
Submitted:
12 July 2024
Posted:
13 July 2024
You are already at the latest version
Abstract
Early reporting of the introduction and establishment of exotic species is of paramount importance for successful management. Here we report the introduction and rapid spread of the Bare-eyed Pigeon, Patagioenas corensis, on the binational island of Saint-Martin, West Indies. This range-restricted species naturally occurs in the arid coastal areas of Columbia and Venezuela and nearby islands. Its introduction on Saint-Martin represents an expansion of about 1000 km beyond its established native range. Using observations recorded in e-bird and results from a recent field survey, we show that since its introduction, most probably between late 2012 and early 2013, the species is expanding fast in Saint-Martin and has recently broadened its habitat to include anthropized, built areas. The expansion of Bare-eyed Pigeon on Saint-Martin and the neighbouring Leeward Islands, possibly facilitated by climate change in the future, could be a threat to both native columbid species and other bird species, through competition for resources. We therefore recommend to local authorities and stakeholders to rapidly eradicate the species or at least prevent its further spread on Saint-Martin, possibly though listing it as a game species, while it is still possible to do so.
Keywords:
Biological invasion
; Caribbean
; Columbidae
; eBird
; Wildlife trade
1. Introduction
The introduction and spread of exotic species are a major threat to ecosystems, agriculture, health, and biodiversity, particularly in insular environments [1,2,3], and might be accelerated in the coming decades by change in both land use and climate [4,5]. Because of the huge ecological and economic costs associated with biological invasions [6,7], especially in insular ecosystems [8], preventing the introduction or establishment of exotic species is a major concern today for conservation biologists [9].
Although exotic species can be introduced through multiple pathways, trade and transport are regarded as the major drivers of introduction on islands, while land use/cover change appears to be the main determinant of their establishment and spread [9]. Consequently, several countries and small island developing states have reinforced their control at points of entry [10], but only with limited success as records of new exotic species on islands are still regularly reported [11,12,13,14]. Because in most cases, eradication of exotic species on islands is possible only soon after the onset of the invasion process, early reporting of the introduction and establishment of exotic species on islands is of paramount importance for successful management of biological invasions [15]. Early detection of invasive species can be enhanced through various techniques, such as environmental DNA, chemoreception or remote sensing [16]. More recently, the combination of citizen science with professional expertise has been identified as an efficient way of documenting the introduction, establishment, and expansion of exotic species [16,17]. This is particularly true of invasive or expanding bird species that can be easily observed and identified by amateur ornithologists, as illustrated by several studies [18,19,20].
Here we document the recent introduction, establishment, and rapid expansion of the Bare-eyed Pigeon, Patagioenas corensis, originating from north-west South America, on the binational island of Saint-Martin, West Indies, about 1000 km beyond its native range. Using a combination of citizen science and field survey, we quantified the demographic and spatial expansion of the species on the island, based on the chronology of reported observations in eBird, and characterized its habitat occupancy. We discuss the potential consequences of this invasion at the local and regional level and make recommendations for rapid management actions.
2. Materials and Methods
2.1. Study Area
The island of Saint-Martin (hereafter called Saint-Martin) lies between 18.0° and 18.1° North latitude and between 63.2° and 63.0° West longitude, approximately 300 km east of Puerto Rico. It belongs to the Leeward Islands, a group of islands with similar ecological characteristics situated where the northeastern Caribbean Sea meets the western Atlantic Ocean. The island is administratively divided into two parts. The northern part, called Saint-Martin, is a French overseas territory, while the southern part, called Sint Maarten, is a Dutch overseas constituent country. The island landscape is characterized by undulating coastline with sandy beaches and impounded salt ponds, offshore islets, dry landscapes, with some water-dependent vegetation at higher elevation [21].
2.2. Study Species
The Bare-eyed Pigeon has an overall pale, sandy-coloured, plumage, a large pale blue ocular ring with a wider outer area of reddish brown, orange to orange-brown iris, and conspicuous white crescent wing patches contrasting with black flight feathers [22]. The head, neck and breast are mauve-pink, with blue-grey on top of the head, and a pale pinkish bill. Neck feathers are edged with some pinkish-bronze, black bands, and pale brown margins, producing a scaled effect. Back, rump, and tail are grey, while feet are pink. It has a distinctive call, consisting of two short chucks interspersed between two loud and melodious notes [23]. Its distinctive appearance makes the species easy to identify in the field. However, although much larger, it can be confounded with the White-winged Dove, Zenaida asiatica, especially young individuals.
The species is most often seen in pairs or trios but can form small flocks of about 10-20 individuals and even be part of mixed-species flocks with other columbid species [23]. It feeds upon various seeds and fruits, including cultivated plants [22,24]. Breeding pairs build rudimentary nest platforms made of twigs in trees, bushes, and cacti, where the female lays a single-egg clutch.
2.3. Data Collection
We compiled all records of the Bare-eyed Pigeon on Saint-Martin to track its introduction history and spread on the island, relying on all observations of the species on the island available on e-bird, from 2013 (corresponding to the first reported observation) up to 2024 (eBird Basic Dataset [25]). We checked for double count events, corresponding to two different observers independently reporting the same observation (i.e., same place, same date and same time). As a control, we did the same for the Zenaida Dove, Zenaida aurita, a resident species and one of the most abundant native Columbidae on Saint-Martin. Records from e-bird included our own observations on the two species made during a recent two-day field survey conducted from 5 to 7 July 2023, during which we covered 175 km, mainly on the French part of the island (see supplementary materiels, Table S1) and proceeded to 40 separate 5-min auditory and visual counts to record the presence and abundance of the two species. We also made casual observations of Bare-eyed Pigeons on some occasions during our presence on the island.
2.4. Data Analysis
To analyze variation in the spatial distribution of the Bare-eyed Pigeon through time, we considered three different periods (2015-2019, 2020-2022, and 2023-2024), to have at least 15 observations for each time period. We did not consider the years 2013-2014, as only one observation has been reported in eBird for that period, corresponding to the first observation of the species on Saint Marin (21st January 2013, Observation list ID of eBird : S12697637). We then relied on QGIS 3.36.3 (QGIS.org, [26]) to report all observations on heat maps for both the Bare-eyed Pigeon and the Zenaida Dove. The spatial density of independent observations was represented, for each species and each time period, using heat map function and circles with a radius of 1,000 m around observation locations. We considered this radius size as columbid species similar in size and ecology to the Bare-eyed Pigeon tend to have a good flight ability [27,28]. We further assessed to what extent the Bare-eyed Pigeon’s distribution area on the island increased through time. To that end, we relied on a one-tailed Spearman’s rank-correlation test to assess to what extent the distance between the location of each observation and that of the first observation of the Bare-eyed Pigeon on the island increased through time.
To test for an increase in abundance through time, we classified all observations of Bare-eyed Pigeons and Zenaida Doves in two groups, according to whether a single bird or more than one bird was reported. We then relied on a logistic regression (binomial generalized linear model with a logit link) to assess to what extent the probability of observing more than one Bare-eyed Pigeon or Zenaida Dove in eBird counting events was influenced by the species, the year of observation and their interaction. Because the number of observations differed markedly between the two species, we relied on a Type III analysis of deviance [29,30] and estimated the magnitude of the effects of models parameters using odds ratios and their 95% confidence interval [31,32]. As the interaction was significant (see results), we performed a logistic regression of the proportion of observations with more than one individual as a function of time for each species separately, and tested for the significance of models’ parameters using an analysis of deviance.
We then assessed to what extent observations of the Bare-eyed Pigeon were randomly distributed on Saint-Martin or were associated with particular habitats. To that end, we created a buffer area with a 250 m radius around all observation points. We then overlapped this buffer layer on a land cover raster file created by the European Spatial Agency (ESA WorldCover 10-m 2021 v200; [33]) to count the number of pixels (surface-like estimate) for each category of land cover around all observation points. We first considered all land cover categories characterizing Saint-Martin according to ESA WorldCover 10-m 2021 v200. However, to increase statistical power, we pooled them into four main types of habitats, including Wetland habitats (Permanent water bodies, Mangroves and Herbaceous wetlands), Open habitats (Grasslands, Shrublands and Bare and sparse vegetation), Anthropized habitats (Built-up areas and Croplands), and Tree covered habitats (Tree cover). The number of pixels and their proportion were also estimated for each land cover category at the scale of the entire island. We then relied on a Chi-square goodness-of-fit test to assess to what extent the proportions of the different habitats around observation points differed from what would be expected under the assumption of random habitat selection, and we calculated the Cohen’s W effect size and its bootstrapped confidence intervals (95%; 10,000 iterations) to assess the magnitude of the difference [34]. Finally, we assessed the change in habitat preference over years by reducing the multidimensionality of the land cover dataset around observation points to two major axes, using a Principal Component Analysis. Thereafter, we used Spearman’s rank-correlation tests between contributions of observation events to principal components and time (year of observation) to assess change in habitat preference over time. We further calculated the diversity of habitats around observation points relying on the Shannon index [35], to assess to what extent the diversity of habitats varied over years using a Spearman’s rank-correlation test. All statistical analyses were performed using the R software 4.4.0 [36], with a significance level set at 0.05. When multiple tests were performed (post-hoc tests), we corrected estimated p-values using the Bonferroni’s correction method.
3. Results
From e-bird, we retrieved 108 and 2 043 independent observations of the Bare-eyed Pigeon and the Zenaida Dove, respectively, for the 2013-2024 period. Figure 1 shows the spatial distribution of observations of the two species reported in e-bird for the three considered time periods (i.e., 2015-2019, 2020-2022, and 2023-2024), with the invasive species most often cooccurring with the native species.
According to eBird data, the first observation of Bare-eyed Pigeon on Saint-Martin was made on 21st January 2013, in the eastern part of Saint-Martin, close to the brackish pond known as “Salines d’Orient”. Thereafter, no observation of the species was reported until 4th January 2015. Since that second observation, the number of observations continuously increased (2015-2019 = 19, 2020-2022 = 24, and 2023-2024 = 64), with an evident spatial expansion of the species over time from its supposed point of introduction across the island, first to the north and west, and later to the south (Figure 1, Spearman’s correlation test, rs = 0.355, p < 0.0001). This spatial expansion was accompanied by an increase in the proportion of observations with more than one Bare-eyed Pigeon over time (Table 1AC and Figure 2). In contrast, that probability was stable or tended to slightly decrease for the Zenaida Dove, (Figure 2, Table 1B).
Although all categories of land cover characterizing Saint-Martin were found around observation points of the Bare-eyed Pigeon (Figure 3 and Figure 4), most observations were made along the coastline, corresponding mainly to wetland habitats, anthropized areas and open habitats. Indeed, the spatial distribution of the species differed significantly from random expectation (X² = 9,404.7, df = 3, p < 0.0001 ; Cohen’s W [95% CI]= 0.372 [0.365; 0.378]), with an over-representation of costal habitat (Figure 3).
Principal component analysis reduced variation in land cover to two principal components, explaining respectively 48.89% and 28.66% of the total variation (Figure 4, see in the appendices, Table A1). Larger values of PC1 represented observation points in areas dominated by tree cover and anthropized habitats, while lower values corresponded to observations made in areas dominated by wetland habitat. For PC2, larger values represented observation points made in areas dominated by open habitat, while lower values corresponded to observations of pigeons in areas dominated by anthropized habitats (Figure 4).
The contribution of observation points to PC1 increased significantly through time (Spearman’s correlation test, rs = 0.412, p < 0.0001). In contrast, the contribution of observation points to PC2 was independent of time (rs = -0.004, p = 0.970). This corresponds to the fact that Bare-eyed Pigeons were more likely to be observed in tree-covered and anthropized areas in recent times, whereas initially the species was mainly observed in coastal wetland habitat (Figure 4). In addition, the diversity of habitats around observation points increased significantly through time (rs = 0.204; p = 0.018), evidencing that the Bare-eyed Pigeon diversified its habitat use during its establishment.
4. Discussion
Our results provide the first quantitative assessment of the invasion dynamics of the Bare-eyed Pigeon on Saint-Martin. Although the precise date of introduction of the species on Saint-Martin cannot be firmly established, we can use observations of the Zenaida Dove to estimate a time window. Indeed, our results show that both species tend to occupy the same areas on the island (see Figure 1). In addition, the risk of the two species being confused by birdwatchers is very low, given the obvious morphological differences between them, such that we can reasonably consider that if the invasive species had been present at the same time observations of the native species were made in the past, it would very probably have been detected. We therefore estimate that the introduction of the Bare-eyed Pigeon on Saint-Martin most likely occurred between October 10, 2012 (last eBird survey made in the area of first observation of the Bare-eyed Pigeon) or December 12, 2012 (last observation of the Zenaida Dove reported on eBird for 2012) and January 21, 2013 (date of the first sighting of the Bare-eyed Pigeon reported on eBird).
To the best of our knowledge, there is no information on the origin of the introduction of the Bare-eyed Pigeon to Saint-Martin. The most likely hypothesis is therefore that the animals escaped accidentally or were released following voluntary introduction as pets [37]. Indeed, relatively rare pigeon species are particularly sought after by collectors (e.g. [38]) and illegal trade is considered a major threat for several of them [39]. In addition, bird species caught in the wild and offered for sale on the market have a particularly high invasion success rate [40]. The presence of the species on Aruba, Curaçao and Bonaire [41], three other islands belonging to the Netherlands Antilles, may explain the origin of the introduced individuals. However, captive breeding of wild pigeon species is also practiced illegally in the French West Indies (C. Cambrone pers. obs.). Specifying the origin of the introduction of the Bare-eyed Pigeon to Saint-Martin would be important to ensure that this route of arrival can be neutralized to prevent any further introduction.
Our analysis shows that the Bare-eyed Pigeon population on Saint-Martin has increased in size and has diversified its habitat use since its introduction. Interestingly, the species progressively modified its land use from traditional coastline habitat to anthropized parts of the island, including the urban or suburban habitat, like what has been reported for other expanding Columbidae species [42,43,44,45]. To what extent this increase in numbers and spatial expansion is the consequence of the reproductive activity of a few individuals introduced on one single occasion or is indicative of recurrent introductions since first observation is unclear at the moment. However, although our field survey was of too short duration to investigate the reproduction of the Bare-eyed Pigeon on Saint-Martin, we casually observed and photographed an individual nesting in an Indian jujube tree, Ziziphus mauritiana, on a busy street lined with numerous houses (Figure 5), which corresponds to the main road connecting the French and Dutch parts of the island. The location of this nest was at about 300 m west of the brackish pond “Etang aux poissons” and less than two kilometers southwest of the “Salines d’Orient”, where the first observation of the Bare-eyed Pigeon was made (Figure 1). This is direct evidence of the ability of the species to reproduce, or at least attempt to, on Saint-Martin.
The establishment and rapid spread of the Bare-eyed Pigeon of Saint-Martin deserves particular attention from local and regional authorities and managers. First, although there is so far no evidence for interference competition between the Bare-eyed Pigeon and native columbid species on Saint-Martin, the invasive species may compete with them for access to food and nesting sites in the future. Second, the species tends to be quite common and ubiquitous in its natural distribution area, including around human settlements [41]. Given the recent intensification of urbanization on Saint-Martin [46], the Bare-eyed Pigeon may flourish on the island as tolerance to urbanization is one of the main drivers of invasion success in birds [47], and particularly in Columbidae [42,45]. In addition, the expansion of the Bare-eyed pigeon might not be limited to Saint-Martin if some individuals manage to reach neighbouring islands in the range of their flying ability, such as Anguilla or Saint-Barthélemy distant of about 19 and 30 km, respectively, from Saint-Martin. In addition, hurricanes, which are relatively frequent in the Caribbean, can facilitate their movement over long distances, as already observed for other Caribbean Patagioenas spp.[48]. As the species is well adapted to arid and semi-arid environments [49], its regional expansion might also be further favoured by climate change [5], since climate models predict a significant drying in the insular Caribbean during the 21st century [50].
5. Conclusion
Successful management of invasive species critically depends on early detection and engagement of stakeholders [51,52]. Failure to respond quickly can lead to a situation beyond control, as recently happened on the Caribbean island of Barbados where the introduced Eurasian Collared-Dove, Streptopelia decaocto, has rapidly spread across the island at the beginning of the 21st century to become today the most dominant dove, particularly outcompeting the local Zenaida Dove, and the most numerous bird species in some areas of the island (F. Cézilly, unpublished data).
Therefore, we strongly recommend eradicating the exotic Bare-eyed Pigeon on Saint-Martin before it is too late to do so. Baited closing net bird traps could be used to capture birds on their feeding grounds, as the technique has previously been used efficiently to capture Columbidae species on other Caribbean islands [53,54]. However, this non-selective technique is time consuming, such that, based on previous success with eradicating columbid species on islands [55], shooting might actually be the most efficient removal technique. In that regard, as the Bare-eyed Pigeon is of potential hunting interest, it could be rapidly added to the official list of game species on Saint-Martin, alike the Eurasian Collared-Dove. In particular, hunters should be trained to identify Bare-eyed pigeons without ambiguity and be able to differentiate juveniles of the species from those of the White-winged Dove. We recommend close cooperation between administrative authorities on the French and Dutch parts of the island to coordinate eradication and public awareness campaigns. As the risk of rapid spread and fast population growth is very high, control actions should be initiated as early as possible.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: List of identification codes corresponding to eBird count events performed by authors.
Author Contributions
C.C. was involved in study design, methodology, data collection, statistical analysis, and writing up of the article. F.C. was involved in study design, statistical analysis and writing up of the article. A.L. was involved in data collection. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Fédération départementale des chasseurs de Guadeloupe and the Direction de l'environnement, de l'aménagement et du logement de Guadeloupe.
Data Availability Statement
Raw data are available on eBird.org
Acknowledgments
I also acknowledge Sylvio Plaisant for his advice concerning field survey.
Conflicts of Interest
The authors declare no conflicts of interest
Appendix A

Table A1.
Factor loadings of the four types of land cover on the components 1 (PC 1) and 2 (PC 2). See Figure 4 for PCA plots.
Table A1.
Factor loadings of the four types of land cover on the components 1 (PC 1) and 2 (PC 2). See Figure 4 for PCA plots.
| Habitat | PC1 | PC2 |
|---|---|---|
| Tree cover | 0.657 | -0.277 |
| Open habitat | 0.426 | 0.895 |
| Wetland habitat | -0.994 | -0.104 |
| Anthropized habitat | 0.595 | -0.508 |
| Eigenvalues | 1.956 | 1.147 |
| Cumulative var (%) | 48.893 | 28.663 |
References
- Clavero, M.; Brotons, L.; Pons, P.; Sol, D. Prominent role of invasive species in avian biodiversity loss. Biol. Conserv. 2009, 142, 2043–2049. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Delean, S.; Pyšek, P.; Cassey, P. On the island biogeography of aliens: a global analysis of the richness of plant and bird species on oceanic islands. Glob. Ecol. Biogeogr. 2015, 25, 859–868. [Google Scholar] [CrossRef]
- Bellard, C.; Rysman, J.-F.; Leroy, B.; Claud, C.; Mace, G.M. A global picture of biological invasion threat on islands. Nat. Ecol. Evol. 2017, 1, 1862–1869. [Google Scholar] [CrossRef] [PubMed]
- Soares, F.C.; Panisi, M.; Sampaio, H.; Soares, E.; Santana, A.; Buchanan, G.M.; Leal, A.I.; Palmeirim, J.M.; de Lima, R.F. Land-use intensification promotes non-native species in a tropical island bird assemblage. Anim. Conserv. 2020, 23, 573–584. [Google Scholar] [CrossRef]
- Naimi, B.; Capinha, C.; Ribeiro, J.; Rahbek, C.; Strubbe, D.; Reino, L.; Araújo, M.B. Potential for invasion of traded birds under climate and land-cover change. Glob. Chang. Biol. 2022, 28, 5654–5666. [Google Scholar] [CrossRef] [PubMed]
- Diagne, C.; Leroy, B.; Vaissière, A.-C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.-M.; Bradshaw, C.J.A.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Zenni, R.D.; Essl, F.; García-Berthou, E.; McDermott, S.M. The economic costs of biological invasions around the world. NeoBiota 2021, 67, 1–9. [Google Scholar] [CrossRef]
- Soto, I.; Balzani, P.; Oficialdegui, F.J.; Molinero, C.; Kouba, A.; Ahmed, D.A.; Turbelin, A.J.; Hudgins, E.J.; Bodey, T.W.; Gojery, S.A.; et al. The wild cost of invasive feral animals worldwide. Sci. Total. Environ. 2024, 912, 169281. [Google Scholar] [CrossRef] [PubMed]
- Lenzner, B.; Latombe, G.; Capinha, C.; Bellard, C.; Courchamp, F.; Diagne, C.; Dullinger, S.; Golivets, M.; Irl, S.D.H.; Kühn, I.; et al. What Will the Future Bring for Biological Invasions on Islands? An Expert-Based Assessment. Front. Ecol. Evol. 2020, 8. [Google Scholar] [CrossRef]
- Matos, J.; Little, A.; Broome, K.; Kennedy, E.; Sánchez, F.A.M.; Latofski-Robles, M.; Irvine, R.; Gill, C.; Espinoza, A.; Howald, G.; et al. Connecting Island Communities on a Global Scale: Case Studies in Island Biosecurity. West. North Am. Nat. 2018, 78, 959–972. [Google Scholar] [CrossRef]
- Nowakowski, J.; Dulisz, B. The Red-vented Bulbul Pycnonotus cafer (Linnaeus, 1766) – a new invasive bird species breeding in Europe. BioInvasions Rec. 2019, 8, 947–952. [Google Scholar] [CrossRef]
- Lin, Y.-S.; Liao, J.-R.; Shiao, S.-F.; Ko, C.-C. Origin and Potential Expansion of the Invasive Longan Lanternfly, Pyrops candelaria (Hemiptera: Fulgoridae) in Taiwan. Biology 2021, 10, 678. [Google Scholar] [CrossRef] [PubMed]
- Behm, J.; Busala, G.; Helmus, M. First records of three new lizard species and a range expansion of a fourth lizard species introduced to Aruba. BioInvasions Rec. 2022, 11, 296–306. [Google Scholar] [CrossRef]
- Liu, Y.; Thomas, M.L.; Coupland, G.T.; Wang, P.; Zheng, D.; McKirdy, S.J. Info-gap theory to determine cost-effective eradication of invasive species. Sci. Rep. 2023, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.C.; Meyer, J.-Y.; Holmes, N.D.; Pagad, S. Invasive alien species on islands: impacts, distribution, interactions and management. Environ. Conserv. 2017, 44, 359–370. [Google Scholar] [CrossRef]
- Larson, E.R.; Graham, B.M.; Achury, R.; Coon, J.J.; Daniels, M.K.; Gambrell, D.K.; Jonasen, K.L.; King, G.D.; LaRacuente, N.; Perrin-Stowe, T.I.; et al. From eDNA to citizen science: emerging tools for the early detection of invasive species. Front. Ecol. Environ. 2020, 18, 194–202. [Google Scholar] [CrossRef]
- Dickinson, J.L.; Shirk, J.; Bonter, D.; Bonney, R.; Crain, R.L.; Martin, J.; Phillips, T.; Purcell, K. The current state of citizen science as a tool for ecological research and public engagement. Front. Ecol. Environ. 2012, 10, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Gjerdrum, C.; Loch, J.; Fifield, D.A. The Recent Invasion of Cory's Shearwaters into Atlantic Canada. Northeast. Nat. 2018, 25, 532–544. [Google Scholar] [CrossRef]
- Thibault, M.; Vidal, E.; Potter, M.A.; Dyer, E.; Brescia, F. The red-vented bulbul (Pycnonotus cafer): serious pest or understudied invader? Biol. Invasions 2017, 20, 121–136. [Google Scholar] [CrossRef]
- Arazmi, F.N.; Ismail, N.A.; Daud, U.N.S.; Abidin, K.Z.; Nor, S.M.; Mansor, M.S. Spread of the invasive Javan myna along an urban–suburban gradient in Peninsular Malaysia. Urban Ecosyst. 2022, 25, 1007–1014. [Google Scholar] [CrossRef]
- Fielding, R. Saint Martin/Sint Maarten and Saint Barthélemy. In Landscapes and Landforms of the Lesser Antilles; Allen, C.D., Ed.; Springer: Cham, Switzerland, 2017; pp. 45–59. [Google Scholar]
- Baptista, L.F.; Trail, P.W.; Horblit, H.M.; Boesman, P.F.D. Bare-eyed Pigeon (Patagioenas corensis), Version 1. 0. Birds of the World 2020. [CrossRef]
- Friedmann, H.; Foster, D.S. A Contribution to the Ornithology of Northeastern Venezuela. Proc. United States Natl. Mus. 1950, 100, 411. [Google Scholar] [CrossRef]
- Verea, C.; Aponte, O.; Vargas, G.; Mendoza, A. Primer reporte de daño producido por la Paloma Ala Blanca Patagioenas corensis (Columbiformes: Columbidae) en viñedos del área agrícola de Altagracia, Estado Lara, Venezuela. Memoria de la Fundación La Salle de Ciencias Naturales 2011, 71, 141–145. [Google Scholar]
- eBird Basic Dataset Version: EBD_relApr-2024; Ithaca, New York, 2024.
- QGIS Development Team QGIS Geographic Information System 2024.
- McNair, D.B. Conservation implications of the current breeding distribution and abundance of the White-crowned Pigeon Patagioenas leucocephala at St. Croix, US Virgin Islands. Caribb J Sci 2008, 44, 311–320. [Google Scholar]
- Collins, D.P.; Carleton, S.A.; Coxen, C.L. Movement patterns of adult Interior Band-tailed Pigeons (Patagioenas fasciata) in New Mexico. Wilson J. Ornithol. 2019, 131, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Singmann, H.; Kellen, D. An introduction to mixed models for experimental psychology. In New Methods in Cognitive Psychology; Routledge, 2019; pp. 4–31.
- Hector, A.; Von Felten, S.; Schmid, B. Analysis of variance with unbalanced data: an update for ecology & evolution. J. Anim. Ecol. 2010, 79, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.C.D.; Barry, S.J.E.; Ferguson, H.M.; Müller, P. Power analysis for generalized linear mixed models in ecology and evolution. Methods Ecol. Evol. 2014, 6, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Nagel, R.; Ruxton, G.D.; Morrissey, M.B. Classical tests, linear models and their extensions for the analysis of 2 × 2 contingency tables. Methods Ecol. Evol. 2024, 15, 843–855. [Google Scholar] [CrossRef]
- Zanaga, D.; Van De Kerchove, R.; Daems, D.; De Keersmaecker, W.; Brockmann, C.; Kirches, G.; Wevers, J.; Cartus, O.; Santoro, M.; Fritz, S. ESA WorldCover 10 m 2021 V200 2022.
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Routledge, 2013; ISBN 0203771583.
- Cambrone, C.; Bezault, E.; Cezilly, F. Efficiency of the call-broadcast method for detecting two Caribbean-endemic columbid game species. Eur. J. Wildl. Res. 2021, 67, 1–18. [Google Scholar] [CrossRef]
- R Core Team R: A language and environment for statistical computing 2024.
- Lockwood, J.L.; Welbourne, D.J.; Romagosa, C.M.; Cassey, P.; Mandrak, N.E.; Strecker, A.; Leung, B.; Stringham, O.C.; Udell, B.; Episcopio-Sturgeon, D.J.; et al. When pets become pests: the role of the exotic pet trade in producing invasive vertebrate animals. Front. Ecol. Environ. 2019, 17, 323–330. [Google Scholar] [CrossRef]
- Bruslund, S.; Leupen, B.; Shepherd, C.R.; Nelson, S.S. Online trade as a serious additional threat to the Critically Endangered silvery pigeon Columba argentina in Indonesia. Nat. Conserv. 2022, 46, 41–48. [Google Scholar] [CrossRef]
- IUCN SSC IUCN SSC Pigeon and Dove Specialist Group: Position Statement on the Value of Ex Situ Populations in Conserving Threatened Columbiformes and Recommendations on Enhanced Use of CITES to Address Unsustainable Trade. Available at: www.iucn.org/SSC; 2023.
- Carrete, M.; Tella, J. Wild-bird trade and exotic invasions: a new link of conservation concern? Front. Ecol. Environ. 2008, 6, 207–211. [Google Scholar] [CrossRef]
- Prins, T.; Reuter, J.; Debrot, A.; Wattel, J.; Nijman, V. Checklist of the Birds of Aruba, Curaçao and Bonaire, South Caribbean. Ardea 2009, 97, 137–268. [Google Scholar] [CrossRef]
- Mulhall, S.; Lill, A. What Facilitates urban colonisation by Crested pigeons Ochyphaps lophotes? Corella 2011, 35, 73–81. [Google Scholar]
- Veech, J.A.; Small, M.F.; Baccus, J.T. The effect of habitat on the range expansion of a native and an introduced bird species. J. Biogeogr. 2010, 38, 69–77. [Google Scholar] [CrossRef]
- Bendjoudi, D.; Voisin, J.-F.; Doumandji, S.; Merabet, A.; Benyounes, N.; Chenchouni, H. Rapid increase in numbers and change of land-use in two expanding Columbidae species (Columba palumbus and Streptopelia decaocto) in Algeria. Avian Res. 2015, 6, 18. [Google Scholar] [CrossRef]
- Luna, A.; Romero-Vidal, P.; Hiraldo, F.; Tella, J. Cities favour the recent establishment and current spread of the Eurasian collared dove Streptopelia decaocto (Frivaldszky, 1838) in Dominican Republic. BioInvasions Rec. 2018, 7, 95–99. [Google Scholar] [CrossRef]
- Pasquon, K.; Jouannic, G.; Gargani, J.; Minh, C.T.D.; Crozier, D. Évolution de l’urbanisme et exposition au risque cyclonique à Saint-Martin de 1954 à 2017. VertigO-la revue électronique en sciences de l’environnement 2022, 22.
- González-Lagos, C.; Cardador, L.; Sol, D. Invasion success and tolerance to urbanization in birds. Ecography 2021, 44, 1642–1652. [Google Scholar] [CrossRef]
- Wiley, J.W.; Wunderle, J.M. The effects of hurricanes on birds, with special reference to Caribbean islands. Bird Conserv. Int. 1993, 3, 319–349. [Google Scholar] [CrossRef]
- Gibbs, D.; Barnes, E.; Cox, J. Pigeons and Doves: A Guide to the Pigeons and Doves of the World; A&C Black, 2001; Vol. 13; ISBN 1873403607.
- Herrera, D.A.; Ault, T.R.; Carrillo, C.M.; Fasullo, J.T.; Li, X.; Evans, C.P.; Alessi, M.J.; Mahowald, N.M. Dynamical Characteristics of Drought in the Caribbean from Observations and Simulations. J. Clim. 2020, 33, 10773–10797. [Google Scholar] [CrossRef]
- Kluever, B.; Avery, M.; Gawlik, D.; Hall, P.; Humphrey, J.; Pernas, T.; Ridgley, F. Eradication of African sacred ibis (Threskiornis aethiopicus) from South Florida, USA: a collaborative early detection and rapid response case study. Manag. Biol. Invasions 2023, 14, 123–132. [Google Scholar] [CrossRef]
- Mazzamuto, M.V.; Wauters, L.A.; Koprowski, J.L. Exotic Pet Trade as a Cause of Biological Invasions: The Case of Tree Squirrels of the Genus Callosciurus. Biology 2021, 10, 1046. [Google Scholar] [CrossRef] [PubMed]
- Cézilly, F.; Quinard, A.; Motreuil, S.; Pradel, R. Adult survival selection in relation to multilocus heterozygosity and body size in a tropical bird species, the Zenaida dove, Zenaida aurita. Oecologia 2015, 180, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Cambrone, C.; Cézilly, F.; Wattier, R.; Eraud, C.; Bezault, E. Levels of genetic differentiation and gene flow between four populations of the Scaly-naped Pigeon, Patagioenas squamosa: implications for conservation. Stud. Neotropical Fauna Environ. 2021, 57, 1–13. [Google Scholar] [CrossRef]
- Phillips, R.B.; Cooke, B.D.; Carrión, V.; Snell, H.L. Eradication of rock pigeons, Columba livia, from the Galápagos Islands. Biol. Conserv. 2012, 147, 264–269. [Google Scholar] [CrossRef]
Figure 1.
Heat maps showing spatial distribution changes of the Bare-eyed Pigeon (left) and the Zenaida Dove (right) over three time periods. The top map is a satellite view of Saint-Martin (Sources: ESRI) showing the first record of the Bare-eyed Pigeon on the 21st January 2013, indicated by a red arrow. The yellow arrow indicated the location of the nest observed (Figure 5) on the 7th July 2023, at 03:30 pm.
Figure 1.
Heat maps showing spatial distribution changes of the Bare-eyed Pigeon (left) and the Zenaida Dove (right) over three time periods. The top map is a satellite view of Saint-Martin (Sources: ESRI) showing the first record of the Bare-eyed Pigeon on the 21st January 2013, indicated by a red arrow. The yellow arrow indicated the location of the nest observed (Figure 5) on the 7th July 2023, at 03:30 pm.
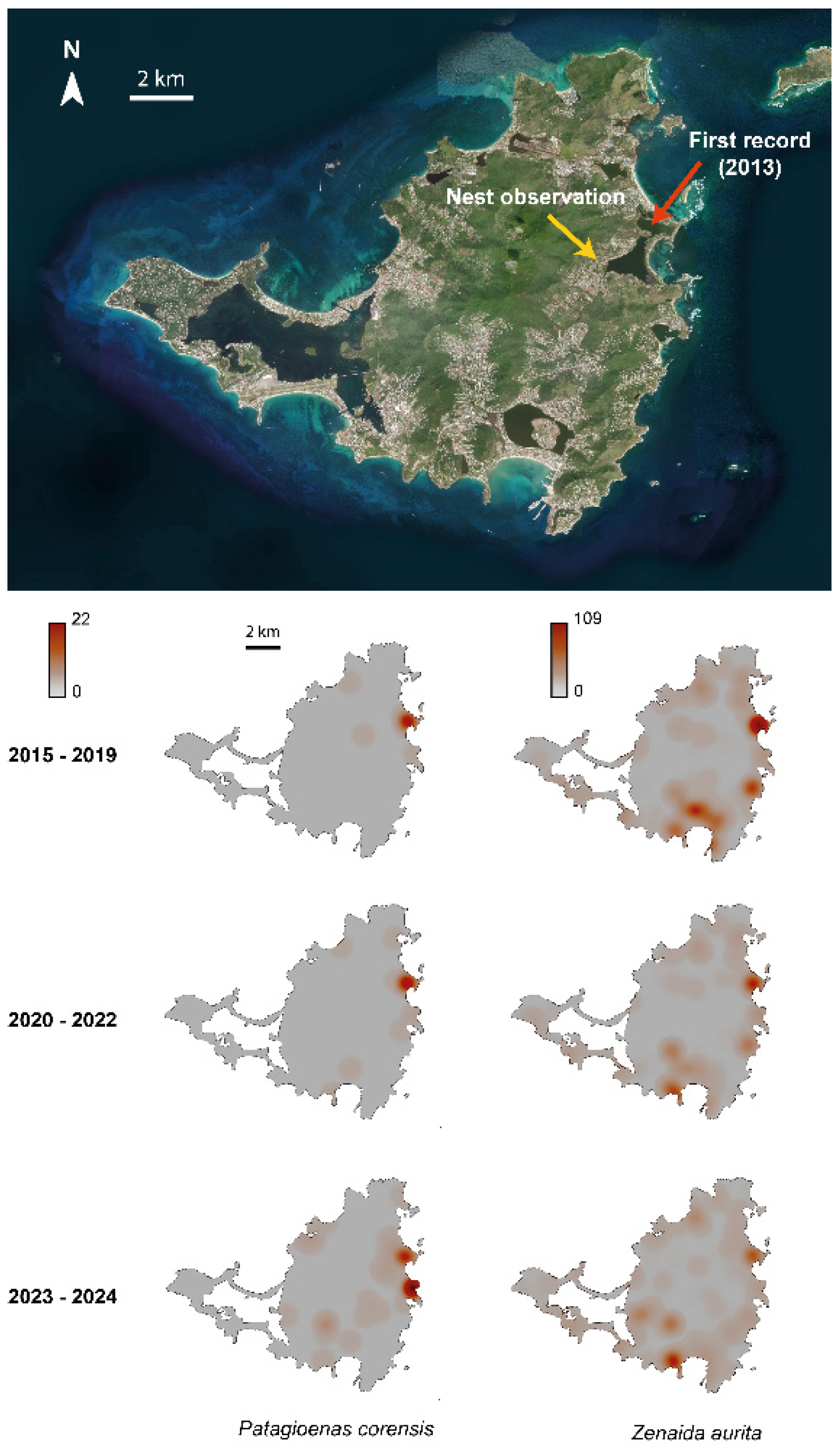
Figure 2.
Variation in the probability of detecting more than one individual on each observation reported in eBird for Patagioenas corensis and Zenaida aurita between 2013 and 2024. The lines represent the predictive trends for each species according to model A (Table 1), with 95% confidence intervals indicated by grey ribbons around the lines.
Figure 2.
Variation in the probability of detecting more than one individual on each observation reported in eBird for Patagioenas corensis and Zenaida aurita between 2013 and 2024. The lines represent the predictive trends for each species according to model A (Table 1), with 95% confidence intervals indicated by grey ribbons around the lines.
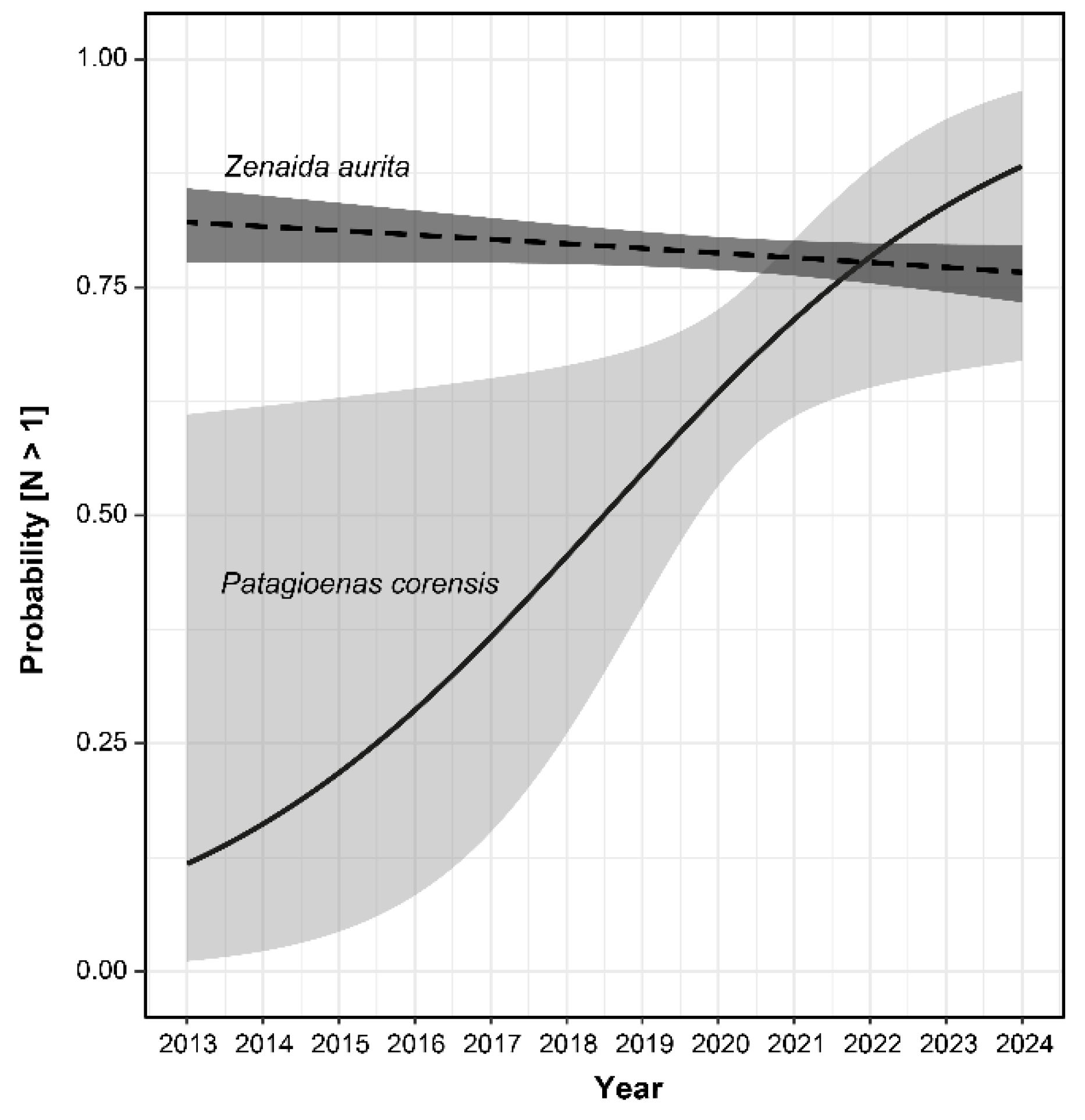
Figure 3.
Proportion of habitat types around observation points of the Bare-eyed Pigeon reported in eBird.org and for the entire island of Saint-Martin.
Figure 3.
Proportion of habitat types around observation points of the Bare-eyed Pigeon reported in eBird.org and for the entire island of Saint-Martin.
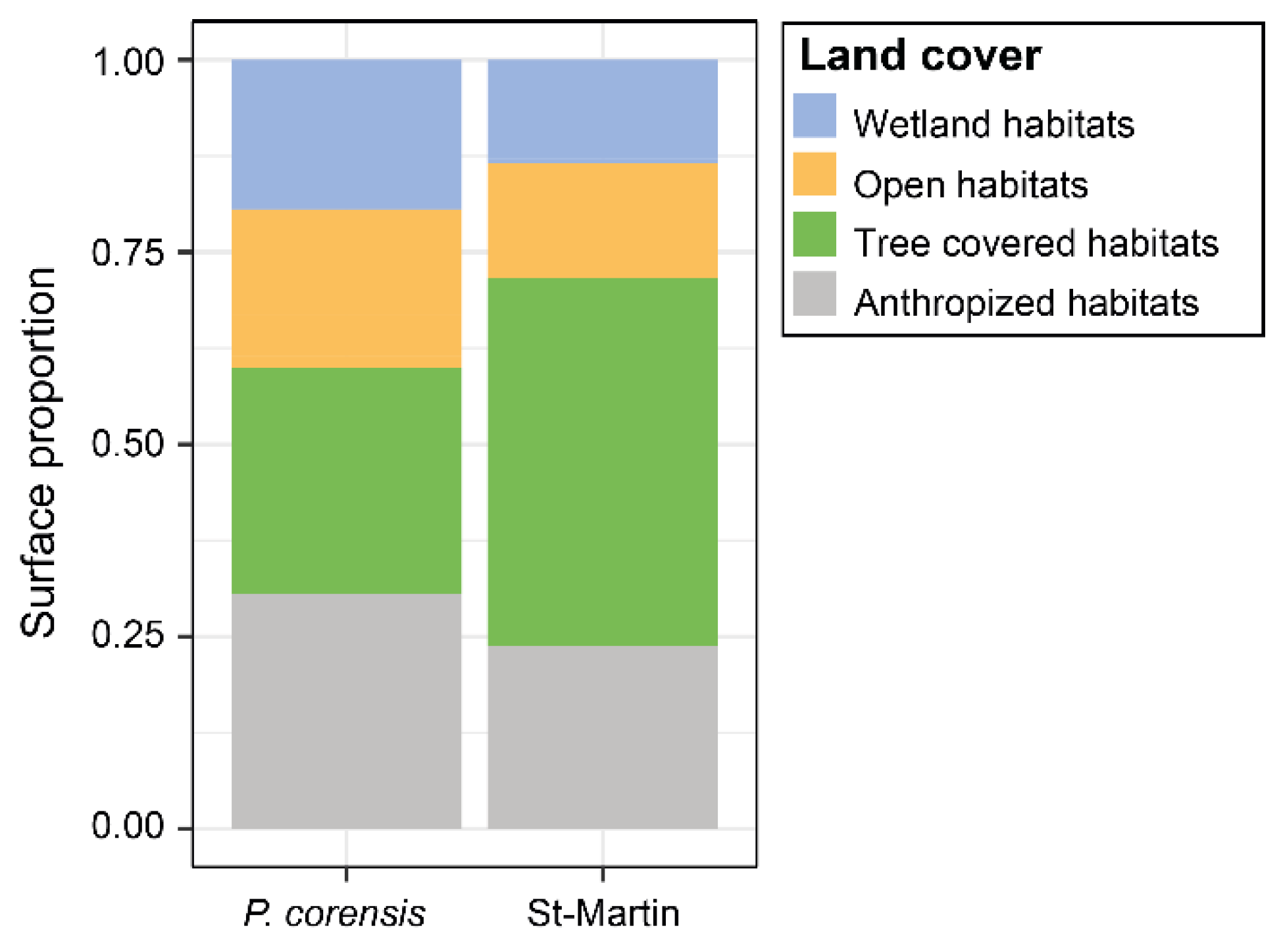
Figure 4.
Plots showing ACP results. Plot (a) displays the contributions of different of the four land cover types to the first two principal components (PC1 and PC2), explaining 48.89% (PC1) and 28.66% (PC2) of the total variance. See in the appendices, Table A1, for factor loadings of the four types of land cover on the PC1 and PC2. The color gradient represents the magnitude of contribution from each land cover type, with red indicating higher contributions and green indicating lower contributions. Plot (b) shows the distribution of eBird observation along PC1 and PC2 for different time periods. Ellipses give a visual indication of the spread and central tendency for each time period, with a confidence level at 95%.
Figure 4.
Plots showing ACP results. Plot (a) displays the contributions of different of the four land cover types to the first two principal components (PC1 and PC2), explaining 48.89% (PC1) and 28.66% (PC2) of the total variance. See in the appendices, Table A1, for factor loadings of the four types of land cover on the PC1 and PC2. The color gradient represents the magnitude of contribution from each land cover type, with red indicating higher contributions and green indicating lower contributions. Plot (b) shows the distribution of eBird observation along PC1 and PC2 for different time periods. Ellipses give a visual indication of the spread and central tendency for each time period, with a confidence level at 95%.
Figure 5.
Photography of a Bare-eyed Pigeon nesting in an Indian jujube tree, Ziziphus mauritiana, on a very lively street (18.073410, -63.031927).
Figure 5.
Photography of a Bare-eyed Pigeon nesting in an Indian jujube tree, Ziziphus mauritiana, on a very lively street (18.073410, -63.031927).

Table 1.
Significance of model parameters for three logistic regressions. The model (A) analyses how the probability of detecting more than one individual (Y) varies according to the year, the species, and their interaction. The model (B) and (C) analyses how the probability of detecting more than one Zenaida Dove and Bare-eyed Pigeon varies according to the year, respectively.
Table 1.
Significance of model parameters for three logistic regressions. The model (A) analyses how the probability of detecting more than one individual (Y) varies according to the year, the species, and their interaction. The model (B) and (C) analyses how the probability of detecting more than one Zenaida Dove and Bare-eyed Pigeon varies according to the year, respectively.
| Model | Parameter | Odds Ratio [95% CI] | X² | p |
|---|---|---|---|---|
| (A) Y ~Year + Species + Year x Species | Year | 1.183 [1.016; 1.432] | 4.742 | 0.029 |
| Species | 2.148 [1.388; 3.323] | 6.817 | 0.009 | |
| Year x Species | 1.219 [1.047; 1.477] | 6.801 | 0.009 | |
| (B) Y ~Year (Zenaida aurita) | Year | 0.970 [0.935; 1.005] | 2.832 | 0.092 |
| (C) Y ~Year (Patagioenas corensis) | Year | 1.442 [1.066; 2.112] | 5.821 | 0.016 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



