
Submitted:
08 July 2024
Posted:
09 July 2024
You are already at the latest version
Abstract
Abstract: RNase Y is an endoribonuclease that governs global mRNA turnover/processing in Ba-cillus subtilis and likely many other bacteria. The enzyme is anchored at the cell membrane, a pseudo-compartmentalization that appears coherent with its task to initiate decay of mRNAs translated predominantly at the cell periphery. However, the reason and consequence for RNase Y attachment to the membrane are largely unknown. We have studied a strain expressing wild-type levels of a cytoplasmic form of RNase Y from the chromosomal locus. This strain exhibits a slow growth phenotype similar to an RNase Y null mutant. Genome-wide data show a significant effect on the expression of hundreds of genes. Certain RNA substrates clearly require RNase Y membrane attachment but others do not. We found no correlation between mRNA stabilization and the cellular location or function of encoded proteins. The Y-complex, an RNase Y specificity factor, appears to also recognize the cytoplasmic form of the enzyme which is able to restore wild-type levels of the corresponding transcripts. We propose that membrane attachment of RNase Y is required for the functional interaction with many coding and non-coding RNAs, for limiting the cleavage of certain substrates and potentially to avoid unfavorable competition between RNase Y and other ribonucleases like RNase J that have a similar evolutionarily conserved cleavage speci-ficity.
Keywords:
Bacillus subtilis
; RNA degradation
; RNase Y
; membrane localization
1. Introduction
The instability of messenger RNA is fundamental to the control of gene expression in all organisms. It enables bacteria to rapidly adapt to a changing environment, produce the correct amount of a given protein and recycle ribonucleotides for incorporation into new RNA molecules. To maximize the competitivity of bacteria, mRNA degradation must be tightly controlled and the only efficient way to do this is to control the steps initiating mRNA decay. The important role of the key endo-ribonucleases RNase Y (B. subtilis) and RNase E (E. coli) in producing short-lived decay intermediates is now well established and can be summarized as “different enzymes-similar strategies” [1].
In B. subtilis, RNase Y cleaves its substrates with an RNase E-like UA-rich single strand-specific cleavage specificity on preferably 5’ monophosphorylated substrates and its depletion increases global mRNA stability [2]. This RNA decay pathway affects the levels of a majority of transcripts in B. subtilis [3,4,5] and S. pyogenes [6] but has a much more limited effect in S. aureus [7]. A search and identification of RNase Y cleavage sites in B. subtilis and other Gram-positive organisms confirms a preference for UA-rich single-stranded sequences often flanked by secondary structures [2,7,8,9,10,11].
Degradosome-like complexes based on RNase Y have been proposed [12,13] but are generally not stable without cross-linking except maybe under certain stress conditions [14]. Whether other ribonucleases can form meaningful interactions with RNase Y in vivo is still an open question [1,15,16,17]. At present, in B. subtilis, the most significant effect on RNase Y activity in vivo appears to be mediated by three small proteins, YaaT, YlbF and YmcA [18]. They can stably bind to each other, forming the so-called Y-complex [19,20] which is required for the efficient maturation of many operon mRNAs and determines the abundance of certain riboswitch RNAs [9,21]. Since it does not affect all RNase Y targets, the Y-complex can be considered as a specificity factor for this globally acting endoribonuclease [9].
RNase Y is tethered to the inner side of the membrane by a single-pass N-terminal helix ([22], named YmdA in this reference). This pseudo-compartmentalization, a feature shared with E. coli RNase E [23,24] is coherent with the predominant distribution of translating ribosomes along the cell periphery [25,26]. Available data provide some insight on the importance of membrane tethering of enzymes initiating RNA decay. Detachment of RNase E from the inner membrane in E. coli results in a global slow-down of RNA degradation and an increased turn-over of ribosome-free transcripts [27]. In S. aureus, where RNase Y plays only a minor role in initiating RNA decay compared to B. subtilis, release of the enzyme from the membrane slows growth but does not significantly alter its activity profile [7].
In B. subtilis, RNase Y moves rapidly along the membrane in the form of dynamic short-lived foci. Upon transcription arrest, these foci become more abundant and increase in size, suggesting they do not represent the most active form of the enzyme [21]. This contrasts with a similar formation of foci by RNase E in E. coli which is dependent on the presence of RNA substrates [24]. The YaaT component of the Y-complex also localizes to the cell periphery [28] in a manner that depends on the presence of RNase Y [9]. Y-complex mutations actually have an even stronger effect than that of RNA depletion in increasing the size and number of RNase Y foci at the membrane. This suggested that the Y-complex could modify RNase Y activity by shifting the assembly status of the enzyme towards more active smaller membrane complexes [21].
Here, we analyse the importance of the attachment of RNase Y to the membrane by characterizing effects of a rny∆TMD mutation on RNA degradation. This allele expresses an RNase Y mutant lacking the N-terminal trans-membrane domain at a physiological level from the wild-type chromosomal locus. This cytoplasmic version of the enzyme which is uniformly distributed in the cell leads to slower growth and extensive modification of the levels of hundreds of transcripts. We highlight a variety of RNAs that require tethering of RNase Y to the membrane and others that do not. Our observations indicate that the subcellular localization of wild-type RNase Y does not correlate with its activity towards a certain class or type of RNA substrates. We discuss the functional consequences and rational for attachment of RNase Y to the membrane.
2. Results
2.1. Experimental Strategy
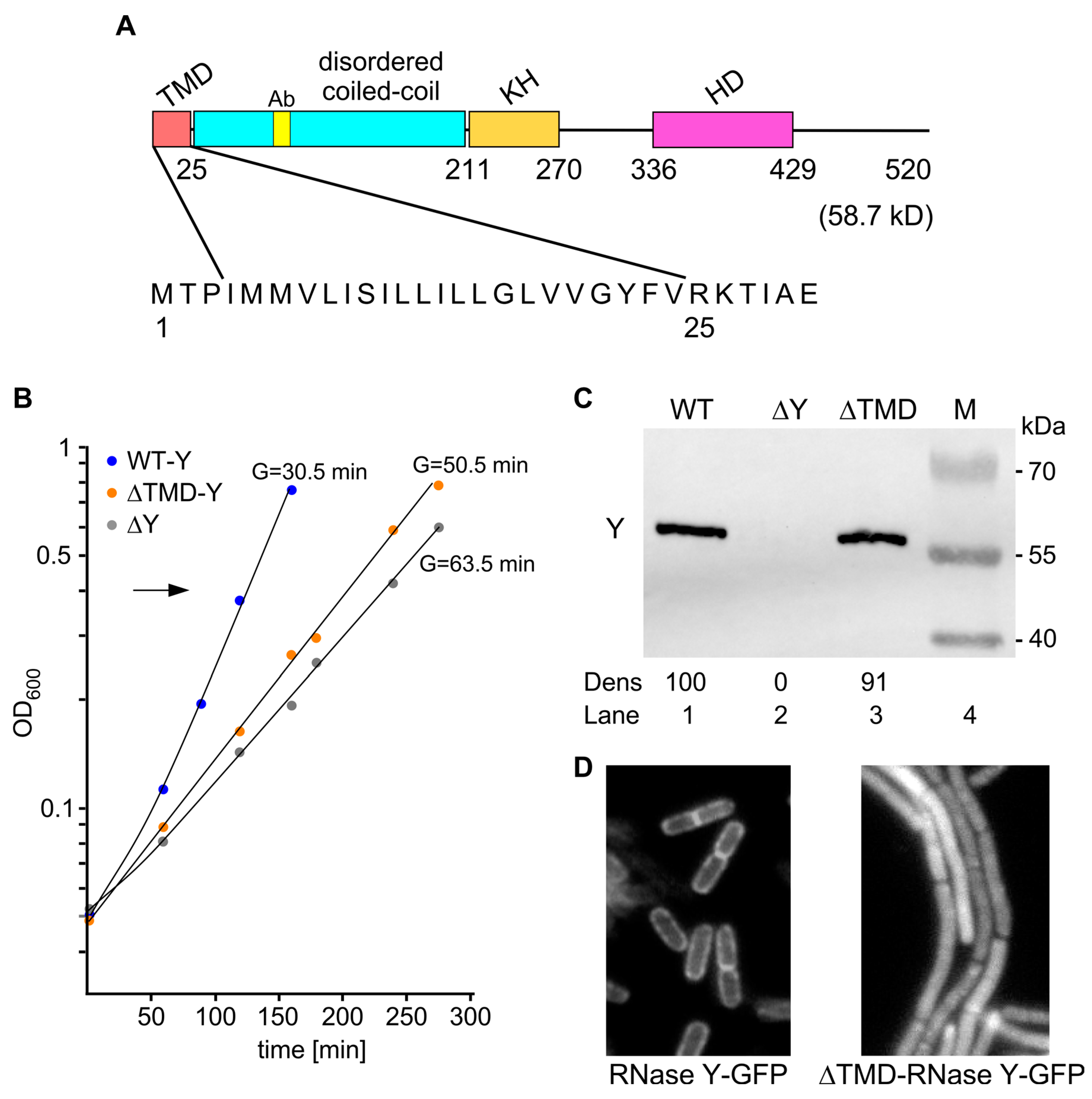
In order to study how the membrane localization of RNase Y might affect its biological activity in vivo we constructed a strain expressing a cytoplasmic form of the enzyme. B. subtilis RNase Y contains a stretch of 21 predominantly hydrophobic residues beginning at residue 4 (Figure 1A). There are no charged residues at the N terminus, but the hydrophobic region is immediately followed by two basic residues (R, K). According to the positive inside rule of von Heinje (1992), these features are strongly suggestive of a single transmembrane region with the C-terminal part of the protein located in the cytosol. In agreement, an RNase Y-GFP fusion protein lacking the aforementioned 21 amino acids (∆TMD) clearly localizes to the cytoplasm, in contrast to wild-type RNase Y-GFP (Figure 1D).
In complex medium (LB), a strain lacking RNase Y (∆rny, SSB508) grew with a > 2-fold longer generation time compared to the wild-type strain, 63.5 min versus 30.5 min, respectively. A similar growth defect was observed in a strain expressing exclusively the cytoplasmic form of RNase Y (Figure 1B). In this strain, ∆TMD-RNase Y was present at a level equivalent to that observed for wild-type RNase Y (Figure 1C) as expected since the 63 nt (21 aa) deletion was introduced in the wild-type rny gene without any additional alteration. Since the enzyme retains all structural domains besides the membrane anchor and is active in this form in vitro [2], the localization of RNase Y itself seems to be important for its biological activity and hence to support efficient growth. The ∆TMD strain grows as chains while wild-type cells grow as single or dividing cells (Figure 3D. Nevertheless, cytoplasmic RNase Y can restore the severe defects in cell morphology of a ∆rny strain that under the same growth conditions forms spirals interspersed with long chains [29].
Figure 1.
Culture conditions and analysis of RNase Y expression. (A). Domains composing B. subtilis RNase Y (520 aa) include an N-terminal transmembrane domain (TMD, aa 1–25), followed by a large region predicted to be disordered (aa ~30–210), an RNA binding KH domain (aa 211–270), and a metal-chelating HD domain (aa 336–429) containing the conserved His/Asp motif required for RNase activity [2,22,30,31,32]. Ab indicates the position of the 12 aa peptide used for monoclonal antibody production. The hydrophobic 21 amino acid stretch composing the RNase Y transmembrane region is shown below the domain structure scheme. The cytoplasmic version of RNase Y (∆TMD-RNase Y) expressed in strain SSB574 lacks aa 2 to 24. (B). Growth curves in LB medium at 37°C. Strains expressing wild-type RNase Y (SSB507), cytoplasmic ∆TMD-RNase Y (SSB574) or no RNase Y (SSB508) were harvested at OD600 ~0.4 (arrow). (C). Expression of RNase Y and ∆TMD-RNase Y as well as the absence of RNase Y in the different strains was analyzed by Western blot. (D) ∆TMD-RNase Y localizes in the cytoplasm. Wild-type RNase Y-sfGFP (SSB2048) and ∆TMD-RNase Y-sfGFP (SSB2066) expression was analyzed by epifluorescence microscopy in cells grown to mid-log phase.
Figure 1.
Culture conditions and analysis of RNase Y expression. (A). Domains composing B. subtilis RNase Y (520 aa) include an N-terminal transmembrane domain (TMD, aa 1–25), followed by a large region predicted to be disordered (aa ~30–210), an RNA binding KH domain (aa 211–270), and a metal-chelating HD domain (aa 336–429) containing the conserved His/Asp motif required for RNase activity [2,22,30,31,32]. Ab indicates the position of the 12 aa peptide used for monoclonal antibody production. The hydrophobic 21 amino acid stretch composing the RNase Y transmembrane region is shown below the domain structure scheme. The cytoplasmic version of RNase Y (∆TMD-RNase Y) expressed in strain SSB574 lacks aa 2 to 24. (B). Growth curves in LB medium at 37°C. Strains expressing wild-type RNase Y (SSB507), cytoplasmic ∆TMD-RNase Y (SSB574) or no RNase Y (SSB508) were harvested at OD600 ~0.4 (arrow). (C). Expression of RNase Y and ∆TMD-RNase Y as well as the absence of RNase Y in the different strains was analyzed by Western blot. (D) ∆TMD-RNase Y localizes in the cytoplasm. Wild-type RNase Y-sfGFP (SSB2048) and ∆TMD-RNase Y-sfGFP (SSB2066) expression was analyzed by epifluorescence microscopy in cells grown to mid-log phase.
2.2. Cytoplasmic Localization of RNase Y Affects Gene Expression on a Genome-Wide Scale
To better apprehend the role of RNase Y membrane localization on global gene expression, we sequenced and compared the transcriptomes of the wild-type, an RNase Y null mutant strain (∆rny) and a strain expressing a cytoplasmic version of RNase Y (∆TMD) using stranded RNA-Seq. A total of ~145 million raw sequence reads was obtained from the analysis of 9 samples, with an average per sample of ~16 million reads (Table S1) mapped to the B. subtilis genome. The absence of reads corresponding to the entire rny ORF or the 63 nt of the TMD region confirmed the integrity of the ∆rny and ∆TMD strains, respectively (Figure 2A).
Figure 2.
Global effects on the transcriptome in strains expressing ∆TMD-RNase Y or no RNase Y. (A). Browser view of RNA-Seq reads of the rny locus of the wild-type and mutant strains expressing the ∆TMD version of RNase Y or lack the rny ORF. The red box indicates the gene sequences coding for the N-terminal TMD domain. Reads from a single replicate of each strain are shown as representative examples. (B). Multidimensional scaling (MDS) plot of WT, ∆TMD and ∆rny triplicate libraries. Distance between samples is based on the leading log2 fold-change (FC) of the top 500 most differentially regulated genes. (C). MA plots of differentially expressed genes (red dots) at FDR ≤ 0.05 identified in the two mutant srains compared to WT.
Figure 2.
Global effects on the transcriptome in strains expressing ∆TMD-RNase Y or no RNase Y. (A). Browser view of RNA-Seq reads of the rny locus of the wild-type and mutant strains expressing the ∆TMD version of RNase Y or lack the rny ORF. The red box indicates the gene sequences coding for the N-terminal TMD domain. Reads from a single replicate of each strain are shown as representative examples. (B). Multidimensional scaling (MDS) plot of WT, ∆TMD and ∆rny triplicate libraries. Distance between samples is based on the leading log2 fold-change (FC) of the top 500 most differentially regulated genes. (C). MA plots of differentially expressed genes (red dots) at FDR ≤ 0.05 identified in the two mutant srains compared to WT.
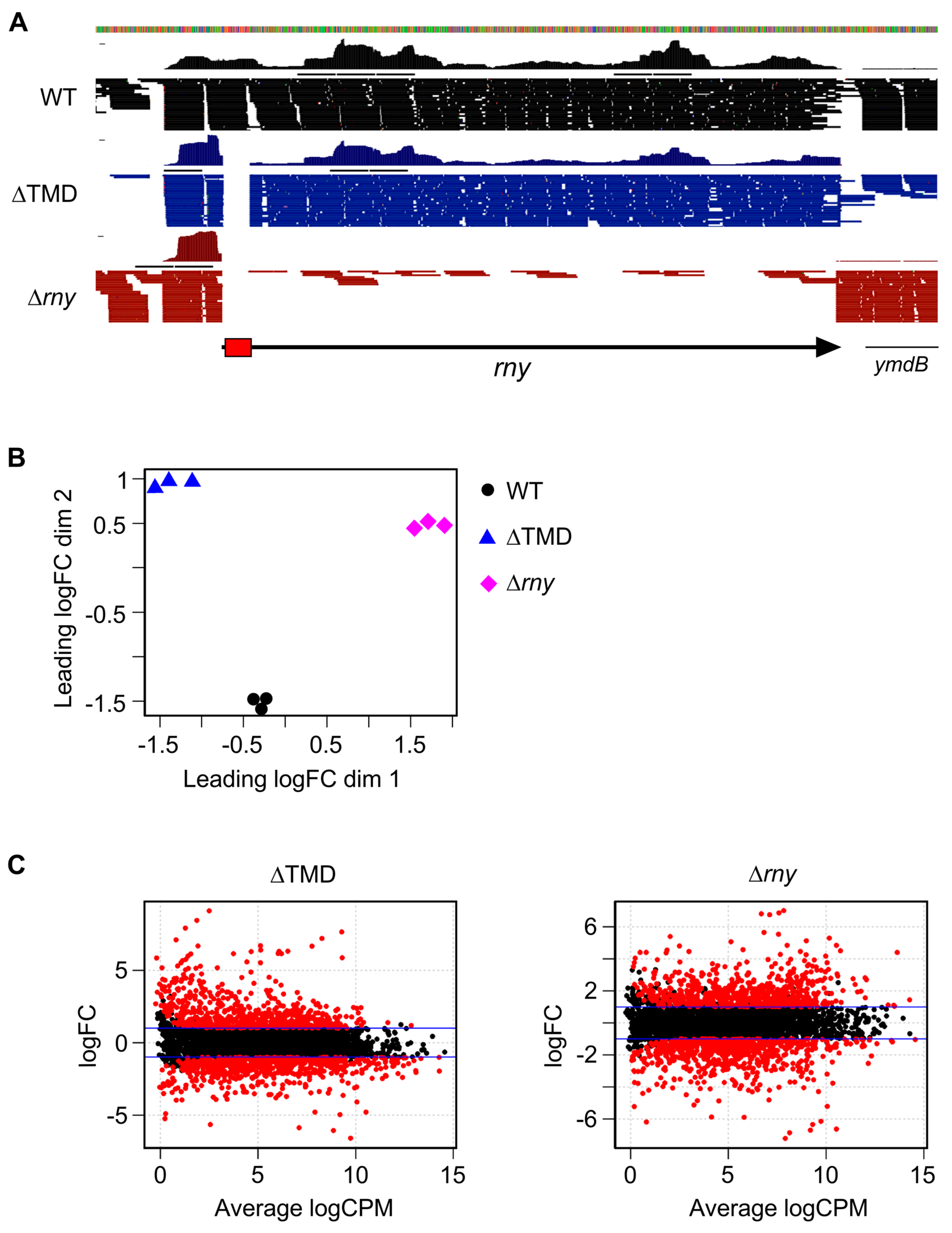
Differential gene expression between all strains was explored by measuring the distance of gene expression values between any pair of samples. The data sets, filtered for poorly or not expressed genes, consisted of 4105 RNA features including open reading frames and untranslated regions (UTRs, riboswitches, small RNAs) derived from the chromosome and retrieved from Subtiwiki [33]. On a multidimensional scaling (MDS) plot (Figure 2B), the triplicates within each group (strain) clustered closely, indicating similar gene expression levels and consistency between biological replicates. By contrast, both the ∆rny and the ∆TMD strains were clearly separated from the WT samples. This suggested that localization to the membrane via the TMD is important for the normal function of RNase Y. These observations are corroborated by the MA plots of the expression data (Figure 2C) which relate the ratio of level counts for each RNA feature between WT and mutant strains against the average level counts for each feature from all libraries. We applied a cutoff of FC ≥ 2 and ≤ 2 at a FDR ≤ 0.05 to call for differentially expressed gene (DEG) features (red dots above and below the blue line in Figure 2C) and we categorized them into functional groups according to Gene Ontology (GO) annotation (Supplementary Tables S2 and S3).
Compared to WT, the ∆rny and the ∆TMD-RNase Y expressing strains had similar numbers of DEG’s (Table 1): 712 and 665 upregulated features and 751 and 652 downregulated features, respectively. An increased mRNA level in the absence of a given RNase is often an indication of direct action of the enzyme on the substrate RNA while down-regulated RNAs are generally caused by indirect effects as observed in most RNase related transcriptome studies. Expression of cytoplasmic ∆TMD-RNase Y might be expected to increase cleavage of natural RNase Y substrates. Indeed, 72 RNAs that are up-regulated in the ∆rny mutant have significantly lower levels in the ∆TMD strain compared to the WT strain (Table S3). However, the majority of the down-regulated RNAs in the ∆TMD strain (462 out of 652) are likely no genuine RNase Y substrates because they are also present at lower levels in the ∆rny mutant. In both mutant strains most of the up-regulated genes belonged to the GO categories « undefined », « unknown » and « general function prediction », 41 % (293 in total) and 42.6 % (292 in total) in the ∆rny and ∆TMD-RNase Y strains, respectively (Table S2). There were several functional categories where significantly more genes were induced in the ∆TMD strain compared to the ∆rny strain : “Carbohydrate transport and metabolism” (53 vs 24 transcripts), “Lipid transport and metabolism” (27 vs 14 mRNAs) and “Energy production and conversion” (24 vs 13 mRNAs). About 30% of the transcripts with significantly increased levels in the ∆TMD strain (203) compared to a WT strain were also induced in the ∆rny knock-out strain. Of those, 45 mRNAs are prophage related (SBbeta and PBSX) and together with the remaining 158 transcripts are good candidates for RNase Y substrates that require localization of the enzyme at the membrane for efficient cleavage (Table S4). An analysis of 55 riboswitch, 5’ leader sequences and other non-coding RNAs showed that more than half (29) have higher levels in the ∆rny strain and 11 are also induced in the presence of cytoplasmic RNase Y (Table S5). There is no general pattern as to which type of non-coding RNA is an RNase Y substrate and/or requires membrane localization of the enzyme; e.g., T-box or guanine riboswitches can be found in all categories (Table S5).
The detection of transcripts that are induced only in the ∆TMD strain but not in the ∆rny strain is intriguing (Table S6). One possibility could be that cytoplasmic RNase Y masks cleavage sites prone to be cleaved by other endoribonucleases (see below).
2.3. Modulation of Specific Transcript Levels by Cytoplasmic RNase Y
We performed Northern blots of specific transcripts to confirm some of the RNA-Seq data and to analyze more precisely if and how cytoplasmic RNase Y can alter transcription patterns. The RNA substrates were devided into two categories, one that requires and one that is insensitive to membrane localization of RNase Y.
2.3.1. RNA Substrates that Require Membrane Localization of RNase Y
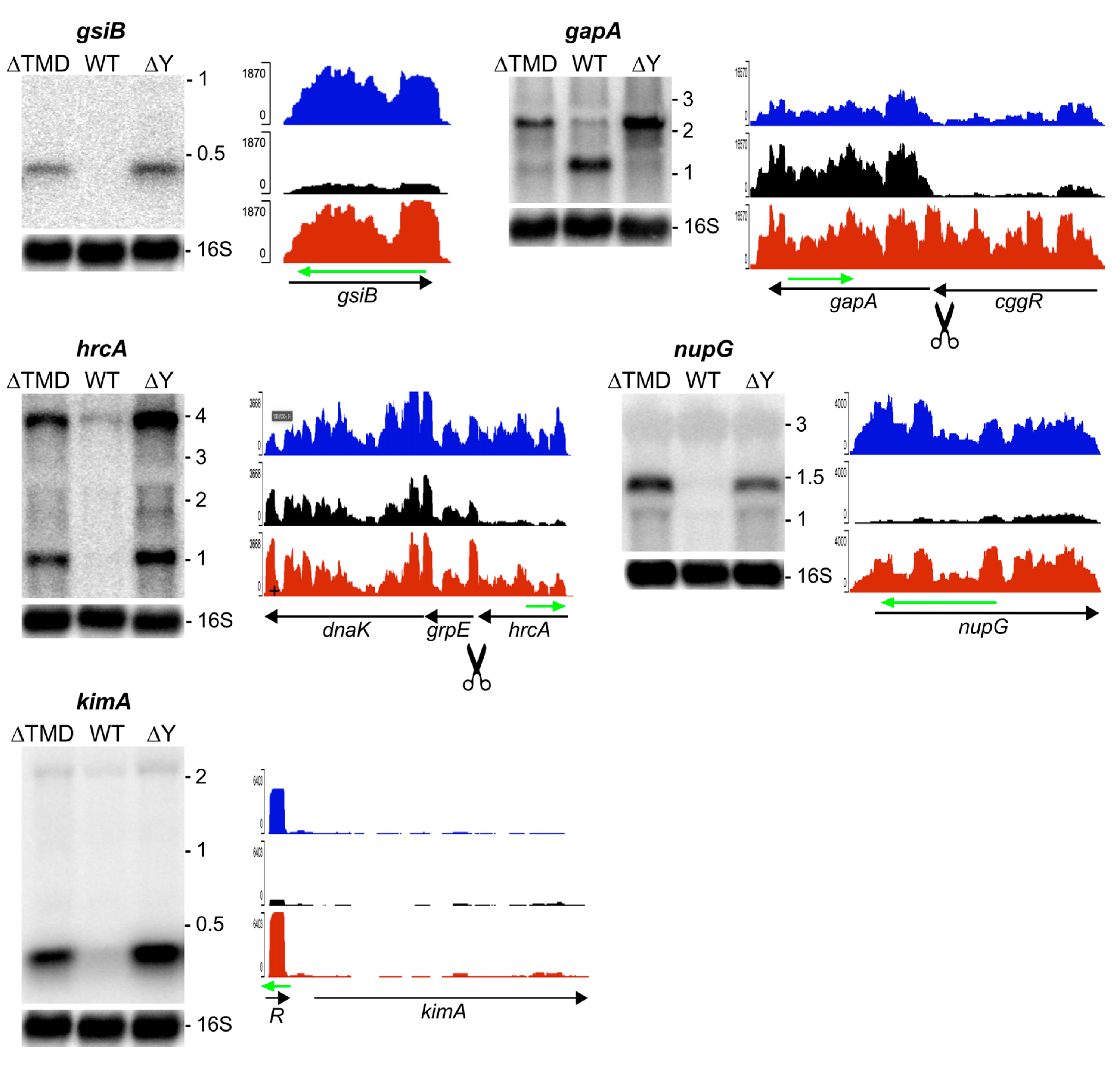
Transcripts that require RNase Y to be anchored at the membrane for cleavage should be up-regulated not only in the ∆rny mutant but also in a strain expressing the cytoplasmic version of the nuclease. In Figure 3, several examples of transcripts that have elevated levels in both mutant strains compared to a wild-type strain are shown.
The gsiB gene encodes a small general stress protein. The monocistronic 0.4 kb transcript is transcribed from a σB promoter under various stress conditions like glucose starvation or heat shock [34]. The very low gsiB mRNA level present in exponentially growing WT cultures used here is strongly increased in the absence of RNase Y and to almost the same level in the strain expressing cytoplasmic RNase Y (see gsiB in Figure 3). This indicates that cytoplasmic ∆TMD-RNase Y cannot replace the membrane tethered wild-type enzyme to act on the gsiB transcript.
A regulatory processing by RNase Y in the cggR mRNA 3’ end uncouples gapA and cggR expression [12]. In the absence of RNase Y, the 2.2 kb gapA-cggR transcript accumulates (Figure 3). As judged by the ratio of reads corresponding to the cggR and gapA ORFs, some cleavage still occurs in the ∆TMD strain but cytoplasmic RNase Y is clearly less efficient in this processing event (Figure 3, gapA).
A cleavage with similar outcome occurs in the intergenic region between hrcA and grpE. It uncouples expression of the transcriptional repressor HrcA from the other components of the dnaK heat shock operon that it controls [35]. As expected for cultures grown at 37°C, the transcript levels from this operon are very low in the wild-type strain. However, the levels of both the monocistronic hrcA and the tricistronic hrcA-grpE-dnaK transcripts are strongly increased in the RNase Y knock-out strain (Figure 3, hcrA) highlighting the importance of the mRNA processing step in the DnaK mediated heat shock response. Localization of RNase Y at the membrane was critically important as a strain expressing the enzymatically active ∆TMD-RNase Y had the same elevated transcript pattern as the knock-out mutant (Figure 3, hrcA).
Another example is the nupG mRNA encoding a purine nucleoside transporter (Figure 3, nupG). Transcription is under control of a guanine dependent riboswitch (G-box) [36] which is strongly induced in both the ∆rny and the ∆TMD strains (Table S5). Whether cleavage of the riboswitch is sufficient to maintain nupG mRNA wild-type levels or also requires a cleavage by membrane bound RNase Y within the nupG ORF is unknown.
RNase Y is known to initiate the degradation of many riboswitches [2,5] and here we found that several of those sensitive to RNase Y also require membrane localization of the enzyme. This is the case for the cyclic di-AMP riboswitch that controls the expression of the high-affinity K+ transporter KimA (Figure 3, [37,38]. In contrast, the second cyclic di-AMP riboswitch upstream of the ktrA gene does not appear to be an RNase Y substrate. Again, there is no rule as to which class of riboswitches is susceptible to RNase Y cleavage and/or a requirement for its localization at the membrane (confer Table S5).
Figure 3.
Examples of coding and non-coding RNAs requiring RNase Y membrane attachment for maintaining wild-type transcript patterns. The left panels show a Northern analysis of total RNA isolated from WT, ∆TMD and ∆rny strains. The blots were hybridized to specific riboprobes complementary to the regions indicated by a green arrow above the indicated gene. The RNAseq profiles (right panels) for the ∆TMD (blue), WT (black) and ∆Y (red) strains are at the same scale for a given gene. When known, the identified RNase Y cleavage site is indicated by a scissors symbol.
Figure 3.
Examples of coding and non-coding RNAs requiring RNase Y membrane attachment for maintaining wild-type transcript patterns. The left panels show a Northern analysis of total RNA isolated from WT, ∆TMD and ∆rny strains. The blots were hybridized to specific riboprobes complementary to the regions indicated by a green arrow above the indicated gene. The RNAseq profiles (right panels) for the ∆TMD (blue), WT (black) and ∆Y (red) strains are at the same scale for a given gene. When known, the identified RNase Y cleavage site is indicated by a scissors symbol.
2.3.2. RNA Substrates Not Sensitive to RNase Y Localization
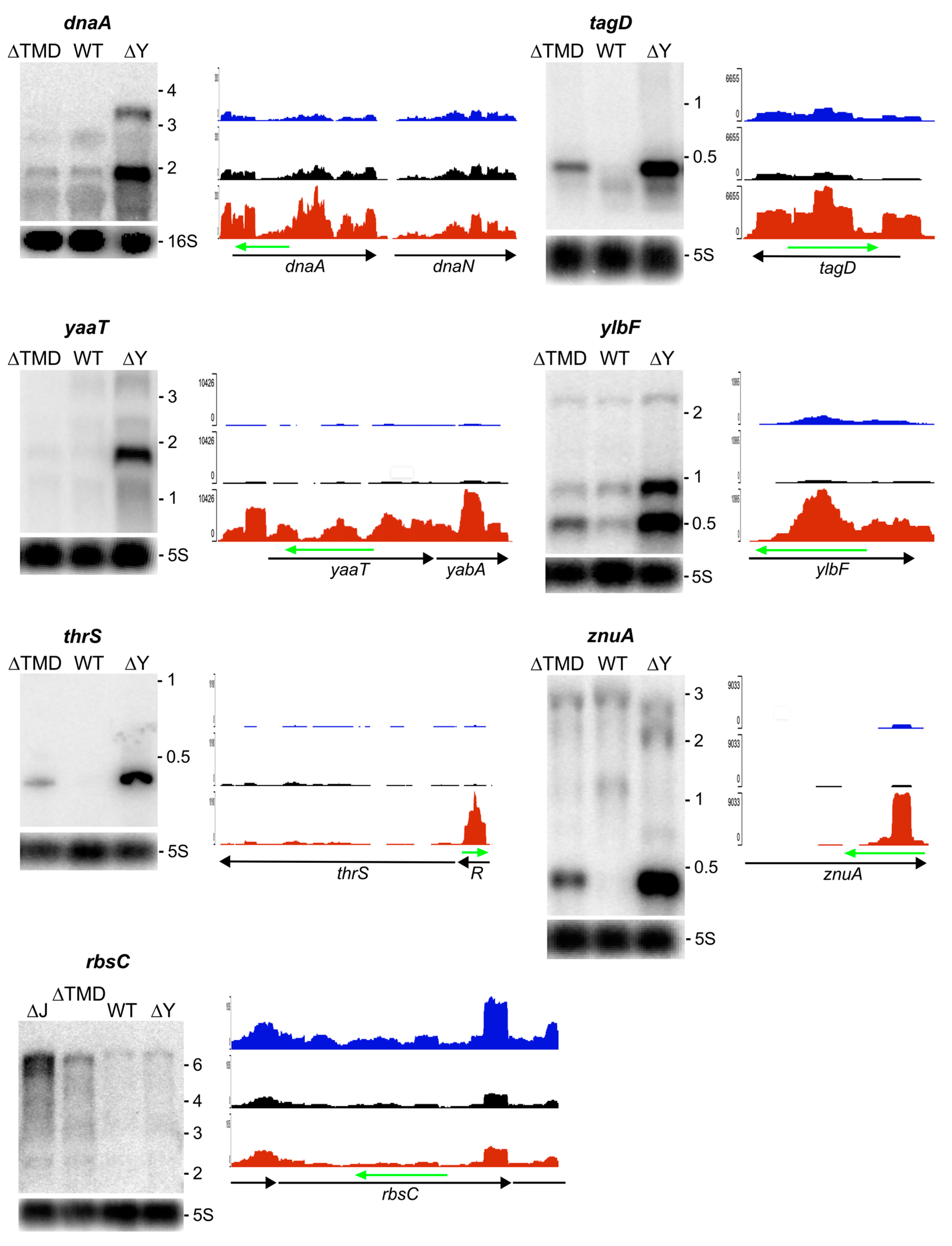
Transcripts that do not require RNase Y to be tethered to membrane for cleavage should be present at the same or a similar level in the ∆TMD strain expressing cytoplasmic RNase Y when compared to the wild-type strain. Roughly 60-70 % of the transcripts increased in the ∆rny mutant fall into this category; many of these transcripts show actually lower levels in the ∆TMD strain suggesting a more efficient cleavage (Table S3). We have analyzed several examples by Northern blot (Figure 4), some of which have previously been shown to be a direct target of RNase Y [5]. Expression of the entire tagD and the divergent tagA operon as well as the tagO gene, all of which are crucial for cell wall synthesis, is strongly induced in the ∆rny strain (Table S3). All of these transcripts are present close to wild-type levels in the ∆TMD strain. As shown for tagD, the 0.4 kb mRNA is strongly reduced compared to the ∆rny strain but remains slightly more abundant than in the wid-type strain (Figure 4).
The essential dnaA-dnaN operon encoding the replication initiator protein (DnaA) and the β-subunit of DNA polymerase III (the beta clamp, DnaN) is transcribed into mono- and bicistronic mRNAs (~1.6 and ~3 kb) that are strongly stabilized in the absence of RNase Y [5], Figure 4). In this case, cytoplasmic ∆TMD-RNase Y is able to maintain exactly the transcript pattern and levels observed in the wild-type strain (Figure 4, dnaA).
The so-called Y-complex composed of three small proteins YaaT, YlbF and YmcA [20] was shown to bind to and modify RNase Y activity and assembly status in vivo [9,18,21]. The transcript levels of all three genes are up-regulated in the ∆rny strain (Figure 4, yaaT, ylbF and Table S3) as observed previously [5,9]. Since individual Y-complex mutations alone affect the transcript levels of the remaining Y-complex proteins [9] it is likely that the Y-complex autoregulates its expression through its interaction with RNase Y. As shown in Figure 4, RNase Y does not need to be at the membrane to maintain wild-type levels of the yaaT and ylbF transcripts (see also discussion).
As mentionned above, many riboswitches and non-coding RNAs do not require RNase Y to be at the membrane to initiate their degradation. One example is the T-box riboswitch controlling threonyl-tRNA synthetase (thrS) expression. The prematurely terminated riboswitch RNA (~0.3 kb) is present only at very low levels in the ∆TMD strain, similar to the wild-type strain (Figure 4, thrS).
A peculiar non-coding RNA is transcribed from an internal sigma A promoter in the znuA (adcA) gene encoding a zinc-binding lipo-protein [39]. Comprised of the 400 3’ proximal nucleotides, it is only detected in the absence of RNase Y. Again, cytoplasmic RNase Y can efficiently replace the wild-type enzyme (Figure 4, znuA).
There are a number transcripts that are only induced in the presence of cytoplasmic RNase Y but not in the absence of the enzyme (Table S6). There is no straight forward explanation for this observation. We have previously shown that RNase Y and RNase J1/J2 (and also E. coli RNase E) can cleave the thrS leader sequence at the same site [29,40] suggesting a convergent evolution toward a common enzymatic activity. However, cleavage efficiency can be very different in vitro and in vivo. For example, RNase J1 cleaves the thrS leader much better in vitro than RNase Y, yet in vivo cleavage is mediated almost exclusively by RNase Y [29]. Since both enzymes RNase Y and RNase J can obviously recognize but not necessarily cleave with the same efficiency in a given sequence context, normal cleavage patterns could be perturbed by the increased concentration of one or the other enzyme. Expression of ∆TMD-RNase Y even at wild-type levels increases the local concentration of RNase Y in the cytoplasm and could lead to increased binding and masking of cleavage sites on RNAs that are normally a substrate for RNase J. An example for such a configuration is the rbsC transcript encoding a ribose ABC transporter [41]. The level of the major polycistronic ribose operon transcript including rbsC (~6.3 kb) is increased in the ∆TMD but not the ∆rny strain (Figure 4, rbsC). If cytoplasmic RNase Y acts by masking a potential RNase J cleavage site in this mRNA, the transcript level should be equal or higher in a RNase J mutant. This is exactly what we observed in the RNase J1/J2 double mutant (Figure 4, rbsC).
Figure 4.
Examples of coding and non-coding RNAs sensitive to both the membrane attached and the cytoplasmic form of RNase Y. The left panels show a Northern analysis of total RNA isolated from WT, ∆TMD and ∆rny strains. The blots were hybridized to specific riboprobes complementary to the regions indicated by a green arrow above the indicated gene. The RNAseq profiles (right panels) for the ∆TMD (blue), WT (black) and ∆Y (red) strains are at the same scale for a given gene. For the rbsC gene, the Northern analysis also includes RNA isolated from the ∆rnjA/∆rnjB double mutant expressing no RNase J.
Figure 4.
Examples of coding and non-coding RNAs sensitive to both the membrane attached and the cytoplasmic form of RNase Y. The left panels show a Northern analysis of total RNA isolated from WT, ∆TMD and ∆rny strains. The blots were hybridized to specific riboprobes complementary to the regions indicated by a green arrow above the indicated gene. The RNAseq profiles (right panels) for the ∆TMD (blue), WT (black) and ∆Y (red) strains are at the same scale for a given gene. For the rbsC gene, the Northern analysis also includes RNA isolated from the ∆rnjA/∆rnjB double mutant expressing no RNase J.
3. Discussion
In this study, we have shown that cytoplasmic expression of RNase Y has a profound effect on the physiology and the global transcription pattern in B. subtilis. In LB medium the strain with the rny∆TMD allele grows 1.7 -fold more slowly than the wild-type control which is similar to the 2-fold effect observed for the ∆rny null mutant. The pleiotropic changes induced by the ∆TMD mutation do not allow to pinpoint a single cause for the growth defect. An obvious reason for the observed phenotype might reside in an altered activity of the mutant enzyme. However, we have previously measured comparable enzyme activities in vitro for ∆TMD-RNase Y and the wild-type enzyme bound to multilamellar vesicles prepared from B. subtilis native lipids [2,42]. Moreover, the wild-type and the cytoplasmic versions of RNase Y are both expressed from the chromosomal wild-type locus and present in equivalent amounts in the cell. We therefore favor the hypothesis that differences observed for many potential RNA substrates are not due to the enzyme activity per se but rather to other factors: i) an altered accessibility of the RNA substrate by the cytoplasmic enzyme and ii) a potential difference in the interaction of the two RNase Y forms with auxiliary factors like the Y-complex (see below).
Among the transcripts that were up-regulated in the ∆rny strain and which are potential direct RNase Y targets, 203 also had higher levels in the presence of cytoplasmic RNase Y. We concentrated on the 158 transcripts that were not prophage related. A Northern analysis for several of them confirmed that cleavage/maturation of the RNA requires or is strongly stimulated by RNase Y tethered to the membrane. We have previously shown that E. coli RNase E can quite efficiently replace RNase Y in B. subtilis and that the single most important parameter to efficient complementation was the requirement for RNase E to localize to the membrane [29]. It is noteworthy that about one third (48) of the 158 transcripts highlighted to depend on RNase Y membrane localization also required RNase E to be localized at the membrane for restoring wild-type levels in a ∆rny mutant. Even though the experimental conditions were similar but not identical in the two studies, this suggests that accessibility of the substrate to RNase Y can be an important factor for the cleavage of a considerable number of RNA substrates. An interesting example is the HrcA repressor protein that controls nine class I heat shock genes organized in two operons. The complex regulation of this regulon which also involves the activation of HrcA by one of the regulated genes [35,43] is not fully understood. The strong up-regulation of the hrcA mRNA in the presence of cytoplasmic RNase Y nevertheless suggests an important role for the membrane localization of RNase Y in the heat shock response of B. subtilis.
The conditioned interaction with partner proteins could be another reason why certain RNA substrates require membrane localization of RNase Y. The Y-complex (YaaT, YlbF and YmcA) that localizes to the membrane in a manner dependent on RNase Y, has previously been shown to act as an auxiliary factor required for the efficient cleavage of many polycistronic mRNAs [9]. It can shift the assembly status of membrane bound higher order structures of RNase Y toward fewer and smaller complexes which are likely the more active form of the enzyme [21]. Referring to the available data from the Y-complex transcriptome study [9] we have checked whether the five transcripts that required membrane localization of RNase Y (Figure 3) might also be sensitive to Y-complex mutations. With the exception of the kimA (ydaO) riboswitch, which was not even sensitive to the absence of RNase Y in that study, the other transcripts all required the Y-complex for maintaining wild-type transcript levels. This supports the hypothesis that in some cases RNase Y membrane attachment is important for cleavage because binding of the Y-complex might require the higher order structures of RNase Y found at the membrane. When comparing the data on a larger scale, we found that only about 20 % of transcripts preferentially cleaved by membrane tethered RNase Y are also up-regulated in Y-complex mutant strains. However, one has to consider that the compared data sets were derived from different strains, B. subtilis 168 used in our study and less domesticated B. subtilis 3610 in the Y-complex study. Together with the fact that many of the transcripts were present at very low levels (Tables S3 and S4 in [9]) and that membrane dependent cleavage is rarely an all or nothing effect, this might also explain the limited accordance.
More than two thirds of the transcripts that are up-regulated in the ∆rny strain are maintained at near wild-type levels in the strain expressing cytoplasmic RNase Y. They include a number of essential genes involved in initiation of DNA replication (e.g., dnaA, Figure 4) and biosynthesis of cell wall teichoic acids (e.g., tagABDGH, Figure 4). For several of them RNase Y has been shown to act directly by modulating the half-life of the respective transcripts [5]. Deregulation of these genes has been linked to aberrant cell morphology [44,45]. Restoring their correct expression could explain the almost normal cell shape observed in the ∆TMD strain compared to the RNase Y null mutant.
Interestingly, maintaining wild-type levels of the Y-complex transcripts can also be achieved by cytoplasmic RNase Y. Since the putative autoregulation of the Y-complex proteins likely occurs via RNase Y cleavage of the respective mRNAs, this implies that the Y-complex should also be able to interact with ∆TMD-RNase Y. It is unclear which oligomeric state RNase Y might have in the cell when expressed as a cytoplasmic form of the enzyme. The N-terminal intrinsically unstructured domain of RNase Y by itself (~200 aa) lacking the TMD can form dimers that contribute to the formation of ∆TMD-RNase Y dimeric forms [32]. However, when overexpressed and purified from E. coli, ∆TMD-RNase Y is always present in various amounts of high molecular weight oligomers in addition to the dimeric form [2,32]. Further studies are required to determine whether the function of the Y-complex involves interactions with an RNase Y dimer or requires higher order structures like those present at the membrane [21].
We found no obvious reason or common attributes that could indicate why a given RNA substrate would require RNase Y to be attached to the membrane or not. Nevertheless, we have determined the presence of transmembrane domains for all protein encoding gene products but found no significant link between RNase Y membrane localization and a potential preference for RNA substrates encoding membrane proteins (Tables S3–S6). This is in agreement with a previous study that found no strong bias for localization of membrane protein-encoding transcripts at the cell periphery [46]. In B. subtilis, transcription and translation occur predominantly in separate functional domains with ribosomes distributed at the inner membrane and the cell poles [25,26]. It makes thus good sense that the mRNA degradation machinery is localized at the membrane where it can monitor suboptimal translation and initiate mRNA decay to maintain efficient gene expression. At the same time, unrelated observations from E. coli suggest that localization of the decay initiating RNase at the membrane might not necessarily require a substrate RNA to diffuse to the cell periphery in order to be cleaved. Indeed, an artificially membrane-attached transcriptional antiterminator protein (i.e., E. coli BglG) is capable of interacting fast enough with its chromosomally encoded nascent mRNA target sequence to promote transcriptional read-through [47].
The subcellular organization of the major decay initiating endoribonucleases RNases Y and E in bacteria can be achieved by different strategies like membrane anchoring [21,24] or formation of bacterial ribonucleoprotein bodies (BR-bodies, [48]). This pseudo-compartmentalization is generally advantageous for cell growth, but its importance varies in different organisms. In E. coli, attachment of RNase E to the inner membrane assures optimal rates of global mRNA degradation and protects ribosome-free transcripts from increased turnover [27]. In S. aureus, detachment of RNase Y from the membrane slows growth but does not alter the number and cleavage profiles of the roughly hundred RNase Y substrates identified in this organism [7]. In B. subtilis, where RNase Y has a much more important role in global mRNA turnover [5], cytoplasmic expression of RNase Y influenced the expression of hundreds of genes. These include three types of transcripts: i) those that require membrane localization of RNase Y ii) those that do not, including RNAs downregulated compared to the wild-type strain, most likely due to a more efficient access of the enzyme to the substrate and iii) potential RNA substrates that are specifically up-regulated in the presence of ∆TMD-RNase Y. The number of 295 transcripts might be over-estimated as some values for the ∆rny strain are close to the 2x cut-off (Table S6). Nevertheless, this could highlight another potentially important aspect of pseudo-compartmentalization which is competition between enzymes with similar activity. We have previously shown that RNase J1/J2 and RNase Y have evolved towards an evolutionarily conserved endo-nucleolytic activity [29,49]. The individual effects of RNase J1 and J2 mutations on the transcriptome have been analyzed [50] but the extent of endo-nucleolytic cleavages mediated by RNase J1/J2 is unknown. A plausible explanation for the specific induction of transcripts in the presence of ∆TMD-RNase Y would be that cytoplasmic RNase Y can recognize but not efficiently cleave certain RNase J cleavage sites. The increased level of the rbsC mRNA in the ∆J mutant and the ∆TMD strain tends to confirm this hypothesis. However, a more detailed analysis of this and other potential RNA substrates are required to validate this competition model. Some bacteria like many firmicutes or delta-proteobacteria have all three RNases Y, J and E. In these organisms, homeostatic mechanisms including subcellular location of major RNA decay enzymes with similar specificity are likely important to optimize RNA metabolism.
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
The B. subtilis strains used in this work are derivatives of strain SSB1002, a wild-type laboratory stock strain derived from strain 168. E. coli strain JM109 [51] was used for plasmid constructions. B. subtilis strain SSB507 is wild-type for rny and contains the empty pDR160T vector integrated at amyE. SSB508 is derived from SSB507 where the rny ORF has been replaced in frame with that of chloramphenicol acetyltransferase (651 nt) from the S. aureus plasmid pC194 [52] as described [21]. SSB574 carries the same rny deletion as SSB508 and a xylose-inducible copy of the rny gene lacking the 5’ terminal transmembrane domain (aa 2-24) integrated at amyE (plasmid pHMD40). Primers used in this study are listed in Table 2. B. subtilis and E. coli strains were grown at 37°C in LB medium with aeration. Expression of cytoplasmic (∆TMD) RNase Y was induced by the addition of 50 mM xylose to the medium. When required, the following antibiotics were added to the medium: chloramphenicol (5 μg/ml) and spectinomycin (100 μg/ml).
Strains SSB2048 and SSB2066 express RNase Y-sfGFP and ∆TMD-RNase Y-sfGFP from the native rny locus as described [21]. Briefly, they were constructed by markerless allelic replacement using the thermoexcisable plasmid pMAD [53] and epifluorescence images were taken from mid-log cultures with a 100x oil objective on a Zeiss AxioImager M1 as decribed [21].

Table 2.
B. subtilis strains used in this study.
| B. subtilis Strain | Relevant Genotype | Reference |
|---|---|---|
| SSB1002 | Wild type strain | Lab stock |
| SSB507 | ∆amyE::pDR160T | This work |
| SSB508 | ∆rny::cat, ∆amyE::pDR160T | This work |
| SSB574 | ∆rny::cat, ∆amyE::pHMD40 | This work |
Table 3.
Oligonucleotides used in this study.
| Oligonucleotide | Sequence 5’-3’ |
|---|---|
| HP1696 | GACTCGAGCCGTAGAGTATGCAAAATAAAGGATCCTATC |
| HP1827 | AATGATTAATTAACAACAACCAAGTTCATAGCAAGAGGAGGTGAAAGTATGCGTAAAACCATTGCCGAAGCG |
4.2. Plasmid Constructs
pDR160T. Ectopic integration vector (at amyE) expressing inserted genes from the xylose-inducible Psweet promoter [54]. The original vector was modified to contain the rnjA 3’ transcription terminator in order to terminate transcripts transcribed from the xylose inducible promoter [29].
pHMD40. Plasmid pHMD40 contains a mutated rny gene lacking amino acids 2 to 24 (∆TMD), encoding a cytoplasmic version of RNase Y under the control of the xylose-inducible promoter and the wild-type rny Shine-Dalgarno sequence. A 1.5 kb PCR fragment (oligonucleotides HP1827-HP1696) was cleaved with PacI and BamHI and ligated into the respective sites of plasmid pDR160T.
4.3. Epi-Fluorescence Microscopy
GFP fluorescent images were taken with the Zeiss Axio Imager M1 microscope equipped with an AxioCam MRm camera (Zeiss) using filter set 10 (Zeiss). For the visualization of cells from exponentially growing cultures, overnight cultures in LB medium were diluted to OD600 ∼0.1 and grown at 37◦C in fresh LB medium for at least three generations. Cells were mounted on 1% (w/v) agarose pads and images acquired by an AxioCam camera MRm (Zeiss) using a 1.3 NA ×100 oil objective.
4.4. Northern Blot
RNA blot analysis was carried out using 5 μg of total RNA separated on 0.8 or 1.2 % formaldehyde-agarose gels. The RNA was transferred by capillary blotting to a Hybond N+ membrane (GE Healthcare) and UV cross-linked at 120 mJ cm-2 for 1 min. Hybridization to specific RNA probes was carried out as described previously [29].
4.5. Western Blot
For Western blot analysis, a monoclonal RNase Y antibody directed against a 12 aa peptide (residues 79-90) was used to detect RNase Y in 20 μg of protein extract as described previously [55].
4.6. Transcriptome Analysis
The RNAseq analysis (library preparation and sequencing) was performed at the iGE3 Genomics Platform of the University of Geneva (hhhps://ige3.genomics.unige.ch). RNA samples were treated with the Ribo-Zero Kit for Bacteria (Illumina) to remove ribosomal RNA (rRNAs). Multiplex RNA-Seq libraries were prepared with the Illumina TruSeq Stranded Total RNA Sample Preparation and sequenced (HiSeq2000). The complete genome sequence of the Bacillus subtilis subsp. subtilis str. 168 (NCBI accession number for chromosome: NC 000964.3) and its annotation were retrieved from the NCBI: (https://www.ncbi.nlm.nih.gov/genome/?term=Bacillus±subtilis) and from Subtiwiki [33]. Reads processing, mapping and differential gene expression analysis were performed according to [56]. Briefly, single-end 100-nt reads were mapped to the reference genome using bwa and allowing soft-clipping in the alignment parameters (bwa mem) [57]. Analysis of the mappings used samtools for processing, sorting and indexing of the alignment files; BEDtools for computing coverage; the IGV browser for displaying the alignment files [57,58,59]. Reads counts were normalized either as reads per million (RPM) or as transcripts for million (TPM). Differential gene expression analysis between strains was performed with the EdgeR package [60] using the Trimmed Mean of M-values (TMM) scaling factor as normalization method. Gene features were considered as significantly up- or down-regulated in the mutants versus WT samples if the log2 fold-change (FC) ratio was ≥1 or ≤–1 and the p-value adjusted for multiple testing False Discovery Rate (FDR) calculated using the Benjamini-Hochberg (BH) method in edgeR was equal or lower than 5% (FDR ≤ 0.05). Multi-dimensional scaling (MDS) and MA plots (visualizing measurement differences between two samples by transforming the data onto M (log ratio) and A (mean average scales) were generated, respectively, through the ‘plotMDS.dge’ and the plotSmear functions of the edgeR package. Raw sequencing data will be deposited at the NCBI Sequence Read Archive accession number (SRA).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Tables S1 to S6.
Author Contributions
Conceptualization, S.L. and H.P.; methodology, S.L, M.C. and J.O.; validation, S.L., M.C., and H.P.; formal analysis, S.L. and H.P.; investigation, S.L., M.C. and J.O.; resources, H.P.; data curation, S.L., M.C., J.O. and H.P.; writing—original draft preparation, S.L.; writing—review and editing, H.P..; supervision, S.L. and H.P.; project administration, H.P.; funding acquisition, H.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Centre National de la Recherche Scientifique [UMR 8261], University Paris Cité, Agence Nationale de la Recherche [IB-mRND] and Labex Dynamo ANR-11- LABX-0011-01.
Data Availability Statement
Data sharing is not applicable for this article as no datasets were generated or analyzed during the current study.
Acknowledgments
We are thankful to S. Ngo for excellent technical support.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Laalami, S.; Zig, L.; Putzer, H. Initiation of mRNA decay in bacteria. Cellular and molecular life sciences: CMLS 2014, 71, 1799-1828. [CrossRef]
- Shahbabian, K.; Jamalli, A.; Zig, L.; Putzer, H. RNase Y, a novel endoribonuclease, initiates riboswitch turnover in Bacillus subtilis. EMBO J 2009, 28, 3523-3533. [CrossRef]
- Lehnik-Habrink, M.; Schaffer, M.; Mader, U.; Diethmaier, C.; Herzberg, C.; Stulke, J. RNA processing in Bacillus subtilis: identification of targets of the essential RNase Y. Mol Microbiol 2011, 81, 1459-1473. [CrossRef]
- Durand, S.; Gilet, L.; Bessieres, P.; Nicolas, P.; Condon, C. Three essential ribonucleases-RNase Y, J1, and III-control the abundance of a majority of Bacillus subtilis mRNAs. PLoS Genet 2012, 8, e1002520. [CrossRef]
- Laalami, S.; Bessieres, P.; Rocca, A.; Zig, L.; Nicolas, P.; Putzer, H. Bacillus subtilis RNase Y activity in vivo analysed by tiling microarrays. PLoS One 2013, 8, e54062. [CrossRef]
- Chen, Z.; Itzek, A.; Malke, H.; Ferretti, J.J.; Kreth, J. Multiple Roles of RNase Y in Streptococcus pyogenes mRNA Processing and Degradation. J Bacteriol 2013, 195, 2585-2594. [CrossRef]
- Khemici, V.; Prados, J.; Linder, P.; Redder, P. Decay-Initiating Endoribonucleolytic Cleavage by RNase Y Is Kept under Tight Control via Sequence Preference and Sub-cellular Localisation. PLoS Genet 2015, 11, e1005577. [CrossRef]
- Marincola, G.; Wolz, C. Downstream element determines RNase Y cleavage of the saePQRS operon in Staphylococcus aureus. Nucleic Acids Res 2017, 45, 5980-5994. [CrossRef]
- DeLoughery, A.; Lalanne, J.B.; Losick, R.; Li, G.W. Maturation of polycistronic mRNAs by the endoribonuclease RNase Y and its associated Y-complex in Bacillus subtilis. Proc Natl Acad Sci U S A 2018, 115, E5585-E5594. [CrossRef]
- Broglia, L.; Lecrivain, A.L.; Renault, T.T.; Hahnke, K.; Ahmed-Begrich, R.; Le Rhun, A.; Charpentier, E. An RNA-seq based comparative approach reveals the transcriptome-wide interplay between 3'-to-5' exoRNases and RNase Y. Nature communications 2020, 11, 1587. [CrossRef]
- Taggart, J.C.; Lalanne, J.-B.; Durand, S.; Braun, F.; Condon, C.; Li, G.-W. A high-resolution view of RNA endonuclease cleavage in <em>Bacillus subtilis</em>. bioRxiv 2023, 2023.2003.2012.532304. [CrossRef]
- Commichau, F.M.; Rothe, F.M.; Herzberg, C.; Wagner, E.; Hellwig, D.; Lehnik-Habrink, M.; Hammer, E.; Völker, U.; Stülke, J. Novel activities of glycolytic enzymes in Bacillus subtilis: interactions with essential proteins involved in mRNA processing. Mol Cell Proteomics 2009, 8, 1350-1360. [CrossRef]
- Lehnik-Habrink, M.; Pfortner, H.; Rempeters, L.; Pietack, N.; Herzberg, C.; Stülke, J. The RNA degradosome in Bacillus subtilis: identification of CshA as the major RNA helicase in the multiprotein complex. Mol Microbiol 2010, 77, 958-971. [CrossRef]
- Haq, I.U.; Muller, P.; Brantl, S. A comprehensive study of the interactions in the B. subtilis degradosome with special emphasis on the role of the small proteins SR1P and SR7P. Mol Microbiol 2024, 121, 40-52. [CrossRef]
- Newman, J.A.; Hewitt, L.; Rodrigues, C.; Solovyova, A.S.; Harwood, C.R.; Lewis, R.J. Dissection of the network of interactions that links RNA processing with glycolysis in the Bacillus subtilis degradosome. J Mol Biol 2012, 416, 121-136. [CrossRef]
- Cascante-Estepa, N.; Gunka, K.; Stulke, J. Localization of Components of the RNA-Degrading Machine in Bacillus subtilis. Frontiers in microbiology 2016, 7, 1492. [CrossRef]
- Redder, P. Molecular and genetic interactions of the RNA degradation machineries in Firmicute bacteria. Wiley interdisciplinary reviews. RNA 2018, 9. [CrossRef]
- DeLoughery, A.; Dengler, V.; Chai, Y.; Losick, R. Biofilm formation by Bacillus subtilis requires an endoribonuclease-containing multisubunit complex that controls mRNA levels for the matrix gene repressor SinR. Mol Microbiol 2016, 99, 425-437. [CrossRef]
- Adusei-Danso, F.; Khaja, F.T.; DeSantis, M.; Jeffrey, P.D.; Dubnau, E.; Demeler, B.; Neiditch, M.B.; Dubnau, D. Structure-Function Studies of the Bacillus subtilis Ric Proteins Identify the Fe-S Cluster-Ligating Residues and Their Roles in Development and RNA Processing. mBio 2019, 10. [CrossRef]
- Carabetta, V.J.; Tanner, A.W.; Greco, T.M.; Defrancesco, M.; Cristea, I.M.; Dubnau, D. A complex of YlbF, YmcA and YaaT regulates sporulation, competence and biofilm formation by accelerating the phosphorylation of Spo0A. Mol Microbiol 2013, 88, 283-300. [CrossRef]
- Hamouche, L.; Billaudeau, C.; Rocca, A.; Chastanet, A.; Ngo, S.; Laalami, S.; Putzer, H. Dynamic Membrane Localization of RNase Y in Bacillus subtilis. mBio 2020, 11. [CrossRef]
- Hunt, A.; Rawlins, J.P.; Thomaides, H.B.; Errington, J. Functional analysis of 11 putative essential genes in Bacillus subtilis. Microbiology 2006, 152, 2895-2907.
- Khemici, V.; Poljak, L.; Luisi, B.F.; Carpousis, A.J. The RNase E of Escherichia coli is a membrane-binding protein. Mol Microbiol 2008, 70, 799-813, doi:MMI6454 [pii] 10.1111/j.1365-2958.2008.06454.x.
- Strahl, H.; Turlan, C.; Khalid, S.; Bond, P.J.; Kebalo, J.M.; Peyron, P.; Poljak, L.; Bouvier, M.; Hamoen, L.; Luisi, B.F.; et al. Membrane recognition and dynamics of the RNA degradosome. PLoS Genet 2015, 11, e1004961. [CrossRef]
- Lewis, P.J.; Thaker, S.D.; Errington, J. Compartmentalization of transcription and translation in Bacillus subtilis. EMBO J 2000, 19, 710-718. [CrossRef]
- Mascarenhas, J.; Weber, M.H.; Graumann, P.L. Specific polar localization of ribosomes in Bacillus subtilis depends on active transcription. EMBO Rep 2001, 2, 685-689. [CrossRef]
- Hadjeras, L.; Poljak, L.; Bouvier, M.; Morin-Ogier, Q.; Canal, I.; Cocaign-Bousquet, M.; Girbal, L.; Carpousis, A.J. Detachment of the RNA degradosome from the inner membrane of Escherichia coli results in a global slowdown of mRNA degradation, proteolysis of RNase E and increased turnover of ribosome-free transcripts. Mol Microbiol 2019, 111, 1715-1731. [CrossRef]
- Hosoya, S.; Asai, K.; Ogasawara, N.; Takeuchi, M.; Sato, T. Mutation in yaaT leads to significant inhibition of phosphorelay during sporulation in Bacillus subtilis. J Bacteriol 2002, 184, 5545-5553.
- Laalami, S.; Cavaiuolo, M.; Roque, S.; Chagneau, C.; Putzer, H. Escherichia coli RNase E can efficiently replace RNase Y in Bacillus subtilis. Nucleic Acids Res 2021, 49, 4643-4654. [CrossRef]
- Aravind, L.; Koonin, E.V. The HD domain defines a new superfamily of metal-dependent phosphohydrolases. Trends Biochem Sci 1998, 23, 469-472.
- Lehnik-Habrink, M.; Newman, J.; Rothe, F.M.; Solovyova, A.S.; Rodrigues, C.; Herzberg, C.; Commichau, F.M.; Lewis, R.J.; Stulke, J. RNase Y in Bacillus subtilis: a Natively disordered protein that is the functional equivalent of RNase E from Escherichia coli. J Bacteriol 2011, 193, 5431-5441. [CrossRef]
- Hardouin, P.; Velours, C.; Bou-Nader, C.; Assrir, N.; Laalami, S.; Putzer, H.; Durand, D.; Golinelli-Pimpaneau, B. Dissociation of the Dimer of the Intrinsically Disordered Domain of RNase Y upon Antibody Binding. Biophys J 2018, 115, 2102-2113. [CrossRef]
- Zhu, B.; Stulke, J. SubtiWiki in 2018: from genes and proteins to functional network annotation of the model organism Bacillus subtilis. Nucleic Acids Res 2018, 46, D743-D748. [CrossRef]
- Maul, B.; Volker, U.; Riethdorf, S.; Engelmann, S.; Hecker, M. sigma B-dependent regulation of gsiB in response to multiple stimuli in Bacillus subtilis. Molecular & general genetics: MGG 1995, 248, 114-120. [CrossRef]
- Homuth, G.; Mogk, A.; Schumann, W. Post-transcriptional regulation of the Bacillus subtilis dnaK operon. Mol Microbiol 1999, 32, 1183-1197.
- Mulhbacher, J.; Lafontaine, D.A. Ligand recognition determinants of guanine riboswitches. Nucleic Acids Res 2007, 35, 5568-5580. [CrossRef]
- Nelson, J.W.; Sudarsan, N.; Furukawa, K.; Weinberg, Z.; Wang, J.X.; Breaker, R.R. Riboswitches in eubacteria sense the second messenger c-di-AMP. Nature chemical biology 2013, 9, 834-839. [CrossRef]
- Gundlach, J.; Herzberg, C.; Kaever, V.; Gunka, K.; Hoffmann, T.; Weiss, M.; Gibhardt, J.; Thurmer, A.; Hertel, D.; Daniel, R.; et al. Control of potassium homeostasis is an essential function of the second messenger cyclic di-AMP in Bacillus subtilis. Sci Signal 2017, 10. [CrossRef]
- Shin, J.H.; Helmann, J.D. Molecular logic of the Zur-regulated zinc deprivation response in Bacillus subtilis. Nature communications 2016, 7, 12612. [CrossRef]
- Even, S.; Pellegrini, O.; Zig, L.; Labas, V.; Vinh, J.; Brechemmier-Baey, D.; Putzer, H. Ribonucleases J1 and J2: two novel endoribonucleases in B. subtilis with functional homology to E. coli RNase E. Nucleic Acids Res 2005, 33, 2141-2152.
- Woodson, K.; Devine, K.M. Analysis of a ribose transport operon from Bacillus subtilis. Microbiology (Reading) 1994, 140 (Pt 8), 1829-1838. [CrossRef]
- Mora, L.; Ngo, S.; Laalami, S.; Putzer, H. In Vitro Study of the Major Bacillus subtilis Ribonucleases Y and J. Methods Enzymol 2018, 612, 343-359. [CrossRef]
- Mogk, A.; Homuth, G.; Scholz, C.; Kim, L.; Schmid, F.X.; Schumann, W. The GroE chaperonin machine is a major modulator of the CIRCE heat shock regulon of Bacillus subtilis. Embo J 1997, 16, 4579-4590.
- Ogura, Y.; Imai, Y.; Ogasawara, N.; Moriya, S. Autoregulation of the dnaA-dnaN operon and effects of DnaA protein levels on replication initiation in Bacillus subtilis. J Bacteriol 2001, 183, 3833-3841. [CrossRef]
- Brown, S.; Santa Maria, J.P., Jr.; Walker, S. Wall teichoic acids of gram-positive bacteria. Annu Rev Microbiol 2013, 67, 313-336. [CrossRef]
- Sattler, L.; Graumann, P.L. Real-Time Messenger RNA Dynamics in Bacillus subtilis. Frontiers in microbiology 2021, 12, 760857. [CrossRef]
- Gorke, B.; Rak, B. Efficient transcriptional antitermination from the Escherichia coli cytoplasmic membrane. J Mol Biol 2001, 308, 131-145. [CrossRef]
- Al-Husini, N.; Tomares, D.T.; Bitar, O.; Childers, W.S.; Schrader, J.M. alpha-Proteobacterial RNA Degradosomes Assemble Liquid-Liquid Phase-Separated RNP Bodies. Mol Cell 2018, 71, 1027-1039 e1014. [CrossRef]
- Laalami, S.; Putzer, H. mRNA degradation and maturation in prokaryotes: the global players. Biomolecular Concepts 2011, 2, 491-506. [CrossRef]
- Mäder, U.; Zig, L.; Kretschmer, J.; Homuth, G.; Putzer, H. mRNA processing by RNases J1 and J2 affects Bacillus subtilis gene expression on a global scale. Mol Microbiol 2008, 70, 183-196. [CrossRef]
- Yanisch-Perron, C.; Vieira, J.; Messing, J. Improved M13 phage cloning vectors and host strains: nucleotide sequences of the M13mp18 and pUC19 vectors. Gene 1985, 33, 103-119.
- Horinouchi, S.; Weisblum, B. Nucleotide sequence and functional map of pC194, a plasmid that specifies inducible chloramphenicol resistance. J Bacteriol 1982, 150, 815-825.
- Arnaud, M.; Chastanet, A.; Debarbouille, M. New vector for efficient allelic replacement in naturally nontransformable, low-GC-content, gram-positive bacteria. Appl Environ Microbiol 2004, 70, 6887-6891. [CrossRef]
- Rudner, D.Z.; Breger, K.S.; Rio, D.C. Molecular genetic analysis of the heterodimeric splicing factor U2AF: the RS domain on either the large or small Drosophila subunit is dispensable in vivo. Genes Dev 1998, 12, 1010-1021.
- Korobeinikova, A.; Laalami, S.; Berthy, C.; Putzer, H. RNase Y Autoregulates Its Synthesis in Bacillus subtilis. Microorganisms 2023, 11. [CrossRef]
- Cavaiuolo, M.; Chagneau, C.; Laalami, S.; Putzer, H. Impact of RNase E and RNase J on Global mRNA Metabolism in the Cyanobacterium Synechocystis PCC6803. Frontiers in microbiology 2020, 11, 1055. [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754-1760. [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: a flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841-842. [CrossRef]
- Robinson, J.T.; Thorvaldsdottir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat Biotechnol 2011, 29, 24-26. [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139-140. [CrossRef]
Table 1.
Number of up- and downregulated transcripts with a fold-change (FC) ≥ 2 or ≤ -2 (FDR ≤ 0.05) in mutant strains compared to WT.
Table 1.
Number of up- and downregulated transcripts with a fold-change (FC) ≥ 2 or ≤ -2 (FDR ≤ 0.05) in mutant strains compared to WT.
| Strain, Relevant Genotype | No. Upregulated Transcripts |
No. Downregulated Transcripts |
|---|---|---|
| SSB574, ∆TMD-RNase Y | 665 | 652 |
| SSB508, ∆rny | 712 | 751 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



