Submitted:
10 July 2024
Posted:
12 July 2024
You are already at the latest version
Abstract
Abstract: Little is known about landbird group dynamics in response to predation. Here, we describe the interactions between the Helmeted Guineafowl (Numida meleagris) and its predators regarding the survival advantages grouping behaviour may provide. Livestream webcam observations were conducted in Madikwe Game Reserve (South Africa) from August 2020 to December 2021. Emphasis was placed on predator–prey interaction and its effect on group size and structure in a spatial framework. Although Black-Backed Jackals (Lupulella mesomelas) are not adept at hunting birds, they might negatively impact guineafowl by curtailing their feeding time during winter with constant harassment. We found that these canids often approach larger and denser groups in the morning and late evenings. Surprisingly, guineafowl most often responded by standing nearby. On the contrary, a higher predator-induced response was associated with the presence of predatory birds, arguably as part of an evolutionary stable prey-predator system. This line of thought is also supported by the lack of a precise defensive mechanism elicited by jackal attacks, unlike those of aerial predators. Nevertheless, we contend that grouping in Helmeted Guineafowl is a defensive strategy to respond to predator attacks while individuals engage in daily activities.
Keywords:
antipredator model
; collective behaviour
; terrestrial and aerial predation
; Lupulella mesomelas
; predator-prey ecology
1. Introduction
Collective behaviour in birds has traditionally been ascribed to feeding strategies, reproduction, parasite control, thermal benefits and energy saving [1,2,3]. Nevertheless, little effort has been made to understand how anti-predator strategies shape and drive collective behaviour [4]. In previous studies, predation has been primarily considered in ecological terms, with predators limiting the numbers of their prey in a regulated (density-dependent) or unregulated (density-independent) manner [5,6]. Only recently has growing attention been paid to the costs and benefits underlying collective behaviour as a defensive strategy [7,8].
Increased protection from predators has been invoked as the rationale behind group formation in several avian predator-prey systems. For example, the Common Redshank (Tringa totanus), is known to benefit from the lower risk of Sparrowhawk (Accipiter nisus) and Peregrine Falcon (Falco peregrinus) attacks by living in larger groups due to the ‘dilution’ effect, increased vigilance (which was a direct consequence of group size) and possibly the ‘confusion’ effect [9]. Likewise, significantly higher frequencies of large and dense flocks were found in European starling (Sturnus vulgaris) roosts under high predation pressure [10]. However, if predator presence has been described as a crucial factor behind grouping behaviour as anti-predator strategy in several animal species [11], other hypotheses suggest that grouping serves important functions, such as defence against parasites. For example, a specific case of what is generally referred to as the selfish herd effect [12] postulates that individuals in the edges of an aggregation will be less frequently attacked by predators and some parasites. Moreover, grouping behaviour is often associated with thermoregulation and, more specifically, with heat loss reduction, as seen in birds huddling during cold spells [13]. Additionally, foraging in a group could reduce the energetic costs of movement [14] and increase individual feeding rates thanks to the information shared among group members [15], as well as the reduction of time allocated to vigilance [16]. Nevertheless, there is a trade-off between resource intake rate and both vigilance rate and group size, suggesting the occurrence of an optimum number of foraging individuals that maximises individual fitness [17]. Indeed, if constantly harassed by predators while having to compete for food, prey may substantially decrease their feeding rates, with detrimental effects on their fitness [15]. While antipredator strategies seem to be influenced by specific predator traits, it was also argued that the selection and intensity of antipredator behaviours are also influenced by hunting style, such as, for instance, that of aerial or terrestrial predators stalking groups of birds on the ground [18]. Moreover, habitat structure can affect anti-predator behaviour [19]. For example, landscape fragmentation impacts the structure of mixed-species flocks of forest passerines, as well as bird anti-predator behaviour and energy management strategies [20]. Finally, the activities of prey being attacked by a predator may also influence the anti-predator response, whether collective or not [21]. More specifically, vegetation structure and landscape configuration are crucial factors in determining how prey responds to predators. For example, in territories with lush vegetation, small prey like the Rufous-and-White Wren (Thryophilus rufalbus) and the Banded Wren (T. pleurostictus) rely on acoustic, rather than visual, signals [22]. Yet another experiment on the foraging of granivorous birds evidenced that more food leftovers occurred in the open than under cover, meaning that feeding therein was preferred presumably because this provided an effective anti-predator shield [23].
However, grouping behaviour responding to predation is still poorly known in landfowl [24,25]. This also applies to the Helmeted Guineafowl (Numida meleagris), a widespread Afrotropical galliform that responds overtly to predator attacks by adopting a suite of defensive strategies including loud alarm calls, fleeing, mobbing and associating with other species behaviour in response to predation [26,27,28,29]. This species is territorial and highly sociable, living in close-knit groups of 8 to 15 birds and seasonally joining neighbouring groups that can sum up to 50 birds [30]. These units are stable social systems led by a dominant male initiating the foraging direction and warding off male intruders into their territory, wherein breeding cooperation maximises offspring survival [31,32]. Although social interactions between individuals within these aggregates have been previously investigated [30,31,33], little attention has been paid to collective response to predation and group formation. Filling this knowledge gap is a management concern because the Helmeted Guineafowl is a valuable game bird in South Africa and neighbouring countries [34]. Noteworthy, this species has recently expanded to anthropised areas – such as the South African inland plateau known as Highveld [35] – which might lack the resources they need to escape predators [25,34]. Moreover, by being large-sized, noisy, and mostly foraging in the open, the Helmeted Guineafowl represents a good model for examining the driving forces underlying grouping and predation [3,36]. The challenge in addressing this topic in these birds is to determine how these behaviourally respond to multiple predators [26], with some mainly inducing a non-lethal effect and others a lethal effect.
We hypothesise that grouping behaviour is a defensive mechanism in the Helmeted Guineafowl. The hypothesis will be supported if the following predictions are met: i) guineafowl change their group structure to optimise feeding, drinking, sunning, resting and roosting while busy with constant predator surveillance and avoidance behaviour; ii) group protection allows guineafowl to exploit scarce food patches optimally even under predator pressure; iii) groups serve as an information centre to maximise predator avoidance and individual fitness. However, group formation bears additional costs as groups can attract predators’ attention and thus iv) there would be an optimum group size beyond which predatory attack probability will increase asymptotically with group size [7].
2. Material and Methods
2.1. Study Area
All the data was obtained from videos taken with a stationary Hikvision livestream webcam (model DS-2DF8236IX-AELW) installed at Tau Waterhole, located in the 75,000 ha Madikwe Game Reserve (hereafter referred to as Tau: 24 42′33.24 S 26 12′19.17 E), from August 2020 to August 2021. The camera was installed by AfriCam and is operated remotely by volunteers. At the same time, audible sounds were recorded through Sennheiser HD 435 headphones plugged into an HP (Pavilion) computer fitted with a Bang and Olufsen (B&O) sound system.
The study site, covering 6.6 ha, consists of a waterhole, an open plain, roosting tree, a scrub area, and dense bush of Resin Tree (Grewia spp.), Acacia Thorn Tree (Acacia karoo), Umbrella Thorn Tree (A. tortilis), Black Thorn Tree (A. mellifera), Camel Thorn Tree (A. erioloba) and Sickle Bush (Dichrostachys cinerea) (Figure 1). Local vegetation is classified as Dwaalboom Thornveld [37]. The plain was visited daily by herds of 30 to 100 African Bush Elephants (Loxodonta africana), 20 to 100 African Buffalos (Syncerus caffer), 10 to 15 Greater Kudus (Tragelaphus strepsiceros), 50 to 100 Impalas (Aepyceros melampus), 30 to 50 Blue Wildebeests (Connochaetes taurinus), 10 to 30 Burchell’s Zebras (Equus burchelli) and Giraffes (Giraffa giraffa). Chacma Baboons (Papio comatus) and carnivores such as African Lions (Panthera leo), Spotted Hyenas (Crocuta crocuta) and Black-backed Jackals (Lupulella mesomelas) also roamed the plain.
2.2. Livestream Camera Observations
The fixed livestream webcam at Tau had been previously used to observe guineafowl drinking at the waterhole between 7:00 and 10:00 and between 16:00 and 18:00, where they primarily formed guilds with Impala and Blue Wildebeest [38]. The webcam scanned the entire plain and neighbouring scrub from left to right and back again with 5-minute sweeps (Figure 1). If a sweep was unsatisfactory (for instance, when the camera was not rolling correctly or when the inclement weather caused temporary interference leading to a blurry optic), we logged in (that were conducted nine times daily, from 6:00 to 19:00) again to find a suitable sweep from which it was possible to collect data consistently.
From the overall number of guineafowl groups recorded from August 2020 to August 2021 (see Results), suitable (i.e., images not blurry with easily countable individuals) sub-samples were selected to conduct downstream statistical analyses. Different groups generally remained from 50 m to 200 m apart, with the camera usually spotting several during the five 5-minute sweeps. Group size and location were recorded daily. The birds were easily observed on the plain and in the scrub further away from the water during the dry winter (see Figure 1). Group sizes in the scrub could be determined accurately as the camera captured the entirety of the birds that periodically moved in and out of the scrub cover. Predator approaches by jackals and birds of prey and guineafowl response behaviours were recorded during the scans. More specifically, the camera was brought to a standstill by the remote controller to capture the scene when a predator-prey interaction was about to occur. This extra time was added to the standardised 5-minute calculation sweeps. Suitable photos were selected to determine the social distance (SD) in groups (Supplementary S1). The following data were recorded in a matrix for each SD calculated (see below): date, time of day, number of birds, guineafowl scale (i.e., the broadside bird length in mm), habitat and number of birds forming a straight line. In this case, a vernier measured group spread (i.e., the distance between two proximal guineafowl on either side of a straight line, as seen in mm on the computer screen). For drinking birds, the scale referred to the (frontal) breadth of the birds. In this case, SD was determined between birds standing beside one another.
To study predator surveillance behaviour, the heads-up postures adopted by members of guineafowl groups were photographed from April to August 2021 [39]. An independent Pentax K-5 camera with a Sigma EX 10- to 20-mm wide-angle lens was used to take and download photographs via the computer screen. For this purpose, a random group was sampled in the scrub, on the open plain, during a huddle, and when drinking water from the webcam. Every 2 seconds, a photograph was taken. Although screenshots would have also been possible, the camera automatically supplied a 2-second interval between the images for consistency. For each sampling effort, 30 photographs were taken in 60 seconds (with the visibility of a group during the sweep generally being of about 60 seconds). All the data was used to convert the heads-up postures to percentages.
2.3. Statistical Methods
Kruskal-Wallis tests were used to determine differences in: i) group size at different times of the day; ii) average social distance between birds within group; iii) group size of roosting birds across winter months; iv) time needed for a group to ascend a communal roosting tree (from the first to the last bird in the group) with time needed to descend from it in the morning. We focused on iii) and iv) since grouping behaviour at roost and roost departure time are known as anti-predator responses in other species [40,41]. We also performed a multinomial model with predator presence type (i.e., predatory bird, jackal, no predator) as response variable and group size and month as predictors.
Surveillance behaviour was measured in terms of head-up counts per group, which were converted to percentages (head-up counts/number of birds x 100) to compare their counts in groups engaging in different activities and across different habitats. Furthermore, to compare head-ups in different habitats (i.e., scrub vs open plain) or when engaging in different activities (i.e., during drinking or huddling) using chi-square tests (https://www.socscistatistics.com/tests/kruskal/default.aspx). Spearman rank correlation (Statistics Kingdom.com) was used to correlate jackals/hour vs mean sizes of groups stalked by jackals. The number of guineafowl groups in different time slots of the day was compared using chi-square tests. SD between individuals was calculated using Excel using the following algorithm:
{[((s/scale)*constant)-(f*constant)]/(f-1)} = SD,
where “s” is the spread of the birds standing in a straight line, “scale” is the size of one or more representative guineafowl in the line determined using measuring the broadside size or the frontal breadth of the bird(s) (with the average being calculated in the case of the measuring more than one bird); “constant” is 0.350 m, corresponding to the average broadside size of a live guineafowl, and the constant for drinking birds being 0.175 m (the frontal breadth of the birds); and“f-1” is the number of birds in a straight line minus 1, due to the focus being on the average distance between birds in the group (Supplementary S2).
3. Results
In total 411 groups (mean group size = 14.26; range 2 - 70; Std. D = 12.61) were observed and recorded over the entire study period. The number of groups selected for each theme that was analysed is given in Table 1. Livestream camera observation times are revealed in Table 2.
3.1. Spatiotemporal Group Size Dynamic
Average group sizes were minimal from November to February (mean = 2.5, range = 2-5, Std. D = 1, n = 13) during the breeding season, which occurs during summer when very few birds could be spotted. During winter, the group dynamics changed monthly and daily, being maximal in midwinter late afternoons and evenings (Figure 2; Supplementary S3). During the day, the webcam clearly showed how the birds descended from their roost as a relatively large collective group, breaking down into smaller ones, with the opposite behaviour occurring during the late afternoon when the birds prepared to roost again (Figure 2). The difference between the mean group sizes in the morning (9:00), compared to in the afternoons (16:00 to 18:00), was statistically significant (Table 3).
3.2. Group Sizes on the Plain and in the Scrub
One or two hours after the birds descended from their roost onto the plain, most of them were invariably dispersed in the scrub and the dense thickets further away from the plain (Figure 1). The mean group sizes of guineafowl recorded in the scrub (mean = 7.64, Std. D = 7.80, range = 1 - 55, n = 73) and on the plain (mean = 14.88, Std. D = 12.24, range = 2 - 40, n = 25) from July to August were significantly different (H = 7.049, n = 98, p = 0.025) (Supplementary S5).
3.3. Social Distance
The mean SD between group members was minimal when they stood shoulder to shoulder while drinking water (0.071 m). The SD was even smaller (0.034 m) when guineafowl huddled together after being attacked by a jackal and just before or after roosting. A larger mean SD (0.835 m) was calculated for birds foraging on the open plain than for birds feeding in the scrub (0.279 m). SD was significantly higher between birds feeding in the open relative to those engaging in all other activities elsewhere in the study area (Table 4; Supplementary S6). Likewise, the mean SD was significantly higher in the afternoons than in the morning and even in the late afternoon compared to early afternoon (p < 0.05: Table 5).
3.4. Roosting Behaviour
From March to August 2021, the groups on the plain were found to roost communally in a single clump of trees, with most birds roosting in three or four adjacent Sweet Thorn trees opposite the waterhole (40 m across, from tree to launching pad) (see Figure 1). The average roosting group size, that was relatively smaller in March, increased in June and July but then tapered off as smaller units peeled off to roost elsewhere during wintertime (in secondary roosts; see Figure 1) when the amount of residual tree foliage diminished. Accordingly, the mean group size in April (47.5) was significantly smaller than in June (109.2) and July (91.31), respectively, shrinking significantly again in August (53.37) (Table 6: Supplementary S6).
Birds ascended the trees one by one or in small groups of three to four, but when they descended, they did so in groups of 20 to 40 per batch. On average, the time between the first and the last bird to fly into the roosting tree from the launching pad (regular ascending patch) in the late afternoon/evening was 8.23 min. Such duration was significantly shorter in the mornings when the birds were found to leave the trees in 2.44 minutes (Table 7; H = 49.059, n = 71, p < 0.5). The time needed for the group to enter the trees was not correlated with the number of birds roosting together (R2 = 0.000095; F (1,38) = 0.006; p = 0.952), nor was the time needed for birds to descend the trees (R2 = 0.035; F (1,26); p = 0.341) (Table 7). Guineafowl were found to ascend into their roost 20 or more minutes after sunset. As this occurred later with the approaching summer, the birds were seen to follow suit with their timing (Figure 3). On 20 July 2021, an unidentified predatory bird attacked the roosting birds; in response, about 30 of them flew down to the launching pad again. After remaining on the ground for about 15 to 20 minutes, they flew back into the trees about 40 minutes after sunset.
3.5. Surveillance Behaviour
The number of suitable photographs selected for counting heads-up was 190, with 3816 birds appearing in the images (Table 8; Supplementary S7). The least head-ups were measured in groups in scrub (5.3%) or on the open plain (1.01%). Comparatively, more head-up counts were recorded in groups drinking water (40%) or huddling together (45%) before roosting in the evenings (chi-square = 185.655, df 1,3, p < 0.001). There was a non-significant correlation between head-ups and group sizes (rs = -0.011, p = 0.91, n = 101, Table 8). In a few instances (n = 5), the heads-up posture in a group was performed by the same member identified based on its unique helmet shape, but when an alarm call was emitted, several birds matched it with a heads-up posture.
3.6. Vocalisations
While foraging, guineafowl invariably emitted a soft contact whistle, sounding like “so we we we” (at about 40 dB), which served to keep group members together while keeping their heads down [42]. When an intruder was detected at a distance, one or two members in a group usually emitted a low-intensity alarm call sounding like a medium-loud “chit” (at between 50 and 60 dB) to signal a potential threat, with such an occurrence being recorded on 40 different occasions [43]. Finally, the call would transform into a loud “chit chit chiiir” (at between 70 and 85 dB) emitted by several birds while adopting a high alert posture, with their heads and necks stretched out to signal the presence of a dangerous intruder [43]. This occurred 35 times overall, not only when predatory birds and jackal chases but also when a lion pride or a baboon troop with a dominant male approached the birds in the scrub in the early morning.
3.7. Interspecific association
During August 2021, Blue Wildebeest (Connochaetes taurinus) daily came out into the open to lie down or stand still for hours in a herd with guineafowl foraging nearby or even within it (see Table 9). All such associations were observed between 11:00 and 16:00 daily, with the guild lasting up to an hour each time. In one case, a Blue Wildebeest chased an approaching jackal away. After drinking water, groups of Burchell’s Zebras, Impalas, Greater Kudus and African Elephants also spent lengthy periods (30 minutes to several hours) on the plain. Yet, guineafowl showed no propensity to associate with them.
3.8. Jackal-Targeted Guineafowl Groups
On 26 occasions, jackals were observed to attack guineafowl, but only one kill was reported. Roaming jackals were frequently spotted on the plain, in full view of guineafowl groups throughout the day (0.874 jackals/hour recorded on average; Supplementary S8 and S9). Photographs showing unique markings on individual jackals (JHvN, unpublished work) suggested that at least five individuals lived permanently in and around the plain during winter. Overall, we observed at least five different hunting strategies by the jackals, each eliciting a different response by the guineafowl (see details in Table 10). On 20 occasions, jackals were seen strolling past or sitting 10 to 15 m away from feeding guineafowl with their heads down. In one case, a jackal chased a guineafowl, which caused others to flow up and land repeatedly across a 100 m straight line. Jackals were more visible on the plain early morning and late afternoons (Table 11).
3.9. Guineafowl Response to Predatory Bird Attacks
On 14 occasions, predatory birds were observed to attack with only one kill reported by a Martial Eagle (Polemaetus bellicosus). The approaching predatory birds caused guineafowl groups to fly low over the ground (3 to 4 m above ground level) and hide under trees with overhanging branches, where they remained for up to an hour. On five occasions under these conditions, some group members repeatedly moved about 5 meters away from the shelter as an exit strategy. In other words, if no further attempts at aerial attack occurred, the rest of the group also emerged, and all individuals commingle to their feeding spot. Of about 100 birds, 30 flew down to the plain when attacked by a predatory bird, remaining there for about 15 to 20 minutes before flying back to the trees 40 minutes after sunset.
Deducing predatory birds’ particular hunting strategy, as done for the jackals, was not feasible. However, guineafowl was attacked either on the ground or in a tree. During one unidentified predatory bird overflight at Tau, the group flushed out into cover in all directions. On three occasions, such overflights prompted the groups to gather in one unit and to produce ear-deafening “chit chit chirr” calls, as they remained on the same spot.
3.10. Group Size as Predictor of Predator Attack
Group size (χ2 = 32.6, P < 0.001) was a significant predictor of attack type when the model was controlled by the significant effect of month (χ2 = 22.2, P = 0.004) because February witnessed significantly more predator attacks. Mean group size during predatory bird and jackal attack was 34.86 (range = 5-90, Std. D = 24.55, n = 14) and 39.73 (range = 2-200, Std. D = 40.14, n = 81), respectively, while it was 15.78 (range = 0-150, Std. D = 30.49, n = 817) in the absence of predators. Thus, only jackal attack likelihood increased significantly with larger guineafowl groups (Figure 4).
3.11. Estimated Avoidance Time
The average composite group size attacked, resulting in evasive action due to predators, was 43.62 birds (range = 1-200, standard deviation = 41.06, n = 43). The mean avoidance (i.e., position manoeuvring by prey before predator attack) [44] time for jackals (15.13 min) and predatory birds (14.66 min) after an attack was not statistically different (Table 12). The estimated daily feeding time that guineafowl lost due to jackal and predatory bird harassment was 30.90 min and 16.49 min, respectively (Table 12). The rate of jackal presence on the plain far outnumbered that of the predatory birds, being 113 vs 14 (Table 12).
4. Discussion
Grouping behaviour in Helmeted Guineafowl serves various vital functions such as food finding and cooperative breeding [31]. However, its role in an anti-predator strategy has never been studied. These birds lived under constant threat from predators, especially Black-backed Jackals. In Borakalalo Game Reserve (North-west province, South Africa), larger guineafowl groups are known to be attacked more frequently than the smaller ones, especially in February when juvenile birds join the group. Jackals also stalked guineafowl in the Krugersdorp Game Reserve (Gauteng province, South Africa) during winter and, in one case, was seen killing a guineafowl (JHvN, unpublished work).
Territorial jackals were often found to roam across the favourable foraging grounds for guineafowl, resulting in a permanent spatiotemporal predator–prey overlap. In such a condition, it is conceivable that prey species developed specific defensive strategies to survive, if trade-offs are likely to occur in a predator-riddled environment [45,46]. Predatory birds neither kept guineafowl away from their favourite grounds. In Krugersdorp Game Reserve, the birds showed strict site fidelity while killing some of them, probably carried out by predatory birds in most cases. However, the birds kept on coming back to the old vegetable gardens, which was a favourable 6-ha feeding spot [47] where 27 guineafowl were killed at a rate of about two birds per month from April 1981-June 1982 (JHvN, unpublished work).
4.1. Habitat Use and Lethal Space
Madiwke plain is a favourable feeding ground for guineafowl due to the high trophic level caused by elephant and antelope dung and droppings, respectively. Guineafowl daily presence during winter suggests that the surrounding bushes do not offer enough nourishment. Not only do these droppings have seeds picked out to consume, but they were also overturned by guineafowl as they searched for invertebrates below them [48] In addition, the manure fertilised the soil, and during the rainy season, it became a lust land with grass seeds for guineafowl to feed on.
We believe that guineafowls have the skills to counteract predation in and around the plains, which enables them to remain in a lethal space (open plain). Averting predators and aerial predators was possible through behavioural compensation and specific landscape features such as nearby bushes with overhanging branches, foliage, and grass cover to conceal during attacks. In other study areas, such as Krugersdorp Game Reserve (South Africa), Helmeted Guineafowl gather in small open vegetable gardens to feed during winter, even though predatory kills were recorded regularly on this site [47]. Ostensibly, this site fidelity is subject to the net sum of kills being equal to or lower than group reproductive output. Otherwise, the birds would roam over larger areas [49].
4.2. Terrestrial Predation
Although jackals have been mainly described as scavengers, they are also active hunters [50]. We found a correlation between guineafowl group size and the presence of jackals, suggesting that these canids are attracted by larger groups of this bird. Even though this positive relationship followed an exponential trend, it did not reach the asymptote, suggesting that the maximum group size in this bird population was either not large enough or may be constrained by predation because too large groups exponentially increase conspicuousness and harassment from predators [51]. Moreover, based on our observations, jackals were not perceived as hazardous predators, which accounts for the guineafowl tolerating their continuous presence 5 to 10 m away while foraging. However, the birds kept contact with each other with their soft “so wee wee wee” calls or clicks, which were likely to increase in volume and urgency whenever a jackal made a sudden dash towards them. This behaviour allowed guineafowl to evade the attack and continue foraging a few meters away, pointing to the importance of exploiting productive patches without having the urge to move away from their predators [52]. The jackals preferentially approached guineafowl when they occurred in larger groups, usually before roosting or sunning in the morning. In the current study, the guineafowl could be seen to become increasingly agitated when jackals hunted in pairs, especially when the groups flushed under cover. On such occasions, the guineafowl often formed tight groups of five to ten birds, mobbing the jackals with loud cackle calls, in response to which the predators could be seen to move away [26,53]. The knock-on effect of the loud calls often alerted other groups, much to the disadvantage of the jackals involved, which tended to stand still in bewilderment for up to 10 m before walking away. The poor skills that jackals possess to catch wild birds are also evident from the relatively low occurrence (3.1%) of their remains in their stomachs [54,55,56]. However, their constant harassment of guineafowl groups curtailed the birds’ feeding time by about 29.72 min daily; therefore, the extent of predation to which guineafowl are exposed could be critical for their survival, especially during dry years marked by poor recruitment in food-depleted habitats [6,57]. Group harassment is known as a long-term strategy aimed at weakening the birds over time [7], which is possibly why the jackals persist despite the low hunting success. It is also arguable that guineafowl have not yet evolved proper defensive mechanisms against an opportunistic predator. The same consideration applies to baboons, which mainly were tolerated by the birds even at short distances even if, on at least one occasion, they were seen to prey upon a guineafowl in Krugersdorp Game Reserve [42]. Not surprisingly, the two species share the same feeding patch in other South African reserves in winter [58]. On the contrary, the bold response to approaching lion pride indicates the felids are adept at hunting guineafowl, as witnessed on one occasion before the onset of this study.
4.3. Aerial Predation
Attacks by predatory birds were primarily stealth and rare, and their presence was not explained by prey group size. However, guineafowl did not tolerate it as they did with the jackals and invariably fled. Guineafowl were particularly wary of predators when ascending to their roost. Unlike other birds, such as the Brown-headed Cowbirds (Molothrus ater), the guineafowl did not enter their roosting trees in a circadian rhythm; instead, they stayed out on the plain until the light was dim [59]. The guineafowl followed sunset timing by ascending trees later during winter, staying still in the darkness, possibly maximising inconspicuousness. Furthermore, they ascended the trees in small groups/ of 2 to 3 birds at a time to prevent attracting predatory birds. They huddled at the launching pad for 20-30 minutes without feeding before ascending trees, suggesting the birds did not stay out longer to increase feeding time.
When a predatory bird alerted an individual or group, other individuals were encouraged to take evasive action together, such as flying off low over the ground in the same direction for cover and huddling under the shade of trees with overhanging branches. This explains why predatory birds mostly attacked composite groups larger than the year-round mean group sizes (40.68 vs 14.26), similar to jackals. Comparatively speaking, larger groups have also been observed to be attacked in other predator-prey systems [7] since this strategy maximises the chances of success for the predator.
Evading predatory birds also prevented the guineafowl from foraging. However, it was less intense than in the case of jackal (16.49 min per day vs. 29 min per day, respectively). The aggregation of smaller guineafowl groups moving in one direction in response to predatory bird attacks seem congruent with the self-organisation theory [60]. It is likely that many guineafowl did not see the predatory birds but still reacted to the flee signals issued by their neighbours and the fluttering of other avian species in nearby trees as warnings. This coordinated movement reduced the cognitive cost of tracking down predator locations, reducing the risk of being caught. The evasive action was coordinated without centralised decision-making (i.e., one or two dominant birds leading the group away from danger) since groups joined and flew immediately to reach cover.
4.4. Surveillance
Guineafowl were exposed to predators during water drinking, standing shoulder to shoulder to minimise risk. They alternated with each other, with some maintaining a stretched neck and heads-up motion to survey the area for predators. Interestingly, the birds must also perform a heads-up movement to swallow water, which seems functional for surveillance. Comparatively, spurfowl - medium-sized galliforms often coexisting with guineafowl in the same habitat - do not hold their heads high when swallowing water at drinking points but withdraw the neck with their beak remaining parallel with their crouching body when swallowing water (JvN, personal observation). Moreover, they often visit drinking points singularly, possibly because single birds attract fewer predators than groups.
As opposed to drinking, the percentage of heads-up postures was less frequent in the scrub (5.3%) and on the open plain (1.10%) rather than at the waterhole (35%) (Table 8). In the scrub, where visual contact is limited, guineafowl foraged relatively close to each other (0.4 m apart vs 0.8 m in the open), relying on the contact calls emitted by group members, more specifically a continuous soft “so wee wee wee” call. Hence, the head-up posture is seemingly less critical when the birds are in close contact with cover. Crested Guineafowl (Guttera edouardi) also emits these contact calls in social groups [43].
When the guineafowl foraged in the open in late afternoons, the heads-up rate was expected to be higher than 1.10%, since predator exposure is high. In such a situation, groups are larger (14.88 vs 7.64) but less dense (0.8 m vs 0.1 m SD) than in the scrub (Table 8). The plausible explanation is that guineafowl keep their heads down to maximise food intake and keep their metabolic rate high when roosting in trees during cold winter nights [61]. Instead of heads up, guineafowl rely on a louder call, consisting of a “chit chit chit”, to warn their neighbours when a threat is detected. Likely, guineafowl were often found to forage close to Blue Wildebeests because these antelopes chase away approaching jackals, thus creating a safe environment for the birds to optimise feeding unhindered. The larger SD that guineafowl kept in a composite group when out in the open - granting them enough space to take flight and avoid collisions with their neighbours when escaping - possibly discourages sudden attacks by jackals. However, when they huddle to prepare for roosting or sunning, they form a more compact group with less social distance, making them vulnerable but allowing for better predator detection. The birds also stand shoulder to shoulder when drinking water and maintain a short social distance during idling from 11:00 to 12:00, allowing effective group communication when a predator approaches.
5. Conclusions
Our findings concluded that guineafowl grouping behaviour has evolved in this species to withstand predation. This conclusion is based on the following results: 1) guineafowl change their group structure, which allows them to maximise predator surveillance and avoidance during their daily activities required for survival across their home range encompassing different habitat features. 2) The limited individual investment for predator surveillance and avoidance allows guineafowl to remain on feeding patches even in the presence of jackals, whose threat level was perceived as tolerable (as supported by the low killing rate). 3) The high attack rate on larger groups ascending roost trees is mitigated by the many-eyes advantage for predator detection. 4) Groups serve as information centres for their members, which, with their physical signs (e.g., heads-up) and calls, not only warn about predation presence but also provide guidance for its avoidance, such as fleeing towards the canopy of a tree. 5) Finally, grouping allows guineafowl to confront a predator directly by mobbing and ear-deafening cackles.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplementary S1. Photograph indicating the measurements collected to calculate the mean social distance (SD) between guineafowl in groups. A = group spread; B = scale. The size of the group below equates to eight birds (in a straight line). Supplementary S2. Measurements (obtained from a livestream webcam as reported on the computer screen) used to determine the social distance between guineafowl in groups. S = spread in mm (two farthest guineafowl in a straight line – see Supplementary S2); scale (mm) = broadside size of a guineafowl on the screen; group size = the number of birds in a straight (horizontal) line through the group (for the sake of clarity, residual birds in front of, or behind, the line of the group were excluded). Constant = broadside length of a live bird (0.350 m) or frontal breadth (0.175 m) of a drinking bird. Data were ordered in this table based on habitat (scrub or open) and activity (huddling, sunning, water drinking), instead of on date of observation. Supplementary S3. Guineafowl group sizes and descriptive statistics used to formulate a two-dimensional graph showing how group sizes varied during consecutive months and at various times of the day. The graph also allowed monthly and hourly mean group sizes to be formulated from March to August 2021. Supplementary S4. Guineafowl group sizes recorded in four sequential time slots during August 2021. Supplementary S5. Guineafowl group sizes in the scrub and open plain recorded simultaneously on different days and hours. Supplementary S6. Group size, just before roosting in the trees. Counts were made at the launching pad in the evening. Supplementary S6. Webcam data used to determine the percentage of heads-up counts of guineafowl in groups. Supplementary S7. Hourly Black-backed Jackal sightings and mean guineafowl group sizes in Tau (Madikwe Game Reserve) from April to August 2021 (see Supplementary S3 and S4 for social distance data and mean group sizes, respectively). Supplementary S8. Correlation between mean guineafowl group size, jackal/hour and social distance between group members at Tau from April to August 2021.
Author Contributions
Johann van Niekerk: conceptualisation, data curation, formal analysis, funding acquisition, investigation, writing – original draft, writing – review and editing; Giovanni Forcina: formal analysis, methodology, writing – review and editing, funding acquisition, visualisation. All authors read and approved the final manuscript; Rodrigo Megía-Palma: conceptualisation, writing – review and editing
Funding
This research was partly funded by a research grant from Department of Environmental Sciences, University of South Africa. Partial support was provided by the Spanish Government, Ministry of Universities (“María Zambrano” – Next Generation EU) to GF.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The Authors are thankful to AfriCam and his volunteers for their assistance with livestream cameras.
Conflicts of Interest
The author declares no conflicts of interest. The author is responsible for the results and the interpretations of the materials in this article.
References
- Lovette, I.J.; Fitzpatrick, J.W. Handbook of Bird Biology. Cornell Lab of Ornithology: Ithaca, NY, USA, 2016.
- Ling, H.; Mclvor, G.E.; van der Vaart, K.; Vaughan, R.T.; Thornton, A.; Ouellette, N.T. Local interactions and their group-level consequences in flocking jackdaws. Proc. R. Soc. B: Biol. Sci. 2019, 286, 20190865. [Google Scholar] [CrossRef]
- Storms, R.F.; Carere, C.; Zoratto, F.; Hemelrijk, C.K. Complex patterns of collective escape in starling flocks under predation. Behav. Ecol. Sociobiol. 2019, 73, 10. [Google Scholar] [CrossRef] [PubMed]
- Cresswell, W. Predation in bird populations. J. Ornithol. 2010, 152, 251–263. [Google Scholar] [CrossRef]
- Crowe, T.M. Limitation of population in the helmeted guineafowl. S. Afr. J. Wildl. Res. 1978, 8, 117–126. [Google Scholar]
- Newton, I. Group Limitation in Birds. Academic Press: London, UK, 2003.
- Cresswell, W.; Quinn, J.L. Predicting the optimal prey group size from predator hunting behaviour. J. Anim. Ecol. 2011, 80, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, C. Grouping and predation. In Encyclopedia of Evolutionary Psychological Science; Shackelford, T., Weekes-Shackelford, V., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- 9Cresswell,W. Flocking is an effective anti-predator strategy in redshanks, Tringa tetanus. Anim. Behav. 1994, 47, 433–442.
- 1Carere, C.; Montanino, S.; Moreschini, F.; Zoratto, F.; Chiarotti, F.; Santucci, D.; Alleva, E. Aerial flocking patterns of wintering starlings, Sturnus vulgaris, under different predation risk. Anim. Behav. 2009, 77, 101–107. [Google Scholar] [CrossRef]
- Morrell, L.J.; Ruxton, G.D.; James, R. Spatial positioning in the selfish herd. Behav. Ecol. 2011, 22, 16–22. [Google Scholar] [CrossRef]
- Hamilton, W.D. Geometry for the selfish herd. J. Theor. Biol. 1971, 31, 295–311. [Google Scholar] [CrossRef]
- Ancel, A.; Gilbert, C.; Poulin, N.; Beaulieu, M.; Thierry, B. New insights into the huddling dynamics of emperor penguins. Anim. Behav. 2015, 110, 91–98. [Google Scholar] [CrossRef]
- Colorado Zuluaga, G.L. Why animals come together, with the special case of mixed-species bird flocks. Rev. EIA. Esc. Ing. Antioq. 2013, 146, 999–1023. [Google Scholar]
- Clark, C.W.; Mangel, M. Foraging and Flocking Strategies: Information in an Uncertain Environment. Am. Nat. 1984, 123, 626–641. [Google Scholar] [CrossRef]
- Lima, S.L. Vigilance while feeding and its relation to the risk of predation. J. Theor. Biol. 1987, 124, 303–316. [Google Scholar] [CrossRef]
- Jackson, A.L.; Beauchamp, G.; Broom, M.; Ruxton, G.D. Evolution of anti-predator traits in response to a flexible targeting strategy by predators. Proc. R. Soc. B: Biol. Sci. 2006, 273, 1055–1062. [Google Scholar] [CrossRef]
- Palmer, M.S.; Packer, C. Reactive anti-predator behavioral strategy shaped by predator characteristics. PLoS ONE 2021, 16, e0256147. [Google Scholar] [CrossRef] [PubMed]
- Whittingham, M.J.; Evans, K.L. The effects of habitat structure on predation risk of birds in agricultural landscapes. Ibis 2004, 146, 210–220. [Google Scholar] [CrossRef]
- Tellería, J.L.; Virgós, E.; Carbonell, R.; Pérez-Tris, J.; Santos, J. Behavioural responses to changing landscapes: Flock structure and antipredator strategies of tits wintering in fragmented forests. OIKOS 2001, 95, 253–264. [Google Scholar] [CrossRef]
- Jovani, R.; Schielzeth, H.; Mavor, R.; Oro, D. Specificity of grouping behaviour: Comparing colony sizes for the same seabird species in distant populations. J. Avian Biol. 2012, 43, 397–402. [Google Scholar] [CrossRef]
- Ramírez-Santos, P.L.; Enríquez, P.; Raúl Vázquez-Pérez, J.; Rangel-Salazar, J.L. Bird behaviour during prey–predator interaction in a tropical forest in Mexico. In Owls; Mikkola, H (Ed.) IntechOpen, 2018. [CrossRef]
- Abdulwahab, U.A.; Osinubi, S.T.; Abalaka, J. Risk of predation: A critical force driving habitat quality perception and foraging behavior of granivorous birds in a Nigerian forest reserve. Avian Res. 2019, 10, 33. [Google Scholar] [CrossRef]
- Park, K.J.; Graham, J.; Calladine, J.; Wernham, C.W. Impacts of birds of prey on gamebirds in the UK: A review. Ibis 2008, 150, 9–26. [Google Scholar] [CrossRef]
- Berruti, A. The AGRED Guide to Gamebird Management in South Africa. African Gamebird Research Education and Development Trust, Johannesburg, South Africa, 2011.
- Skead, C.J. A study of the Crowned Guineafowl Numida meleagris coronata Gurney. Ostrich 1962, 33, 51–65. [Google Scholar] [CrossRef]
- Rose, M. ; The guineafowl and the buzzard. Witwatersrand Bird Club. News Sheet 1967, 58, 7–8. [Google Scholar]
- Rose, M. Crowned guineafowl behaviour. Witwatersrand Bird Club. News Sheet 1966, 55, 2. [Google Scholar]
- Monty, K. Helmeted guineafowl spars with black sparrowhawk. Albatross 1983, 271, 4. [Google Scholar]
- Van Niekerk, J.H. Interflock movement in a population of Helmeted Guineafowl Numida meleagris at the Krugersdorp Game Reserve, Gauteng province, South Africa. Ostrich 2009, 80, 201–204. [Google Scholar] [CrossRef]
- Van Niekerk, J.H. Social organization of a flock of Helmeted Guineafowl (Numida meleagris) at the Krugersdorp Game Reserve, South Africa. Chin. Birds 2010, 1, 22–29. [Google Scholar] [CrossRef]
- Papageorgiou, D.; Christensen, D.C.; Gabriella, E.C.; Gall, J.A.; Klarevas-Irby, B.; Nyaguthii, B.; Couzin, I.D.; Farine, D.R. The multilevel society of a small-brained bird. Curr. Biol. 2019, 29, 1120–1121. [Google Scholar] [CrossRef] [PubMed]
- Van Niekerk, J.H. Notes on habitat use by guineafowl in the Krugersdorp Game Reserve, South Africa. S. Afr. J. Wildl. 2002, 2, 166–168. [Google Scholar]
- Viljoen, P.J. AGRED’s Gamebirds of South Africa. African Gamebird Research Education and Development Trust, Johannesburg, South Africa, 2005.
- Van Niekerk, J.H. Roosting requirements of Helmeted Guineafowl Numida meleagris on Highveld grain and livestock farms with alien tree groves, Gauteng province, South Africa. Ostrich 2019, 90, 37–40. [Google Scholar] [CrossRef]
- Papadopoulou, M.; Hildenbrandt, H.; Sankey, D.W.E.; Portugal, S.J.; Hemelrijk, C.K. Self-organization of collective escape in pigeon flock. PLoS Comput. Biol. 2022, 18, e1009772. [Google Scholar] [CrossRef]
- Mucina, L.; Rutherford, M.C. (Eds.), The Vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19. South African National Biodiversity Institute, Pretoria. 2006.
- Hayward, M.W.; Hayward, M.D. Waterhole use by African fauna. S. Afr. J. Wildl. 2012, 42, 117–127. [Google Scholar] [CrossRef]
- Magige, F.J. Flock-size effect on scanning behaviour of Maasai Ostrich Struthio camelus massaicus. Scopus 2017, 37, 38–41. [Google Scholar]
- Petrželková, K.J.; Zukal, J. Does a live barn owl (Tyto alba) affect emergence behavior of serotine bats (Eptesicus serotinus)? Acta Chiropt. 2003, 5, 177–184. [Google Scholar] [CrossRef]
- Kelm, D.H.; Langheld, M.; Nogueras, J.; Popa-Lisseanu, A.G.; Ibáñez, C. Continuous low-intensity predation by owls (Strix aluco) on bats (Nyctalus lasiopterus) in Spain and the potential effect on bat colony stability. R. Soc. Open Sci. 2023, 10, 230309. [Google Scholar] [CrossRef] [PubMed]
- Van Niekerk, J.H. Some socio-biological features of Crowned Guineafowl in the Krugersdorp Game Reserve. Bokmakierie 1980, 32, 102–108. [Google Scholar]
- Van Niekerk, J.H. Vocal behavior of Crested Guineafowl (Guttera edouardi) based on visual and sound playback surveys in the Umhlanga Lagoon Nature Reserve, KwaZulu-Natal province, South Africa. Avian Res. 2015, 6, 13. [Google Scholar] [CrossRef]
- Weihs, D.; Webb, P.W. Optimal avoidance and evasion tactics in predator-prey interactions. J. Theor. Biol. 1984, 106, 189–206. [Google Scholar] [CrossRef]
- Bonter, D.N.; Zuckerberg, B.; Sedgwick, C.W.; Hochachka, W.M. Daily foraging patterns in free-living birds: Exploring the predation–starvation trade-off. Proc. Royal Soc. B. 2013, 280, 20123087. [Google Scholar] [CrossRef]
- Olson, R.S.; Haley, P.B.; Dyer, F.C.; Adami, C. Exploring the evolution of a trade-off between vigilance and foraging in group-living organisms. R. Soc. Open Sci. 2015, 2, 150135. [Google Scholar] [CrossRef]
- Van Niekerk, J.H. Habitat preferences of Helmeted Guineafowl Numida meleagris in the Krugersdorp Game Reserve, Gauteng province, South Africa. Ostrich 2013, 84, 153–156. [Google Scholar] [CrossRef]
- Van Niekerk, J.H. Observations of Red-billed Spurfowl (Pternistis adspersus) in the arid Molopo Nature Reserve, North West Province, South Africa. Chin. Birds 2011, 2, 117–124. [Google Scholar] [CrossRef]
- Newton, I. Predation and limitation of bird numbers. In Current Ornithology; Power, D.M. (Ed.)., vol. 11. Springer, Boston, MA, USA,, 1993, pp 143–198. [CrossRef]
- Avery, G.; Avery, D.M.; Braine, S.; Loutit, R. Prey of coastal black-backed jackal Canis mesomelas (Mammalia: Canidae) in the Skeleton Coast Park, Namibia. J. Zool. 1987, 213, 81–94. [Google Scholar] [CrossRef]
- Alexander, R.D. The evolution of social behavior. Annu. Rev. Ecol. Syst. 1974, 5, 325–383. [Google Scholar] [CrossRef]
- Creswell, W. Non-lethal effects of predation in birds. Ibis 2008, 150, 3–17. [Google Scholar] [CrossRef]
- Kalb, N.; Randler, C. Behavioral responses to conspecific mobbing calls are predator-specific in great tits (Parus major). Ecol. Evol. 2019, 9, 9207–9213. [Google Scholar] [CrossRef] [PubMed]
- Grafton, R.N. Food of the black-backed jackal: A preliminary report. Afr. Zool. 1965, 1, 41–53. [Google Scholar] [CrossRef]
- Humphries, D.H. The Ecology of Black-Backed Jackal (Canis mesomelas) on Farmlands in the Midlands of KwaZulu-Natal, South Africa. Master of Science in the Discipline of Ecology, School of Life Sciences, College of Agriculture, Science and Engineering, University of KwaZulu-Natal, Pietermaritzburg Campus, 2014.
- Minnie, l.; Avenant, N.L.; Kamler, J.; Butler, H.; Parker, D.; Rouilly, M.; du Plessis, J.; Do Linh San, E. A conservation assessment of Canis mesomelas. In The Red List of Mammals of South Africa, Swaziland and Lesotho; Child, M.F., Roxburgh, l., Do Linh San, E., Raimondo, D., Daviesmostert, H.T. (Eds.); South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa. 2016.
- Potts, G.R. Partridges. Collins. London, UK, 2013.
- Branch, W.R. Predation on Helmeted Guineafowl by Grey-footed Chacma Baboon in Mashatu Game Reserve, north east Botswana. Biodivers. Observations 2017, 8, 1–5. [Google Scholar]
- Woller, A.; Gonze, D. The Bird Circadian Clock: Insights from a Computational Model. J. Biol. Rhythms 2013, 28, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Couzin, I.; Krause, J. Self-organisation and collective behavior in vertebrates. Adv. Study Behav. 2003, 32, 1–75. [Google Scholar] [CrossRef]
- Sweet, K.A.; Sweet, B.P.; Gomes, D.G.E.; Francis, C.D.F.; Barber, J.R. Natural and anthropogenic noise increase vigilance and decrease foraging behaviors in song sparrows. Behav. Ecol. 2022, 33, 288–297. [Google Scholar] [CrossRef]
Figure 1.
Aerial footage of the study area at Tau Waterhole (Madikwe Game Reserve), showing different habitat features: water body (2.5 ha), open plain (ca. 3 ha) and the scrub (ca. 2.5 ha). The distance from the primary roosting trees (where the guineafowl roosted together from April to June) to the secondary roosting trees (where 40 birds roosted after breaking away from the roosting composite group during July and August) was about 230 m. The distance from the launching pad to the primary roosting trees was about 40 m. The launching pad was situated where all the groups converged to fly up into the primary trees and roost communally. The insets at the top right corner show the map of the study area and the position of Madikwe Game Reserve in South Africa.
Figure 1.
Aerial footage of the study area at Tau Waterhole (Madikwe Game Reserve), showing different habitat features: water body (2.5 ha), open plain (ca. 3 ha) and the scrub (ca. 2.5 ha). The distance from the primary roosting trees (where the guineafowl roosted together from April to June) to the secondary roosting trees (where 40 birds roosted after breaking away from the roosting composite group during July and August) was about 230 m. The distance from the launching pad to the primary roosting trees was about 40 m. The launching pad was situated where all the groups converged to fly up into the primary trees and roost communally. The insets at the top right corner show the map of the study area and the position of Madikwe Game Reserve in South Africa.
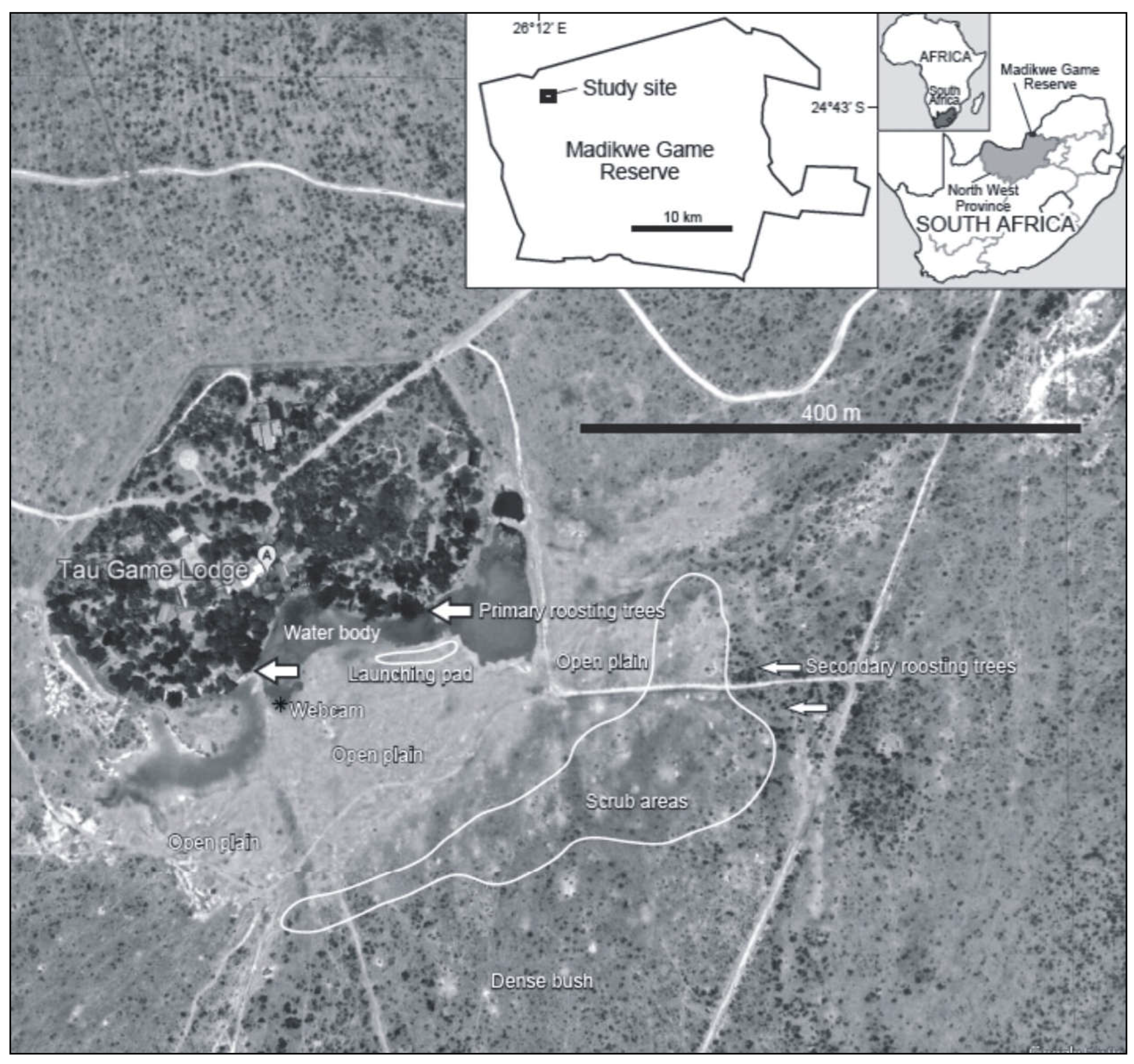
Figure 2.
Two-dimensional temporal illustration of group dynamics of the Helmeted Guineafowl population across months (1st dimension) and daytime (2nd dimension) during winter.
Figure 2.
Two-dimensional temporal illustration of group dynamics of the Helmeted Guineafowl population across months (1st dimension) and daytime (2nd dimension) during winter.
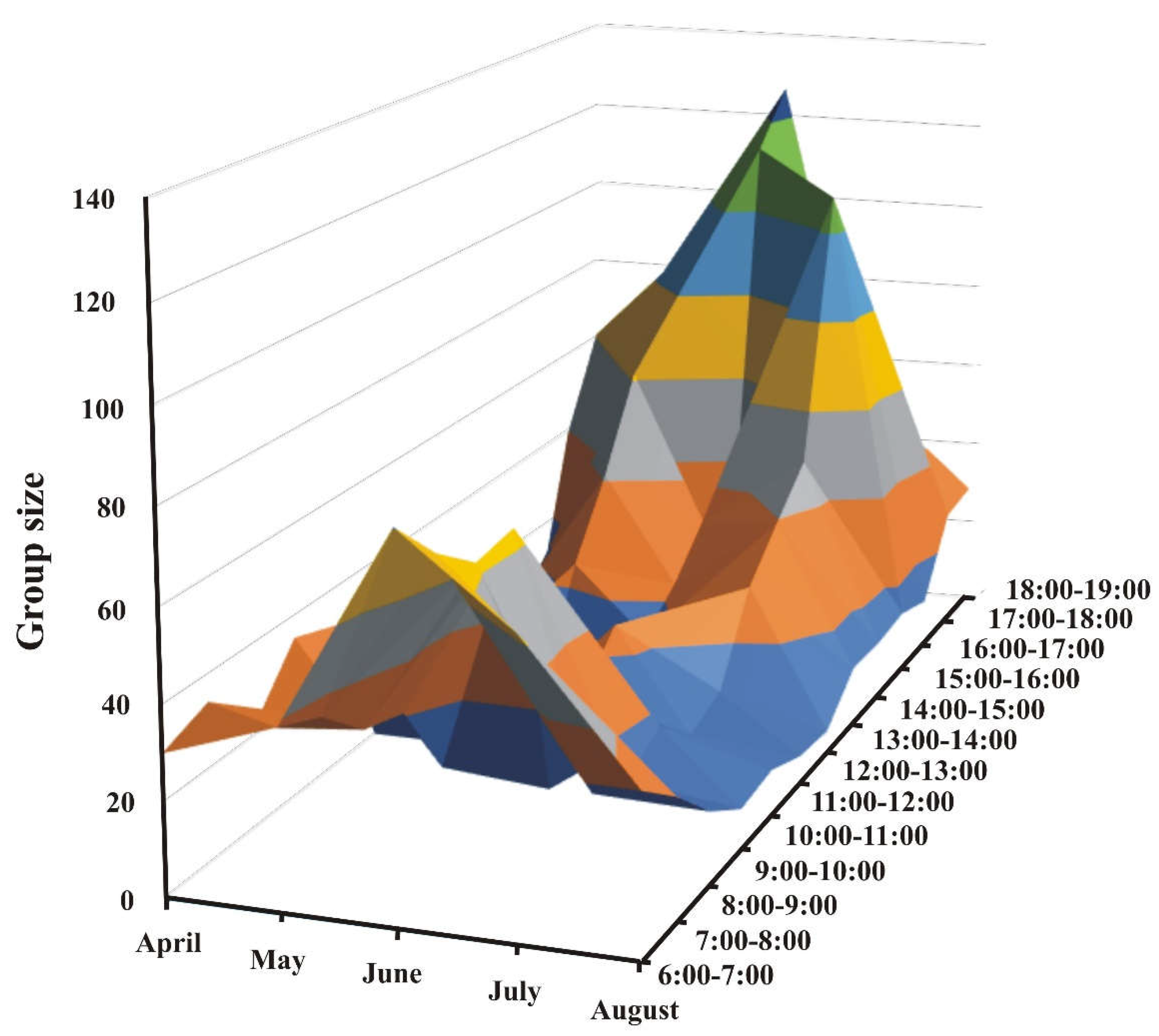
Figure 3.
Winter roosting times of guineafowl following changes in sunset times. Start: when group members start flying into the roosting trees; end: when the last bird of the group flies into the roosting trees.
Figure 3.
Winter roosting times of guineafowl following changes in sunset times. Start: when group members start flying into the roosting trees; end: when the last bird of the group flies into the roosting trees.
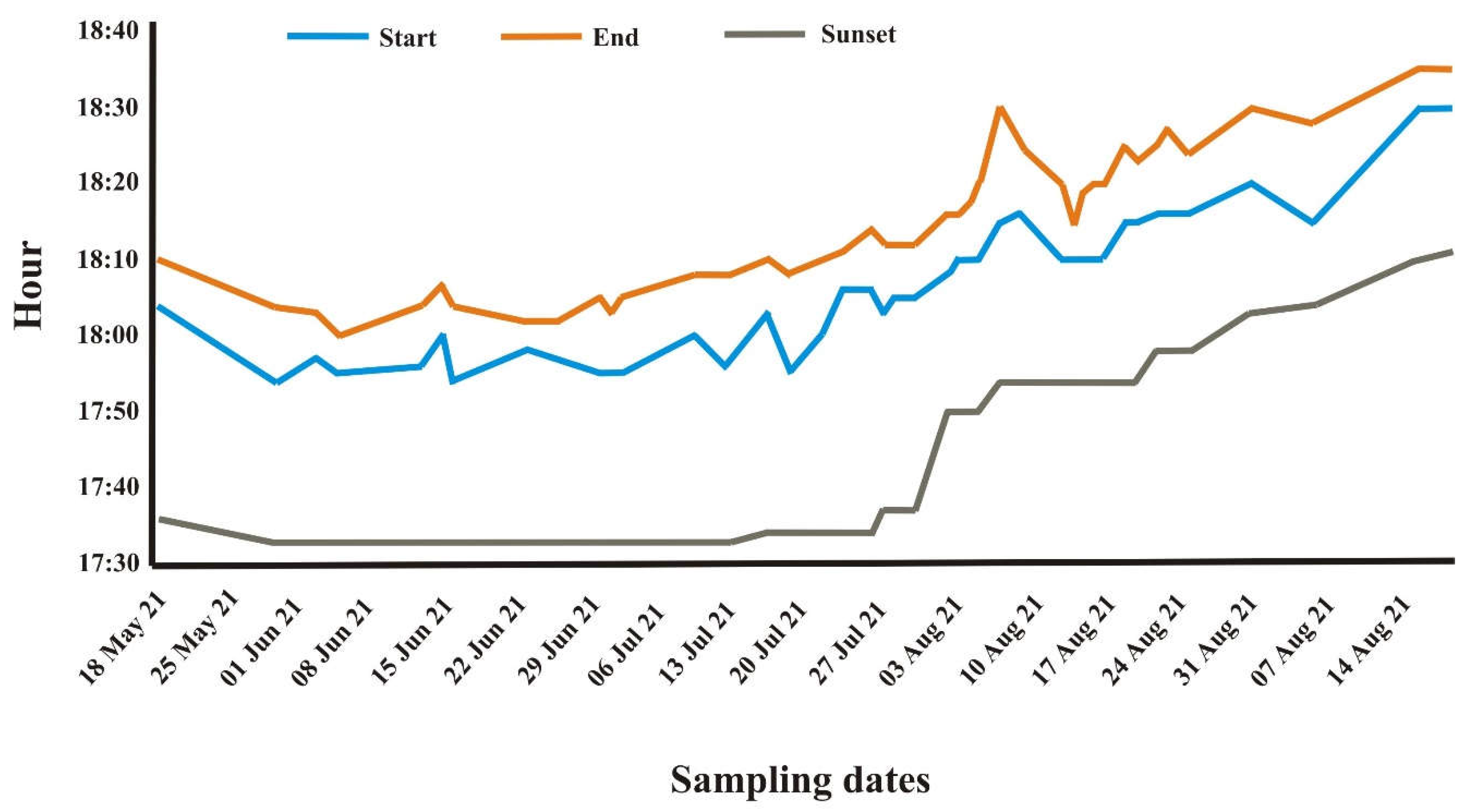
Figure 4.
Predator attack likelihood as a function of guineafowl group size. Shadow area represents standard error.
Figure 4.
Predator attack likelihood as a function of guineafowl group size. Shadow area represents standard error.

Table 1.
Sub-sampling from 411 groups observed from August 2020 to August 2021 for various themes investigated.
Table 1.
Sub-sampling from 411 groups observed from August 2020 to August 2021 for various themes investigated.
| Description | Suitable sub-samples |
|---|---|
| Calculation of summer group sizes | 13 |
| Calculation of mean group sizes at different times of the day | 174 |
| SD Calculation in groups at different times of the day | 381 |
| Calculation of mean monthly group sizes getting ready to roost | 82 |
| Calculation of time it takes for communal rooting group to ascend rooting tree | 42 |
| Calculation of time it takes for communal rooting group to descend rooting tree | 29 |
| Selected photographs of groups to conduct head-up counts (surveillance behaviour) | 190 |
| Observations revealing associations between guineafowl and Blue Wildebeest | 17 |
| Groups selected to correlate jackal/hour vs. mean group sizes of guineafowl | 393 |
| Calculation of SD in groups in different parts of the site, drinking and huddling | 411 |
Table 2.
Sampling days and times at Tau Waterhole (Madikwe Game Reserve). The low sampling time in December 2020 is ascribed to the absence of guineafowl from the study site due to breeding. Login time was 5 minutes, while “additional time” refers to the extra time added to monitor predator–prey interaction.
Table 2.
Sampling days and times at Tau Waterhole (Madikwe Game Reserve). The low sampling time in December 2020 is ascribed to the absence of guineafowl from the study site due to breeding. Login time was 5 minutes, while “additional time” refers to the extra time added to monitor predator–prey interaction.
| Month | Number of days | Number of logins | Mins observed | Additional time/mins |
|---|---|---|---|---|
| Aug. 2020 | 4 | 36 | 180 | 0 |
| Sep. 2020 | 6 | 54 | 270 | 0 |
| Oct. 2020 | 16 | 144 | 720 | 0 |
| Nov. 2020 | 16 | 144 | 720 | 0 |
| Dec. 2020 | 6 | 54 | 270 | 0 |
| Jan. 2021 | 19 | 171 | 855 | 0 |
| Feb. 2021 | 18 | 162 | 810 | 0 |
| Mar. 2021 | 28 | 252 | 1260 | 0 |
| Apr. 2021 | 25 | 225 | 1125 | 0 |
| May 2021 | 26 | 234 | 1170 | 60 |
| Jun. 2021 | 20 | 180 | 900 | 40 |
| Jul. 2021 | 23 | 207 | 1035 | 155 |
| Aug. 2021 | 28 | 252 | 1260 | 245 |
| Total | 235 | 2115 | 10575 | 500 |
Table 3.
Descriptive statistics and Kruskal-Wallis tests comparing group sizes during different times of the day (Supplementary S4).
Table 3.
Descriptive statistics and Kruskal-Wallis tests comparing group sizes during different times of the day (Supplementary S4).
| Time | Mean group size | Std. D | Sample | Range |
| 07:00 | 15.61 | 16.77 | 18 | 2-55 |
| 09:00 | 6.35 | 4.42 | 39 | 1-22 |
| 16:00 | 12.08 | 11.55 | 67 | 2-80 |
| 18:00 | 20.16 | 19.86 | 50 | 3-90 |
| Pair | Difference | H-statistics | Critical value | p-value |
| 7:00 – 9:00 | 2.5 | 4.01 | 6.23 | 0.04 |
| 7:00-16:00 | 2.5 | 0.00 | 6.23 | 0.96 |
| 7:00-18:00 | 3.5 | 2.38 | 6.23 | 0.12 |
| 9:00-16:00 | 5 | 11.40 | 6.23 | 0.00 |
| 9:00-16:00 | 6 | 23.95 | 6.23 | 0 |
| 16:00-18:00 | 1 | 5.90 | 6.23 | 0.015 |
Table 4.
Descriptive statistics and Kruskal-Wallis test comparing group sizes in different habitats and engaging into different activities.
Table 4.
Descriptive statistics and Kruskal-Wallis test comparing group sizes in different habitats and engaging into different activities.
| Activity/Place | Mean social distance | Std. D | Sample | Range |
| Plain | 0.835 | 1.53 | 249 | - 0.280 - 11.550 |
| Scrub | 0.274 | 0.537 | 144 | - 0.277 - 3.080 |
| Huddling | 0.034 | 0.135 | 8 | - 0.194 - 0.210 |
| Drinking | 0.071 | 0.091 | 10 | - 0.021 - 0.190 |
| Pair | Difference | H-statistics | Critical value | p-value |
| Plain-scrub | 0.22 | 22.88 | 6.23 | 0.00 |
| Plain-huddling | 0.28 | 6.42 | 6.23 | 0.01 |
| Plain-drinking | 0.31 | 6.26 | 6.23 | 0.01 |
| Scrub-huddling | 0.06 | 0.90 | 6.23 | 0.34 |
| Scrub-drinking | 0.09 | 0.11 | 6.23 | 0.73 |
| Huddling-drinking | 0.03 | 0.95 | 6.23 | 0.32 |
Table 5.
Descriptive statistics and Kruskal-Wallis tests comparing social distances at different times of the day.
Table 5.
Descriptive statistics and Kruskal-Wallis tests comparing social distances at different times of the day.
| Time | Mean | Std. D | Sample | Range |
| 7:00-8:00 | 0.233 | 0.407 | 44 | - 0.24 - 1.61 |
| 8:00-9:00 | 0.382 | 0.594 | 53 | - 0.28 - 2.23 |
| 9:00-10:00 | 0.614 | 1.43 | 21 | - 0.233 - 6.3 |
| 10:00-11:00 | 0.653 | 0 | 10 | - 0.81 - 1.42 |
| 11:00-12:00 | 0.316 | 0.589 | 5 | - 0.142 - 1.14 |
| 12:00-13:00 | 0.408 | 0.54 | 9 | - 0.13 - 1.42 |
| 13:00-14:00 | 0.546 | 0.77 | 8 | - 0.58 - 2.28 |
| 14:00-15:00 | 1.09 | 2.06 | 14 | - 0.19 - 7.91 |
| 15:00-16:00 | 0.889 | 1.47 | 42 | - 0.21 - 6.25 |
| 16:00-17:00 | 0.806 | 1.07 | 73 | - 0.19 - 4.2 |
| 17:00-18:00 | 0.806 | 1.69 | 81 | - 0.24 - 11.55 |
| 18:00-19:00 | 0.439 | 0.72 | 21 | - 0.16 - 2.64 |
| Pair | Difference | H-statistics | Critical value | p-value |
| 6:00-7:00 vs 14:00-15:00 | 0.34 | 8.90 | 8.35 | 0.00 |
| 7:00-8:00 vs 14:00-15:00 | 0.28 | 5.08 | 8.35 | 0.02 |
| 7:00-8:00 vs 15:00 -16:00 | 0.18 | 6.77 | 8.35 | 0.00 |
| 8:00-9:00 vs 11:00 - 12:00 | 0.21 | 1.34 | 8.35 | 0.00 |
| 11:00-12:00 vs 12:00-13:00 | 0.22 | 4.17 | 8.35 | 0.04 |
Table 6.
Descriptive statistics and Kruskal-Wallis tests comparing roosting group sizes during winter.
Table 6.
Descriptive statistics and Kruskal-Wallis tests comparing roosting group sizes during winter.
| Month(s) | Mean | Std. D | Sample | Range |
|---|---|---|---|---|
| March | 60 | 45.82 | 3 | 10-100 |
| April | 47.5 | 21.1 | 10 | 20-80 |
| May | 80.53 | 45.57 | 15 | 30-160 |
| June | 109.21 | 43.36 | 14 | 29-200 |
| July | 91.31 | 39.38 | 16 | 16-140 |
| August | 53.37 | 19.12 | 24 | 30-100 |
| Pair | Difference | H-statistics | Critical value | p-value |
| March-April | 25 | 0.25 | 6.96 | 0.61 |
| March-May | 10 | 0.17 | 6.96 | 0.67 |
| March-June | 40 | 2.71 | 6.96 | 0.09 |
| March-July | 30 | 1.86 | 6.96 | 0.17 |
| March-August | 20 | 0.33 | 6.96 | 0.56 |
| April-May | 15 | 3.27 | 6.96 | 0.07 |
| April-June | 65 | 10.84 | 6.96 | 0.00 |
| April-July | 55 | 7.02 | 6.96 | 0.00 |
| April-August | 5 | 0.60 | 6.96 | 0.43 |
| May-June | 50 | 2.41 | 6.96 | 0.12 |
| May-July | 40 | 0.45 | 6.96 | 0.49 |
| May-August | 10 | 2.67 | 6.96 | 0.10 |
| June-July | 10 | 0.93 | 6.96 | 0.33 |
| June-August | 60 | 14.14 | 6.96 | 0.00 |
| July-August | 50 | 8.95 | 6.96 | 0.00 |
Table 7.
Time difference (in minutes) between the first and last bird that flew into trees to roost (42 observations) and again when the first and last bird descended from the trees (29 observations) in the mornings from 18 May to 14 September 2021. Some fly-down data are missing due to misty mornings and bad weather, which impaired recording.
Table 7.
Time difference (in minutes) between the first and last bird that flew into trees to roost (42 observations) and again when the first and last bird descended from the trees (29 observations) in the mornings from 18 May to 14 September 2021. Some fly-down data are missing due to misty mornings and bad weather, which impaired recording.
| Fly up to roost | Fly down from trees | ||||||
|---|---|---|---|---|---|---|---|
| Star | End | Duration | No. of birds | Start | End | Duration | No. of birds |
| 18:04 | 18:10 | 6 | 50 | 06:31 | 06:35 | 4 | 70 |
| 17:54 | 18:04 | 10 | 120 | 06:30 | 06:35 | 5 | 80 |
| 17:57 | 18:03 | 6 | 120 | 06:32 | 06:35 | 3 | 100 |
| 17:55 | 18:00 | 5 | 100 | 06:36 | 06:38 | 2 | 100 |
| 17:54 | 18:03 | 7 | 130 | 06:38 | 06:40 | 2 | 100 |
| 17:56 | 18:04 | 8 | 130 | 06:38 | 06:40 | 2 | 50 |
| 18:00 | 18:07 | 7 | 100 | 06:40 | 06:44 | 4 | 75 |
| 17:54 | 18:04 | 10 | 100 | 06:38 | 06:43 | 5 | 100 |
| 17:58 | 18:02 | 4 | 120 | 06:39 | 06:42 | 3 | 100 |
| 17:57 | 18:05 | 8 | 70 | 06:40 | 06:44 | 4 | 100 |
| 17:53 | 18:03 | 10 | 100 | 06:44 | 06:46 | 2 | 100 |
| 17:55 | 18:03 | 8 | 100 | 06:42 | 06:44 | 2 | 110 |
| 17:57 | 18:05 | 8 | 140 | 06:43 | 06:55 | 2 | 120 |
| 18:00 | 18:08 | 10 | 100 | 06:40 | 06:42 | 2 | 100 |
| 17:58 | 18:08 | 12 | 100 | 06:42 | 06:44 | 2 | 130 |
| 18:03 | 18:10 | 7 | 50 | 06:39 | 06:42 | 3 | 70 |
| 17:55 | 18:08 | 13 | 120 | 06:37 | 06:40 | 3 | 100 |
| 18:00 | 18:10 | 10 | 110 | 06:36 | 06:38 | 2 | 100 |
| 18:06 | 18:11 | 5 | 90 | 06:35 | 06:37 | 2 | 120 |
| 18:06 | 18:14 | 8 | 50 | 06:32 | 06:34 | 2 | 80 |
| 18:03 | 18:12 | 9 | 100 | 06:38 | 06:39 | 1 | 80 |
| 18:05 | 18:12 | 7 | 100 | 06:34 | 06:35 | 1 | 80 |
| 18:05 | 18:12 | 7 | 70 | 06:36 | 06:38 | 2 | 80 |
| 18:08 | 18:16 | 8 | 120 | 06:31 | 06:33 | 2 | 40 |
| 18:10 | 18:16 | 6 | 110 | 06:29 | 06:30 | 1 | 40 |
| 18:10 | 18:17 | 7 | 130 | 06:21 | 06:22 | 1 | 40 |
| 18:10 | 18:20 | 10 | 100 | 06:10 | 06:14 | 4 | 45 |
| 18:15 | 18:30 | 15 | 90 | 06:10 | 06:11 | 1 | 40 |
| 18:16 | 18:25 | 9 | 70 | 06:00 | 06:02 | 2 | 46 |
| 18:10 | 18:20 | 10 | 65 | ||||
| 18:10 | 18:15 | 5 | 60 | ||||
| 18:10 | 18:19 | 9 | 70 | ||||
| 18:14 | 18:20 | 6 | 80 | ||||
| 18:10 | 18:20 | 10 | 40 | ||||
| 18:15 | 18:25 | 10 | 50 | ||||
| 18:15 | 18:23 | 8 | 50 | ||||
| 18:16 | 18:27 | 11 | 50 | ||||
| 18:16 | 18:24 | 8 | 50 | ||||
| 18:20 | 18:30 | 10 | 40 | ||||
| 18:15 | 18:28 | 9 | 40 | ||||
| 18:30 | 18:35 | 5 | 50 | ||||
| 18:30 | 18:35 | 5 | 50 | ||||
Table 8.
Descriptive statistics and study periods during which data were obtained photographs for head-up counts in groups under different conditions.*= no of groups; **(birds: head-ups); Std. D: standard deviation.
Table 8.
Descriptive statistics and study periods during which data were obtained photographs for head-up counts in groups under different conditions.*= no of groups; **(birds: head-ups); Std. D: standard deviation.
| Activity | Date | No. photos* | No. birds | Head-ups |
Group size | ||||
|---|---|---|---|---|---|---|---|---|---|
| Counts | Ratios** | % | Mean | Range | Std. D | ||||
| Huddle | May-August 2021 | 56 | 2037 | 917 | 1:2.22 | 45.01 | 36.6 | 2-190 | 42.92 |
| Drinking | September 2020 - August 2021 | 33 | 547 | 220 | 1:2.45 | 40.21 | 16.63 | 2-190 | 32.9 |
| Foraging in scrub | August 2020-August 2021 | 29 | 241 | 13 | 1:18.53 | 5.39 | 8.31 | 2-40 | 8.28 |
| Foraging in open | August 2020-August 2021 | 72 | 991 | 11 | 1:90.09 | 1.1 | 13.76 | 2-55 | 12.42 |
| Totals | 190 | 3816 | 1161 | ||||||
Table 9.
Observations of guineafowl groups foraging close to (within 15 m) and within Blue Wildebeest herd on Tau open plain. The table shows the group sizes of both species recorded during such associations.
Table 9.
Observations of guineafowl groups foraging close to (within 15 m) and within Blue Wildebeest herd on Tau open plain. The table shows the group sizes of both species recorded during such associations.
| Date | Guineafowl | Blue Wildebeest |
|---|---|---|
| 9 August 2021 | 20 | 20 |
| 20 | 15 | |
| 10 August 2021 | 30 | 10 |
| 10 | 3 | |
| 15 | 20 | |
| 11 August 2021 | 25 | 24 |
| 5 | 20 | |
| 4 | 30 | |
| 13 August 2021 | 10 | 20 |
| 15 | 20 | |
| 19 August 2021 | 15 | 15 |
| 22 August 2021 | 10 | 9 |
| 24 August 2021 | 7 | 5 |
| 4 | 10 | |
| 27 August 2021 | 10 | 10 |
| Average | 13.33 | 15.4 |
| Ratio | 1 | 1.15 |
Table 10.
Hunting strategies of jackal and guineafowl response strategies observed in Madikwe Game Reserve.
Table 10.
Hunting strategies of jackal and guineafowl response strategies observed in Madikwe Game Reserve.
| Name | Observations | Predator attack strategy | Prey response |
|---|---|---|---|
| Bombshell in the scrub | 3 | Jackals approached a group of guineafowl in pairs, chasing them until they scattered in all directions. | After that, some birds flew to the tops of nearby trees, making a loud “chit chit chirr” call. This distracted one predator while another searched for guineafowl hidden in the grass. The birds in the tree attracted the jackals, which allowed the hidden birds in the grass to fly up into the trees. |
| Surprise attack | 4 | Lone jackals were seen approaching a group of six to ten guineafowl in a non-threatening manner on the plain, lying 5 to 10 meters away from the group. When a bird turned its back on the jackal, it would dash towards it to catch it. | The guineafowl responded by running towards each other first, then all together 10 to 20 meters away, producing a loud “chit chit chirr” call before continuing to feed. |
| The challenge | 4 | A pair of jackals dashed into a group, stopping near the birds and waiting to catch one. | The birds gathered in a dense group, confronting predators by mobbing them and emitting loud calls. The jackals stood motionless for 10 minutes, appearing confused, before walking away as the birds continued foraging. |
| Herding | 20 | Groups were chased by a jackal at the launch pad, where they gathered from different directions before roosting. | The guineafowl responded by forming a close-knit group of about 20 to 30 birds and running short distances, staying close to the launching pad, just ahead of the jackal for several minutes before flying into the roosting trees. |
| Eavesdropping | 4 | This type of hunting occurred when the groups of guineafowl in the scrub were first disturbed or chased by other animals perceived to be hazardous, such as lions and hyenas. The threatening animals caused the guineafowl to flush out, and the jackal quickly appeared on the scene to pursue the hidden birds in the grass cover. | Like the first response above. Some birds distracted predators by making loud calls, allowing others to escape into trees. |
Table 11.
Hourly associations between jackal and guineafowl groups during the day. Std. D: standard deviation.
Table 11.
Hourly associations between jackal and guineafowl groups during the day. Std. D: standard deviation.
| Time of day | Jackal counts | Jackal/hour | Guineafowl/group (mean) | Std. D | Range | Groups | Hours |
|---|---|---|---|---|---|---|---|
| 6:00-7:00 | 18 | 1.81 | 40.91 | 31.85 | 9-135 | 28 | 9.93 |
| 7:00-8:00 | 4 | 0.42 | 30.58 | 30.21 | 1-100 | 43 | 9.93 |
| 8:00-9:00 | 3 | 0.30 | 21.17 | 19.82 | 5-100 | 41 | 9.93 |
| 9:00-10:00 | 4 | 0.40 | 24.1 | 18.2 | 2-85 | 19 | 9.93 |
| 10:00-11:00 | 0 | 0 | 12.54 | 11.82 | 5-35 | 12 | 9.93 |
| 11:00-12:00 | 0 | 0 | 7.63 | 4.65 | 5 - 15 | 12 | 9.93 |
| 12:00-13:00 | 1 | 0.1 | 13.78 | 11.8 | 6-43 | 13 | 9.93 |
| 13:00-14:00 | 4 | 0.40 | 19.58 | 17.29 | 7-80 | 20 | 9.93 |
| 14:00-15:00 | 4 | 0.40 | 21.9 | 19.06 | 5-90 | 35 | 9.93 |
| 15:00-16:00 | 10 | 1 | 34.02 | 17.63 | 3-90 | 40 | 9.93 |
| 16:00-17:00 | 35 | 3.52 | 28.5 | 25.31 | 7-130 | 62 | 9.93 |
| 17:00-18:00 | 30 | 3.02 | 39.92 | 33.5 | 20-150 | 68 | 9.93 |
Table 12.
Guineafowl avoidance behaviour resulting from jackal and predatory bird attacks at Tau waterhole from April to August 2021. Statistical tests include the chi-square (difference between jackal and predatory bird attacks) and Kruskal–Wallis tests (D, E and F). The avoidance resulting from 3 attack attempts in C could not be seen hence the difference between C and D.
Table 12.
Guineafowl avoidance behaviour resulting from jackal and predatory bird attacks at Tau waterhole from April to August 2021. Statistical tests include the chi-square (difference between jackal and predatory bird attacks) and Kruskal–Wallis tests (D, E and F). The avoidance resulting from 3 attack attempts in C could not be seen hence the difference between C and D.
| Code | Parameters | Black-backed Jackal | Predatory birds | Totals | Statistical test |
|---|---|---|---|---|---|
| A | Total webcam observation minutes during winter (plus ad-hoc time added for predator incidents observations) | 4375 plus 208 | 4375 plus 105 | - | - |
| B | Foraging time available per day in min | 360 | 360 | - | - |
| C | Number of attacks recorded | 26 | 14 | 39 | - |
| D | Avoidance attempts recorded | 22 | 12 | 34 | χ2= 3.102, p = 0.07 |
| E | Jackal and predatory birds noted on the plain | 113 | 14 | 127 | χ2= 77.17, p = 0. 00 |
| F | Mean avoidance time per attack in min | 15.13 | 14.66 | - | H= 0.32811, n =35; p = 0.56 |
| G | Avoidance time: The mean avoidance time x the number of attacks = (F x D) (min) | 332.99 | 175.92 | 508.91 | |
| H | Estimated minutes lost foraging per day = B/(A/C) x F | 30.90 | 16.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



