) in presence of root exudates (fumaric, citric and malic acid) in the liquid medium after 96 h in dependence of the pH value for (a) the 200 µM experiment; (b) the 20 µM experiment and (c) the 2 µM experiment.
Figure 7.
Calculated speciation of the remaining Eu(III) in presence of root exudates (fumaric, citric and malic acid) in the liquid medium after 96 h in dependence of the pH value for (a) the 200 µM experiment; (b) the 20 µM experiment and (c) the 2 µM experiment.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



