
Submitted:
03 July 2024
Posted:
04 July 2024
You are already at the latest version
Abstract
Kynurenic acid (KYNA), a tryptophan metabolite, is believed to exert neuromodulatory and neuroprotective effects in the brain. The study aimed to examine the KYNA’s capacity to modify gene expression and the activity of cellular antioxidant enzymes in specific structures of the sheep brain. Anestrous sheep were infused intracerebroventricularly with two KYNA doses: lower (4×5 μg/60 μL/30 min, KYNA20) and higher (4×25 μg/60 μL/30 min, KYNA100) at 30min intervals. The abundance of superoxide dismutase (SOD2), catalase (CAT) and glutathione peroxidase (GPx1) mRNA, as well as enzyme activities (µM/µg of protein), were determined in the hypothalamic medial-basal (MBH) and preoptic (POA) areas, and in the hippocampal CA1 field. Both doses of KYNA caused a decrease (P<0.01) in the expression of SOD2 and CAT mRNA in all structures examined compared to the control group (except for CAT in the POA at the KYNA100 dose). Furthermore, lower levels of SOD2 mRNA (P<0.05) and CAT mRNA (P<0.01) were found in the MBH and POA and in the POA and CA, respectively, in sheep administered with the KYNA20 dose. Different stimulatory effects on GPx1 mRNA expression were observed for both doses (P<0.05-P<0.01). KYNA exerted stimulatory, but dose-dependent effects on SOD2, CAT and GPx1 activities (P<0.05-P<0.01) in all brain tissues examined. The results indicate that KYNA may influence the level of oxidative stress in individual brain structures in sheep by modulating the expression of genes and activity of at least SOD2, CAT and GPx1. The present findings also expand the general knowledge about the potential neuroprotective properties of KYNA in the central nervous system.
Keywords:
kynurenic acid
; superoxide dismutase
; catalase
; glutathione peroxidase
; hypothalamus
; hippocampus
; sheep
1. Introduction
Kynurenic acid (KYNA) is one of the highly neuroactive products formed during the enzymatic transformation of tryptophan in the kynurenine metabolic pathway (KP) [1]. Within the central nervous system (CNS), KYNA acts as a non-selective antagonist of ionotropic receptors for excitatory amino acids, including glutamate N-methyl-D-aspartate (NMDA) and α7 nicotinic acetylcholine (α7nACh) receptors [1,2]. The compound is also synthesized in peripheral organs and can be found in various dietary food products [3,4]. However, due to its low efficiency of penetrating the blood-brain barrier, the concentration of KYNA in the CNS depends on its local synthesis [5]. KYNA is produced in all types of cells present in the brain, including neurons, oligodendrocytes and glial cells, with the involvement of kynurenine aminotransferases [6,7]. Fluctuations in KYNA concentrations have been observed in the mammalian brain during the pre- and postnatal periods, as well as in adulthood [8]. It is believed that high levels of KYNA in the fetal brain may play a specific role during neurodevelopment, particularly in antagonizing both NMDA and α7nACh receptors [9,10]. Metabolic alterations in KP during adulthood are usually associated with a broad spectrum of neurological and psychiatric disorders. Increased KYNA concentrations have been demonstrated in various brain structures or cerebrospinal fluid (CSF) in Alzheimer’s disease, schizophrenia, bipolar disorder, meningitis, autoimmune diseases and inflammatory processes. Conversely, decreased levels of this compound have been found in Huntington’s disease, Parkinson’s disease and multiple sclerosis [3]. Pharmacological treatments increasing brain KYNA concentration have been shown to reduce excitatory glutamatergic and cholinergic neurotransmission, whereas those capable of decreasing brain KYNA levels facilitated transmission and increased the excitotoxic damage [11,12,13]. Modulating KYNA levels in the brain through pharmacological interventions also affects transmission activity within the dopaminergic and gamma-aminobutyric acid systems [14,15,16]. Consequently, considering the multifaceted action of KYNA, elucidating the mechanisms behind the neuroprotective effects of this compound poses a considerable challenge, often contingent on its concentration in the CNS.
Given the endogenous nature of KYNA and its association with various neuropathologies, the antioxidant activity of this compound has also been demonstrated. Several authors have found its effective ability to scavenge hydroxyl radicals in various non-biological experimental systems [17,18]. KYNA has also been shown to decrease the levels of important markers of oxidative damage produced by different pro-oxidants in tissue preparations [19]. The intensity of neurogenesis and the high sensitivity of the developing nervous system to reactive oxygen species (ROS) may explain the high levels of KYNA in the fetal brain [8], while CNS exposure to oxidative stress during adulthood is one of the factors contributing to neurological disorders [20]. However, KYNA is not a major antioxidant in the brain. Mammalian cells are well adapted to combat oxidative stress and neutralize ROS through the activity of antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) [21]. SOD is a group of enzymes that catalyzes the dismutation of the highly reactive superoxide anion to O2 and the less reactive species hydrogen peroxide (H2O2). CAT decomposes H2O2 into water and molecular oxygen, while the main biochemical function of GPx is to reduce the concentrations of hydroperoxides derived from unsaturated fatty acids. The expression of antioxidant enzyme genes and their activity can be regulated by many factors, with cell oxidative status being a key determinant [22]. These processes are also affected by various endogenous molecules, such as cytokines, neurotrophins and hormones [23,24,25]. Measuring the activity of these major antioxidant enzymes serves as a useful biomarker for the body’s exposure to oxidative stress [26]. Considering the multiple mechanisms relating to the neuroprotective effects of KYNA in the CNS [27], we hypothesized that this compound could modulate the expression of genes and activity of antioxidant enzymes in specific sheep brain structures, such as the hypothalamus and hippocampus. The sheep brain shares many structural similarities with the human brain and can be utilized in studies on mental disorders and neurodegenerative diseases [28]. Among the known isoforms of SOD and GPx, we selected SOD2 and GPx1 for our study, because both isoforms of enzymes are characteristic of tissues rich in mitochondria, including the brain, and provide basic protection of neurons and astrocytes against oxidative stress [21].
2. Results
2.1. SOD2, CAT and GPx1 mRNA Expression Levels in the Hypothalamus and Hippocampus
The expression of gene transcripts of all antioxidant enzymes examined was found in both hypothalamic areas, the POA and MBH, as well as in the hippocampal CA1 field. Differences in the abundance of these transcripts among the treatment groups are shown for individual tissues in Figure 1A,C,E (for MBH), Figure 2A,C,A (for POA) and Figure 3A,C,E (for CA1).
Both doses of KYNA caused a significant decrease (P<0.01) in SOD2 mRNA expression in the MBH, POA and CA1 field of the hippocampus compared to the control group. Furthermore, lower (P<0.05) SOD2 transcript abundance was observed in the MBH and POA of sheep infused with the dose KYNA20 compared to those infused with the dose KYNA100. On the other hand, sheep treated with different KYNA doses expressed SOD2 mRNA at similar levels in the hippocampal CA1 field.
A consistent pattern in response to KYNA infusion was detected for CAT mRNA expression. Evident and highly significant (P<0.01) decreases in the expression were recorded in the MBH and hippocampal CA1 in response to both KYNA doses in comparison to the control group; however, CAT transcript abundance in the CA1 was lower (P<0.05) in sheep infused with the dose KYNA20 than in those receiving the KYNA100 dose. In the POA, the dose KYNA20 reduced (P<0.01) CAT mRNA levels, but the KYNA100 was ineffective compared to controls.
Distinct stimulatory effects of the two KYNA doses were observed for GPx1 mRNA expression. While the dose KYNA20 increased (P<0.01) GPx1 mRNA expression in the POA and hippocampal CA1 field, the dose KYNA100 elevated (P<0.01) the expression in the MBH and CA1 compared to the control group. Moreover, a higher abundance (P<0.05) of GPx1 transcript was noted in the CA1 field of sheep infused with the dose KYNA100 compared to those receiving the dose KYNA20.
2.2. SOD, CAT and GPx Activities in the Hypothalamus and Hippocampus
Differences in the activity of the enzymes examined across the treatment groups are depicted for individual tissues in Figure 1B,D,F (for MBH), Figure 2B,D,F (for POA) and Figure 3B,D,F (for CA1). It should be emphasized that the enzymatic efficiency found in individual brain structures may apply to all isoforms of superoxide dismutase and glutathione peroxidase together.
SOD enzymatic activity in hypothalamic and hippocampal tissues increased (P<0.05 and P<0.01) in response to infusion of both KYNA doses compared to the control group. The increase was more pronounced in the MBH and POA after infusion of the dose KYNA20.
For CAT, a progressive, dose-dependent increase (P<0.05 and P<0.01) in enzyme activity in the MBH and POA was observed compared to the control group. However, in all tissues (MBH, POA and CA1), the highest enzymatic activity was recorded in the group infused with the dose KYNA100 (P<0.01) compared to the other groups.
The effect of KYNA administration on the enzymatic activity of GPx varied. The dose KYNA20 stimulated GPx activity in the POA and CA1 (P<0.01), while the dose KYNA100 exerted stimulatory effect in the MBH and also in the CA1 (P<0.01) compared to the control group. Additionally, the stimulatory effect of the dose KYNA100 was stronger (P<0.05) in the CA1 than that of the KYNA20 dose.
3. Discussion
The primary objective of this study was to analyze the effect of KYNA on the expression of genes and activity of antioxidant enzymes in the hypothalamus and hippocampus in sheep under physiological conditions. These two structures, which in humans and other mammalian species consist of different anatomical areas and small neuronal nuclei, play crucial roles in regulating various physiological processes, such as metabolism and reproduction, as well as cognitive functions like learning, memory and emotion. Furthermore, the hippocampus is recognized for sustaining physiological neurogenesis that continues into adulthood across mammalian species, including humans [29]. Due to their high biochemical activity, these brain regions are continually exposed to highly reactive oxidants generated during normal cellular metabolism, particularly through the mitochondrial energy production pathway [30]. Many studies have demonstrated that excessive accumulation of ROS in nerve cells can lead to oxidative stress, which in turn contributes to various neurological dysfunctions and neurodegenerative diseases [20,31]. Regarding the protection of CNS structures against the harmful effects of oxidants, we observed relatively high and comparable levels of SOD2, CAT and GPx1 mRNA expression in the examined medial-basal and preoptic areas of the hypothalamus, as well as in the CA1 field of the hippocampus. What is more, KYNA appeared to be an important and effective modulator of this defense system: increasing brain KYNA levels by intracerebroventricular infusion resulted in significant and dose-dependent changes in gene expression and the activity of antioxidant enzymes in all structures examined. The potency of the observed response in the tissues may have been influenced by both the dose administered and the distance from the KYNA infusion site, located in the third ventricle. As described previously, the organization of CSF circulation in the sheep brain allows administered compounds to reach more distant areas, among others, the vicinity of the lateral ventricles [32]. Although the dose of infused KYNA appears to be higher than the physiological concentration described in the mammalian brain [27,33], a significant portion of KYNA was expected to flow out of the third ventricle in the mainstream and be absorbed into capillaries, e.g. in the median eminence or other circumventricular organs and the choroid plexus.
The expression of SOD2 and CAT gene transcripts decreased in response to KYNA administration in the MBH, POA and CA1. These two enzymes are a very important pair in the cellular antioxidant defense system, converting superoxide anion (O2-) into H2O2, and subsequently to water and oxygen [21,34]. The observed downregulation of mRNA expression of both antioxidant enzymes could suggest an adverse effect of the compound under study, leading to lower brain tissue protection against oxidative stress. However, considering the KYNA’s ability to independently scavenge free radicals [17,18], it could be speculated that the increased brain KYNA levels resulted in a decreased demand for SOD2 and CAT in the cells. On the other hand, the natural decline in antioxidant enzymes in the body may be associated with age-related decreases in KYNA levels [8,35]. Interestingly, the decrease in SOD2 and CAT mRNA expression was in most cases stronger at the lower dose, indicating differences in sensitivity to KYNA between individual brain structures and/or a possible efficacy threshold for the doses applied. However, a more plausible explanation for the observed changes in the expression of SOD2 and CAT transcripts is increased translation, resulting in higher synthesis of individual enzymes. This, in turn, was reflected by the enhancement of the efficiency of both antioxidant enzymes. While the enhancement of SOD activity was stronger in response to the lower dose, especially in the hypothalamic tissue, CAT activity was significantly more pronounced at the higher one in all tissues tested. Studies have shown that increasing tissue SOD level protects neurons against the death caused by oxidative injury and restores healthy mitochondrial morphology in a monosodium glutamate-induced excitotoxicity disease model [36]. It is believed that impaired SOD2 is a potential pathogenesis related to oxidative stress in PD and AD [37], whereas deficiency or malfunction of CAT is related to the pathogenesis of many age-associated degenerative diseases [38].
An entirely different pattern of response to KYNA was observed for GPx1 mRNA expression, where both the lower and higher dose exerted a stimulatory effect. Particularly, a marked gradual increase in GPx1 mRNA expression level was found in the CA1 field of the hippocampus. Furthermore, changes in the abundance of GPx1 mRNA in the tissues tested were reflected by the increased enzymatic efficiency of GPx. This enzyme removes H2O2 by coupling its reduction with glutathione (GSH) oxidation and helps prevent lipid peroxidation, maintaining intracellular homeostasis and redox balance [21,34]. GSH is therefore an important endogenous compound, highly abundant in the brain, whose primary role is to neutralize free radicals in the cell. Loss of GSH in hippocampal neurons has been shown to lead to dendrite disruption and cognitive impairment [39]. Research by Silva Ferreira et al. [40] demonstrated that KYNA administration in rats was able to prevent an increase in ROS production and SOD/CAT ratio as well as a decrease in GPx activity caused by another excitotoxic metabolite of tryptophan, i.e., quinolinic acid. More recently, sub-chronic administration of L-kynurenine, an intermediate in KYNA synthesis, was found to elevate the GSH content and GPx activity in rat brain tissue, as well as to prevent oxidative damage induced by ex vivo exposure of brain cells to pro-oxidants [41]. Additionally, a moderate elevation of brain KYNA levels was shown to reduce the histopathological and biochemical outcomes of experimentally induced ischemia [42]. Therefore, the observed increase in GPx synthesis and activity in our study may be associated with an increase in cellular GSH concentration and consequently a mobilization of antioxidant protection, particularly in the hippocampal CA1 field. Interestingly, increasing SOD levels in the body, e.g., through dietary supplementation, results in the increase in GPx activity in animal tissues [43]. Relationships between the activities of individual enzymes may also be visible in our study. Importantly, accumulating evidence indicates that GPx-1 has multiple cellular functions: it is involved not only in the protection of cells against oxidative damage, but also in the regulation of metabolism and mitochondrial function, as well as in the control of cellular processes, such as apoptosis, growth, and signaling [44].
However, there is also evidence indicating harmful effects of KYNA on nerve cells, potentially leading to their damage [45,46]. It has been shown that intrathecal infusion of KYNA in the spinal cord, lasting for several days, caused damage and loss of myelin [45]. Moreover, rats infused with the highest concentrations of KYNA demonstrated adverse neurological signs, such as weakness and quadriplegia, linked to diffuse myelin damage. Another in vitro study [46] showed that KYNA administered at high levels reduced the viability of oligodendrocytes, and the mechanism of this action was distinct from that mediated by glutamatergic receptors. It is important to note that these deleterious effects of KYNA were associated with long-term treatment of nervous tissue and extremely high concentrations of the compound applied. Considering the low nanomolar range of extracellular KYNA concentration in the mammalian brain, and the relatively fast turnover rate [27,33], the total concentration of the compound infused in our study could reach levels of several micromoles, predisposing it to interact with various types of receptors.
Since KYNA has several biological targets, understanding its neuroprotective effects is challenging, and multiple mechanisms have been proposed [47]. Apart from its well-established antagonistic effects on NMDA and α7nACh receptors [1,2], KYNA can activate the G-protein-coupled receptor 35 located in the CNS [48], with its potential relevance to the regulation of brain functions still under discussion [49]. KYNA is also an agonist for the aryl hydrocarbon receptor, an attractive target in neurodegenerative diseases, and may therefore contribute to immune and inflammatory regulation [47]. However, the effect of KYNA observed in our study could be largely related to the activation of the transcription factor Nrf2, which is responsible for the expression of a multitude of endogenous antioxidant agents [50]. As shown by the previously cited Silva Ferreira [40], the restoration of the activity of antioxidant enzymes by KYNA was associated with an increase in the cytoplasmic and nuclear Nrf2 levels. Moreover, the ability of KYNA to alter cellular redox balance [17,18] may indicate mechanisms beyond known receptors. Therefore, a comprehensive understanding of the mechanism of KYNA's antioxidant action in the CNS remains to be elucidated.
4. Materials and Methods
4.1. Animal Maintenance
Eighteen Polish Longwool sheep (a breed showing reproductive seasonality), aged 1 year and weighing 55 ± 2 kg, were used in the experiment. The animals were bred at the Sheep Breeding Center of the Kielanowski Institute of Animal Physiology and Nutrition, Polish Academy of Sciences (Jablonna near Warsaw, Poland) under natural lighting conditions (52°N, 21°E). They were fed twice daily according to their physiological status, with a diet based on pellet concentrate, following the recommendations of the National Research Institute of Animal Production in Krakow-Balice (Poland), and the National Institute for Agricultural Research (France) [51]. During the experimental period, sheep were housed in individual pens, ensuring visual, olfactory and tactile contact, and provided free access to water and mineral licks.
4.2. Third Ventricle Cannulation
One month before the experiment, the sheep underwent surgical implantation of a stainless-steel guide cannula into the third ventricle (IIIv) of the brain (outer diameter: 1.2 mm, frontal position: 31.0 mm). Implantation was performed under general anesthesia (xylazine: 40 mg/kg body mass, intravenously; xylapan and ketamine: 10-20 mg/kg body weight, intravenously; Bioketan, Vetoquinol Biowet, Pulawy, Poland), in accordance with the stereotaxic coordinate system for sheep hypothalamus [52] and the procedure described by Traczyk and Przekop [53]. The guide cannula was secured to the skull with stainless-steel screws and dental cement, and the external orifice of the canal was sealed with a stylet. After surgery, the sheep were injected for 4 days with antibiotics (1 g streptomycin and 1,200,000 IU benzylpenicillin; Polfa, Poland) and analgesics (metamizole sodium: 50 mg/animal; Biovetalgin, Biowet Drwalew, Poland, or meloxicam: 1.5 mg/animal; Metacam, Boehringer Ingelheim, Germany). The correct positioning of the cannula in the ventricle was confirmed for all sheep through the efflux of CSF during surgery and examination of the brain after slaughter.
4.3. Experimental Design and Tissue Collection
The experiment was performed in March during the natural anestrous season for this breed of sheep. The animals were randomly divided into three groups (n = 6 each) and infused into the IIIv with Ringer-Locke solution (RLs, control) or with one of two KYNA doses (Sigma Chemical Co., St Louis, MO, USA) dissolved in RLs. The treatment was performed in a series of four 30-min infusions, at 30-min intervals, from 10:00 to 14:00. KYNA doses: lower, 4×5 μg/60 μL/30 min (KYNA20) and higher, 4×25 μg/60 μL/30 min (KYNA100) were selected on the basis of scientific literature [27,33]. All infusions were performed using a BAS Bee microinjection pump (Bioanalytical Systems Inc., West Lafayette, IN, USA) and calibrated 1.0-mL gas-tight syringes. During the treatments, the sheep were kept in pairs in the experimental room in comfortable cages, where they could lie down and to which they had been previously acclimated for three days. Directly after the experiment, the sheep were slaughtered after prior pharmacological stunning (xylazine 0.2 mg/kg body weight and ketamine: 3 mg/kg body weight, intravenously), and the brains were promptly removed from the skull. Following the separation of the median eminence and cerebellum, each brain was sagittally dissected into cerebral hemispheres. The extracted blocks of the hypothalamus (cut to a depth of 2 mm) were dissected into two parts: the medial-basal hypothalamus (MBH) and the preoptic area (POA) [54], according to ovine brain stereotaxic atlas [52]. The optic chiasm, thalamus and mammillary body were utilized as landmarks. The hippocampus was dissected from the medial part of the temporal lobe of the right hemisphere, beginning from the floor of the lateral ventricle through ventral and dorsal parts, according to the sheep brain atlas [55]. Approximately 2-3 mm sections were cut out from the CA1 field of the hippocampus. All tissue incisions were performed on sterile glass plates placed on ice, and subsequently the collected structures were immediately frozen in liquid nitrogen and stored at -80°C.
4.4. Relative mRNA Abundance
Total RNA from hypothalamic and hippocampal tissues was isolated using the NucleoSpin RNA II kit (Macherey-Nagel, Düren, Germany), according to the manufacturer’s protocol. The concentration and purity of isolated RNA were quantified using a NanoDrop ND-1000 spectrophotometer (Thermo Fisher Scientific). RNA integrity was electrophoretically verified on a 1.5% agarose gel stained with ethidium bromide. The TranScriba Kit (A&A Biotechnology, Gdynia, Poland) was used to synthesize cDNA according to the manufacturer’s instructions, utilizing 1 µg of total RNA in a reaction volume of 20 µL. Quantitative polymerase chain reaction (qPCR) was performed using 5× HOT FIREPol® EvaGreen qPCR Mix Plus (Solis BioDyne, Tartu, Estonia). The PCR amplification mix contained 2 µL of cDNA template, 1 µL of primers (0.5 µL each at 10 pmol/mL), 3 µL of buffer PCR Master Mix and 9 µL of dd H2O. The reaction conditions were as follows: initial denaturation at 95°C for 15 min, denaturation at 95°C for 15 s, annealing at 60°C for 20 s, and elongation at 72°C for 20 s (40 cycles). Specific primers for determining the expression of the SOD2, CAT and GPx1 genes, as well as endogenous control genes: glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and peptidylprolyl isomerase C (PPIC), were designed using Primer3 software (The Whitehead Institute, Boston, MA, USA) (Table 1). Amplification specificity was further validated by electrophoresis of the obtained amplicons in a 2% agarose gel and visualized under a UV light camera. Data were analyzed with Rotor Gene 6000 v. 1.7 software (Qiagen, Hilden, Germany) using a comparative quantification option and Relative Expression Software Tool, based on the PCR efficiency correction algorithm developed by Pfaffl et al. [56,57]. The expression levels of the tested genes were normalized using geometrical means of the expression of reference genes. Endogenous con trol genes were assayed in each sample to compensate for variation in cDNA concentration and PCR efficiency between individual tubes.
4.5. Determination of the Antioxidant Enzyme Activity
Total enzymatic activity of SOD was determined using the method described by Fridovich et al. [58], involving superoxide anion generation in the “xanthine-xanthine oxidase system”, with NBT as the O2- detector. The principle of the method is based on two reactions. The first involves superoxide radical generation through the action of xanthine oxidase on xanthine. Subsequently, NBT reacts with the superoxide radical to form a blue-navy formazone dye, whose incrementing level is recorded spectrophotometrically at 540 nm. SOD competes with NBT for the superoxide radical, inhibiting the rate of NBT breakdown. The amount of xanthine oxidase is adjusted so that, in the absence of SOD, the change in absorbance is 0.020 units/minute. A unit of SOD activity is defined as the amount of enzyme that reduces the reaction rate of NBT conversion to formazone by 50%.
CAT activity was determined spectrophotometrically by following the degradation of H2O2 at 240 nm for 2 minutes, following the method outlined by Aebi [59]. Triton X-100 solution was added to phosphate buffer to a final concentration of 0.1%.
Total enzymatic activity of GPx was determined using the method of Hopkins and Tudhope [60], involving the measurement of NADPH oxidation coupled with glutathione disulfide reduction by glutathione reductase. Changes in NADPH concentration were measured at 340 nm for 5 minutes. Enzyme activity was expressed as micromoles of oxidized NADPH2 per µg of protein.
Determinations of all three enzymatic activities were performed using a Freedom EVO® series pipetting station (Tecan, Mannedorf, Switzerland), a modular system with an incubator, shaker and sample plate reader, tailored to the specific application and required throughput. The Freedom Evo system is operated using the Evo software, enabling control of the entire process carried out at the workstation.
In addition, the total protein concentration in tissue homogenates was analyzed spectrophotometrically using the Bradford method with the Bio-Rad Protein Assay Kit II (Bio-Rad, Hercules, CA, USA) according to the manufacturer’s instruction. All enzyme activities, reflecting the rate of substrate conversion into product, are expressed in relative units per µg of protein per unit of time.
4.6. Statistical Analysis
Initially, the Shapiro-Wilk test of normality was carried out to extract parametric and non-parametric data. Statistical assessments of differences in mRNA expression levels of SOD2, CAT and GPx1 in the POA, MBH and CA1 between the treatment groups were carried out using non-parametric statistics, including the Kruskal-Wallis with multiple comparisons of average ranks, and subsequently the Mann-Whitney U test for individual groups. Enzyme activity differences in the brain tissues tested between the groups were analyzed using one-way analysis of variance (STATISTICA, Stat Soft, Tulsa, OK, USA). Subsequently, each analysis was followed by the least significant difference post-hoc test. Differences were considered significant at p < 0.05, and all data are presented as a mean ± standard error of the mean (SEM).
5. Conclusions
In line with KYNA's antioxidant activity, our study in the sheep model, emphasizes the potential of this compound to modulate the expression of genes and activity of cellular antioxidant enzymes in specific brain structures. Considering our and existing data, it can be assumed that KYNA plays a crucial role in maintaining redox homeostasis in brain cells. Furthermore, these results confirm the usefulness of the sheep model in studies concerning the development and/or prevention of neurological disorders in the CNS.
Author Contributions
Conceptualization, T.M. and K.R.-O.; investigation and methodology, T.M., K.R.-O., P.K., P.M. and E.M.; data curation, T.M., K.R.-O and P.K.; writing—original draft preparation, T.M. and K.R.-O.; writing—review and editing, P.K., P.M. and E.M. All authors have read and agreed to the published version of the manuscript.
Funding
The research was supported by a statutory subsidy from the Ministry of Education and Science, Poland, and the Institute’s funds allocated for the 5th edition of “Grant for a Start.”.
Institutional Review Board Statement
All animal procedures were conducted in accordance with the Polish Act on the Protection of Animals Used for Scientific or Educational Purposes (2015) and were approved by the 2nd Local Ethics Committee for Animal Experiments, Warsaw University of Life Sciences – SGGW, Warsaw, Poland (Resolution No. WAW2/128/2020).
Data Availability Statement
The datasets analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors thank W. Mrozek and R. Druchniak for animal care.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Stone, T.W. Kynurenines in the CNS: from endogenous obscurity to therapeutic importance. Prog. Neurobiol. 2001, 64, 185–218. [Google Scholar] [CrossRef] [PubMed]
- Hilmas, C.; Pereira, E.F.; Alkondon, M.; Rassoulpour, A.; Schwarcz, R.; Albuquerque, E.X. The brain metabolite kynurenic acid inhibits alpha7 nicotinic receptor activity and increases non-alpha7 nicotinic receptor expression: physiopathological implications. J. Neurosci. 2001, 21, 7463–7473. [Google Scholar] [CrossRef]
- Kozłowska, M.; Kozłowski, P. Kynurenine acid - metabolism and regulation of kynurenine pathway. J. Edu. Health Sport. 2017, 7, 888–895. [Google Scholar]
- Turski, M.P.; Turska, M.; Zgrajka, W.; Kuc, D.; Turski, W.A. Presence of kynurenic acid in food and honeybee products. Amino Acids 2009, 36, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Fukui, S.; Schwarcz, R.; Rapoport, S.I.; Takada, Y.; Smith, Q.R. Blood-brain barrier transport of kynurenines: implications for brain synthesis and metabolism. J. Neurochem. 1991, 56, 2007–2017. [Google Scholar] [CrossRef]
- Du, F.; Schmidt, W.; Okuno, E.; Kido, R.; Köhler, C.; Schwarcz, R. Localization of kynurenine aminotransferase immunoreactivity in the rat hippocampus. J. Comp. Neurol. 1992, 321, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Wejksza, K.; Rzeski, W.; Okuno, E.; Kandefer-Szerszeń, M.; Albrecht, J. Turski, W.A. Demonstration of kynurenine aminotransferases I and II and characterization of kynurenic acid synthesis in oligodendrocyte cell line (OLN-93). Neurochem. Res. 2005, 30, 963–968. [Google Scholar] [CrossRef]
- Cannazza, G.; Chiarugi, A.; Parenti, C.; Zanoli, P.; Baraldi, M. Changes in kynurenic, anthranilic, and quinolinic acid concentrations in rat brain tissue during development. Neurochem. Res. 2001, 26, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Ceresoli-Borroni, G.; Schwarcz, R. Perinatal kynurenine pathway metabolism in the normal and asphyctic rat brain. Amino Acids. 2000, 19, 311–323. [Google Scholar] [CrossRef]
- Notarangelo, F.M.; Pocivavsek, A. Elevated kynurenine pathway metabolism during neurodevelopment: Implications for brain and behavior. Neuropharmacology 2017, 112, 275–285. [Google Scholar] [CrossRef]
- Du, F.; Eid, T.; Schwarcz, R. Neuronal damage after the injection of aminooxyacetic acid into the rat entorhinal cortex: a silver impregnation study. Neuroscience 1998, 82, 1165–1178. [Google Scholar] [CrossRef] [PubMed]
- Carpenedo, R.; Pittaluga, A.; Cozzi, A.; Attucci, S.; Galli, A.; Raiteri, M.; Moroni, F. Presynaptic kynurenate-sensitive receptors inhibit glutamate release. Eur. J. Neurosci. 2001, 13, 2141–2147. [Google Scholar] [CrossRef] [PubMed]
- Zmarowski, A.; Wu, H.Q.; Brooks, J.M.; Potter, M.C.; Pellicciari, R.; Schwarcz, R.; Bruno, J.P. Astrocyte-derived kynurenic acid modulates basal and evoked cortical acetylcholine release. Eur. J. Neurosci. 2009, 29, 529–538. [Google Scholar] [CrossRef]
- Rassoulpour, A.; Wu, H.Q.; Ferré, S.; Schwarcz, R. Nanomolar concentrations of kynurenic acid reduce extracellular dopamine levels in the striatum. J. Neurochem. 2005, 93, 762–765. [Google Scholar] [CrossRef]
- Amori, L.; Wu, H.Q.; Marinozzi, M.; Pellicciari, R.; Guidetti, P.; Schwarcz, R. Specific inhibition of kynurenate synthesis enhances extracellular dopamine levels in the rodent striatum. Neuroscience 2009, 159, 196–203. [Google Scholar] [CrossRef]
- Beggiato, S.; Tanganelli, S.; Fuxe, K.; Antonelli, T.; Schwarcz, R.; Ferraro, L. Endogenous kynurenic acid regulates extracellular GABA levels in the rat prefrontal cortex. Neuropharmacology 2014, 82, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R.; Zsizsik, B.K. Kynurenic acid as a free radical scavenger: measurements of educt and product fluorescence and of light emission from an excited intermediate state. In: Hardeland R, ed. Biological rhythms and antioxidative protection. Göttingen: Cuvillier. 1997, p. 153-160.
- Zsizsik, B.K.; Hardeland, R. A putative mechanism of kynurenic acid oxidation by free radicals: scavenging of two hydroxyl radicals and superoxide anion, release of •NO and CO2. In: Hardeland R, ed. Actions and redox properties of melatonin and other aromatic amino acid metabolites. Göttingen: Cuvillier; 2001, p. 164-167.
- Lugo-Huitrón, R.; Blanco-Ayala, T.; Ugalde-Muñiz, P.; Carrillo-Mora, P.; Pedraza-Chaverrí, J.; Silva-Adaya, D.; Maldonado, P.D.; Torres, I.; Pinzón, E.; Ortiz-Islas, E.; López, T.; García, E.; Pineda, B.; Torres-Ramos, M.; Santamaría, A.; La Cruz, V.P. On the antioxidant properties of kynurenic acid: Free radical scavenging activity and inhibition of oxidative stress. Neurotoxicol. Teratol. 2011, 33, 538–347. [Google Scholar] [CrossRef]
- De Felice, A.; Ricceri, L.; Venerosi, A.; Chiarotti, F.; Calamandrei, G. Multifactorial origin of neurodevelopmental disorders: approaches to understanding complex etiologies. Toxics 2015, 3, 89–129. [Google Scholar] [CrossRef]
- Deska, M. Activity of antioxidant enzymes under induced oxidative stress. J. Ecol. Engineer. 2020, 21, 42–51. [Google Scholar] [CrossRef]
- Pahl, H.L.; Baererle, P.A. Oxygen and the control of gene expression. BioEssays 1994, 16, 497–502. [Google Scholar] [CrossRef]
- Rogers, R.J.; Chesrown, S.E.; Kuo, S.; Monnier, J.M.; Nick, H.S. Cytokine-inducible enhancer with promoter activity in both the rat and human manganese-superoxide dismutase genes. Biochem. J. 2000, 347, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Sampath, D.; Perez-Polo, R. Regulation of antioxidant enzyme expression by NGF. Neurochem. Res. 1997, 22, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Antolin, I.; Rodriguez, C.; Sainz, R.M.; Mayo, J.C.; Uría, H.; Kotler, M.L.; Rodríguez-Colunga, M.J.; Tolivia, D.; Menéndez-Peláez, A. Neurohormone melatonin prevents cell damage: effect on gene expression for antioxidant enzymes. FASEB J. 1996, 10, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Czerska, M.; Mikołajewska, K.; Zieliński, M.; Gromadzińska, J.; Wąsowicz, W. Today’s oxidative stress markers. Medycyna Pracy (in Polish). 2015, 66, 393–405. [Google Scholar] [CrossRef]
- Moroni, F.; Cozzi, A.; Sili, M.; Mannaioni, G. Kynurenic acid: a metabolite with multiple actions and multiple targets in brain and periphery. J. Neural Transm. 2012, 119, 133–139. [Google Scholar] [CrossRef]
- Murray, S.J.; Mitchell, N.L. The translational benefits of sheep as large animal models of human neurological disorders. Front. Vet. Sci. 2022, 9, 831838. [Google Scholar] [CrossRef] [PubMed]
- van Praag H, Schinder AF, Christie BR, Toni N, Palmer TD, Gage FH. Functional neurogenesis in the adult hippocampus. Nature 2002, 415, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Cadanes, E.; Davies, K.J. Mitochondrial free radical generation, oxidative stress, and aging. Free Radic. Biol. Med. 2000, 29, 222–230. [Google Scholar]
- Espinet, C.; Gonzalo, H.; Fleitas, C.; Menal, M.J.; Egea, J. Oxidative stress and neurodegenerative diseases: A neurotrophic approach. Cur. Drug Targ. 2015, 16, 20–30. [Google Scholar] [CrossRef]
- Roszkowicz-Ostrowska, K.; Młotkowska, P.; Kowalczyk, P.; Marciniak, E.; Barszcz, M.; Misztal, T. Central stimulatory effect of kynurenic acid on BDNF-TrkB signaling and BER enzymatic activity in the hippocampal CA1 field in sheep. Int. J. Mol. Sci. 2023, 24, 136. [Google Scholar] [CrossRef]
- Stone, T.W. Development and therapeutic potential of kynurenic acid and kynurenine derivatives for neuroprotection. Trends Pharmacol. Sci. 2000, 21, 149–154. [Google Scholar] [CrossRef]
- Bratovcic, A. Antioxidant enzymes and their role in preventing cell damage. Acta Sci. Nutrition. Health 2020, 4, 132–138. [Google Scholar] [CrossRef]
- Inal, M.E.; Kanbak, G.; Sunal, E. Antioxidant enzyme activities and malondialdehyde levels related to aging. Clin. Chim. Acta 2001, 305, 75–80. [Google Scholar] [CrossRef]
- Liao, R.; Wood, T.R.; Nance, E. Superoxide dismutase reduces monosodium glutamate-induced injury in an organotypic whole hemisphere brain slice model of excitotoxicity. J. Biol. Eng. 2020, 14, 1–12. [Google Scholar] [CrossRef]
- Niedzielska, E.; Smaga, I.; Gawlik, M.; Moniczewski, A.; Stankowicz, P.; Pera, J.; Filip, M. Oxidative stress in neurodegenerative diseases. Mol. Neurobiol. 2016, 53, 4094–4125. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Cha, M.; Lee, B.H. Neuroprotective Effect of Antioxidants in the Brain. Int. J. Mol. Sci. 2020, 21, 7152. [Google Scholar] [CrossRef]
- Fernandez-Fernandez, S.; Bobo-Jimenez, V.; Requejo-Aguilar, R.; Gonzalez-Fernandez, S.; Resch, M.; Carabias-Carrasco, M.; Ros, J.; Almeida, A.; Bolaños, J.P. Hippocampal neurons require a large pool of glutathione to sustain dendrite integrity and cognitive function. Redox Biology 2018, 19, 52–61. [Google Scholar] [CrossRef]
- Silver Ferreira, F.; Schmitz, F.; Marques, E.P.; Siebert, C.; Wyse, A.T.S. Intrastriatal quinolinic acid administration impairs redox homeostasis and induces inflammatory changes: Prevention by kynurenic acid. Neurotox. Res. 2020, 38, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Ramírez Ortega, D.; Ugalde Muñiz, P.E.; Blanco Ayala, T.; Vázquez Cervantes, G.I.; Lugo Huitrón, R.; Pineda, B.; González Esquivel, D.F.; Pérez de la Cruz, G.; Pedraza Chaverrí, J.; Sánchez Chapul, L.; Gomez-Manzo, S.; Perez de la Cruz, V. On the antioxidant properties of L-kynurenine: An efficient ROS scavenger and enhancer of rat brain antioxidant defense. Antioxidants 2022, 11, 31. [Google Scholar] [CrossRef]
- Bratek-Gerej, E.; Ziembowicz, A.; Godlewski, J.; Salinska, E. The mechanism of the neuroprotective effect of kynurenic acid in the experimental model of neonatal hypoxia–ischemia: The link to oxidative stress. Antioxidants 2021, 10, 1775. [Google Scholar] [CrossRef]
- Yan, Z.; Liu, S.; Liu, Y.; Zheng, M.; Peng, J.; Chen, Q. Effects of dietary superoxide dismutase on growth performance, antioxidant capacity and digestive enzyme activity of yellow-feather broilers during the early breeding period (1-28d). J. Anim. Feed Sci. 2022, 31, 232–240. [Google Scholar] [CrossRef]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: from molecular mechanisms to therapeutic opportunities. Antioxid Redox Signal. 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [PubMed]
- Dabrowski, W.; Kwiecien, J.M.; Rola, R.; Klapec, M.; Stanisz, G.; Kotlinska-Hasiec, E.; Oakden, W.; Janik, R.; Coote, M.; Frey, B.N.; Turski, W.A. Prolonged subdural infusion of kynurenic acid is associated with dose-dependent myelin damage in the rat spinal cord. PLoS ONE 2015, 10, e0142598. [Google Scholar] [CrossRef]
- Langner, E.; Lemieszek, M.K.; Kwiecień, J.M.; Rajtar, G.; Rzeski, W.; Turski, W.A. Kynurenic acid induces impairment ofoligodendrocyte viability: On the role of glutamatergic mechanisms. Neurochem. Res. 2017, 42, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Schwarcz, R.; Stone, T.W. The kynurenine pathway and the brain: challenges, controversies and promises. Neuropharmacology 2017, 112, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Simonavicius, N.; Wu, X.; Swaminath, G.; Reagan, J.; Tian, H.; Ling, L. Kynurenic acid as a ligand for orphan G protein-coupled receptor GPR35. J. Biol. Chem. 2006, 281, 22021–22028. [Google Scholar] [CrossRef] [PubMed]
- Alkondon, M.; Pereira, E.F.; Todd, S.W.; Randall, W.R.; Lane, M.V.; Albuquerque, E.X. Functional G-protein-coupled receptor 35 is expressed by neurons in the CA1 field of the hippocampus. Biochem. Pharmacol. 2015, 93, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Ngo, V.; Duennwald, M.L. Nrf2 and oxidative stress: A general Overview of mechanisms and implications in human disease. Antioxidants 2022, 11, 2345. [Google Scholar] [CrossRef] [PubMed]
- Strzetelski, J. IZ PIB–INRA Feeding recommendations for ruminants and feed tables. Kraków, Poland 2014 (in Polish).
- Welento, J.; Szteyn, S.; Milart, Z. Observations on the stereotaxic configuration of the hypothalamus nuclei in the sheep. Anat. Anz. 1969, 124, 1–27. [Google Scholar]
- Traczyk, W.; Przekop, F. Methods of investigation of the function of the hypothalamus and hypophysis in chronic experiments in sheep. Acta Physiol. Pol. 1963, 14, 227–236. [Google Scholar]
- Ciechanowska, M.; Łapot, M.; Malewski, T.; Misztal, T.; Mateusiak, K.; Przekop, F. Effect of stress on the expression of GnRH gene and GnRH receptor (GnRH-R) genes in the preoptic area-hypothalamus and GnRH-R gene in the stalk/median eminence and anterior pituitary gland in the ewes during follicular phase of the estrous cycle. Acta Neurobiol. Exp. 2007, 67, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.I.; Sudheimer, K.D.; Davis, K.K.; , Kerndt, G.M.; Winn, B.M. The sheep brain atlas. Michigan State University, Brain Biodiversity Bank. Available online: http://brains.anatomy.msu.edu/brains/sheep/index.html (accessed on 5 March 2021).
- Pfaffl, M.W.; Horgan, G.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, 36. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper – Excel-based tool using pairwise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Fridovich, I. Oxygen radicals, hydrogen peroxide and oxygen toxicity. In Free Radicals in Biology, Pryor W.A., Ed., New York, Academic Press, 1976, 239.
- Aebi, H. Methods Enzymol; Bergmeyer, H.U., Ed.; Academic Press: New York, London, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Hopkins, J.; Tudhope, GR. Glutathione peroxidase in human red cells in health and disease. Br. J. Haematol. 1973, 25, 563–575. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Relative mRNA abundance (mean ± SEM, left panel) and enzyme activity (mean ± SEM, right panel) of superoxide dismutase (SOD, A, B - respectively), catalase (CAT, C, D - respectively) and glutathione peroxidase (GPx, E, F - respectively) in the medial-basal hypothalamus of sheep infused into the third ventricle with control solution and the lower (total 20 µg, KYNA20) and higher (total 100 µg, KYNA100) dose of kynurenic acid (KYNA). Significance of differences: AB, CD, p < 0.01, ab, cd, p < 0.05.
Figure 1.
Relative mRNA abundance (mean ± SEM, left panel) and enzyme activity (mean ± SEM, right panel) of superoxide dismutase (SOD, A, B - respectively), catalase (CAT, C, D - respectively) and glutathione peroxidase (GPx, E, F - respectively) in the medial-basal hypothalamus of sheep infused into the third ventricle with control solution and the lower (total 20 µg, KYNA20) and higher (total 100 µg, KYNA100) dose of kynurenic acid (KYNA). Significance of differences: AB, CD, p < 0.01, ab, cd, p < 0.05.
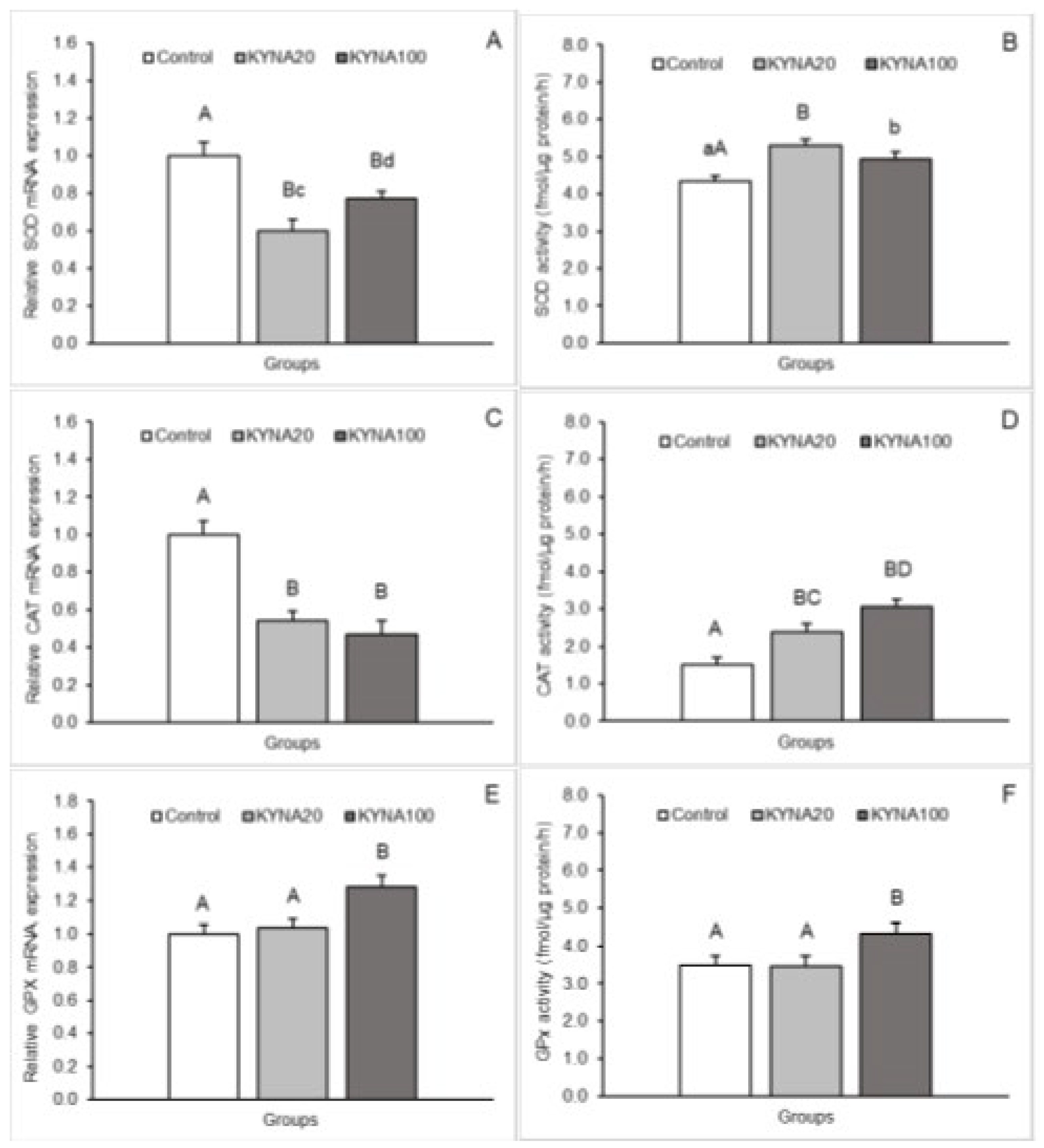
Figure 2.
Relative mRNA abundance (mean ± SEM, left panel) and enzyme activity (mean ± SEM, right panel) of superoxide dismutase (SOD, A, B - respectively), catalase (CAT, C, D - respectively) and glutathione peroxidase (GPx, E, F - respectively) in the hypothalamic preoptic area of sheep infused into the third ventricle with control solution and the lower (total 20 µg, KYNA20) and higher (total 100 µg, KYNA100) dose of kynurenic acid (KYNA). Significance of differences: AB, CD, p < 0.01, ab, cd, p < 0.05.
Figure 2.
Relative mRNA abundance (mean ± SEM, left panel) and enzyme activity (mean ± SEM, right panel) of superoxide dismutase (SOD, A, B - respectively), catalase (CAT, C, D - respectively) and glutathione peroxidase (GPx, E, F - respectively) in the hypothalamic preoptic area of sheep infused into the third ventricle with control solution and the lower (total 20 µg, KYNA20) and higher (total 100 µg, KYNA100) dose of kynurenic acid (KYNA). Significance of differences: AB, CD, p < 0.01, ab, cd, p < 0.05.
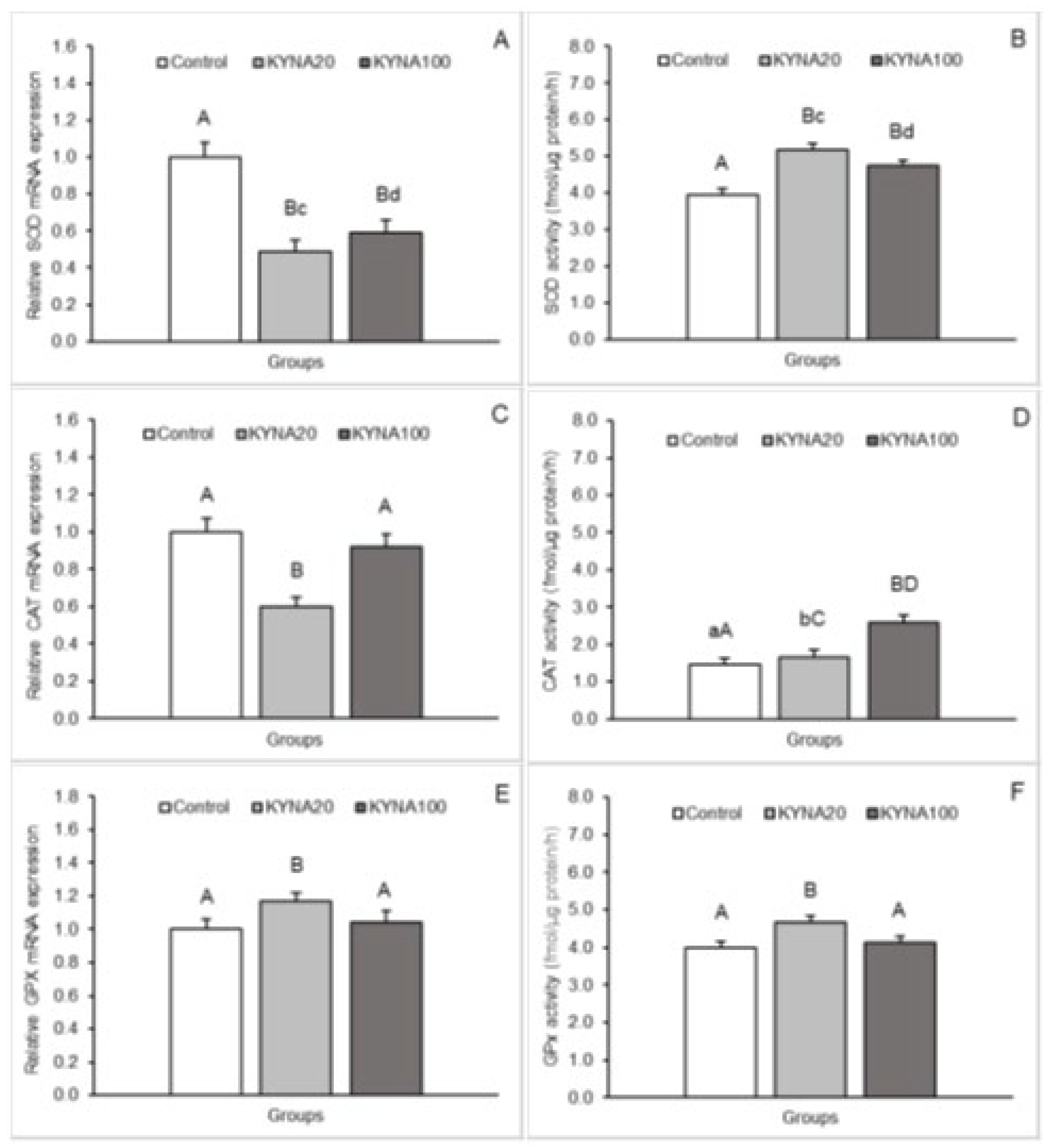
Figure 3.
Relative mRNA abundance (mean ± SEM, left panel) and enzyme activity (mean ± SEM, right panel) of superoxide dismutase (SOD, A, B - respectively), catalase (CAT, C, D - respectively) and glutathione peroxidase (GPx, E, F - respectively) in the hippocampal CA1 field of sheep infused into the third ventricle with control solution and the lower (total 20 µg, KYNA20) and higher (total 100 µg, KYNA100) dose of kynurenic acid (KYNA). Significance of differences: AB, p < 0.01, cd, p < 0.05.
Figure 3.
Relative mRNA abundance (mean ± SEM, left panel) and enzyme activity (mean ± SEM, right panel) of superoxide dismutase (SOD, A, B - respectively), catalase (CAT, C, D - respectively) and glutathione peroxidase (GPx, E, F - respectively) in the hippocampal CA1 field of sheep infused into the third ventricle with control solution and the lower (total 20 µg, KYNA20) and higher (total 100 µg, KYNA100) dose of kynurenic acid (KYNA). Significance of differences: AB, p < 0.01, cd, p < 0.05.
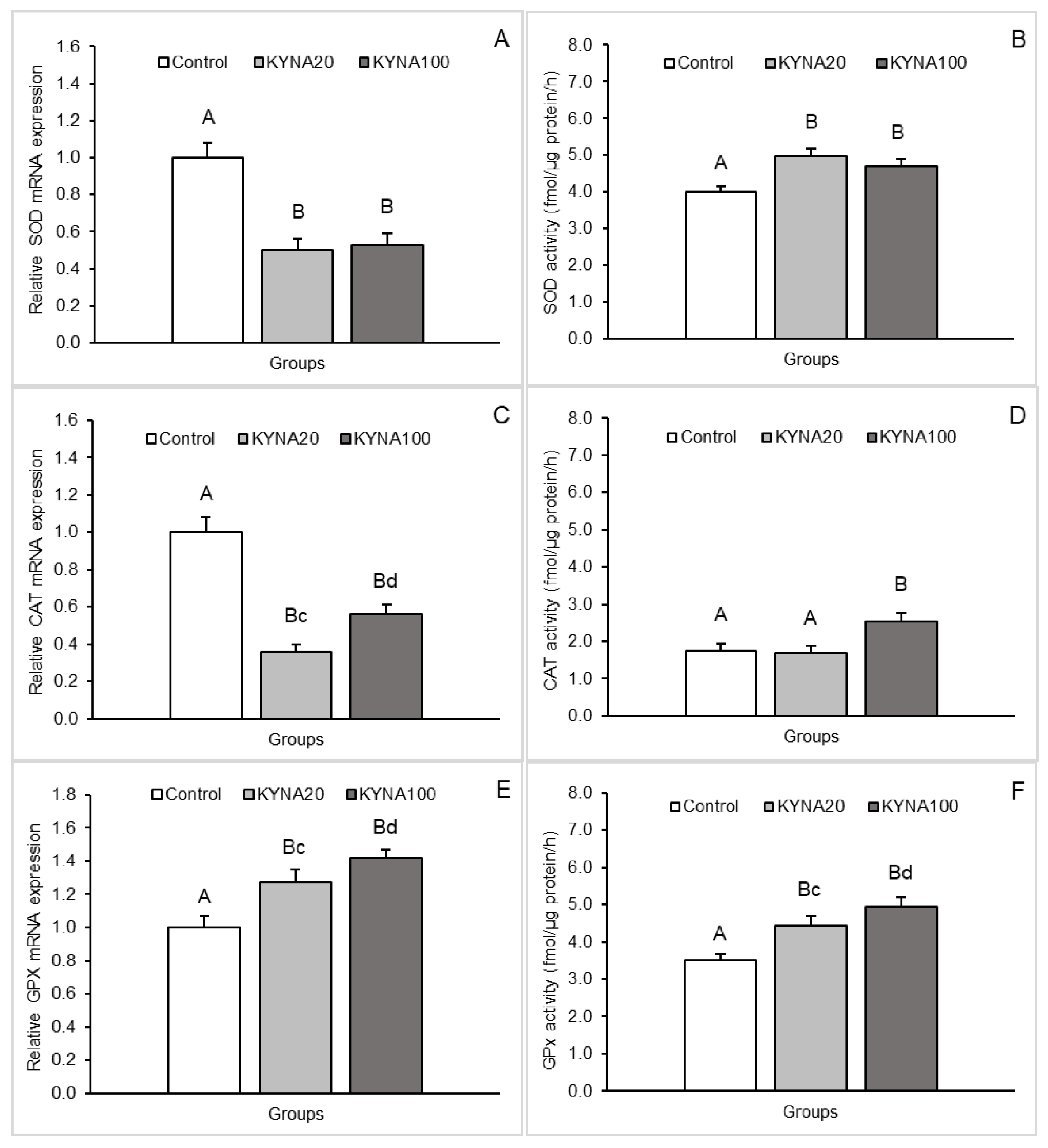

Table 1.
Sequences of primer pairs used in the study.
| Gene | Primers (5’ – 3’) | Genbank Acc. No. | Amplicon size |
|---|---|---|---|
| SOD2 | F: GCAAGGAACAACAGGTCTTATCC R: ACTTGGTGTAAGGCTGACGG |
NM_001280703.1 | 181 |
| CAT | F: GAGCCCACCTGCAAAGTTCT R: CTCCTACTGGATTACCGGCG |
XM_004016396.6 | 148 |
| GPx1 | F: TGTCGTACTCGGCTTCCC R: AGCGGATGCGCCTTCTCG |
XM_004018462.1 | 163 |
| GAPDH | F: GGGTCATCATCTCTGCACCT R: GGTCATAAGTCCCTCCACGA |
NM_001190390.1 | 131 |
| PPIC | F: TGGAAAAGTCGTGCCCAAGA R: TGCTTATACCACCAGTGCCA |
XM_004008676.1 | 158 |
SOD2: superoxide dismutase-2, CAT: catalase, GPx1: glutathione peroxidase-1, GAPDH: glyceraldehyde-3-phosphate dehydrogenase, PPIC: peptidylprolyl isomerase C, F: forward primer, R: reverse primer. Real-Time PCR amplification efficiencies of target and reference genes was 96 - 101%.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



