Submitted:
09 July 2024
Posted:
11 July 2024
You are already at the latest version
Abstract
Grass pea (Lathyrus sativus L.) is a protein-rich crop that is resilient to various abiotic stresses, including drought. However, it is not cultivated widely for human consumption due to the neurotoxin β-N-oxalyl-L-α, β-diaminopropionic acid (β-ODAP), and its association with neurolathyrism. Though some varieties with low β-ODAP have been developed through classical breeding, the β-ODAP content is increasing due to genotype x environment interactions. This review covers grass pea nutritional quality, β-ODAP biosynthesis, mechanism of paralysis, traditional ways to reduce β-ODAP, candidate genes for boosting sulfur-containing amino acids, and the potential and targets of gene editing to reduce β-ODAP content. Recently, two key enzymes (β-ODAP synthase and β-cyanoalanine synthase) have been identified in the biosynthetic pathway of β-ODAP. We proposed four strategies through which the genes encoding these enzymes can be targeted and suppressed using CRISPR/Cas9 gene editing. Compared to its homology in Medicago truncatula, the grass pea β-ODAP synthase gene sequence and β-cyanoalanine synthase showed 62.9% and 95% similarity, respectively. The β-ODAP synthase converts the final intermediate L-DAPA into toxic β-ODAP whist β-cyanoalanine synthase converts O-Acetylserine into β-isoxazolin-5-on-2-yl-alanine. Since grass pea is low in methionine and cysteine amino acids, improvement of these amino acids is also needed to boost its protein content.
Keywords:
Grass pea
; β-ODAP
; β-ODAP synthase
; β-cyanoalanine synthase
; stress tolerant
1. Background
Grass pea (Lathyrus sativus L.) is a legume crop that belongs to the Fabaceae family. It has high adaptability and tolerance to abiotic stresses such as drought, salinity, and poor soil [1,2,3]. It is cultivated in various areas of the world, including Eastern Africa, Eurasia, North America, and South America, for both animal feed and human consumption [4]. It requires an average annual rainfall between 300 and 1500 mm [5]. In South Asia and in East Africa, grass pea is cultivated on a total of 1.5 million ha of land with an annual production of 1.2 million tons [6]. However, this figure might have increased recently as its popularity is increasing in developing countries [7] for its tolerance to severe drought.
The grass pea is a diploid plant species with 2n=2x=14 chromosomes [8]. In addition to abiotic factors, it is also resistant to several biotic stresses such as insects and diseases and has high protein content for human and animal feed [9]. Furthermore, grass pea is superior in yield and nitrogen fixation compared to other legume crops such as beans, peas, and chickpeas [10]. It has immense potential to be expanded to dry and drought-prone areas with increased salinity or an increased tendency to suffer from biotic stresses [11]. Due to its inherent tolerance to harsh weather conditions, some researchers consider grass pea a survival food in drought times [12] that has saved thousands of lives during droughts in Ethiopia, India, Bangladesh, China, and other developing countries [1]. Though not as commonly cultivated as other crops, grass pea production is increasing, especially in developing countries and drought-affected areas among which Ethiopia, India, and Bangladesh are major producers [7]. In Ethiopia, grass pea became a crop of choice as an off-season crop because of its higher tolerance to adverse environmental conditions, like drought, soil infertility, and waterlogging [13], and more recently due to severe and devastating common bean diseases such as anthracnose, rust, bacterial blight and halo blight that cause 100%, 85%, 62%, and 45% yield losses, respectively [14].
Crops like grass pea that tolerate both abiotic and biotic stress are much needed for global agriculture to ensure nutritional security in the face of climate change [15]. Despite having great economic and nutritional values, grass pea has a major drawback. It contains a major neurotoxic anti-nutritional compound known as β-N-oxalyl-L-α, β di-amino propionic acid (β-ODAP) that causes leg paralysis, that is irreversible spastic paraparesis of the lower limbs in humans [2,16,17]. Studies indicated that grass pea cultivation is limited to certain countries mainly due to the concerns of β- ODAP toxicity [18,19], which hinders its large-scale cultivation and utilization for nutritional purposes. Hence, developing grass pea varieties with low seed anti-nutritional factors but with high seed protein content would be used for human consumption and nutritional choice. Therefore, reducing β-ODAP content in seeds of this crop becomes pertinent to ensure that it continues to provide food and nutritional security to low-income communities [2,20].
Since the discovery of neuro-excitatory amino acid β-ODAP [21], several screening strategies have been employed to select grass pea varieties with low β-ODAP. However, β-ODAP toxicity is still common problem in developing countries due to the absence of varieties that are free from unwanted toxic substances. Advances in molecular genetics have shed light on some key enzymes of the β-ODAP biosynthetic pathway [22,23], opening new avenues for targeted knockout of key genes to eliminate this toxin. In this review, classical approaches used to reduce the biosynthesis of β-ODAP and its level, mechanism of action, and toxicity to humans, as well as permanent and more specific biotechnological approaches that can be applied to reduce or eliminate the β-ODAP are reviewed.
2. Nutritional Benefits and Side Effects of the Grass Pea
Grass pea is a valuable source of protein for low-income families, contributing to their daily diets and ensuring balanced nutrition. Research has shown that grass pea has great potential as a food crop, providing not only protein but also essential micronutrients for both humans and animals [24]. The protein found in grass pea contains 17 out of the 20 amino acids, with a particularly high lysine content [18,20]. In fact, its protein content ranges from 28% to 49%, surpassing other legumes like chickpeas, peas, faba beans, lupine, and kidney beans [20,25]. Additionally, grass pea seeds are rich in carbohydrates, with up to a content of 58% reported [25].
Grass pea-based dishes are not only nutritious but also delicious, making them popular in various regions including Europe, Africa, and Asia [2]. In Ethiopia, for example, grass pea is prepared in different ways, such as boiling the whole seeds (nifro), roasting them (kollo), using them in traditional sauces (shiro wot and kik wot), or consuming the green unripe seeds as a snack (eshet) by cattle keepers [16]. However, it is important to note that like other legumes, grass pea seeds have a low level of methionine, which is crucial for human nutrition and brain function [26]. Despite this limitation, the nutritional benefits and versatility of grass pea make it a valuable addition to the diets of low-income families, providing them with much-needed protein and essential nutrients.
Conversely, the grass pea seed harbors a distinctive non-protein amino acid called L-homoarginine (hArg) [27]. Studies have demonstrated that hArg is utilized to diminish the production of nitric oxide and plays a pivotal role in the management of cardiovascular ailments, as well as the inhibition of tissue-nonspecific alkaline phosphatase (TNAP), an enzyme that fosters vascular calcification [2,28,29]. Consequently, hArg serves as an endogenous safeguard for cardiovascular and metabolic factors [30]. A recent investigation also revealed that hArg supplementation prevents the enlargement of the left ventricle and preserves systolic function in a model of coronary artery disease [29]. Moreover, hArg is implicated in curtailing the expansion of cancerous tumors [31]. The concentration of hArg in human plasma is approximately 1 to 2 μmol/L [32], and it is believed to be synthesized from arginine and L-lysine by the mitochondrial enzyme argiFfigure 2nine: glycine amidino transferase [33]. Therefore, grass pea exhibits potential as an exceptional functional food ingredient [34], provided that its antinutritional factor is minimized.
The presence of β-ODAP in grass pea seeds has limited its consumption since high consumption may initiates the development of neurolathyrism, a disease that results in degeneration of pyramidal-tract neurons in the spinal cord [35] and legs cortex [36]. The neurodegenerative process results in a spastic paraparesis of the legs without affecting sensory systems [37]. Such a problem can be observed in communities where the consumption of unprocessed grass pea is common. For instance, over 2000 people were affected in Ethiopia in a single village in 1996 [38,39]. The incidence of paralysis mostly occurs during drought season when younger people eat the green ripe seeds on their way to school or when they are watching cattle. Early studies showed that neurolathyrism was caused by the long-term overconsumption of grass pea seeds, containing up to 1% of β-ODAP in the seeds [21].
According to a study conducted by Grima et al [40], farmers in Ethiopia hold the belief that the consumption of unripe grass pea seeds, particularly when combined with milk and milk products, can potentially worsen the occurrence of paralysis. The study aimed to explore the perceptions of farmers regarding the consumption and toxicity of grass pea, and the findings indicated that farmers in the study area perceive a higher risk of toxicity when consuming grass pea ‘kita’ (softened bread) or porridge with milk, butter, or oil. However, Hoque et al [41] reached a different conclusion, suggesting that the addition of milk can effectively reduce the toxicity of β-ODAP in grass pea. Nevertheless, further laboratory-based investigations are necessary to validate these claims. Moreover, Ethiopian farmers who have family members affected by β-ODAP toxicity propose that sleeping on grass pea straw overnight during the harvesting process could potentially contribute to paralysis. Additionally, farmers associate lathyrism with exposure to vapors emitted from cooking of grass pea or when harvesting on a cloudy day [42].
The grass pea toxin exists in α and β isomers [43]. The β-isomer accounts for 99% of the ODAP content and the α isomers are non-toxic [22]. The presence of β-ODAP in seeds as free amino acid especially in drought-tolerant lines is believed to be responsible for the irreversible paralysis observed in humans [2], which is more prevalent when grass pea is a main source of diet [44,45]. Studies have suggested that deficiencies of certain sulfur-rich amino acids such as cysteine and methionine increase the neurotoxicity level of ODAP [18,46]. According to Fikre et al. [16], dietary supplementation with methionine and cysteine may significantly lower the risk for neurolathyrism. Studies also indicated that the deprivation of both methionine and cysteine increased the toxicity of l-β-ODAP by 66% [47]. Hence, the development of grass pea rich in these two amino acids can be another way to reduce the toxic effects of grass pea.
3. Conventional Improvement for β-ODAP Reduction and Influence of Environmental Factors
The global importance of grass pea as a promising food source has been recognized by the Kew’s Millennium Seed Bank [2,48]. Figure 1 Illustrates specimen records of grass pea using global mapper (https://www.discoverlife.org/mp/20m?act=make_map). There is a mild increase in the cultivation of grass pea in some countries, especially in drought-prone regions, due to its remarkable ability to adapt to difficult soil conditions and emerging ecological environments [15]. While toxin-free varieties have yet to be identified, research has indicated that germplasm from South Asia contains relatively high levels of ODAP (0.7–2.4%), whereas those from North Africa, Syria, Turkey, and Cyprus exhibit significantly lower quantities of ODAP (0.02–1.2%) [49]. Despite its ability to thrive in harsh and water-limited environments, only a limited number of scientific approaches have been employed thus far to improve grass pea traits. Through classical screening methods, several grass pea accessions with low ODAP content (0.04–0.1%) have been developed in various parts of the world, including India, Bangladesh, Nepal, Ethiopia, Australia, Canada, Poland, and Turkey [28]. However, it is important to note that this approach may not provide a sustainable and reliable solution to fully address the issue of grass pea toxicity for large-scale production and consumption.
Conventional breeding is centralized fundamentally on the hybridization of selected genotypes followed by screening and evaluation of the subsequent progenies for the traits of interest. Breeding programs for grass pea improvement are being carried out in several countries, including Australia [50], Bangladesh [51], Canada [52], China [53], Ethiopia [54], India [55], Nepal [56], Syria [49], Poland [57], Italy [58], USA [59], and Chile [60]. Several varieties and lines have been developed by combining low β-ODAP (<0.1%) with high-yielding and disease-tolerant traits [61,62]. Most of the initial progress in grass pea research aimed to develop low ODAP varieties were made through direct selection from landraces and lines [18].
Various countries have conducted traditional germplasm selection to identify varieties with low ODAP (0.04–0.3%) content. The recommended varieties include Bair Khesari and Bina Khesari in Bangladesh [63,64], Gurbuz-1 in Turkey [65], Clima pink in Nepal [56], as well as LS 8246 and AC-Greenfix in Canada [52]. In 1966, India introduced the first conventionally enhanced variety called Pusa 24 (P-24), which exhibited a low ODAP content (0.3%) in its seeds [18]. Nonetheless, the instability of the low ODAP trait was a major bottleneck of the P-24 cultivar [66]. In Chile, a cultivar Quila-blanco was also developed to have lower ODAP and higher protein content [67]. In Ethiopia, a low ODAP (0.08%) variety known as Wassie was released in 2006 [68] though its ODAP content was not found to be consistent and has even increased in subsequent field and laboratory tests [42]. Though several germplasm collections and introductions were conventionally screened, varietal performances were found to be inconsistent with ODAP reduction [42] and ODAP content varies from location to location, indicating a strong G x E interaction in determining ODAP content.
Conventional breeding has been unsuccessful in reducing ODAP at a lower level due to the complex inheritance of ODAP, which is greatly influenced by the genotype, environment, and their interactions [18]. An example of this is seen in the variety Wassie, developed in Ethiopia, where the ODAP content has increased from 0.08% to over 0.2% in certain lowland areas [16]. The problem becomes even more severe when the plant is cultivated in a stressed environment [42]. Therefore, recent biotechnological approaches may provide a more dependable strategy to decrease the ODAP content and address the persistent toxicity associated with this compound.
Several studies have shown that various environmental factors such as drought, salinity, minerals, and heavy metals affect the biosynthesis of β-ODAP [69,70,71]. The study on the effect of drought, salinity, and deficiency or oversupply of micronutrients on the biosynthesis of β-ODAP by feeding callus tissue of Lathyrus sativus with precursor BIA (β-isoxazolin-5-on-2-yl-alanine) was conducted by Hoque et al. [72]. According to this study, lower concentrations of NaCl decreased the conversion of BIA into β-ODAP, while higher doses of NaCl showed the opposite effects. Mannitol has been reported to increase β-ODAP production, suggesting that drought may increase the biosynthesis of β-ODAP. Furthermore, β-ODAP accumulation in grass pea is thought to be due to the level of total free nitrogenous compounds [18]. According to Jiao et al. [71], the accumulation pattern of β-ODAP in seven-day-old grass pea seedlings grown in the nitrogen-deficient solution was higher as compared to the control. Hence, a nitrogen-poor environment might aggravate β-ODAP content in grass pea. Micronutrients such as Al3+, B3+, and Co2+ have also been reported to increase β-ODAP production, whilst Cu2+ and Zn2+ played opposite roles [72]. According to Takarz et al. [73], a higher accumulation of β-ODAP was observed in both shoots and roots of grass pea seedlings under PEG-induced drought stress. This may suggest that β-ODAP or its intermediate product may have a role in drought stress tolerance.
4. Biosynthesis of β-ODAP in Grass Pea
Despite the discovery of β-ODAP’s association with neurolathyrism 50 years ago, the complete understanding of its biosynthetic pathway remains elusive. Nevertheless, researchers have made progress in unraveling certain crucial steps in the biosynthesis of β-ODAP by using enzyme extract [74,75,76] and radioisotope studies [77]. The biosynthesis of β-ODAP is initiated with the formation of BIA from isoxazoline-5-one and cysteine, facilitated by the enzyme β-cyanoalanine synthase (CAS) under conditions of high sulfur [78]. However, under low sulfur conditions, BIA is primarily synthesized from isoxazoline-5-one and O-acetyl-serine, catalyzed by cysteine synthase (CS) [76]. According to the previous study, a gene encoding CAS and at least four other genes encoding CS have been identified from grass pea, and their activities have been characterized [79]. Both CAS and CS are important enzymes in β-ODAP synthesis. Studies by Song et al. [78] indicated that β-Cyanoalanine synthase regulates the accumulation of β-ODAP through interaction with serine acetyltransferase. An attempt to characterize the activity of genes encoding CAS and CS was recently reported [79]. Figure 2 shows a proposed biosynthetic pathway of β-ODAP reviewed by Yan et al. [80] and how its biosynthesis switches between CS and CAS based on the availability of sulfur. Figure 2B below shows L-Glutamic acid analogues that can interfere with glutamic acid action [2]. Since the trend in sulfur content on agriculture farms is decreasing [81], CS could be a key enzyme to improve cysteine in the future.
β-ODAP synthase (BOS), a crucial enzyme responsible for catalyzing the last stage of β-ODAP formation in grass pea was isolated and characterized by Glodsmith et al. [22]. Additionally, Edwards et l. [23] employed a genomics approach to identify the CS in grass pea. These discoveries have opened new possibilities for utilizing gene editing techniques to decrease the β-ODAP content in grass pea. Both CAS and BOS can be targeted for genome editing to reduce the level of β-ODAP. Although L-DAPA is stable in vitro, it is a short-lived metabolic intermediate in grass pea and cannot be extracted from the grass pea [23]. This might be due to the faster reaction of β-ODAP synthase in converting the metabolite intermediate L-DAPA into β-ODAP. According to Goldsmith et al. [22], more than 99% of the L-DAPA is converted to β-ODAP, whilst fα-ODAP is less than 1%. They suggested that mutating the highly conserved Asp166 and His162 residues in β-ODAP synthase may result in an inactive form of this enzyme. Even though mutating β-ODAP synthase may help in studying L-DAPA intermediate and the nature of its stability in grass pea, this may not address the overall trends as its substrate may also have adverse effects.
5. Mechanism of β-ODAP Action and Toxicity
According to Krogsgaard-Larsen et al. [82], L-Glutamic acid (Glu) is the major excitatory amino acid neurotransmitter in the central nervous system (CNS) that activates at least three types of receptors. These are the N-Methyl-D-aspartic acid receptor (NMDA), α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPA), and kainic acid receptor (KAR) that mediate the neurotoxicity of several naturally occurring Glu analogs including β-ODAP. β-ODAP was reported to act as an agonist of AMPA receptors, which resulted in the accumulation of intracellular calcium ions up to a toxic level [82]. Since the 1960s, efforts have been made to investigate the potential role of β -ODAP as a glutamate analogy and its interaction with glutamate receptors, and neuron signalling. However, an attempt to fully understand the molecular mechanism of neurolathyrism pathology at the cellular level is still lagging [1].
Though detailed in vivo mechanism of paralysis triggered by β-ODAP is still not fully understood, studies have used an animal cell model and animals to decipher its mechanism of action. β-ODAP acts as an excitatory amino acid in the same way as L-glutamic acid and AMPA receptors and triggers motor neuron degeneration by inducing excitotoxic cell death and increasing oxidative stress [37]. The toxin is known to disturb the mitochondrial respiration chain and inhibit the uptake of cystine thereby affecting the cells’ abilities to cope with oxidative stress [37]. One of the mechanisms of glutamate excitotoxicity is Ca2+ overload-induced cell death. Ca2+ was proposed as [37] a key player of β-ODAP-induced excitotoxicity and oxidative stress due to increased reactive oxygen species (ROS) that results in motor neuron degeneration paralysis. Accordingly, β-ODAP disturbs the cellular Ca2+ homeostatic machinery with increased Ca2+ loading in the endoplasmic reticulum (ER)-mitochondrial axis [37]. Motor neurons are very vulnerable to an increase in intracellular calcium due to the low content of Ca2+ binding proteins [83]. β-ODAP triggers Ca2+ accumulation and cell death in primary motor neurons through transient receptor potential channels and metabotropic glutamate receptors [84].
The experimental study conducted by Tan et al. [1] has widened our understanding of the molecular mechanism of β-ODAP toxicity using a cell model. Tan et al showed that human glioma cell line treated with β-ODAP resulted in decreased mitochondrial membrane potential by increasing Ca2+ in the cellular matrix and resulted in overexpression of β1 integrin on cytomembrane surface and resulted in oxidative stress due to the interruption of electron transport chain in the mitochondria. Specifically, β-ODAP combines with AMPA receptors on the cell surface and abnormally activates the receptors and intracellular homeostasis through the inflow of Ca2+[1]. Most β-ODAP affected people in Ethiopia believe that milk consumption aggravates β-ODAP toxicity (personal communication), which may be due to the high amount of calcium in milk that needs further investigation. The neurolathyrism work conducted by Khandare et al. [85] has shown that milk fed goats supplemented with 0.17 - 0.96 % of β-ODAP containing grass pea powder were affected by paralysis.
In normal physiological glutamate signaling (in the absence of β-ODAP), presynaptically released glutamate binds to surface receptors on postsynaptic neurons and activates them by inducing depolarization and an intracellular Ca2+ increase. Glutamate is then removed from the synaptic cleft by glutamate transporters (excitatory amino acid transporters; EAAT), located in the plasma membrane of both neurons and astrocytes. However, β-ODAP cannot be removed by glutamate transporters, and its binding allows more Ca2+ influx [86, 87}. In astrocytes, glutamate is converted into glutamine-by-glutamine synthetase. Glutamine is released into the extracellular space, taken up by presynaptic neurons, and converted into glutamate by glutaminase. Glutamate is then stored in presynaptic vesicles, ready for the next glutamate–glutamine cycle [86,87]. β-ODAP competes with L-glutamic acid (Glu) to bind to the AMPA receptor. Furthermore, activation of AMPA receptors is reported to enhance β1integrin expression, and further increase the focal adhesion kinase (FAK) phosphorylation level [1]. On the other hand, β-ODAP toxicity was shown to be reduced by overexpression of Glutaredoxin 1 (Grx1), an important cytosolic thiol-disulfide oxidoreductase, which reduces glutathionylated proteins to protein thiols and helps to maintain redox status of proteins during oxidative stress [88]. Figure 3 below summarizes the possible mechanism of β-ODAP action [1] and Figure 4 summarizes overall toxicity from the cellular to organismal level.
6. Reducing β-ODAP Using Traditional Food Processing Strategies
Farmers usually use traditional food processing strategies to reduce toxic compounds in grass pea. Studies have shown that pre-processing by long soaking (>18hrs) and fermentation significantly reduce the toxicant [91]. Using condiments such as onion and ginger in grass pea food preparation has been traditionally used to reduce its toxicity [92]. Furthermore, the traditional way of soaking grass pea seeds in various media has been shown to reduce the contents of β-ODAP to a varying and significant extent [91]. For instance, the loss of β-ODAP was significantly higher when soaked in boiling water (65–70%) than in cold water [91]. some chemicals like polyphenols have been suggested to lower the toxicity of grass pea though it is feasible and an effective way to apply [91]. While Tadele et al. [93] suggested that cooking grass pea for 20 minutes and roasting is important in reducing and eliminating anti-nutritional factors like β-ODAP. According to a study by Akalu et al [94], roasting and autoclaving grass pea reduced the content of β-ODAP by up to 30% and 50%, respectively. While Buta et al (2019) reported that soaking grass pea seeds under high hydrostatic pressure resulted in 36-71% reduction in β-ODAP. However, a more feasible, durable, and scalable approach is needed for farmers to reduce or eliminate toxic compounds. Hence, the production of toxic-free grass pea is paramount important to address all safety concerns related to this important stress-resilient crop without compromising its nutritional value.
7. Enhancing Sulfur-Containing Amino Acids (SCA) through Genetic Modification of Grass Pea
Despite its high protein content, grass pea is limited by its low levels of sulfur-containing amino acids (SCA) such as methionine and cysteine. As mentioned earlier, under low sulfur conditions, the intermediate BIA is predominantly synthesized from isoxazoline-5-one and O-acetyl-serine with the help of cysteine synthase. To address this limitation, genetic engineering of grass pea can be explored by introducing genes that can enhance SCA levels. This approach may lead to the development of biofortified and toxin-free grass pea. A reproducible transformation protocol has been developed for Indian grass pea accession using epicotyl segment co-cultivation with Agrobacterium strains EHA 105 and LBA 4404 [95]. The biolistic transformation of grass pea somatic embryos was also reported as a potential tool for grass pea transformation [96]. Transient genetic transformation of two grass pea varieties has also been reported in Ethiopia [97]. Therefore, by introducing candidate genes that can boost SCA through heterologous expression, the amino acid content of grass pea can be improved.
Various genes, including Brazil nut 2S protein (BN2S) [98], sunflower 2S albumin (SFA8) [99], and δ-zein protein [101], have been identified and reported as potential candidates for boosting SCA. For instance, the introduction of a methionine-rich 10-kDa δ-zein gene into transgenic maize resulted in an increase of 57.6% in methionine content [102]. Similarly, engineering of the 15-kDa δ-zein gene in soybean led to a 12-20% increase in methionine and a 15-35% increase in cysteine [103]. Another promising candidate gene is the 11 kDa δ-zein protein, which contains 32 methionine and 7 cysteine residues, accounting for 25% of the total amino acids in the protein [104]. Similarly, the introduction of the β-zein gene into soybean resulted in an increase of up to 15% in methionine content [105].
The utilization of enzymatic genes such as ATP sulfurylase, which aids in the conversion of sulfate to adenosine 5′-phosphosulfate (APS), may play a role in regulating sulfur metabolism and improving SCA. The overexpression of ATP sulfurylase resulted in significant improvements, with a 37–52% increase in protein-bound cysteine content and a 15–19% increase in methionine content [106]. In a study conducted by Kin et al. [107], cytosolic isoform of O-acetylserine sulfhydrylase (OASS) was introduced into transgenic soybeans, leading to a noteworthy 22-32% elevation in free cysteine levels in the soybeans. Furthermore, Avraham et al. [108] conducted another study where they introduced Arabidopsis cystathionine γ-synthase (AtCGS) into alfalfa plants, resulting in a remarkable 32-fold increase in methionine and a 2.6-fold increase in free cysteine, indicating a potential to increase the SCA content in grass pea, thereby generating biofortified grass pea lines.
8. The Potential of Genome Editing Approach for Antinutritional Factors in Crops
Genome editing in plants has opened new opportunities to incorporate novel traits beneficial for human health and the environment by eliminating antinutritional factors. Genome editing encompasses techniques such as oligonucleotide-directed mutagenesis (ODM) and site-directed nucleases (SDNs) like zinc finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), mega-nucleases and clustered regularly interspaced short palindromic repeats/CRISPR-associated (CRISPR/Cas) techniques [109]. Unlike the random traditional mutagenesis approach, site-directed nucleases (SDN) are playing a pivotal role in precise genome edition as it is faster and more targeted [110]. Depending on the nature of the edit, genome editing with the SDN process can be categorized under SDN1, SDN2, and SDN3 [111]. The SDN1 involves the unguided repair of a targeted double-strand break (DSB) with small indels, and joins ends in a non-homologous manner. The repair of this break can lead to a mutation causing gene silencing through loss of function without the addition of foreign DNA but due to a shift in the reading frame. SDN2 involves a template-guided repair of a targeted DSB, typically short single-stranded DNA while SDN3 involves a template-guided repair of a targeted DSB, typically with double-stranded DNA with longer genetic elements [112]. Studies indicate that SDN1 and SDN2 do not require foreign DNA for genome editing whilst the SDN3 require foreign DNA segment [111, 112 and 113]].
CRISPR-associated Cas9 endonuclease is becoming available in genome editing technology that enables precise modification in the genome. CRISPR/Cas9 has advantages over other gene editing because it is relatively less costly, more precise, the operation is easier, and more efficient, allowing researchers to induce targeted knockouts [113,114]. In recent years, CRISPR/ Cas9 has been used successfully for genome editing in more than 20 crop plants to alter various characteristics such as increasing yield, promoting growth, removing unwanted anti-nutritional factors [115,116,117]. This method can be used to create knockouts of non-important enzymatic functions which in many cases has been difficult to achieve with the traditional genetic engineering approach. In sweet potato, CRISPR/ Cas9 has been used to knockout SBEII and increase the amylose percentage [118,119]. High-amylose and resistant starch wheat was also generated through suppression of TaSBEIIa in a modern winter and spring wheat using CRISPR/Cas9 [120,121]. Very recently, cytochrome P450 genes CYP79D1 and CYP79D2 that accumulate toxic cyanogen in cassava were successfully edited using CRISPR-Cas9-mediated knockout [122].
Phytic acid (PA) is antinutrient in cereal grains that inhibit the absorption of micronutrients like iron and zinc in humans, leading to malnutrition. While PA is a significant source of phosphorus in plants, it is considered detrimental for monogastric animals, including humans, due to its negative effects on essential mineral uptake [123]. Through targeting of the enzyme Inositol-pentakisphosphate 1-kinase (IPK1) which catalysis the final step of PA biosynthesis in soybean (Glycine max L.) using two guide RNAs directed at the second and third exons of GmIPK1, the PA content was reduced by 25% without adverse effects on plant growth or seed development [123]. Likewise, CRISPR-Cas9 was applied to tetraploid Brassica napus to knockout three functional paralogs of BnITPK which that resulted in a higher reduction of PA (~35%) in triple mutants [124].
Despite being an essential amino acid, crops like wheat contain elevated levels of free asparagine in their grains, which can form acrylamide during high-temperature cooking processes, a known carcinogenic compound [124]. By introducing four gRNAs targeting the wheat asparagine synthetase gene (TaASN2) into wheat embryos using CRISPR/Cas9, Raffan et al [125] reported reduction of free asparagine by approximately 90%. Asparagine is also precursor of β-ODAP biosynthesis.
In cotton (Gossypium hirsutum) gossypol is a polyphenol which accumulates in the seeds. Gossypol can lead to acute poisoning when consumed by various animals such as broiler chicks, pigs, dogs, sheep, and goats due to its antispermatogenic activity, raising health concerns [126]. The utilization of CRISPR-Cas9 has been shown to successfully knock out dirigent proteins genes GhDIR5 and GhDIR6, reducing cotton seed toxicity without affecting other phytoalexins [127]. Additionally, CRISPR-Cas9 was employed to significantly decrease nicotine levels in tobacco by 99.6%, aiming to develop nicotine-free tobacco by targeting the flavoproteins of the berberine bridge enzyme-like (BBL) gene responsible for the final oxidation step in nicotine biosynthesis [128]. Furthermore, CRISPR-Cas9 was effectively used in the medicinal plant comfrey (Symphytum officinale) to eliminate toxic pyrrolizidine alkaloids by introducing detrimental mutations into the hss gene encoding homospermidine synthase (HSS) [129]. In legumes, the CRISPR/Cas9-mediated edition of the GmFATB1 gene significantly reduced the amount of saturated fat in soybean [130] and the level of trypsin inhibitor by targeting KTI1 and KTI3 genes [131] in soybean. Hence, CRISPR/ Cas9 can be applied to other crops such as grass to reduce or eliminate anti-nutrient content.
8.1. The Potential of CRISPR/Cas9 to Target β-ODAP Biosynthesis in Grass Pea
The availability of draft genome sequence of grass pea [23,114,132] can now allow researchers to develop a novel strategy to reduce β-ODAP toxicity. It is becoming clearer that enzymes involved in the biosynthesis of β-ODAP are potential candidates for CRISPR/Cas9-based genome editing. β-ODAP synthase (BOS) gene (AC: MT457411) is a 1,320 bp mRNA encoding 439 amino acids. Analysis of conserved domains of BOS has shown that amino acids from position 3-434 have a role in transferase activity (E value =5.11e-75). Hence, a single nucleotide deletion in this gene can lead to frameshift blocking the synthesis of β-ODAP from L-DAPA and oxalyl-CoA following the first strategy. We propose four potential strategies to decrease or eliminate β-ODAP biosynthesis in grass pea. The first strategy is to target the BOS gene only. The second strategy is to target the CAS gene only. The third strategy is to simultaneously target both BOS and CAS genes. The fourth strategy is replacing specific amino acids in either BOS or CAS, either separately or together, to partially reduce β-ODAP biosynthesis. The latter strategy is suggested if the complete or partial knockout of β-ODAP biosynthesis has undesirable phenotypes such as sensitivity to abiotic stresses like drought. In first strategy (targeting BOS), further accumulation of the precursor L-DAPA must also be considered to monitor the level of toxic intermediate. In vitro toxicological effects of isoxazoline amino acids of Lathyrus sativus was conducted by Riepe et al. [133]. Their finding suggested that 2000µM of BIA resulted in cell damage and neurodegeneration in mouse cortical similar to 50µM β-ODAP in a concentration-dependent manner. This is a good indication that the intermediate BIA is 40X less toxic when compared to β-ODAP and may not be a major concern. However, further study is needed determine the level of toxin precursor accumulation after gene knockout. The recent non-peer reviewed bioRxiv article has reported an attempt to reduce β-ODAP by targeting BAHD-AT3 gene [134].
If L-DAPA and BIA show significant toxicity as the intermediate precursors, β-cyanoalanine synthase (CAS) can be another target for gene editing. CAS has been shown to synthesize the intermediate BIA from isoxazoline-5-one and cysteine catalyzed [46] under high-sulfur conditions. In grass pea the CAS gene (AC: KJ563188) is a 1146 bp complete mRNA that encodes 381 amino acids. Cloning of the CAS gene from L. sativus (GenBank: KJ563188) which has 99.74 % identity with KJ563188 at the protein level was reported by Xu et al. [79]. Conserved domain analysis (data not shown) has shown that this gene has a dual function (cysteine synthase and/ L-3-cyanoalanine synthase) and may be able to block L-DAPA synthesis both at the low and high sulfur environment. Figure 5 depicts the possible gene targets for CRISPR/Cas9.
To further understand the similarity of BOS and CAS enzymes in seven legumes and a rosaceae family, we computed multiple sequence alignment using COBALT (constraint-based alignment tool for multiple protein sequences) which computes multiple protein sequence alignment using conserved domain and local sequence similarity information [135]. Figure 6 shows the similarity of grass pea BOS and CAS with homologs in other plant species.
As indicated in Figure 6 above, CAS has shown strong similarity among aligned proteins. The Lathyrus sativus CAS and a model Medicago truncatula showed 95% similarity followed by CAS of Glycine max and Cajanus cajani (94%). The lowest similarity was observed between the Prunus persica and Sesbania bispinosa CAS genes. Furthermore, remarkable variability was shown in the conserved domain of BOS among selected plants. Using the BOS gene, L. sativum, and M. truncatula has a similarity of 62.9% at the protein level indicating the existence of significant variability from model legumes. The BOS of C. cajani and G. max has shown 73.5 % similarity followed by M. truncatula and Cicer arietinum (72.8%). The variability of β-ODAP in grass pea might have resulted in enhanced enzyme activity to convert the intermediate L-DAPA into more toxic β-ODAP selectively in grass pea which needs further empirical study. The scaled and unscaled phylogenetic tree (Figure 7) depicts the genetic relatedness of those two enzymes in different crop plants including model legume. The trees were constructed using MEGA 11 [136] and edited by ITOLv5 (Interactive tree of life) [137]. Overall, the β-ODAP of L. sativus and the model legume M. truncatulaare are strongly related to each other than to other species used fore tree construction.
9. Conclusion and Prospects
Grass pea is stress-tolerant, nutritious crop that has a high level of the antinutritional factor β-ODAP. This review summaries the benefit of grass pea in adapting to abiotic stresses affected ecological zones as future crop of choice in ever changing global environment. As grass pea is poor in sulfur containing amino acids (SCA), the ideal genes capable of boosting SCA were also highlighted. The biosynthesis of toxic compounds in grass pea and molecular mechanisms of their toxicity, and the potential of CRISPR/Cas9 technology in reducing grass pea toxicity has been discussed. To fully benefit from grass pea, toxic compounds need to be reduced or eliminated by targeting enzymes responsible for β-ODAP production via CRISPR/Cas9 genome editing approach. Researchers should take into consideration equally evaluating the level of intermediate products such as L-DAPA and BIA after BOS knockout. There is a need also to target upstream enzymes such as CAS that are involved in the biosynthesis of intermediate. Knocking out of BOS and/or CAS enzymes may help in reducing the β-ODAP biosynthesis. Given its high-yielding potential and stress tolerance, grass pea production needs to be expanded to increase food security in the face of global change. However, the use of modern biotechnologies such as CRISPR/Cas9 is crucial to eliminate antinutrient components for this crop. We believe this is a comprehensive review that covers the limitations and opportunities of grass pea supported by bioinformatics analysis and can be the pertinent document for improving grass pea-related species.
Author Contributions
A.B-A conceived the idea collected the resources, drafted the manuscript, and edited the final version, D. G-T collected the resources and drafted the manuscript. A. L-A edited the draft and incorporated extra resources and edited the final version. All authors read and agreed on the final manuscript.
Funding
This work was supported by NIH (grant #1SC2GM144193-01) to A L-O.
Ethics declarations
Not applicable.
Consent of publication
Not applicable.
Data availability
All data used in this manuscript are included in the main manuscript.
Competing interests
The authors declare no conflict of interest.
Acknowledgments
The author acknowledges Dr. Tilahun Ayane for Chemdrew and the University of North Carolina at Greensboro for an assistantship position to the first author.
References
- Tan, R.-Y.; Xing, G.-Y.; Zhou, G.-M.; Li, F.-M.; Hu, W.-T.; Lambein, F.; Xiong, J.-L.; Zhang, S.-X.; Kong, H.-Y.; Zhu, H.; et al. Plant toxin β-ODAP activates integrin β1 and focal adhesion: A critical pathway to cause neurolathyrism. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Lambein, F.; Travella, S.; Kuo, Y.-H.; Van Montagu, M.; Heijde, M. Grass pea (Lathyrus sativus L.): orphan crop, nutraceutical or just plain food? Planta 2019, 250, 821–838. [Google Scholar] [CrossRef]
- Gonçalves, L.; Rubiales, D.; Bronze, M.R.; Patto, M.C.V. Grass Pea (Lathyrus sativus L.)—A Sustainable and Resilient Answer to Climate Challenges. Agronomy 2022, 12, 1324. [Google Scholar] [CrossRef]
- Smartt, J. (1990). Pulses of the classical world. Grain legumes: evaluation and genetic resources. Cambridge Univ. Press, Cambridge, 190-200.
- Abdipour, M.; Vaezi, B.; Khademi, K.; Ghasemi, S. An optimized artificial intelligence approach and sensitivity analysis for predicting the biological yield of grass pea (Lathyrus sativus L.). Arch. Agron. Soil Sci. 2019, 66, 1909–1924. [Google Scholar] [CrossRef]
- Sammour RH. (2014) Genetic diversity in Lathyrus sativus L. germplasm. Res Rev BioSci 8:325–336.
- Gupta, P.; Udupa, S.M.; Gupta, D.S.; Kumar, J.; Kumar, S. Population structure analysis and determination of neurotoxin content in a set of grass pea (Lathyrus sativus L.) accessions of Bangladesh origin. Crop. J. 2018, 6, 435–442. [Google Scholar] [CrossRef]
- Verma, S.C.; Ohri, D. Chromosome and nuclear phenotype in the legume Lathyrus sativus L. CYTOLOGIA 1979, 44, 77–90. [Google Scholar] [CrossRef]
- Hanbury, C.; White, C.; Mullan, B.; Siddique, K. A review of the potential of Lathyrus sativus L. and L. cicera L. grain for use as animal feed. Anim. Feed. Sci. Technol. 2000, 87, 1–27. [Google Scholar] [CrossRef]
- Patto, M.C.V.; Skiba, B.; Pang, E.C.K.; Ochatt, S.J.; Lambein, F.; Rubiales, D. Lathyrus improvement for resistance against biotic and abiotic stresses: From classical breeding to marker assisted selection. Euphytica 2006, 147, 133. [Google Scholar] [CrossRef]
- Almeida, N. , Rubiales, D., Vaz Patto, M. (2015). Grass Pea. In: De Ron, A. (eds) Grain Legumes. Handbook of Plant Breeding, vol 10. Springer, New York, NY. [CrossRef]
- Shiferaw, E.; Pè, M.E.; Porceddu, E.; Ponnaiah, M. Exploring the genetic diversity of Ethiopian grass pea (Lathyrus sativus L.) using EST-SSR markers. Mol. Breed. 2011, 30, 789–797. [Google Scholar] [CrossRef]
- Dadi, L. , Teklewold, H., Aw-Hassan, A., Moneim, A.A., Bejiga, G., 2003. The Socioeconomic Factors Affecting Grass Pea Consumption and Influence of Lathyrism in Ethiopia. Integr. National Resources Management Res. Report Series, No. 4. ICARDA, Aleppo, Syria, pp. 7–11.
- Gabrekiristos E, Wondimu M (2022) Emerging and Reemerging Diseases of Common Bean (Phaseolus vulgaris L.) in major production areas: In the case of Ethiopia. J Plant Pathol Microbiol. 13:619.
- Sarkar, A.; Emmrich, P.M.F.; Sarker, A.; Zong, X.; Martin, C.; Wang, T.L. (2019). Grass pea: Remodelling an ancient insurance crop for climate resilience. In Genomic Designing of Climate-Smart Pulse Crops; Kole, C., Ed.; Springer International Publishing: Cham, Switzerland, pp.
- Fikre, A.; Van Moorhem, M.; Ahmed, S.; Lambein, F.; Gheysen, G. Studies on neurolathyrism in Ethiopia: Dietary habits, perception of risks and prevention. Food Chem. Toxicol. 2011, 49, 678–684. [Google Scholar] [CrossRef]
- Arslan, M.; Basak, M.; Aksu, E.; Uzun, B.; Yol, E. Genotyping of Low β-ODAP Grass Pea (Lathyrus sativus L.) Germplasm with EST-SSR Markers. Braz. Arch. Biol. Technol. 2020, 63. [Google Scholar] [CrossRef]
- Das, A.; Parihar, A.K.; Barpete, S.; Kumar, S.; Gupta, S. Current Perspectives on Reducing the β-ODAP Content and Improving Potential Agronomic Traits in Grass Pea (Lathyrus sativus L.). Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Banerjee, J. , Das, A., Parihar, A.K., Sharma, R., Pramanik, K., Barpete, S. (2022). Genomic Designing Towards Development of Abiotic Stress Tolerant Grass Pea for Food and Nutritional Security. In: Kole, C. (eds) Genomic Designing for Abiotic Stress Resistant Pulse Crops. Springer, Cham. [CrossRef]
- Rizvi, A.H.; Sarker, A.; Dogra, A. Enhancing grasspea (Lathyrus sativus L.) production in problematic soils of South Asia for nutritional security. Indian J. Genet. Plant Breed. (The) 2016, 76, 583. [Google Scholar] [CrossRef]
- Rao, S. L. , Adiga, P. R. & Sarma, P. S. (1964). The Isolation and Characterization of Beta-N-Oxalyl-L-Alpha,Beta-Diaminopropionic Acid: A Neurotoxin from the Seeds of Lathyrus Sativus. Biochemistry 3, 432–436.
- Goldsmith, M.; Barad, S.; Knafo, M.; Savidor, A.; Ben-Dor, S.; Brandis, A.; Mehlman, T.; Peleg, Y.; Albeck, S.; Dym, O.; et al. Identification and characterization of the key enzyme in the biosynthesis of the neurotoxin β-ODAP in grass pea. J. Biol. Chem. 2022, 298, 101806. [Google Scholar] [CrossRef]
- Edwards, A.; Njaci, I.; Sarkar, A.; Jiang, Z.; Kaithakottil, G.G.; Moore, C.; Cheema, J.; Stevenson, C.E.M.; Rejzek, M.; Novák, P.; et al. Genomics and biochemical analyses reveal a metabolon key to β-L-ODAP biosynthesis in Lathyrus sativus. Nat. Commun. 2023, 14, 1–16. [Google Scholar] [CrossRef]
- Rathi, D.; Chakraborty, S.; Chakraborty, N. Grasspea, a critical recruit among neglected and underutilized legumes, for tapping genomic resources. Curr. Plant Biol. 2021, 26, 100200. [Google Scholar] [CrossRef]
- Urga k, Fufa H, Biratu E, Husain A (2005). Evaluation of Lathyrus sativus cultivated in Ethiopia for proximate composition, mineral and b-ODAP and antinutritional components. African Journal of Food Agriculture and Nutritional Development, 5: 1-15.
- Arslan M (2017) Diversity for vitamin and amino acid content in grass pea (Lathyrus sativus L.). Legume Research 40:803–810.
- Rao, S.L.N.; Ramachandran, L.K.; Adiga, P.R. The Isolation and Characterization of L-Homoarginine from Seeds of Lathyrus sativus*. Biochemistry 1963, 2, 298–300. [Google Scholar] [CrossRef]
- Rao, S. A look at the brighter facets of β-N-oxalyl-l-α,β-diaminopropionic acid, homoarginine and the grass pea. Food Chem. Toxicol. 2011, 49, 620–622. [Google Scholar] [CrossRef]
- Rodionov, R.N.; Begmatov, H.; Jarzebska, N.; Patel, K.; Mills, M.T.; Ghani, Z.; Khakshour, D.; Tamboli, P.; Patel, M.N.; Abdalla, M.; et al. Homoarginine Supplementation Prevents Left Ventricular Dilatation and Preserves Systolic Function in a Model of Coronary Artery Disease. J. Am. Hear. Assoc. 2019, 8, e012486. [Google Scholar] [CrossRef]
- Tsikas D, Wu G. (2015). Homoarginine, arginine, and relatives: analysis, metabolism, transport, physiology, and pathology. Amino Acids. 47:1697–1702.
- Jammulamadaka, N.; Burgula, S.; Medisetty, R.; Ilavazhagan, G.; Rao, S.L.N.; Singh, S.S. β-N-Oxalyl-l-α,β-diaminopropionic acid regulates mitogen-activated protein kinase signaling by down-regulation of phosphatidylethanolamine-binding protein 1. J. Neurochem. 2011, 118, 176–186. [Google Scholar] [CrossRef]
- May, M.; Kayacelebi, A.A.; Batkai, S.; Jordan, J.; Tsikas, D.; Engeli, S. Plasma and tissue homoarginine concentrations in healthy and obese humans. Amino Acids 2015, 47, 1847–1852. [Google Scholar] [CrossRef]
- Tsikas, D.; Bollenbach, A.; Hanff, E.; Kayacelebi, A.A. Asymmetric dimethylarginine (ADMA), symmetric dimethylarginine (SDMA) and homoarginine (hArg): the ADMA, SDMA and hArg paradoxes. Cardiovasc. Diabetol. 2018, 17, 1–4. [Google Scholar] [CrossRef]
- Llorent-Martínez, E.J.; Zengin, G.; Fernández-de Córdova, M.L.; Bender, O.; Atalay, A.; Ceylan, R.; Mollica, A.; Mocan, A.; Uysal, S.; Guler, G.O.; et al. Traditionally Used Lathyrus Species: Phytochemical Composition, Antioxidant Activity, Enzyme Inhibitory Properties, Cytotoxic Effects, and in silico Studies of L. czeczottianus and L. nissolia. Front. Pharmacol. 2017, 8, 83. [Google Scholar] [CrossRef]
- Striefler M, Cohn D, Hirano A, Schujman E. (1977). The central nervous system in a case of neurolathyrism Neurology, 27 (12) (1977), pp. 1176-1178.
- Haimanot H, Kidane Y, Wuhib E, Kalissa A, Alemu T, Zein Z, Spencer P. (1990). Lathyrism in rural northwestern Ethiopia: a highly prevalent neurotoxic disorder Int. J. Epidemiol., 19 (3) (1990), pp.
- Van Moorhem, M.; Lambein, F.; Leybaert, L. Unraveling the mechanism of β-N-oxalyl-α,β-diaminopropionic acid (β-ODAP) induced excitotoxicity and oxidative stress, relevance for neurolathyrism prevention. Food Chem. Toxicol. 2011, 49, 550–555. [Google Scholar] [CrossRef]
- Getahun, H.; Mekonnen, A.; TekleHaimanot, R.; Lambein, F. Epidemic of neurolathyrism in Ethiopia. Lancet 1999, 354, 306–307. [Google Scholar] [CrossRef]
- Woldeamanuel, Y.W.; Hassan, A.; Zenebe, G. Neurolathyrism: two Ethiopian case reports and review of the literature. J. Neurol. 2011, 259, 1263–1268. [Google Scholar] [CrossRef]
- Girma, A.; Tefera, B.; Dadi, L. Grass Pea and Neurolathyrism: Farmers’ perception on its consumption and protective measure in North Shewa, Ethiopia. Food Chem. Toxicol. 2011, 49, 668–672. [Google Scholar] [CrossRef]
- Hoque, H., Jamali, S., Akther, J., & Prodhan, S. H. (2012). Computational analysis of milk sources from different domestic animals as supplementary food source to protect Lathyrism. International Journal of Biosciences, 2(7), 74-82.
- Girma, D.; Korbu, L. Genetic improvement of grass pea (Lathyrus sativus) in Ethiopia: an unfulfilled promise. Plant Breed. 2012, 131, 231–236. [Google Scholar] [CrossRef]
- Bell, E.; O'Donovan, J.P. The isolation of α- and γ-oxalyl derivatives of α,γ-diaminobutyric acid from seeds of Lathyrus latifolius, and the detection of the α-oxalyl isomer of the neurotoxin α-amino-β-oxalylaminopropionic acid which occurs together with the neurotoxin in this and other species. Phytochemistry 1966, 5, 1211–1219. [Google Scholar] [CrossRef]
- Patto, M.C.V.; Rubiales, D. Lathyrus diversity: available resources with relevance to crop improvement – L. sativus and L. cicera as case studies. Ann. Bot. 2014, 113, 895–908. [Google Scholar] [CrossRef]
- Girish Prasad Dixit, Ashok Kumar Parihar, Abhishek Bohra, Narendra Pratap Singh (2016) Achievements and prospects of grass pea (Lathyrus sativus L.) provement for sustainable food production., ICAR-Indian Institute of Pulses Research (IIPR), Kanpur 208024, India.
- Song, Y.; Wang, L.; Liu, F.; Jiao, C.; Nan, H.; Shen, X.; Chen, H.; Li, Y.; Lei, B.; Jiang, J.; et al. β-Cyanoalanine Synthase Regulates the Accumulation of β-ODAP via Interaction with Serine Acetyltransferase in Lathyrus sativus. J. Agric. Food Chem. 2021, 69, 1953–1962. [Google Scholar] [CrossRef]
- Kusama-Eguchi, K.; Yoshino, N.; Minoura, A.; Watanabe, K.; Kusama, T.; Lambein, F.; Ikegami, F. Sulfur amino acids deficiency caused by grass pea diet plays an important role in the toxicity of l-β-ODAP by increasing the oxidative stress: Studies on a motor neuron cell line. Food Chem. Toxicol. 2011, 49, 636–643. [Google Scholar] [CrossRef]
- Gonçalves, L.; Rubiales, D.; Bronze, M.R.; Patto, M.C.V. Grass Pea (Lathyrus sativus L.)—A Sustainable and Resilient Answer to Climate Challenges. Agronomy 2022, 12, 1324. [Google Scholar] [CrossRef]
- Kumar, S.; Bejiga, G.; Ahmed, S.; Nakkoul, H.; Sarker, A. Genetic improvement of grass pea for low neurotoxin (β-ODAP) content. Food Chem. Toxicol. 2010, 49, 589–600. [Google Scholar] [CrossRef]
- McCutchan, J. S. (2003). A brief history of grasspea and its use in crop improvement. Lath. Newsl. 3, 18–23.
- Rahman, M.; Ali, M.; Alam, F.; Banu, M.; Faruk, M.; Bhuiyan, M. Biocontrol of Foot and Root Rot Disease of Grasspea (Lathyrus sativus) by Dual Inoculation with Rhizobium and Arbuscular Mycorrhiza. Bangladesh J. Microbiol. 2019, 34, 109–117. [Google Scholar] [CrossRef]
- Campbell, C.G.; Briggs, C.J. Registration of Low Neurotoxin Content Lathyrus Germplasm LS 8246. Crop. Sci. 1987, 27, 821–821. [Google Scholar] [CrossRef]
- Yang, H. M., & Zhang, X. Y. (2005). Considerations on the reintroduction of grass pea in China. Lathyrus Lathyrism Newsletter, 4, 22-26.
- Tadesse, W.; Bekele, E. Variation and association of morphological and biochemical characters in grass pea (Lathyrus sativus L.). Euphytica 2003, 130, 315–324. [Google Scholar] [CrossRef]
- Santha, I. M. , and Mehta, S. L. (2001). Development of low ODAP somaclones of Lathyrus sativus. Lath. Lath. Newsl. 2:42.
- Yadav, C. R. (1996). “Genetic evaluation and varietal improvement of grasspea in Nepal,” in: Lathyrus Genetic Resources in Asia: Proceedings of a Regional Workshop (Raipur:.
- Grela, E.R.; Rybiński, W.; Klebaniuk, R.; Matras, J. Morphological characteristics of some accessions of grass pea (Lathyrus sativus L.) grown in Europe and nutritional traits of their seeds. Genet. Resour. Crop. Evol. 2010, 57, 693–701. [Google Scholar] [CrossRef]
- Granati, E.; Bisignano, V.; Chiaretti, D.; Crinò, P.; Polignano, G.B. Characterization of Italian and exotic Lathyrus germplasm for quality traits. Genet. Resour. Crop. Evol. 2003, 50, 273–280. [Google Scholar] [CrossRef]
- Krause, D., and Krause, I. (2003). New green manuring Lathyrus sativus variety AC Greenfix available in USA. Lath. Lath. Newsl. 3, 13–14.
- Mera, M. , Tay, J., France, A., Montenegro, A., Espinoza, N., and Gaete, N. (2003). Luanco-INIA, a large-seeded cultivar of Lathyrus sativus released in Chile. Lath. Lath. Newsl. 3:26.
- Dixit, G.P.; Parihar, A.K.; Bohra, A.; Singh, N.P. Achievements and prospects of grass pea ( Lathyrus sativus L.) improvement for sustainable food production. Crop. J. 2016, 4, 407–416. [Google Scholar] [CrossRef]
- Sen Gupta, D. , Barpete, S., Kumar, J., and Kumar, S. (2021). “Breeding for better grain quality in lathyrus,” in Breeding for Enhanced Nutrition and Bio-Active Compounds in Food Legumes, eds D. S. Gupta, S. Gupa, and J. Kumar (Switzerland: Springer), 131–156. [CrossRef]
- Malek, M. A. , Sarwar, C. D. M., Sarker, A., and Hassan, M. S. (1996). “Status of grass pea research and future strategy in Bangladesh”, in Lathyrus Genetic Resources in Asia, ed. R. K., Arora, P. N., Mathur, K. W., Riley, Y., Adham (Rome: International Plant Genetic Resources Institute), 7–12.
- Kumar, S. , Gupta P., Barpete S., Sarker A., Amri, A., Mathur, P. N., et al. (2013). “Grass pea,” in Improvement, Genetic and Genomic Resources for Grain Legume eds M. Singh, H. D. Upadhyaya, I. S. Bisht (London: Elsevier), 269-293. [CrossRef]
- ICARDA (2007). ICARDA Annual report. International Center for Agricultural Research in the Dry Areas.
- Mehta, S. L. , and I. M. Santha, 2008: Somaclonal variation and genetic transformation in Lathyrus sativus. In: P. B. Kirti (eds), Handbook of New Technologies for Genetic Improvement of Legumes, 177— 184. CRS Press, New York.
- Campbell, C.G.; Mehra, R.B.; Agrawal, S.K.; Chen, Y.Z.; El Moneim, A.M.A.; Khawaja, H.I.T.; Yadov, C.R.; Tay, J.U.; Araya, W.A. Current status and future strategy in breeding grasspea (Lathyrus sativus). Euphytica 1993, 73, 167–175. [Google Scholar] [CrossRef]
- MoARD, 2008: Animal and plant health regulatory directorate. Crop Variety Register, 11. Addis Ababa, Ethiopia.
- Hussain, M. , Chowdhury, B., Hoque, R., Lambein, F., 1997. Effect of water stress, salinity, interaction of cations, stage of maturity of seeds and storage devices on the ODAP content of Lathyrus sativus. In: Tekle Haimanot, R., Lambein, F. (Eds.), Lathyrus and Lathyrism a Decade of Progress. University of Ghent, Belgium, pp.
- Xing, G.; Cui, K.; Li, J.; Wang, Y.; Li, Z. (2001). Water stress and accumulation of beta- N-oxalyl-L-alpha,beta-diaminopropionic acid in grass pea (Lathyrus sativus). J. Agric. Food Chem. 49, 216–220.
- Jiao, C.J. , Xu, Q.L., Wang, C.Y., Li, Z.X., Wang, Y.F., 2006. Accumulation pattern of toxin beta-ODAP during lifespan and effect of nutrient elements on beta-ODAP content in Lathyrus sativus seedlings. J. Agric. Sci. 144, 369–375.
- Haque, R.M.; Kuo, Y.-H.; Lambein, F.; Hussain, M. Effect of environmental factors on the biosynthesis of the neuro-excitatory amino acid β-ODAP (β-N-oxalyl-l-α,β-diaminopropionic acid) in callus tissue of Lathyrus sativus. Food Chem. Toxicol. 2010, 49, 583–588. [Google Scholar] [CrossRef]
- Tokarz, B.; Wójtowicz, T.; Makowski, W.; Jędrzejczyk, R.J.; Tokarz, K.M. What is the Difference between the Response of Grass Pea (Lathyrus sativus L.) to Salinity and Drought Stress?—A Physiological Study. Agronomy 2020, 10, 833. [Google Scholar] [CrossRef]
- Malathi, K. , Padmanaban, G., Rao, S. L., and Sarma, P. S. (1967) Studies on the biosynthesis of beta-N-oxalyl-L-alpha, beta-diaminopropionic acid, the Lathyrus sativus neurotoxin. Biochim. Biophys. Acta 141, 71–78.
- Malathi, K., Padmanaban, G., and Sarma, P. S. (1970) Biosynthesis of beta-N-Oxalyl-L-Alpha,Beta-Diaminopropionic acid, Lathyrus-sativus neurotoxin. Phytochemistry 9, 1603±.
- Ikegami, F.; Ongena, G.; Sakai, R.; Itagaki, S.; Kobori, M.; Ishikawa, T.; Kuo, Y.-H.; Lambein, F.; Murakoshi, I. Biosynthesis of β-(isoxazolin-5-on-2-yl)-l-alanine by cysteine synthase in Lathyrus sativus. Phytochemistry 1993, 33, 93–98. [Google Scholar] [CrossRef]
- Kuo, J. M. , & Raushel, F. M. (1994) Biochemistry 33, 4265—4272.
- Song, Y.; Wang, L.; Liu, F.; Jiao, C.; Nan, H.; Shen, X.; Chen, H.; Li, Y.; Lei, B.; Jiang, J.; et al. β-Cyanoalanine Synthase Regulates the Accumulation of β-ODAP via Interaction with Serine Acetyltransferase in Lathyrus sativus. J. Agric. Food Chem. 2021, 69, 1953–1962. [Google Scholar] [CrossRef]
- Xu, Q.; Liu, F.; Chen, P.; Jez, J.M.; Krishnan, H.B. β-N-Oxalyl-l-α,β-diaminopropionic Acid (β-ODAP) Content in Lathyrus sativus: The Integration of Nitrogen and Sulfur Metabolism through β-Cyanoalanine Synthase. Int. J. Mol. Sci. 2017, 18, 526. [Google Scholar] [CrossRef]
- Yan, Z.-Y.; Spencer, P.S.; Li, Z.-X.; Liang, Y.-M.; Wang, Y.-F.; Wang, C.-Y.; Li, F.-M. Lathyrus sativus (grass pea) and its neurotoxin ODAP. Phytochemistry 2006, 67, 107–121. [Google Scholar] [CrossRef]
- Feinberg, A.; Stenke, A.; Peter, T.; Hinckley, E.-L.S.; Driscoll, C.T.; Winkel, L.H.E. Reductions in the deposition of sulfur and selenium to agricultural soils pose risk of future nutrient deficiencies. Commun. Earth Environ. 2021, 2, 1–8. [Google Scholar] [CrossRef]
- Krogsgaard-Larsen, P.; Hansen, J.J. Naturally-occurring excitatory amino acids as neurotoxins and leads in drug design. Toxicol. Lett. 1992, 64-65, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Grosskreutz J, Van Den Bosch L, Keller B. (2010). Calcium dysregulation in amyotrophic lateral sclerosis Cell Calcium, 47 (2010), pp. 165-174.
- Kusama-Eguchi, K.; Miyano, T.; Yamamoto, M.; Suda, A.; Ito, Y.; Ishige, K.; Ishii, M.; Ogawa, Y.; Watanabe, K.; Ikegami, F.; et al. New insights into the mechanism of neurolathyrism: L-β-ODAP triggers [Ca2+]i accumulation and cell death in primary motor neurons through transient receptor potential channels and metabotropic glutamate receptors. Food Chem. Toxicol. 2014, 67, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Khandare, A.L.; B, K.; Validandi, V.; Ssyh, Q.; Harishankar, N.; Singh, S.S.; Kodali, V. Neurolathyrism in goat (Capra hircus) kid: Model development. Res. Veter- Sci. 2020, 132, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Doble A. (1999).The role of excitotoxicity in neurodegenerative disease: implications for therapy. Pharmacol. Ther., 81 (3) (1999), pp. 163-221.
- Hertz L. (2006). Glutamate, a neurotransmitter–and so much more A synopsis of Wierzba III. Neurochem. Int., 48 (6–7) (2006), pp. 416-425.
- Saeed, U.; Durgadoss, L.; Valli, R.K.; Joshi, D.C.; Joshi, P.G.; Ravindranath, V. Knockdown of Cytosolic Glutaredoxin 1 Leads to Loss of Mitochondrial Membrane Potential: Implication in Neurodegenerative Diseases. PLOS ONE 2008, 3, e2459–e2459. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, V. (2002). Neurolathyrism: mitochondrial dysfunction in excitotoxicity mediated by L-β-oxalyl aminoalanine. Neurochemistry international, 40(6), 505-509.
- López-Martín, M.C.; Becana, M.; Romero, L.C.; Gotor, C. Knocking Out Cytosolic Cysteine Synthesis Compromises the Antioxidant Capacity of the Cytosol to Maintain Discrete Concentrations of Hydrogen Peroxide in Arabidopsis. Plant Physiol. 2008, 147, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Hailu D, Abera S, Abera T. (2015), Effects of Processing on Nutritional Composition and Anti-Nutritional Factors of Grass pea (Lathyrus Sativus L) journal of Food Science and Quality Management www.iiste.org ISSN 2224-6088 (Paper) ISSN 2225-0557, Vol.36.
- Haileyesus Getahun, Fernand Lambein, Michel Vanhoorne and Patrick Van der Stuyft, (2005). Neurolathyrism risk depends on type of grass pea preparation and on mixing with cereals and antioxidants volume 10no 2 pp 169–178.
- Tadele, D. , Alemu, Y., Nigusie, D. and Peters, K. J. (2003). Evaluation of Processing Methods on the Feeding Value of Grass Pea to Broilers. Inter. J. Poult. Sci., 2(2): 120- 127.
- Akalu, G. , Johansson, G., & Nair, B. M. (1998). Effect of processing on the content of β-N-oxalyl-α, β-diaminopropionic acid (gb-ODAP) in grass pea (Lathyrus sativus) seeds and flour as determined by flow injection analysis. Food Chemistry, 62(2), 233-237.
- Barik, D.P.; Mohapatra, U.; Chand, P.K. Transgenic grasspea (Lathyrus sativus L.): Factors influencing Agrobacterium-mediated transformation and regeneration. Plant Cell Rep. 2005, 24, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Barna, K.S.; Mehta, S.L. Genetic Transformation and Somatic Embryogenesis in Lathyrus sativus. J. Plant Biochem. Biotechnol. 1995, 4, 67–71. [Google Scholar] [CrossRef]
- Girma, D. (2010). Ethiopian grass pea (Lathyrus sativus L.) started the genomics era: transient genetic transformation of grass pea. LAP Lambert Academic Publishing, Köln.
- Krishnan, H.B. Engineering Soybean for Enhanced Sulfur Amino Acid Content. Crop. Sci. 2005, 45, 454–461. [Google Scholar] [CrossRef]
- Kortt, A.A.; Caldwell, J.B.; Lilley, G.G.; Higgins, T.J.V. Amino acid and cDNA sequences of a methionine-rich 2S protein from sunflower seed (Helianthus annuus L.). Eur. J. Biochem. 1991, 195, 329–334. [Google Scholar] [CrossRef]
- Kirihara, J.A.; Petri, J.B.; Messing, J. Isolation and sequence of a gene encoding a methionine-rich 10-kDa zein protein from maize. Gene 1988, 71, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Chui, C.F.; Falco, S.C. A New Methionine-Rich Seed Storage Protein from Maize. Plant Physiol. 1995, 107, 291–291. [Google Scholar] [CrossRef] [PubMed]
- Planta, J.; Xiang, X.; Leustek, T.; Messing, J. Engineering sulfur storage in maize seed proteins without apparent yield loss. Proc. Natl. Acad. Sci. 2017, 114, 11386–11391. [Google Scholar] [CrossRef] [PubMed]
- Dinkins, R.D.; Reddy, M.S.S.; Meurer, C.A.; Yan, B.; Trick, H.; Thibaud-Nissen, F.; Finer, J.J.; Parrott, W.A.; Collins, G.B. Increased sulfur amino acids in soybean plants overexpressing the maize 15 kDa zein protein. Vitr. Cell. Dev. Biol. - Plant 2001, 37, 742–747. [Google Scholar] [CrossRef]
- Kim, W.S.; Krishnan, H.B. Allelic variation and differential expression of methionine-rich δ-zeins in maize inbred lines B73 and W23a1. Planta 2003, 217, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Liu, X.; Chen, L.; Cai, Y.; Yao, W.; Yuan, S.; Wu, C.; Han, T.; Sun, S.; Hou, W. Elevated methionine content in soybean seed by overexpressing maize β-zein protein. Oil Crop. Sci. 2020, 5, 11–16. [Google Scholar] [CrossRef]
- Kim, W.-S.; Sun-Hyung, J.; Oehrle, N.W.; Jez, J.M.; Krishnan, H.B. Overexpression of ATP sulfurylase improves the sulfur amino acid content, enhances the accumulation of Bowman–Birk protease inhibitor and suppresses the accumulation of the β-subunit of β-conglycinin in soybean seeds. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.-S.; Chronis, D.; Juergens, M.; Schroeder, A.C.; Hyun, S.W.; Jez, J.M.; Krishnan, H.B. Transgenic soybean plants overexpressing O-acetylserine sulfhydrylase accumulate enhanced levels of cysteine and Bowman–Birk protease inhibitor in seeds. Planta 2011, 235, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Avraham, T. , Badani, H., Galili, S., & Amir, R. (2005). Enhanced levels of methionine and cysteine in transgenic alfalfa (Medicago sativa L.) plants over-expressing the Arabidopsis cystathionine γ-synthase gene. Plant Biotechnology Journal, 3(1), 71-79.
- Endo, M.; Mikami, M.; Toki, S. Biallelic Gene Targeting in Rice. Plant Physiol. 2015, 170, 667–677. [Google Scholar] [CrossRef]
- Hilscher, J.; Bürstmayr, H.; Stoger, E. Targeted modification of plant genomes for precision crop breeding. Biotechnol. J. 2016, 12. [Google Scholar] [CrossRef]
- Friedrichs, S.; Takasu, Y.; Kearns, P.; Dagallier, B.; Oshima, R.; Schofield, J.; Moreddu, C. An overview of regulatory approaches to genome editing in agriculture. Biotechnol. Res. Innov. 2019, 3, 208–220. [Google Scholar] [CrossRef]
- Zannoni, L. Evolving Regulatory Landscape for Genome-Edited Plants. CRISPR J. 2019, 2, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Hussain, A.; Manghwar, H.; Xie, K.; Xie, S.; Zhao, S.; Larkin, R.M.; Qing, P.; Jin, S.; Ding, F. Genome editing with the CRISPR-Cas system: an art, ethics and global regulatory perspective. Plant Biotechnol. J. 2020, 18, 1651–1669. [Google Scholar] [CrossRef] [PubMed]
- Emmrich, P. M. , Sarkar, A., Njaci, I., Kaithakottil, G. G., Ellis, N., Moore, C.,... & Wang, T. L. (2020). A draft genome of grass pea (Lathyrus sativus), a resilient diploid legume. BioRxiv, 2020-04.
- Friedrichs S, Takasu Y, Kearns P et al (2019) Meeting report of the OECD conference on “genome editing: applications in agriculture—implications for health, environment and regulation.” Transgenic Res 28:419–463.
- Chilcoat D, Liu ZB, Sander J (2017) Use of CRISPR/Cas9 for crop improvement in maize and soybean. Prog Mol Biol Transl Sci 149:27–46.
- Fiaz, S.; Ahmad, S.; Noor, M.A.; Wang, X.; Younas, A.; Riaz, A.; Riaz, A.; Ali, F. Applications of the CRISPR/Cas9 System for Rice Grain Quality Improvement: Perspectives and Opportunities. Int. J. Mol. Sci. 2019, 20, 888. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wu, Y.; Zhang, Y.; Yang, J.; Fan, W.; Zhang, H.; Zhao, S.; Yuan, L.; Zhang, P. CRISPR/Cas9-Based Mutagenesis of Starch Biosynthetic Genes in Sweet Potato (Ipomoea Batatas) for the Improvement of Starch Quality. Int. J. Mol. Sci. 2019, 20, 4702. [Google Scholar] [CrossRef] [PubMed]
- Tuncel, A.; Corbin, K.R.; Ahn-Jarvis, J.; Harris, S.; Hawkins, E.; Smedley, M.A.; Harwood, W.; Warren, F.J.; Patron, N.J.; Smith, A.M. Cas9-mediated mutagenesis of potato starch-branching enzymes generates a range of tuber starch phenotypes. Plant Biotechnol. J. 2019, 17, 2259–2271. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Jiao, G.; Sun, Y.; Chen, J.; Zhong, Y.; Yan, L.; Jiang, D.; Ma, Y.; Xia, L. Modification of starch composition, structure and properties through editing of TaSBEIIa in both winter and spring wheat varieties by CRISPR/Cas9. Plant Biotechnol. J. 2020, 19, 937–951. [Google Scholar] [CrossRef] [PubMed]
- Nieves-Cordones, M.; Mohamed, S.; Tanoi, K.; Kobayashi, N.I.; Takagi, K.; Vernet, A.; Guiderdoni, E.; Périn, C.; Sentenac, H.; Véry, A. Production of low-Cs+ rice plants by inactivation of the K+ transporter OsHAK1 with the CRISPR-Cas system. Plant J. 2017, 92, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Gomez, M.A.; Berkoff, K.C.; Gill, B.K.; Iavarone, A.T.; Lieberman, S.E.; Ma, J.M.; Schultink, A.; Karavolias, N.G.; Wyman, S.K.; Chauhan, R.D.; et al. CRISPR-Cas9-mediated knockout of CYP79D1 and CYP79D2 in cassava attenuates toxic cyanogen production. Front. Plant Sci. 2023, 13, 1079254. [Google Scholar] [CrossRef]
- Song, J.H.; Shin, G.; Kim, H.J.; Lee, S.B.; Moon, J.Y.; Jeong, J.C.; Choi, H.-K.; Kim, I.A.; Song, H.J.; Kim, C.Y.; et al. Mutation of GmIPK1 Gene Using CRISPR/Cas9 Reduced Phytic Acid Content in Soybean Seeds. Int. J. Mol. Sci. 2022, 23, 10583. [Google Scholar] [CrossRef]
- Sashidhar, N.; Harloff, H.J.; Potgieter, L.; Jung, C. Gene editing of three BnITPK genes in tetraploid oilseed rape leads to significant reduction of phytic acid in seeds. Plant Biotechnol. J. 2020, 18, 2241–2250. [Google Scholar] [CrossRef]
- Raffan, S.; Sparks, C.; Huttly, A.; Hyde, L.; Martignago, D.; Mead, A.; Hanley, S.J.; Wilkinson, P.A.; Barker, G.; Edwards, K.J.; et al. Wheat with greatly reduced accumulation of free asparagine in the grain, produced by CRISPR/Cas9 editing of asparagine synthetase gene TaASN2. Plant Biotechnol. J. 2021, 19, 1602–1613. [Google Scholar] [CrossRef] [PubMed]
- Kenar, J.A. Reaction chemistry of gossypol and its derivatives. J. Am. Oil Chem. Soc. 2006, 83, 269–302. [Google Scholar] [CrossRef]
- Lin, J.-L.; Fang, X.; Li, J.-X.; Chen, Z.-W.; Wu, W.-K.; Guo, X.-X.; Liu, N.-J.; Huang, J.-F.; Chen, F.-Y.; Wang, L.-J.; et al. Dirigent gene editing of gossypol enantiomers for toxicity-depleted cotton seeds. Nat. Plants 2023, 9, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Schachtsiek, J.; Stehle, F. Dataset on nicotine-free, nontransgenic tobacco (Nicotiana tabacum l.) edited by CRISPR-Cas9. Data Brief 2019, 26, 104395. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, M.; Schemmerling, B.; Ober, D. CRISPR/Cas9-Mediated Genome Editing in Comfrey (Symphytum officinale) Hairy Roots Results in the Complete Eradication of Pyrrolizidine Alkaloids. Molecules 2021, 26, 1498. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Sun, S.; Whelan, J.; Shou, H. CRISPR/Cas9-Mediated Knockout of GmFATB1 Significantly Reduced the Amount of Saturated Fatty Acids in Soybean Seeds. Int. J. Mol. Sci. 2021, 22, 3877. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Shea, Z.; Rosso, L.; Shang, C.; Li, J.; Bewick, P.; Li, Q.; Zhao, B.; Zhang, B. Development of new mutant alleles and markers for KTI1 and KTI3 via CRISPR/Cas9-mediated mutagenesis to reduce trypsin inhibitor content and activity in soybean seeds. Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Rajarammohan, S.; Kaur, L.; Verma, A.; Singh, D.; Mantri, S.; Roy, J.K.; Sharma, T.R.; Pareek, A.; Kandoth, P.K. Genome sequencing and assembly of Lathyrus sativus - a nutrient-rich hardy legume crop. Sci. Data 2023, 10, 1–9. [Google Scholar] [CrossRef]
- Riepe, M.; Spencer, P.S.; Lambein, F.; Ludolph, A.C.; Allen, C.N. In vitro toxicological investigations of isoxazolinone amino acids of Lathyrus sativus. Nat. Toxins 1995, 3, 58–64. [Google Scholar] [CrossRef]
- Saha, T. , Shee, R., Sahid, S., Shee, D., Roy, C., Sharma, R.,... & Datta, R. (2023). Designer grass pea for transgene-free minimal neurotoxin-containing seeds with CRISPR-Cas9. bioRxiv, 2023-03.
- Papadopoulos, J.S.; Agarwala, R. COBALT: constraint-based alignment tool for multiple protein sequences. Bioinformatics 2007, 23, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Global specimen registry (yellow circles) of grass pea (Lathyrus sativus). The map was taken from global mapper on 28 March, 2024 (https://www.discoverlife.org/mp/20m?act=make_map&kind=Lathyrus+sativus).
Figure 1.
Global specimen registry (yellow circles) of grass pea (Lathyrus sativus). The map was taken from global mapper on 28 March, 2024 (https://www.discoverlife.org/mp/20m?act=make_map&kind=Lathyrus+sativus).
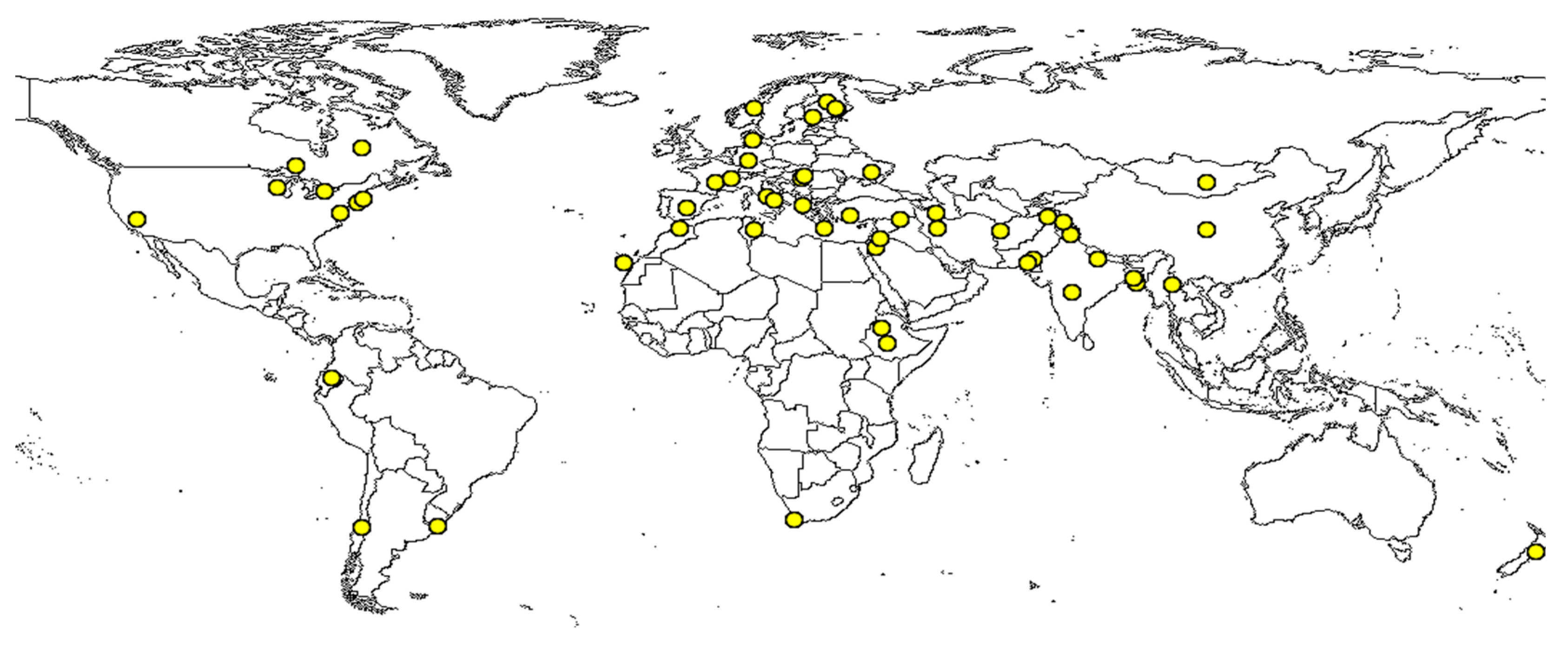
Figure 2.
(A) Illustration of the proposed biosynthetic pathway of β-ODAP biosynthesis (ChemDraw) adopted from [80]. The products in brackets (Isoxazolin-5-one) are hypothetical and has not been detected in grass pea. BIA is β-(isoxazolin-5-on-2-yl) alanine) a precursor of L-DAPA. L-DAPA is L-2,3-Diaminopropionic Acid a final precursor of β-ODAP. The light green line indicated the formation of BIA from Isoxazolin-5-one and cysteine by the CAS enzyme at high sulfur. The blue line indicates the formation of BIA from Isoxazolin-5-one and O-Acetyl serine at low sulfur by CS enzyme. According to our conserved domain analysis, CAS is bifunctional enzyme that may also act as CS. (B) Structural illustration of L-Glutamic acid analog that competes for AMPA receptor. The structures were drawn using ChemDraw.
Figure 2.
(A) Illustration of the proposed biosynthetic pathway of β-ODAP biosynthesis (ChemDraw) adopted from [80]. The products in brackets (Isoxazolin-5-one) are hypothetical and has not been detected in grass pea. BIA is β-(isoxazolin-5-on-2-yl) alanine) a precursor of L-DAPA. L-DAPA is L-2,3-Diaminopropionic Acid a final precursor of β-ODAP. The light green line indicated the formation of BIA from Isoxazolin-5-one and cysteine by the CAS enzyme at high sulfur. The blue line indicates the formation of BIA from Isoxazolin-5-one and O-Acetyl serine at low sulfur by CS enzyme. According to our conserved domain analysis, CAS is bifunctional enzyme that may also act as CS. (B) Structural illustration of L-Glutamic acid analog that competes for AMPA receptor. The structures were drawn using ChemDraw.
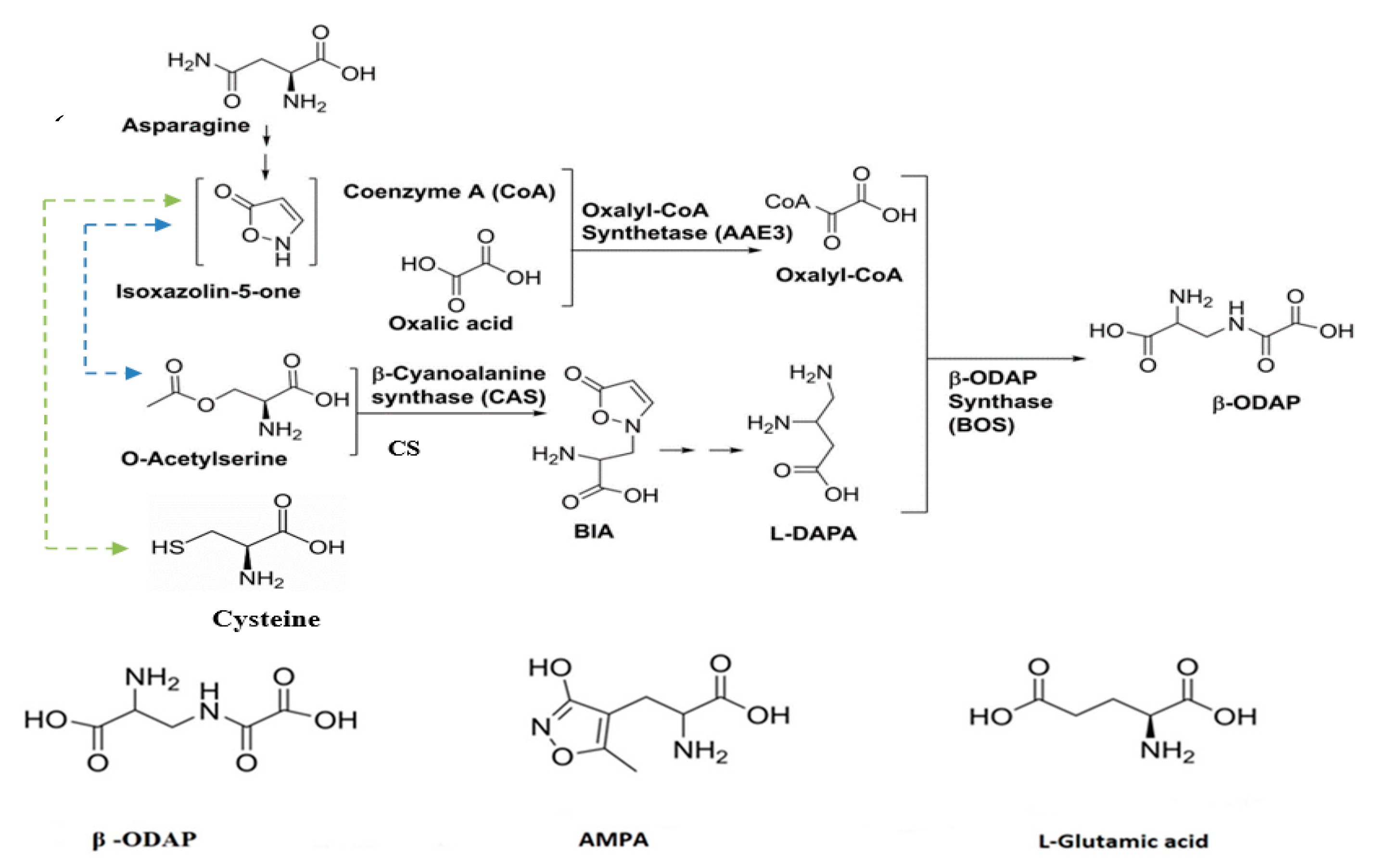
Figure 3.
The mechanism of degeneration induced by β-ODAP proposed by[1]with modification of figure design. Figure (A) illustrates the removal of excitatory amino acid glutamate by glutamate transporter and its storage in a vesicle until needed. Figure (B) indicates the binding of β -ODAP to the AMPA receptor and allows the influx of Ca2+ into the cell. The binding of β -ODAP and over-activating AMPA receptors triggers the signaling cascade of reactions with intracellular Ca2+ low. Meanwhile, the increased expression level of β1 integrin on the cell surface substantially induces the phosphorylation level of FAK and overexpression of paxillin. This causes massive aggregation of FA units on cell actin filaments, interferes with the assembly of cell microfilaments, and ultimately results in structural damage to the cells [1]. .
Figure 3.
The mechanism of degeneration induced by β-ODAP proposed by[1]with modification of figure design. Figure (A) illustrates the removal of excitatory amino acid glutamate by glutamate transporter and its storage in a vesicle until needed. Figure (B) indicates the binding of β -ODAP to the AMPA receptor and allows the influx of Ca2+ into the cell. The binding of β -ODAP and over-activating AMPA receptors triggers the signaling cascade of reactions with intracellular Ca2+ low. Meanwhile, the increased expression level of β1 integrin on the cell surface substantially induces the phosphorylation level of FAK and overexpression of paxillin. This causes massive aggregation of FA units on cell actin filaments, interferes with the assembly of cell microfilaments, and ultimately results in structural damage to the cells [1]. .
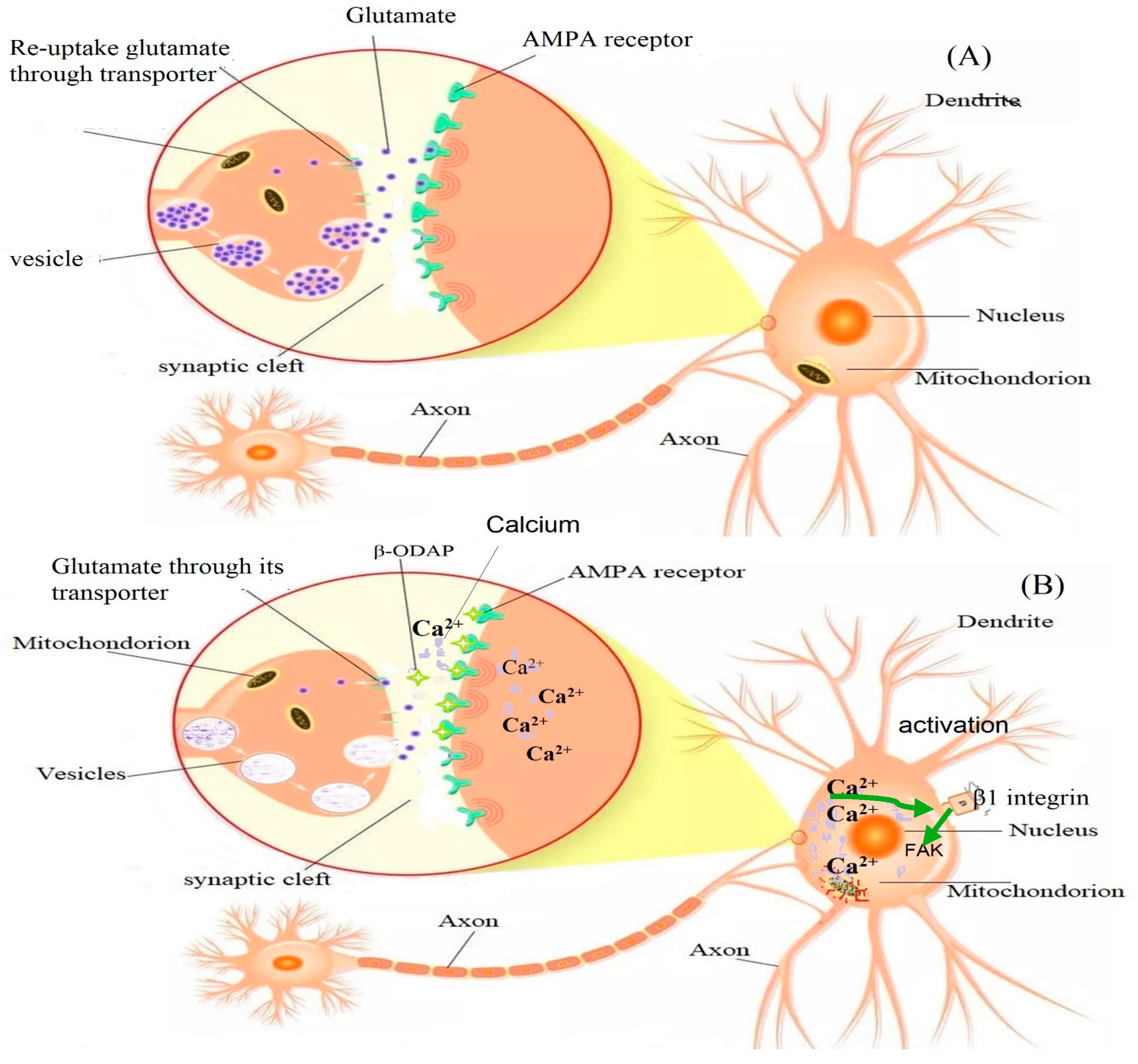
Figure 4.
Illustration of β-ODAP toxicity and irreversible paralysis caused by extensive consumption of grass pea as reported by a number of studies [79,89,90]. The model of lathyrism in humans was taken from mind and body: holistic health center (https://www.mindbodyheal.co.uk/lathyrism/).
Figure 4.
Illustration of β-ODAP toxicity and irreversible paralysis caused by extensive consumption of grass pea as reported by a number of studies [79,89,90]. The model of lathyrism in humans was taken from mind and body: holistic health center (https://www.mindbodyheal.co.uk/lathyrism/).
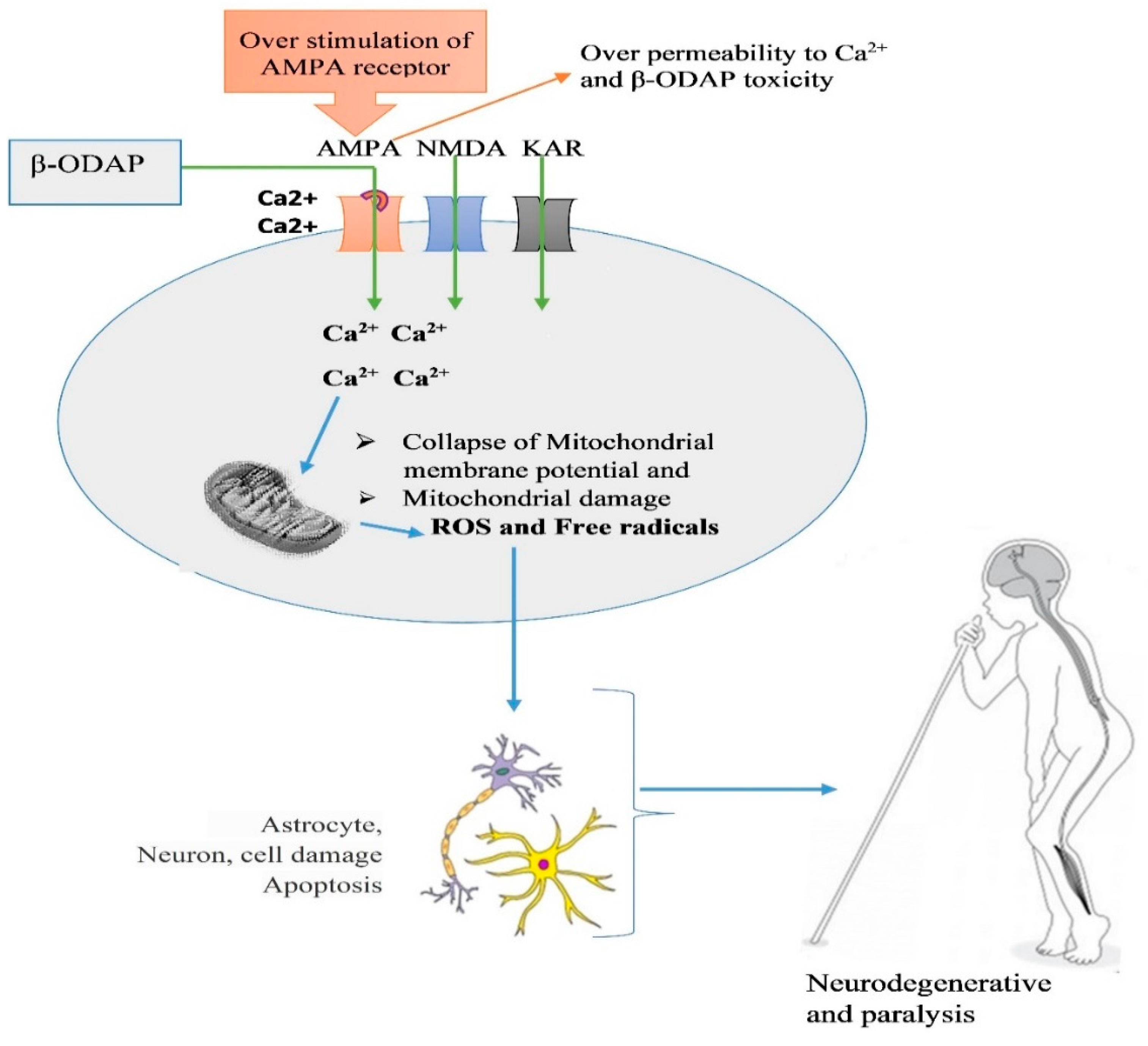
Figure 5.
Potential target for CRISPR/cas9 for β-ODAP reduction in grass pea. Abbreviations L-DAPA represent L-2,3-Diaminopropionic Acid; BIA is β-(isoxazolin-5-on-2-yl) alanine); BOS is β-ODAP synthase and CAS is β-cyanoalanine synthase. CAS and BOS are potential target genes that can be either knockout simultaneously or separately.
Figure 5.
Potential target for CRISPR/cas9 for β-ODAP reduction in grass pea. Abbreviations L-DAPA represent L-2,3-Diaminopropionic Acid; BIA is β-(isoxazolin-5-on-2-yl) alanine); BOS is β-ODAP synthase and CAS is β-cyanoalanine synthase. CAS and BOS are potential target genes that can be either knockout simultaneously or separately.
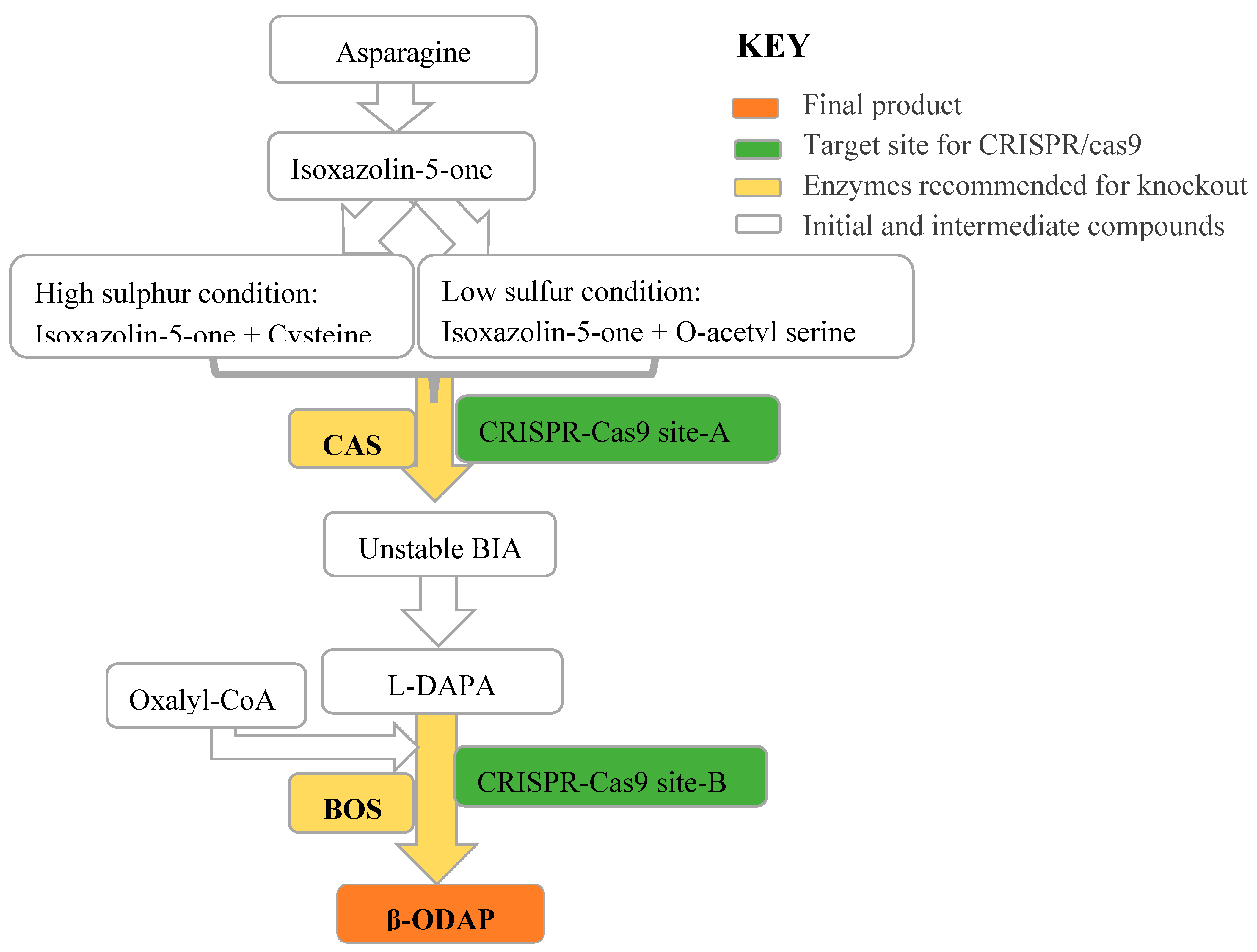
Figure 6.
Similarity of grass pea (lathyrus sativus) β-ODAP synthase enzymes (A) and β-cyanoalanine synthase (B) with other plants. Solid red color indicates conserved amino acids in all the eight species, the grey and white columns represent indels, and the blue line represents the substitution of aligned sequences. Amino acid length and names of all the species were listed on the right side of the figure.
Figure 6.
Similarity of grass pea (lathyrus sativus) β-ODAP synthase enzymes (A) and β-cyanoalanine synthase (B) with other plants. Solid red color indicates conserved amino acids in all the eight species, the grey and white columns represent indels, and the blue line represents the substitution of aligned sequences. Amino acid length and names of all the species were listed on the right side of the figure.
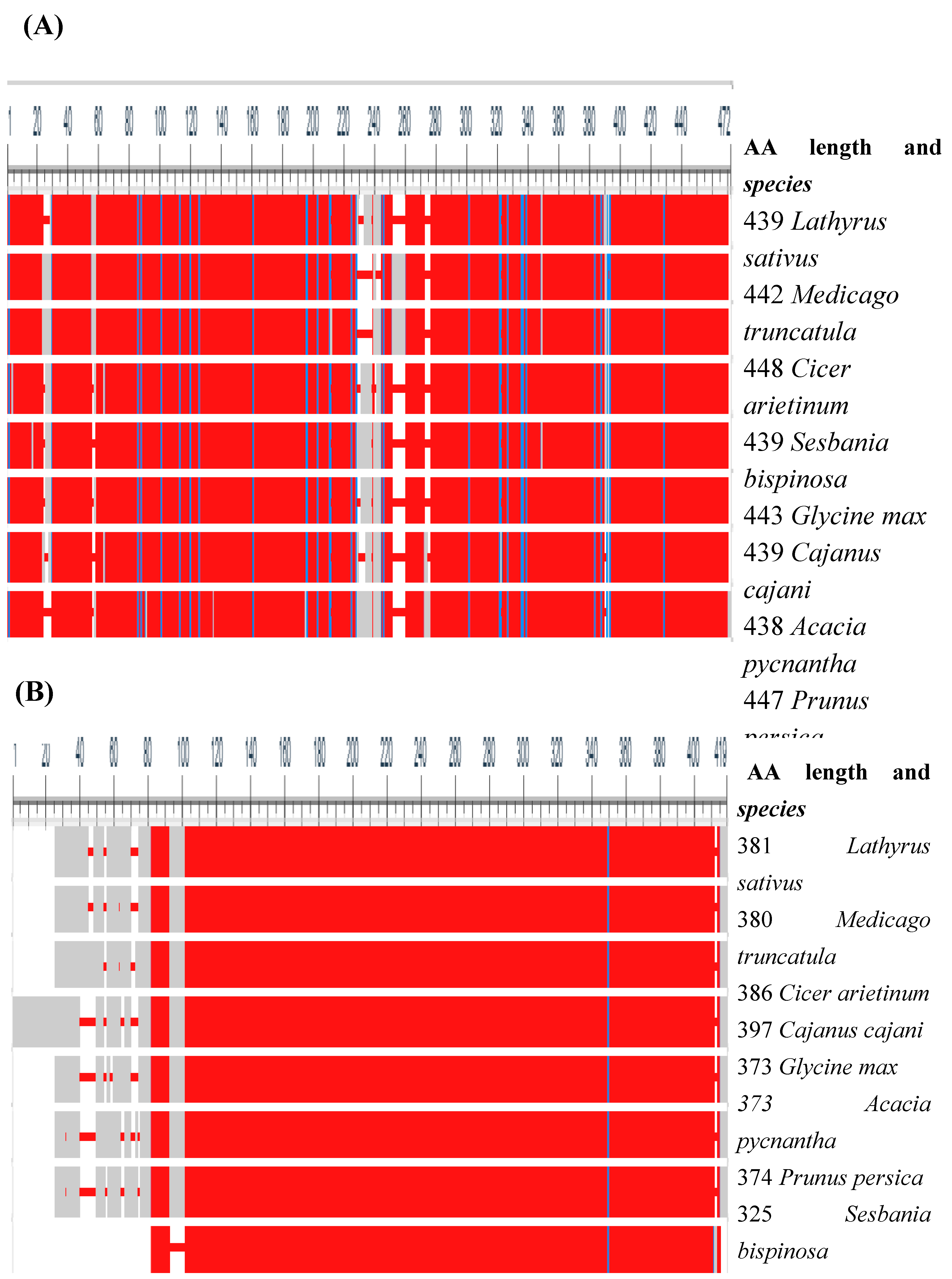
Figure 7.
Genetic relatedness of β-ODAP (A, scaled tree) and CAS enzyme (B, unscaled tree) as compressed to legume model and other plants. With both enzymes, L. satvus and M. truncatula have shown close relationship though the there are some sort of amino acids differences in conserved areas.
Figure 7.
Genetic relatedness of β-ODAP (A, scaled tree) and CAS enzyme (B, unscaled tree) as compressed to legume model and other plants. With both enzymes, L. satvus and M. truncatula have shown close relationship though the there are some sort of amino acids differences in conserved areas.
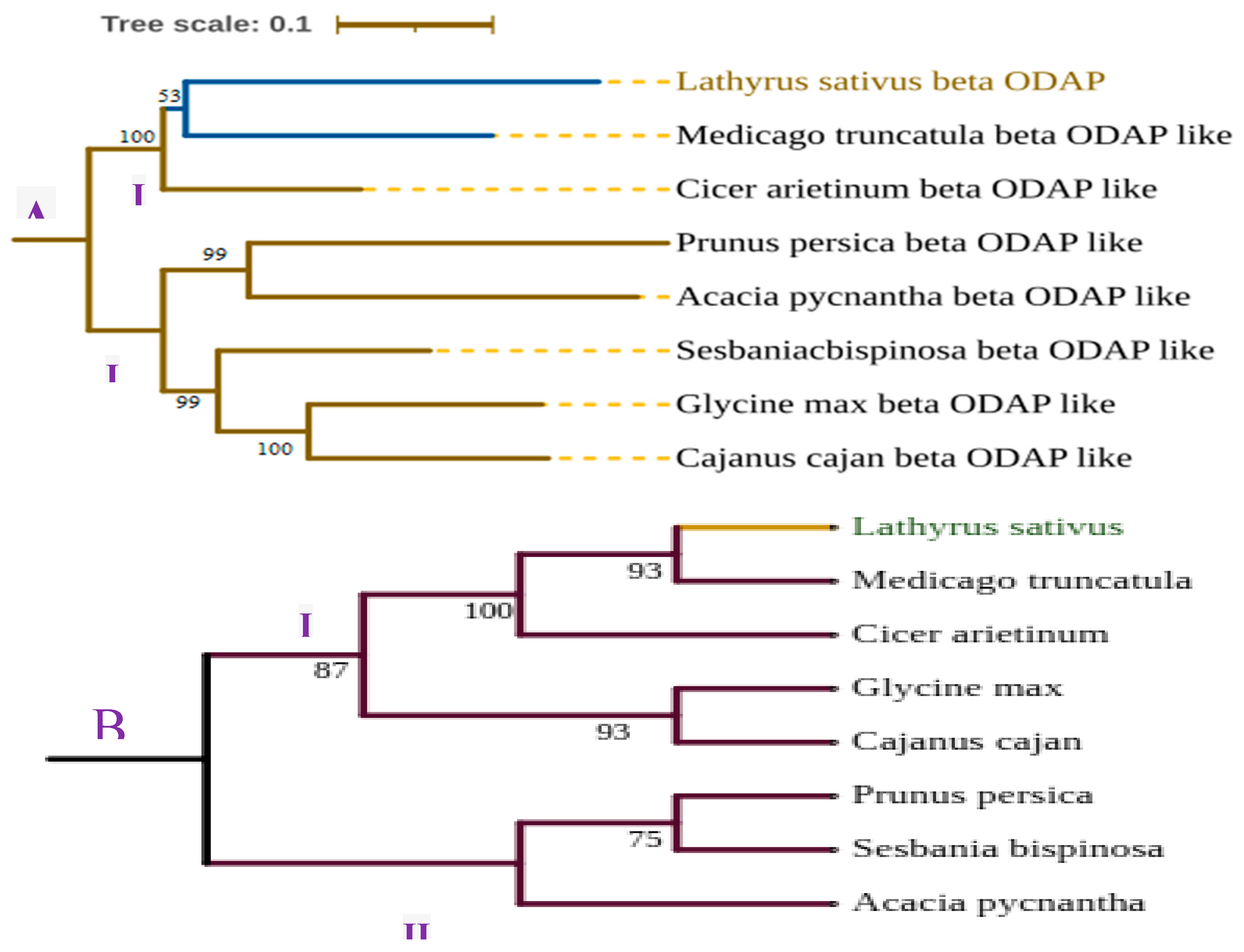
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



