
Submitted:
11 September 2024
Posted:
12 September 2024
You are already at the latest version
Abstract
People with aphantasia exhibit the inability to voluntarily generate or form mental imagery in their minds. Since the term “aphantasia” was proposed by Zeman et al. in 2015 to describe this, it has gained increasing attention from psychiatrists, neuroscientists, and clinicians. Previous studies have mainly focused on definition, prevalence, and measurement, its impacts on individuals’ cognitive and emotional processing, and theoretical frameworks synthesizing existing findings, which have contributed greatly to our understanding of aphantasia. However, there are still some debates regarding the conclusions derived from existing research and the theories that were constructed from various sources of evidence. Building upon existing endeavors, the current systematic review emphasizes that future research is much needed to refine definition and diagnosis of aphantasia, strengthen empirical investigations at behavioral and neural levels, and more importantly, develop or update theories. These multiple lines of efforts could lead to a deeper understanding of aphantasia and further guide researchers in future research directions.
Keywords:
aphantasia
; visual imagery
; mental imagery
; vividness
; cognitive functioning
; mental health
1. Introduction
In 1880, Galton first systematically elucidated and studied individual differences in visual imagery, designing specific questions to investigate related phenomena [1]. Visual imagery ability refers to the capacity of individuals to create mental images of objects or scenes that are not in front of their eyes [2], such as closing one’s eyes and imagining a brightly colored apple. Visual imagery has been shown to be closely associated with various cognitive functions and everyday activities. The active generation of visual imagery is an important cognitive ability of humans. However, the human capacity for visual imagery can be considered a spectrum with some individuals exhibiting exceptionally vivid visual imagery, which is referred to as hyperphantasia [3,4], but other individuals can only produce limited or even deficient visual imagery. In 2015, Zeman and colleagues introduced the term “aphantasia” to describe absent or marked reduced mental imagery in individuals [5].
Presently, investigations into aphantasia concentrate on delineating its nature, assessing its frequency, devising measurement methods, examining its influence on cognitive and emotional functions, identifying related disorders, probing its neural underpinnings, and developing conceptual models. In particular, studies examining cognitive functions primarily explore how aphantasia impacts memory, atemporal and future imagination, spatial imagery, mental rotation abilities, and visual search abilities (e.g., [6,7]). Neuroimaging studies indicate a possible link between the lack of visual imagery and the visual cortex (e.g., [4,7,8]).
Aphantasia is still a relatively new topic in psychological research and has not received widespread attention from researchers. This review aims to summarize the existing empirical studies and theoretical viewpoints on aphantasia. First, our discussion will concentrate on the definition, prevalence, heritability, and assessment of aphantasia. Second, we will synthesize findings regarding aphantasia's influence on cognitive functions. Third, we will examine its potential associations with related disorders. Fourth, we will summarize existing evidence on the neural basis of aphantasia. Fifth, we will critically appraise the principal theoretical models. Last, we provide future research prospects for aphantasia, aiming at further guiding researchers in possible research directions.
2. Literature Retrieval and Screening
The present study undertook a systematic search of English articles published between 2015 and the present (Figure 1). The commencement year of 2015 was selected due to its significance as the year Zeman et al. first coined the term “aphantasia” to characterize the individual difference in the absence and marked reduction of visual imagery. due to its significance as the year Zeman et al. first coined the term “aphantasia” to characterize the databases, including Scopus, PubMed, ProQuest, and Web of Science. All journal articles, master’s and doctoral theses, conference papers, and preprints were included in the search. As of August 14, 2024, an initial retrieval yielded 255 literature sources, from which 113 duplicates were removed. Other screening methods and a careful examination of the references cited in pertinent articles further added 28 more literature sources to the pool. An in-depth review of titles, abstracts, and full texts for these 170 literature sources led to the exclusion of 10 inaccessible papers, 76 papers not centrally concerned with aphantasia, and 19 non-empirical studies, finally resulting in a curated collection of 65 pertinent literature sources.
We evaluated the quality of these studies and classified their evidence. Study quality evaluation was conducted based on the criteria used in previous works [9,10]. The details of the criteria for quality assessment are presented in Supplementary Table 1. Out of the 65 studies included, 58 studies were cross-sectional or case-control study. Seven studies were case reports or ecological studies. The following overall quality scores (%) were categorized as follows: excellent quality (score ≥81%), good quality (between 61 and 80%), fair quality (between 41 and 60%), poor quality (between 21 and 40%) and very poor quality (≤20%) (Table 1). The evaluation results showed that 58 studies were assessed as “excellent” (N = 22) or “good” (N = 36), 8 studies were assessed as “fair”, and only one study was identified as “poor” quality (Table 1).
Based on these 65 papers, the current review provides a detailed analysis, evaluation, and summary of the existing empirical research on aphantasia. It should be emphasized that the key studies published before 2015, though not using the term “aphantasia”, have also been considered in discussions if necessary. Reviews and commentaries are further incorporated into the current work. In this review, we used “aphantasia” to refer to an absence of mental imagery in any modality for consistency, although the majority of the reviewed studies focused on visual imagery deficits, The detailed content is summarized as follows (see Supplementary Materials for the details of each study included).
3. Definition, Measurement, and Prevalence of Aphantasia
Aphantasia is conceptually defined as the inability to generate mental imagery. However, variations exist in the details of the conceptual definitions [11,12]. Blomkvist reviewed various definitions of aphantasia [4,5,6,7,13,14,15,16,17,18,19]and identified three primary areas of disagreement: whether aphantasics exhibit impairments solely in visual imagery, whether a distinction is made between the absence of voluntary and involuntary imagery, and whether aphantasia primarily concerns the generation of imagery [11]. Regarding the first aspect, aphantasia has also been associated with deficits in non-visual modalities (see section 4.1). Some studies do not emphasize “visual imagery” when describing its absence, instead using the broader term “mental imagery” to refer to various modalities [7,15,18]. It is recommended to use the consistent term “aphantasia” and specify the modality when necessary, such as “auditory aphantasia” [20]. Concerning voluntariness, some researchers explicitly defined aphantasia as the lack of voluntary visual imagery [6,7,17], while others did not specifically address the aspect [14,16]. Krempel & Monzel call for simply using a broader characterization of aphantasia in the field as there is no solid evidence of preserved involuntary imagery in aphantasics [21]. Regarding the third aspect, increasing evidence suggests that aphantasia is linked to other deficits in cognitive and emotional processes (see section 4).
In measuring visual imagery deficits, most research utilizes scores from the Vividness of Visual Imagery Questionnaire (VVIQ, [22]) as the primary identification and diagnostic tool [12]. The VVIQ comprises 16 questions and employs a 5-point Likert scale. Participants are asked to rate their visual imagery ability on a scale ranging from “No imagery at all, you only ‘know’ that you are thinking of the object” to “Perfectly clear and as vivid as real seeing”, to differentiate between individuals with visual imagery deficits, those with normal imagery abilities, and those with excessive visual imagery [22]. According to the criteria, a score of 16 denotes a complete absence of visual imagery, with a score of 17-32 indicating vague/dim imagery. In recent years, a new reliable method for identifying visual imagery deficits has emerged, known as the binocular rivalry task [13,23]. Prior instructions to imagine one of the images increase the likelihood of perceiving that image during the subsequent binocular rivalry task. Participants with visual imagery deficits exhibit minimal evidence of image-based binocular rivalry priming [13,23]. The VVIQ-2, an extended version of the VVIQ containing 32 items [24], and a variant of the VVIQ have also been used in the field [4]. Recent research has also found that pupillary responses to light can indicate visual imagery deficits, as affected individuals lack an imagery pupillary light response but still exhibit a perceptual pupillary light response [25]. Additionally, rhythmic visual flickering typically induces illusory percepts, and compared to control groups, individuals with visual imagery deficits are less likely to experience complex and vivid illusions [26,27]. These findings further highlight the physiological differences between individuals with visual imagery deficits and control groups, offering an objective measure for assessing visual imagery intensity.
The vividness of the image can be viewed as a continuous characteristic forming a normal distribution curve with aphantasia occupying the left tail and hyperphantasia occupying the right tail. Before the term “aphantasia” was introduced, the most cited work based on a single question about visual imagery indicated a prevalence rate of 2.1 – 2.7% for a total absence of visual imagery in the general population [28]. In the field, the VVIQ was widely used for measuring the vividness of visual imagery. Different operational definitions have led to varying prevalence rates. Zeman et al. found that the prevalence rate of aphantasia, where imagery is completely absent, was 0.7% (VVIQ = 16), and that of moderate aphantasia with a VVIQ score of 16–23 was 2.6% [4]. Using a VVIQ score of 16-32 covering “total absent” to “vague/dim” imagery ratings, Dance et al. found a prevalence rate of 3.9% across two separate samples [29]. When using a score of 16, the rate was only 0.8%. Later, Monzel, Vetterlein & Reuter conducted meta-analyses and found a prevalence rate of 4.8% across all the included studies and 3.5% in studies using the VVIQ [30]. Recently, Takahashi et al. found in a large sample of Japanese individuals, the rate was 0.07% when using a score of 16 and 3.6% when using a score of 17-32 [31]. A study with a large sample of Brazilian university students reported a prevalence rate of 5.9% based on VVIQ scores of 16-32 [32]. However, another study noted a self-reported prevalence rate of 8.9% in the general adult population, but not all self-reported aphantasics showed low imagery scores on the VVIQ, resulting in only 1.5% [33]. Overall, the prevalence rates of aphantasia are inconsistent due to the use of different criteria across studies. If the strictest criterion is used, the rate would be extremely low. It is important to use consistent standards when discussing the prevalence rates.
For the effects of gender, studies have not found any gender differences in the prevalence of aphantasia [29,34]. Although previous studies found that women tend to score higher on object imagery measures compared to men [35,36], this difference is not statistically significant enough to conclude that men are more likely to be aphantasics. Age has also been considered as a factor influencing aphantasia. However, there is no consensus on how age affects aphantasia [34,37]. Additionally, whether the prevalence of aphantasia varies across different professions remains controversial [4,37,38], despite Galton’s initial suggestion that scientists might have weaker visual imagery abilities as a group [1]. This controversy could be explained by Blajenkova et al.’s opinion that scientists may not lack all types of visual imagery but may be specifically deficient in object imagery [39,40]. Similarly, visual artists tend to excel in spatial imagery rather than object imagery [40].
Aphantasia can be either congenital or acquired [41,42]. It is more common in individuals with a family history of lacking visual imagery, suggesting a genetic basis. However, a recent study found no significant genetic association, indicating the need for further research to explore the genetic evidence of aphantasia [43]. Acquired aphantasia can provide insights into the causes and mechanisms of aphantasia. This form of aphantasia can be triggered by various events, including craniocerebral injuries, emotional disorders, stroke, and postoperative complications [41,42,44]. A case study reported a female who developed aphantasia after contracting COVID-19 [45]. Psychological and psychiatric factors should be carefully considered in the assessment of aphantasia [46,47] as mental illnesses such as depression and anxiety can significantly impact the vividness of visual imagery [46].
4. Aphantasia and Cognitive Processing
4.1. Visual and Non-Visual Imagery Ability
Visual imagery ability is associated with other forms of imagery, such as olfactory and auditory imagery. In a pioneering study by Zeman et al., 10 out of 21 individuals with visual imagery deficits reported impairments in all forms of imagery, including auditory, olfactory, gustatory, tactile, motor, and bodily imagery, to some extent [5]. A certain number of aphantasics reported a lack of all forms of mental imagery, while some only have deficits in visual imagery [31]. In a large-scale study, 54.2% of aphantasics with visual deficits reported deficits in all types of imagery [4]. Similar findings have been confirmed in other studies, revealing that some aphantasics with visual deficits also reported deficits in other forms of mental imagery (34%, [48]; 26%, [6]). Takahashi & Gyoba reported a person with aphantasia who exhibited a complete deficit in various types of images (e.g., visual, olfactory, pain, tactile, gustatory, and somatic images) in vividness and a substantial deficit in auditory images [49]. Furthermore, individuals with hyperphantasia, who exhibit exceptionally vivid visual imagery, tend to experience more vivid imagery in other types of imagery as well [4]. There may be overlapping mechanisms underlying visual and auditory imagery, as many aphantasia participants report weak or absent auditory imagery [50,51,52] and individuals who lack auditory imagery, which is termed “anauralia”, often show visual imagery deficits [51]. However, some aphantasics self-reported higher auditory imagery than the mean score of the control group [53]. Using cluster analysis, Dawes et al. recently found that aphantasia is heterogeneous and has two subtypes: visual aphantasia, which selectively shows an absence of visual imagery, and multisensory aphantasia, which shows an inability to generate any sensory modality of mental imagery [54].
Overall, some individuals with aphantasia may also experience deficits in all types of mental imagery. However, there is still a lack of large-scale studies to establish a reliable and consistent picture, as well as plausible explanations of the neural mechanisms for understanding these phenomena. It is worth noting that most of the previous measurements of visual and other forms of mental imagery in research have relied on self-report questionnaires (e.g., VVIQ, Auditory Imagery Questionnaire, and The Questionnaire upon Mental Imagery). The latest research revealed no significant difference between aphantasics and control group in auditory imagery task, even though most of the aphantasic participants reported an impairment in auditory imagery [53]. This suggests that, although self-report questionnaires are convenient for large samples, there may be a meta-cognitive effect in the data. Participants may not actually lack genuine mental imagery but rather report deficits based on cognitive differences and misunderstandings of the questionnaire items. More precise, reliable, and objective measurement methods could help clarify the relationships between different forms of mental imagery deficits by mitigating the risks of participants’ meta-cognitive influences. Alternatively, when using self-report questionnaires, participants should be provided with clear explanations and clarifications of the items to minimize misunderstandings about the content of the questionnaires.
4.2. Aphantasia and Memory
Existing research on the impact of visual imagery impairment on memory has mainly focused on working memory, episodic memory, autobiographical memory, and object and spatial memory. Overall, aphantasics do not exhibit deficits in simple working memory tasks [55,56,57,58]. However, it has been reported that aphantasics’ perform worse than control groups in tasks requiring fine-grained visual working memory, which are cognitively more demanding and require participants to remember smaller differences [55,56,58]. Jacobs et al. reported an aphantasia individual who performed worse than controls only on the most difficult visual working memory trials requiring a high level of precision [15]. The difficulty level appears to be a crucial factor in visual working memory tasks. In terms of episodic memory, individuals with aphantasia demonstrated significantly lower performance than the control group [6,59,60]. Similarly, aphantasia individuals have been reported to experience difficulties in autobiographical memory [4,5,6,16,55,60,61,62]. Regarding spatial memory, which involves the processing of an object’s location information, two recent studies did not find any deficits in aphantasics [6,17]. For object memory, aphantasics showed poorer performance in object memory in a drawing task that requires object processing [17]. Siena & Simons used both subjective and objective measures of object and spatial memory and found that aphantasia participants showed no objective deficits in memory performance, indicating some aphantasics might show a deficit in the awareness of mental imagery [63].
It should be noted that new measurements of memory performance have been used in existing research, deviating from traditional scales and experimental tasks. For instance, drawing has been used to complement the assessment of memory performance in participants [17]. In the case of drawing, aphantasics and control groups were instructed to draw scene images from real-world memory. It was found that aphantasics remembered fewer objects and used fewer colors to draw, and relied more on verbal scaffolding to compensate for the lack of visual imager [17].
Overall, memory is a relatively well-researched aspect in the field of aphantasia. Many studies have employed questionnaires and experiments to measure participants’ memory performance. In visual working memory, aphantasics exhibit similar performance to control groups, which could be explained by their potential use of semantic encoding or other representational strategies to aid them in completing working memory tasks [7]. However, some studies indicate that aphantasics perform worse in these visual working memory tasks [15]. The inconsistency of these findings may be attributed to task characteristics, measurement methods, and individual differences among participants. For instance, the heterogeneous outcome included only one individual with aphantasia, suggesting that the observed differences in working memory could be influenced by sample size or the unique characteristics of the case study [15]. As for autobiographical memory, all results seem to converge on the conclusion that individual with aphantasia have poorer autobiographical memory. Differences in object memory and spatial memory performance between the aphantasics group and the control group may be related to their abilities in object imagery and spatial imagery. These differences may be explained through the neural mechanisms of the ventral and dorsal pathways of visual processing, which will be discussed more comprehensively later.
4.3. Aphantasia and Object and Spatial Imagery
Object imagery refers to visualizing the image appearance of objects and scenes, such as their shape, color, and brightness, while spatial imagery refers to visualizing the spatial relationships and movements of objects and their components, and spatial transformation [64]. Existing research on the performance of individuals with aphantasia in object and spatial imagery abilities has not reached a consensus and can be categorized into three types of studies. The first type of study indicates that aphantasia individuals score lower in object imagery abilities but show no significant differences from control groups in spatial imagery performance [6,17,52,60]. The second type of study shows that aphantasics score significantly lower in object imagery than control groups but perform better in spatial imagery tests [13,55]. In the last category of results, individuals with aphantasia demonstrate poorer performance in both object and spatial imagery abilities [65,66]. Based on these findings and review of the literature, researchers have proposed two types of visual imagery impairments, object aphantasia and spatial aphantasia [39,67]
The finding that object imagery is impaired while spatial imagery remains unaffected can be also explained by the functional division of visual pathways. Aphantasics’ ventral pathway for object processing may be impaired, while the dorsal pathway for spatial processing remains intact ([7,68], also see below). To better explain spatial aphantasia, researchers have proposed that this type of aphantasia may be related to functional changes in the visual dorsal stream projecting to the frontal lobe [67]. However, this explanation still lacks further support from neuroimaging studies.
4.4. Aphantasia and Atemporal and Future Imagination
Atemporal and future imagination are related to voluntarily imagining general events and future events, respectively. Autobiographical interviews on atemporal and future imagination show that aphantasia participants have poorer abilities in atemporal and future imagination, with fewer details in their imagined scenarios [16,60]. Evidence suggests that atemporal and future imagination abilities are related to autobiographical memory [16]. This may help explain why aphantasics show poorer abilities in atemporal and future imagination, as their autobiographical memory abilities are also lower compared to control groups.
4.5. Aphantasia and Mental Rotation Task Performance
The mental rotation task has been widely used to investigate individuals’ spatial abilities [69]. It has been reported that, compared to control groups, individuals with aphantasia showed equal [16,70] or even higher accuracy in mental rotation tasks [53,65,66,71]. However, aphantasics also required longer response times in these tasks [58,65,71].
Aphantasia and control groups performed similarly in mental rotation tasks, possibly due to the use of different strategies to solve the tasks [66,71]. Pounder and his colleagues suggested that aphantasics may employ non-visual processes in these tasks [58]. For example, the patient in Furman et al.’s study self-reported using a spatial and kinesthetic strategy instead of low-level visual object imagery in the mental rotation task [13,66]. Specifically, he used a first-person path-following strategy (i.e., spatial strategy) containing kinesthetic features, using his body as a reference. This hypothesis is supported by the performance of congenitally or early blind individuals in mental rotation tasks, as they did not show differences from control groups in completing complex spatial tasks, indicating that mental rotation tasks can be completed independently of visual processes [72]. Kay and his colleagues suggested that aphantasics are more likely to use analytic strategies, which do not depend on features and orientation [71]. Additionally, the mental rotation task is considered to rely on an individual’s spatial imagery ability, and many individuals with aphantasia report having similar or even better spatial imagery abilities compared to control groups [13,60].
4.6. Aphantasia and Visual Searching Ability
Visual search is a commonly used skill in everyday life that helps us find the things we need, even if the desired target is within our current field of vision. When performing visual search tasks, aphantasics exhibit significantly slower speeds compared to control groups [73,74]. This difference could be explained by the varying involvement of visual imagery in top-down strategies between the two groups [74,75]. However, in the Moriya’s task, which examines participants’ attentional guidance in visual search tasks ([76]; participants are first asked to imagine color primed by a color word such as “blue” and then to indicate whether one of the two colored squares presented on the left and right sides has an opening at the top or bottom), the priming process seems to be similar between the aphantasia group and the control group [73]. This can be explained by the characteristics of the task, where the instructions in the Moriya’s task are overly complex, resulting in a lack of visual priming and only non-visual priming [73]. These findings suggest that visual imagery could affect the way people perceive the world. Future research would benefit from designing materials that are more ecologically valid, such as incorporating environmental cues into the experimental design.
5. Aphantasia and Disorders and Emotional Processing
5.1. Emotion
Mood may influence the vividness of mental imagery [4]. By contrast, mental imagery was also examined to affect individuals’ emotion experience. Due to absent or reduced visual imagery, aphantasics experience reduced emotional engagement and less sympathy for characters in stories [77] and exhibit lower levels of fear response when reading scary materials [78]. Mental imagery ability was found to be positively associated with empathy elicited by descriptions, but not associated with empathy elicited by pictures [79]. Additionally, aphantasics seem to experience lower emotional intensity when listening to music [80]. This may be because vivid imagery makes thoughts more realistic, triggering stronger responses in the brain’s emotional circuitry and thereby amplifying emotions [81]. Future research can continue to explore different types of emotions and incorporate neuroimaging methods to validate previous findings.
5.2. Mental Health
Although aphantasia may impact individuals’ cognition and emotional processing and potentially cause certain personal distress, it is not currently included in any common clinical diagnostic systems. Furthermore, the impact of aphantasia on individuals’ daily lives and personal distress is relatively minimal and not sufficient to classify it as a psychological disorder [30.82]. However, research has found associations between aphantasia and certain conditions or diseases [83]. Besides, scientists have proposed that progressive aphantasia may be a precursor to dementia [84].
Existing research has focused on the impact of aphantasia on mental health, such as depression, anxiety, distress, and well-being. Research has revealed that the aphantasia group showed no significant differences compared to the control group in terms of depression, anxiety, and state-trait anxiety [16,81]. However, Monzel, Vetterlein & Reuter found that 34.7% of aphantasia participants reported distress caused by the lack of visual imagery [30]. Nevertheless, the researchers also suggested that these negative emotions were relatively weak and did not significantly affect individuals’ daily lives [30].
5.3. Post-Traumatic Stress Disorder (PTSD)
Previous research has established a link between mental imagery and PTSD, a condition characterized by re-experiencing traumatic events through unwanted and recurring intrusive memories and nightmares [85,86]. Aphantasics have been found to experience fewer intrusions and exhibit less avoidance behaviors following trauma [6], both of which are predictors of PTSD. Additionally, the aforementioned lack of visual imagery and its association with reduced emotional intensity may also contribute to their lower likelihood of developing PTSD. However, it is important to note that researchers emphasize the limited protective effect of visual imagery deficits when individuals with such deficits face stressful events in their daily lives [6].
5.4. Autism
Individuals with aphantasia frequently score higher on the Autism Quotient Questionnaire (AQ) and are more likely to be classified within the autism spectrum [16,48,87]. This association may be related to the connection between imagery and mentalizing. Compared to control groups, aphantasics exhibit slightly reduced atemporal and future imagination, as well as episodic memory [6,16,60], which are also characteristics of Autistic Spectrum Disorder [88,89]. Additionally, limited or absent visual imagery can impair the theory of mind ability in aphantasics, leading to difficulties in social skills [87]. These factors may explain why individuals with aphantasia tend to have higher scores on the AQ, indicating a potential connection between visual imagery deficits and Autistic Spectrum Disorder.
5.5. Prosopagnosia
Aphantasia has been associated with reduced facial recognition ability [31]. In a recent study, 5.9% of participants in the spatial aphantasia group were identified as having prosopagnosia [67], and face recognition difficulties were found to be more common in individuals with aphantasia [16,90]. Aphantasics also reported difficulties with facial recognition task [41]. Additionally, aphantasics have been associated with lower confidence in tasks involving facial perception [91]. In a study of developmental prosopagnosia, about 20% participant with developmental prosopagnosia reported a comorbidity with aphantasia [92]. However, aphantasia does not significantly impact accuracy in tasks involving the construction of facial composites and face recognition tasks [93], as aphantasia individuals can still create facial composites from memory similar to control groups [90]. Nevertheless, some researchers argued that the scores of aphantasics in tests are not sufficient to diagnose them with prosopagnosia [66]. Overall, individuals with aphantasia tend to exhibit poorer face recognition abilities. However, this may be influenced by the scales used in experiments, as these scales may elicit metacognitive effects [16]. In other words, individuals with aphantasia may have reduced confidence in their own facial recognition abilities due to their lack of vivid facial imagery, which can manifest as relatively lower scores on self-report measures.
6. Neural Basis of Aphantasia
Although numerous studies have focused on the neural basis of mental imagery (7,8,94,95), there are limited studies utilizing modern neuroimaging techniques such as magnetic resonance imaging (MRI) and electroencephalography (EEG) to explore the neural basis of visual imagery deficits. Using resting-state and task-based MRI, Milton et al. recently compared the brain activities of three groups, aphantasics, typical controls, and hyperphantasics [16]. The results showed that individuals with hyperphantasia exhibit stronger connectivity between the prefrontal regions and visual-occipital network compared to the aphantasia group. When comparing visualization and perception of famous faces and places, individuals with hyperphantasia and the control group exhibited greater frontal and parietal activation compared to the aphantasia group [16]. Another functional MRI study showed that the aphantasic group exhibited decreased activation in the hippocampus and increased activation in the visual-perceptual cortex during an autobiographical memory task [96]. However, the control group displayed strong negative task-based functional connectivity between the hippocampus and the visual cortex during the task, and the resting-state functional connectivity between these two areas could predict visualization skills [96]. Furthermore, using transcranial magnetic stimulation to induce changes in brain activity, Dupont, Papaxanthis, Madden-Lombardi & Lebonet found that there was no increase in the amplitude of motor-evoked potentials triggered in the target right index finger in the aphantasia group, indicating a lack of corticospinal excitability in individuals with aphantasia during motor simulation [97]. Similar findings were also observed in an action reading task, which involves motor simulation [98].
Case studies are also a crucial source of neural evidence. Lesion studies have found that patients showed intact visual imagery after brain lesions restricted to the occipital cortex [99,100,101,102], suggesting that early visual areas are not involved in visual imagery. Moreover, patients with damage to the anterior part of the temporal lobe, particularly in the left hemisphere, often report an inability to generate visual images [103,104,105], which is consistent with observations of a left hemisphere bias [106]. Zeman et al. reported a patient who showed reduced activation in the occipitotemporal regions during an imagery task [107]. In a recent study, a patient with an absence of visual imagery ability shows selective lesions in a specific area of the left fusiform gyrus and a portion of the right lingual gyrus, demonstrating a causal role of the left fusiform gyrus in visual imagery [41]. Therefore, the fusiform region “might act as a neural interface between sensory information coming from the occipital cortex and semantic processing in the anterior temporal lobe” during visual perception, and “could endow semantic memories with visual information during visual imagery” ([100], p517). An EEG study with source reconstruction reported that during a visual imagery task, an aphantasic begins the evoking phase from the left temporal area while lacking activation of the occipital and parietal lobes, which are associated with visual image vividness [66].
Due to the limited evidence in these studies, it is challenging to draw a consistent picture of the aberrant brain activity in aphantasia. However, we can still draw inspiration from neuroimaging studies based on individual differences in mental imagery. Spagna et al. recently conducted a meta-analysis of fMRI studies of visual imagery and found that visual imagery recruits several fronto-parietal areas and a specialized area in the left fusiform gyrus [94]. The specialized area was labeled the fusiform imagery node (FIN), referring to a brain network node specifically responsible for voluntary visual mental imagery [94,108,109]. Liu et al. found that imagery tasks activated the left frontal-parietal regions, the FIN, and areas in the ventral temporal cortex, which were similarly activated in the aphantasia and control groups [110]. However, the connectivity between the FIN and frontoparietal regions is reduced in aphantasics [110]. Together, these brain-lesion cases and fMRI studies demonstrate that the fusiform gyrus is a core area for visual imagery, and damage or impairment to it could lead to deficits in visual imagery. Additionally, abnormalities in the fusiform gyrus have been associated with prosopagnosia, which may explain the facial recognition deficits observed in individuals with visual imagery disorders [111,112].
A related debate concerns the involvement of the early visual cortex in (impaired) visual imagery [100,113]. Using stimulation, Kosslyn et al., found that the early visual cortex is causally involved in visual imagery [114]. Bergmann et al.documented that a smaller V1 size is associated with stronger but less precise imagery, indicating an anatomical basis [115]. Keogh et al. suggested a causal relationship between cortical excitability in the early visual cortex and the intensity of visual imagery [116]. However, Meng et al. suggested that an imagery-related representation exists in the primary visual cortex of aphantasics, despite the absence of visual imagery, though the representation contains less or transformed sensory information [117]. Result from Cabbai et al. showed a dissociation between V1 representations and subjective imagery [118]. Bartolomeo et al. [100] argued that the left fusiform gyrus plays a crucial role in visual imagery, rather than the early visual cortex, and that the involvement of the early visual cortex [114] might be modulated by downstream areas. Dijkstra recently reviewed existing evidence and proposed that “imagery can recruit the early visual cortex, but that does not mean that it always does so” [95].
7. Theory Development
Individuals with imagery deficits can still have imagination capabilities similar to those without such deficits, such as creating novels and movies. Based on this, Arcangeli attempts to distinguish between mental imagery and sensory imagination [119]. Mental imagery can be considered a type of mental content (e.g., the appearance of an apple). In contrast, sensory imagination is a special psychological attitude that involves the recreation of perceptual experiences. According to this theory, most individuals previously defined as having aphantasia in past research may actually have a deficiency in sensory imagination rather than in mental imagery, which could explain why individuals with visual imagery deficits can still perform imaginative tasks.
Nanay explains aphantasia from the perspective of both conscious and unconscious visual imagery [18]. Previous research has found that individuals with aphantasia do not exhibit the imagery priming effect in binocular rivalry tasks; yet, some participants can generate vivid dreams and perform visual imagery tasks similar to control groups. To explain this phenomenon, Nanay distinguishes between conscious and unconscious visual imagery abilities, both of which could be generated voluntarily or involuntarily [18]. One type of aphantasia involves a fundamental lack of visual imagery, while the other type involves the ability to generate visual imagery without conscious awareness [73]. However, this theory is challenged by Blomkvist [11], who argues that it does not fully explain the issues of episodic memory or imagination of atemporal and future events in visual imagery deficits, as it does not provide a link between mental imagery and the episodic processes in episodic memory and episodic imagination. Additionally, there is research suggesting that imagery tasks can be accomplished through cognitive strategies that do not rely on imagery [91,120,121,122]. A recent study using both implicit and explicit priming tasks found that participants with aphantasia did not show priming effects, suggesting that aphantasia is an inability to generate visual imageries rather than an impairment in conscious awareness of images [123].
To provide a better explanation for aphantasia, Blomkvist proposed enhancements to the Constructive Episodic Simulation Hypothesis (CESH) model [11]. The original CESH posits that memory and imagination involve three key processes: semantic retrieval process, episodic retrieval process, and (re)combination process [124]. Blomkvist expanded on this model by adding three new components: “memory indices, differing episodic retrieval mechanisms for all kinds of sensory information, and spatial retrieval mechanisms” [11]. These additions aim to provide a new theoretical explanation for visual imagery deficits. According to this updated theory, the mechanism of the episodic system is deficient, resulting in the loss of visual imagery. However, this theory still lacks empirical research data to support it further.
The theoretical framework of the ventral and dorsal pathways in the visual system has been proposed for many years [125,126,127] (although there are ongoing debates about this framework, [128,129]. Pearson proposed that the ventral (or “what”) pathway is associated with object information, while the dorsal (or “where”) pathway is associated with location and spatial features [7]. The two aspects of visual imagery, object imagery and spatial imagery, are also likely generated through the ventral and dorsal pathways [39]. Damage to the ventral pathway could impair individuals’ ability to visualize objects’ appearance, while damage to the dorsal pathway is associated with a disrupted ability in spatial imagery. These processes can be dissociated in aphantasia, which may explain why individuals with visual imagery deficits perform similarly to, or even better than, control groups in spatial imagery tasks [7,68]. Bergmann and Ortiz-Tudela also suggested that visual feedback pathways used during episodic and schematic memory retrieval may be different depending on the two visual processing streams: episodic memory retrieval involves both “what” and “where” streams, while schematic memory retrieval primarily involves the “where” stream [130]. In other words, aphantasia may be associated with differences in the “what” stream.
Inspired by neural models of mental imagery [7,8] and existing sources of neural evidence, Zeman tried to depict candidate neural mechanisms of extreme imagery, aphantasia, and hyperphantasia [2]. In the brain mapping picture, there are five functional clusters, each involved in a unique role in mental imagery: the frontal cortex for initiating imagery generation, the parietal cortex for interacting with the frontal cortex to generate imagery during which attentional and spatial aspects of imagery are mediated, the temporal cortex including limbic structures for enabling access to the semantic and episodic memories that determine what to visualize, higher-order visual areas (e.g., the FIN) for visualizing imagery. The extent to which activity in the early visual cortex (e.g., the V1) is required for imagery is still debated. Moreover, the connections between these clusters are also essential for imagery generation. From this mapping picture, Zeman [2]proposes five candidates showing neural atypicality in extreme imageries, variations in the strengths of the top-down feedback connection between higher-order regions (the frontal cortex) serving as cognitive control and modality-specific areas (e.g., the visual cortex) activated by sensory imagery, variations in the structure and function of the frontal cortex involved in generating imagery, variations at the level of higher-order visual areas including the FIN proposed by Spagna et al., [94], variations in the anatomical structure of the early visual cortex including the V1, variations in the saliency involves the parietal areas of the frontoparietal control system. In this proposal, Zeman provided future directions for researchers in the field [2]. A neural model constructed and refined from accumulated evidence would add to a better understanding of aphantasia.
8. Summary and Future Directions
This review primarily examines the definition, prevalence, and measurement methods of aphantasia, as well as its impacts on individual cognitive and emotional processing, associated disorders, neural basis, and theory development. Research on aphantasia is continuously progressing, and its descriptions are becoming increasingly clear. However, several debated areas remain, and the study of its impacts on individuals and underlying mechanisms faces several challenges. Future investigations into aphantasia should focus on the following aspects.
8.1. Clarify Definition and Diagnosis
It is evident that the current research on aphantasia is still in its early infancy. One of the primary concerns is the definition of aphantasia. Clear definitions serve as the foundation for advancing further research and facilitating effective communication among researchers. The existing literature presents variations in the definition of aphantasia [12]. For example, some definitions do not explicitly distinguish between voluntary and involuntary imagery (see a review by [21]). The cut scores for the diagnosis of aphantasia vary across studies (see section 3). The inconsistent use of the arbitrary cut scores is considered a hindrance to cross-study comparison and communication among researchers. Moreover, the use of cut scores may also discourage efforts to examine individual differences in imagery. Additionally, more effective, objective tools for non-clinical and clinical purposes should be developed [131,132] rather than relying predominantly on the VVIQ or its variants [133].
The terminology in the field has recently been debated [20,134,135,136]. While the debate continues on whether the term “aphantasia” specifically refers to the absence of visual mental imagery or encompasses a deficit of all types of mental imagery (see section 3), most current research on aphantasia focuses on visual imagery. Other terms have been proposed to refer to an absence of other modalities of imagery. For auditory imagery, Hinwar and Lambert introduced the term “anauralia” to refer to an absence of auditory imagery and “hyperauralia” to refer to the experience of extremely vivid auditory imagery [51]. Dance, Ward & Simner proposed the term “dysikonesia” to refer to multisensory or global aphantasia [48]. In this context, Monzel, Mitchell et al. believe that these new terms complicate the field, making communication less effective for researchers and the public, and therefore advocate for the use of the simple term “aphantasia,” which is widely known [20]. When referring to an absence of a specific modality of imagery, it is easy to use modality-specific terms (e.g., “visual/auditory/multisensory aphantasia”).
These divergencies across studies create difficulties in the classification of participants and scientific communications among researchers and the public, therefore hindering progress toward a better understanding of aphantasia. Researchers in the field should reach a consensus in these issues to facilitate further development.
8.2. Strengthen Behavioral Research
Aphantasia research is still at its early stages, and there is limited literature in several research directions. Future studies should build upon existing efforts to systematically examine this phenomenon to better understand aphantasia. At the behavioral level, more research is urgently needed within and beyond existing research directions. First of all, together with a clear and consistent definition of aphantasia, researchers should focus on examining the nature of aphantasia, including but not limited to investigating effects of demographic variables (e.g., age and gender) and their interactions in large datasets, developing new assessment or diagnostic tools with high reliability and validity, elucidating perceptual and cognitive processes required by tasks used to elicit mental imagery, and determining the comorbidity rate of two or more subtypes of aphantasia in large samples. Second, researchers should investigate how aphantasia is associated with other psychological aspects (e.g., conscious experience, [18,137,138,139], the influences of aphantasia on emotional processing and disorders (see section 4), and possible mechanisms underlying these relationships (e.g., [140]). Third, research on hyperphantasia, the opposite extreme characterized by vivid mental imagery, can also help contribute to a better understanding of aphantasia. Viewing mental imagery ability as a continuum could be more realistic and scientifically useful in understanding the absence or reduction of imagery in people with aphantasia. Fourth, researchers should use these different sources of evidence as a foundation to propose or create intervention programs for people with aphantasia. The effectiveness of these interventions can also help researchers better understand the nature of this phenomenon, fostering further advances.
8.3. Discover Neural Basis
Unraveling the neural bases of aphantasia has recently become a prominent research topic. Neuroimaging evidence has been instrumental in understanding the manifestations of aphantasia and in constructing neural models. Employing neuroimaging methods, future research can explore various variables to gather more neural evidence. Specifically, researchers can utilize the high spatial resolution of MRI techniques to investigate regional activation and inter-regional connections (structural and functional connectivity) involved in the absence or reduction of mental imagery. Previous functional MRI studies have identified areas involved in mental imagery, based on which neural models have also been proposed (e.g., [14,115,116]; see a review by [7]). Although some studies of aphantasia have been conducted [16,91,107], it is difficult to accumulate convergent evidence for localizing aberrant brain regions and connections. Using EEG and magnetoencephalography (MEG) techniques with high temporal resolution [141,142], researchers can examine the neural dynamics of the processes involving mental imagery in people with and without aphantasia (e.g., [66]). For example, Xie et al. found shared alpha-band neural representations in visual imagery and perception [143]. Combining spatial and temporal neural evidence across different studies, or even using fusion techniques (e.g., EEG-MRI fusion, [144]) can contribute to a better understanding of aphantasia by incorporating different types of information into a single picture, further facilitating the construction of neural models. Recent technical advances in multi-variate pattern analysis (e.g., [145,146]) may also be powerful in understanding the neural correlates of aphantasia, as demonstrated in previous studies of mental imagery [111,117,143]. Non-invasive brain stimulation techniques [147] can be powerful tools for discovering causal evidence of the involvement of brain areas in mental imagery. Together, the convergence of behavioral and neural evidence not only helps elucidate various aspects of aphantasia but also is useful in building behavioral and neural models that explain this phenomenon [2]. In addition, evidence from other sources of investigation should also be encouraged. It has been reported that aphantasia has a genetic basis [4] and dopamine plays an important role in generating mental imagery [148].
8.4. Construct and Refine Theories
Good theories help humans systematically and scientifically understand their minds [149,150] Although aphantasia has garnered increasing attention over the past decade, and it also gained interest from art, philosophical, and theoretical discussions [80,83,151,152,153,154,155,156,157,158,159,160], empirical studies examining this phenomenon remain limited. The field is still in its early stages. A lack of theories for synthesizing existing literature is one of the field’s significant characteristics. Although some accounts and models have been proposed to interpret behavioral findings [11,18,119], they have focused on certain aspects. Neural models for synthesizing neural evidence (e.g., [2]) are largely unknown or require further examination. One of the main reasons is that both behavioral and neural empirical studies are very limited in number and diversity (see above), resulting in insufficient evidence to be integrated into a theory or model.
In addition to proposing new theories or models, we advocate for efforts to use existing theories or models of mental imagery [7,8,11,109] as a foundation to theoretically guide future directions on researching aphantasia. Modifications to these models or theories will be made as more evidence is collected. We believe these changes will either refine existing theories or help foster new ones. For instance, theories that explain the mechanisms of visual imagery can be adapted to account for the absence of such imagery in individuals with aphantasia. By modifying these models to include the cognitive and neural underpinnings of aphantasia, researchers can create a more robust theoretical framework that encompasses both the presence and absence of mental imagery.
Given that aphantasia may extend beyond visual imagery to include other modalities, such as auditory or tactile imagery, theories should be developed to address these broader aspects. This could involve creating a unified model that categorizes different types of aphantasia (e.g., visual, auditory, multisensory) and explores how these modalities interact within cognitive processes. Such a model would not only clarify the nature of aphantasia but also facilitate comparisons with individuals who possess typical imagery capabilities.
8.5. Encourage Direct and Conceptual Replications
In recent years, concerns about replication in psychology have increased (e.g., [161,162]). Replicability is a crucial indicator of the reliability and stability of research findings [163]. Conducting replicable experiments to demonstrate the replicability of the findings lays a solid foundation for further investigations. There are two types of replications, each with different requirements. Conceptual replication deliberately modifies key elements of the original program to test the robustness of the phenomenon or the generalizability of the theoretical claims, while direct replication involves recreating the original experiment, which has been questioned regarding its impossibility of implementation [164].
The inconsistencies in research findings on aphantasia, which can undermine confidence in the literature, calls for considering replicability in existing studies, requiring numerous replication studies to confirm the reliability and stability of results. For example, there are limited studies on the relationship between aphantasia and atemporal and future imagination, as well as visual search. Although consistent conclusions have been reached, different experimental designs are still needed to validate the accuracy and replicability of the results (conceptual replications). Moreover, researchers have explored the relationships between attentional templates [19], sensory sensitivity [48], verbal overshadowing [120], mid-level characteristics of drawings [165], and aphantasia, but these are isolated studies lacking cross-validation from other research, indicating the necessity of conducting direct and conceptual replication studies.
To facilitate replication efforts, aphantasia researchers should adopt open science practices, such as sharing data, materials, and protocols. By making these more transparent, the scientific community of aphantasia can more easily conduct replications and build upon existing work. Open access to the datasets of research published or to be published can also encourage more collaboration and foster a collective effort to understand this phenomenon more deeply.
9. Conclusion
In this work, we provide a comprehensive review of aphantasia, encompassing its definition, prevalence, measurement methods, empirical research, and relevant theories. The existing literature has contributed to a good understanding of aphantasia. The absence of visual imagery can have significant implications for an individual’s cognitive and emotional processing and is also associated with certain psychological disorders. Studies on the neural bases of aphantasia are gradually increasing, especially in recent years. Following these endeavors, theories and neural models have been proposed to summarize existing behavioral and neural evidence. The research of visual imagery deficits is continually growing, although there are still areas of debate. However, a consensus regarding the theoretical framework has not yet been achieved. The theories still lack sufficient empirical support, leaving significant gap. We emphasize several points researchers can make contributions to, such as clarifying diagnosis, strengthening behavioral and neuroimaging research, constructing and refining theories, and encouraging replications. New works on the different aspects of aphantasia is not only beneficial for expanding researchers’ understanding of cognitive and emotional processes and their relationships with other disorders, but also helpful in building theories or model.
Author Contributions
F.J., study design, investigation, methodology, software, visualization, writing - original draft, writing - review and editing; H.S. investigation, writing - review and editing. Y.L., study design, funding acquisition, investigation, methodology, resources, supervision, writing - original draft, writing - review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by BNU-HKBU United International College Re- search Grants (R72021207, R202102, R202011,) and in part by the Guangdong Provincial Key Laboratory IRADS (2022B1212010006, R0400001-22).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Galton, F. I.--Statistics of mental imagery. mind 1880, os-V, 301–318. [CrossRef]
- Zeman, A. Aphantasia and hyperphantasia: exploring imagery vividness extremes. Trends Cogn. Sci. 2024, 28, 467–480. [CrossRef]
- Keogh, R., Pearson, J., & Zeman, A. (2021). Aphantasia: The science of visual imagery extremes. Handbook of Clinical Neurology (pp. 277-296).
- Zeman, A.; Milton, F.; Della Sala, S.; Dewar, M.; Frayling, T.; Gaddum, J.; Hattersley, A.; Heuerman-Williamson, B.; Jones, K.; MacKisack, M.; et al. Phantasia–The psychological significance of lifelong visual imagery vividness extremes. Cortex 2020, 130, 426–440. [CrossRef]
- Zeman, A.; Dewar, M.; Della Sala, S. Lives without imagery – Congenital aphantasia. Cortex 2015, 73, 378–380. [CrossRef]
- Dawes, A.J.; Keogh, R.; Andrillon, T.; Pearson, J. A cognitive profile of multi-sensory imagery, memory and dreaming in aphantasia. Sci. Rep. 2020, 10, 1–10. [CrossRef]
- Pearson, J. The human imagination: the cognitive neuroscience of visual mental imagery. Nat. Rev. Neurosci. 2019, 20, 624–634. [CrossRef]
- Dijkstra, N.; Bosch, S.E.; van Gerven, M.A. Shared Neural Mechanisms of Visual Perception and Imagery. Trends Cogn. Sci. 2019, 23, 423–434. [CrossRef]
- Luchini, C.; Veronese, N.; Nottegar, A.; Shin, J.I.; Gentile, G.; Granziol, U.; Soysal, P.; Alexinschi, O.; Smith, L.; Solmi, M. Assessing the quality of studies in meta-research: Review/guidelines on the most important quality assessment tools. Pharm. Stat. 2021, 20, 185–195. [CrossRef]
- Shaheen, N.; Shaheen, A.; Ramadan, A.; Hefnawy, M.T.; Ramadan, A.; Ibrahim, I.A.; Hassanein, M.E.; Ashour, M.E.; Flouty, O. Appraising systematic reviews: a comprehensive guide to ensuring validity and reliability. Front. Res. Metrics Anal. 2023, 8, 1268045. [CrossRef]
- Blomkvist, A. Aphantasia: In search of a theory. Mind Lang. 2022, 38, 866–888. [CrossRef]
- Blomkvist, A., & Marks, D. F. (2023). Defining and 'diagnosing' aphantasia: Condition or individual difference?. Cortex, 169, 220–234. [CrossRef]
- Keogh, R.; Pearson, J. The blind mind: No sensory visual imagery in aphantasia. Cortex 2018, 105, 53–60. [CrossRef]
- Fulford, J.; Milton, F.; Salas, D.; Smith, A.; Simler, A.; Winlove, C.; Zeman, A. The neural correlates of visual imagery vividness – An fMRI study and literature review. Cortex 2018, 105, 26–40. [CrossRef]
- Jacobs, C.; Schwarzkopf, D.S.; Silvanto, J. Visual working memory performance in aphantasia. Cortex 2018, 105, 61–73. [CrossRef]
- Milton, F.; Fulford, J.; Dance, C.; Gaddum, J.; Heuerman-Williamson, B.; Jones, K.; Knight, K.F.; MacKisack, M.; Winlove, C.; Zeman, A. Behavioral and Neural Signatures of Visual Imagery Vividness Extremes: Aphantasia versus Hyperphantasia. Cereb. Cortex Commun. 2021, 2, tgab035. [CrossRef]
- Bainbridge, W.A.; Pounder, Z.; Eardley, A.F.; Baker, C.I. Quantifying aphantasia through drawing: Those without visual imagery show deficits in object but not spatial memory. Cortex 2021, 135, 159–172. [CrossRef]
- Nanay, B. (2021). Unconscious mental imagery: Unconscious mental imagery. Philosophical Transactions of the Royal Society B: Biological Sciences, 376(1817). [CrossRef]
- Keogh, R., & Pearson, J. (2021). Attention driven phantom vision: Measuring the sensory strength of attentional templates and their relation to visual mental imagery and aphantasia: Measuring attentional templates. Philosophical Transactions of the Royal Society B: Biological Sciences, 376(1817). [CrossRef]
- Monzel, M., Mitchell, D., Macpherson, F., Pearson, J., & Zeman, A. (2022a). Aphantasia, dysikonesia, anauralia: Call for a single term for the lack of mental imagery - Commentary on dance et al. (2021) and Hinwar and lambert (2021). Cortex, 150, 149-152. [CrossRef]
- Krempel, R.; Monzel, M. Aphantasia and involuntary imagery. Conscious. Cogn. 2024, 120, 103679. [CrossRef]
- Marks, D.F. Visual Imagery Differences in the Recall of Pictures. Br. J. Psychol. 1973, 64, 17–24. [CrossRef]
- Keogh, R.; Pearson, J. Revisiting the blind mind: Still no evidence for sensory visual imagery in individuals with aphantasia. Neurosci. Res. 2024, 201, 27–30. [CrossRef]
- Marks, D. F. (1995). New directions for mental imagery research. Journal of Mental Imagery, 19(3e4), 153e167.
- Kay, L., Keogh, R., Andrillon, T., & Pearson, J. (2022). The pupillary light response as a physiological index of aphantasia, sensory and phenomenological imagery strength. Elife, 11, e72484. [CrossRef]
- Königsmark, V.T.; Bergmann, J.; Reeder, R.R. The Ganzflicker experience: High probability of seeing vivid and complex pseudo-hallucinations with imagery but not aphantasia. Cortex 2021, 141, 522–534. [CrossRef]
- Reeder, R.R. Ganzflicker Reveals the Complex Relationship Between Visual Mental Imagery and Pseudo-Hallucinatory Experiences: A Replication and Expansion. Collabra: Psychol. 2022, 8. [CrossRef]
- Faw, B. (2009). Conflicting intuitions may be based on differing abilities: Evidence from mental imaging research. Journal of Consciousness Studies, 16(4), 45-68.
- Dance, C.; Ipser, A.; Simner, J. The prevalence of aphantasia (imagery weakness) in the general population. Conscious. Cogn. 2022, 97, 103243. [CrossRef]
- Monzel, M.; Vetterlein, A.; Reuter, M. No general pathological significance of aphantasia: An evaluation based on criteria for mental disorders. Scand. J. Psychol. 2022, 64, 314–324. [CrossRef]
- Takahashi, J.; Saito, G.; Omura, K.; Yasunaga, D.; Sugimura, S.; Sakamoto, S.; Horikawa, T.; Gyoba, J. Diversity of aphantasia revealed by multiple assessments of visual imagery, multisensory imagery, and cognitive style. Front. Psychol. 2023, 14, 1174873. [CrossRef]
- Gouveia, L. A. (2023). Imagination capacity in university students: aphantasia and its possible changes in personal development. [CrossRef]
- Beran, M.J.; James, B.T.; French, K.; Haseltine, E.L.; Kleider-Offutt, H.M. Assessing aphantasia prevalence and the relation of self-reported imagery abilities and memory task performance. Conscious. Cogn. 2023, 113, 103548. [CrossRef]
- Gulyás, E.; Gombos, F.; Sütöri, S.; Lovas, A.; Ziman, G.; Kovács, I. Visual imagery vividness declines across the lifespan. Cortex 2022, 154, 365–374. [CrossRef]
- Campos, A. (2014). Gender differences in imagery. Personality and Individual Differences, 59, 107-111. [CrossRef]
- Aydin, Ç. Gender Differences in Visual Imagery: Object and Spatial Imagery. 2020, 22, 1045–1064. [CrossRef]
- Smyth, R.S.; Acharya, P.; Hunt, N.P. Is visual imagery ability higher for orthodontic students than those in other disciplines? A cross-sectional questionnaire-based study. J. Orthod. 2019, 46, 205–211. [CrossRef]
- Fielding, D.; Kahui, V.; Wesselbaum, D. Visual imagination and the performance of undergraduate economics students. New Zealand Econ. Pap. 2020, 54, 127–137. [CrossRef]
- Blazhenkova, O.; Pechenkova, E.; National Research University Higher School of Economics The Two Eyes of the Blind Mind: Object vs. Spatial Aphantasia?. Russ. J. Cogn. Sci. 2019, 6, 51–62. [CrossRef]
- Blajenkova, O., Kozhevnikov, M., & Motes, M. A. (2006). Object-spatial imagery: A new self-report imagery questionnaire. Applied Cognitive Psychology: The Official Journal of the Society for Applied Research in Memory and Cognition, 20(2), 239-263. [CrossRef]
- Thorudottir, S.; Sigurdardottir, H.M.; Rice, G.E.; Kerry, S.J.; Robotham, R.J.; Leff, A.P.; Starrfelt, R. The Architect Who Lost the Ability to Imagine: The Cerebral Basis of Visual Imagery. Brain Sci. 2020, 10, 59. [CrossRef]
- Knowles, L.; Jones, K.; Zeman, A. #3112 Acquired aphantasia in 88 cases: a preliminary report. J. Neurol. Neurosurg. Psychiatry 2021, 92, A6.3–A7. [CrossRef]
- Day, J.; Frayling, T.; Wood, A.; Zeman, A. 130 Does visual imagery vividness have a genetic basis? A genome-wide associa- tion study of 1019 individuals. J. Neurol. Neurosurg. Psychiatry 2022, 93, A51.1–A51. [CrossRef]
- Bumgardner, A.L.; Yuan, K.; Chiu, A.V. I cannot picture it in my mind: acquired aphantasia after autologous stem cell transplantation for multiple myeloma. Oxf. Med Case Rep. 2021, 8, 158–160. [CrossRef]
- Gaber, T. A. K., & Eltemamy, M. (2021). Post-COVID-19 aphantasia. Progress in Neurology and Psychiatry, 25(3), 16-17.
- De Vito, S., & Bartolomeo, P. (2016). Refusing to imagine? on the possibility of psychogenic aphantasia. A commentary on Zeman et al. (2015). Cortex, 74, 334-335.
- Zeman, A.; Dewar, M.; Della Sala, S. Reflections on aphantasia. Cortex 2016, 74, 336–337. [CrossRef]
- Dance, C.J.; Ward, J.; Simner, J. What is the Link Between Mental Imagery and Sensory Sensitivity? Insights from Aphantasia. Perception 2021, 50, 757–782. [CrossRef]
- Takahashi, J., & Gyoba, J. (2021). A preliminary single-case study of aphantasia in Japan. Tohoku psychologica folia, 79, 26-32.
- Bouyer, L.N.; Arnold, D.H. Deep Aphantasia: a visual brain with minimal influence from priors or inhibitory feedback?. Front. Psychol. 2024, 15, 1374349. [CrossRef]
- Hinwar, R.P.; Lambert, A.J. Anauralia: The Silent Mind and Its Association With Aphantasia. Front. Psychol. 2021, 12. [CrossRef]
- Wittmann, B.C.; Şatırer, Y. Decreased associative processing and memory confidence in aphantasia. Learn. Mem. 2022, 29, 412–420. [CrossRef]
- Pounder, Z.; Eardley, A.F.; Loveday, C.; Evans, S. No clear evidence of a difference between individuals who self-report an absence of auditory imagery and typical imagers on auditory imagery tasks. PLOS ONE 2024, 19, e0300219. [CrossRef]
- Dawes, A.J.; Keogh, R.; Pearson, J. Multisensory subtypes of aphantasia: Mental imagery as supramodal perception in reverse. Neurosci. Res. 2024, 201, 50–59. [CrossRef]
- Ganczarek, J., Żurawska-Żyła, R., & Rolek, A. (2020). “I remember things, but I can’t picture them.” what can a case of aphantasia tell us about imagery and memory? Psychiatria i Psychologia Kliniczna, 20(2), 134-141. [CrossRef]
- Keogh, R.; Wicken, M.; Pearson, J. Visual working memory in aphantasia: Retained accuracy and capacity with a different strategy. Cortex 2021, 143, 237–253. [CrossRef]
- Knight, K. F., & Milton, F. (2022). Memory without Imagery: No Evidence of Visual Working Memory Impairment in People with Aphantasia. In Proceedings of the Annual Meeting of the Cognitive Science Society (Vol. 44, No. 44).
- Pounder, Z.; Jacob, J.; Evans, S.; Loveday, C.; Eardley, A.F.; Silvanto, J. Only minimal differences between individuals with congenital aphantasia and those with typical imagery on neuropsychological tasks that involve imagery. Cortex 2022, 148, 180–192. [CrossRef]
- Dando, C.J.; Nahouli, Z.; Hart, A.; Pounder, Z. Real-world implications of aphantasia: episodic recall of eyewitnesses with aphantasia is less complete but no less accurate than typical imagers. R. Soc. Open Sci. 2023, 10, 231007. [CrossRef]
- Dawes, A.J.; Keogh, R.; Robuck, S.; Pearson, J. Memories with a blind mind: Remembering the past and imagining the future with aphantasia. Cognition 2022, 227, 105192. [CrossRef]
- Monzel, M.; Vetterlein, A.; Reuter, M. Memory deficits in aphantasics are not restricted to autobiographical memory – Perspectives from the Dual Coding Approach. J. Neuropsychol. 2021, 16, 444–461. [CrossRef]
- Watkins, N.W. (A)phantasia and severely deficient autobiographical memory: Scientific and personal perspectives. Cortex 2018, 105, 41–52. [CrossRef]
- Siena, M.J.; Simons, J.S. Metacognitive Awareness and the Subjective Experience of Remembering in Aphantasia. J. Cogn. Neurosci. 2024, 36, 1–21. [CrossRef]
- Kozhevnikov, M.; Kosslyn, S.; Shephard, J. Spatial versus object visualizers: A new characterization of visual cognitive style. Mem. Cogn. 2005, 33, 710–726. [CrossRef]
- Crowder, A. (2018). Differences in spatial visualization ability and vividness of spatial imagery between people with and without aphantasia. Virginia Commonwealth University.
- Furman, M., Fleitas-Rumak, P., Lopez-Segura, P., Furman, M., Tafet, G., de Erausquin, G. A., & Ortiz, T. (2022). Cortical activity involved in perception and imagery of visual stimuli in a subject with aphantasia. an EEG case report. Neurocase, 28(4), 344-355. [CrossRef]
- Palermo, L.; Boccia, M.; Piccardi, L.; Nori, R. Congenital lack and extraordinary ability in object and spatial imagery: An investigation on sub-types of aphantasia and hyperphantasia. Conscious. Cogn. 2022, 103, 103360. [CrossRef]
- Keogh, R.; Pearson, J. The perceptual and phenomenal capacity of mental imagery. Cognition 2017, 162, 124–132. [CrossRef]
- Shepard, R.N.; Metzler, J. Mental Rotation of Three-Dimensional Objects. Science 1971, 171, 701–703. [CrossRef]
- Zhao, B.; Della Sala, S.; Zeman, A.; Gherri, E. Spatial transformation in mental rotation tasks in aphantasia. Psychon. Bull. Rev. 2022, 29, 2096–2107. [CrossRef]
- Kay, L., Keogh, R., & Pearson, J. (2024). Slower but more accurate mental rotation performance in aphantasia linked to differences in cognitive strategies. Consciousness and cognition, 121, 103694. [CrossRef]
- Eardley, A.F.; Pring, L. Spatial processing, mental imagery, and creativity in individuals with and without sight. Eur. J. Cogn. Psychol. 2007, 19, 37–58. [CrossRef]
- Monzel, M., Keidel, K., & Reuter, M. (2021). Imagine, and you will find – lack of attentional guidance through visual imagery in aphantasics. Attention, Perception, and Psychophysics, 83(6), 2486-2497. [CrossRef]
- Monzel, M.; Reuter, M. Where’s Wanda? The influence of visual imagery vividness on visual search speed measured by means of hidden object pictures. Attention, Perception, Psychophys. 2024, 86, 22–27. [CrossRef]
- Pearson, J.; Clifford, C.W.; Tong, F. The Functional Impact of Mental Imagery on Conscious Perception. Curr. Biol. 2008, 18, 982–986. [CrossRef]
- Moriya, J. Visual mental imagery influences attentional guidance in a visual-search task. Attention, Perception, Psychophys. 2018, 80, 1127–1142. [CrossRef]
- Speed, L.J.; Eekhof, L.S.; Mak, M. The role of visual imagery in story reading: Evidence from aphantasia. Conscious. Cogn. 2024, 118, 103645. [CrossRef]
- Wicken, M.; Keogh, R.; Pearson, J. The critical role of mental imagery in human emotion: insights from fear-based imagery and aphantasia. Proc. R. Soc. B: Biol. Sci. 2021, 288, 20210267. [CrossRef]
- Monzel, M.; Keidel, K.; Reuter, M. Is it really empathy? The potentially confounding role of mental imagery in self-reports of empathy. J. Res. Pers. 2023, 103. [CrossRef]
- Hashim, S.; Pulcini, C.; Jansari, A.; Küssner, M.B.; Omigie, D. The Experience of Music in Aphantasia: Emotion, Reward, and Everyday Functions. Music. Sci. 2024, 7. [CrossRef]
- Keogh, R., Wicken, M., & Pearson, J. (2023). Fewer intrusive memories in aphantasia: using the trauma film paradigm as a laboratory model of PTSD. PsyArxiv, 2023.
- Monzel, M.; Dance, C.; Azañón, E.; Simner, J. Aphantasia within the framework of neurodivergence: Some preliminary data and the curse of the confidence gap. Conscious. Cogn. 2023, 115, 103567. [CrossRef]
- Cavedon-Taylor, D. Aphantasia and psychological disorder: Current connections, defining the imagery deficit and future directions. Front. Psychol. 2022, 13, 822989. [CrossRef]
- Sitek, E.J.; Konieczna, S. Does progressive aphantasia exist? The hypothetical role of aphantasia in the diagnosis of neurodegenerative diseases. Behav. Brain Sci. 2022, 45, e299–51. [CrossRef]
- Pearson, J.; Naselaris, T.; Holmes, E.A.; Kosslyn, S.M. Mental Imagery: Functional Mechanisms and Clinical Applications. Trends Cogn. Sci. 2015, 19, 590–602. [CrossRef]
- Yehuda, R., Hoge, C. W., McFarlane, A. C., Vermetten, E., Lanius, R. A., Nievergelt, C. M., ... & Hyman, S. E. (2015). Post-traumatic stress disorder. Nature Reviews Disease Primers, 1(1), 1-22. [CrossRef]
- Dance, C. J., Jaquiery, M., Eagleman, D. M., Porteous, D., Zeman, A., & Simner, J. (2021). What is the relationship between aphantasia, synaesthesia and autism? Consciousness and Cognition, 89, 103087. [CrossRef]
- Crespi, B.; Leach, E.; Dinsdale, N.; Mokkonen, M.; Hurd, P. Imagination in human social cognition, autism, and psychotic-affective conditions. Cognition 2016, 150, 181–199. [CrossRef]
- Lord, C., Elsabbagh, M., Baird, G., & Veenstra-Vanderweele, J. (2018). Autism spectrum disorder. The Lancet, 392(10146), 508-520. [CrossRef]
- Dance, C.J.; Hole, G.; Simner, J. The role of visual imagery in face recognition and the construction of facial composites. Evidence from Aphantasia. Cortex 2023, 167, 318–334. [CrossRef]
- Liu, J.; Bartolomeo, P. Probing the unimaginable: The impact of aphantasia on distinct domains of visual mental imagery and visual perception. Cortex 2023, 166, 338–347. [CrossRef]
- Svart, N.; Starrfelt, R. Is It Just Face Blindness? Exploring Developmental Comorbidity in Individuals with Self-Reported Developmental Prosopagnosia. Brain Sci. 2022, 12, 230. [CrossRef]
- Monzel, M.; Vetterlein, A.; Hogeterp, S.A.; Reuter, M. No increased prevalence of prosopagnosia in aphantasia: Visual recognition deficits are small and not restricted to faces. Perception 2023, 52, 629–644. [CrossRef]
- Spagna, A.; Hajhajate, D.; Liu, J.; Bartolomeo, P. Visual mental imagery engages the left fusiform gyrus, but not the early visual cortex: A meta-analysis of neuroimaging evidence. Neurosci. Biobehav. Rev. 2021, 122, 201–217. [CrossRef]
- Dijkstra, N. Uncovering the Role of the Early Visual Cortex in Visual Mental Imagery. Vision 2024, 8, 29. [CrossRef]
- Monzel, M., Leelaarporn, P., Lutz, T., Schultz, J., Brunheim, S., Reuter, M., & McCormick, C. (2023). Hippocampal-occipital connectivity reflects autobiographical memory deficits in aphantasia. bioRxiv, 2023-08. [CrossRef]
- Dupont, W.; Papaxanthis, C.; Madden-Lombardi, C.; Lebon, F. Explicit and implicit motor simulations are impaired in individuals with aphantasia. Brain Commun. 2024, 6, fcae072. [CrossRef]
- Dupont, W.; Papaxanthis, C.; Lebon, F.; Madden-Lombardi, C. Mental Simulations and Action Language Are Impaired in Individuals with Aphantasia. J. Cogn. Neurosci. 2024, 36, 1–11. [CrossRef]
- Bartolomeo, P. The Relationship Between Visual Perception and Visual Mental Imagery: A Reappraisal of the Neuropsychological Evidence. Cortex 2002, 38, 357–378. [CrossRef]
- Bartolomeo, P.; Hajhajate, D.; Liu, J.; Spagna, A. Assessing the causal role of early visual areas in visual mental imagery. Nat. Rev. Neurosci. 2020, 21, 517–517. [CrossRef]
- Bartolomeo, P. Visual agnosia and imagery after Lissauer. Brain 2021, 144, 2557–2559. [CrossRef]
- Bartolomeo, P.; Bachoud-Lévi, A.-C.; De Gelder, B.; Denes, G.; Barba, G.D.; Brugières, P.; Degos, J.-D. Multiple-domain dissociation between impaired visual perception and preserved mental imagery in a patient with bilateral extrastriate lesions. Neuropsychologia 1998, 36, 239–249. [CrossRef]
- Bartolomeo, P. The neural correlates of visual mental imagery: An ongoing debate. Cortex 2008, 44, 107–108. [CrossRef]
- Moro, V.; Berlucchi, G.; Lerch, J.; Tomaiuolo, F.; Aglioti, S.M. Selective deficit of mental visual imagery with intact primary visual cortex and visual perception. Cortex 2008, 44, 109–118. [CrossRef]
- Riley, S.N.; Davies, J. Vividness as the similarity between generated imagery and an internal model. Brain Cogn. 2023, 169, 105988. [CrossRef]
- Arnold, D.H.; Andresen, I.; Anderson, N.; Saurels, B.W. Commonalities between the Berger Rhythm and spectra differences driven by cross-modal attention and imagination. Conscious. Cogn. 2023, 107, 103436. [CrossRef]
- Zeman, A.Z.; Della Sala, S.; Torrens, L.A.; Gountouna, V.-E.; McGonigle, D.J.; Logie, R.H. Loss of imagery phenomenology with intact visuo-spatial task performance: A case of ‘blind imagination’. Neuropsychologia 2010, 48, 145–155. [CrossRef]
- Hajhajate, D.; Kaufmann, B.C.; Liu, J.; Siuda-Krzywicka, K.; Bartolomeo, P. The connectional anatomy of visual mental imagery: evidence from a patient with left occipito-temporal damage. Anat. Embryol. 2022, 227, 3075–3083. [CrossRef]
- Spagna, A.; Heidenry, Z.; Miselevich, M.; Lambert, C.; Eisenstadt, B.E.; Tremblay, L.; Liu, Z.; Liu, J.; Bartolomeo, P. Visual mental imagery: Evidence for a heterarchical neural architecture. Phys. Life Rev. 2024, 48, 113–131. [CrossRef]
- Liu, J., Zhan, M., Hajhajate, D., Spagna, A., Dehaene, S., Cohen, L., & Bartolomeo, P. (2023). Ultra-high field fMRI of visual mental imagery in typical imagers and aphantasic individuals. BioRxiv, 2023-06.
- Boccia, M.; Sulpizio, V.; Bencivenga, F.; Guariglia, C.; Galati, G. Neural representations underlying mental imagery as unveiled by representation similarity analysis. Anat. Embryol. 2021, 226, 1511–1531. [CrossRef]
- McCarthy, G.; Puce, A.; Gore, J.C.; Allison, T. Face-Specific Processing in the Human Fusiform Gyrus. J. Cogn. Neurosci. 1997, 9, 605–610. [CrossRef]
- Pearson, J. Reply to: Assessing the causal role of early visual areas in visual mental imagery. Nat. Rev. Neurosci. 2020, 21, 517–518. [CrossRef]
- Kosslyn, S.M.; Pascual-Leone, A.; Felician, O.; Camposano, S.; Keenan, J.P.; L., W.; Thompson, W.L.; Ganis, G.; Sukel, K.E.; Alpert, N.M. The Role of Area 17 in Visual Imagery: Convergent Evidence from PET and rTMS. Science 1999, 284, 167–170. [CrossRef]
- Bergmann, J.; Genç, E.; Kohler, A.; Singer, W.; Pearson, J. Smaller Primary Visual Cortex Is Associated with Stronger, but Less Precise Mental Imagery. Cereb. Cortex 2016, 26, 3838–3850. [CrossRef]
- Keogh, R.; Bergmann, J.; Pearson, J. Cortical excitability controls the strength of mental imagery. eLife 2020, 9. [CrossRef]
- Meng, M., Chang, S., Zhang, X., & Pearson, J. (2023). Imageless imagery in aphantasia: decoding non-sensory imagery in aphantasia. [CrossRef]
- Cabbai, G., Racey, C., Simner, J., Dance, C., Ward, J., & Forster, S. (2024). Sensory representations in primary visual cortex are not sufficient for subjective imagery. bioRxiv, 2024-01. [CrossRef]
- Arcangeli, M. Aphantasia demystified. Synthese 2023, 201, 1–20. [CrossRef]
- Monzel, M.; Handlogten, J.; Reuter, M. No verbal overshadowing in aphantasia: The role of visual imagery for the verbal overshadowing effect. Cognition 2024, 245, 105732. [CrossRef]
- Park, T. Why successful performance in imagery tasks does not require the manipulation of mental imagery. AVANT. J. Philos. Vanguard 2019, 10. [CrossRef]
- Toftness, A. R. (2022). Clarifying aphantasia (Doctoral dissertation, Iowa State University).
- Purkart, R., Delem, M., Cavalli, E., Ranson, V., Andrey, C., Versace, R., & Plancher, G. Are There Unconscious Visual Images in Aphantasia? Development of an Implicit Priming Paradigm. https://ssrn.com/abstract=4755823.
- Perrin, D., & Michaelian, K. (2017). Memory as mental time travel. The Routledge handbook of philosophy of memory, 228-239.
- Ungerleider, L. G., & Haxby, J. V. (1994). ‘What’and ‘where’in the human brain. Current opinion in neurobiology, 4(2), 157-165. [CrossRef]
- Goodale, M. A., & Milner, A. D. (1992). Separate visual pathways for perception and action. Trends in neurosciences, 15(1), 20-25. [CrossRef]
- Milner, A.; Goodale, M. Two visual systems re-viewed. Neuropsychologia 2008, 46, 774–785. [CrossRef]
- de Haan, E. H. F., Jackson, S. R., & Schenk, T. (2018). Where are we now with ‘What’ and ‘How’? Cortex, 98, 1–7. [CrossRef]
- Grünbaum, T. The two visual systems hypothesis and contrastive underdetermination. Synthese 2021, 198, 4045–4068. [CrossRef]
- Bergmann, J.; Ortiz-Tudela, J. Feedback signals in visual cortex during episodic and schematic memory retrieval and their potential implications for aphantasia. Neurosci. Biobehav. Rev. 2023, 152, 105335. [CrossRef]
- Suica, Z.; Behrendt, F.; Gäumann, S.; Gerth, U.; Schmidt-Trucksäss, A.; Ettlin, T.; Schuster-Amft, C. Imagery ability assessments: a cross-disciplinary systematic review and quality evaluation of psychometric properties. BMC Med. 2022, 20, 1–124. [CrossRef]
- Schwarzkopf, D.S. What is the true range of mental imagery?. Cortex 2024, 170, 21–25. [CrossRef]
- Larner, A.; Leff, A.; Nachev, P. Phantasia, aphantasia, and hyperphantasia: Empirical data and conceptual considerations. Neurosci. Biobehav. Rev. 2024, 164, 105819. [CrossRef]
- Monzel, M.; Mitchell, D.; Macpherson, F.; Pearson, J.; Zeman, A. Proposal for a consistent definition of aphantasia and hyperphantasia: A response to Lambert and Sibley (2022) and Simner and Dance (2022). Cortex 2022, 152, 74–76. [CrossRef]
- Lambert, A.J.; Sibley, C.G. On the importance of consistent terminology for describing sensory imagery and its absence: A response to Monzel et al. (2022). Cortex 2022, 152, 153–156. [CrossRef]
- Simner, J.; Dance, C. Dysikonesia or aphantasia? Understanding the impact and history of names. A reply to Monzel et al. (2022). Cortex 2022, 153, 220–223. [CrossRef]
- D’aNgiulli, A. Vividness, Consciousness and Mental Imagery: A Start on Connecting the Dots. Brain Sci. 2020, 10, 500. [CrossRef]
- Küssner, M.B.; Taruffi, L. Modalities and causal routes in music-induced mental imagery. Trends Cogn. Sci. 2023, 27, 114–115. [CrossRef]
- Lau, H.; Michel, M.; LeDoux, J.E.; Fleming, S.M. The mnemonic basis of subjective experience. Nat. Rev. Psychol. 2022, 1, 479–488. [CrossRef]
- Muraki, E.J.; Speed, L.J.; Pexman, P.M. Insights into embodied cognition and mental imagery from aphantasia. Nat. Rev. Psychol. 2023, 2, 591–605. [CrossRef]
- Cohen, M.X. Where Does EEG Come From and What Does It Mean?. Trends Neurosci. 2017, 40, 208–218. [CrossRef]
- Baillet, S. Magnetoencephalography for brain electrophysiology and imaging. Nat. Neurosci. 2017, 20, 327–339. [CrossRef]
- Xie, S., Kaiser, D., & Cichy, R. M. (2020). Visual imagery and perception share neural representations in the alpha frequency band. Current Biology, 30(13), 2621–2627. [CrossRef]
- Cichy, R. M., & Oliva, A. (2020). A M/EEG-fMRI fusion primer: resolving human brain responses in space and time. Neuron, 107(5), 772-781. [CrossRef]
- Kriegeskorte, N.; Mur, M.; Bandettini, P. Representational similarity analysis – connecting the branches of systems neuroscience. Front. Syst. Neurosci. 2008, 2, 4–4. [CrossRef]
- Haynes, J.-D. A Primer on Pattern-Based Approaches to fMRI: Principles, Pitfalls, and Perspectives. Neuron 2015, 87, 257–270. [CrossRef]
- Polanía, R.; Nitsche, M.A.; Ruff, C.C. Studying and modifying brain function with non-invasive brain stimulation. Nat. Neurosci. 2018, 21, 174–187. [CrossRef]
- Monzel, M.; Karneboge, J.; Reuter, M. The role of dopamine in visual imagery—An experimental pharmacological study. J. Neurosci. Res. 2024, 102. [CrossRef]
- Fried, E.I. Theories and Models: What They Are, What They Are for, and What They Are About. Psychol. Inq. 2020, 31, 336–344. [CrossRef]
- Gigerenzer, G. Personal Reflections on Theory and Psychology. Theory Psychol. 2010, 20, 733–743. [CrossRef]
- Cavedon-Taylor, D. Mental imagery: pulling the plug on perceptualism. Philos. Stud. 2021, 178, 3847–3868. [CrossRef]
- Cavedon-Taylor, D. Untying the knot: imagination, perception and their neural substrates. Synthese 2021, 199, 7203–7230. [CrossRef]
- Cavedon-Taylor, D. (2022b). Predictive processing and perception: What does imagining have to do with it? Consciousness and Cognition, 106, 103419. [CrossRef]
- D’aLoisio-Montilla, N. Imagery and overflow: We see more than we report. Philos. Psychol. 2017, 30, 545–570. [CrossRef]
- Duch, W. Imagery Agnosia and its Phenomenology. Rocz. Psychol. 2021, 24, 307–323. [CrossRef]
- Fox-Muraton, M. Aphantasia and the Language of Imagination: A Wittgensteinian Exploration. Anal. i Egzystencja 2021, 55, 5–24. [CrossRef]
- Fox-Muraton, M. A world without imagination? Consequences of aphantasia for an existential account of self. Hist. Eur. Ideas 2021, 47, 414–428. [CrossRef]
- Lorenzatti, J.J. Aphantasia: a philosophical approach. Philos. Psychol. 2023, 1–29. [CrossRef]
- MacKisack, M.; Aldworth, S.; Macpherson, F.; Onians, J.; Winlove, C.; Zeman, A. Plural Imagination: Diversity in Mind and Making. Art J. 2022, 81, 70–87. [CrossRef]
- Whiteley, C.M.K. Aphantasia, imagination and dreaming. Philos. Stud. 2021, 178, 2111–2132. [CrossRef]
- Open Science Collaboration Estimating the reproducibility of psychological science. Science 2015, 349, aac4716. [CrossRef]
- Tackett, J. L., Brandes, C. M., King, K. M., & Markon, K. E. (2019). Psychology's replication crisis and clinical psychological science. Annual Review of Clinical Psychology, 15, 579-604. [CrossRef]
- Nosek, B.A.; Hardwicke, T.E.; Moshontz, H.; Allard, A.; Corker, K.S.; Dreber, A.; Fidler, F.; Hilgard, J.; Struhl, M.K.; Nuijten, M.B.; et al. Replicability, Robustness, and Reproducibility in Psychological Science. Annu. Rev. Psychol. 2022, 73, 719–748. [CrossRef]
- Miłkowski, M.; Hensel, W.M.; Hohol, M. Replicability or reproducibility? On the replication crisis in computational neuroscience and sharing only relevant detail. J. Comput. Neurosci. 2018, 45, 163–172. [CrossRef]
- Balas, B. Mid-level Characteristics of Drawings Made by Observers with Aphantasia. J. Percept. Imaging 2024, 7, 1–8. [CrossRef]
Figure 1.
Flow diagram for illustrating research searching and screening. Adapted from Moher et al. (2009).
Figure 1.
Flow diagram for illustrating research searching and screening. Adapted from Moher et al. (2009).
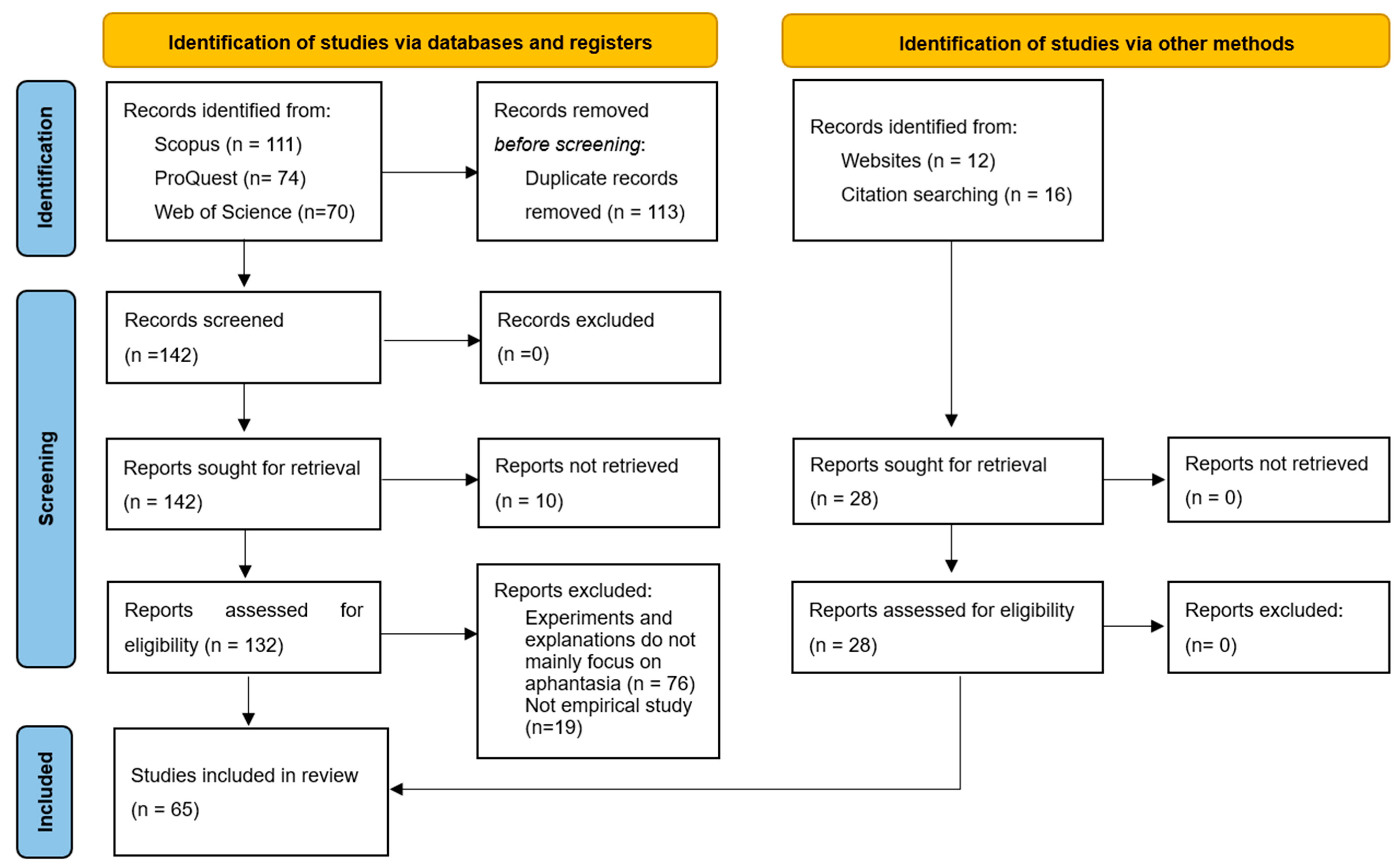

Table 1.
Scores and quality categories of the 65 studies evaluated.
| Study design (0-2) |
Confounding factors (0-2) |
Statistics (0-1) |
Potential bias (0-1) |
Multiplicity (0-1) |
Outcome assessment (0-2) |
Aphantasia assessment (0-2) |
Effect size (0-1) |
Score (absolute) |
Score (%) | Quality category | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Balas, 2024 [165] | 1 | 0 | 1 | 1 | 1 | 2 | 0 | 0 | 6 | 50 | Fair |
| Bainbridge et al., 2021 [17] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Beran et al., 2023 [33] | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 9 | 75 | Good |
| Bumgardner et al., 2021 [44] | 0 | 1 | NA | 1 | 1 | NA | 1 | NA | 4 | 50 | Fair |
| Cabbai et al., 2024 [118] | 1 | 1 | 1 | 1 | 1 | 2 | 1 | NA | 8 | 73 | Good |
| Crowder, 2018 [65] | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 9 | 75 | Good |
| Dance et al., 2023 [90] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Dance et al., 2022 [29] | 0 | 2 | 1 | 1 | 1 | NA | 1 | 1 | 7 | 70 | Good |
| Dance et al., 2021a [48] | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 75 | Good |
| Dance et al., 2021b [87] | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 75 | Good |
| Dando et al., 2023 [59] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Dawes et al., 2020 [6] | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 75 | Good |
| Dawes et al., 2023 [54] | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 7 | 58 | Fair |
| Dawes et al., 2022 [60] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Dupont et al., 2024a [97] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Dupont et al., 2024b [98] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Fielding et al., 2020 [38] | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 8 | 67 | Good |
| Fulford et al., 2018 [14] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | NA | 9 | 82 | Excellent |
| Furman et al., 2022 [66] | 0 | 2 | 1 | 1 | 1 | 2 | 1 | NA | 8 | 73 | Good |
| Gaber et al., 2021 [45] | 0 | 1 | NA | 1 | 1 | NA | 0 | NA | 3 | 38 | Poor |
| Ganczarek et al., 2020 [55] | 1 | 0 | 0 | 1 | 1 | 2 | 1 | 0 | 6 | 50 | Fair |
| Gouveia, 2023 [32] | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 5 | 42 | Fair |
| Gulyas et al., 2022 [34] | 0 | 2 | 1 | 1 | 1 | NA | 1 | 1 | 7 | 70 | Good |
| Hashim et al., 2024 [80] | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 8 | 67 | Good |
| Hinwar & Lambert, 2021 [51] | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 8 | 67 | Good |
| Jacobs et al., 2018 [15] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Kay et al., 2024 [71] | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 9 | 75 | Good |
| Kay et al., 2022 [25] | 1 | 1 | 1 | 1 | 1 | 2 | 2 | NA | 9 | 82 | Excellent |
| Keogh & Pearson, 2024 [23] | 1 | 1 | 1 | 1 | 1 | NA | 2 | 0 | 7 | 70 | Good |
| Keogh & Pearson, 2021 [19] | 1 | 2 | 1 | 1 | 1 | 2 | 2 | 0 | 10 | 83 | Excellent |
| Keogh & Pearson, 2018 [13] | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 0 | 8 | 67 | Good |
| Keogh et al., 2023 [81] | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 7 | 58 | Fair |
| Keogh et al., 2021 [56] | 1 | 2 | 1 | 1 | 1 | 2 | 2 | 1 | 11 | 92 | Excellent |
| Knight & Milton, 2022 [57] | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 9 | 75 | Good |
| Knowles et al., 2021 [42] | 0 | 1 | 1 | 1 | 1 | NA | 1 | 0 | 5 | 50 | Fair |
| Konigsmark et al., 2021 [26] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Liu & Bartolomeo, 2023 [91] | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 9 | 75 | Good |
| Liu et al., 2023 [110] | 1 | 2 | 1 | 1 | 1 | 2 | 2 | NA | 10 | 91 | Excellent |
| Meng et al., 2023 [117] | 1 | 1 | 1 | 1 | 1 | 2 | 1 | NA | 8 | 73 | Good |
| Milton et al., 2021 [16] | 1 | 2 | 1 | 0 | 1 | 2 | 1 | NA | 8 | 73 | Good |
| Monzel et al., 2024 [120] | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 75 | Good |
| Monzel & Reuter, 2024 [74] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Monzel et al., 2023a [79] | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 75 | Good |
| Monzel et al., 2023b [93] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Monzel et al., 2023c [30] | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 75 | Good |
| Monzel et al., 2023e [96] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | NA | 9 | 82 | Excellent |
| Monzel et al., 2022 [61] | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 75 | Good |
| Monzel et al., 2021 [73] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Palermo et al., 2022 [67] | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 0 | 8 | 67 | Good |
| Pounder et al., 2024 [53] | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 75 | Good |
| Pounder et al., 2022 [58] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Purkart et al., n.d. [123] | 1 | 0 | 1 | 1 | 1 | 2 | 1 | 1 | 8 | 67 | Good |
| Reeder, 2022 [27] | 1 | 2 | 1 | 1 | 1 | 1 | 0 | 1 | 8 | 67 | Good |
| Siena & Simons, 2023 [63] | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 9 | 75 | Good |
| Smyth et al, 2019 [37] | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 75 | Good |
| Speed et al., 2024 [77] | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 8 | 67 | Good |
| Takahashi & Gyoba, 2021 [49] | 0 | 1 | 1 | 1 | 1 | 1 | 1 | NA | 6 | 55 | Fair |
| Takahashi et al., 2023 [31] | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 75 | Good |
| Thorudottir et al., 2020 [41] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 0 | 9 | 75 | Good |
| Toftness, 2022 [122] | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 8 | 67 | Good |
| Wicken et al., 2021 [78] | 1 | 2 | 1 | 1 | 1 | 2 | 2 | 1 | 11 | 92 | Excellent |
| Wittmann & Satirer, 2022 [52] | 1 | 2 | 1 | 1 | 1 | 2 | 1 | 1 | 10 | 83 | Excellent |
| Zeman et al., 2020 [4] | 1 | 2 | 1 | 1 | 1 | 0 | 1 | 1 | 8 | 67 | Good |
| Zeman et al., 2015 [5] | 1 | 2 | 1 | 1 | 1 | 0 | 1 | 1 | 8 | 67 | Good |
| Zhao et al., 2020 [70] | 1 | 2 | 1 | 1 | 1 | 2 | 2 | 1 | 11 | 92 | Excellent |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



