
Submitted:
06 July 2024
Posted:
08 July 2024
You are already at the latest version
Abstract
Gestational trophoblastic diseases (GTDs) encompass a spectrum of conditions characterized by abnormal trophoblastic cell growth, ranging from benign molar pregnancies to malignant trophoblastic neoplasms. This systematic review explores the molecular underpinnings of GTDs, focusing on genetic and epigenetic factors that influence disease progression and clinical outcomes. Based on 65 studies identified through systematic search and selection criteria, key findings include dysregulations in tumor suppressor genes such as p53, aberrant apoptotic pathways involving BCL-2, and altered expression of growth factor receptors and microRNAs. These molecular alterations not only differentiate molar pregnancies from normal placental development but also contribute to their clinical behavior, from benign moles to potentially malignant forms. The review synthesizes insights from immunohistochemical studies and molecular analyses to provide a comprehensive understanding of GTD pathogenesis and implications for personalized care strategies.
Keywords:
Gestational trophoblastic disease
; Hydatidiform Mole
; Complete molar pregnancy
; Partial molar pregnancy
; Choriocarcinoma
; Molecular basis of molar pregnancy
1. Introduction
Gestational trophoblastic diseases (GTDs) encompass a spectrum of conditions that range from benign to malignant forms and are characterized by abnormal growth of trophoblastic cells. The benign forms of GTD comprise molar pregnancies, which include complete hydatidiform moles (CHMs) and partial hydatidiform moles (PHMs). These two entities have varying clinical presentations and outcomes [1]. The malignant forms of GTD, also known as gestational trophoblastic neoplasms (GTN), include choriocarcinoma (CC) and invasive moles (IM), epithelioid trophoblastic tumor (ETT), and placental-site trophoblastic tumor (PSTT). These neoplasms can arise de novo, after a normal pregnancy, or, most commonly, following a molar pregnancy [2]. [Figure 1]
Molar pregnancies typically result from errors in gamete formation and the fertilization process, which lead to the formation of an abnormal zygote with an abnormal amount of parental genetic material contributions. In CHM, the total amount of genetic material is normal, but all chromosomes are paternal without maternal chromosomes [2]. In CHM, an enucleated egg is either fertilized by one sperm whose genetic material then duplicates (more common) or is fertilized by two sperms (less common). The result is a diploid zygote with 46 paternal chromosomes and no maternal genetic material (i.e., androgenetic diploid) [3]. However, there is a subtype of CHM called familial recurrent hydatidiform moles (FRHM) where cells are biparental diploid (i.e., cells contain 23 paternal chromosomes and 23 maternal chromosomes) [4,5]. This entity will be discussed in detail later in this review. In PHM, a normal egg (1n) is fertilized by more than one sperm, resulting in polyploidy [3]. Usually, there is triploidy with an extra paternal haploid set of chromosomes, with the most common type being 69, XXX (90% of cases) [6].
The incidence of molar pregnancies varies across different world regions, ranging from 0.2 to 9.9 per 1000 pregnancies. However, its incidence is higher in Asian and African ethnicities [7]. The possibility of developing post-molar GTN is the primary reason for extended follow-up with serial beta-HCG monitoring in patients after the evacuation of molar pregnancies. The incidence of post-molar pregnancy is around 15-20% in CHM but is only 0.5-5% in PHM [8,9]. This discrepancy is the reason follow-up protocols after the evacuation of a molar pregnancy differ for each subtype. Thus, it is essential to differentiate between the two entities during histopathological testing. The diagnosis of molar pregnancy follows specific morphological criteria through histopathological examination. The microscopic appearance differs between CHMs, PHMs, and hydropic abortions (HAs) [10]. However, challenges arise in differentiating between CHM and PHM, and even between molar pregnancy and hydropic abortions (HA), due to overlapping histological signs, especially in the early stages of pregnancy [11,12]. The advent of ultrasonographic machines has made it possible to diagnose molar pregnancies in the early stages, which has paradoxically led to a more confusing morphological assessment and sometimes erroneous interpretation of tissue specimens during histopathological examination [11,12]. Multiple ancillary techniques utilizing immunohistochemical staining and genetic analyses have emerged to help differentiate between CHMs, PHMs, and HAs [13].
Research into these ancillary techniques has shed light on the intricate molecular mechanisms driving molar pregnancies. These include dysregulations in tumor suppressor genes such as p53, apoptotic pathways involving BCL-2, and aberrant expression of growth factor receptors and microRNAs [14,15]. These molecular alterations not only differentiate molar pregnancies from normal placental development but also contribute to their clinical behavior, ranging from benign moles to potentially malignant forms [16,17,18]. This review explores the current understanding of molecular markers and genetic mutations in molar pregnancies, emphasizing their roles in cell proliferation, apoptosis, and trophoblastic differentiation. By synthesizing findings from immunohistochemical studies and molecular analyses, this article aims to provide insights into the pathogenesis of GTDs.
2. Materials and Methods
2.1. Search Strategy
A systematic search was conducted across electronic databases including PubMed, Scopus, and CINAHL EBSCO. Medical Subject Headings (MeSH) terms and free-text keywords such as “Hydatidiform mole,” “gestational trophoblastic disease,” and “molar pregnancy” were used in conjunction with “gene,” “genome,” “genetic”, “immunohistochemistry” or “molecular” in February 2024 to retrieve relevant data from January 2004 to January 2024. Additionally, the references of pertinent studies were manually searched if they were not included in these databases.
2.2. Eligibility Criteria:
2.2.1. Inclusion Criteria
Studied were included if they met all the following criteria-1,2, and 3:
1. Articles written in English language or English translation is available;
2. Full-text articles reporting on human genes, genome, genetics, or molecular bases;
3. Articles containing information on gestational trophoblastic disease or hydatidiform mole or molar pregnancy
All eligible studies published between January 2014 and January 2024 were included for review.
2.2.2. Exclusion Criteria
Exclusions criteria included duplicate studies, review articles, non-genetic and non-immunohistochemical studies, articles not available in the English language, and studies where full-text articles were only available with payment. Conference abstracts, expert opinions, and critical appraisals were also excluded from the review.
2.3. Study Selection
After a comprehensive search of the databases, a total of 5259 results were initially retrieved. All the abstracts and study titles were screened, and duplicates were removed. Furthermore, there were a total of 5194 studies excluded, as they either did not fit the inclusion criteria, were animal studies, included only gestational trophoblastic neoplasia, placental abnormalities other than hydatidiform mole, or did not explore the genetic basis of the disease. Eventually, 65 articles met all criteria and were included in the review.
2.4. Data Collection
All authors (SB, RD, MA, MG, RS, RK, SSK) independently reviewed all titles. The potential relevance of studies for inclusion in the review was determined through discussion. Selected titles and abstracts were further screened to avoid overlap of cases. Full-text copies of the selected papers were obtained, and the same reviewers independently extracted relevant data regarding study characteristics, results of molecular testing, and significant associations with GTD outcomes. Whenever possible, single case reports were cross-checked with other reports from the same location and hospital. Finally, studies were screened by assessing their suitability for inclusion in the evidence acquisition of the molecular basis of hydatidiform mole. Figure 2 illustrates the Preferred Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA).
3. Results and Discussion
3.1. Genomic Imprinting
Genomic imprinting, regulated by DNA methylation at differentially methylated regions (DMRs), governs the expression of genes based on their parental origin [19]. In complete moles, which are predominantly androgenetic diploids, the absence of maternal genomic contribution results in significant implications for imprinted genes such as p57kip2 [5,6,20]. P57kip2, paternally imprinted and expressed exclusively from the maternal allele, exhibits markedly reduced expression in CHMs compared to partial moles and normal placentas [21,22,23,24,25]. This distinct immunohistochemical pattern of p57kip2 has become crucial for distinguishing between different types of hydatidiform moles [11,12,26]. Studies using human trophoblast stem cells have highlighted p57kip2’s role in regulating contact inhibition, suggesting that its diminished expression contributes to the uncontrolled proliferation characteristic of CHMs [27]. Despite rare instances of biparental diploid CHMs [5,28], and retained maternal copy of respective chromosomes the majority exhibit abnormal genomic imprinting and epigenetic dysregulation, including down-regulation of DNMT3A and LIN28B, key enzymes for DNA methylation and parental imprinting maintenance [30,31,32]. Sanchez-Delgado et al. confirmed abnormalities in epigenomic regulation of placenta-specific maternally inherited genes, characterized by atypical methylation patterns [33]. Abnormal methylation patterns affect genes like ERVWE1 (Syncyntin-1), implicated in syncytiotrophoblast function and apoptosis regulation, further underlining the impact of epigenetic changes in CHM pathogenesis [34,35]. Other similar genes affected by abnormal methylation are LINE-1 and indirectly STAT5A [36,37]. While typically p57-negative, rare cases of CHMs show aberrant p57 expression linked to retained maternal chromosomes or biparental diploidy, emphasizing the utility of genotyping in diagnosing and understanding the molecular basis of molar pregnancies [38,39,40,41]. Similarly, although partial hydatidiform moles are typically p57-positive, rare instances (1.3%) have been reported with negative p57 results [42]. Genomic imprinting in gametogenesis is responsible for genetic expression in the off-spring depending on parent-of-origin and the human placenta has high and prolific expression of imprinted genes [43,44]. The genotype of molar pregnancies, particularly heterozygous/dispermic complete moles, correlates with a higher risk of developing post-molar gestational trophoblastic neoplasia, suggesting a potential role of additional paternal imprints in promoting trophoblastic proliferation [16]. Further research is needed to elucidate the precise mechanisms underlying these genomic imprinting abnormalities and their clinical implications in molar pregnancy pathogenesis [1,2,3,4].
3.2. Recurrent Hydatidiform Mole
Recurrent hydatidiform moles (RHM) encompass two distinct categories: sporadic cases and familial forms. Sporadic RHM typically occur with a recurrence rate of 1-6% among individuals with a history of molar pregnancy [45]. These cases often exhibit a monospermic or dispermic androgenetic diploid genotype [5]. In contrast, familial RHM (FRHM) are generally biparental diploids and characterized by two or more affected female family members experiencing recurrent molar pregnancies [4,5]. These familial cases are considered autosomal recessive inherited disorders and are associated with abnormal CpG methylation in imprinted genes [46].
The predominant genetic culprits linked to FRHM include mutations in NLRP7 (Nucleotide-Binding Oligomerization Domain, Leucine Rich Repeat and Pyrin Domain-Containing 7) and KHDC3L (KH Domain Containing 3 Like), with NLRP7 mutations found in 40-80% and KHDC3L mutations in 10-14% of affected individuals [45,47]. A mutation analysis by Nguyen et al. of 113 patients with recurrent molar pregnancies showed that mutations with NLRP7 and KHDC3L were associated with diploid biparental HM, while recurrent molar pregnancies without mutations were associated mostly with diploid androgenic monospermic and triploid biparental dispermic [48].
Multiple case reports and case series identified mutations in homozygous state or combined heterozygous states involving NLRP7 and KHDC3L genes associated with recurrent hydatidiform moles [46,49,50,51,52,53]. NLRP7, located on chromosome 19q13.4, is a maternal effect gene, which means that the phenotype is influenced by mutations in maternal genes only, crucial for oocyte and embryo development [54]. It localizes in the cytoskeleton which contains microtubules essential for cellular division [54]. Homozygous mutations in NLRP7 disrupt embryo development by impairing cellular division and organization within the cytoskeleton, leading to developmental arrest and failed embryo progression. In a study investigating 10 embryos from a woman with recurrent hydatidiform mole and a homozygous pathogenic variant in NLRP7 (c.2810+2T>G), all embryos exhibited developmental arrest and did not advance to a stage suitable for transfer [55]. This mutation primarily affects embryos when inherited maternally, owing to its maternal inheritance pattern [46]. Additionally, NLRP7 interacts with YY1 (Yin Yang 1) to activate BMP4 (Bone Morphogenetic Protein 4) signaling pathway, stimulating embryonic cell to differentiate into trophoblasts rather than maintaining pluripotency to differentiate into various cell types, thereby contributing to mole formation [21,47].
KHDC3L, though less frequently implicated in biparental complete hydatidiform mole (BICHM), likely forms complexes with NLRP7 to assist in oogenesis and early embryonic development [37]. Mutations in these genes can dysregulate inflammatory cytokine secretion within the endometrial cavity, potentially altering the implantation microenvironment and contributing to molar pregnancy pathogenesis [37]. Specifically, mutations in NLRP7 have been associated with lower levels of immune signaling molecules like IL-1β [56]. The altered immune function observed in patients with NLRP7 mutations may be relevant to understanding the pathogenesis of molar pregnancy. Reddy et al. identified 11 NLRP7 variants in homozygous or compound heterozygous state in individuals with a history of recurrent hydatidiform mole. Some of these variants were associated with abnormalities in transcription and post-transcriptional modifications of mRNA affecting splicing and resulting in either the absence of gene transcription or the formation of abnormally long or truncated proteins [57]. Reddy et al. NLRP7 variants along with other variants associated with RHM are presented in Table 1.
In addition to NLRP7 and KHDC3L, other gene variants have been identified in RHM cases, such as MEI1 (meiotic double-stranded break formation protein 1), TOP6BL/C11orf80 (type 2 DNA topoisomerase 6 subunit B-like), and REC114 (REC114 meiotic recombination), which affect oocyte meiosis and early embryonic development [59]. These mutations can result in abnormal spindle morphology, misaligned chromosomes, extrusion of all chromosomes into the polar body, and other defects that can lead to empty oocytes, androgenetic zygote formation, and subsequent molar pregnancy [59].
Understanding the genetic underpinning of diseases through comprehensive mutation analysis provides insights into the complex mechanisms involved and has the potential to offer avenues for personalized care in affected individuals. Similarly, the genetic and molecular basis of different molar pregnancies has a pivotal role in decisions about treatment protocols and follow-ups.
3.3. Molecular Dysregulations in GTD
Immunohistochemical studies on hydatidiform moles offer valuable insights into the molecular underpinnings that distinguish them from normal placentas and hydropic abortions. These investigations reveal significant alterations in markers crucial for cell cycle regulation, apoptosis, and trophoblast differentiation, shedding light on the pathogenesis of molar pregnancies. Table 2 summarizes the molecular dysregulations associated with Gestational Trophoblastic Disease.
The expression of p53 in cytotrophoblasts is markedly elevated in molar pregnancies compared to hydropic abortions [14,83,84]. Studies indicate higher levels of p53 expression in patients with invasive moles and choriocarcinoma compared to those with non-invasive hydatidiform moles [74,80,85]. Hadi et al. demonstrated that more than 55% positive staining for the TP53 gene can effectively distinguish non-invasive hydatidiform mole from invasive forms and choriocarcinoma with 100% sensitivity and 92.9% specificity, albeit based on a small sample size [85].
P53, recognized as a tumor suppressor gene and often referred to as the “guardian of the genome,” is normally expressed in cytotrophoblasts and infrequently in stromal cells [84]. Its primary functions include inducing cell cycle arrest and apoptosis [84,90]. This is achieved through the transcriptional activation of p21/WAF1, which interacts with cyclin E/Cdk2 and cyclin D/Cdk4 complexes, leading to G1 arrest in the cell cycle [90]. Therefore, p53 serves as a marker of proliferative activity, crucial for regulating proliferation by inducing either cell cycle arrest or apoptosis. The overexpression of p53 in molar tissues reflects the heightened proliferative capacity of trophoblastic cells [84]. Similarly, Studies have demonstrated significantly higher expression of p63, a tumor suppressor gene from p53 family, in molar pregnancies compared to hydropic abortions (p-value < 0.05) [70,87].
The ASPP (Ankyrin-repeat, SH3-domain, and proline-rich region containing protein) family, including ASPP1 and ASPP2, modulates p53 activity and is found to be dysregulated in gestational trophoblastic diseases (GTD) [91,92]. Normally functioning as tumor suppressors, ASPP1 and ASPP2 stimulate p53-mediated transcriptional modifications of p21. However, Mak et al. have reported their downregulation in GTD [91,92]. Another crucial member of this family is iASPP (inhibitory iASPP), which exhibits increased expression in complete hydatidiform mole and choriocarcinoma compared to normal placental tissue [76]. Chan et al. demonstrated that silencing iASPP was associated with reduced production of autophagy-related proteins (LC3) and increased susceptibility to oxidative stress in choriocarcinoma cells [76]. These findings underscore the role of imbalanced expression of ASPP1/2 (downregulation) and iASPP (upregulation) in the pathogenesis of GTD [76,91,92].
Several mutations in p53 genes have also been identified in patients with molar pregnancy. Chan et al. identified two missense mutations (p.R249S and p.R248Q) that disrupt p53 DNA binding sites, impairing its ability to control cell proliferation [62]. Additionally, a nonsense mutation (p.R213X) was reported to prematurely truncate the protein, resulting in loss of its normal function [62]. Several studies showed that Ki-67 exhibits heightened expression levels in hydatidiform moles compared to both hydropic abortions and normal placental tissues [70,78]. This protein plays a crucial role in trophoblast differentiation and is primarily expressed by cytotrophoblasts. Notably, studies have shown significant overexpression of Ki-67 in CHM when analyzed in conjunction with Cyclin E, a key promoter of cell cycle progression, demonstrating statistically significant difference compared to normal and hydropic placentas (p-value < 0.05) [93].
Apart from aberrant cell proliferation, dysregulation of apoptosis also plays a pivotal role in the pathogenesis of GTD. Studies examining the apoptotic index in molar pregnancies yield disparate findings, primarily due to methodological variations in apoptosis assessment. One study focused on caspase-3, a critical enzyme in caspase-dependent apoptosis, and reported diminished expression levels in GTN, suggesting reduced apoptosis [72]. In contrast, another study utilizing the TUNEL assay (terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick end labeling), which detects DNA fragmentation as a late-stage apoptosis marker, indicated heightened apoptotic activity in GTD [94]. These findings suggest that while apoptosis levels are elevated in GTD, it may predominantly occur through caspase-independent pathways rather than the traditional caspase-dependent pathway. This underscores the necessity for further research to elucidate the specific apoptotic mechanisms at play in molar pregnancies and their clinical implications.
BCL-2 (B-Cell Lymphoma-2), an anti-apoptotic gene, plays a crucial role in regulating caspase-dependent apoptosis and acts as an anti-proliferative protein by inhibiting cell transition from quiescence to S-phase [95]. In the placenta, BCL-2 expression in syncytiotrophoblasts controls apoptosis in these multinuclear cells. It prevents the spread of apoptotic changes and fragmentation to other nuclei sharing the same cytoplasm when apoptosis occurs in one nucleus of the syncytiotrophoblast [94]. Studies assessing BCL-2 expression in molar pregnancies have yielded conflicting results. Several studies have demonstrated significantly reduced BCL-2 immunohistochemical staining in syncytiotrophoblasts of complete mole compared to control specimens [66,67,68,71]. Lin et al. demonstrated that CHM progressing to gestational trophoblastic neoplasia expressed higher levels of miR-181b-5p and miR181d-5p and lower levels of their target, BCL-2, compared to those that regressed following evacuation [69]. Conversely, other studies have reported increased BCL-2 expression in molar pregnancies or found insignificant staining differences between molar and non-molar placentas [70,96,97]. Missaoui et al., in their evaluation of 220 specimens classified based on morphological appearance and molecular markers into 140 CHMs, 41 PHMs, and 39 HAs, found significantly higher BCL-2 immunostaining in partial mole (61%) and CHM (73.6%) compared to hydropic abortions (7.7%, p = 0.001, p < 0.0001 respectively), which may explain earlier findings of reduced caspase expression in GTD [80,90]. Larger studies incorporating reliable diagnostic criteria and ploidy analysis are warranted to accurately delineate the alterations in BCL-2 expression in molar pregnancy. In contrast, the immunoexpression of Bax, another regulator gene in the apoptosis pathway, did not show significant differences between molar pregnancies and non-molar pregnancies according to Reza et al. [68].
The molecular distinctions between different types of GTDs are crucial in identifying markers for the behavior and aggressiveness of molar pregnancies. Research has identified distinct patterns of growth factor receptor expression in various trophoblast types. Specifically, Epidermal Growth Factor Receptor (EGFR) and ERBB4 are notably overexpressed in actively proliferating trophoblasts [16]. Additionally, trophoblasts exhibiting invasive characteristics demonstrate heightened expression of ERBB2 (also known as HER2/neu or C-erbB-2) and ERBB3 [98,99]. Epidermal Growth Factor (EGF) and Heparin-Binding EGF-like Growth Factor (HB-EGF) enhance cell signaling pathways in EGFR-expressing trophoblasts, thereby promoting cell cycle progression and contributing to proliferative activity in hydatidiform moles [16].
C-erbB-2 was also found by Erol et al. to be significantly overexpressed in complete moles compared to partial moles and hydropic abortions (HA) [78]. Previous studies suggest that this overexpression may correlate with aggressive behavior in CHM [17,18]. Another tyrosine receptor kinase, CD117 (c-KIT), present on various cell types including mast cells, hematopoietic stem cells, and germ cells [100], exhibits increased activation in molar tissues compared to hydropic abortions and is associated with aggressive CHM behavior [71]. Activation of CD117 through stem cell factor (SCF) regulates processes such as proliferation, cell differentiation, apoptosis, and cell adhesion [100,101].
MicroRNAs (miRNAs) have been implicated in gestational trophoblastic diseases (GTDs) based on their distinct expression profiles in trophoblastic tissues. Among these, miR-371a-5p is notable for its oncogenic properties in various cancers and is found to be upregulated in progressive complete hydatidiform moles (CHMs), suggesting a potential role in the progression of malignancy [15]. Another microRNA, miR-196b-5p (miR-196b), functions as a tumor suppressor and exhibits reduced expression in CHM tissues compared to normal placentas [79]. This decrease correlates with elevated levels of MAP3K1, a protein promoting cell proliferation and differentiation [79]. Increasing miR-196b expression in hydatidiform mole cells has shown to diminish their proliferation and invasion, suggesting its potential as both a diagnostic marker and therapeutic target [79]. Furthermore, miR-21 is significantly overexpressed in tissues from hydatidiform moles, where it enhances aggressive behaviors such as proliferation, migration, and invasion of trophoblastic cells. This characteristic makes miR-21 a promising candidate for targeted approaches in the diagnosis and treatment of gestational trophoblastic neoplasms [81].
Twist-1, a negative regulator of E-cadherin, shows significantly elevated levels in CHM and correlates with disease invasiveness [75,88,89]. E-cadherin, crucial for cell-to-cell adhesion, exhibits reduced expression associated with GTD and is associated with increased invasiveness [14,73,75,88] although one study with a small sample failed to show a significant association [73].
4. Conclusions
In conclusion, the molecular landscape of molar pregnancies unveils a complex interplay of genetic and epigenetic factors that influence disease progression and clinical outcomes. The overexpression of p53 in cytotrophoblasts and dysregulation of its modulators, ASPP1/2 and iASPP, highlights the proliferative potential and invasive nature of molar tissues. Moreover, discrepancies in proteins involved in apoptotic pathways such as BCL-2 and caspases highlight the dysregulation of the balance between cell survival and death in GTDs. The differential expression of growth factor receptors such as EGFR, ERBB2, and CD117 in trophoblasts reflects their roles in promoting cell proliferation and invasion, contributing to the aggressive behavior observed in some molar pregnancies.
Despite these advancements, challenges remain in standardizing diagnostic criteria and therapeutic approaches for molar pregnancies. Future research should focus on elucidating the specific molecular pathways driving molar pregnancy pathogenesis and developing targeted interventions to improve clinical outcomes. It may be specifically helpful in preventing the recurrence of GTD and GTNs. By integrating molecular insights with clinical observations, we can advance our understanding of GTDs and enhance patient care through personalized medicine strategies.
Supplementary Materials
S1- Details of included studies. The supporting information can be downloaded at the website of this paper post on Preprints.org.
Author Contributions
Conceptualization- R.D., R.K., S.B.; methodology, R.D., S.B. and S.S.K.; software, S.B., M.G., and M.A.; validation, S.B., R.D., and R.S..; formal analysis, S.B; investigation, S.B., M.A., M.G., S.S.K., R.S., R.K., and R.D.; resources, S.B., R.D., R.K., M.A., M.G., and S.S.K.; data curation, R.D., and S.B.; writing— original draft preparation, S.B., and R.D.; writing—review and editing, R.D., R.K. and R.S.; visualization, S.B. and R.D.; supervision, R.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not Applicable
Informed Consent Statement
Not Applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lurain, J.R. Gestational trophoblastic disease I: epidemiology, pathology, clinical presentation and diagnosis of gestational trophoblastic disease, and management of hydatidiform mole. American journal of obstetrics and gynecology, 2010, 203(6), 531–539. [CrossRef]
- Bruce, S.; Sorosky, J. Gestational Trophoblastic Disease. StatPearls, Feb. 2024, Accessed: Jun. 26, 2024. [Online]. Available: https://www.ncbi.nlm.nih.gov/books/NBK470267/.
- Ning, F.; Hou, H.; Morse, A.N.; Lash, G.E. Understanding and management of gestational trophoblastic disease. F1000Research, 2019, 8, F1000 Faculty Rev–428. [Google Scholar] [CrossRef]
- Zhao, J.; Moss, J.; Sebire, N.J.; Cui, Q.C.; Seckl, M.J.; et al. Analysis of the chromosomal region 19q13.4 in two Chinese families with recurrent hydatidiform mole. Human Reproduction, 2006, 21(2), 536–541. [CrossRef]
- Kalogiannidis, I.; Kalinderi, K.; Kalinderis, M.; Miliaras, D.; Tarlatzis, B.; et al. Recurrent complete hydatidiform mole: where we are, is there a safe gestational horizon? Opinion and mini-review. Journal of assisted reproduction and genetics, 2018, 35(6), 967–973. [CrossRef]
- Capozzi, V.A.; Butera, D.; Armano, G.; Monfardini, L.; Gaiano, M.; et al. Obstetrics outcomes after complete and partial molar pregnancy: Review of the literature and meta-analysis. European journal of obstetrics, gynecology, and reproductive biology, 2021, 259, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Tantengco, O.A.G.; De Jesus, F.C. C. 2nd; Gampoy, E.F. S.; Ornos, E.D. B.; Vidal, M.S. Jr; et al. Molar pregnancy in the last 50 years: A bibliometric analysis of global research output. Placenta, 2021, 112, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Ngan, H.Y.S.; Seckl, M.J.; Berkowitz, R.S.; Xiang, Y.; Golfier, F.; et al. Diagnosis and management of gestational trophoblastic disease: 2021 update. International Journal of Gynecology & Obstetrics, 2021, 155 (S1), 86-93. [CrossRef]
- Albright, B.B.; Shorter, J.M.; Mastroyannis, S.A.; Ko, E.M.; Schreiber, C.A.; et al. Gestational Trophoblastic Neoplasia After Human Chorionic Gonadotropin Normalization Following Molar Pregnancy: A Systematic Review and Meta-analysis. Obstetrics and gynecology, 2020, 135(1), 12–23. [CrossRef]
- Sebire, N.J.; Fisher, R.A.; Rees, H.C. Histopathological diagnosis of partial and complete hydatidiform mole in the first trimester of pregnancy. Pediatric and developmental pathology: the official journal of the Society for Pediatric Pathology and the Paediatric Pathology Society, 2003, 6(1), 69–77. [CrossRef]
- Ronnett, B.M. Hydatidiform Moles: Ancillary Techniques to Refine Diagnosis. Archives of pathology & laboratory medicine, 2018, 142(12), 1485–1502. [CrossRef]
- Khawajkie, Y.; Mechtouf, N.; Nguyen, N.M.P.; Rahimi, K.; Breguet, M.; et al. Comprehensive analysis of 204 sporadic hydatidiform moles: revisiting risk factors and their correlations with the molar genotypes. Modern pathology: an official journal of the United States and Canadian Academy of Pathology, Inc, 2020, 33(5), 880–892. [CrossRef]
- Joyce, C.M.; Fitzgerald, B.; McCarthy, T.V.; Coulter, J.; O’Donoghue, K. Advances in the diagnosis and early management of gestational trophoblastic disease. BMJ Medicine, 2022, 1, e000321. [Google Scholar] [CrossRef] [PubMed]
- Erol, O.; Süren, D.; Tutuş; B; Toptaş; T; Gökay, A.A.; et al. Immunohistochemical Analysis of E-Cadherin, p53 and Inhibin-α Expression in Hydatidiform Mole and Hydropic Abortion. Pathology oncology research: POR, 2016, 22(3), 515–521. [CrossRef]
- Zhao, J.R.; Cheng, W.W.; Wang, Y.X.; Cai, M.; Bin Wu, W.; et al. Identification of microRNA signature in the progression of gestational trophoblastic disease. Cell Death & Disease, 2018, 9 (2), 94. [CrossRef]
- Zheng, X.Z.; Qin, X.Y.; Chen, S.W.; Wang, P.; Zhan, Y.; et al. Heterozygous/dispermic complete mole confers a significantly higher risk for post-molar gestational trophoblastic disease. Modern pathology: an official journal of the United States and Canadian Academy of Pathology, Inc, 2020, 33(10), 1979–1988. [CrossRef]
- Yang, X.; Zhang, Z.; Jia, C.; Li, J.; Yin, L.; et al. The relationship between expression of c-ras, c-erbB-2, nm23, and p53 gene products and development of trophoblastic tumor and their predictive significance for the malignant transformation of complete hydatidiform mole. Gynecologic oncology, 2002, 85(3), 438–444. [CrossRef]
- Yazaki-Sun, S.; Daher, S.; De Souza Ishigai, M.M.; Alves, M.T.S.; Mantovani, T.M.; et al. Correlation of c-erbB-2 oncogene and p53 tumor suppressor gene with malignant transformation of hydatidiform mole. The Journal of Obstetrics and Gynaecology Research, 2006, 32 (3). 265–272. [CrossRef]
- Kobayashi, E.H.; Shibata, S.; Oike, A.; Kobayashi, N.; Hamada, H.; et al. Genomic imprinting in human placentation. Reproductive Medicine and Biology, 2022, 21(1). [CrossRef]
- Atabaki Pasdar, F.; Khooei, A.; Fazel, A.; Rastin, M.; Tabasi, N.; et al. DNA flow cytometric analysis in variable types of hydropic placentas. Iranian journal of reproductive medicine 2015, 13, 269–274. [Google Scholar] [PubMed]
- Ndukwe, C.O.; Ukah, C.O. Epidemiological Aspects and Diagnostic Accuracy of Morphological Diagnosis of Hydatidiform Mole Using p57kip2 Immunostain in Nnewi, South-East Nigeria – A Multicenter Study. Journal of Nature and Science of Medicine 2021, 4, 281–287. [Google Scholar]
- Khashaba, M.; Arafa, M.; Elsalkh, E.; Hemida, R.; Kandil, W. Morphological Features and Immunohistochemical Expression of p57Kip2 in Early Molar Pregnancies and Their Relations to the Progression to Persistent Trophoblastic Disease. Journal of pathology and translational medicine, 2017, 51(4), 381–387. [CrossRef]
- Lelic, M.; Fatusic, Z.; Iljazovic, E.; Ramic, S.; Markovic, S.; et al. Challenges in the Routine Praxis Diagnosis of Hydatidiform Mole: a Tertiary Health Center Experience. Medical archives (Sarajevo, Bosnia and Herzegovina), 2017, 71(4), 256–260. [CrossRef]
- Sasaki, S.; Sasaki, Y.; Kunimura, T.; Sekizawa, A.; Kojima, Y.; et al. Clinical Usefulness of Immunohistochemical Staining of p57 kip2 for the Differential Diagnosis of Complete Mole. BioMed research international, 2015, 905648. [CrossRef]
- Xing, D.; Adams, E.; Huang, J.; Ronnett, B.M. Refined diagnosis of hydatidiform moles with p57 immunohistochemistry and molecular genotyping: updated analysis of a prospective series of 2217 cases. Modern pathology: an official journal of the United States and Canadian Academy of Pathology, Inc, 2021, 34(5), 961–982. [CrossRef]
- Zainal, N.; Kampan, N.C.; Rose, I.M.; Ghazali, R.; Shafiee, M.N.; et al. Complementary role of p57kip2 immunostaining in diagnosing hydatidiform mole subtypes. Hormone Molecular Biology and Clinical Investigation, 2021, 42 (3), 311-316. [CrossRef]
- Takahashi, S.; Okae, H.; Kobayashi, N.; Kitamura, A.; Kumada, K.; et al. Loss of p57KIP2 expression confers resistance to contact inhibition in human androgenetic trophoblast stem cells. Proceedings of the National Academy of Sciences, 2019, 116(52), 26606-26613. [CrossRef]
- Zheng, X.Z.; Hui, P.; Chang, B.; Gao, Z.B.; Li, Y.; et al. STR DNA genotyping of hydatidiform moles in South China. Int J Clin Exp Pathol. 2014, 7, 4704–4719. [Google Scholar] [PubMed]
- Banet, N.; DeScipio, C.; Murphy, K.M.; Beierl, K.; Adams, E.; et al. Characteristics of hydatidiform moles: analysis of a prospective series with p57 immunohistochemistry and molecular genotyping. Modern Pathology, 2014, 27 (2), 238–254. [CrossRef]
- King, J.R.; Wilson, M.L.; Hetey, S.; Kiraly, P.; Matsuo, K.; et al. Dysregulation of Placental Functions and Immune Pathways in Complete Hydatidiform Moles. Int. J. Mol. Sci. 2019, 20, 4999. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.F.; Chan, W.Y. The de novo DNA methyltransferase DNMT3A in development and cancer. Epigenetics, 2014, 9(5), 669–677. [CrossRef]
- Mahadevan, S; Wen, S.; Wan, Y.; Peng, H.; Otta, S.; et al. NLRP7 affects trophoblast lineage differentiation, binds to overexpressed YY1 and alters CpG methylation. Hum Mol Genet, 2014, 23 (3), 706–716. [CrossRef]
- Sanchez-Delgado, M.; Martin-Trujillo, A.; Tayama, C.; Vidal, E.; Esteller, M.; et al. Absence of Maternal Methylation in Biparental Hydatidiform Moles from Women with NLRP7 Maternal-Effect Mutations Reveals Widespread Placenta-Specific Imprinting. PLoS Genetics, 2015, 11(11), e1005644. [CrossRef]
- Bolze, P.A.; Patrier, S.; Cheynet, V.; Oriol, G.; Massardier, J.; et al. Expression patterns of ERVWE1/Syncytin-1 and other placentally expressed human endogenous retroviruses along the malignant transformation process of hydatidiform moles. Placenta, 2016, 39, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Langbein, M.; Strick, R.; Strissel, P.L.; Vogt, N.; Parsch, H.; et al. Impaired cytotrophoblast cell-cell fusion is associated with reduced Syncytin and increased apoptosis in patients with placental dysfunction. Molecular reproduction and development, 2008, 75(1), 175–183. [CrossRef]
- Lertkhachonsuk, R.; Paiwattananupant, K.; Tantbirojn, P.; Rattanatanyong, P.; Mutirangura, A. LINE-1 Methylation Patterns as a Predictor of Postmolar Gestational Trophoblastic Neoplasia, BioMed Research International, 2015, 421747 (7), 2015. [CrossRef]
- Rahat, B.; Thakur, S.; Bagga, R.; Kaur, J. Epigenetic regulation of STAT5A and its role as fetal DNA epigenetic marker during placental development and dysfunction. Placenta, 2016, 44, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Triratanachat, S.; Nakaporntham, P.; Tantbirojn, P.; Shuangshoti, S.; Lertkhachonsuk, R. Role of P57KIP2 Immunohistochemical Expression in Histological Diagnosis of Hydatidiform Moles. Asian Pac J Cancer Prev, 2016, 17 (4), 2061–2066, 2016. [CrossRef]
- Samadder, A.; Kar, R. Utility of p57 immunohistochemistry in differentiating between complete mole, partial mole & non-molar or hydropic abortus. Indian J Med Res, 2017, 145 (1), 133–137. [CrossRef]
- McConnell, T.G.; Murphy, K.M.; Hafez, M.; Vang, R.; Ronnett, B.M. Diagnosis and Subclassification of Hydatidiform Moles Using p57 Immunohistochemistry and Molecular Genotyping: Validation and Prospective Analysis in Routine and Consultation Practice Settings With Development of an Algorithmic Approach. The American Journal of Surgical Pathology, 2009, 33(6), 805-817. [CrossRef]
- LeGallo, R.D.; Stelow, E.B.; Ramirez, N.C.; Atkins, K.A. Diagnosis of hydatidiform moles using p57 immunohistochemistry and HER2 fluorescent in situ hybridization. Am J Clin Pathol, 2008, 129 (5), 749–755. [CrossRef]
- Diwa, M.H.; Kim, M.A.; Avila, J.M.C.; Pedroza, D.G.; Encinas-Latoy, M.A.M. Utility of p57KIP2 and Her-2 Fluorescence in Situ Hybridization in Differentiating Partial from Complete Hydatidiform Mole. Acta Med Philipp, 2016, 50(4), 318–325. [CrossRef]
- Frost, J.M.; Moore, G.E. The importance of imprinting in the human placenta. PLoS Genet, 2010, 6 (7), 1–9. [CrossRef]
- Tycko, B.; Morison, I.M. Physiological functions of imprinted genes. Journal of Cellular Physiology, 2002, 192 (3), 245–258. [CrossRef]
- Reddy, R.; Akoury, E.; Nguyen, N.M.P.; Abdul-Rahman, O.A.; Dery, C.; et al. Report of four new patients with protein-truncating mutations in C6orf221/KHDC3L and colocalization with NLRP7. Eur J Hum Genet, 2013, 21(9), 957–964. [CrossRef]
- Ji, M.; Shi, X.; Xiang, Y.; Cui, Q.; Zhao, J. NLRP7 and KHDC3L variants in Chinese patients with recurrent hydatidiform moles. Japan Journal of Clinical Oncology, 2019, 49(7), 620–627. [CrossRef]
- Nguyen, N.M.P.; Slim, R. Genetics and Epigenetics of Recurrent Hydatidiform Moles: Basic Science and Genetic Counselling. Curr Obstet Gynecol Rep, 2014, 3(1), 55. [CrossRef]
- Nguyen, N.M.P.; Khawajkie, Y.; Mechtouf, N.; Rezaei, M.; Breguet, M.; et al. The genetics of recurrent hydatidiform moles: new insights and lessons from a comprehensive analysis of 113 patients. Mod Pathol, 2018, 31(7), 1116–1130. [CrossRef]
- Fallahi, J.; Anvar, Z.; Razban, V.; Momtahan, M.; Namavar-Jahromi, B.; et al. Founder Effect of KHDC3L, p.M1V Mutation, on Iranian Patients with Recurrent Hydatidiform Moles,” Iran J Med Sci, 2020, 45(2), 118. [CrossRef]
- Fallahi, J.; Alashti, S.K.; Aliabadi, B.E.; Mohammadi, S.; Fardaei, M. Recurrent pregnancy loss in the female with a heterozygous mutation in KHDC3L gene. Gene Rep, 2020, 20, 100721. [Google Scholar] [CrossRef]
- Rezaei, M.; Nguyen, N.M.P.; Foroughinia, L.; Dash, P.; Ahmadpour, F.; et al. Two novel mutations in the KHDC3L gene in Asian patients with recurrent hydatidiform mole. Human Genome Variation, 2016, 3(1), 1–5. [CrossRef]
- Shalabi, T.A.; Abdel-Hamid, M.S.; Shaker, M.M. Two Novel Variants in NLRP7 Gene in an Egyptian Female Patient with Consecutive Molar Pregnancies Complicated by Choriocarcinoma. International Journal of Infertility and Fetal Medicine, 2019, 10(3), 54-57. [CrossRef]
- Fallahi, J.; Razban, V.; Momtahan, M.; Akbarzadeh-Jahromi, M.; Namavar-Jahromi, B.; et al. A novel mutation in NLRP7 related to recurrent hydatidiform mole and reproductive failure. Int J Fertil Steril, 2019, 13(2), 135-138. [CrossRef]
- Messaed, C.; Akoury, E.; Djuric, U.; Zeng, J.; Saleh, M.; et al. NLRP7, a nucleotide oligomerization domain-like receptor protein, is required for normal cytokine secretion and co-localizes with Golgi and the microtubule-organizing center. J Biol Chem, 2011, 286(50), 43313–43323. [CrossRef]
- Sills, E.S.; Obregon-Tito, A.J.; Gao, H.; McWilliams, T.K.; Gordon, A.T.; et al. Pathogenic variant in NLRP7 (19q13.42) associated with recurrent gestational trophoblastic disease: Data from early embryo development observed during in vitro fertilization.Clinical and experimental reproductive medicine,2017,44(1),40–46. [CrossRef]
- Zhang, P.; Zhu, X.; Yu, X.; Huang, B.; Jiang, T.; et al. Abnormal processing of IL-1β in NLRP7-mutated monocytes in hydatidiform mole patients. Clinical and Experimental Immunology, 2020, 202(1), 72–79. [CrossRef]
- Reddy, R.; Nguyen, N.M.P.; Sarrabay, G.; Rezaei, M.; Rivas, M.C.G.; et al. The genomic architecture of NLRP7 is Alu rich and predisposes to disease-associated large deletions. European Journal of Human Genetics, 2016, 24(10), 1445–1452. [CrossRef]
- Rezaei, M.; Suresh, B.; Bereke, E.; Hadipour, Z.; Aguinaga, M.; et al. Novel pathogenic variants in NLRP7, NLRP5, and PADI6 in patients with recurrent hydatidiform moles and reproductive failure. Clin Genet, 2021, 99(6), 823–828. [CrossRef]
- Buza, N.; McGregor, S.M.; Barroilhet, L.; Zheng, X.; Hui, P. Paternal uniparental isodisomy of tyrosine hydroxylase locus at chromosome 11p15.4: spectrum of phenotypical presentations simulating hydatidiform moles. Modern pathology: an official journal of the United States and Canadian Academy of Pathology, Inc, 2019, 32(8), 1180–1188. [CrossRef]
- Yu, Y.; Lu, B.; Lu, W.; Li, S.; Li, X.; et al. Whole-exome sequencing reveals genetic variants in ERC1 and KCNG4 associated with complete hydatidiform mole in Chinese Han women.Oncotarget. 2017; 8(3): 75264-75271. https://www.oncotarget.com/article/20769/text/.
- Chan, K.K.; Wong, E.S.Y.; Wong, O.G. W.; Ngan, H.Y. S.; Cheung, A.N. Y. Identification of nonsynonymous TP53 mutations in hydatidiform moles. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis, 2018, 809, 20–23. [Google Scholar] [CrossRef]
- Hemida, R.; van Doorn, H.; Fisher, R. A Novel Genetic Mutation in a Patient With Recurrent Biparental Complete Hydatidiform Mole: A Brief Report. International journal of gynecological cancer: official journal of the International Gynecological Cancer Society, 2016, 26(7), 1351–1353. [CrossRef]
- Ito, Y.; Maehara, K.; Kaneki, E.; Matsuoka, K.; Sugahara, N.; et al. Novel Nonsense Mutation in the NLRP7 Gene Associated with Recurrent Hydatidiform Mole. Gynecol Obstet Invest, 2016, 81(4), 353–358. [CrossRef]
- Nguyen, N.M.P.; Ge, Z.J.; Reddy, R.; Fahiminiya, S.; Sauthier, P.; et al. Causative Mutations and Mechanism of Androgenetic Hydatidiform Moles. Am J Hum Genet, 2018, 103(5), 740–751. [CrossRef]
- Dube, R.; Kar, S.S.; Jhancy, M.; George, B.T. Molecular Basis of Müllerian Agenesis Causing Congenital Uterine Factor Infertility—A Systematic Review. Int. J. Mol. Sci. 2024, 25, 120. [Google Scholar] [CrossRef]
- Al-Jabri, M.; Al-Badi, S.; Al-Kindi, H.; Arafa, M. Immunohistochemical expression of BCL-2 in hydatidiform moles: a tissue microarray study. Pathologica - Journal of the Italian Society of Anatomic Pathology and Diagnostic Cytopathology, 2023, 115(3), 148–154. [CrossRef]
- Wargasetia, T.L.; Shahib, M.N.; Martaadisoebrata, D.; Dhianawaty, D.; Hernowo, B. Characterization of apoptosis and autophagy through Bcl-2 and Beclin-1 immunoexpression in gestational trophoblastic disease. Iran J Reprod Med 2015, 13, 413. [Google Scholar]
- Khooei, A.; Pasdar, F.A.; Fazel, A.; Mahmoudi, M.; Reza, N.M.; et al. View of Expression of Pro-Apoptotic Bax and Anti-Apoptotic Bcl-2 Proteins in Hydatidiform Moles and Placentas With Hydropic Changes. Acta Medica Iranica, 2019,57(1), 27-32. [CrossRef]
- Lin, L.H.; Maestá, I.; St Laurent, J.D.; Hasselblatt, K.T.; Horowitz, N.S.; et al. Distinct microRNA profiles for complete hydatidiform moles at risk of malignant progression. American journal of obstetrics and gynecology, 2021, 224(4), 372.e1–372.e30. [CrossRef]
- Missaoui, N.; Landolsi, H.; Mestiri, S.; Essakly, A.; Abdessayed, N.; et al. Immunohistochemical analysis of c-erbB-2, Bcl-2, p53, p21WAF1/Cip1, p63 and Ki-67 expression in hydatidiform moles. Pathology, research and practice, 2019, 215(3), 446–452. [CrossRef]
- Erol, O.; Suren, D.; Tutus, B.; Yararbas, K.; Sayiner, A.; et al. Comparison of p57, c-erbB-2, CD117, and Bcl-2 expression in the differential diagnosis of hydatidiform mole and hydropic abortion. European journal of gynaecological oncology 2016, 37, 522–529. [Google Scholar] [PubMed]
- Braga, A.; Maestá, I.; Rocha Soares, R.; Elias, K.M.; Custódio Domingues, M.A.; et al. Apoptotic index for prediction of postmolar gestational trophoblastic neoplasia. American journal of obstetrics and gynecology, 2016, 215(3), 336.e1–336.e12. [CrossRef]
- Wang, J.; Zhao, M.; Xiao, J.; Wu, M.; Song, Y.; et al. E-Cadherin, CD44v6, and Insulin-Like Growth Factor-II mRNA-Binding Protein 3 Expressions in Different Stages of Hydatidiform Moles. Journal of biochemical and molecular toxicology, 2016, 30(9), 455–461. [CrossRef]
- Hasanzadeh, M.; Sharifi, N.; Farazestanian, M.; Nazemian, S.S.; Sani, F.M. Immunohistochemistry Study of P53 and C-erbB-2 Expression in Trophoblastic Tissue and Their Predictive Values in Diagnosing Malignant Progression of Simple Molar Pregnancy. Iran J Cancer Prev, 2016, 9(3), 1–7. [CrossRef]
- Moussa, R.A.; Eesa, A.N.; Abdallah, Z.F.; Abdelmeged, A.; Mahran, A.; et al. Diagnostic Utility of Twist1, Ki-67, and E-Cadherin in Diagnosing Molar Gestations and Hydropic Abortions. Am J Clin Pathol, 2018, 149(5), 442–455. [CrossRef]
- Chan, K.K.; Wong, E.S.; Wong, I.T.; Cheung, C.L.; Gee-Wan, O.; et al. Overexpression of iASPP is required for autophagy in response to oxidative stress in choriocarcinoma. BMC Cancer, 2019, 19(1), 1–13. [CrossRef]
- Yucel Cicek, O.S.; Hekimoglu, E.R.; Turgal, M.; Atilla, P.; Cakar, A.N.; et al. Differential expression of leukemia inhibitory factor and insulin like growth factor-1 between normal pregnancies, partial hydatidiform moles and complete hydatidiform moles. Placenta, 2018, 69, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Deka FF, H.A.; ZH, A.A. A. S.; Z, K.A. Role of the Immunohistochemical Marker (Ki67) in Diagnosis and Classification of Hydatidiform Mole. IIUM Medical Journal Malaysia, 2019, 18 (3). [CrossRef]
- Guo, Z.; Sui, L.; Qi, J.; Sun, Q.; Xu, Y.; et al. miR-196b inhibits cell migration and invasion through targeting MAP3K1 in hydatidiform mole. Biomedicine & pharmacotherapy, 2019, 113, 108760. [Google Scholar] [CrossRef]
- Sun, P.; Wu, Q.; Ruan, G.; Zheng, X.; Song, Y.; et al. Expression patterns of maspin and mutant p53 are associated with the development of gestational trophoblastic neoplasia. Oncology Letters, 2016, 12(5), 3135-3142. [CrossRef]
- Wang, Y.X.; Zhao, J.R.; Xu, Y.Y.; Wu, W.B.; Zhang, H.J. miR-21 Is Overexpressed in Hydatidiform Mole Tissues and Promotes Proliferation, Migration, and Invasion in Choriocarcinoma Cells. Int J Gynecol Cancer, 2017, 27(2), 364–374. [CrossRef]
- Hoeijmakers, Y.M.; Gorris, M.A.J.; Sweep, F.C. G. J.; Bulten, J.; Eysbouts, Y.K.; et al. Immune cell composition in the endometrium of patients with a complete molar pregnancy: Effects on outcome. Gynecologic oncology, 2021, 160(2), 450–456. [CrossRef]
- Kheradmand, P.; Goudarzi, M.; Tavakoli, M. Analysis of p53 expression in partial hydatidiform mole and hydropic abortion. Front Biol (Beijing), 12(5), 357–360. [CrossRef]
- Khooei, A.; Pasdar, F.A.; Fazel, A.; Mahmoudi, M.; Nikravesh, M.R.; et al. P53 expression in various types of hydropic placentas (through ploidy analysis as a complementary tool in diagnosis of samples). Caspian J Intern Med, 2019, 10(2), 205. [CrossRef]
- Hadi, F.; Kazemi, N.; Hosseini, M.S.; Ebrahimi, A. Evaluation of TP53 and HER-2/neu Genes Expression Levels in Gestational Trophoblastic Diseases Cases and Determining Their Predictive Value in Diagnosis of Malignancy and Disease Progression. International Journal of Cancer Management, 2022, 15(8), 119264. [CrossRef]
- Kubelka-Sabit, K.; Prodanova, I.; Jasar, D.; Bozinovski, G.; Filipovski, V.; et al. Molecular and immunohistochemical characteristics of complete hydatidiform moles. Balkan Journal of Medical Genetics, 2017, 20(1) 27-34. [CrossRef]
- Masood, S.; Kehar, S.I.; Shawana, S.; Aamir, I. Differential expression of p63 in hydropic and molar gestations. Journal of the College of Physicians and Surgeons Pakistan 2015, 25, 198–202. [Google Scholar] [PubMed]
- Jahanbin, B.; Sarmadi, S.; Ghasemi, D.; Nili, F.; Moradi, J.A.; et al. Pathogenic role of Twist-1 protein in hydatidiform molar pregnancies and investigation of its potential diagnostic utility in complete moles. Diagn Pathol, 2023, 18(1), 1–8. [CrossRef]
- Luchini, C.; Parcesepe, P.; Mafficini, A.; Nottegar, A.; Parolini, C.; et al. Specific expression patterns of epithelial to mesenchymal transition factors in gestational molar disease. Placenta, 2015, 36(11), 1318–1324. [CrossRef]
- Chen, J. The Cell-Cycle Arrest and Apoptotic Functions of p53 in Tumor Initiation and Progression. Cold Spring Harbor perspectives in medicine, 2016, 6(3), a026104. [CrossRef]
- Mak, V.C.Y.; Lee, L.; Siu, M.K.Y.; Wong, O.G.W.; Lu, X.; et al. Downregulation of ASPP2 in choriocarcinoma contributes to increased migratory potential through Src signaling pathway activation. Carcinogenesis, 2013, 34(9), 2170–2177. [CrossRef]
- Mak, V.C.Y.; Lee, L.; Siu, M.K.Y.; Wong, O.G.W.; Lu, X.; et al. Downregulation of ASPP1 in gestational trophoblastic disease: correlation with hypermethylation, apoptotic activity and clinical outcome. Mod Pathol, 2011, 24(4), 522–532. [CrossRef]
- Kar, A.; Mishra, C.; Biswal, P.; Kar, T.; Panda, S.; et al. Differential expression of cyclin E, p63, and Ki-67 in gestational trophoblastic disease and its role in diagnosis and management: A prospective case-control study. Indian J Pathol Microbiol, 2019, 62(1), 54–60. [CrossRef]
- Toki, T.; Horiuchi, A.; Ichikawa, N.; Mori, A.; Nikaido, T.; et al. Inverse relationship between apoptosis and Bcl-2 expression in syncytiotrophoblast and fibrin-type fibrinoid in early gestation. Mol Hum Reprod, 1999, 5(3), 246–251. [CrossRef]
- Ronnett, B.M.; Descipio, C.; Murphy, K.M. Hydatidiform moles: Ancillary techniques to refine diagnosis. International Journal of Gynecological Pathology, 2011, 30(2), 101–116. [CrossRef]
- Fulop, V.; Mok, S.C.; Genest, D.R.; Szigetvari, I.; Cseh, I.; et al. c-myc, c-erbB-2, c-fms and bcl-2 oncoproteins. Expression in normal placenta, partial and complete mole, and choriocarcinoma. J Reprod Med. 1998, 43, 101–110. [Google Scholar] [PubMed]
- Al-Bozom, I.A. P53 and Bcl-2 oncoprotein expression in placentas with hydropic changes and partial and complete moles. APMIS, 2000, 108(11), 756-760. [CrossRef]
- Pinkas-Kramarski, R.; Alroy, I.; Yarden, Y. ErbB receptors and EGF-like ligands: Cell lineage determination and oncogenesis through combinatorial signaling. J Mammary Gland Biol Neoplasia, 1997, 2(2), 97–107. [CrossRef]
- Fock, V.; Plessl, K.; Fuchs, R.; Dekan, S.; Milla, S.K.; et al. , Trophoblast subtype-specific EGFR/ERBB4 expression correlates with cell cycle progression and hyperplasia in complete hydatidiform moles. Human Reproduction, 2015, 30(4), 789–799. [CrossRef]
- Miettinen, M.; Lasota, J. KIT (CD117): a review on expression in normal and neoplastic tissues, and mutations and their clinicopathologic correlation. Appl Immunohistochem Mol Morphol, 2005, 13(3), 205–220. [CrossRef]
- Sheikh, E.; Tran, T.; Vranic, S.; Levy, A.; Bonfil, R.D. Role and significance of c-KIT receptor tyrosine kinase in cancer: A review. Bosn J Basic Med Sci, 2022, 22(5), 683. [CrossRef]
Figure 1.
Classification of Gestational Trophoblastic Diseases (GTD).
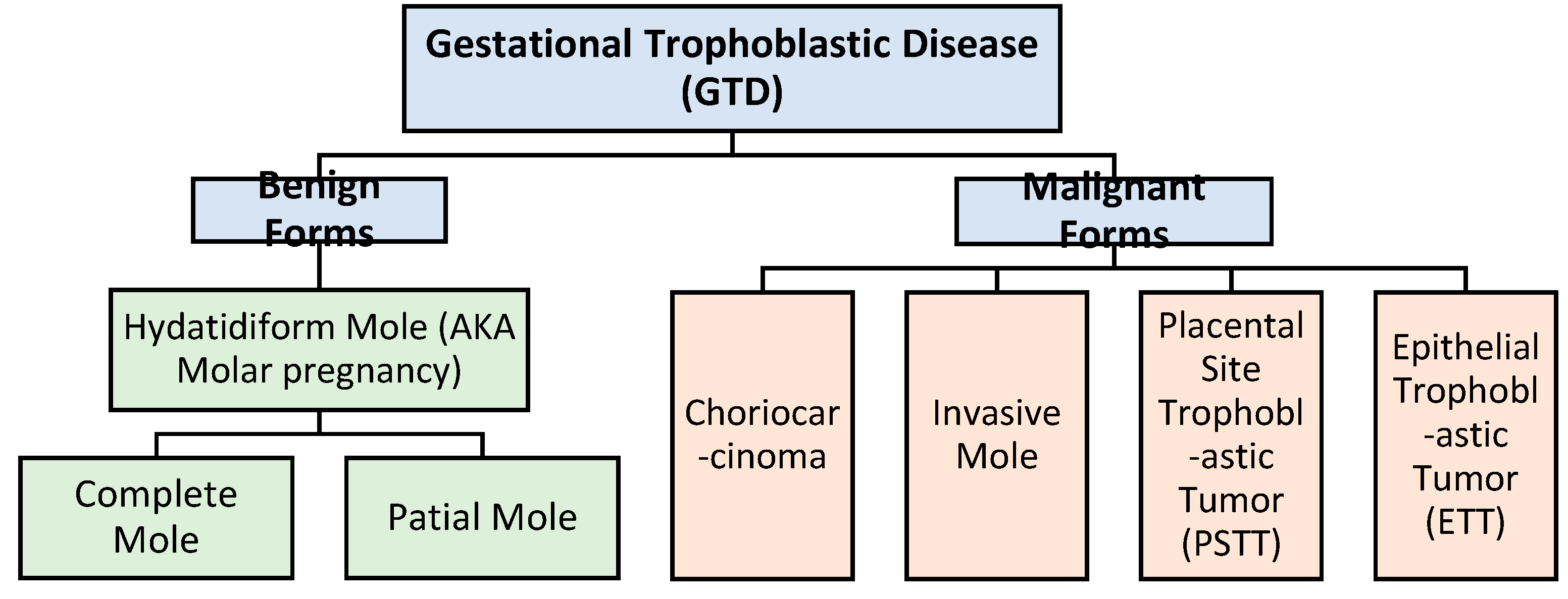
Figure 2.
PRISMA flow diagram for study inclusions.
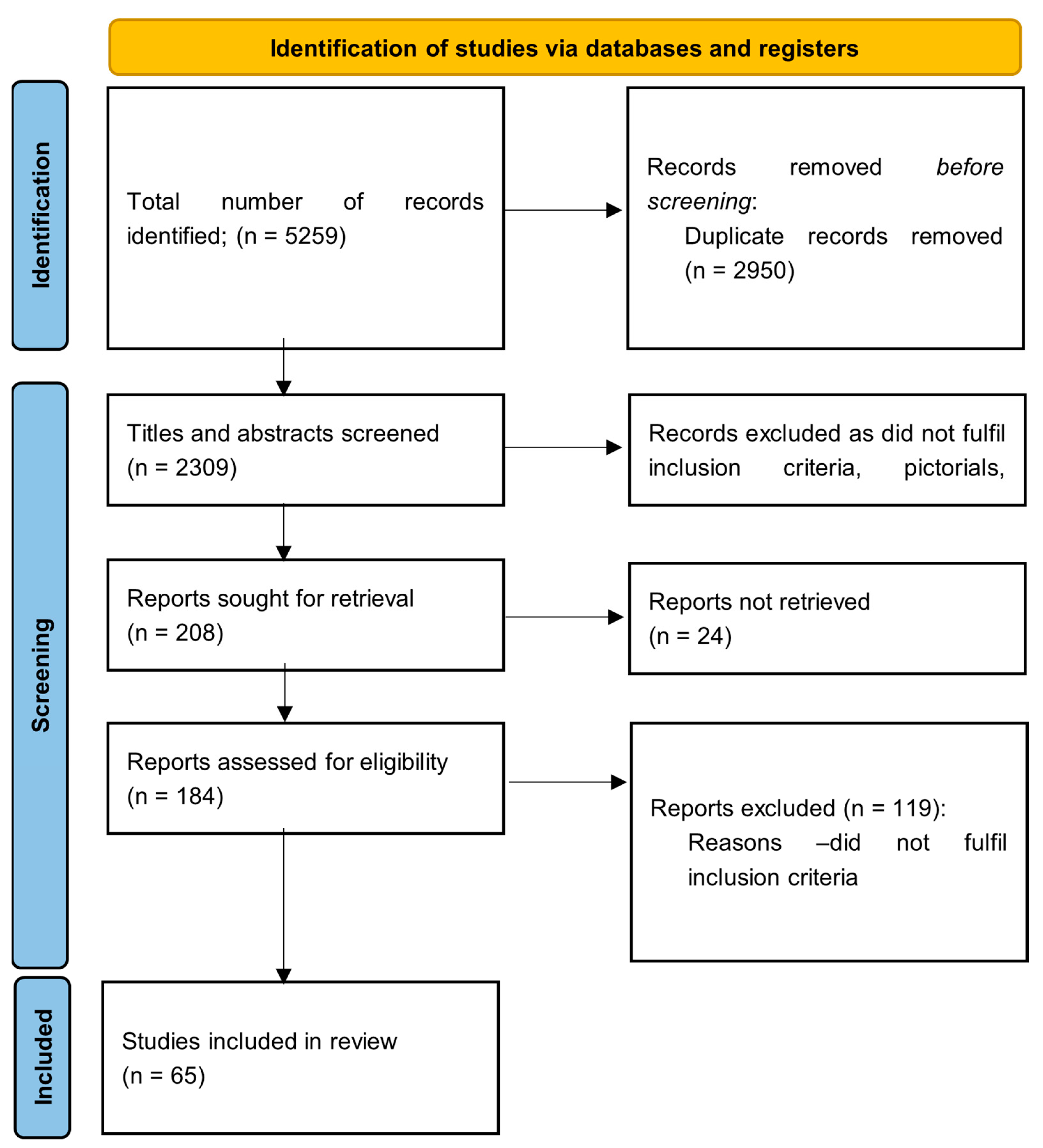

Table 1.
Genetic Variants and Mutations Associated with Recurrent Hydatidiform Mole (RHM).
| Chromosome Number | Mutation [Reference] |
Gene Involved |
| 1 | c.[1796T > A]; [1796T > A] [58] |
PADI6 |
| 6 | 6p21.33 Mutation p.M1V, c.1A>G [50] |
KHDC3L |
| 6 | c.322_325delGACT [6,15,16,17] |
KHDC3L |
| 6 | c.602 C>G [46] |
KHDC3L |
| 6 | c.299_302delTCAA, p.Ile100Argfs*2 c.322_325delGACT, p.Asp108Ilefs*30 [45] |
KHDC3L |
| 6 | c.17_20delGGTT, p.Arg6Leufs*7 c.349+1G4A [51] |
KHDC3L |
| 6 | c.334 1G>A [59] |
REC114 |
| 11 | pUPD 11p15.4 [60] |
TH01 |
| 11 | c.783dup (p.Glu262*) missense variant c.1501T>C (p.Ser501Pro) [59] |
TOP6BL/C11orf80 |
| 13 | G48C(p.Q16H) [61] |
ERC1c |
| 16 | c.G1114A(p.G372S) [61] |
KCNG4 |
| 17 | Exon6-213(nonsense) p.Arg213X Exon6-220(missense) p.Tyr220Cys Exon7-245(missense) p.Gly245Ser Exon7-248(missense) p.Arg248Gln Exon7-249(missense) p.Arg249Ser Exon8-295(missense) p.Pro295Leu [62] |
TP53 |
| 19 | c.1441 G>A [46] |
NLRP7 |
| 19 | Exon 4 (missense) c.1358T>G, p.Ile453Ser Exon 7 (Frameshift) c.2655dupC, p.Ile886HisfsTer11 [52] |
NLRP7 |
| 19 | c.555_557delCAC, p.Thr185del [53] |
NLRP7 |
| 19 | c.2810+2T>G [55] |
NLRP7 |
| 19 | Exon 2, c.197G>A [63] |
NLRP7 |
| 19 | c.584G>A; p.W195X [64] |
NLRP7 |
| 19 | Exon 6/intron 9 (c.[2248C4G]; [2810+2T4G]) Exon 4 (c.[1374_1375delAG]; [1374_1375delAG]) Exon 4/6 (c.[1908dup]; [2161C4T]) Exon4/intron 10 (c.[939_952dup14]; [2982-2A4G]) Exon 9 (c.[2759G4A]; [2759G4A]) Exon 9 (c.[2777T4G]; [2777T4G]) Intron 5 (c.[2130-6_2132del]; [ = ]) Intron 5 & 6 (c.[2130-266_2300+782del]; [2130-266_2300+782del]) Before exon 1 & intron 5 (c.[ −39-231_2130-510del]; [-39-231_2130-510del]) Intron 1 & intron 5/exon 8 (c.[-40+251_2130-681del];[2571dup]) Intron 1 & intron 5 (c.[-3998_2130-668del]; [-3998_2130-668del]) Before exon 1/exon 6 (c.[-13413_2982-344del];[2248C4G]) [57] |
NLRP7 |
| 19 | c.[1812_1837dup]; [1812_1837dup] c.[2162G > A]; [2162G > A] c.[2204A > C]; [2204A > C] c.[−40 + 3G > C]; [−40 + 3G > C] c.[−6831_-39–1586]; [2248C > G] [58] |
NLRP7 |
| 19 |
c.1093G > A, p.(Asp365Asn) c.[1093G > A]; [1093G > A] [58] |
NLRP5 |
| 22 | c.3452G>A c.1196þ1G>A, affecting the splice donor of exon 10, and a 1-bp deletion, c.2206del (p.Val736Serfs*31), in exon 19 [59] |
MEI1 |
Table 2.
Molecular dysregulation in GTD.
| Gene | Reference | Detection method | GTD Diagnosis method | Number of cases | Result |
| BCL-2 | [66] | TMA | Morphological appearance and p57 IHC | Control 202 PHM 95 CHM 142 |
Decreased expression in CHM compared to PHM and control |
| BCL-2 | [67] | IHC | Not specified | Control 6 PHM 11 CHM 11 IM 11 CC 9 |
Decreasing expression from PHM, to CHM, to invasive mole, to choriocarcinoma compared to normal placenta |
| BCL-2 | [68] | IHC | Morphological appearance and ploidy analysis by flow cytometry | HA 10 PHM 8 CHM 11 |
Decreased expression in CHM compared to PHM and HA |
| BCL-2 | [69] | IHC | Morphological appearance | Regressed HM 30 Post-molar GTN 9 |
Decreased expression is associated with progression from CHM to GTN |
| BCL-2 | [70] |
IHC | Morphological appearance and p57 IHC nuclear DNA micro- satellite polymorphism for discordant cases |
HA 39 PHM 41 CHM 140 |
Increased expression in CHM and PHM compared to HA |
| BCL-2 | [71] | IHC | Morphological appearance and STR genotyping | HA 17 PHM 23 CHM 20 |
Increased expression in HA compared to CHM and PHM |
| Beclin-1 | [67] | IHC | Not specified | Control 6 PHM 11 CHM 11 IM 11 CC 9 |
Increased expression in choriocarcinoma |
| Capase-3 | [72] | IHC | Morphological appearance and p57 IHC | Regressed CHM 590 Post-CHM GTN 190 |
The NPV for GTN of apoptotic index ≥ 4.0% was 97% |
| CD117 | [71] | IHC | Morphological appearance and STR genotyoing | HA 17 PHM 23 CHM 20 |
Decreased staining percentage in HA compared to CHM and PHM |
| CD44v6 | [73] | IHC | Not specified | Control 36 PHM 25 CHM 48 IM 12 |
No significant difference in CD44v6 expression |
| c-erB-2 | [71] | IHC | Morphological appearance and STR genotyoing | HA 17 PHM 23 CHM 20 |
Increased expression in CHM compared to PHM and HA |
| c-erB-2 | [74] |
IHC | Morphological appearance and immunohistochemistry and serial beta HCG | PHM 10 CHM 18 GTN 30 |
Increased expression in GTN compared to simple molar pregnancy. |
| E-cadherin | [14] | IHC | Morphological appearance and molecular genotyping | HA 23 PHM 24 CHM 23 |
Decreasing expression from HA to PHM to CHM |
| E-cadherin | [75] | IHC | Morphological appearance, ploidy analysis by flow cytometry, and p57 IHC | HA 16 PHM 17 CHM 16 |
Decreased expression in HM compared to HA |
| E-cadherin | [73] | IHC | Not specified | Control 36 PHM 25 CHM 48 IM 12 |
No significant difference in E-cadherin expression |
| iASPP | [76] | IHC siRNA transfection |
Not specified | First trimester placenta 10 Term placenta 11 HM 63 CC 7 |
Over-expression might be related to development of GTN Overexpression is associated with increased autophagy related protein expression Silencing is associated with cellular senescence |
| IGF-1 | [77] | IHC | Morphological appearance and p57 IHC | Control 8 PHM 8 CHM 8 |
Downregulation in CHM decidua and chorionic villi |
| IMP3 | [73] | IHC | Not specified | Control 36 PHM 25 CHM 48 IM 12 |
Decreasing expression from normal placental tissues, to PHM, to CHM, to IM |
| Inhibin-alpha | [14] | IHC | Morphological appearance and molecular genotyping | HA 23 PHM 24 CHM 23 |
Increased expression in molar pregnancy compared with HA |
| Ki-67 | [70] |
IHC | Morphological appearance and p57 IHC | HA 39 PHM 41 CHM 140 |
Increased expression in CHM compared to PHM and HA |
| Ki-67 | [78] | IHC | Morphological appearance | HA 30 PHM 30 CHM 30 |
Expression in cytotrophoblasts is significantly higher in CHM than PHM than HA Expression in stromal cells is significantly higher in molar pregnancy than HA |
| Ki-67 | [75] | IHC | Morphological appearance, ploidy analysis by flow cytometry, and p57 IHC | HA 16 PHM 17 CHM 16 |
Significantly increased expression in PHM compared to HA |
| LIF | [77] | IHC | Morphological appearance and p57 immunostaining | Control 8 PHM 8 CHM 8 |
Downregulated in CHM decidua but upregulated in CHM trophoblasts. |
| MAP3K1 | [79] | IHC | Not specified | Controls 15 CHM 20 |
Significantly higher expression in CHM compared to control |
| Maspin m-p53 |
[80] | IHC | Not specified | Control 48 Regressed HM 49 Progressed HM 39 |
Maspin was inversely correlated with FIGO prognostic score whereas expression of m-p53 was positively correlated with FIGO stage. |
| miR-181 | [69] |
miRNA-sequencing and qRT-PCR and western blot | Morphological appearance | Regressed HM 30 Post-molar GTN 9 |
Increased miR-181 expression in post-molar GTN. |
| miR-371a-5p and miR-518a-3p | [15] | miRNA array and qRT-PCR | Morphological appearance | Control 6 Regressed CHM 35 Post-CHM GTN 21 |
miR-371a-5p and miR-518a-3p were upregulated in progressed CHMs (GTN) |
| miRNA-21 | [81] | qRT-PCR | Morphological appearance | Control 20 HM 16 |
miRNA-21 is upregulated in HM |
| miR-196b | [79] | qRT-PCR | Not specified | Control 15 CHM 20 |
Significantly lower expression in CHM compared to control. |
| NKT-like cells | [82] | mIHC | The FIGO 2000 guideline | 16 CHM with spontaneous regression 16 CHM with progress to post-molar GTN |
The density of NKT-like cells was significantly higher in patients with spontaneous regression compared to those who progressed to GTN |
| P53 | [70] |
IHC | Morphological appearance and p57 immunostaining | HA 39 PHM 41 CHM 140 |
Increased expression in CHM compared to PHM and HA |
| P53 | [83] | IHC | Morphological appearance | HA 20 PHM 20 |
Significantly higher positive rate and grade of staining in PHM compared to HA. |
| P53 | [14] | IHC | Morphological appearance and molecular genotyping | HA 23 PHM 24 CHM 23 |
Increased expression of p53 in CHM compared to PHM and HA |
| P53 | [74] |
IHC | Morphological appearance and immunohistochemistry and serial beta HCG | PHM 10 CHM 18 GTN 30 |
Increased expression of p53 in GTN compared to simple HM. |
| P53 | [84] |
IHC | Morphological appearance and ploidy analysis by flow cytometry | HA 10 PHM 8 CHM 11 |
Increased expression in HM compared to HA |
| P53 | [85] | IHC | Not specified | PHM 32 CHM 24 4 IM 2 CC |
Expression was significantly associated with tendency to invasion and metastatic behaviours |
| P57 | [86] | IHC | Not specified | CHM 8 | Absent expression in all CHM (both androgenetic diploidy and biparental diploidy) |
| P57 | [71] | IHC | Morphological appearance and STR genotyoing | HA 17 PHM 23 CHM 20 |
Absent expression in CHM No significant difference between PHM and HA |
| P63 | [70] | IHC | Morphological appearance, p57 immunostaining, and nuclear DNA micro- satellite polymorphism for discordant cases |
HA 39 PHM 41 CHM 140 |
Increased expression in CHM and PHM compared to HA |
| P63 | [87] | IHC | Morphological appearance | HA 30 PHM 30 CHM 30 |
Increased intensity of staining in HM compared to HA |
| Syncyntin-1 |
[34] | IHC | Not specified | Control 8 PHM 6 CHM 12 IM 1 CC 1 PSTT 1 |
the staining intensity of the surface subunit C-terminus was significantly higher in HM, especially those with malignant transformation on follow up |
| Twist-1 | [88] | IHC | Morphological appearance and p57 immunostaining | CHM 47 PHM 40 |
Expression is significantly higher villous stromal cells of CHM compared to PHM |
| Twist-1 | [89] | IHC | Morphological appearance and p57 immunostaining and ploidy analysis | Abortion 23 PHM 10 CHM 12 Term placenta 7 |
Expression is significantly higher in CHM compared to PHM and HA |
| Twist-1 | [75] | IHC | Morphological appearance, ploidy analysis by flow cytometry, and p57 immunostaining | CHM 16 PHM 17 HA 16 |
Expression is significantly higher in CHM compared to PHM and HA |
BCL-2= B cell lymphoma-2, TMA = Tissue Microarray, PMH = Partial hydatidiform mole, CHM = Complete hydatidiform mole, IHC = Immunohistochemistry, IM = Invasive mole, CC = Choriocarcinoma, HA = Hydropic abortion, HM = Hydatidiform mole, GTN = Gestational trophoblastic neoplasia, STR = Short tandem repeat, NPV = Negative predictive value, Apoptotic index = Positive caspase -3 staining cells / negative caspase-3 staining cells x 100, FIGO = International Federation of Gynecology and Obstetrics, miRNA = Micro-Ribonucleic acid, qRT-PCR = quantitative Reverse Transcription Polymerase Chain Reaction, quantitative real-time polymerase chain reaction, mIHC = multiplex Immunohistochemistry, PSTT = Placental site trophoblastic tumor.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



