
Submitted:
10 July 2024
Posted:
11 July 2024
You are already at the latest version
Abstract
Iron is a critical yet limited nutrient for microbial growth. To scavenge iron, most microbes produce siderophores—diverse small molecules with high iron affinities. Different siderophores are specifically recognized and uptaken by corresponding recognizers, enabling targeted interventions and intriguing cheater-producer dynamics. We propose constructing a comprehensive iron interaction network, or "iron-net," across the microbial world. Such a network offers the potential for precise manipulation of the microbiota, with conceivable applications in medicine, agriculture, and industry, as well as advancing microbial ecology and evolution theories. Previously, our successful construction of an iron-net in the Pseudomonas genus demonstrated the feasibility of co-evolution-inspired digital siderophore typing. Enhanced by machine learning techniques and expanding sequencing data, forging such an iron-net calls for multidisciplinary collaborations, and holds significant promise in addressing critical challenges in microbial communities.
Keywords:
iron-net
; siderophores
; microbial community
; ecology
; evolution
1. Poking Into the Microbial Community by Siderophore-Mediated Iron Interaction Network
Ironically, most microbes on Earth are limited by iron [1]. Since the Great Oxygenation Event, the oxidation of iron has led to its precipitation as insoluble iron(III) oxides, significantly reducing the availability of bioavailable iron in most natural habitats [2]. Consequently, the concentration of free iron in these environments is much lower than the levels needed by microbes for optimal growth (Figure 1A, upper left). For example, while bacteria generally require iron concentrations around 10-7-10-5 M to thrive [1], the available iron in the ocean ranges from 10-11-10-10 M [3], in soil around 10-18 M [4], and in human extracellular environments as low as 10-24 M [5].
To overcome this scarcity, microbes have evolved to produce siderophores, a class of small molecules with high affinity for iron [6]. Siderophores chelate iron from the environment, forming siderophore-iron complexes that can be specifically recognized and taken up by microbial cells [7] (Figure 1A, lower left). This process involves two key components: (1) the production and (2) the recognition and uptake of diverse siderophores.
Despite being essential for microbial survival, siderophores exhibit remarkable structural diversity (Figure 1A, upper right). At least 700 siderophore structures have been experimentally determined [8], and there could be thousands of iron-binding structures in the vast natural products repository [9]. Moreover, bioinformatic approaches have revealed that structural diversities, even within well-studied genera like Pseudomonas, are largely hidden [10]. Moreover, siderophore is among the largest classes of microbial secondary metabolites [11], and our preliminary computational analysis indicates that around 60% of microbial genomes contain at least one siderophore synthetase pathway, highlighting the vast potential for siderophore diversity.
The other key components, recognition and uptake, differ significantly between Gram-negative and Gram-positive microbes (Figure 1A, lower right) [12]. In Gram-negative bacteria, siderophore-iron complexes are recognized by TonB-dependent receptors (TonBDR) and transported across the outer membrane, through the periplasm, and into the cytoplasm via ATP binding cassette (ABC) transporters [13]. Conversely, Gram-positive bacteria, which lack an outer membrane, use membrane-bound periplasmic binding proteins (PBP) to recognize the siderophore-iron complexes, then the ABC transporters to directly transport them across their cell wall [14,15]. Intriguingly, in our preliminary estimation, while 40% of microbes are incapable of producing siderophores, only around 8% lack these siderophore recognition machinery (does not possess TonBDR nor PBP in genome). This suggests interesting ecological interplay, where non-producing microbes may rely on siderophores produced by others within their community.
The specific recognition relationship between various siderophores and their corresponding recognizers is crucial in microbial iron scavenging. Intriguingly, previous research has shown that different siderophores are preferentially recognized and taken up by specific recognizers [16,17,18]. For example, experimental evidence indicates that three different strains of Pseudomonas aeruginosa each produce three types of pyoverdines (the typical siderophore in Pseudomonas genus) and can only utilize the pyoverdine they produce, with minimal cross-recognition or cross-uptake (Figure 1B) [17,18]. Despite the structural similarities among pyoverdine receptors (FpvAs) in these strains, subsequent structural studies identified sites in the loop L7 and the plug domain that influence receptor selectivity [19,20]. Recent bioinformatic analyses suggested that sequence regions near the plug domain are most strongly associated with pyoverdine selectivity in Pseudomonas [10]. The pyoverdines and their matching receptors form specific recognition relationships, much like keys fitting into locks.
Meanwhile, siderophore piracy, where microbes utilize siderophores not produced by themselves, is widespread. This phenomenon occurs not only within phylogenetically close species but also extends across genera, families, and even kingdoms [21,22,23]. Studies investigating natural habitats have shown that uncultured strains frequently utilize siderophores produced by nearby species [24]. Other examples come from well-characterized model organisms. For instance, Escherichia coli can uptake not only the enterobactin it produces, but also aerobactin(produced by Klebsiella and some pathegenic E. coli), ferrichrome (produced by multiple fungi), coprogen (produced by Penicillium), and heterobactin A (produced by Rhodococcus) [25,26] (Figure 1C). Similarly, the Gram-positive Bacillus subtilis produces bacillibactin and can also uptake enterobactin, ferrichrome, and ferrioxamine E (produced by Streptomycetes) [27,28,29]. Pseudomonas aeruginosa produces pyoverdine and pyochelin and can pirate ferrichrome and enterobactin [30,31,32], though it cannot utilize the structurally similar bacillibactin [33]. Within the diverse types of pyoverdines produced by the Pseudomonas genus, most Pseudomonas aeruginosa strains are stringent in only uptaking self-produced pyoverdines [17,34]. However, other Pseudomonas species, such as Pseudomonas fluorescens and Pseudomonas putida, frequently uptake different pyoverdines produced by other strains [22,35].
How can extensive cross-utilization between microbes occur despite the stringent specific recognition between siderophores and their recognizers? The answer lies in the genetic makeup of most microbes, which usually possess multiple recognizers in their genomes [36]. Besides the self-recognizers for their own siderophores, these additional recognizers can match the lock-key relationship of various target siderophores, allowing a strain to uptake siderophores from other producers [18,26,28,30,37] (Figure 1C).
Taken together, we envision a comprehensive “iron-net” (Figure 1D), an iron interaction network mediated by various siderophores across microbes. We believe that such network would provide a sharp cutting point to “poke” into the microbial community. The universal need for iron and the widespread presence of siderophore production and uptake genes ensure the broad applicability of this approach. Meanwhile, the diversity of siderophores, coupled with their specific lock-key relationships with recognizers, provides room for differentiation. Although depicted ideally in Figure 1D, these relationships are not strictly one-to-one; for instance, recognizers in E. coli and B. subtilis can uptake multiple siderophore types, and a single siderophore, such as enterobactin, can be recognized by different recognizers in gram-positive and gram-negative bacteria (Figure 1C). Despite this, recognizers generally show strong preferences for specific siderophores. This discriminative power allows for differentiation of microbes in the siderophore chemical space. For example, in a microbiota with varying health states, such as different enterotypes, modifying siderophore sets could potentially shift it from a diseased to a healthier state (Figure 1E) [38]. These differences create opportunities for targeted interventions in microbial communities, enabling the rational manipulation and design of complex microbiotas.
A. Overview of Siderophore-Microbe Relationships. Ferric iron concentrations in various environments are significantly below microbial requirements (upper left). Microbes produce siderophores to chelate and uptake iron (lower left). The chemical diversity of known siderophores is illustrated with examples (upper right). Different siderophore uptake mechanisms in Gram-negative and Gram-positive bacteria are shown, highlighting the distribution of TonB-dependent receptors (TonBDRs) and periplasmic binding proteins (PBPs) (lower panel).
B. Recognition of pyoverdine siderophores by specific receptors in different Pseudomonas. The names of three siderophores can be searched in Siderophore Information Database (SIDERITE, https://siderite.bdainformatics.org/) [9]. Pyoverdine receptors are indicated by by notched rectangles.
C. Cross-utilization of siderophores among different species. Double arrow indicates that this species can produce and update this siderophore, and the corresponding recognizers are marked with edges. Meanwhile, single arrow indicates that this species only can update (but not produce) this siderophore, and the corresponding recognizers have no edges.
D. A scheme of microbial interaction networks medicated by siderophores. Different types of siderophores (indicated by colors) and their corresponding recognizers on microbial membranes are shown. Microbes are represented by rounded rectangles, siderophores by colored hexagons, and recognizers by notched rectangles. Recognizers can uptake siderophores of the same color. Production is indicated by black arrows, and utilization by arrows in the corresponding siderophore colors. The same settings are used for illustrating siderophore interactions in subsequent figures.
E. Schematic of using different siderophores to regulate microbial community states.
2. From Sequence to Ecology: Harness the Molecular Coevolution to Predict Siderophore-Mediated Interactions
The fundamental step to construct such an iron-net is to perform “siderophore-typing” for each microbe [17,39]. This involves identifying which types of siderophores a microbe produces (if any) and which types of siderophores it can uptake. In this context, we use the term “type” to refer to a set of structurally similar siderophores that can be uptaken by a set of highly homologous recognizers.
The primary challenges in siderophore-typing all microbes are the limited experimental capacities and the bioinformatic complexities. Bioinformatic analyses often identify multiple recognizers within a genome [36], making it challenging to distinguish between “self-” recognizers (those that uptake the types of siderophores produced by the same strain) and “cheating-” recognizers (those that uptake types of siderophores not produced by this strains) [37]. The widespread cross-interactions between phylogenetically distant species add further complexity. However, the correspondence between genetic sequences and functional specificity, such as recognizers specificity and its genetic sequences, and siderophore products with their synthetase pathways, offers potential solutions. By these gene-function mappings, it is possible to reconstruct the iron net from microbial sequences.
We propose that molecular coevolution can be harnessed to identify the lock-key pairs of siderophores and their recognizers. The basis for such coevolution lies in the fact that siderophores are costly secondary metabolites, and cells must ensure that they can uptake the siderophores they produce. Microbes generally use two main pathways for siderophore production [6]: the Non-Ribosomal Peptide Synthetase (NRPS) pathway and the NRPS-independent Siderophore (NIS) synthetase pathway (Figure 2A). NRPS pathways are utilized more frequently than the NIS pathway, responsible for around 70% of structurally known siderophores [9]. The NRPS pathway operates in an assembly line manner, where each module selects and adds specific components to the growing peptide chain, resulting in a diverse array of siderophores [11]. Of note, each module is approximately 1000-1300 amino acids in length, and a siderophore NRPS pathway typically contains three to eight modules, making this gigantic enzyme expensive to synthesize. The NIS pathways are shorter in nucleotide length in general, yet still involve multiple enzymes that catalyze the synthesis of siderophores from carboxylic acid substrates [40,41]. Besides enzyme synthesis, both NRPS and NIS pathways may compete with protein synthesis for the amino acid pool, imposing further metabolic costs on the cells. Experimental evidence have suggested that forced production of siderophores when iron is non-limiting decreases the fitness of microbes [42], supporting the high cost of synthesizing siderophores.
Given the cost of synthesis, structural coevolution between siderophores and their corresponding recognizers is reinforced by natural selection [43]. A microbe with matching siderophores and recognizers can efficiently uptake the necessary iron, maintaining its fitness (Figure 2B, top panel, with both siderophore and recognizers shown in green). However, mutations in the siderophore synthetase (left arrow, resulting in new type of siderophores illustrated in blue) or in the recognizers (right arrow, changing recognizers into a new yellow type) lead to mismatches. Unmatched siderophores and recognizers fail to transport iron into the cell, causing the microbe to incur the costs of siderophore synthesis without reaping the benefits, thereby reducing its fitness. Appropriate mutations that either revert the changes in siderophore or recognizer structures back to the original type or further mutate the sequences to create new matching lock-key pairs can restore the iron scavenging machinery. This ensures that the microbe’s investment in siderophore production is compensated (Figure 2B, bottom panel). Under the assumption that “cells need to be able to uptake the siderophores they produce,” various lock-key pairs of siderophores and recognizers evolve, with concurrent changes in their key sites demonstrating coevolution.
Multiple sequence-based computational approaches can help establish the lock-key relationships between siderophores and their recognizers. The first is by genome approximity (Figure 2C, first panel). In many microbes, siderophore synthetase genes are located close to their corresponding recognizer genes, facilitating coevolution through genome rearrangements [44]. For instance, in E. coli, the enterobactin synthetase and its receptor FepA are within the same biosynthetic gene cluster [45]. In Pseudomonas, the majorities of self-recognizers lie within 10 kb of the synthetase [46]. However, this proximity rule has exceptions, such as in B. subtilis, where bacillibactin synthetase is over 1000 kb away from the periplasmic binding protein FeuA [47].
Recently, our work exploited the sequence co-variation between synthetase and recognizer genes to reconstruct the pyoverdine-mediated interaction network in Pseudomonas [46] (Figure 2C, second panel). The approach assumes that the strongest coevolution signals occur between matching synthetase and recognizer pairs. Algorithms maximizing coevolution strength can identify these “self-recognizer” from the many possible recognizer in the genome. This method is not limited by genome co-localization of the iron scavenging machineries, yet requires a large number of sequences for accurate detection of coevolution signals. It has shown high prediction accuracy in experimental validations.
Leveraging advances in large models of structure prediction and molecule docking [48,49], a third approach can be developed in the near future to predict interactions at a molecular level (Fig. 2C, last panel). Two levels of protein-ligand dockings are required. Initially, the substrates of each module in the NRPS can be inferred from its genome sequence, allowing the structure of the small molecule siderophore to be determined [10]. Subsequently, the structure of the siderophore recognizers (TONBDR or PBP) can be predicted from their sequences, and docking between recognizers and siderophores enables the determination of lock-key relationships from a mechanistic perspective.
The rapid expansion of microbial sequence data offers new opportunities to understand the iron interaction behaviors of microbes [50]. We envision a pipeline to predict these behaviors from any sequenced microbial genome (Figure 2D). The input would be the genome sequence, processed by a pre-trained model that has learned lock-key relationships from extensive microbial sequence data, using the three aforementioned methods or a combination thereof. This pre-trained model should connect lock-key pairs in two sequence spaces: synthetases producing similar siderophores and recognizers selecting these siderophores. Although interactions occur in the chemical space, we assume that sequence similarity correlates with functional similarity. Thus, any query gene sufficiently similar to one type of lock-key pair in the sequence space would behave as a member of this type.
The output is the “microbial iron functional group”, indicating which types of siderophores the microbe produces and uptakes. By this output, recognizers in a microbe can be classified into self-recognizers (uptaking its own siderophores) and cheating recognizers (uptaking siderophores from other organisms). Mathematically, it can be represented by a two-element digital vector, characterizing the microbe’s iron acquisition and competition strategies. If such “from sequence to functional group” digital siderophore-typing can be realized in general, the expanding sequence data will enable researchers to investigate iron interaction networks across the microbial world, opening up intriguing explorations into ecological and evolutionary dynamics.
A. Overview of the two main classes of siderophore synthesis pathways: Non-Ribosomal Peptide Synthetase (NRPS) (upper) and NRPS-independent Siderophore (NIS) (lower). Approximately 70% of microbial siderophores are synthesized by NRPS, while about 30% are synthesized by NIS.
B. Illustration of the selection pressure driving structural co-evolution between siderophores and their recognizers. This plot uses the same settings as Figure 1D for illustrating siderophore interactions.
C. Three computational methods for detecting co-evolution between siderophore synthetases and recognizers.
D. Proposed pipeline for predicting siderophore functional groups from microbial genomes. The functional group is characterized by a two-element vector identifying the types of siderophores produced and utilized by each strain.
3. The Iron Interactions Network as a Playground for Microbial Game Theories
Iron interactions among microorganisms create complex and fascinating ecological dynamics. Iron, being the only limiting nutrient in this game, is divided into various “sectors” by different types of siderophores [51]. For each type of siderophore-iron complexes, microbes possessing the matching recognizers uptake them as public goods, whereas the others see them as “public bads” that prevent them from accessing iron [52]. From the perspective of production and cheating, multiple iron strategies can emerge. In Pseudomonas, we observed three classes of iron acquisition strategies [46] (Figure 3A): 1. “Pure Producers” that produce siderophores and only uptake the classes of siderophores they produce, lacking cheating recognizers. 2. “Partial Producers” that produce/uptake their own siderophores and also absorb siderophores produced by others, possessing both self- and cheating-recognizers. 3. “Pure Cheaters” that do not produce siderophores while uptake siderophores produced by others, possessing only cheating recognizers. Different habitats and lifestyles exhibit distinct preferences for iron strategies. Notably, various pathogenic lifestyles consistently correlate with the lack of partial producers in the community, raising further curiosity about the ecological interactions mediated by these iron acquisition strategies [46].
We can utilize a siderophore interaction matrix (SIM) to analyze the complex interplays in siderophore-mediated iron interactions between two microbes (Figure 3B). Each strain’s iron functional group is characterized by a two-element vector identifying its self and cheating recognizers. This creates a 2x2 matrix describing the overlaps between two strains’ self and cheating recognizers. Each element of this matrix represents different types of interactions.
For example, if the self-recognizers of two microbes belong to the same type, the self1-self2 element of the matrix will be non-zero, indicating the convergent production of the same siderophores (Figure 3C, upper left plot). Conversely, if the self1-self2 element is zero, it indicates that the two microbes produce different siderophores, leading to direct iron competition (Figure 3C, upper right plot). When the self-recognizer of microbe 2 overlaps with the cheating-recognizer of microbe 1, cheating occurs, where microbe 2 exploits the siderophore production of microbe 1 (Figure 3C, lower left plot). When both microbes share the same cheating recognizers (Figure 3C, lower right plot), they can compete to exploit another microbe. Multiple interactions can occur simultaneously between two microbes, illustrating the complexity of microbial iron interactions.
Many classical scenarios in microbial game theory can be realized through different combinations of self- and cheating-recognizers in microbiota, alongside intriguing new games that warrant further interdisciplinary investigations (Figure 3D). Classical competitive exclusion [53], or “winner takes all,” is demonstrated by microbes producing different siderophores and engaging in direct iron competition (Figure 3D(1)). Without cross-utilization, the strain that isolates iron more effectively or has a higher initial inoculation wins, eliminating the others [54].
The “tragedy of the commons” [55] occurs when a producer is exploited by a pure cheater strain, leading to the extinction of both microbes (Figure 3D(2)). When two microbes produce different siderophores with one cheating on the other, the scenario remains “winner takes all,” but the partial producer (microbe 2) gains an advantage over the pure producer (microbe 1). This makes microbe 2 more resistant to invasion by microbe 1, and microbe 1 more capable of invading microbe 2 (Figure 3D(3)). This biased advantage towards partial cheaters can be utilized for pathogen control [46,56].
Interestingly, when both strains cheat on each other while producing their own siderophores, mutual piracy allows coexistence under appropriate cheating strength [57] (Figure 3D(4)). This opens up intriguing discussions on how cheating may facilitate biodiversity.
The “rock-paper-scissors” dynamic, a nonlinear phenomenon with significant implications for dynamic coexistence, raises researchers’ interest in both synthetic and natural systems [58,59,60]. Although it has been challenging to find microbes with such cyclic relationships in nature [59,61], we propose that this dynamic might be easily realized in siderophore-mediated interactions. Three partial producers, each producing a siderophore type that can be utilized by the following strain, could exhibit this relationship (Figure 3D(5)).
Taken together, the intricate iron interactions among microbes create an exciting playground for microbial game theory, offering profound insights into ecological dynamics and biodiversity. This framework sets the stage for further interdisciplinary research and innovative approaches to understand the design principle and to manipulate microbial communities.
A. Three classes of strategies for iron acquisition are differentiated by each microbe’s siderophore production and uptake behaviors, with their functional groups illustrated in the bottom panel. Self-recognizers are marked with edges, and cheating-recognizers have no edges. A box with a cross indicates “non-existence”. All plots in this figure use the same settings as Figure 1D for siderophore-mediated interactions, with the edge colors of microbes indicating the classes of strategies (Green: pure producer; orange: partial producer; red: pure cheater).
B. The Siderophore-Interaction Matrix (SIM) characterizes the interactions mediated by siderophores between two microbial strains. Each of the four elements of the matrix indicates whether or not the self- and cheating-recognizers of the two strains overlap in type (indicated by 1 and 0 values), or if strains do not have such recognizers (indicated by an NA value).
C. Demonstration of the interactions characterized by different elements in the SIM matrix, including convergent production (upper left, by SIM(1,1)=1), iron isolation (upper right, by SIM(1,1)=0), cheating (lower left, by SIM(1,2)=1 or SIM(2,1)=1), and cheater competition (lower right, by SIM(2,2)==1).
D. Classical scenarios of ecological interplays that can be realized by siderophore-mediated interactions, such as competitive exclusion, the tragedy of the commons, partial cheating, and mutual cheating (from left column to right column, D(1) to D(4)). Each column shows, from top to bottom, the strains’ production and utilization relationships, the corresponding SIM, the simplified network representation using arrows from strain i to j to indicate that the siderophore made by strain i can be utilized by strain j, and the schematic phase-diagrams of the dynamic model. Additionally, a three-strain system has the potential to exhibit a ‘rock-paper-scissors’ dynamic, which is also shown in D(5).
4. Fighting for Diversity: The Eco-Evolutionary Implications of Microbial Chemical Innovations via Secondary Metabolism
High diversity of siderophore types is crucial for the complex ecological interactions described. Under intense selection pressure for iron acquisition, how and why have so many different siderophores evolved instead of converging on the most effective siderophore? What are the ecological and evolutionary implications of this diversity.
At the molecular level, the repetitive modular structure and assembly-line working style of NRPS provide a excellent platform for evolving chemical diversity (Figure 4A) [11,62,63]. Rearrangement within this modular enzyme system is enabled by conserved motifs in each module, while mutations in adenylation (A) domains alter substrate selectivity [64,65]. These changes synergistically contribute to the amazing diversity of non-ribosomal peptides (NRPs). Insertion, deletion, or replacement of large DNA fragments drive backbone alterations, while point mutations fine-tune substrate specificity. This high diversity fuels microbial arms races, explaining why NRPS pathways are among the most abundant secondary metabolism pathways in microbes utilized for numerous competitive agents including siderophores and antibiotics [62,66,67,68]. Conversely, the less common NIS pathway for siderophore synthesis remains less explored in terms of chemical diversity and underlying mechanisms.
The practical implications of this molecular diversification extend to the rational re-engineering of NRPSs. Inspired by the pharmaceutical potential of natural products and the “lego-like” assembly of modular enzymes, researchers have been attempting to manipulate NRPS enzymes to create novel products [69]. However, piecing together NRPS modules from different sources has proven notoriously difficult, with directed mutations in substrate-specifying regions often resulting in dysfunctional enzymes [70]. In contrast, natural processes have continually re-engineered NRPSs to produce diverse siderophores. For instance, nearly 200 pyoverdines have been predicted in the Pseudomonas genus, with some differing by only one or two modules, suggesting they are connected by single recombination events [46]. These numerous evolutionary events that shape diverse siderophores provide a wealth of information on natural fusion hotspots and the rules of combination that produce viable products (Figure 4A). By studying these natural processes, researchers can gain valuable insights to guide the engineering of NRPSs [71]. Nature remains a vital teacher in the quest for innovation.
At the level of driving forces, a longstanding question is whether and why generating new siderophores confers evolutionary benefits. The simplest possibility is no—neutral drift can generate new types of siderophores and recognizers without involving selections [72]. However, the relative conservation of other resource uptake pathways suggests an exceptional diversification rate in the iron-scavenging system [73]. Positive selection pressure among FpvA receptors in the Pseudomonas genus further indicates that neutral drift alone may not be sufficient to explain this phenomenon [74].
One explanation involving selection is that the diversification of siderophores provides an advantage by helping producers evade cheaters [74,75] (Figure 4B). Even small changes in the siderophore structure can substantially alter its recognizer specificity, escaping the piracy from other competitors [76]. Given the prevalence of piracy in the siderophore world, this is a plausible explanation [43]. Nevertheless, some highly conserved siderophores, like enterobactin and ferrichromes [23,77,78,79], are frequently exploited by cheaters without evolving into new forms, making researchers wonder their mechanisms of resisting cheating.
Another possible explanation is that it is the recognizers, rather than the siderophores themselves, that need to change due to phage selection (Figure 4C) [74]. Phages can utilize the iron-scavenging pathway to enter microbes [80,81,82]. E.coli has been shown to produce siderophore and even other metabolites to occupy the TONB-dependent receptors to prevent them from being utilized by phage [83,84]. Under such selection pressure, recognizers can mutate to escape phages, which then subsequently lead to changes in siderophore structures. Although multiple theories all provide some compelling narrative, no definitive answer has yet been achieved due to lack of a global overview on the diversity of siderophores.
Additionally, as many siderophores are considered virulence factors for various pathogens and are crucial for some to colonize their hosts [52], the host-microbe interactions can impose positive selection pressures on siderophores [5,43]. Host immune systems have evolved ways to eliminate iron from pathogens, like that neutrophils express lipocalin-2 to sequester enterobactin [85]. Conversely, enterobacteria evade this sequestration by producing glucosylated derivatives of enterobactin [86]. This ongoing tug-of-war between host and bacteria exerts selection pressures on microbial siderophores to adapt and change.
Irrespective of the potential driving forces, the established fact that siderophores exhibit significant diversity has substantial implications for microbial chemical innovations. “Chemical innovation” is a comprehensive term describing how life generates molecules not yet present in its environment, including new siderophores, novel antibiotics, and any other new metabolites [87]. Stuart Kauffman, in his book “Reinventing the Sacred: A New View of Science, Reason, and Religion,” discussed the idea that innovation occurs when life explores the immediate possibilities just beyond the current boundaries of what is known or possible, which expands the “sampling space” of life and fosters diversity [88,89,90]. This concept is demonstrated by the way microbes shape their own niches, not only consuming existing resources but also keeping contributing new chemicals [51]. The emergence of new metabolites exponentially expands the chemical space, mediating new interactions that may drive phase transitions in the community [57]. With increased chemical dimensions, competitive principles may no longer limit biodiversity, as life itself pushes up the boundaries (Figure 4D). Investigating the driving forces and subsequent consequences of chemical innovation, both within and beyond the realm of siderophores, can provide profound insights into the self-organization principles of the microbial world and beyond.
A. Illustration of how siderophore diversification can help us understand nature’s method of re-engineering. Natural NRPS diversification, such as the rearrangement and replacement of modular building blocks, inspires rational re-engineering of NRPS to create new products by understanding fusion hotspots and combination rules.
B-C. Illustration of how the cheater-cooperator game between microbes (B) and the phage selection (C) favors the creation of new siderophore lock-key pairs. Plots use the same settings as Figure 1D for siderophore-mediated interactions.
D. The concept of “Adjacent Possible” in microbial chemical innovations. The cartoon in the upper panel shows how a existing siderophore lock-key pair (green type) expand into its adjacent possibles (different colors) due to certain evolutionary driving forces. The lower panel demonstrates how this diversification of siderophores increases the upper limit of competitive exclusion.
5. Discussion
From a practical perspective, the envisioned iron-net has the potential to provide targeted intervention in microbial communities. Microbes and microbial communities are crucial to human health, agriculture, and industry, yet we lack precise methods to manipulate microbiota due to their astonishing complexity. Indeed, iron interaction is just one of the many ways microbes interact with each other. Competition for primary nutrients like carbon and nitrogen, antibiotic and toxin wars, and prey-predator interactions between microbes and phages are all significant factors shaping microbial communities [91,92,93]. Nevertheless, the critical importance of iron’s catalytic functions in respiration and DNA replication makes iron acquisition an essential condition for the growth of most microbes [94,95], on top of these different interactions [96]. Additionally, the diverse lock-key pairs in the siderophore world enable targeted interventions, and digital siderophore typing provides a feasible method to infer these iron interactions from ever-expanding sequence data. Currently, there are already reports of microbiota interventions through siderophore-mediated interactions [97,98]. In the age of data and machine learning, constructing a comprehensive iron-net becomes possible and will provide us with means for precise manipulation in this invisible world.
From a theoretical perspective, secondary metabolism may introduce some “new ecology,” as it represents life actively expanding its possibility space. Nearly all life forms actively create new ways to survive, from the microbial arms race of antibiotics to human innovations. Previous ecological theories are largely based on predefined dimensions, with interaction methods predetermined in the model. The proposed “adjacent possible” remains largely conceptual, with limited realization in concrete mathematical models. Given the crucial importance of dimensions in theoretical models, investigating how this behavior of actively expanding the possibility space may bring novel insights into ecological theories.
Despite these practical and theoretical potentials, constructing and understanding the iron-net is still a long journey. Beyond the current progress in the Pseudomonas pyoverdine iron-net, many puzzles remain. For example, why are different siderophores so varied, that some siderophores act as universal currencies in the microbial world, while others are privatized? How specific is the siderophore-recognizer lock-key relationship across the microbial world, and why hasn’t a super recognizer that uptakes all siderophores evolved? How do cells regulate their iron strategies in response to different environments and competitors? Can we develop a general algorithm for constructing the iron-net, or do we need to build it genus by genus? How do Gram-positive and Gram-negative microbes cross-talk? Forging the iron-net is a vast project that requires interdisciplinary collaborations, spanning molecular structures, bioinformatics, ecological modeling, and medical and agricultural experiments. We call for researchers from all related fields who are interested to join this endeavor.
Author Contribution
Zhiyuan Li drafted the manuscript; Shaohua Gu, Jiqi Shao, Ruolin He, Guanyue Xiong, Zeyang Qu, and Yuanzhe Shao participated in figure and text revisions. Linlong Yu, Di Zhang, Fanhao Wang, Ruichen Xu, Peng Guo, Ningbo Xi, Yinxiang Li, Yanzhao Wu participated in data curation and literature searches; Zhong Wei revised the manuscript.
Ethics Statement
This article does not contain any studies with human or animal subjects performed by any of the authors.
Acknowledgments
This work was supported by National Natural Science Foundation of China (No. 32071255, No. T2321001). LZ was supported in part by the Peking-Tsinghua Center for Life Sciences.
Conflicts of Interest
All authors of this manuscript declare that they have no conflict of interest or financial conflicts to disclose.
References
- Andrews SC, Robinson AK, Rodríguez-Quiñones F: Bacterial iron homeostasis. FEMS microbiology reviews 2003, 27:215-237. [CrossRef]
- Ilbert M, Bonnefoy V: Insight into the evolution of the iron oxidation pathways. Biochimica et Biophysica Acta (BBA)-Bioenergetics 2013, 1827:161-175. [CrossRef]
- Boyd PW, Ellwood MJ: The biogeochemical cycle of iron in the ocean. Nature Geoscience 2010, 3:675-682. [CrossRef]
- Boguta P, D’Orazio V, Senesi N, Sokołowska Z, Szewczuk-Karpisz K: Insight into the interaction mechanism of iron ions with soil humic acids. The effect of the pH and chemical properties of humic acids. Journal of Environmental Management 2019, 245:367-374. [CrossRef]
- Seyoum Y, Baye K, Humblot C: Iron homeostasis in host and gut bacteria–a complex interrelationship. Gut Microbes 2021, 13:1874855. [CrossRef]
- Hider RC, Kong X: Chemistry and biology of siderophores. Natural product reports 2010, 27:637-657. [CrossRef]
- Krewulak KD, Hans J. Vogel: Structural biology of bacterial iron uptake. 2008, 1778:1781-1804. [CrossRef]
- Siderophore Information Database (SIDERTE). 2024.
- He R, Gu S, Xu J, Li X, Chen H, Shao Z, Wang F, Shao J, Yin WB, Qian L: SIDERITE: Unveiling hidden siderophore diversity in the chemical space through digital exploration. iMeta 2024, 3:e192. [CrossRef]
- Gu Shaohua SY, Rehm Karoline, Bigler Laurent, Zhang Di, He Ruolin, Xu Ruichen, Shao Jiqi, Jousset Alexandre, Friman Ville-Petri, Bian Xiaoying, Wei Zhong, Kümmerli Rolf, Li Zhiyuan: From sequence to molecules: Feature sequence-based genome mining uncovers the hidden diversity of bacterial siderophore pathways eLife 13:RP96719. eLife 2024, 13:RP96719. [CrossRef]
- He R, Zhang J, Shao Y, Gu S, Song C, Qian L, Yin W-B, Li Z: Knowledge-guided data mining on the standardized architecture of NRPS: Subtypes, novel motifs, and sequence entanglements. PLOS Computational Biology 2023, 19:e1011100. [CrossRef]
- Timofeeva AM, Galyamova MR, Sedykh SE: Bacterial siderophores: Classification, biosynthesis, perspectives of use in agriculture. Plants 2022, 11:3065. [CrossRef]
- Schalk IJ, Mislin GL, Brillet K: Structure, function and binding selectivity and stereoselectivity of siderophore–iron outer membrane transporters. Current topics in membranes 2012, 69:37-66. [CrossRef]
- Clarke TE, Ku S-Y, Dougan DR, Vogel HJ, Tari LW: The structure of the ferric siderophore binding protein FhuD complexed with gallichrome. Nature structural biology 2000, 7:287-291. [CrossRef]
- Fukushima T, Allred BE, Sia AK, Nichiporuk R, Andersen UN, Raymond KN: Gram-positive siderophore-shuttle with iron-exchange from Fe-siderophore to apo-siderophore by Bacillus cereus YxeB. Proceedings of the National Academy of Sciences 2013, 110:13821-13826. [CrossRef]
- Grigg JC, Cheung J, Heinrichs DE, Murphy ME: Specificity of Staphyloferrin B recognition by the SirA receptor from Staphylococcus aureus. Journal of Biological Chemistry 2010, 285:34579-34588. [CrossRef]
- Meyer J-M, Stintzi A, De Vos D, Cornelis P, Tappe R, Taraz K, Budzikiewicz H: Use of siderophores to type pseudomonads: the three Pseudomonas aeruginosa pyoverdine systems. Microbiology 1997, 143:35-43. [CrossRef]
- De Chial M, Ghysels B, Beatson SA, Geoffroy V, Meyer JM, Pattery T, Baysse C, Chablain P, Parsons YN, Winstanley CJM: Identification of type II and type III pyoverdine receptors from Pseudomonas aeruginosa. 2003, 149:821-831. [CrossRef]
- Greenwald J, Nader M, Celia H, Gruffaz C, Geoffroy V, Meyer JM, Schalk IJ, Pattus F: FpvA bound to non-cognate pyoverdines: molecular basis of siderophore recognition by an iron transporter. Molecular microbiology 2009, 72:1246-1259. [CrossRef]
- Bouvier B, Cézard C, Sonnet P: Selectivity of pyoverdine recognition by the FpvA receptor of Pseudomonas aeruginosa from molecular dynamics simulations. Physical Chemistry Chemical Physics 2015, 17:18022-18034. [CrossRef]
- Byun H, Jung I-J, Chen J, Larios Valencia J, Zhu J: Siderophore piracy enhances Vibrio cholerae environmental survival and pathogenesis. Microbiology 2020, 166:1038-1046. [CrossRef]
- Butaitė E, Baumgartner M, Wyder S, Kümmerli R: Siderophore cheating and cheating resistance shape competition for iron in soil and freshwater Pseudomonas communities. Nature communications 2017, 8:414. [CrossRef]
- Rutz J, Abdullah T, Singh S, Kalve V, Klebba P: Evolution of the ferric enterobactin receptor in gram-negative bacteria. Journal of bacteriology 1991, 173:5964-5974. [CrossRef]
- D’Onofrio A, Crawford JM, Stewart EJ, Witt K, Gavrish E, Epstein S, Clardy J, Lewis K: Siderophores from neighboring organisms promote the growth of uncultured bacteria. Chemistry & biology 2010, 17:254-264. [CrossRef]
- Carrano CJ, Jordan M, Drechsel H, Schmid DG, Winkelmann G: Heterobactins: a new class of siderophores from Rhodococcus erythropolis IGTS8 containing both hydroxamate and catecholate donor groups. Biometals 2001, 14:119-125. [CrossRef]
- Raymond KN, Allred BE, Sia AK: Coordination chemistry of microbial iron transport. Accounts of chemical research 2015, 48:2496-2505. [CrossRef]
- Grandchamp GM, Caro L, Shank EA: Pirated Siderophores Promote Sporulation in. Applied and Environmental Microbiology 2017, 83. [CrossRef]
- Dertz EA, Xu JD, Stintzi A, Raymond KN: Bacillibactin-mediated iron transport in. Journal of the American Chemical Society 2006, 128:22-23. [CrossRef]
- Zawadzka AM, Abergel RJ, Nichiporuk R, Andersen UN, Raymond KN: Siderophore-mediated iron acquisition systems in Bacillus cereus: identification of receptors for anthrax virulence-associated petrobactin. Biochemistry 2009, 48:3645-3657. [CrossRef]
- Moynié L, Milenkovic S, Mislin GLA, Gasser V, Malloci G, Baco E, McCaughan RP, Page MGP, Schalk IJ, Ceccarelli M, Naismith JH: The complex of ferric-enterobactin with its transporter from Pseudomonas aeruginosa suggests a two-site model. Nature Communications 2019, 10. [CrossRef]
- Ankenbauer RG, Quan HN: FptA, the Fe (III)-pyochelin receptor of Pseudomonas aeruginosa: a phenolate siderophore receptor homologous to hydroxamate siderophore receptors. Journal of bacteriology 1994, 176:307-319. [CrossRef]
- Chan DC, Burrows LL: Pseudomonas aeruginosa FpvB is a high-affinity transporter for xenosiderophores ferrichrome and ferrioxamine B. Mbio 2023, 14:e03149-03122. [CrossRef]
- Dimopoulou A, Theologidis I, Benaki D, Koukounia M, Zervakou A, Tzima A, Diallinas G, Hatzinikolaou DG, Skandalis NJM: Direct antibiotic activity of bacillibactin broadens the biocontrol range of Bacillus amyloliquefaciens MBI600. 2021, 6:10.1128/msphere. 00376-00321. [CrossRef]
- Bodilis J, Ghysels B, Osayande J, Matthijs S, Pirnay JP, Denayer S, De Vos D, Cornelis PJEM: Distribution and evolution of ferripyoverdine receptors in Pseudomonas aeruginosa. 2009, 11:2123-2135. [CrossRef]
- Visca P, Imperi F, Lamont IL: Pyoverdine siderophores: from biogenesis to biosignificance. Trends in microbiology 2007, 15:22-30. [CrossRef]
- Crits-Christoph A, Bhattacharya N, Olm MR, Song YS, Banfield JFJGr: Transporter genes in biosynthetic gene clusters predict metabolite characteristics and siderophore activity. 2021, 31:239-250. [CrossRef]
- Barelmann I, Taraz K, Budzikiewicz H, Geoffroy V, Meyer J-MJZfNC: The structures of the pyoverdins from two Pseudomonas fluorescens strains accepted mutually by their respective producers. 2002, 57:9-16. [CrossRef]
- Costea PI, Hildebrand F, Arumugam M, Bäckhed F, Blaser MJ, Bushman FD, De Vos WM, Ehrlich SD, Fraser CM, Hattori M: Enterotypes in the landscape of gut microbial community composition. Nature microbiology 2018, 3:8-16. [CrossRef]
- Meyer J-M, Geoffroy VA, Baida N, Gardan L, Izard D, Lemanceau P, Achouak W, Palleroni NJJA, Microbiology E: Siderophore typing, a powerful tool for the identification of fluorescent and nonfluorescent pseudomonads. 2002, 68:2745-2753. [CrossRef]
- Li C, Pan D, Li M, Wang Y, Song L, Yu D, Zuo Y, Wang K, Liu Y, Wei ZJFim: Aerobactin-mediated iron acquisition enhances biofilm formation, oxidative stress resistance, and virulence of Yersinia pseudotuberculosis. 2021, 12:699913. [CrossRef]
- Carroll CS, Moore MMJCrib, biology m: Ironing out siderophore biosynthesis: a review of non-ribosomal peptide synthetase (NRPS)-independent siderophore synthetases. 2018, 53:356-381. [CrossRef]
- Griffin AS, West SA, Buckling AJN: Cooperation and competition in pathogenic bacteria. 2004, 430:1024-1027. [CrossRef]
- Kramer J, Özkaya Ö, Kümmerli RJNRM: Bacterial siderophores in community and host interactions. 2020, 18:152-163. [CrossRef]
- Patel P, Song L, Challis GLJB: Distinct extracytoplasmic siderophore binding proteins recognize ferrioxamines and ferricoelichelin in Streptomyces coelicolor A3 (2). 2010, 49:8033-8042. [CrossRef]
- Coderre PE, Earhart CF: The entD gene of the Escherichia coli K12 enterobactin gene cluster. Microbiology 1989, 135:3043-3055. [CrossRef]
- Gu S, Shao Z, Qu Z, Zhu S, Shao Y, Zhang D, Allen R, He R, Shao J, Xiong GJb: Siderophore-receptor coevolution analysis reveals habitat-and pathogen-specific bacterial iron interaction networks. 2023:2023.2011. 2005.565711. [CrossRef]
- Hotta K, Kim C-Y, Fox DT, Koppisch AT: Siderophore-mediated iron acquisition in Bacillus anthracis and related strains. Microbiology 2010, 156:1918-1925. [CrossRef]
- Abramson J, Adler J, Dunger J, Evans R, Green T, Pritzel A, Ronneberger O, Willmore L, Ballard AJ, Bambrick J: Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024:1-3. [CrossRef]
- Zhu J, Gu Z, Pei J, Lai L: DiffBindFR: an SE (3) equivariant network for flexible protein–ligand docking. Chemical Science 2024, 15:7926-7942. [CrossRef]
- Wensel CR, Pluznick JL, Salzberg SL, Sears CLJTJoci: Next-generation sequencing: insights to advance clinical investigations of the microbiome. 2022, 132. [CrossRef]
- Shao J, Rong N, Wu Z, Gu S, Liu B, Shen N, Li Z: Siderophore-mediated iron partition promotes dynamical coexistence between cooperators and cheaters. Iscience 2023, 26. [CrossRef]
- Wu Z, Shao J, Zheng J, Liu B, Li Z, Shen N: A zero-sum game or an interactive frame? Iron competition between bacteria and humans in infection war. Chinese Medical Journal 2022, 135:1917-1926. [CrossRef]
- Hibbing ME, Fuqua C, Parsek MR, Peterson SB: Bacterial competition: surviving and thriving in the microbial jungle. Nature reviews microbiology 2010, 8:15-25. [CrossRef]
- Niehus R, Picot A, Oliveira NM, Mitri S, Foster KR: The evolution of siderophore production as a competitive trait. Evolution 2017, 71:1443-1455. [CrossRef]
- Smith P, Schuster M: Public goods and cheating in microbes. Current biology 2019, 29:R442-R447. [CrossRef]
- Gu S, Yang T, Shao Z, Wang T, Cao K, Jousset A, Friman V-P, Mallon C, Mei X, Wei ZJM: Siderophore-mediated interactions determine the disease suppressiveness of microbial consortia. 2020, 5:10.1128/msystems. 00811-00819. [CrossRef]
- Shao J, Li Y, Lu J, Gu S, Li Z: Siderophore Piracy Promotes Dynamical Coexistence in Microbial Community. bioRxiv 2023:2023.2011. 2021.568182. [CrossRef]
- Li Z, Liu B, Li SH-J, King CG, Gitai Z, Wingreen NS: Modeling microbial metabolic trade-offs in a chemostat. PLoS computational biology 2020, 16:e1008156. [CrossRef]
- Kerr B, Riley MA, Feldman MW, Bohannan BJ: Local dispersal promotes biodiversity in a real-life game of rock–paper–scissors. Nature 2002, 418:171-174. [CrossRef]
- Liao MJ, Din MO, Tsimring L, Hasty J: Rock-paper-scissors: Engineered population dynamics increase genetic stability. Science 2019, 365:1045-1049. [CrossRef]
- Kirkup BC, Riley MA: Antibiotic-mediated antagonism leads to a bacterial game of rock–paper–scissors in vivo. Nature 2004, 428:412-414. [CrossRef]
- Bushley KE, Ripoll DR, Turgeon BG: Module evolution and substrate specificity of fungal nonribosomal peptide synthetases involved in siderophore biosynthesis. BMC Evolutionary Biology 2008, 8:1-24. [CrossRef]
- Baunach M, Chowdhury S, Stallforth P, Dittmann E: The landscape of recombination events that create nonribosomal peptide diversity. Molecular Biology and Evolution 2021, 38:2116-2130. [CrossRef]
- Fan J, Ren J, He R, Wei P-L, Li Y, Li W, Chen D, Druzhinina IS, Li Z, Yin W-B: Biosynthetic diversification of peptaibol mediates fungus-mycohost interactions. bioRxiv 2022:2022.2006. 2005.494846. [CrossRef]
- Fischbach MA, Walsh CT: Assembly-line enzymology for polyketide and nonribosomal peptide antibiotics: logic, machinery, and mechanisms. Chemical reviews 2006, 106:3468-3496. [CrossRef]
- Bruns H, Crüsemann M, Letzel A-C, Alanjary M, McInerney JO, Jensen PR, Schulz S, Moore BS, Ziemert N: Function-related replacement of bacterial siderophore pathways. The ISME journal 2018, 12:320-329. [CrossRef]
- Soutar CD, Stavrinides J: The evolution of three siderophore biosynthetic clusters in environmental and host-associating strains of Pantoea. Molecular genetics and genomics 2018, 293:1453-1467. [CrossRef]
- Cimermancic P, Medema MH, Claesen J, Kurita K, Brown LCW, Mavrommatis K, Pati A, Godfrey PA, Koehrsen M, Clardy J: Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2014, 158:412-421. [CrossRef]
- Wang X, Zhou H, Ren X, Chen H, Zhong L, Bai X, Bian X: Recombineering enables genome mining of novel siderophores in a non-model Burkholderiales strain. Engineering Microbiology 2023, 3:100106. [CrossRef]
- Brown AS, Calcott MJ, Owen JG, Ackerley DF: Structural, functional and evolutionary perspectives on effective re-engineering of non-ribosomal peptide synthetase assembly lines. Natural product reports 2018, 35:1210-1228. [CrossRef]
- Bozhüyük KA, Präve L, Kegler C, Schenk L, Kaiser S, Schelhas C, Shi Y-N, Kuttenlochner W, Schreiber M, Kandler J: Evolution-inspired engineering of nonribosomal peptide synthetases. Science 2024, 383:eadg4320. [CrossRef]
- Barona-Gómez F, Chevrette MG, Hoskisson PA: On the evolution of natural product biosynthesis. In Advances in Microbial Physiology. Volume 83: Elsevier; 2023: 309-349. [CrossRef]
- Martiny AC, Treseder K, Pusch GJTIj: Phylogenetic conservatism of functional traits in microorganisms. 2013, 7:830-838. [CrossRef]
- Smith EE, Sims EH, Spencer DH, Kaul R, Olson MVJJob: Evidence for diversifying selection at the pyoverdine locus of Pseudomonas aeruginosa. 2005, 187:2138-2147. [CrossRef]
- Lee W, van Baalen M, Jansen VAJEl: An evolutionary mechanism for diversity in siderophore-producing bacteria. 2012, 15:119-125. [CrossRef]
- Rütschlin S, Gunesch S, Böttcher TJACB: One enzyme to build them all: ring-size engineered siderophores inhibit the swarming motility of Vibrio. 2018, 13:1153-1158. [CrossRef]
- Thulasiraman P, Newton SM, Xu J, Raymond KN, Mai C, Hall A, Montague MA, Klebba PEJJob: Selectivity of ferric enterobactin binding and cooperativity of transport in gram-negative bacteria. 1998, 180:6689-6696. [CrossRef]
- Chan DC, Burrows LLJM: Pseudomonas aeruginosa FpvB is a high-affinity transporter for xenosiderophores ferrichrome and ferrioxamine B. 2023, 14:e03149-03122. [CrossRef]
- Winkelmann G, Braun VJFML: Stereoselective recognition of ferrichrome by fungi and bacteria. 1981, 11:237-241. [CrossRef]
- Denayer S, Matthijs S, Cornelis PJJob: Pyocin S2 (Sa) kills Pseudomonas aeruginosa strains via the FpvA type I ferripyoverdine receptor. 2007, 189:7663-7668. [CrossRef]
- Bonnain C, Breitbart M, Buck KNJFiMS: The Ferrojan horse hypothesis: iron-virus interactions in the ocean. 2016, 3:82. [CrossRef]
- Rabsch W, Ma L, Wiley G, Najar FZ, Kaserer W, Schuerch DW, Klebba JE, Roe BA, Gomez JAL, Schallmey MJJob: FepA-and TonB-dependent bacteriophage H8: receptor binding and genomic sequence. 2007, 189:5658-5674. [CrossRef]
- Vasse M, Torres-Barceló C, Hochberg MEJPotRSBBS: Phage selection for bacterial cheats leads to population decline. 2015, 282:20152207. [CrossRef]
- Ge H, Hu M, Zhao G, Du Y, Xu N, Chen XJMr: The” fighting wisdom and bravery” of tailed phage and host in the process of adsorption. 2020, 230:126344. [CrossRef]
- Murdoch CC, Skaar EPJNRM: Nutritional immunity: the battle for nutrient metals at the host–pathogen interface. 2022, 20:657-670. [CrossRef]
- Sargun A, Gerner RR, Raffatellu M, Nolan EMJTJoID: Harnessing iron acquisition machinery to target Enterobacteriaceae. 2021, 223:S307-S313. [CrossRef]
- Fischbach MA, Walsh CT, Clardy J: The evolution of gene collectives: How natural selection drives chemical innovation. Proceedings of the National Academy of Sciences 2008, 105:4601-4608. [CrossRef]
- Kauffman SA: Reinventing the sacred: A new view of science, reason, and religion. Basic Books; 2008.
- Mizuno K, Mizuno M, Yamauchi M, Takemura AJ, Medrano Romero V, Morikawa K: Adjacent-possible ecological niche: growth of Lactobacillus species co-cultured with Escherichia coli in a synthetic minimal medium. Scientific reports 2017, 7:12880. [CrossRef]
- Testa B, Vistoli G, Pedretti A: Small Molecules as Exemplars of Emergent Properties and Diversification into the ‘Adjacent Possible’. Chemistry & Biodiversity 2014, 11:1309-1329. [CrossRef]
- Pearl Mizrahi S, Goyal A, Gore J: Community interactions drive the evolution of antibiotic tolerance in bacteria. Proceedings of the National Academy of Sciences 2023, 120:e2209043119. [CrossRef]
- Lee H, Bloxham B, Gore J: Resource competition can explain simplicity in microbial community assembly. Proceedings of the National Academy of Sciences 2023, 120:e2212113120. [CrossRef]
- Pilosof S, Alcala-Corona SA, Wang T, Kim T, Maslov S, Whitaker R, Pascual M: The network structure and eco-evolutionary dynamics of CRISPR-induced immune diversification. Nature Ecology & Evolution 2020, 4:1650-1660. [CrossRef]
- Puig S, Ramos-Alonso L, Romero AM, Martínez-Pastor MT: The elemental role of iron in DNA synthesis and repair. Metallomics 2017, 9:1483-1500. [CrossRef]
- Pau MY, Lipscomb JD, Solomon EI: Substrate activation for O2 reactions by oxidized metal centers in biology. Proceedings of the National Academy of Sciences 2007, 104:18355-18362. [CrossRef]
- Gu S, Wei Z, Shao Z, Friman V-P, Cao K, Yang T, Kramer J, Wang X, Li M, Mei XJNM: Competition for iron drives phytopathogen control by natural rhizosphere microbiomes. 2020, 5:1002-1010. [CrossRef]
- Sargun A, Gerner RR, Raffatellu M, Nolan EM: Harnessing iron acquisition machinery to target Enterobacteriaceae. The Journal of Infectious Diseases 2021, 223:S307-S313. [CrossRef]
- Ellermann M, Arthur JC: Siderophore-mediated iron acquisition and modulation of host-bacterial interactions. Free Radical Biology and Medicine 2017, 105:68-78. [CrossRef]
Figure 1.
An iron interaction network mediated by diverse siderophores offers insights and controls over microbial communities.
Figure 1.
An iron interaction network mediated by diverse siderophores offers insights and controls over microbial communities.
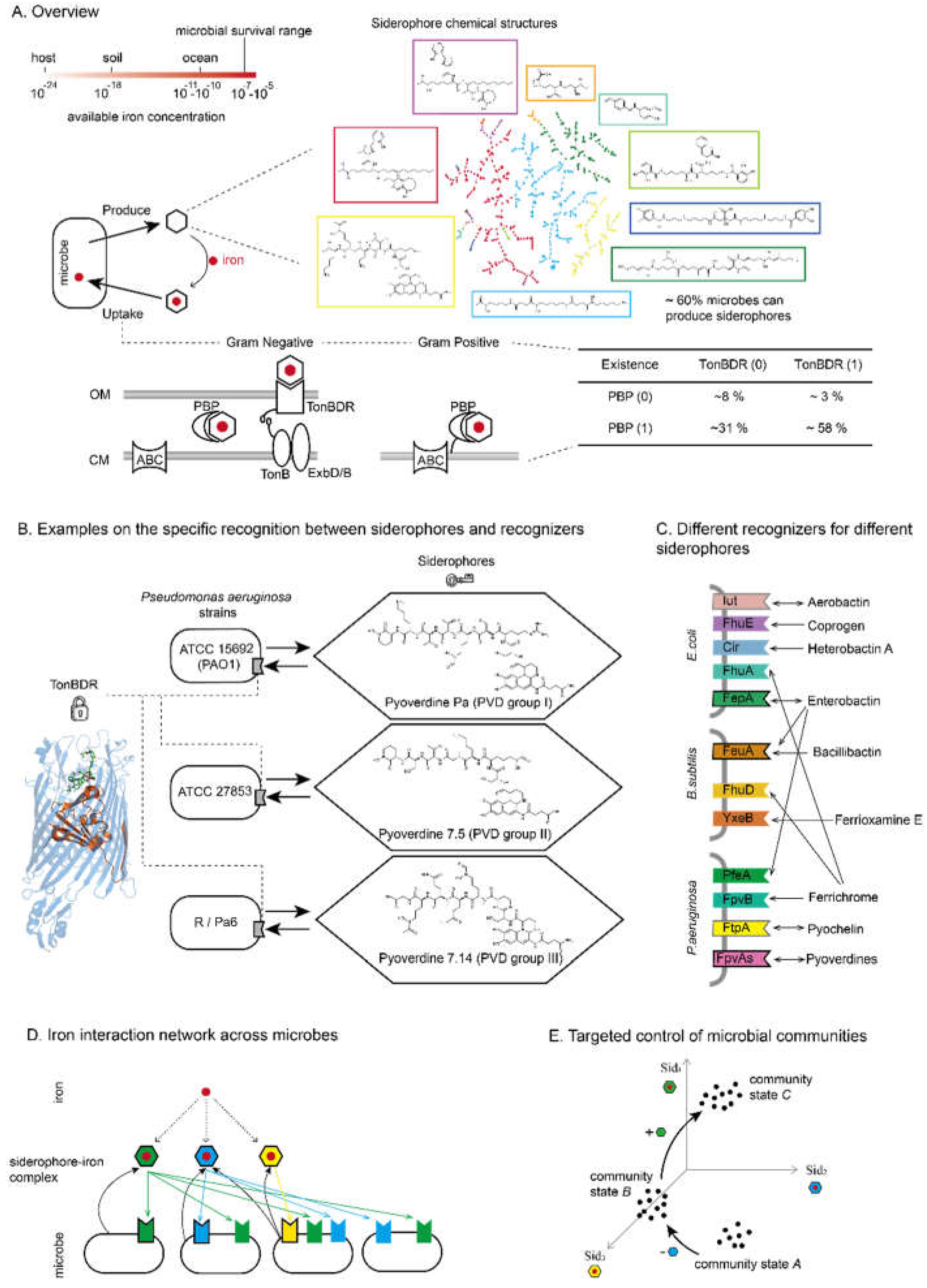
Figure 2.
Digital siderophore-typing based on sequences.
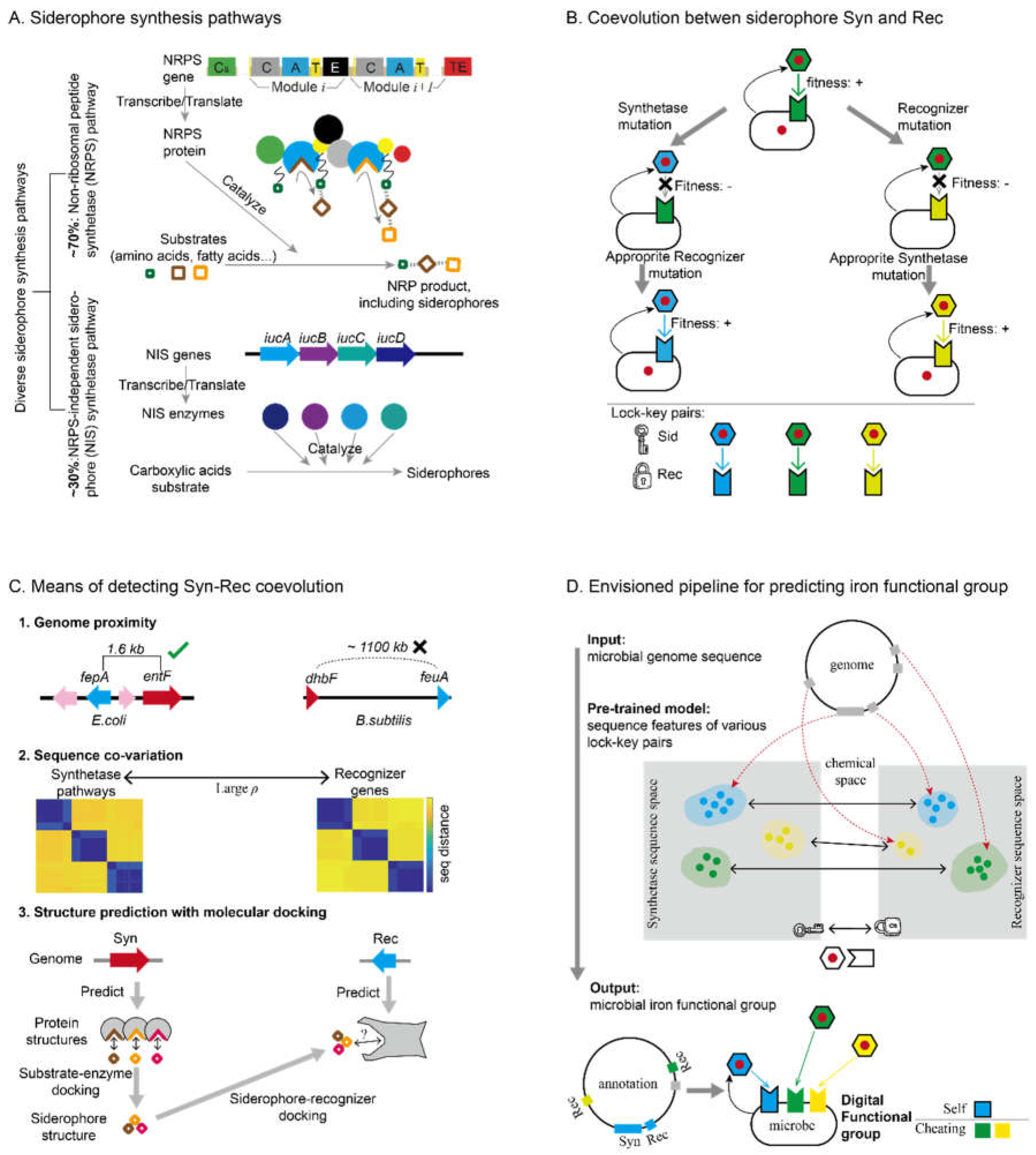
Figure 3.
Siderophore-mediated interactions can manifest diverse ecological interaplays.
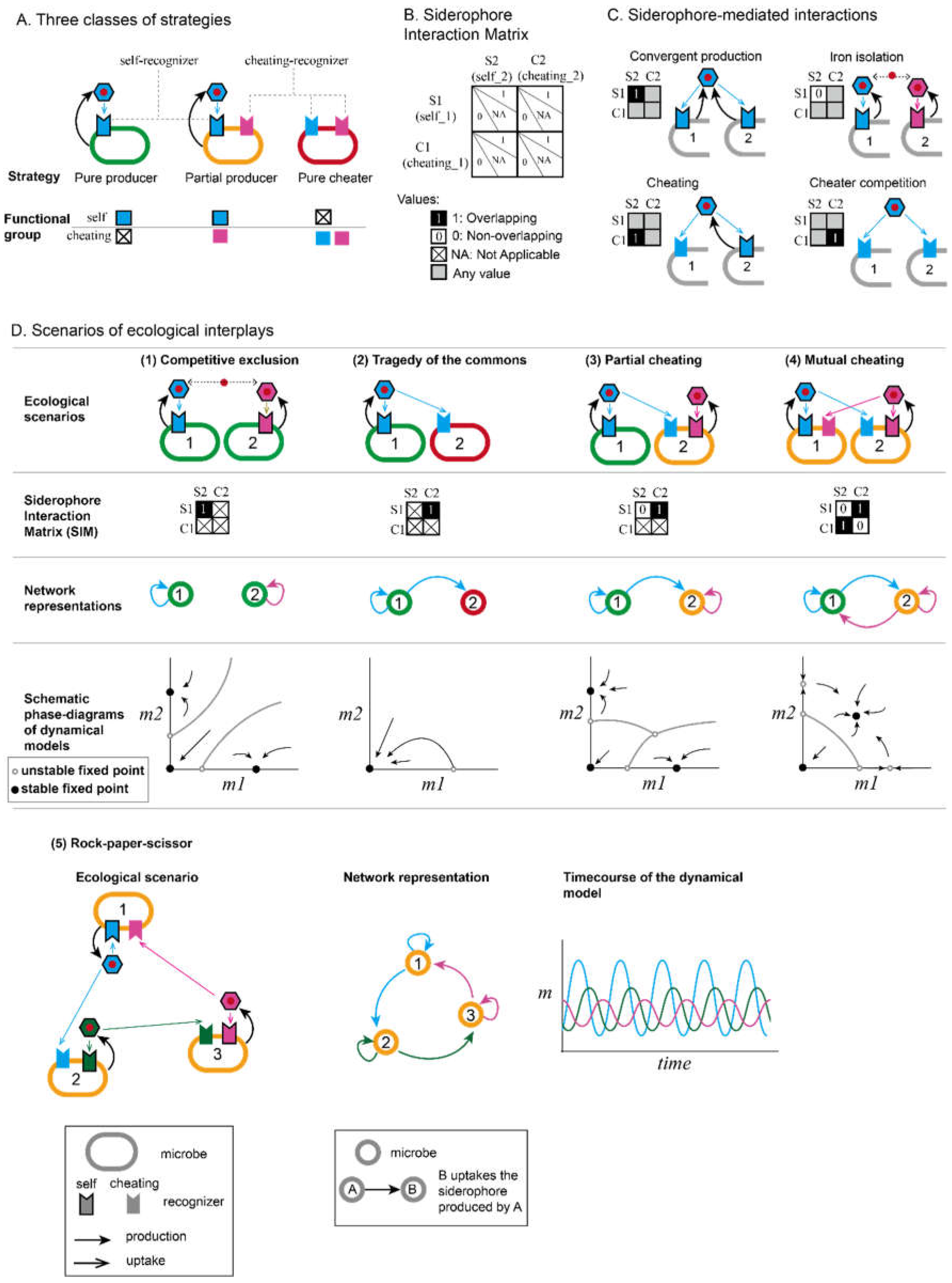
Figure 4.
Evolutionary and molecular basis, and implications of siderophore diversification.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



