
Submitted:
03 July 2024
Posted:
04 July 2024
You are already at the latest version
Abstract
Two new species of free-living nematode, Metachromadora parobscura sp. nov. and Molgolaimus longicaudatus sp. nov., from mangrove wetlands of Beihai, Guangxi province in China, are described. Metachromadora parobscura sp. nov. is characterized by eight longitudinal rows of somatic setae arranged from the posterior part of the body, loop-shaped amphidial foveae with an open top and double contours, pharynx with bipartite cuticularized internal cavity, spicules with well-developed capitulum, gubernaculum extended ventrally to support spicule, 6-8 precloacal tubular supplements, and short, conical tail with two protuberances. Molgolaimus longicaudatus sp. nov. is characterized by short cephalic setae, relatively small amphidial fovea, ventrally bent spicules with pronounced hamose curved proximal terminus, two undulate precloacal supplements, and relatively long conico-cylindrical tail. Nearly full-length SSU sequences of the two species were provided, and phylogenetic tree based on maximum likelihood analyses supported the taxonomic position of the two new species. Combined use of traditional morphology-based taxonomy and molecular technology would be a good choice for identification of free-living nematodes.
Keywords:
Metachromadora parobscura sp. nov.
; Molgolaimus longicaudatus sp. nov.
; taxonomy
; Beibu Gulf of the South China Sea
; Small subunit (SSU) 18S rDNA gene
1. Introduction
The Mangrove is one of the most productive ecosystems in the world and plays an important role in the maintenance of coastal biodiversity. Mangrove sediments harbor numerous marine nematodes, which play a key role in nutrient cycling and affect carbon fixation in mangrove wetlands [1]. Furthermore, nematodes are sensitive to physicochemical conditions of sediment, thus being potential bioindicators for conservation purposes [2]. Hence, the identification of nematode species would improve our understanding of ecosystem’s functional roles [3]. In the past years, community structure of nematodes was investigated, and also some nematode species were found and described from mangrove wetlands of China [4,5,6].
The family Desmodoridae belongs to the superfamily Desmodoroidea Filipjev, 1922, and it is composed of six subfamilies: Desmodorinae Micoletzky, 1924, Molgolaiminae Jensen, 1978, Prodesmodorinae Lorenzen, 1981, Pseudonchinae Gerlach & Riemann, 1973, Spiriniinae Gerlach & Murphy, 1965 and Stilbonematinae Chitwood, 1936 [7]. The genus Metachromadora Filipjev, 1918 belongs to subfamily Spiriniinae Gerlach & Murphy, 1965. Now, it is comprised of 32 valid species which are divided into five subgenera: Bradulaimus Stekhoven, 1951, Metachromadora Filipjev, 1918, Metachromadoroides Timm, 1961, Metonyx Chitwood, 1936, and Neonyx Cobb, 1933. And Maria et al. provided a revised dichotomous key to the genus [8]. The genus Molgolaimus Ditlevsen, 1921 belongs to subfamily Molgolaiminae Jensen, 1978. It was divided into four species groups by Fonseca et al. based on spicule length, and scatter-plot graphs of morphometric data, and illustrations of the different 33 Molgolaimus species were provided in the study [9]. Furthermore, Molgolaimus haakonmosbiensis sp. n. [10], Molgolaimus kaikouraensis sp. nov. [11] and Molgolaimus euryformis sp. nov. [6] were described. Additionally, two Microlaimus species were transfered to the genus by Shi and Xu [12]. Molgolaimus now contains 38 valid species.
Choice of molecular markers in nematode barcoding studies was controversial. COI, 18S and 28S nuclear ribosomal DNA (rDNA) have been suggested as ideal gene markers to discriminate marine nematodes [13,14]. And also some scientists proposed the combined use of different genes would get better results.18S rRNA sequences contain both conserved and variable regions, suitable for primer design and taxonomic distinction. Therefore, the 18S rRNA sequence was a potential useful marker for the rapid molecular identification of nematode. Desmodoridae is complicated due to lack of synapomorphy and it is also highly polyphyletic based upon small subunit of ribosomal DNA [15]. So in this study, we described two new species, one belonging to the genus Metachromadora and another to Molgolaimus, based on combined morphological and molecular techniques. Nearly full-length SSU sequences of this two species were obtained for phylogenetic analysis. This study will help to improve our assessment of marine nematode diversity and their evolutionary relationships.
2. Materials and Methods
2.1. Sampling and Morphological Analysis
Sediments were obtained from mangrove wetland located in Tieshan Harbor of Beihai, Guangxi province (109°36’ E, 21°36’ N) in October 2023. Samples were collected by pushing syringe (2.6 cm internal diameter) into the sediment to a depth of 5 cm. The sediment was then fixed in 5% formalin. Samples used for molecular analysis was stored in 95% ethanol and frozen at − 20 °C. The nematodes were sorted using differential interference contrast microscope (Leica DM 2500) (CMS Gmbh, Wetzlar, Germany and assembled in Shanghai, China). Line drawings were made with the aid of a camera lucida (Leica, 10×). All measurements were taken using Leica LAS X version 3.3.3, and all curved structures were measured along the arc or median line.
2.2. Molecular Analysis
Specimens were rinsed with phosphate buffer three times to remove ethanol. Subsequently, nematodes were cut into several pieces with a sterile scalpel, transferred into PCR tubes and DNA was extracted from nematodes following the protocol of the DNeasy Blood & Tissue kit (Qiagen, Germany). PCR was performed using an Ex taq polymerase kit (TaKaRa). Nearly full-length of the 18S rDNA gene was amplified using 18S-G18S4 (5’-GCTTGTCTCAAAGATTAAGCC-3’) and 18S-18P (5’-TGATCCWKCYGCAGGTTCAC-3’) [16]. Each PCR mixture (50 μl) contained 5µl DNA template, 200 μmol/L of each dNTP, 1.5 μmol/L MgCl2, 0.3 μmol/L of each primer, 2.5 U of Ex Taq DNA polymerase, and 5 µl of 10× reaction buffer. The PCR program was as follows: denaturation at 94°C for 5 min; 38 cycles of denaturation at 94°C for 30s, annealing at 56°C for 1 min 30s, and extension at 72°C for 30s; and a final extension at 72°C for 7 min. PCR products were gel purified, cloned and sequenced. All sequences were deposited in GenBank. Diplopeltis cirrhatus was selected as the outgroup. A total of 23 18S rDNA sequences, belonging to the family Desmodoridae and Microlaimidae, were downloaded from the GenBank database. The sequences were subsequently edited and aligned using the program Geneious v. 9.0.2 (Biomatters Ltd., Auckland, New Zealand), Maximum likelihood bootstrapping analysis was performed with RAxML HPC v. 8, using the locus-specific model partitions with the default parameters and the GTRGAMMA model as implemented on the CIPRES, NSF XSEDE resource with bootstrap statistics calculated from 1000 bootstrap replicates [17].
2.3. Abbreviations
De Man (1884) ratios a = body length divided by body maximum width, b = body length divided by the pharynx length, c = body length divided by the tail length, c’ = tail length divided by the anal body diameter, c.b.d.=corresponding body diameter, V% = distance from the vulva from the anterior end divided by the total body length [18].
3. Results
3.1. Systematics of Metachromadora Parobscura sp. Nov.
Class Chromadorea Inglis, 1983
Subclass Chromadoria Pearse, 1942
Order Desmodorida De Coninck, 1965
Superfamily Desmodoroidea Filipjev, 1922
Family Desmodoridae Filipjev, 1922
Subfamily Spiriniinae Gerlach & Murpy, 1965
Genus Metachromadora Filipjev, 1918
Diagnosis of the genus Metachromadora Filipjev, 1918 (Modified from Maria et al. 2013)
Cuticle finely or coarsely striated, extreme anterior end of the head unstriated, longitudinal lateral ridges present or absent. Somatic setae present. Cryptospiral amphidial fovea partly or not surrounded by striations. Inner labial sensilla papilliform, outer labial and cephalic sensilla papilliform or setiform, buccal cavity with large dorsal tooth, and subventral teeth present or absent. Pharynx with well developed posterior bulb with a thick cuticular lining and partitioned into two or three sections; male with one testis at left of the intestine; spicules with well developed velum and capitulum; precloacal supplements of various forms or absent; tail conical [8].
3.1.1. Type material
Four males and two females were measured and studied. Holotype: ♂1 on slide C3-1-5. Paratypes: ♂2 on slide C3-1-5, ♂3 and ♂4 on slides C3-1-15, ♀1 on slide C3-1-7, and ♀2 on slide C3-1-8.
3.1.2. Type Locality and Habitat
Muddy surface sediment (0–5 cm) in mangrove wetland, located in Tieshan Harbor in Beihai, Guangxi province (109°36’ E, 21°36’ N).
3.1.3. Etymology
The species epithet comes from the species name Metachromadora bscura, referring to the new species is similar to Metachromadora bscura in most morphological characters.
3.1.4. Measurements
All measurement data are given in Table 1.
3.1.5. Description of Metachromadora Parobscura sp. nov. (Figure 1 and Figure 2 )
Male: Body cylindrical 896–1020 µm long,finely striated, with striation starting immediately after the amphidial foveae. Lateral ridges, 2.0–3.0 µm wide (~1/30 of the body diameter at mid-body), beginning at the anterior end of the pharynx bulb and extending as far as middle of the tail. Blunt conical head. Six internal and six external labial sensillae prominent papilliform. Four cephalic setae, 5.0–6.0 µm long. No cervical setae. Somatic setae, short and stout, arranged in eight longitudinal rows from the posterior part of the body (two rows at each lateral side, two rows subventral and two rows sudorsal). Amphideal fovea loop-shaped, with an open top and have double contours, positioned anterior to the first head cuticular annulus.Width of amphidial foveae is 13.0 - 15.0µm,about 50.0–62.5% of corresponding body diameter. Buccal cavity 33.0 - 38.0μm deep, with three teeth, a large dorsal tooth and two minute subventral teeth, dorsal tooth bigger. Bulb length is 39–43% of the total pharynx length, with bipartite cuticularized internal cavity. Cardia muscular, small. Nerve ring anterior to the pharynx bulb. Secretory–excretory pore not observed. Reproductive system monorchic with outstretched testis. Vas deferens right of intestine. Two equal spicules (63–68 µm), slender, ventrally curved with well-cephalated capitulum. Gubernaculum (30-31 µm) boat-shaped, parallel to spicule, with pointed end extended ventrally to support spicule, without apophyses. Six tubular precloacal supplements, about 11-17µm apart, first supplement 5-6 µm distant from the cloaca. Tail conical (1.7–2.0 times cloacal body diameter) with spinneret (7-8 µm). Terminal part of tail smooth, 8-10 µm long, 10 –14% of total tail length.Two protuberances positioned on ventral side of the tail, the first one 38-44 µm distant from the cloaca. Caudal glands present, but poorly visible.
Female: Similar to male in general morphology. Few and scattered somatic setae, without a pattern of organization. Reproductive system amphidelphic with two ovaries opposed and reflexed, anterior ovary positioned to the left of the intestine, posterior ovary to right.Vagina very sclerotized. Two glands present.Vulva in the shape of transversal slit, located posterior to mid-body (56–59% of total length), occupying about five annules. Sperm cells are always found within the uterus. Tail without protuberance and setae.
3.1.6. Diagnosis
Metachromadora parobscura sp. nov. is characterized by eight longitudinal rows somatic setae arranged from the posterior part of the body, loop-shaped amphidial foveae with an open top and double contours, pharynx with bipartite cuticularized internal cavity, spicules with well-developed capitulum, gubernaculum extended ventrally to support spicule, 6-8 precloacal tubular supplements, and short, conical tail with two protuberances.
3.1.7. Differential Diagnosis
Due to the morphological characters, the new species is placed in Metachromadoroides Timm, 1961. There are nine valid species within the subgenus Metachromadoroides Timm, 1961: M. (M.) complexa, M. (M.) pulvinata, M. (M.) remanei, M. (M.) vulgaris, M. (M.) zaixsi, M. (M.) minor, and M. (M.) orientalis, M. (M.) obscura, and M. (M.) xiamenensis. Description of Subgenus Metachromadoroides Timm, 1961 (Modified from Timm 1961) is as follows: cuticle heavily annulated,without longitudinal striation on head. Lateral ridges present. Stoma cylindrical, with dorsal tooth. Smaller subventral teeth present or absent. Amphidial foveae circular or loopshaped, having double contour. Esophageal bulb with two or three internal division. Precloacal supplemental male organs in the shape of short faintly cuticularized tubules or absent [19].
Metachromadoroides could be subdivided into two groups based on the precloacal supplements. Supplements absent: M. (M.) zaixsi, M. (M.) orientalis and M. (M.) vulgaris. These three species could be easily differentiated by internal division of pharyngeal bulb (bipartite in M. (M.) orientalis vs. tripartite in M. (M.) zaixsi and M. (M.) vulgaris) and length of anterior non-striated part (one amphidial fovea lengths in M. (M.) zaixsi vs. two in M. (M.) vulgaris). Supplements present: M. (M.) complexa, M. (M.) xiamenensis and M. (M.) minor have unique characteristics separately (setae absent,three small protuberances on ventral side of the tail and short slim body); M. (M.) pulvinata, M. (M.) remanei and M. (M.) obscura differed from each other by body length, and numbers of precloacal supplements (1720mm and 23 vs. 1100-1300mm and 5 vs. 786mm and 13-14).
Metachromadora parobscura sp. nov. is closely resembles M. (M.) remanei in De Man ratios. New species could be distinguished from it by body length (896-1020mm vs.1100-1300mm in M. (M.) remanei ) , spicules length (63-68 µm vs. 49-55 µm in M. (M.) remanei ) and numbers of precloacal supplements ( 6-8 vs. 5 in M. (M.) remanei ). Otherwise, the new species is morphologically similar to M. (M.) obscura. But differs from it by relatively shorter and thinner tail (c = 11.5–14.8, с’ = 1.7–2.0 vs. c = 10.7-12.1, с’ = 1.4-1.6 in M. (M.) obscura), amphid diameter as a percentage of the corresponding body diameter (50-62.5% vs. 65-75% in M. (M.) obscura), shorter spicules (63-68 µm vs. 75-78 µm in M. (M.) obscura), shorter gubernaculum (30-31 µm vs. 38-40 µm in M. (M.) obscura), smaller number of precloacal supplements ( 6-8 vs. 13-14 tubular supplements in M. (M.) obscura). Morphological differentiating characters of the species of Metachromadoroides are proposed (Table 2).
3.2. Systematics of Molgolaimus Longicaudatus sp. nov.
Class Chromadorea Inglis, 1983
Subclass Chromadoria Pearse, 1942
Order Desmodorida De Coninck, 1965
Superfamily Desmodoroidea Filipjev, 1922
Family Desmodoridae Filipjev, 1922
Subfamily Molgolaiminae Jensen, 1978
Genus Molgolaimus Ditlevsen, 1921
Description of the Genus Molgolaimus Ditlevsen, 1921 (Modified from Fonseca et al., 2006)
Cuticle finely striated or apparently smooth. Amphidial fovea round and posterior to the cephalic constriction. Cephalic setae close to the cephalic constriction. Buccal cavity small, narrow and with small teeth. Pharynx with a pronounced spherical bulb. Male reproductive system monorchic. Spicules of variable length and shape from short and bent to long and sinusoidal or straight. Gubernaculum with or without apophysis. Precloacal supplements often present. Female reproductive system didelphic-amphidelphic, with ovaries reflexed. Tail of varying shape and length [9].
3.2.1. Type Material
Four males and four females were measured and studied. Holotype: ♂1 on slide C3-1-49. Paratypes: ♂2 on slide C3-1-48, ♂3 on slide C3-1-50, ♂4 on slide C3-1-51, ♀1 on slide C3-1-47, ♀2 on slide C3-1-49, and ♀3 on slide C3-1-50.
3.2.2. Type Locality and Habitat
Muddy surface sediment (0–5 cm) in mangrove wetland, located in Tieshan Harbor in Beihai, Guangxi province (109°36’ E, 21°36’ N).
3.2.3. Etymology
The species name is composed of the Latin adjectives longus and caudatus, referring to the new species with a relatively long tail.
3.2.4. Measurements
All measurement data are given in Table 3.
3.2.5. Description of Molgolaimus Longicaudatus sp. nov. (Figure 3 and Figure 4 )
Male: Body slender and cylindrical, tapering gradually towards both ends. Cuticle annules not obvious. Head separated from body by a faint constriction. Inner labial and outer labial sensilla indistinct. Four cephalic setae, 2 μm long, located posterior to cephalic constriction. Amphideal fovea round, posterior to the cephalic constriction, 3-4 µm in diameter, corresponding to 33.3-44.4 % of c.b.d., about 6-7 µm from the anterior end. Buccal cavity small and narrow, without teeth.Buccal cavity small, weakly sclerotized, narrow and with small teeth. Pharynx corpus narrow cylindrical, ending in apronounced spherical muscular bulb (17-19µm diameter), and the bulb heavily cuticularised. Nerve ring located slightly anterior to middle of pharynx length. Excretory pore not observed. Cardia cylindrical, 6-9 μm long. Reproductive system monorchic, with a single anterior outstretched testis left of the intestine. Vas deferens long and thin. Spicules 32-35 µm long (1.8-1.9 abd), ventrally bent and pronounced hamose curved proximal terminus. Gubernaculum simple and parallel to the spicules, without apophyses, its length corresponding to one third of the spicule length. Two precloacal supplements, 25-41 µm and 43-68 µm distant from the cloaca. Tail conico-cylindrical, conical in the proximal half and distally characterized by a cylindrical prolongation. Caudal glands not observed.
Female: Similar to male, but with larger maximum body diameter and shorter tail. Reproductive system didelphic with reflexed ovaries, the anterior branch right of the intestine and posterior branch to the left. Vulva located anterior to mid-body (42.7-44.1% of total length).
3.2.6. Diagnosis
Molgolaimus longicaudatus sp. nov. is characterized by short cephalic setae (2 μm), relative small amphidial fovea (3-4 μm in diameter), ventrally bent spicules with pronounced hamose curved proximal terminus, two undulate precloacal supplements, long conico-cylindrical tail (115-138 μm). Female reproductive system didelphic with both ovaries reflexed.
3.2.7. Differential Diagnosis
Fonseca et al. provides an illustrated polytomous identification key to 33 species of the genus, which were classified into four groups according to absolute spicule length [9]. This new species is morphologically closely related to the species in group 1b1 of the identification key, characterized by short spicules (< 35 μm), ratio spicules divided by the anal body diameter ranging from 1 to 3. There are seven species in the group: M. drakus, M. exceptionregulum, M. gazii, M. pecticauda, M. mareprofundus,M. sapiens,M. spirifer. Molgolaimus could be subdivided into three groups based on the number of precloacal supplements. Supplements absent: Molgolaimus pecticauda and Molgolaimus spirifer could be easily differentiated clearly from other species of the group by its amphid ( amphid with a tubiform structure and multispiral amphid fovea, vs unispiral or circular). M. sapiens differs in spicule with S-shaped blade. One supplements: M. drakusthe is characterized by slim body, the shortest spicules and amphidial fovea at 3 hd from the anterior end. M. mareprofundus resembles M. exceptionregulum, but differs from it by the smaller body and slender spicules. Two supplements: M. gazii is more shorter and characterized by sexual dimorphism amphids. Molgolaimus longicaudatus sp. nov. can be distinguished by the longest tail (130-138 μm) and the spicules shape (pronounced hamose curved proximal terminus). Regardless of precloacal supplements, Molgolaimus longicaudatus sp. nov. is morphologically close to M. exceptionregulum and M. mareprofundus, but differs from them by the larger body length( 735-793 µm vs. 500-605 μm and 610–630 μm), longer tail (c = 5.4-6.9 vs. c =7.0-8.9, c =7.2-7.6) , and the presence of two precloacal supplement (one in M. exceptionregulum and M. mareprofundus). Morphological differentiating characters of the Molgolaimus species of the group1b1( based on classification of Fonseca et al. [9] )was provided (Table 4).
3.3. Molecular Phylogenetic Analysis
In this study, nearly full-length SSU sequences(1542-1592 bp) of the two new specieswere were provided. All sequences were deposited in GenBank, and the accession numbers were shown in Figure 5. Sequences alignment indicated that the amplified 18S rRNA fragments contained some variable regions, and it is suitable for DNA barcoding studies of Desmodoridae. Then a taxonomic phylogenetic tree was constructed based on maximum likelihood analyses. The new species Molgolaimus longicaudatus sp. nov. formed a separate clade and represented by a bootstrap support of 100%. It showed a close relationship with Molgolaimus kaikouraensis(MK446235.1) and exhibied good differentiation from species of other subfamilies. The other species of Desmodoridae comprised another well-supported monophyletic clade (bootstrap support of 100%). Metachromadora parobscura sp. nov. in this clade formed a high-support clade clustering with other Metachromadora species (bootstrap = 98%).Thus the phylogenetic positions of these two species were confirmed.
4. Discussion
Metachromadora is a heterogeneous genus and was not well defined [30]. It has been reviewed by Gerlach (1951), Wieser & Hopper (1967), Gerlach & Riemann (1972), Lorenzen (1981) and Maria (2013) [8,31]. It has been first divided into five subgenera: Bradylaimus, Chromadoropsis, Metonyx, Metachromadora, and Neonyx by Gerlach (1951)[21]. Subsequently, Timm (1961) erected the sixth subgenus, Metachromadoroides, with species M. vulgaris and M. complexa [19]. Later, Chromadoropsis was reinstated to the genus level by Furstenberg & Vincx, in 1988 [32]. Until now, Metachromadora consists of five subgenera and is still a large genus, containing 32 valid species. Although Metachromadora serrata Gerlach, 1963 and Metachromadora spiralis Gerlach, 1955 are considered valid species, they were regarded belonging to a doubtful genus or as incertae sedis [8,20]. The sequences of M. (M.) parobscura sp. nov. were blasted with sequences in the NCBI genebank. M. (M.) remanei (Accession number AM234620.1), which was identified to species level, was with the highest similarity to the new species, percentage of identity was 94.75% and percentage of base differences was 5.2%(82/1574). Because M. (M.) parobscura sp. nov. most closely resembles M. (M.) remanei in De Man ratios, different geographic speciation of M. (M.) remanei, from estuarine waters around southwest Britain and Mira estuary of Portugal separately (Accession number AM234620.1 and KX944141.1), were also aligned with MegAlign (DNAStar), and percentage of base differences between these two geographic speciation was only 2.4% (33/1389). So Possibility that the new species was the geographic existence of M. (M.) remanei was excluded.The phylogenetic tree based on 18S rRNA sequences also showed the taxonomic relationships between M. (M.) parobscura sp. nov. and M. (M.) remanei.
The taxonomic status of Molgolaimus was complicated and emended several times. Molgolaimus was firstly placed in family Microlaimidae by Gerlach and Riemann in 1973[33]. The family Microlaimidae is characterized by two outstretched ovaries and two opposed testes or only anterior testis. Subsequently, Jensen (1978) erected the family Molgolaimidae based on female and male reproductive systems (a single anterior testis in males and two reflexed ovaries in females) [34]. Then, Lorenzen (1994) downgraded the family Molgolaimidae as a subfamily Molgolaiminae in Desmodoridae Filipjev, 1922 [35]. Shi et al. (2016) transferred Microlaimus pecticauda and Microlaimus spirifer from the family Microlaimidae to Molgolaimus of the family Desmodoridae [12]. But recently, Leduc et al. (2018) again placed Molgolaimus demani Jensen, 1978. in the Microlaimida [36]. Sometimes, it is hard to determine a new species belong to which of these two families. In order to confirm the taxonomic position of M.longicaudatus sp. nov., the phylogenetic tree included members belonging to the the family Desmodoridae and Microlaimidae(Figure 5). As shown in the phylogenetic tree, the family Microlaimidae and Desmodoridae exhibits two monophyletic lineage, and the larger Desmodoridae clade encompassing three subfamilies (Desmodorinae, Molgolaiminae and Spiriniinae). Results showed that the new species Molgolaimus longicaudatus sp. nov. clustered within the Desmodoridae clade, supporting the opinion of Lorenzen.
5. Conclusions
Two new species of free-living nematode, Metachromadora parobscura sp. nov. and Molgolaimus longicaudatus sp. nov. , from mangrove wetlands in Beihai were discovered. And differentiating morphometric data were given. Nearly full-length SSU sequences of the two species were provided. ML topology performed with SSU sequences supported the taxonomic position of the two new species, and they were both assigned a firm place within the phylogeny. Because vailable sequences in databases were seldom, most of sequences were too short or not assigned to species levels, it limited the resolving abilities of molecular tools for nematodes identification. With the enrichment of nematodes DNA database, it would be particularly valuable in identifying those closely related species, and contributes to a more robust nematodes taxonomy.
Author Contributions
preparation of specimens, data measurement and writing---original draft preparation, J.S.; writing---review and editing,Y.H.; funding acquisition, J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by Shandong Provincial Natural Science Foundation, China
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank Chunyan Ji for sample collection and are also grateful to Xin Zhao for providing valuable suggestions for the phylogenetic analysis.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Naidoo, G.; Naidoo, K.; Swart, A. Oil Pollution Effects on Nematodes in Mangrove Sediment: A Microcosm Study. 2021. Preprint (Version 1) at Research Square.doi:10.21203/rs.3.rs-749759/v1. [CrossRef]
- Spedicato, A.; Zeppilli, D.; Thouzeau, G.; Michaud, E. Nematode diversity patterns in mangroves: a review of environmental drivers at different spatial scales. Biodivers Conserv. 2023, 32, 1451–1471.Author 1, A.; Author 2, B. Book Title, 3rd ed.; Publisher: Publisher Location, Country, 2008; pp. 154–196.
- Sahraean, N.; Van Campenhout, J.; Rigaux, A.; Mosallanejad, H.; Leliaert, F.; Moens, T. Lack of population genetic structure in the marine nematodes Ptycholaimellus pandispiculatus and Terschellingia longicaudata in beaches of the Persian Gulf, Iran. Mar. Ecol. 2017, 38, e12426.https://doi.org/10.1111/maec.12426. [CrossRef]
- Xiao, Y.P.; Guo, Y.Q. Three new free-living marine nematode species of Subsphaerolaimus Lorenzen, 1978, Halichoanolaimus de Man, 1886 and Belbolla Andrássy, 1973 from mangrove wetlands in Taiwan. Zootaxa. 2023, 5361(3), 301–322.
- Wang, Y.; Pang, X. P.; GUO Guo, Y.Q. Study on marine nematodes community in Jinhai bay mangrove wetland, Beihai, Guangxi. Oceanologia Et Limnologia Sinica. 2020, 51(3), 583–590.
- Zhou, X.P.; Zeng, J.L.; Cai, L.Z.; Fu, S.J.; Tan, W.J. Two New Species of Free-Living Marine Nematodes of the Desmodoridae from Mangrove Wetlands of Xiamen Bay, China. J. Ocean Univ. China. 2020, 19, 143–150. https://doi.org/10.1007/s11802-020-4080-6. [CrossRef]
- Armenteros, M.; Ruiz-Abierno, A.; Decraemer, W. Revision of Desmodorinae and Spiriniinae (Nematoda: Desmodoridae) with redescription of eight known species. European Journal of Taxonomy. 2014, 96, 1–32. http://doi.org/10.5852/ejt.2014.96. [CrossRef]
- Maria, T.F.; Smol, N.; Esteves, AM. Two new species of Metachromadora (Nematoda: Desmodoridae) from Guanabara Bay, Rio de Janeiro, Brazil, and a revised dichotomous key to the genus. Journal of the Marine Biological Association of the United Kingdom. 2014, 94(1),105–114. doi:10.1017/S0025315413001161. [CrossRef]
- Fonseca, G.; Vanreusel, A.; Decraemer, W. Taxonomy and biogeography of Molgolaimus Ditlevsen, 1921 (Nematoda: Chromadoria) with reference to the origins of deep sea nematodes. Antarctic Science. 2006, 18(1), 23–50. doi:10.1017/S0954102006000034. [CrossRef]
- Portnova, D. Free-living nematodes from the deep-sea Håkon Mosby Mud Volcano, including the description of two new and three known species. Zootaxa. 2019, 2096, 197–213.
- Leduc, D.; Fu, S.; Zhao, Z.Q. New nematode species from the continental slope of New Zealand (Chromadorea, Microlaimida, and Chromadorida), and unexpected placement of the genus Molgolaimus Ditlevsen, 1921. Mar. Biodiv. 2019, 49, 2267–2280. https://doi.org/10.1007/s12526-019-00961-z. [CrossRef]
- Shi, B.; Xu, K. Spirobolbolaimus undulatus sp. nov. in intertidal sediment from the East China Sea, with transfer of two Microlaimus species to Molgolaimus (Nematoda, Desmodorida). Journal of the Marine Biological Association of the United Kingdom. 2017, 97 (6), 1335–1342.
- Macheriotou, L.G.K.B.; Nguyen, D.T.; Phuong Nguyen, T.X.; Noppe, F.; Armenteros, M.; Boufahja, F.; Rigaux, A.; Vanreusel, A.; Derycke, S. Metabarcoding free-living marine nematodes using curated 18S and CO1 reference sequence databases for species-level taxonomic assignments. Ecol. Evol. 2019, 9, 1211–1226.
- Avó, A.P.; Daniell, T.J.; Roy, N.; Solange, O.; Jordana, B.; Helena, A. DNA barcoding and morphological identification of benthic nematodes assemblages of estuarine intertidal sediments: Advances in molecular tools for biodiversity assessment. Front. Mar. Sci. 2017, 4, 66.
- van Megen, H.; van den Elsen, S.; Holterman, M.; Karssen, G.; Mooyman, P.; Bongers, T.; Holovachov, O.; Bakker, J.; Helder, J. A phylogenetic tree of nematodes based on about 1200 full-length small subunit ribosomal DNA sequences. Nematology. 2009, 11, 927–950.
- De Ley, I.T.; De Ley, P.; Vierstraete, A.; Karssen, G.; Moens, M.; Vanfleteren, J. Phylogenetic analysis of Meloidogyne small subunit rDNA. Journal of Nematology. 2002, 34, 319–327.
- Stamatakis, A. RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014, 30, 1312–1313.
- Platt, H.M.; Warwick, R.M. Free-living Marine Nematodes, Part I, British Enoplids. In Synopses of the British Fauna (New Series). The Linnean Society of London: London, UK. 1998, p. 248.
- Timm, R.W. The marine nematodes of the Bay of Bengal. Proceedings of the Pakistan Academy of Sciences. 1961, 1, 1–88.
- Wieser, W.; Hopper, B. Marine nematodes of the east coast of North America. I Florida. Bulletin Museum of Comparative Zoology. 1967, 135 (5), 239–344.
- Gerlach, S. A. Nematoden aus der familie der chromadoridae von den deutschen Küsten. Kieler Meeresforschungen. 1951, 8, 106–132.
- Pastor, C. T.; Ward, D. New species of Hopperia (Nematoda, Comesomatidae) and Metachromadora (Nematoda, Desmodoridae) from Patagonia, Chubut. Zootaxa. 2004, 542, 1–15.
- Gagarin, V. G.; Nguyen V. T. Three new species of free-living marine nematodes of the order Desmodorida (Nematoda) from Vietnam. Zoologicheskii Zhurnal. 2010, 89(4), 398–406.
- Gagarin, V. G.; Tu, N. D. Two new species of free living nematodes (Nematoda and Chromadorea) from mangrove thicket in Vietnam. Inland Water Biology. 2014, 7 (4), 338–347.
- Nguyen, D. Tu.; Gagarin V.G.; Phan K. Long.; Nguyen T. X. P.; Nguyen V. T. Two new species of free-living marine nematodes (Nematoda) from an area near the mouth of the Yen River of Vietnam. Biologiya Morya. 2016, 42(6), 439–448.
- Zhou, X.; Zeng, J.; Cai, L.; Fu, S.; Tan, W. Two New Species of Free-Living Marine Nematodes of the Desmodoridae from Mangrove Wetlands of Xiamen Bay, China. Journal of Ocean University of China. 2020, 19(1), 143–150.
- Murphy, D. G. An initial report on a collection of Chilean marine nematodes. Mitt. hamb. zool. Mus. Inst. 1966, 63, 29–50.
- Warwick, R. M. Fourteen new species of free-living marine nematodes from the Exe estuary. Bull. Br. Mus. nat. Hist. (Zool.). 1970, 19 (4), 137–177.
- Muthumbi, A. W.; Vincx, M. Nematodes from the Indian Ocean: description of six new species of the genus Molgolaimus Ditlevsen, 1921 (Nematoda: Desmodoridae). Bulletin de l’institut Royal dae Sciences Naturelles de Belgique. Biologie. 1996, 66, 17–28.
- Vincx, M. Free-living marine nematodes from the Southern Bight of the North Sea. Ph. D. thesis, State University of Ghent. 678 pp.
- De Ward, C. T. P. New species of Hopperia (Nematoda, Comesomatidae) and Metachromadora (Nematoda, Desmodoridae) from Patagonia, Chubut, Argentina. Zootaxa. 2004, 542, 1–15.
- Furstenberg, J.P.; Vincx, M. Three new species of Chromadoropsis species (Nematoda, Desmodoridae) from Southern Africa and the North Sea. South African Journal of Zoology. 1988, 23, 215–223. [CrossRef]
- Gerlach, A.; Riemann, F. The Bremerhaven checklist of aquatic nematodes. A catalogue of Nematoda Adenophorea excluding the Dorylaimida. Part 1. Veröff Inst Meer Bremerhaven. 1973, 4, 1–736.
- Jensen, P. Revision of Microlaimidae, erection of Molgolaimidae fam. N., and remarks on the systematic position of Paramicrolaimus (Nematoda, Desmodorida). Zool Scr . 1978, 7, 159–173. [CrossRef]
- Lorenzen, S. The phylogenetic systematics of freeliving nematodes. London: The Ray Society, 1994, 383 pp.
- Leduc, D.; Verdon, V.; Zhao, Z.Q. Phylogenetic position of the Paramicrolaimidae, description of a new Paramicrolaimus species and erection of a new order to accommodate the Microlaimoidea (Nematoda: Chromadorea). Zool J Linnaean Soc. 2018, 183, 52–69. [CrossRef]
Figure 1.
Drawing of Metachromadora parobscura sp. nov. (A) Entire view of male Anterior end of male, showing outer labial setae, cephalic setae, amphid (arrow 1) and epidermal gland cells (arrow 2); (B) entire view of female; (C) anterior end of holotype; (D) anterior end of holotype, showing amphid and tooth; (E) posterior end of male, showing spicule, gubernaculum, precloacal supplements and tail papillae; (F) magnifying spicule and gubernaculum. Scale bar: (A,B) = 50 μm; (C,D,E,F) = 20 μm.
Figure 1.
Drawing of Metachromadora parobscura sp. nov. (A) Entire view of male Anterior end of male, showing outer labial setae, cephalic setae, amphid (arrow 1) and epidermal gland cells (arrow 2); (B) entire view of female; (C) anterior end of holotype; (D) anterior end of holotype, showing amphid and tooth; (E) posterior end of male, showing spicule, gubernaculum, precloacal supplements and tail papillae; (F) magnifying spicule and gubernaculum. Scale bar: (A,B) = 50 μm; (C,D,E,F) = 20 μm.

Figure 2.
Micrograph of Metachromadora parobscura sp. nov. (A) Pharyngeal region of holotype, showing pharyngeal bulbs; (B) lateral ridge (C) lateral view of holotype anterior end, showing anterior sensilla and dorsal tooth; (D) amphid of holotype; (E) posterior end of male, showing spicule and tail papillae; (F) lateral view of male cloacal region, showing spicule, gubennaculum and precloacal supplements. Scale bar: (A,E) = 20 μm; (C,D,F) = 10 μm .
Figure 2.
Micrograph of Metachromadora parobscura sp. nov. (A) Pharyngeal region of holotype, showing pharyngeal bulbs; (B) lateral ridge (C) lateral view of holotype anterior end, showing anterior sensilla and dorsal tooth; (D) amphid of holotype; (E) posterior end of male, showing spicule and tail papillae; (F) lateral view of male cloacal region, showing spicule, gubennaculum and precloacal supplements. Scale bar: (A,E) = 20 μm; (C,D,F) = 10 μm .
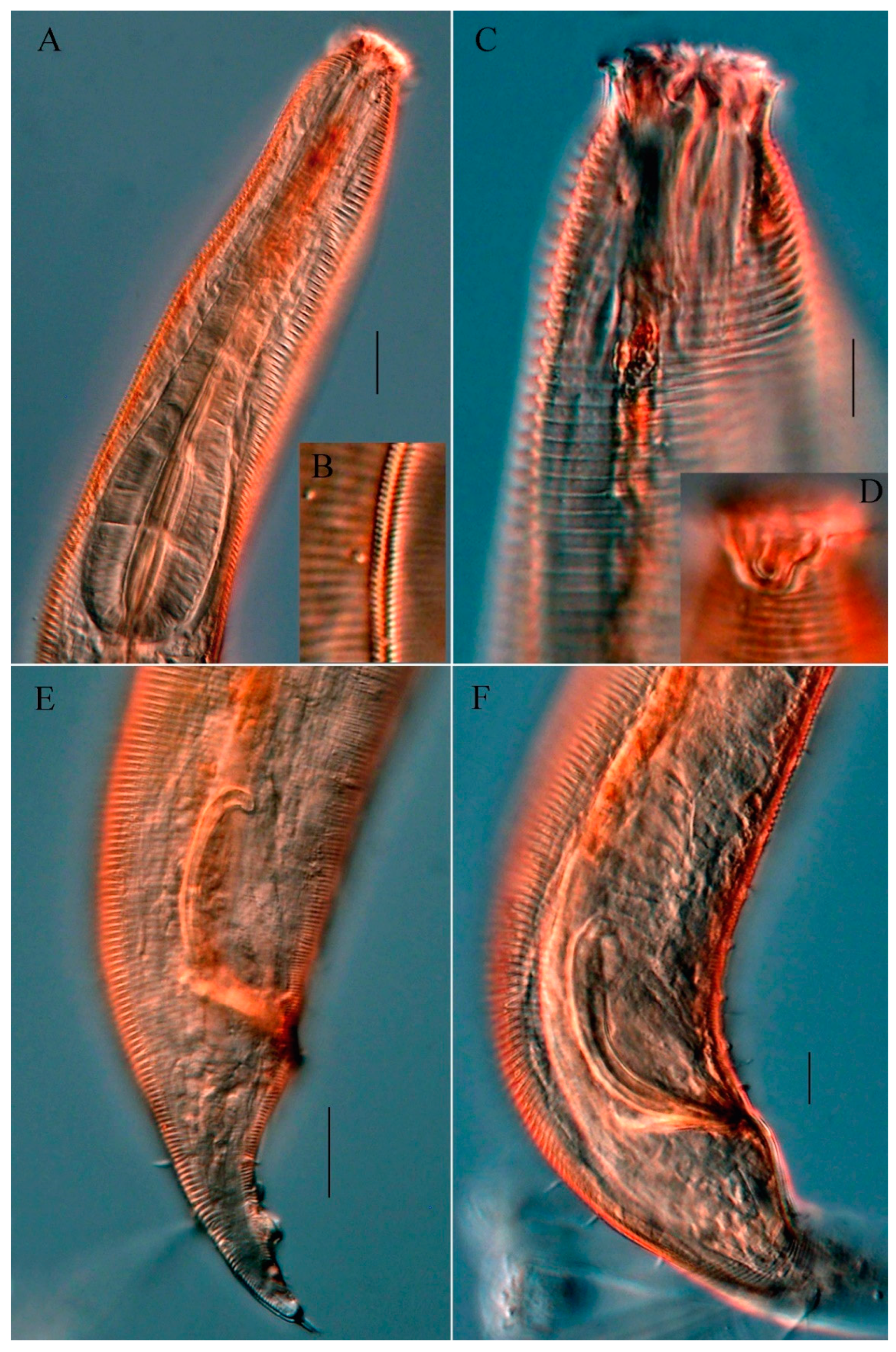
Figure 3.
Micrograph of Molgolaimus longicaudatus sp. nov. (A) Pharyngeal region of holotype, showing amphid, nerve ring and pharyngeal bulb; (B) lateral view of holotype posterior end, showing spicule, gubennaculum and precloacal supplements; (C) lateral view of holotype anterior end, showing cephalic setae, buccal cavity with a dorsal tooth, and amphidial fovea; (D) entire view of female; (E) entire view of male; (F) magnifying spicule and gubernaculum. Scale bar: (A,B,C,F) = 10 μm; (D,E) = 30 μm.
Figure 3.
Micrograph of Molgolaimus longicaudatus sp. nov. (A) Pharyngeal region of holotype, showing amphid, nerve ring and pharyngeal bulb; (B) lateral view of holotype posterior end, showing spicule, gubennaculum and precloacal supplements; (C) lateral view of holotype anterior end, showing cephalic setae, buccal cavity with a dorsal tooth, and amphidial fovea; (D) entire view of female; (E) entire view of male; (F) magnifying spicule and gubernaculum. Scale bar: (A,B,C,F) = 10 μm; (D,E) = 30 μm.
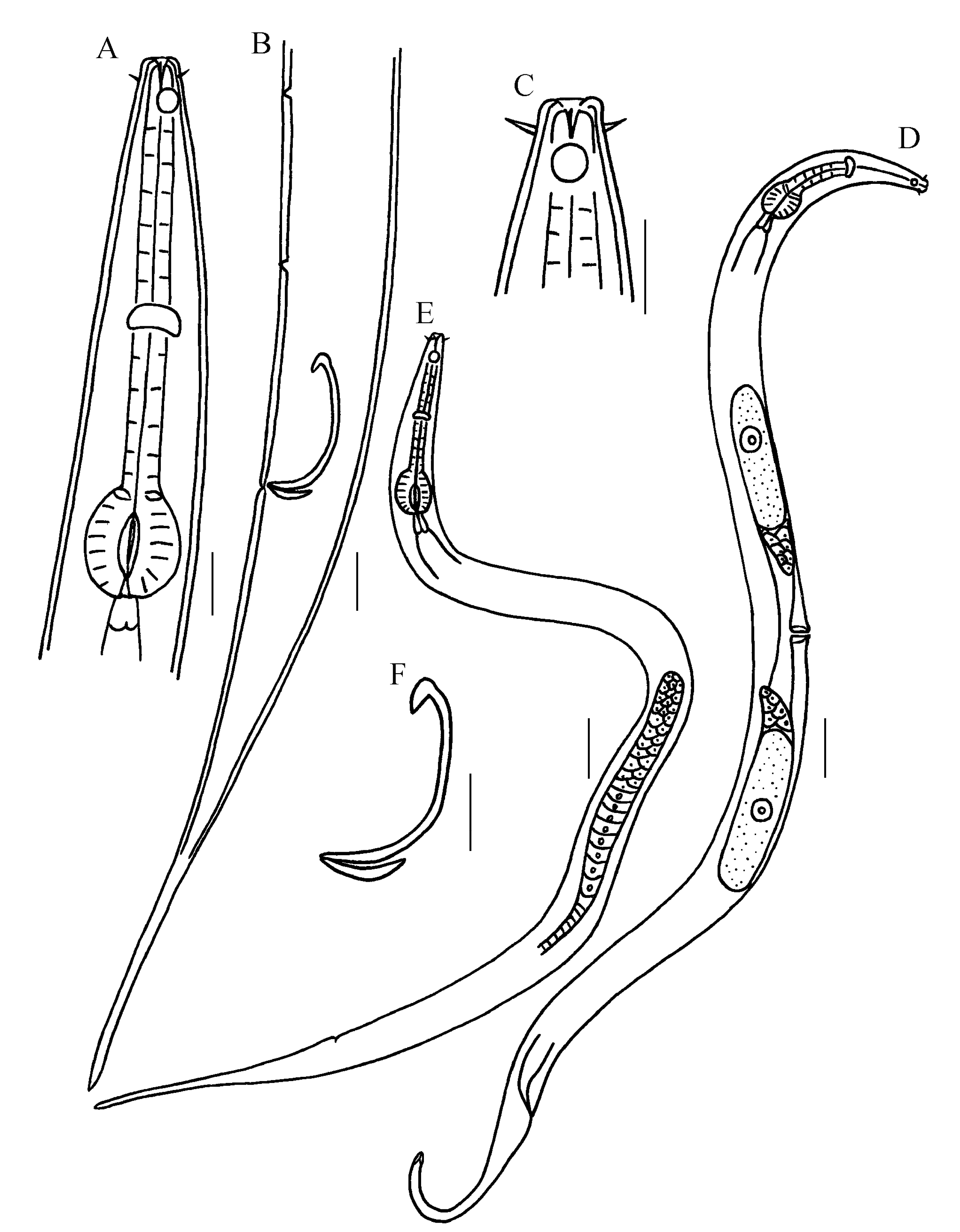
Figure 4.
Micrograph of Molgolaimus longicaudatus sp. nov. (A) Pharyngeal region of holotype, showing amphid (arrow) and pharyngeal bulb; (B) lateral view of holotype anterior end, showing cephalic setae (arrow) and buccal cavity; (C) lateral view of holotype cloacal region, showing spicule, gubennaculum and precloacal supplements (arrows); (D) lateral view of holotype cloacal region, showing spicule. Scale bar: (A,B,C,D) = 10 μm.
Figure 4.
Micrograph of Molgolaimus longicaudatus sp. nov. (A) Pharyngeal region of holotype, showing amphid (arrow) and pharyngeal bulb; (B) lateral view of holotype anterior end, showing cephalic setae (arrow) and buccal cavity; (C) lateral view of holotype cloacal region, showing spicule, gubennaculum and precloacal supplements (arrows); (D) lateral view of holotype cloacal region, showing spicule. Scale bar: (A,B,C,D) = 10 μm.

Figure 5.
Phylogenetic tree generated from maximum likelihood (ML) analysis based on 18S rRNA sequences. ML bootstrap values are displayed above each branch. New species are shown in bold.
Figure 5.
Phylogenetic tree generated from maximum likelihood (ML) analysis based on 18S rRNA sequences. ML bootstrap values are displayed above each branch. New species are shown in bold.
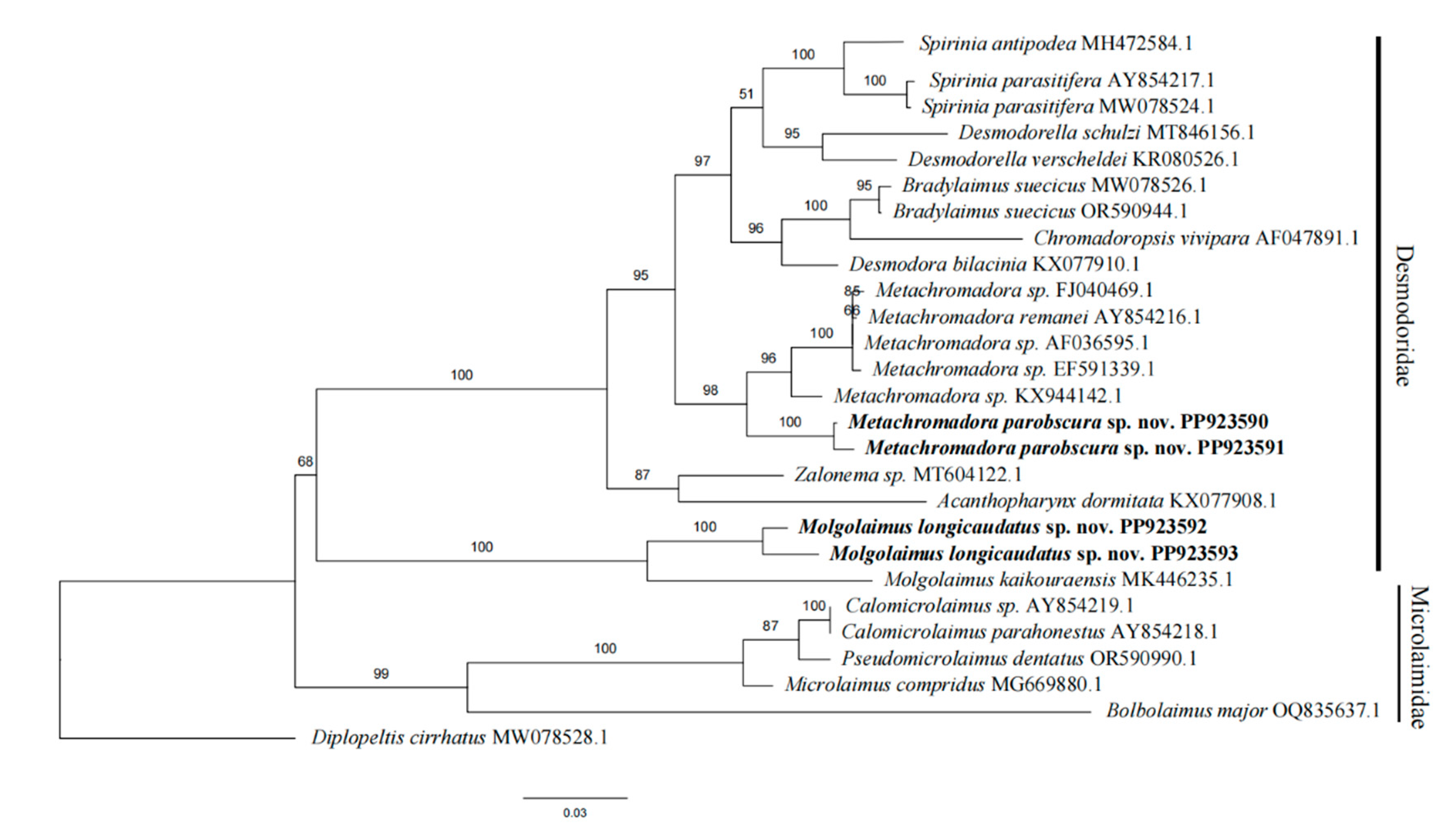

Table 1.
Measurements of individuals of Metachromadora parobscura sp. nov. (in µm).
| Morphological Characters | Holotype | Paratype | ||||
|---|---|---|---|---|---|---|
| ♂1 | ♂2 | ♂3 | ♂4 | ♀1 | ♀2 | |
| a | 14.0 | 14.0 | 17.3 | 15.3 | 11.7 | 12.2 |
| b | 4.1 | 4.2 | 4.4 | 4.4 | 4.3 | 4.2 |
| c | 11.5 | 13.2 | 14.8 | 12.5 | 15.6 | 17.7 |
| c’ | 2.0 | 1.9 | 2.0 | 1.7 | 2.0 | 2.2 |
| Total body length | 896 | 977 | 1020 | 915 | 935 | 976 |
| Maximum body diameter | 64 | 70 | 59 | 60 | 80 | 80 |
| Head diameter | 22 | 24 | 24 | 23 | 26 | 28 |
| Amphideal fovea width | 15 | 13 | 13 | 15 | 17 | 16 |
| Amphideal fovea c.b.d. | 25 | 24 | 26 | 24 | 27 | 24 |
| Amphideal fovea as % of c.b.d. | 60 | 54 | 50 | 63 | 63 | 67 |
| Pharynx length | 216 | 233 | 234 | 210 | 220 | 232 |
| Pharynx c.b.d. | 59 | 67 | 57 | 60 | 71 | 75 |
| Length of double bulb | 77 | 84 | 91 | 90 | 95 | 92 |
| Bulb as % of pharynx length | 35.6 | 36.1 | 38.9 | 42.9 | 43.0 | 40.0 |
| Spicule length as arch | 68 | 63 | 68 | 64 | ||
| Gubernacular length | 31 | 30 | 31 | 31 | ||
| Cloacal /Anal body diameter | 39 | 39 | 39 | 43 | 30 | 25 |
| Distance from first precloacal supplement to anus | 5 | 5 | 5 | 6 | ||
| Distance from last precloacal supplement to anus | 85 | 101 | 114 | 106 | ||
| Precloacal supplement number | 6 | 8 | 6 | 8 | ||
| Distance from the first protuberance to cloaca | 38 | 43 | 44 | 38 | ||
| Distance from the second protuberance to cloaca | 50 | 52 | 52 | 48 | ||
| Tail length | 78 | 74 | 69 | 73 | 60 | 55 |
| Vulva from anterior end | 520 | 575 | ||||
| Vulva c.b.d. | 77 | 80 | ||||
| V% | 55.6 | 58.9 | ||||
Table 2.
Morphological differentiating data of male Metachromadora (Metachromadoroides) species.
| species | Body Length (µm) |
Spicule Length (µm) |
numbers of supplements | internal division of pharyngeal bulb | protuberances on the tail | a | b | c | c’ | References |
|---|---|---|---|---|---|---|---|---|---|---|
| M.(M.) complexa | 758-940 | 74 | 17 | tripartite | 2 | 8.4-11.3 | 3.4-4.4 | 7.3-12.1 | 1.3 | [19] |
| M.(M.) pulvinata | 1720 | 55 | 23 | tripartite | 2 | 18.1 | 5.4 | 15.6 | 2.0 | [20] |
| (M.) remanei | 1100-1300 | 49-55 | 5 | bipartite | * | 14 | 4.9 | 14 | 1.9-2.2 | [21] |
| M.(M.) vulgaris | 1100-1200 | 72 | 0 | tripartite | 0 | 14.1-20.0 | 4.4-4.9 | 12.1-14.5 | 1.8-2.8 | [19] |
| M.(M.) zaixsi | 890-1160 | 40-52 | 0 | tripartite | 0 | 10.5-19.3 | 4.7-7.2 | 6.8-10.1 | 1.7-2.8 | [22] |
| M.(M.) minor | 594-641 | 35-36 | 12-16 | tripartite | * | 12-15 | 4.2-4.7 | 13.5-14.9 | 1.4-1.5 | [23] |
| M.(M.) orientalis | 859-1133 | 59-63 | 0 | bipartite | ** | 13-15 | 4.8-5.3 | 15.3-19.2 | 1.0-1.3 | [24] |
| M.(M.) obscura | 786-1008 | 75-78 | 13-14 | bipartite | 2 | 11-15 | 4.2-4.5 | 10.7-12.1 | 1.4-1.6 | [25] |
| M.(M.) xiamenensis | 741-888 | 46-56 | 18 | bipartite | 3 | 11.0-14.3 | 4.4-5.2 | 10.1-12.2 | 1.5-1.7 | [26] |
| M. (M.) parobscura sp. nov. | 896-1020 | 63-68 | 6-8 | bipartite |
2 | 14.0-17.3 | 4.1-4.4 | 11.5-14.8 | 1.7-2.0 | this study |
* not described, ** described as: Small protuberance with three or four setae.
Table 3.
Measurements of individuals of Molgolaimus longicaudatus sp. nov. (in µm).
| Morphological Characters | Holotype | Paratype | |||||
|---|---|---|---|---|---|---|---|
| ♂1 | ♂2 | ♂3 | ♂4 | ♀1 | ♀2 | ♀3 | |
| a | 31.0 | 30.9 | 28.3 | 30.1 | 28.2 | 24.8 | 28.3 |
| b | 8.0 | 7.4 | 7.7 | 8.2 | 7.9 | 7.9 | 8.2 |
| c | 5.7 | 5.4 | 5.7 | 5.6 | 6.7 | 6.9 | 6.1 |
| c’ | 7.6 | 7.7 | 7.2 | 7.4 | 6.9 | 6.4 | 7.2 |
| Total body length | 775 | 742 | 735 | 752 | 789 | 793 | 791 |
| Maximum body diameter | 25 | 24 | 26 | 25 | 28 | 32 | 28 |
| Head diameter | 6 | 6 | 6 | 6 | 6 | 7 | 5 |
| Cephalic setae length | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Amphideal fovea width | 3 | 3 | 4 | 4 | 4 | 3 | 3 |
| Amphideal fovea c.b.d. | 9 | 8 | 9 | 9 | 9 | 8 | 8 |
| Amphidial fovea from anterior end | 6 | 6 | 6 | 7 | 8 | 5 | 6 |
| Amphideal fovea as % of c.b.d. | 33.3 | 37.5 | 44.4 | 44.4 | 44.4 | 37.5 | 37.5 |
| Nerve ring width | 14 | 15 | 15 | 14 | 15 | 16 | 15 |
| Nerve ring c.b.d. | 20 | 20 | 21 | 20 | 20 | 21 | 20 |
| Pharynx length | 97 | 100 | 96 | 92 | 100 | 101 | 97 |
| Pharynx c.b.d. | 22 | 23 | 22 | 23 | 23 | 25 | 22 |
| Diameter of esophageal bulb | 17 | 19 | 18 | 18 | 17 | 19 | 17 |
| Spicule length as arch | 33 | 35 | 33 | 32 | |||
| Gubernacular length | 11 | 11 | 11 | 12 | |||
| Cloacal or anal body diameter | 18 | 18 | 18 | 18 | 17 | 18 | 18 |
| Cloacal /Anal body diameter | 1.8 | 1.9 | 1.8 | 1.8 | |||
| Distance from first precloacal supplement to anus | 34 | 31 | 41 | 25 | |||
| Distance from second precloacal supplement to anus | 62 | 68 | 72 | 43 | |||
| Tail length | 136 | 138 | 130 | 134 | 117 | 115 | 129 |
| Vulva from anterior end | 348 | 339 | 343 | ||||
| Vulva c.b.d. | 28 | 30 | 26 | ||||
| V% | 44.1 | 42.7 | 43.3 | ||||
Table 4.
Morphological differentiating data of male Molgolaimus. (belonging to the group1b1 based on classification of Fonseca et al. [9])
Table 4.
Morphological differentiating data of male Molgolaimus. (belonging to the group1b1 based on classification of Fonseca et al. [9])
| species | Body Length (µm) |
Spicule Length (µm) |
numbers of supplements | a | b | c | c’ | spic/abd | References |
|---|---|---|---|---|---|---|---|---|---|
| M. pecticauda | 750-850 | - | absent | 23.6-30.6 | 6.6-7.2 | 7.7-10.2 | - | - | [27] |
| M. sapiens | 435-620 | 35 | absent | 19.5-24.5 | 5.4-6.8 | 8.8-10.5 | 2.6-3.5 | 1.9 | [9] |
| M. spirifer | 880-1060 | 22-25 | absent | 36.6-38.0 | 6.2-7.3 | 8.5-11.2 | 5.4-5.7 | 1.2-1.5 | [28] |
| M. drakus | 495-575 | 19-20 | 1 | 35.5-42.7 | 6.1-7.0 | 7.0-7.9 | 5.3-6.2 | 1.4-1.6 | [9] |
| M. exceptionregulum | 585-600 | 28-31 | 1 | 28.4-31.5 | 5.7-6.1 | 7.5-8.9 | 3.6-4.6 | 1.6-1.8 | [9] |
| M. mareprofundus | 610-630 | 29-32 | 1 | 24.1-30.7 | 5.8-6.7 | 7.2-7.6 | 4.6-5.1 |
1.6-1.9 | [9] |
| M. gazii | 382-430 | 29 | 2 | 27.5 | 5.6 | 6.0-6.1 | - | 2.2-2.9 | [29] |
| longicaudatus sp. nov. | 735-775 | 32-35 | 2 | 28.3-31.0 | 7.4-8.2 | 5.4-5.7 | 7.2-7.7 | 1.8-1.9 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



