
Submitted:
12 July 2024
Posted:
12 July 2024
You are already at the latest version
Abstract
The nuclear hormone family of receptors regulates gene expression. The androgen receptor (AR) shuttles upon ligand binding and homodimerization from the cytosol into the nucleus to activate gene expression. Thyroid hormone receptors(TR), retinoic acid receptors (RAR), and the vitamin D receptor (VDR) are present in the nucleus bound to chromatin as a heterodimer with the retinoid X receptors (RXR) and repress gene expression. Ligand binding leads to transcription activation. The hormonal ligands for these receptors play crucial roles to ensure the proper conduct of very many tissues and exert effects on prostate cancer (PCa) cells. Androgens support PCa proliferation and deprivation alone or with chemotherapy is the standard therapy for PCa. RAR activation and 3,5,3'-triiodo-L-thyronine (T3) stimulation of TR support the growth of PCa cells. Ligand stimulation of VDR drives growth arrest, differentiation, and apoptosis of PCa cells. Often these receptors are explored as separate avenues to find treatments for PCa and other cancers. However, there is accumulating evidence to support receptor interactions and crosstalk of regulatory events whereby a better understanding might lead to new combinatorial treatments.
Keywords:
Prostate cancer
; nuclear hormone receptors
; androgen receptor
; thyroid hormone receptor
; retinoic acid receptor
; vitamin D receptor
1. Introduction
Prostate cancer (PCa) is the second most common tumor in males. Hormone sensitivity is a trait Achilles heel of PCa whereby androgen deprivation therapy (ADT), which began in 1941, represents the core strategy of PCa treatment [1]. This treatment alone or with chemotherapy remains the standard therapy, but many patients progress to ADT-resistant tumors termed castration-resistant PCa (CRPC) which is mainly associated with amplifications, mutations and gene rearrangements to the androgen receptor (AR) [2,3]. The potentiation of AR in the CRPC setting historically constituted the rationale for combining ADT with drugs that either target androgen synthesis (abiraterone) or block the AR (enzalutamide and bicalutamide), leading to a beneficial clinical outcome [4,5,6]. Other steroidal hormone receptors, including the progesterone receptor [7], estrogen receptors (ERs) [8] and glucocorticoid receptor [9], have been implicated in PCa tumorigenesis and resistance development.
For PCa, we examine the influence of androgens, 3,5,3’-triiodo-L-thyronine (T3) stimulation of the thyroid hormone receptor (TR) β the use of synthetic retinoids to selectively antagonise the retinoic acid receptor (RAR) γ and activation of the vitamin D receptor (VDR) by 1α,25-dihydroxyvitamin D3 (1,25D3), the active metabolite of vitamin D3. We focus on the activation status of these receptors because they influence the proliferation of PCa cells whereby activity of AR, TRβ, and RARγ enhance PCa proliferation and active VDR drives growth arrest followed by apoptosis. We describe the current state of knowledge regarding the modes of action of the hormones and their receptors, particularly their interactions and the extent to which there is also regulatory crosstalk. A better appreciation of crosstalk is likely to offer new therapeutic possibilities, especially for hormone-dependent forms of cancer [10]. Our aim is to highlight the potential of targeting the above nuclear receptors (NRs) to treat PCa. There are extensive reviews of the roles of other NRs in PCa [11] including orphan nuclear receptors (ONR) [12].
2. Nuclear Hormone Receptors
2.1. The Functionalities
Hormones exert continuous control on gene regulation processes to ensure the proper functioning of all tissues and, therefore, the well-being of an organism. A hormonal imbalance due to an excess or a deficiency can lead to severe and sometimes life-threatening diseases. From studies of estrogen, it became clear that hormones act through NRs to regulate gene expression [13,14]. In 1985, the first full-length NR to be cloned was the human glucocorticoid receptor [15] and around the same time the estrogen receptor (ER) α was cloned [16]. Soon it became clear that these receptors are structurally similar and belong to a NR superfamily [10,17].
In addition to NR family members binding classical hormones, they also bind lipid-soluble ligands, including the most active metabilite of vitamin A all-trans retinoic acid (ATRA), 1,25D3, and oxysterols. Binding activates the essential role of NRs as transcription factors (TFs). Many NRs and their isoforms have been cloned and many of their regulatory cofactors have been identified. Presently, there are 48 genes for NRs and closely related orphan NRs (ONRs), and their phylogenetic classification is based on protein sequence homologies [12,14]. They all contain a highly conserved modular structure, and the evolutionary oldest family members function without ligands. Whilst much is known about NRs and their modes of action, information about their tissue- and disease-specific effects and their binding cistromes still needs to be discovered. Other poorly understood areas include the extent to which NRs interact and their potential mutual regulation.
2.2. Phylogenetical Features
NRs contain four main structural elements. They are the N-terminal domain (NTD), a gene transactivation domain which includes the DNA-binding domain (DBD), a flexible hinge region (H) which contains the nuclear localization motif (NLS), and a ligand-binding domain (LBD) [14,18,19]. NRs function as TFs in an allosteric manner, typically switching between an inactive (ligand-free) and an active (ligand-bound) form. An important feature of NRs is that they can act as monomers, homodimers, and heterodimers with other NRs [18,20]. Figure 1 showns how gene expression is driven by type I and type II NRs by canonical dimers.
The NRs are broadly divided into four types based on their functional characteristics and modes of DNA binding [20]. Type 1 are located in the cytosol and shuttle upon ligand-binding and homodimerization into the nucleus. They are steroid hormone-binding receptors and include AR, the glucocorticoid receptor (GR), the progesterone receptor (PR), the mineralocorticoid receptor (MR), and ERs. Type II NRs include the TRs, RARs, VDR, and peroxisome proliferator-activated receptors (PPARs). They are present in the nucleus and localised to chromatin as RXR heterodimers in a repressive mode. They become transcriptionally active when ligands are bound, which leads to the loss of corepressors and the recruitment of coactivators. Type III are the homodimeric orphan receptors, they shuttle as dimers, and bind to direct hormone response element (HRE) repeats. Type IV are the monomeric orphan receptors [18]. The four types of NRs have been considered as a putative target regarding therapies for PCa [21]. Proteomic studies have revealed the capacity of NRs to liaize with one another and various cofactors [22], creating a wide range of NR interactome possibilities. Crystal structure analyses have revealed non-canonical dimerisation and even multimeric forms [19], highlighting a high functional versatility. Figure 2 shows the structures of the AR, TR, RAR, and VDR genes and proteins.
2.3. DNA-Binding Motifs and Gene Expression Regulation
The NR regions relevant for DNA binding and heterodimerization were identified by inserting specific mutations, leading to the elucidation of the nucleotide nature of the DNA-binding motifs and characterisation of HREs [23]. Similar to other TFs, how NRs interact with chromatin depends on simple, highly conserved hexamer DNA-motifs. Ingeniously, their orientation and spacer regions lead to the fine tuning of binding capabilities [17,22,24]. Importantly regarding DNA response element binding, TR, VDR, and RAR bind as heterodimers with retinoid X receptors (RXRs), including pairing with RXRα, β or γ According to the 3-4-5 rule, these are always direct AGGTCA hexamer repeats, spaced by 3 nucleotides for the VDR, 4 nucleotides for TRs, and 5 nucleotides for RARs. There are other binding possibilities, including additions to the known “canonical” motifs. One site sequence would not be sufficient because a single transcription factor can recognise hundreds of DNA sequences, bind within a range of affinities, and local DNA structure is also essential [25]. All these parameters greatly expand the potential scope of gene expression regulation [26]. To add to this complexity, there are isoform variants of NRs and their multitude interactions, ligand- and DNA-binding capabilities result in regulatory possibilities that are still barely defined [27]. Finally, NRs regulate distinct genes in different tissues, and most probably, they are different in diseases due to pathophysiological changes.
3. Androgen Receptor
Androgen deprivation therapy is the gold-standard treatment for most PCa patients. However, CRPC, which is a highly aggressive metastatic hormone-independent form of the disease, develops in a significant number of patients. Treatment options for these patients are limited, even with the recent introduction of improved therapeutics.
3.1. The Functionalities of the AR
The AR gene is located at q11-12 on the X chromosome and encodes a 920 amino acid protein. Alternative splicing results in multiple transcript variants encoding different isoforms. As for all NRs, there are four main structural elements to the protein (Figure 2). Together with the GR), PR, and MR receptors, AR forms the oxosteroid superfamily, which is phylogenetically separated from the ERs and TRs [28]. The AR is a classical steroid hormone receptor whereby functionality is critically dependent on the presence of androgens. They are a group of steroidal sex hormones that are produced in the testes, the ovaries, and the adrenal glands and androgens are the main drivers of male sex development. Testosterone, the major androgen, is converted intracellularly to dihydrotestosterone (DHT), which has the highest affinity for the AR. Post-ligand binding, the AR forms homodimers which then translocate to the nucleus [29] to control gene expression by binding to inverted repeat androgen response elements (AREs) [24].
3.2. AR and Prostate Cancer
The role of AR in PCa is described exhaustively in numerous reviews [30,31]. The AR maintains a healthy prostate epithelium, including its proper differentiation. Overactivation alone does not lead to PCa [11], and instead the AR plays a role in conjunction with other malignant processes. In this case, correlating AR expression levels with the risk of PCa development is not significant.
AR contributes to tumour growth by driving the expression of genes that control cell proliferation. Whilst AR-mediated changes to PCa-associated gene expression are known, their contributions to PCa and dissemination still need to be fully understood. There are changes in extranuclear steroid hormone receptor signalling [32], and the AR, after shuttling to the nucleus, shows altered DNA binding capabilities, which reshapes the AR-driven cistrome. Since the canonical AR binding motif and its structure are highly conserved, this must be due to chromatin landscape changes and its accessibility, as was shown for the cadherin 1 gene [33].
During advanced malignant transformation, the role of the AR becomes increasingly essential. ADT drives the development of active AR variants without ligand binding. After an initial successful ADT treatment phase, PCa becomes “androgen-independent”, but remains dependent on active AR signaling, which is conferred by various mechanisms that include AR point mutations, gene amplifications, activating AR splice variants, truncated AR variants, and upregulation of AR co-activators [34]. Point mutations accumulated at the dimerization interface, which probably reflect higher stability of the resulting monomers [19], and AR splice variants activate distinct transcriptional programs [35]. Very late-stage PCa is characterized by complete loss of the AR. This fatal development has been strongly advanced by the use of highly effective AR inhibitors and degraders. Hence, treatment options that circumvent the development of this situation are urgently needed.
3.3. Crosstalk between AR and Other NRs/Transcription Factors
There are potential non-canonical AR interactions that have implications for PCa. Recent crystal structure data indicate a high flexibility of the core AR dimers which might form a head-to-tail or a head-to-head arrangement [36]. This non-canonical dimeric conformation theoretically opens up many more spatial possibilities for potential interactions with other NRs or the cofactors that associate to form the transcriptionally active AR complex [19]. Atypical dimers are considered very likely to exist [27], though they are less prevalent than the canonical ones. Moreover, when AR function is strongly suppressed during ADT, atypical dimers can become essential and might even overtake AR function. There are likely direct consequences regarding physical NR interactions/competitions at the level of DNA [27] and to the regulation of subsets of genes. Indirect modes of pathway interference might include perturbations to the expression of the other NRs and/or their ligands [27]. All these possibilities might substitute for reduced AR function during ADT or might drive PCa growth per se.
There is a crosstalk between the AR and other NRs. RAR was upregulated post-androgen treatment of LNCaP cells; this was controlled by an ARE promoter element and led to upregulation of the epithelial growth factor receptor [37]. Upregulation of the glucocorticoid receptor was observed as an effect of ADT in PCa [38,39]. Though PCa is primarily an androgen-dependent disease, there is the interplay between AR and ERs because androgens are aromatised to estrogens in adipose tissue, and the testosterone/estradiol ratio is essential to the development of PCa [40] (Figure 2). Low teststerone levels and high levels of estradiol lead to premalignant lesions providing evidence to support the importance of an imbalance to the ratio [41], and a high estradiol level in African-American men has been linked to a greater risk of developing PCa [42]. More recent work has focused on how estrogenic imprinting of the epigenome may contribute to PCa [43] and it has been argued that new compunds that target ERs might lead to therapeutic opportunities [44]. AR interacts with the PR regarding expression of the kallikrein-related peptidase 4 (KLK4). This protein is only expressed in PCa when there is expression of the AR and progesterone receptors. A hormone response element mediates gene expression whereby the progesterone receptor interacts directly with the gene promotor, and the interaction of AR is indirect [7].
There is cooperativity between AR signalling and PPARγ, whereby AR inhibits the expression of PPARγ (Figure 3). At first, PPARγ was viewed as a tumour suppressor for PCa, agonists were shown to inhibit PCa cell growth, but this was then found to be PPARγ independent. Recently, it has been reported that there is increased expression of PPARγ during PCa progression with higher protein levels in advanced PCa compared to low-risk disease and benign hyperplasia [45]. Findings support the view that PPARγ supports PCa cell growth by increasing the machinery for lipid synthesis, including cutaneous fatty acid-binding protein (C-FABP) [46], fatty acid synthase (FASN) and ATP citrate lyase (ACLY), and mitochondrial biosynthesis [47], bringing to attention that NRs regulate PCa metabolism. Upon loss of AR, antagonism of PPARγ may benefit the treatment of advanced stages of PCa. However, there is the need to take into consideration complexities relating to the different splice variants of PPARγ whereby increased PPARγ1 increased PCa tumorigenesis and FASN and ACLY expression whereas overexpression of PPARγ2 decreased LNCaP and PC3 proliferation and invasiveness [47].
Interaction of AR with tumor promoting signaling pathways has been observed, importantly with the Wnt pathway in CRPC [48]. Wnts are an ancient and conserved family of secreted glycoproteins that regulate the fate of developing cells [49]. Downstream of the binding of Wnts to frizzled is the activation of the TF β-catenin [50], which plays a role in various cancers including PCa [51,52]. Recently, a direct interaction of AR with GATA3 has been shown to regulate a luminal epithelial phenotype in breast cancer [53].
4. Thyroid Hormone Receptor
4.1. The Functionalities of TRβ
Thyroid hormone (TH) signaling is a primeval pathway. Though iodine is toxic to pathogens, including viruses, anaerobic bacteria, yeast and fungi, it is a crucial micronutrient [54]. It is taken up by single-cell species, such as photosynthetic bacteria and algae, as iodate which is converted to reactive iodide anions that have a high affinity for tyrosine [54]. Iodide coupling creates the highly reactive iodotyrosine signaling molecule. Virtually all organisms use iodine in this manner for use in essential biochemical tasks. The synthesis of THs evolved from iodotyrosines, which are present in all animal genera except arthropods [54,55]. THs regulate the growth and development in non-bilaterian animals, such as porifera (sponges) and cnidarians (medusae and polyps), and are present throughout lophotrochozoans (molluscs and worms) onwards [55]. Lophotrochozoans have a TH signaling system that is TR-mediated. Essentially, THs are important universally to the conduct of normal development. They provide a pro-survival stimulus and support metamorphosis in amphibians and lampreys and life-transitions in higher animals [56], and their effects are highly time- and dose-specific and depend on tissue type and developmental stage [57].
THs exert their effect in vetebrates via TRs. The TR-coding cDNA was identified as a homologue to the avian erythroblastosis virus v-erb-A oncogene [58].TR proteins are much shorter than the AR; TRα has 490 amino acids, and TRβ has 461amino acids. However, a vast variety of isoforms is generated by alternative splicing. TRs are stimulated by thyroxine (T4) and its active derivative T3. They form homodimers or act as heterodimers in conjunction with RXRs [59]. There are two main TR isoforms: TRα and TRβ. TRα is predominantly expressed in the heart, bone, and brain, whereas TRβ is more abundant in the liver, kidney and thyroid. TRα and TRβ seem to exert distinct tissue-dependent effects which are very likely complementary to each other, as revealed from studies of mutants [60]. Moreover, TRα and TRβ have opposing roles by differences in their molecular properties and modes of action [61], based on diverse DNA binding affinities [62].
Unlike the AR, TRs belong to the class II NRs and reside constantly in the nucleus as a heterodimer with RXR. The canonical TR pathway involves binding of TR/RXR to thyroid response elements (TREs), which are, as mentioned above, direct repeats of two hexamer DNA-motifs spaced by 4 nucleotides (DR4) [24]. This binding mode is highly fine tuned by the more specific DNA motifs [63]. Shuttling of T3 to the nucleus and binding to TRs attached to TRE elements in the genome leads to the loss of corepressors and the subsequent activation of gene transcription [64,65].
Further extensive analyses of TR DNA binding and potential ways of gene expression regulation have led to indications that there is gene regulation beyond canonical binding modes. TRs can bind to DNA as monomers [66], and dimeric variants have been described [19,67]. TRβ is the more interesting TR and the gene that encodes it is located at chromosome 3. It seems to act in a more versatile manner, and several missense mutations are reported, which, as for the AR, map to the homodimer interface and might lead to altered ligand or coregulator binding.
4.2. THs/TRβ and Prostate Cancer
THs contribute to vertebrate development with links to many endocrine systems, as seen from studies of zebrafish, birds, rodents and dogs [68,69,70,71,72]. There are indications that a close interaction between thyroid and gonadal hormone systems controls sex-determination and testicular development [73]. In particular, THs impinge on sexual differentiation and gonadal development in mammals: they are involved in the maturation and functioning of the female [74] and male reproductive systems [71,75], including the stimulation of androgen release in the testis [76]. In men, THs regulate testicular development and function [75]. Thyroid disorders are associated with gonadal dysfunction, hypogonadism and reduced prostate weight [77,78,79].
THs have long been suspected to be involved in the development of malignancies [80], whereby the maintenance of a hypothyroxinemia was suggested to improve the survival of cancer patients [81]. Whether THs, including T3 and T4, play a role in supporting PCa growth, progression, and metastasis has remained largely unexplored. Albeit, THs influence the development and physiology of the human prostate [82] and the incidence of PCa, as seen from studies using a diet-induced regimen [83]. Epidemiological studies have linked low plasma T3 levels with a low incidence of PCa [84,85] and high T4 levels are significantly associated with an increased risk of any solid cancer, particularly PCa, lung, and breast cancer [84,86]. The latter finding led to the hypothesis that high TH levels correlate with tumor progression [87,88,89,90]. Several case reports have reported a correlation between low TH levels and slower cancer growth [64,91,92,93]. Studies of the availability of T3 for binding to TRβ support a role for T3 and TRβ in PCa. μ-Crystallin (CRYM) sequesters T3 in the cytosol to prevent T3 binding to nuclear TRβ [94], thereby controlling downstream target activation [88]. Our study showed that CRYM and TRβ show reciprocal expression in PCa tissue, whereby low CRYM represents a feature of metastatic PCa [95]. We and others have demonstrated that CRYM expression is deficient in hormone-refractory PCa patients [96,97] indicating reciprocal roles for CRYM and TRβ [98]
The literature is more extensive regarding the specific role of TRβ, perhaps reflecting a more significant role in cancer. TRβ is mainly described as a tumor suppressor [28] because expression is often reduced in human tumors due to deletions and epigenetic modifications [99]. A reduction has been linked to a poor prognosis in hepatocellular, renal, thyroid and breast cancers [100,101,102]. TRβ expression is lost upon chromosomal 3p deletions, especially in breast and liver cancers [64]. In addition to somatic loss-of-function mutations that foster thyroid cancers, there is a high incidence of de novo inactivating TRα and TRβ mutations in hepatocellular, renal, and thyroid cancers [99] which may relate to selection pressures during tumor progression [103]. Loss of TRβ dysregulates several growth control pathways, leading to the exaggerated growth of various cancers [104].
In contrast to a tumor suppressor role for TRβ, TRα/TRβ1 double knockout mice develop fewer skin tumors [105], and enhanced TRβ expression has been detected in colon and head and neck cancers [106,107]. A recent study showed that low cytosolic and enhanced nuclear TRβ levels are indicators of a poor outcome in breast cancer [108], highlighting the importance of distinguishing between cytosolic and nuclear TRβ abundance, as recognised previously from studies of HeLa liver cancer cells [109]. Intriguingly, RXRs seem to have an anti-oncogenic role in PCa [110], providing tentative support to a tumor-promoting action of activated TRβ.
Literature regarding the influence of THs on cancer often presents a view of the clinical implications/morbidity relating to hypothyroidism [84,111,112]. THs stimulate the growth of many cancers, including breast, ovarian, pancreatic, hepatocellular, and renal cancer cells [113,114,115,116]. From in vitro studies, T3 supported the growth of PCa, lung, breast, ovarian and squamous cell cancer cells [64,88,117], and THs have a direct stimulatory effect on some key oncogenic signaling pathways, including the phosphatidylinositol-3-kinase (PI3K)- and extracellular signal-regulated kinase (ERK1/2)-mediated pathways. In vitro studies using the androgen-sensitive LNCaP cells revealed that nmolar T3 stimulated cell growth and the expression of prostate specific antigen [118]. Longterm incubation led to an increased androgen binding capacity in the nucleus of LNCaP cells, suggesting that T3 might induce AR expression ([119,120]. It is known that T3 and androgens act cooperatively to affect prostate specific antigen (PSA) expression [95,118]. However, whether there is a direct interaction between AR and TRβ is unknown.
T3/TRβ driven gene regulation has been analyzed in hepatocellular cancer to identify the proteins involved in tumor progression. In particular, T3-mediated upregulation of expression of the protease furin was seen for hepatoma cell lines, and the investigators concluded that this might enhance tumor metastasis [115,121,122,123]. TRβ has been shown to regulate cell metabolism and tumor-relevant genes. They include the hypoxia-inducible factor 1 subunit α [124], the CD44 stemness factor in the brain [125], cathepsin H (CTSH gene) which is a tumor invasive factor in HepG2 [123], and extracellular matrix proteins ([115]. TRβ has been shown to effect mitochondrial respiration directly [126], which probably contributes to senescence, DNA damage and oxidative stress.
Regarding in vivo studies, upregulation of mRNAs for TRβ together with other NRs and ONRs was observed for a PCa cell xenograft model [127]. Administering 6-n-propyl-2-thiouracil (PTU), a thyreostatic agent, reduced the growth of DU145 and PC-3 xenografts in mice [117] and reduced their growth in hypothyroid mice although enhanced aggressive behaviour of hepatocellular and breast cancer cell xenografts was observed [99].
4.3. Crosstalk between TRβ and Other NRs/Transcription Factors
There is evidence to support the view that there is cross-regulation between THs and the androgen axis and that this occurs in cancer [128] (Figure 3). Exposure of the testes of the Western clawed frog (Silurana tropicalis) to T3 led to increased DHT production, and a low level of androgens in the serum of PCa patients correlated with decreased T3/T4 levels [129]. An androgen/TH interplay was recently observed in the PCa microenvironment to exert a cooperative tumor-promoting effect [130]. However, the above hormonal cross-regulation raises the question of whether the downstream effector NRs – in this case, TRβ and AR – control the same or overlapping gene sets and processes [27]. TRβ/AR crosstalk was shown in silico by detailed promoter analysis of TH- and androgen-dependent genes, which revealed binding sites for both receptors (TRE and ARE sites) in the promoters of AR- and TR-regulated genes, indicating mutual or collaborative gene regulation [131]. Accordingly, TRβ-mediated gene expression is stimulated by both T3 and DHT robustly [132]. Other workers showed that THs increased the expression of AR and androgen synthesis enzymes [133]. These findings were corroborated by a recent transcription factor binding site study that detected an overlap of NRs binding to the same regulatory elements in PCa [134], opening the possibility of a mutual or collaborative TR/AR DNA-binding mode.
There is evidence to support the view that TRs interact with RARs. In 1993, Rosen and colleagues characterized a homologous 20-amino acid region conserved in TRβ, RAR and VDR essential for heterodimerization [135]. They postulated the existence of various heterodimeric combinations to allow for enhanced variability and sensitivity of gene expression regulation. Other early studies also claimed that TRs and RAR form heterodimers, supported by overexpression and subsequent cell-free DNA-binding (EMSAs) and luciferase reporter assays in vitro [59,136,137,138]. Where RARs form heterodimers with TR, they have affinities for consensus and natural HREs like those for TR/RXR heterodimers. Therefore, TR/RAR heterodimers can regulate T3-mediated gene expression [138]. For amphibian limb blastemal and COS transfected cells, ATRA was observed to mediate both RAR- and T3-mediated effects [139]. Stromelysin 3, a vital tissue remodeling protease, was found to be regulated by THs and ATRA due to the presence of both TREs and RAREs in the gene promoter [140]. For rat GH3 pituitary gland cells, ATRA has been shown to antagonize T3 action, presumably at a receptor level [141]. In addition to this complex hormonal crosstalk, T3 stimulation has been shown to regulate the expression of the ATRA synthesizing enzymes within mouse brain cells ([142], underscoring the importance of hormonal crosstalk.
An important recent finding regarding PCa relates to the nuclear corepressor 2/silencing mediator (NCOR2) for RAR and TR. The expression of this corepressor is frequently altered in PCa and other cancers [143,144]. Reduced expression of NCOR2 has been shown to accelerate the failure of androgen deprivation. The investigators used the CWR22 xenograft model to show that a stable reduction of expression of NCOR2 accelerated the recurrence of disease post-ADT [[145].
In parallel to the early TR/RAR interaction studies, Schräder and colleagues showed that VDR might directly interact with TRβ, and that polarity directs gene expression [146,147]. Other workers identified interference by 1,25D3 and ATRA regarding TH regulation of glucocorticoid hormone expression, concluding that VDR-TR and VDR-RAR heterodimers act as competitors of TR-RXR and RAR-RXR complexes [148]. In contrast, others have failed to provide evidence for VDR-TR heterodimerization but showed that both VDR and TR compete for RXR binding [149], still highlighting a crosstalk between both receptors. Support to this view by a study that showed that VDR heterodimerizes with RXR, but not with TRβ [150], and that a delicate balance of ligand availability and the limiting amounts of RXR, which would shift between TR and VDR, would affect the repression versus activation of various genes. Beyond the above findings, reports regarding crosstalk between TR and VDR have been limited. However, a very interesting crosstalk was described for adipocyte [151] whereby TRβ expression was controlled by 1,25D3, and vice versa, and that T3 regulates VDR gene expression in mouse photoreceptors has been reported [152].
There is a crosstalk between TRβ and other NRs. TRβ interacts with ER isoforms, resulting in flexible regulation of the consensus estrogen response element [153], and TRβ interaction with ERα has been shown to regulate mitochondrial activity ([154]. TRβ interacts with PPARγ [155] and is then oncogenic [156]. Crosstalk between TRβ and liver X receptor (LXR) has been identified for lipid metabolism-related genes and other physiological systems such as the central nervous system [157]. Regarding other TFs, TRβ interaction with the Wnt pathway is important based on direct interaction between TRβ and β catenin [158]. Overall, TRβ interaction with the Wnt pathway for several organisms and tissues indicates the promotion of a stem cell phenotype [159]. Other crosstalk includes regarding jun and fos oncogene activities, based on the abilities of TRs to inhibit AP-1 binding to DNA [160]. TRβ mediates repression of STAT5 activity [161], and overactivation of STAT5 signaling has been linked to an oncogenic TRβ variant in breast cancer [162].
Diverse findings bring to attention the complex role that TRβ plays in regulating the physiological status of cells. TRβ interacts with intracellular signalling pathways and with PI3K to play a role in the maturation of mouse hippocampal synapses [163]. There is TH regulation of steroid hormone-associated genes, for example, regulation of the expression of kidney androgen-regulated protein in the developing kidney ([164], and T3 regulation of the expression of androgen receptor-associate protein 70 (ARA70) and sex hormone binding globulin (SHBG) in HepG2 cells [122,165]. TRβ modulates the function of the tumor suppressor p53, leading to differential regulation of p53-regulated genes [153]. Early studies revealed a connection between TR-mediated gene expression and changes to the expression of cell cycle regulators [156,166]. Overexpression of TRβ in thyroid cancer cells led to activation of the RhoB signaling pathway and p21-induced cell-cycle arrest [167].
5. Retinoic Acid Receptors
5.1. The Functionalities of RARs
The three distinct RARs, namely RARα, RARβ, and RARγ are encoded by different genes [168]. In addition, each RAR has isoforms, due to the presence of different promoters, alternative splicing, and use of non-AUG start codons [169,170,171,172]. RARγ is of particular interest regarding a potential new avenue for treating PCa (see later). The central portion of the RARγ protein, which contains the DNA and ligand binding domains, is encoded by seven axons. There are separate N-terminal exons for γ1 and γ2 and a major portion of the gene encodes several exons for untranslated regions of γ1 [173].
ATRA is the natural ligand of RARs and is used to control the transcription of target genes [174,175]. RARs form various heterodimers with RXRα, RXRβ, RXRγ or homodimers and the dimers bind to specific retinoic acid response elements (RAREs) within the promoters of genes. Without ATRA binding to RARs, RAR is associated with RAR/RXR dimers to suppress gene expression due to RAR recruitment of transcription corepressors. ATRA binding to RAR leads to gene expression via the release of corepressors, the recruitment of coactivators [176], and the binding of further factors that include histone acetylase or histone methyltransferases, histone demethylases, and DNA-dependent ATPases [177,178].
A crucial consideration of RARs is whether the main isoforms have distinct roles or whether there is functional redundancy. RARs are required for embryonic development, and double knockouts in mice were needed to observe aberrant phenotypes associated with vitamin A deficiency. In this case, other RARs can perform developmental functions in the absence of a particular RAR [179,180,181]. In contrast, the different patterns of expression of main isoforms and developmental changes within embryo and adult tissues support the view that each has a specific role to normal physiology. RARα expression is ubiquitous within embryonic and adult tissues. In contrast, the expression patterns of RARβ and RARγ within the developing embryo are complex and either tissue-specific or regionalised [182,183]. RARγ mRNA expression within embryonic tissues suggested a role in the development of the central nervous system, and early studies indicated a role for RARβ in the differentiation of adult stratified squamous epithelia [184]. At late stages of zebrafish embryo development, RARγ mRNA is highly restricted to primitive cells, including mesodermal and neural crest stem and progenitor cells, in the head area, in the lateral plate mesoderm, and in the pre-somitic mesoderm of the tail bud [185]. These findings suggested a role for RARγ in early morphogenesis, as also concluded from early findings for mouse embryogenesis [186]. Changes to the levels of expression of isotypes are seen during haematopoiesis. Haematopoietic stem cells and their differentiating offspring express RARα with RARα2 increasing dramatically during myeloid cell differentiation of FDCP mixA4 mouse progenitor cells [187]. In contrast, haematopoietic stem cells and primitive progenitors selectively express RARγ [188].
The distinct roles of RARα and RARγ during haematopoiesis regulate the production of mature blood cells alongside the need to maintain stem cells. Active RARα facilitates cell differentiation whereby ATRA activation of RARα promoted neutrophil differentiation of the human promyeloid cell lines HL60 and NB4 [189,190] and normal myeloid progenitors [191]. These findings led to ATRA differentiation therapy for acute promyelocytic leukaemia, with ATRA together with arsenic trioxide providing a cure for this once fatal disease [192]. Even so, RARα is dispensable to myelopoiesis and instead modulates [193] presumably by influencing the frequency of cell decision-making by myeloid progenitor cells to favour differentiation. In contrast, RARγ is needed for the maintenance/self-renewal of haematopoietic stem cells because numbers were markedly reduced in knockout mice [188]. In keeping with this role, treatment of zebrafish embryos with a RARγ agonist in the absence of exogenous ATRA blocked bone and ganglion development from cranial neural crest stem cells and fin development from mesodermal stem cells and the stem cell populations were maintained. The block of fin development was reversible by washout of the agonist or the subsequent addition of a RARγ antagonist [194].
Findings from molecular studies have supported the view that RARγ controls stem cell behaviour, perhaps by imposing stemness and/or by restricting the propensity of stem cells to differentiate including the temporal and/or spatial onset. RAR/RXR dimers were observed to bind to gene loci within undifferentiated F9 embryonal cells that bind the pluripotency-affiliated transcription factors SOX2, NANOG, and POU5f1 [176]. For embryonic stem cells, RARγ was required for chromatin epigenetic marks and transcription activation [195] and epigenetic reorganisation of the Hoxa and Hoxb gene structures [196]. RAR paralog that is expressed by the annelid worm (Platyneris dumarilli) is interesting in terms of its role in signalling. ATRA transactives the paralog but it binds with a low affinity using a different pocket. Therefore, ancestral RAR is likely to act as a permissive sensor and a suggested role is triggering the spatially restricted onset of neurogenesis. High-affinity RARs have evolved at the base of chordates with an instructive role in regulating the Hox gene cluster, with high affinity being important to sensing complex ATRA gradients [197].
5.2. RARs and Prostate Cancer
ATRA differentiation therapy for acute promyelocytic leukaemia has not extended to PCa nor other cancers [198]. Albeit and from immunohistology studies, increased expression of RARα and RARγ has been reported within high-grade PCa [199]. The LNCaP, PC-3, and DU145 prostate cancer cell lines express RARα and RARγ with LNCaP cells additionally expressing RARβ [200]. However, these cell lines are relatively insensitive to ATRA with the induction of growth arrest and apoptosis requiring a pharmacological amount (2 μM) [200,201].
Whether RARβ plays a role in PCa is uncertain. An association between the methylation status of RARβ2 and PCa risk has been reported [202] and a meta-analysis concluded that RARβ promotor methylation may be a correlate of PCa carcinogenesis. Further studies are needed to demonstrate a prognostic value because different methylation rates have been reported for PCa tissue [203]. That RARβ may play a role in controlling the proliferation of PCa has been argued from the finding that stable transfection of RARβ into the RARβ-negative PC-3 cells led to increased sensitivity to the combined use of a RARβγ-selective agonist (SR11262) and a potent vitamin D3 analogue regarding the inhibition of clonal growth. Increased sensitivity was proportional to the level of RARβ expressed [204].
RARγ expression is of prime importance to regulating the growth of PCa cells for the following reasons. PCa cells seem to have evolved to survive in a low ATRA environment. ATRA levels for patients’ cells were close to the limit of detection (~ 1 ng/g tissue), whereas levels were up to eight times higher in surrounding normal tissue and benign prostate hyperplasia [205]. This is important because nM concentrations transactivated RARγ and 100-fold more was needed to activate RARαα [190,206]. The proliferation of the PCa cell lines LNCaP, DU 145, and PC-3 was stimulated by a level of ATRA that was sufficient to activate RARγ but not RARα (10-11-10-9 M), and treatment with 10-10 M ATRA increased colony formation and the percentage of stem cell-like colonies. The RARγ agonist AGN205327 exerted the same effect as low doses of ATRA [207]. Therefore, the proliferation of PCa cells is highly dependent on the activity of RARγ. Findings from knockdown and knockout studies have provided further support to the view that RARγ promotes cancer cell proliferation, for example, from studies of colorectal and pancreatic cancer cells [208,209].
The activity of RARγ is also important to the survival of PCa cells. Treatment of flask cultures of patients’ cells and the LNCaP, PC-3, and DU145 cell lines with the selective RARγ antagonist AGN205728 was highly effective in driving growth arrest in G1 of cell cycle followed by necroptosis. A concentration of 5 nM AGN205728, close to its ED50 for RARγ of 3 nM, prevented colony formation by the cancer stem cell (CSC)-like cells of the PCa cell lines. AGN205728, therefore, targeted both CSC and non-CSC [207]. Normal prostate epithelial cells and non-neoplastic RWPE-1 cells were significantly less sensitive to the action of AGN205728 than patients’ PCa cells. The use of AGN194310 to antagonise all RARs was as effective as the RARγ selective antagonist against patents’ cells and the PCa cell lines, and normal prostate epithelium cells were less sensitive [200,210,211]. Taken together, the findings support the development of an antagonist of RARγ for use to treat PCa.
RARγ is also an oncogene for various carcinomas [212]. Overexpression related to increased cell proliferation, rapid disease progression, and a poor prognosis has been reported for human colorectal cancer, cholangiocarcinoma, hepatocellular cancer, ovarian cancer, pancreatic ductal adenocarcinoma, renal cell cancer, and high-grade PCa. In keeping, RARγ is an oncogene whose expression is downregulated by miR30a-5p [213] and this tumour suppressor miR is commonly at a low level in cancer cells. Acacetin targeting of the non-genomic actions of RARγ was effective against human hepatocellular cancer cell lines [214] and the specific aldehyde dehydrogenase inhibitors 673A, DIMATE, DEAB, NCT-501, silybin, and solomargine, which interfere with the endogenous synthesis of ATRA, were effective against lung, ovarian, prostate, and uterine cancer cells [212].
From all the above, RARγ regulates the behaviour of multipotent stem/ progenitor cells, whereby agonism blocked the onset of their development. That RARγ is an oncogene and that overexpression within cancer cells correlated with a rapid disease progression and poor prognosis fits well with RARγ, ensuring the stemness of cancer stem cells. These cells are primarily responsible for both aggressive and disease relapse, and their frequency within a tumour varies from very few to up to 27% for melanoma [215]. This significant difference reflects the different types of cancer studied. Still, it may also reflect how well the method used to evaluate cancer stem cells within a tumor had efficiently measured their frequency as argued from studies of myeloma [215]. It is noteworthy that antagonising RARγ killed PCa stem cells and all their offspring within flask-cultured cells, whereby offspring seem to have retained expression of RARγ. Antagonising RARγ did not kill haematopoietic stem cells [190]. Like RARα, the action of RARγ is modulatory rather than obligatory to developmental processes. RARγ2 null mice were normal, and the foetuses of mice null for all RARγ isotypes were visibly normal. The latter mice exhibited growth deficiency, early lethality, and interestingly, prostate squamous metaplasia [216]. Moreover, no adverse effects were seen when mice and rats were given substantial doses of a pan-RAR antagonist (BMS-18945) other than the inhibition of spermatogenesis, which was reversible [217,218]. The use of RAR antagonists seems to be safe in an adult organism. Why the cancer stem cells died by necroptosis when treated with very low doses of the RARγ and pan-RA antagonists is surprising and yet to be resolved.
5.3. Crosstalk between RARs and Other Transcription Factors
In addition to RARs competing with other nuclear receptor superfamily members for binding to RXRs and dimerisation with TR (see above), they crosstalk with AR. RARγ impacts androgen signalling because its expression level influences AR activity. ChIP-seq experiments in PCa cell lines showed that RARγ was bound to active chromatin and significantly overlapped with AR binding. Knockdown of RARγ in LNCaP cells affected the expression of genes associated with the AR response. Further experiments, using the HPr1-AR human prostate epithelial cell line showed that RARγ knockdown resulted in a substantial reduction of the transcriptional response to DHT treatment with known AR target genes affected [219]. RARs crosstalk in other ways as seen for ATRA and membrane receptor-provoked events. In 1990, investigators reported a negative cross-modulation between RARs and the activator protein 1, which regulates gene expression in response to cytokines [220].
β-catenin is particularly interesting regarding crosstalk of RARγ with other TFs. RARγ physically interacts with β-catenin, as shown by co-immunoprecipitation studies. Overexpression in chondrocytes strongly inhibited β-catenin signalling, whereas silencing (RNA-mediated) of endogenous RARγ strongly increased signalling. The investigators proposed that unliganded RARγ would associate with β-catenin to inhibit its signalling [221]. Decreases in β-catenin target gene expression may be relevant to the ability of the RARγ antagonist to kill cancer cells. The interaction between RARγ and β-catenin is more complex because Wnt/β-catenin signalling regulates the expression of Yes-associated protein (YAP) as shown for colon cancer cells [222]. YAP is a coactivator of RARγ and acted via RAREs to reinforce stem cell traits within HT-29 colon cancer cells, including self-renewal. Conversely, the use of the pan-RAR antagonist BMS493 to silence RAR signalling downregulated the stem cell traits of HT29 and 5F31 cells, including their renewal capacity [223]. As seen for RARγ, YAP is an oncogene for several cancers, showing elevated expression in bladder, cervical, colon, gastric, non-small cell lung, oesophageal, and ovarian cancers [224]. In keeping with agonism of RARγ blocking stem cell development, YAP expression in human neural in vitro systems negatively correlated with neuronal differentiation and was seen to promote a neural rest/multipotent phenotype [225].
The role of VDR, like that of RARα, is to promote cell differentiation, and there is crosstalk between the actions of RARα and VDR. Studies of acute myeloid leukaemia cell lines revealed that RARα regulated the expression of a VDR transcriptional variant originating in exon 1a. Un-ligated RARα repressed expression of the VDR gene whereby a high basal level of expression of RARα and a lack of RARα agonism correlated with repressed expression of the VDR gene. Downregulation of the level of RARα expression led to increased VDR gene expression. [226]. Whilst RAR and VDR preferentially bind DNA as heterodimers with RXR, a very early, and a perhaps ignored finding was that VDR-RAR and VDR-RXR heterodimers can act functionally on direct repeat, palindrome, and inverted palindrome response elements [227].
6. Vitamin D Receptor
6.1. The Functionalities of VDRs
The two basic forms of vitamin D are D2 (ergocalciferol) and the main form D3 (cholecalciferol). Vitamin D is not a vitamin because vitamin D3 is made in human skin from 5,7-dehydrocholesterol via a non-enzymatic transformation and upon skin exposed to UV-B radiation [228]. Both forms of vitamin D are not biologically active, requiring conversion to the active forms 1α,25-dihydroxyvitamin D2 and 1,25D3, which have a systemic action like classic steroid hormones.
The composition of the VDR genomic region is very complex and covers ~100 kb located on chromosome 12 [229]. There are 14 exons with multiple enhancers regulating the tissue-specific expression of VDR proteins, with translation spanning from exons 2 to 9 [230,231]. In some individuals, there is a T to C polymorphism, which eliminates the most 5’-located ATG codon in exon 2, and translation is from the second in-frame ATG codon. This means that there are two variants of VDR with lengths of 424 and 427 amino acids [232]. Transcription regulation is complex for the VDR gene region that contains the six exons 1a to 1f because various tissues make alternative use of the exons and their corresponding promoters [230]. Three promoter regions have been identified for exons 1a - 1f; exons 1a and 1d are regulated by the promoter upstream to exon 1a, and exons 1f and 1c have their upstream promoters. How the remaining exons are regulated remains to be elucidated [230,231,232].
There is widespread expression of VDR within tissues. Most of the tissues responsive to 1,25D3 express transcripts originating from exon 1a and 1d. Transcripts that start from exon 1d give rise to a longer VDR protein, named VDR B1 [230,233]. VDR plays a vital role in the endocrine control of calcium-phosphate homeostasis [[234]. A transcript that originates in exon 1f is selectively expressed by tissues that play a role in calcium-phosphate homeostasis [230].
Here, we focus on the capacity of 1,25D3 to drive growth arrest, differentiation, and apoptosis of cancer cells. In this regard, VDR binds to thousands of genomic loci to modulate the expression of hundreds of target genes. VDR, after binding of 1,25D3, forms heterodimers with RXRα, RXRβ, and RXRγ to drive gene expression by modifying chromatin structure. The heterodimers bind to vitamin D response elements (VDREs) within the promotors of target genes (VDRE). VDR also forms homodimers that interact with VDREs. Like RARs, VDR is silent in the absence of ligand due to the recruitment of corepressors. When a ligand is bound, gene expression occurs due to the recruitment of coactivators and eventually RNA polymerase II [235,236]. Secondary/indirect 1,25D target genes are regulated by transcription factors that are encoded by the primary target genes.
6.2. VDR and Prostate Cancer
For over four decades, various studies have considered using 1,25D3 to treat multiple cancers [237]. Laboratory studies showed that 1,25D3 drives the growth arrest and apoptosis of a range of cancer cells, particularly PCa, breast cancer, and leukaemia cells. However, the use of vitamin Ds to treat patients, in general, has been greatly hindered by their hypercalcaemic action because increased blood calcium can lead to soft tissue calcification, increased bone resorption, and can even be fatal [238]. Substantial concentrations of 1,25D3 are often used in in vitro and preclinical studies to achieve growth arrest and apoptosis of cancer cells. A recent appraisal of the literature has brought to attention that the positive outcomes from vitamin D clinical studies do not match up with those from in vitro and preclinical studies, whereby reaching an effective dosage in the clinic is likely to be an issue [239].
From mouse model studies of PCa, the growth of cells and metastasis were faster in mice that were fed a vitamin D3 deficient diet as compared to mice that were given 10,000 IU vitamin D3. Deprivation-mediated acceleration of growth and metastasis was attributed to epithelial to mesenchymal transition [240]. A concern regarding administrating vitamin D3 to increase 1,25D3 levels, or 1,25D3, is that 1,25D3 induces the expression of the 24-hydroxylase CYP24A1 which in turn readily catabilizes 1,25D3. Hence, in a PC-3 prostate xenograft model mice were given 1,25D3 and a CYP24A1 inhibitor which resulted in enhanced inhibition of tumor cell growth [241].
A consensus view is that 1,25D3 is able to drive growth arrest and differentiation followed by apoptosis of many different cancer cell lines. Studies of prostate cancer cell lines show that various anticancer actions have been attributed to 1,25D3. For LNCaP cells, 1,25D3 enhanced apoptosis [242], up- and down- regulated the expression of many genes [243], and inhibited AR signalling [244]. 1,25D3-mediated reported effects on LNCaP, PC-3, and DU145 cells have included decreased invasiveness by the selective modulation of proteases [245] and disruption of glucose metabolism and the tricarboxylic acid cycle [246]. For LNCaP, the non-malignant prostate epithelium cells RWPE-1, and RMPE-2 cells (from RWPE-1 by ki-Ras transformation), an effect of 1,25D3 was to upregulate tumour suppressor miRNAs [247]. Additional 1,25D3 effects on RWPE-1 cells included an antioxidative action and the regulation of pathways such as Wnt, Notch, and NFκB1 [[248, 249]. 1,25D inhibited Wnt activity for patient-derived benign prostate epithelial organoids [250].
There has been a substantial effort to synthesise analogues of 1,25D3 that are more potent than the parent hormone and with a much-reduced calcaemic action [251,252]. In particular, the PRI-1906 analogue has antiproliferative activity against PC-3 PCa cells [253]. As mentioned above, there has been a focus of attention on breast cancer and the analogues PRI-1906, PRI-5201, and PRI-5202 were antiproliferative against human breast cancer cell lines [254,255]. PRI-5105, PRI-5106, and PRI-5202 were effective against human colon cancer cell lines [254,256].
1,25D3 does have effects on PCa, breast, and colon cancer cells, but of fundamental importance is whether any benefit might seen from its use to treat patients. A prime consideration regarding PCa is that the aggressive cell lines are somewhat insensitive to the action of 1,25D3. It has been suggested that this unresponsiveness relates to an elevated nuclear expression of corepressors. In this case, 1,25D3 responsiveness is suppressed by a mechanism involving histone deacetylation, which might be overcome by using histone deacetylase inhibitors [257]. Even so, to date, 1,25D3 and their analogues have not provided an effective adjunct to chemotherapy for prostate cancer regarding a reduction of mortality in clinical trials. Though well-guided trials of vitamin D compounds are limited, there is limited data to support their use to treat or prevent PCa [258]. Whether newer 1,25D3 analogues that are much more potent than the parent hormone and with a negligible calcaemic action can be developed and then used to provide some benefit as an adjunct to chemotherapeutics for PCa and other cancers remains to be seen.
Regarding an interest in the use of vitamin D to prevent PCa, an association has been reported from a meta-analysis between circulating 25-hydroxyvitamin D levels (a measure of vitamin D status) and a reduced risk of PCa [259]. However, this association remains unproven because of the variability of findings from both retrospective and prospective studies [260]. Different outcomes may be related to different levels of vitamin D supplementation. Even so, 12,927 persons received daily 2000 IU vitamin D3 and 12,944 persons received a placebo in the large VITAL trial and there was not a significant reduction in the incidence of all cancers nor of PCa [261,262].
8. Concluding Remarks
The search for effective treatment options for advanced PCa, especially after the cessation of the effectiveness of hormonal therapy, has been ongoing for several decades. Management options have become scarce, the prognosis is poor, and further investigations are needed to overcome advanced disease. NRs are a promising class of signaling-effector targets regarding finding new treatments. Epidemiological and experimental findings affirm that NR signalling contributes to PCa pathogenesis and promotes progression. However, a deeper understanding of the molecular actions of NRs is required to define new diagnostic/prognostic and therapeutic targets. This includes the need to extend information regarding whether and how AR-, TRβ-, RARγ-, and VDR-mediated pathways converge in PCa to co-regulate the expression of critical genes, for example, AR and the proteases kallikrein-related peptidase 3 and transmembrane-serine protease 2. On a mechanistic level, there is a need for a better understanding regarding whether AR, TRβ, RARγ-, and VDR undergo hitherto unknown interactions when having when bound their ligands or relating to the recruitment of cofactors. The emergence of new and compelling omics technologies (ChIPseq, CUT&RUN, CUT&TAG, qPLEX-RIME) enables transcription factor complexes to be determined and the regulated cistromes to be analyzed and compared. From rationales developed from such studies, using combinations of agents that target the actions of NRs may provide new therapeutic approaches to treating androgen-dependent and independent PCa, opening paths to treatments beyond androgen blockers. Drugs that interfere with hormones and RARγ and that activate VDR are well established and might be repurposed for PCa treatment.
Author Contributions
G.B., B.H.; writing—original draft preparation, LK, M.H., and V.S.; writing—review and editing.
Funding
This work received funding from Horizon—the Framework Programme for Research and Innovation (2022–2027), Marie Sklodowska-Curie Actions, of the European Union under Research Executive Agency grant agreement eRaDicate No. 101119427. G.B., M.H., and V.S. are funded by UK Research and Innovation (UKRI) under the UK government’s Horizon Europe funding guarantee EP/Y030818/1 and are associate partners to eRaDicate.
Conflicts of Interest
The authors declare that there is no conflict of interest.
References
- Huggins, C.; Hodges, C.V. Studies on Prostatic Cancer I. The Effect of Castration, of Estrogen and of Androgen Injection on Serum Phosphatases in Metastatic Carcinoma of the Prostate. Cancer Res 1941, 1, 293–297. [Google Scholar]
- Watson, P.A.; Chen, Y.F.; Balbas, M.D.; Wongvipat, J.; Socci, N.D.; Viale, A.; Kim, K.; Sawyers, C.L. Constitutively active androgen receptor splice variants expressed in castration-resistant prostate cancer require full-length androgen receptor. Proc Natl Acad Sci U S A 2010, 107, 16759–16765. [Google Scholar] [CrossRef] [PubMed]
- Haile, S.; Sadar, M.D. Androgen receptor and its splice variants in prostate cancer. Cell Mol Life Sci 2011, 68, 3971–3981. [Google Scholar] [CrossRef]
- James, N.D.; de Bono, J.S.; Spears, M.R.; Clarke, N.W.; Mason, M.D.; Dearnaley, D.P.; Ritchie, A.W.S.; Amos, C.L.; Gilson, C.; Jones, R.J.; et al. Abiraterone for Prostate Cancer Not Previously Treated with Hormone Therapy. N Engl J Med 2017, 377, 338–351. [Google Scholar] [CrossRef]
- Fizazi, K.; Tran, N.; Fein, L.; Matsubara, N.; Rodriguez-Antolin, A.; Alekseev, B.Y.; Ozguroglu, M.; Ye, D.; Feyerabend, S.; Protheroe, A.; et al. Abiraterone plus Prednisone in Metastatic, Castration-Sensitive Prostate Cancer. N Engl J Med 2017, 377, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Vaishampayan, U.N. Randomized trial of enzalutamide versus bicalutamide in combination with androgen deprivation in metastatic hormone sensitive prostate cancer: A Prostate Cancer Clinical Trials Consortium trial. J Clin Oncol 2018, 36, 190. [Google Scholar] [CrossRef]
- Lai, J.; Myers, S.A.; Lawrence, M.G.; Odorico, D.M.; Clements, J.A. Direct progesterone receptor and indirect androgen receptor interactions with the kallikrein-related peptidase 4 gene promoter in breast and prostate cancer. Mol Cancer Res 2009, 7, 129–141. [Google Scholar] [CrossRef]
- Setlur, S.R.; Mertz, K.D.; Hoshida, Y.; Demichelis, F.; Lupien, M.; Perner, S.; Sboner, A.; Pawitan, Y.; Andren, O.; Johnson, L.A.; et al. Estrogen-dependent signaling in a molecularly distinct subclass of aggressive prostate cancer. J Natl Cancer Inst 2008, 100, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Puhr, M.; Hoefer, J.; Eigentler, A.; Ploner, C.; Handle, F.; Schaefer, G.; Kroon, J.; Leo, A.; Heidegger, I.; Eder, I.; et al. The Glucocorticoid Receptor Is a Key Player for Prostate Cancer Cell Survival and a Target for Improved Antiandrogen Therapy. Clin Cancer Res 2018, 24, 927–938. [Google Scholar] [CrossRef]
- Zhao, L.; Zhou, S.; Gustafsson, J.A. Nuclear Receptors: Recent Drug Discovery for Cancer Therapies. Endocr Rev 2019, 40, 1207–1249. [Google Scholar] [CrossRef]
- Leach, D.A.; Powell, S.M.; Bevan, C.L. WOMEN IN CANCER THEMATIC REVIEW: New roles for nuclear receptors in prostate cancer. Endocr Relat Cancer 2016, 23, T85–T108. [Google Scholar] [CrossRef]
- Roshan-Moniri, M.; Hsing, M.; Butler, M.S.; Cherkasov, A.; Rennie, P.S. Orphan nuclear receptors as drug targets for the treatment of prostate and breast cancers. Cancer Treat Rev 2014, 40, 1137–1152. [Google Scholar] [CrossRef]
- Means, A.R.; Comstock, J.P.; Rosenfeld, G.C.; O'Malley, B.W. Ovalbumin messenger RNA of chick oviduct: partial characterization, estrogen dependence, and translation in vitro. Proc Natl Acad Sci U S A 1972, 69, 1146–1150. [Google Scholar] [CrossRef] [PubMed]
- Frigo, D.E.; Bondesson, M.; Williams, C. Nuclear receptors: from molecular mechanisms to therapeutics. Essays Biochem 2021, 65, 847–856. [Google Scholar] [CrossRef]
- Hollenberg, S.M.; Weinberger, C.; Ong, E.S.; Cerelli, G.; Oro, A.; Lebo, R.; Thompson, E.B.; Rosenfeld, M.G.; Evans, R.M. Primary structure and expression of a functional human glucocorticoid receptor cDNA. Nature 1985, 318, 635–641. [Google Scholar] [CrossRef]
- Green, S.; Walter, P.; Greene, G.; Krust, A.; Goffin, C.; Jensen, E.; Scrace, G.; Waterfield, M.; Chambon, P. Cloning of the human oestrogen receptor cDNA. J Steroid Biochem 1986, 24, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.M.; Mangelsdorf, D.J. Nuclear Receptors, RXR, and the Big Bang. Cell 2014, 157, 255–266. [Google Scholar] [CrossRef]
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schutz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The nuclear receptor superfamily: the second decade. Cell 1995, 83, 835–839. [Google Scholar] [CrossRef]
- Jimenez-Panizo, A.; Perez, P.; Rojas, A.M.; Fuentes-Prior, P.; Estebanez-Perpina, E. Non-canonical dimerization of the androgen receptor and other nuclear receptors: implications for human disease. Endocr Relat Cancer 2019, 26, R479–R497. [Google Scholar] [CrossRef]
- Sever, R.; Glass, C.K. Signaling by nuclear receptors. Cold Spring Harb Perspect Biol 2013, 5, a016709. [Google Scholar] [CrossRef]
- Porter, B.A.; Ortiz, M.A.; Bratslavsky, G.; Kotula, L. Structure and Function of the Nuclear Receptor Superfamily and Current Targeted Therapies of Prostate Cancer. Cancers (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Helsen, C.; Kerkhofs, S.; Clinckemalie, L.; Spans, L.; Laurent, M.; Boonen, S.; Vanderschueren, D.; Claessens, F. Structural basis for nuclear hormone receptor DNA binding. Mol Cell Endocrinol 2012, 348, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Lazar, M.A. Maturing of the nuclear receptor family. J Clin Invest 2017, 127, 1123–1125. [Google Scholar] [CrossRef] [PubMed]
- Khorasanizadeh, S.; Rastinejad, F. Nuclear-receptor interactions on DNA-response elements. Trends Biochem Sci 2001, 26, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Inukai, S.; Kock, K.H.; Bulyk, M.L. Transcription factor-DNA binding: beyond binding site motifs. Curr Opin Genet Dev 2017, 43, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Bhimsaria, D.; Rodriguez-Martinez, J.A.; Mendez-Johnson, J.L.; Ghoshdastidar, D.; Varadarajan, A.; Bansal, M.; Daniels, D.L.; Ramanathan, P.; Ansari, A.Z. Hidden modes of DNA binding by human nuclear receptors. Nat Commun 2023, 14, 4179. [Google Scholar] [CrossRef] [PubMed]
- De Bosscher, K.; Desmet, S.J.; Clarisse, D.; Estebanez-Perpina, E.; Brunsveld, L. Nuclear receptor crosstalk - defining the mechanisms for therapeutic innovation. Nat Rev Endocrinol 2020, 16, 363–377. [Google Scholar] [CrossRef]
- Font-Diaz, J.; Jimenez-Panizo, A.; Caelles, C.; Vivanco, M.D.; Perez, P.; Aranda, A.; Estebanez-Perpina, E.; Castrillo, A.; Ricote, M.; Valledor, A.F. Nuclear receptors: Lipid and hormone sensors with essential roles in the control of cancer development. Semin Cancer Biol 2021, 73, 58–75. [Google Scholar] [CrossRef]
- Dai, C.; Heemers, H.; Sharifi, N. Androgen Signaling in Prostate Cancer. Cold Spring Harb Perspect Med 2017, 7. [Google Scholar] [CrossRef]
- Culig, Z.; Santer, F.R. Androgen receptor signaling in prostate cancer. Cancer Metastasis Rev 2014, 33, 413–427. [Google Scholar] [CrossRef]
- Mills, I.G. Maintaining and reprogramming genomic androgen receptor activity in prostate cancer. Nat Rev Cancer 2014, 14, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.R.; Hammes, S.R. Nuclear receptors outside the nucleus: extranuclear signalling by steroid receptors. Nat Rev Mol Cell Biol 2016, 17, 783–797. [Google Scholar] [CrossRef]
- Augello, M.A.; Liu, D.; Deonarine, L.D.; Robinson, B.D.; Huang, D.; Stelloo, S.; Blattner, M.; Doane, A.S.; Wong, E.W.P.; Chen, Y.; et al. CHD1 Loss Alters AR Binding at Lineage-Specific Enhancers and Modulates Distinct Transcriptional Programs to Drive Prostate Tumorigenesis. Cancer Cell 2019, 35, 817–819. [Google Scholar] [CrossRef] [PubMed]
- Formaggio, N.; Rubin, M.A.; Theurillat, J.P. Loss and revival of androgen receptor signaling in advanced prostate cancer. Oncogene 2021, 40, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Labaf, M.; Zhao, Y.; Owiredu, J.; Zhang, S.; Patel, K.; Venkataramani, K.; Steinfeld, J.S.; Han, W.; Li, M.; et al. Androgen receptor splice variants drive castration-resistant prostate cancer metastasis by activating distinct transcriptional programs. J Clin Invest 2024, 134. [Google Scholar] [CrossRef] [PubMed]
- Nadal, M.; Prekovic, S.; Gallastegui, N.; Helsen, C.; Abella, M.; Zielinska, K.; Gay, M.; Vilaseca, M.; Taules, M.; Houtsmuller, A.B.; et al. Structure of the homodimeric androgen receptor ligand-binding domain. Nat Commun 2017, 8, 14388. [Google Scholar] [CrossRef] [PubMed]
- Li, M.T.; Richter, F.; Chang, C.; Irwin, R.J.; Huang, H. Androgen and retinoic acid interaction in LNCaP cells, effects on cell proliferation and expression of retinoic acid receptors and epidermal growth factor receptor. BMC Cancer 2002, 2, 16. [Google Scholar] [CrossRef] [PubMed]
- Arora, V.K.; Schenkein, E.; Murali, R.; Subudhi, S.K.; Wongvipat, J.; Balbas, M.D.; Shah, N.; Cai, L.; Efstathiou, E.; Logothetis, C.; et al. Glucocorticoid receptor confers resistance to antiandrogens by bypassing androgen receptor blockade. Cell 2013, 155, 1309–1322. [Google Scholar] [CrossRef]
- Isikbay, M.; Otto, K.; Kregel, S.; Kach, J.; Cai, Y.; Vander Griend, D.J.; Conzen, S.D.; Szmulewitz, R.Z. Glucocorticoid receptor activity contributes to resistance to androgen-targeted therapy in prostate cancer. Horm Cancer 2014, 5, 72–89. [Google Scholar] [CrossRef]
- Di Zazzo, E.; Galasso, G.; Giovannelli, P.; Di Donato, M.; Castoria, G. Estrogens and Their Receptors in Prostate Cancer: Therapeutic Implications. Front Oncol 2018, 8, 2. [Google Scholar] [CrossRef]
- Ellem, S.J.; Risbridger, G.P. Aromatase and regulating the estrogen:androgen ratio in the prostate gland. J Steroid Biochem Mol Biol 2010, 118, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.K.; Bernstein, L.; Lobo, R.A.; Shimizu, H.; Stanczyk, F.Z.; Pike, M.C.; Henderson, B.E. 5-alpha-reductase activity and risk of prostate cancer among Japanese and US white and black males. Lancet 1992, 339, 887–889. [Google Scholar] [CrossRef] [PubMed]
- Nelles, J.L.; Hu, W.Y.; Prins, G.S. Estrogen action and prostate cancer. Expert Rev Endocrinol Metab 2011, 6, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Belluti, S.; Imbriano, C.; Casarini, L. Nuclear Estrogen Receptors in Prostate Cancer: From Genes to Function. Cancers (Basel) 2023, 15. [Google Scholar] [CrossRef]
- Rogenhofer, S.; Ellinger, J.; Kahl, P.; Stoehr, C.; Hartmann, A.; Engehausen, D.; Wieland, W.F.; Muller, S.C.; Hofstadter, F.; Walter, B. Enhanced expression of peroxisome proliferate-activated receptor gamma (PPAR-gamma) in advanced prostate cancer. Anticancer Res 2012, 32, 3479–3483. [Google Scholar] [PubMed]
- Forootan, F.S.; Forootan, S.S.; Malki, M.I.; Chen, D.; Li, G.; Lin, K.; Rudland, P.S.; Foster, C.S.; Ke, Y. The expression of C-FABP and PPARgamma and their prognostic significance in prostate cancer. Int J Oncol 2014, 44, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Hartley, A.; Ahmad, I. The role of PPARgamma in prostate cancer development and progression. Br J Cancer 2023, 128, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Pakula, H.; Xiang, D.; Li, Z. A Tale of Two Signals: AR and WNT in Development and Tumorigenesis of Prostate and Mammary Gland. Cancers (Basel) 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Komiya, Y.; Habas, R. Wnt signal transduction pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev Cell 2009, 17, 9–26. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X. Targeting the Wnt/beta-catenin signaling pathway in cancer. J Hematol Oncol 2020, 13, 165. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, Q.; Xu, H. Wnt/beta-catenin signal transduction pathway in prostate cancer and associated drug resistance. Discov Oncol 2021, 12, 40. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, L.; Kikhtyak, Z.; Laven-Law, G.; Pederson, S.M.; Puiu, C.G.; D'Santos, C.S.; Lim, E.; Carroll, J.S.; Tilley, W.D.; Dwyer, A.R.; et al. The androgen receptor interacts with GATA3 to transcriptionally regulate a luminal epithelial cell phenotype in breast cancer. Genome Biol 2024, 25, 44. [Google Scholar] [CrossRef] [PubMed]
- Crockford, S.J. Evolutionary roots of iodine and thyroid hormones in cell-cell signaling. Integr Comp Biol 2009, 49, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.; Heyland, A. Evolution of thyroid hormone signaling in animals: Non-genomic and genomic modes of action. Mol Cell Endocrinol 2017, 459, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Heyland, A.; Hodin, J.; Reitzel, A.M. Hormone signaling in evolution and development: a non-model system approach. Bioessays 2005, 27, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Heyland, A.; Moroz, L.L. Cross-kingdom hormonal signaling: an insight from thyroid hormone functions in marine larvae. J Exp Biol 2005, 208, 4355–4361. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, C.; Thompson, C.C.; Ong, E.S.; Lebo, R.; Gruol, D.J.; Evans, R.M. The c-erb-A gene encodes a thyroid hormone receptor. Nature 1986, 324, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Lipkin, S.M.; Devary, O.V.; Rosenfeld, M.G. Positive and negative regulation of gene transcription by a retinoic acid-thyroid hormone receptor heterodimer. Cell 1989, 59, 697–708. [Google Scholar] [CrossRef]
- Cheng, S.Y. Thyroid hormone receptor mutations and disease: beyond thyroid hormone resistance. Trends Endocrinol Metab 2005, 16, 176–182. [Google Scholar] [CrossRef]
- Cheng, S.Y.; Leonard, J.L.; Davis, P.J. Molecular aspects of thyroid hormone actions. Endocr Rev 2010, 31, 139–170. [Google Scholar] [CrossRef]
- Chiamolera, M.I.; Sidhaye, A.R.; Matsumoto, S.; He, Q.; Hashimoto, K.; Ortiga-Carvalho, T.M.; Wondisford, F.E. Fundamentally distinct roles of thyroid hormone receptor isoforms in a thyrotroph cell line are due to differential DNA binding. Mol Endocrinol 2012, 26, 926–939. [Google Scholar] [CrossRef]
- Putcha, B.D.; Fernandez, E.J. Direct interdomain interactions can mediate allosterism in the thyroid receptor. J Biol Chem 2009, 284, 22517–22524. [Google Scholar] [CrossRef]
- Aranda, A.; Martinez-Iglesias, O.; Ruiz-Llorente, L.; Garcia-Carpizo, V.; Zambrano, A. Thyroid receptor: roles in cancer. Trends Endocrinol Metab 2009, 20, 318–324. [Google Scholar] [CrossRef]
- Flamant, F.; Cheng, S.Y.; Hollenberg, A.N.; Moeller, L.C.; Samarut, J.; Wondisford, F.E.; Yen, P.M.; Refetoff, S. Thyroid Hormone Signaling Pathways: Time for a More Precise Nomenclature. Endocrinology 2017, 158, 2052–2057. [Google Scholar] [CrossRef] [PubMed]
- Flamant, F. Futures Challenges in Thyroid Hormone Signaling Research. Front Endocrinol (Lausanne) 2016, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Jouravel, N.; Sablin, E.; Togashi, M.; Baxter, J.D.; Webb, P.; Fletterick, R.J. Molecular basis for dimer formation of TRbeta variant D355R. Proteins 2009, 75, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Tovo-Neto, A.; da Silva Rodrigues, M.; Habibi, H.R.; Nobrega, R.H. Thyroid hormone actions on male reproductive system of teleost fish. Gen Comp Endocrinol 2018, 265, 230–236. [Google Scholar] [CrossRef]
- Rodrigues, M.S.; Tovo-Neto, A.; Rosa, I.F.; Doretto, L.B.; Fallah, H.P.; Habibi, H.R.; Nobrega, R.H. Thyroid Hormones Deficiency Impairs Male Germ Cell Development: A Cross Talk Between Hypothalamic-Pituitary-Thyroid, and-Gonadal Axes in Zebrafish. Front Cell Dev Biol 2022, 10, 865948. [Google Scholar] [CrossRef]
- Nabi, G.; Hao, Y.; Liu, X.; Sun, Y.; Wang, Y.; Jiang, C.; Li, J.; Wu, Y.; Li, D. Hypothalamic-Pituitary-Thyroid Axis Crosstalk With the Hypothalamic-Pituitary-Gonadal Axis and Metabolic Regulation in the Eurasian Tree Sparrow During Mating and Non-mating Periods. Front Endocrinol (Lausanne) 2020, 11, 303. [Google Scholar] [CrossRef]
- Kumar, A.; Shekhar, S.; Dhole, B. Thyroid and male reproduction. Indian J Endocrinol Metab 2014, 18, 23–31. [Google Scholar] [CrossRef]
- Quartuccio, M.F., E.; Medica, P.; Cristarella, S.; Emmanuele, G.; Sinagra, L.; Liotta, L. Correlation between sperm parameters and circulating thyroid hormones and testosterone concentrations in Labrador Retriever dog. Ital. J. Anim. Sci. 2021, 20, 947–954. [Google Scholar] [CrossRef]
- Castaneda Cortes, D.C.; Langlois, V.S.; Fernandino, J.I. Crossover of the hypothalamic pituitary-adrenal/interrenal, -thyroid, and -gonadal axes in testicular development. Front Endocrinol (Lausanne) 2014, 5, 139. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Zhu, Y. A New Perspective on Thyroid Hormones: Crosstalk with Reproductive Hormones in Females. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.S.; Wajner, S.M.; Maia, A.L. The role of thyroid hormone in testicular development and function. J Endocrinol 2008, 199, 351–365. [Google Scholar] [CrossRef]
- Aruldhas, M.M.; Ramalingam, N.; Jaganathan, A.; John Sashi, A.M.; Stanley, J.A.; Nagappan, A.S.; Vasavan, J.; Kannan, A.; Seshadri, V.N. Gestational and neonatal-onset hypothyroidism alters androgen receptor status in rat prostate glands at adulthood. Prostate 2010, 70, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.; Chainy, G.B.; Mishro, M.M. Experimentally induced hypo- and hyper-thyroidism influence on the antioxidant defence system in adult rat testis. Andrologia 2003, 35, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Crawford, M.; Kennedy, L. Testosterone replacement therapy: role of pituitary and thyroid in diagnosis and treatment. Transl Androl Urol 2016, 5, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.D.S. Thyroid Disorders and Semen Quality. Biomed. Pharmacol. J. 2018, 11, 01–10. [Google Scholar] [CrossRef]
- Goemann, I.M.; Romitti, M.; Meyer, E.L.S.; Wajner, S.M.; Maia, A.L. Role of thyroid hormones in the neoplastic process: an overview. Endocr Relat Cancer 2017, 24, R367–R385. [Google Scholar] [CrossRef]
- Hercbergs, A. Clinical Implications and Impact of Discovery of the Thyroid Hormone Receptor on Integrin alphavbeta3-A Review. Front Endocrinol (Lausanne) 2019, 10, 565. [Google Scholar] [CrossRef] [PubMed]
- Anguiano, B.; Lopez, A.; Delgado, G.; Romero, C.; Aceves, C. Deiodinase type 1 activity is expressed in the prostate of pubescent rats and is modulated by thyroid hormones, prolactin and sex hormones. J Endocrinol 2006, 190, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Anguiano, B.; Montes de Oca, C.; Delgado-Gonzalez, E.; Aceves, C. Prostate gland as a target organ of thyroid hormones: advances and controversies. Endocr Connect 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.R.; Chaker, L.; Ruiter, R.; Aerts, J.G.; Hofman, A.; Dehghan, A.; Franco, O.H.; Stricker, B.H.; Peeters, R.P. Thyroid Function and Cancer Risk: The Rotterdam Study. J Clin Endocrinol Metab 2016, 101, 5030–5036. [Google Scholar] [CrossRef] [PubMed]
- Mondul, A.M.; Weinstein, S.J.; Bosworth, T.; Remaley, A.T.; Virtamo, J.; Albanes, D. Circulating thyroxine, thyroid-stimulating hormone, and hypothyroid status and the risk of prostate cancer. PLoS One 2012, 7, e47730. [Google Scholar] [CrossRef] [PubMed]
- Voutsadakis, I.A. The TSH/Thyroid Hormones Axis and Breast Cancer. J Clin Med 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Hellevik, A.I.; Asvold, B.O.; Bjoro, T.; Romundstad, P.R.; Nilsen, T.I.; Vatten, L.J. Thyroid function and cancer risk: a prospective population study. Cancer Epidemiol Biomarkers Prev 2009, 18, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Moeller, L.C.; Fuhrer, D. Thyroid hormone, thyroid hormone receptors, and cancer: a clinical perspective. Endocr Relat Cancer 2013, 20, R19–29. [Google Scholar] [CrossRef] [PubMed]
- Krashin, E.; Piekielko-Witkowska, A.; Ellis, M.; Ashur-Fabian, O. Thyroid Hormones and Cancer: A Comprehensive Review of Preclinical and Clinical Studies. Front Endocrinol (Lausanne) 2019, 10, 59. [Google Scholar] [CrossRef]
- Liu, Y.C.; Yeh, C.T.; Lin, K.H. Molecular Functions of Thyroid Hormone Signaling in Regulation of Cancer Progression and Anti-Apoptosis. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef]
- Atkins, M.B.; Mier, J.W.; Parkinson, D.R.; Gould, J.A.; Berkman, E.M.; Kaplan, M.M. Hypothyroidism after treatment with interleukin-2 and lymphokine-activated killer cells. N Engl J Med 1988, 318, 1557–1563. [Google Scholar] [CrossRef] [PubMed]
- Hercbergs, A. Hypothyroidism and tumor regression. N Engl J Med 1988, 319, 1351–1352. [Google Scholar] [CrossRef] [PubMed]
- Miro, C.; Di Cicco, E.; Ambrosio, R.; Mancino, G.; Di Girolamo, D.; Cicatiello, A.G.; Sagliocchi, S.; Nappi, A.; De Stefano, M.A.; Luongo, C.; et al. Thyroid hormone induces progression and invasiveness of squamous cell carcinomas by promoting a ZEB-1/E-cadherin switch. Nat Commun 2019, 10, 5410. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Suzuki, N.; Mori, J.; Oshima, A.; Usami, S.; Hashizume, K. micro-Crystallin as an intracellular 3,5,3'-triiodothyronine holder in vivo. Mol Endocrinol 2007, 21, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, O.; Pencik, J.; Hartenbach, M.; Moazzami, A.A.; Schlederer, M.; Balber, T.; Varady, A.; Philippe, C.; Baltzer, P.A.; Mazumder, B.; et al. Thyroid and androgen receptor signaling are antagonized by mu-Crystallin in prostate cancer. Int J Cancer 2021, 148, 731–747. [Google Scholar] [CrossRef] [PubMed]
- Malinowska, K.; Cavarretta, I.T.; Susani, M.; Wrulich, O.A.; Uberall, F.; Kenner, L.; Culig, Z. Identification of mu-crystallin as an androgen-regulated gene in human prostate cancer. Prostate 2009, 69, 1109–1118. [Google Scholar] [CrossRef]
- Mousses, S.; Bubendorf, L.; Wagner, U.; Hostetter, G.; Kononen, J.; Cornelison, R.; Goldberger, N.; Elkahloun, A.G.; Willi, N.; Koivisto, P.; et al. Clinical validation of candidate genes associated with prostate cancer progression in the CWR22 model system using tissue microarrays. Cancer Res 2002, 62, 1256–1260. [Google Scholar] [PubMed]
- Aksoy, O.; Hantusch, B.; Kenner, L. Emerging role of T3-binding protein mu-crystallin (CRYM) in health and disease. Trends Endocrinol Metab, 1016. [Google Scholar] [CrossRef]
- Kim, W.G.; Cheng, S.Y. Thyroid hormone receptors and cancer. Biochim Biophys Acta 2013, 1830, 3928–3936. [Google Scholar] [CrossRef]
- Muscat, G.E.; Eriksson, N.A.; Byth, K.; Loi, S.; Graham, D.; Jindal, S.; Davis, M.J.; Clyne, C.; Funder, J.W.; Simpson, E.R.; et al. Research resource: nuclear receptors as transcriptome: discriminant and prognostic value in breast cancer. Mol Endocrinol 2013, 27, 350–365. [Google Scholar] [CrossRef]
- Gu, G.; Gelsomino, L.; Covington, K.R.; Beyer, A.R.; Wang, J.; Rechoum, Y.; Huffman, K.; Carstens, R.; Ando, S.; Fuqua, S.A. Targeting thyroid hormone receptor beta in triple-negative breast cancer. Breast Cancer Res Treat 2015, 150, 535–545. [Google Scholar] [CrossRef]
- Park, J.W.; Zhao, L.; Willingham, M.; Cheng, S.Y. Oncogenic mutations of thyroid hormone receptor beta. Oncotarget 2015, 6, 8115–8131. [Google Scholar] [CrossRef] [PubMed]
- Rosen, M.D.; Privalsky, M.L. Thyroid hormone receptor mutations in cancer and resistance to thyroid hormone: perspective and prognosis. J Thyroid Res 2011, 2011, 361304. [Google Scholar] [CrossRef] [PubMed]
- Davidson, C.D.; Gillis, N.E.; Carr, F.E. Thyroid Hormone Receptor Beta as Tumor Suppressor: Untapped Potential in Treatment and Diagnostics in Solid Tumors. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Iglesias, O.; Garcia-Silva, S.; Tenbaum, S.P.; Regadera, J.; Larcher, F.; Paramio, J.M.; Vennstrom, B.; Aranda, A. Thyroid hormone receptor beta1 acts as a potent suppressor of tumor invasiveness and metastasis. Cancer Res 2009, 69, 501–509. [Google Scholar] [CrossRef]
- Horkko, T.T.; Tuppurainen, K.; George, S.M.; Jernvall, P.; Karttunen, T.J.; Makinen, M.J. Thyroid hormone receptor beta1 in normal colon and colorectal cancer-association with differentiation, polypoid growth type and K-ras mutations. Int J Cancer 2006, 118, 1653–1659. [Google Scholar] [CrossRef]
- Schnoell, J.; Kotowski, U.; Jank, B.J.; Stoiber, S.; Gurnhofer, E.; Schlederer, M.; Heiduschka, G.; Kenner, L.; Kadletz-Wanke, L. Prognostic Relevance of Thyroid-Hormone-Associated Proteins in Adenoid Cystic Carcinoma of the Head and Neck. J Pers Med 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Kuhn, C.; Mayr, D.; Ditsch, N.; Kailuweit, M.; Wolf, V.; Harbeck, N.; Mahner, S.; Jeschke, U.; Cavailles, V.; et al. Cytoplasmic and Nuclear Forms of Thyroid Hormone Receptor beta1 Are Inversely Associated with Survival in Primary Breast Cancer. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Baumann, C.T.; Maruvada, P.; Hager, G.L.; Yen, P.M. Nuclear cytoplasmic shuttling by thyroid hormone receptors. multiple protein interactions are required for nuclear retention. J Biol Chem 2001, 276, 11237–11245. [Google Scholar] [CrossRef] [PubMed]
- Shiota, M.; Fujimoto, N.; Kashiwagi, E.; Eto, M. The Role of Nuclear Receptors in Prostate Cancer. Cells 2019, 8. [Google Scholar] [CrossRef]
- Hercbergs, A.H.; Ashur-Fabian, O.; Garfield, D. Thyroid hormones and cancer: clinical studies of hypothyroidism in oncology. Curr Opin Endocrinol Diabetes Obes 2010, 17, 432–436. [Google Scholar] [CrossRef]
- Davis, P.J.; Tang, H.Y.; Hercbergs, A.; Lin, H.Y.; Keating, K.A.; Mousa, S.A. Bioactivity of Thyroid Hormone Analogs at Cancer Cells. Front Endocrinol (Lausanne) 2018, 9, 739. [Google Scholar] [CrossRef] [PubMed]
- Hall, L.C.; Salazar, E.P.; Kane, S.R.; Liu, N. Effects of thyroid hormones on human breast cancer cell proliferation. J Steroid Biochem Mol Biol 2008, 109, 57–66. [Google Scholar] [CrossRef]
- Moriggi, G.; Verga Falzacappa, C.; Mangialardo, C.; Michienzi, S.; Stigliano, A.; Brunetti, E.; Toscano, V.; Misiti, S. Thyroid hormones (T3 and T4): dual effect on human cancer cell proliferation. Anticancer Res 2011, 31, 89–96. [Google Scholar]
- Chen, C.Y.; Tsai, M.M.; Chi, H.C.; Lin, K.H. Biological significance of a thyroid hormone-regulated secretome. Biochim Biophys Acta 2013, 1834, 2271–2284. [Google Scholar] [CrossRef] [PubMed]
- Czarnecka, A.M.; Matak, D.; Szymanski, L.; Czarnecka, K.H.; Lewicki, S.; Zdanowski, R.; Brzezianska-Lasota, E.; Szczylik, C. Triiodothyronine regulates cell growth and survival in renal cell cancer. Int J Oncol 2016, 49, 1666–1678. [Google Scholar] [CrossRef]
- Theodossiou, C.; Skrepnik, N.; Robert, E.G.; Prasad, C.; Axelrad, T.W.; Schapira, D.V.; Hunt, J.D. Propylthiouracil-induced hypothyroidism reduces xenograft tumor growth in athymic nude mice. Cancer 1999, 86, 1596–1601. [Google Scholar] [CrossRef]
- Zhu, W.; Young, C.Y. Androgen-Dependent transcriptional regulation of the prostate-specific antigen gene by thyroid hormone 3,5,3'-L-triiodothyronine. J Androl 2001, 22, 136–141. [Google Scholar] [CrossRef]
- Esquenet, M.; Swinnen, J.V.; Heyns, W.; Verhoeven, G. Triiodothyronine modulates growth, secretory function and androgen receptor concentration in the prostatic carcinoma cell line LNCaP. Mol Cell Endocrinol 1995, 109, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Hsieh, M.L.; Zhu, W.; Klee, G.G.; Tindall, D.J.; Young, C.Y. Interactive effects of triiodothyronine and androgens on prostate cell growth and gene expression. Endocrinology 1999, 140, 1665–1671. [Google Scholar] [CrossRef]
- Chen, R.N.; Huang, Y.H.; Lin, Y.C.; Yeh, C.T.; Liang, Y.; Chen, S.L.; Lin, K.H. Thyroid hormone promotes cell invasion through activation of furin expression in human hepatoma cell lines. Endocrinology 2008, 149, 3817–3831. [Google Scholar] [CrossRef]
- Tai, P.J.; Huang, Y.H.; Shih, C.H.; Chen, R.N.; Chen, C.D.; Chen, W.J.; Wang, C.S.; Lin, K.H. Direct regulation of androgen receptor-associated protein 70 by thyroid hormone and its receptors. Endocrinology 2007, 148, 3485–3495. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.M.; Huang, Y.H.; Yeh, C.T.; Tsai, M.M.; Liao, C.H.; Cheng, W.L.; Chen, W.J.; Lin, K.H. Cathepsin H regulated by the thyroid hormone receptors associate with tumor invasion in human hepatoma cells. Oncogene 2011, 30, 2057–2069. [Google Scholar] [CrossRef] [PubMed]
- Chan, I.H.; Privalsky, M.L. Isoform-specific transcriptional activity of overlapping target genes that respond to thyroid hormone receptors alpha1 and beta1. Mol Endocrinol 2009, 23, 1758–1775. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Yauk, C.L.; Rowan-Carroll, A.; You, S.H.; Zoeller, R.T.; Lambert, I.; Wade, M.G. Identification of thyroid hormone receptor binding sites and target genes using ChIP-on-chip in developing mouse cerebellum. PLoS One 2009, 4, e4610. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, A.; Garcia-Carpizo, V.; Gallardo, M.E.; Villamuera, R.; Gomez-Ferreria, M.A.; Pascual, A.; Buisine, N.; Sachs, L.M.; Garesse, R.; Aranda, A. The thyroid hormone receptor beta induces DNA damage and premature senescence. J Cell Biol 2014, 204, 129–146. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wu, D.; Ng, C.F.; Teoh, J.Y.; Yu, S.; Wang, Y.; Chan, F.L. Nuclear receptor profiling in prostatospheroids and castration-resistant prostate cancer. Endocr Relat Cancer 2018, 25, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.J.; Lin, H.Y.; Mousa, S.A.; Luidens, M.K.; Hercbergs, A.A.; Wehling, M.; Davis, F.B. Overlapping nongenomic and genomic actions of thyroid hormone and steroids. Steroids 2011, 76, 829–833. [Google Scholar] [CrossRef]
- Hoermann, R.; Cheung, A.S.; Milne, M.; Grossmann, M. Hypothalamic-Pituitary-Thyroid Axis Set Point Alterations Are Associated With Body Composition in Androgen-Deprived Men. J Endocr Soc 2017, 1, 874–885. [Google Scholar] [CrossRef]
- Miro, C.; Di Giovanni, A.; Murolo, M.; Cicatiello, A.G.; Nappi, A.; Sagliocchi, S.; Di Cicco, E.; Morra, F.; Celetti, A.; Pacifico, F.; et al. Thyroid hormone and androgen signals mutually interplay and enhance inflammation and tumorigenic activation of tumor microenvironment in prostate cancer. Cancer Lett 2022, 532, 215581. [Google Scholar] [CrossRef]
- Flood, D.E.; Fernandino, J.I.; Langlois, V.S. Thyroid hormones in male reproductive development: evidence for direct crosstalk between the androgen and thyroid hormone axes. Gen Comp Endocrinol 2013, 192, 2–14. [Google Scholar] [CrossRef]
- Campbell, D.E.K.; Langlois, V.S. Thyroid hormones and androgens differentially regulate gene expression in testes and ovaries of sexually mature Silurana tropicalis. Gen Comp Endocrinol 2018, 267, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Torabinejad, S.; Miro, C.; Barone, B.; Imbimbo, C.; Crocetto, F.; Dentice, M. The androgen-thyroid hormone crosstalk in prostate cancer and the clinical implications. Eur Thyroid J 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.A.D.; Lund, D.; Szekeres, F.; Karlsson, S.; Faresjo, M.; Larsson, D. Cis-regulatory elements in conserved non-coding sequences of nuclear receptor genes indicate for crosstalk between endocrine systems. Open Med (Wars) 2021, 16, 640–650. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Beninghof, E.G.; Koenig, R.J. Dimerization interfaces of thyroid hormone, retinoic acid, vitamin D, and retinoid X receptors. J Biol Chem 1993, 268, 11534–11541. [Google Scholar] [CrossRef] [PubMed]
- Yen, P.M.; Sugawara, A.; Chin, W.W. Triiodothyronine (T3) differentially affects T3-receptor/retinoic acid receptor and T3-receptor/retinoid X receptor heterodimer binding to DNA. J Biol Chem 1992, 267, 23248–23252. [Google Scholar] [CrossRef] [PubMed]
- Schrader, M.; Carlberg, C. Thyroid hormone and retinoic acid receptors form heterodimers with retinoid X receptors on direct repeats, palindromes, and inverted palindromes. DNA Cell Biol 1994, 13, 333–341. [Google Scholar] [CrossRef]
- Lee, S.; Privalsky, M.L. Heterodimers of retinoic acid receptors and thyroid hormone receptors display unique combinatorial regulatory properties. Mol Endocrinol 2005, 19, 863–878. [Google Scholar] [CrossRef] [PubMed]
- Schilthuis, J.G.; Gann, A.A.; Brockes, J.P. Chimeric retinoic acid/thyroid hormone receptors implicate RAR-alpha 1 as mediating growth inhibition by retinoic acid. EMBO J 1993, 12, 3459–3466. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.G.; Basset, P.; Anglard, P. Multiple regulatory elements in the murine stromelysin-3 promoter. Evidence for direct control by CCAAT/enhancer-binding protein beta and thyroid and retinoid receptors. J Biol Chem 2000, 275, 39981–39990. [Google Scholar] [CrossRef]
- Davis, K.D.; Lazar, M.A. Selective antagonism of thyroid hormone action by retinoic acid. J Biol Chem 1992, 267, 3185–3189. [Google Scholar] [CrossRef]
- Gil-Ibanez, P.; Bernal, J.; Morte, B. Thyroid hormone regulation of gene expression in primary cerebrocortical cells: role of thyroid hormone receptor subtypes and interactions with retinoic acid and glucocorticoids. PLoS One 2014, 9, e91692. [Google Scholar] [CrossRef] [PubMed]
- Khanim, F.L.; Gommersall, L.M.; Wood, V.H.; Smith, K.L.; Montalvo, L.; O'Neill, L.P.; Xu, Y.; Peehl, D.M.; Stewart, P.M.; Turner, B.M.; et al. Altered SMRT levels disrupt vitamin D3 receptor signalling in prostate cancer cells. Oncogene 2004, 23, 6712–6725. [Google Scholar] [CrossRef] [PubMed]
- Girault, I.; Lerebours, F.; Amarir, S.; Tozlu, S.; Tubiana-Hulin, M.; Lidereau, R.; Bieche, I. Expression analysis of estrogen receptor alpha coregulators in breast carcinoma: evidence that NCOR1 expression is predictive of the response to tamoxifen. Clin Cancer Res 2003, 9, 1259–1266. [Google Scholar] [PubMed]
- Long, M.D.; Jacobi, J.J.; Singh, P.K.; Llimos, G.; Wani, S.A.; Rowsam, A.M.; Rosario, S.R.; Hoogstraat, M.; Linder, S.; Kirk, J.; et al. Reduced NCOR2 expression accelerates androgen deprivation therapy failure in prostate cancer. Cell Rep 2021, 37, 110109. [Google Scholar] [CrossRef] [PubMed]
- Schrader, M.; Muller, K.M.; Carlberg, C. Specificity and flexibility of vitamin D signaling. Modulation of the activation of natural vitamin D response elements by thyroid hormone. J Biol Chem 1994, 269, 5501–5504. [Google Scholar] [CrossRef] [PubMed]
- Schrader, M.; Muller, K.M.; Nayeri, S.; Kahlen, J.P.; Carlberg, C. Vitamin D3-thyroid hormone receptor heterodimer polarity directs ligand sensitivity of transactivation. Nature 1994, 370, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Villalba, P.; Jimenez-Lara, A.M.; Aranda, A. Vitamin D interferes with transactivation of the growth hormone gene by thyroid hormone and retinoic acid. Mol Cell Biol 1996, 16, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Raval-Pandya, M.; Freedman, L.P.; Li, H.; Christakos, S. Thyroid hormone receptor does not heterodimerize with the vitamin D receptor but represses vitamin D receptor-mediated transactivation. Mol Endocrinol 1998, 12, 1367–1379. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.D.; Hsieh, J.C.; Whitfield, G.K.; Haussler, C.A.; Jurutka, P.W.; Galligan, M.A.; Tillman, J.B.; Spindler, S.R.; Haussler, M.R. Vitamin D receptor displays DNA binding and transactivation as a heterodimer with the retinoid X receptor, but not with the thyroid hormone receptor. J Cell Biochem 1999, 75, 462–480. [Google Scholar] [CrossRef]
- Schneider, L.; El-Yazidi, C.; Dace, A.; Maraninchi, M.; Planells, R.; Margotat, A.; Torresani, J. Expression of the 1,25-(OH)2 vitamin D3 receptor gene during the differentiation of mouse Ob17 preadipocytes and cross talk with the thyroid hormone receptor signalling pathway. J Mol Endocrinol 2005, 34, 221–235. [Google Scholar] [CrossRef]
- Gloesmann, M.; Ertl, R.; Himmel, T.; Kummer, S.; Reichart, U. Thyroid Hormone Regulates mRNA Expression of the Vitamin D Receptor in Mouse Photoreceptors. IOVS, ARVO Annual Meeting, Seattle, Wash, (1-5 May 2016). 2016, 57. [Google Scholar]
- Barrera-Hernandez, G.; Zhan, Q.; Wong, R.; Cheng, S.Y. Thyroid hormone receptor is a negative regulator in p53-mediated signaling pathways. DNA Cell Biol 1998, 17, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.K.; Sinha, R.A.; Tripathi, M.; Mendoza, A.; Ohba, K.; Sy, J.A.C.; Xie, S.Y.; Zhou, J.; Ho, J.P.; Chang, C.Y.; et al. Thyroid hormone receptor and ERRalpha coordinately regulate mitochondrial fission, mitophagy, biogenesis, and function. Sci Signal 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Araki, O.; Ying, H.; Furuya, F.; Zhu, X.; Cheng, S.Y. Thyroid hormone receptor beta mutants: Dominant negative regulators of peroxisome proliferator-activated receptor gamma action. Proc Natl Acad Sci U S A 2005, 102, 16251–16256. [Google Scholar] [CrossRef] [PubMed]
- Guigon, C.J.; Cheng, S.Y. Novel oncogenic actions of TRbeta mutants in tumorigenesis. IUBMB Life 2009, 61, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Mori, M. Crosstalk of thyroid hormone receptor and liver X receptor in lipid metabolism and beyond [Review]. Endocr J 2011, 58, 921–930. [Google Scholar] [CrossRef]
- Guigon, C.J.; Kim, D.W.; Zhu, X.; Zhao, L.; Cheng, S.Y. Tumor suppressor action of liganded thyroid hormone receptor beta by direct repression of beta-catenin gene expression. Endocrinology 2010, 151, 5528–5536. [Google Scholar] [CrossRef]
- Skah, S.; Uchuya-Castillo, J.; Sirakov, M.; Plateroti, M. The thyroid hormone nuclear receptors and the Wnt/beta-catenin pathway: An intriguing liaison. Dev Biol 2017, 422, 71–82. [Google Scholar] [CrossRef]
- Zhang, X.K.; Wills, K.N.; Husmann, M.; Hermann, T.; Pfahl, M. Novel pathway for thyroid hormone receptor action through interaction with jun and fos oncogene activities. Mol Cell Biol 1991, 11, 6016–6025. [Google Scholar] [CrossRef]
- Favre-Young, H.; Dif, F.; Roussille, F.; Demeneix, B.A.; Kelly, P.A.; Edery, M.; de Luze, A. Cross-talk between signal transducer and activator of transcription (Stat5) and thyroid hormone receptor-beta 1 (TRbeta1) signaling pathways. Mol Endocrinol 2000, 14, 1411–1424. [Google Scholar] [CrossRef]
- Guigon, C.J.; Kim, D.W.; Willingham, M.C.; Cheng, S.Y. Mutation of thyroid hormone receptor-beta in mice predisposes to the development of mammary tumors. Oncogene 2011, 30, 3381–3390. [Google Scholar] [CrossRef]
- Martin, N.P.; Marron Fernandez de Velasco, E.; Mizuno, F.; Scappini, E.L.; Gloss, B.; Erxleben, C.; Williams, J.G.; Stapleton, H.M.; Gentile, S.; Armstrong, D.L. A rapid cytoplasmic mechanism for PI3 kinase regulation by the nuclear thyroid hormone receptor, TRbeta, and genetic evidence for its role in the maturation of mouse hippocampal synapses in vivo. Endocrinology 2014, 155, 3713–3724. [Google Scholar] [CrossRef]
- Teixido, N.; Soler, M.; Rivera, N.; Bernues, J.; Meseguer, A. CCAAT/enhancer binding protein-mediated role of thyroid hormone in the developmental expression of the kidney androgen-regulated protein gene in proximal convoluted tubules. Mol Endocrinol 2006, 20, 389–404. [Google Scholar] [CrossRef] [PubMed]
- Selva, D.M.; Hammond, G.L. Thyroid hormones act indirectly to increase sex hormone-binding globulin production by liver via hepatocyte nuclear factor-4alpha. J Mol Endocrinol 2009, 43, 19–27. [Google Scholar] [CrossRef]
- Gonzalez-Sancho, J.M.; Garcia, V.; Bonilla, F.; Munoz, A. Thyroid hormone receptors/THR genes in human cancer. Cancer Lett 2003, 192, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Ichijo, S.; Furuya, F.; Shimura, H.; Hayashi, Y.; Takahashi, K.; Ohta, K.; Kobayashi, T.; Kitamura, K. Activation of the RhoB signaling pathway by thyroid hormone receptor beta in thyroid cancer cells. PLoS One 2014, 9, e116252. [Google Scholar] [CrossRef]
- Chambon, P. The nuclear receptor superfamily: a personal retrospect on the first two decades. Mol Endocrinol 2005, 19, 1418–1428. [Google Scholar] [CrossRef]
- Giguere, V.; Shago, M.; Zirngibl, R.; Tate, P.; Rossant, J.; Varmuza, S. Identification of a novel isoform of the retinoic acid receptor gamma expressed in the mouse embryo. Mol Cell Biol 1990, 10, 2335–2340. [Google Scholar] [CrossRef] [PubMed]
- Kastner, P.; Krust, A.; Mendelsohn, C.; Garnier, J.M.; Zelent, A.; Leroy, P.; Staub, A.; Chambon, P. Murine isoforms of retinoic acid receptor gamma with specific patterns of expression. Proc Natl Acad Sci U S A 1990, 87, 2700–2704. [Google Scholar] [CrossRef]
- Leroy, P.; Krust, A.; Zelent, A.; Mendelsohn, C.; Garnier, J.M.; Kastner, P.; Dierich, A.; Chambon, P. Multiple isoforms of the mouse retinoic acid receptor alpha are generated by alternative splicing and differential induction by retinoic acid. EMBO J 1991, 10, 59–69. [Google Scholar] [CrossRef]
- Zelent, A.; Mendelsohn, C.; Kastner, P.; Krust, A.; Garnier, J.M.; Ruffenach, F.; Leroy, P.; Chambon, P. Differentially expressed isoforms of the mouse retinoic acid receptor beta generated by usage of two promoters and alternative splicing. EMBO J 1991, 10, 71–81. [Google Scholar] [CrossRef]
- Lehmann, J.M.; Hoffmann, B.; Pfahl, M. Genomic organization of the retinoic acid receptor gamma gene. Nucleic Acids Res 1991, 19, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Giguere, V.; Ong, E.S.; Segui, P.; Evans, R.M. Identification of a receptor for the morphogen retinoic acid. Nature 1987, 330, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Petkovich, M.; Brand, N.J.; Krust, A.; Chambon, P. A human retinoic acid receptor which belongs to the family of nuclear receptors. Nature 1987, 330, 444–450. [Google Scholar] [CrossRef]
- Chatagnon, A.; Veber, P.; Morin, V.; Bedo, J.; Triqueneaux, G.; Semon, M.; Laudet, V.; d'Alche-Buc, F.; Benoit, G. RAR/RXR binding dynamics distinguish pluripotency from differentiation associated cis-regulatory elements. Nucleic Acids Res 2015, 43, 4833–4854. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Rosenfeld, M.G. The coregulator exchange in transcriptional functions of nuclear receptors. Genes Dev 2000, 14, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Perissi, V.; Jepsen, K.; Glass, C.K.; Rosenfeld, M.G. Deconstructing repression: evolving models of co-repressor action. Nat Rev Genet 2010, 11, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Kastner, P.; Mark, M.; Ghyselinck, N.; Krezel, W.; Dupe, V.; Grondona, J.M.; Chambon, P. Genetic evidence that the retinoid signal is transduced by heterodimeric RXR/RAR functional units during mouse development. Development 1997, 124, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Lohnes, D.; Mark, M.; Mendelsohn, C.; Dolle, P.; Dierich, A.; Gorry, P.; Gansmuller, A.; Chambon, P. Function of the retinoic acid receptors (RARs) during development (I). Craniofacial and skeletal abnormalities in RAR double mutants. Development 1994, 120, 2723–2748. [Google Scholar] [CrossRef]
- Mendelsohn, C.; Lohnes, D.; Decimo, D.; Lufkin, T.; LeMeur, M.; Chambon, P.; Mark, M. Function of the retinoic acid receptors (RARs) during development (II). Multiple abnormalities at various stages of organogenesis in RAR double mutants. Development 1994, 120, 2749–2771. [Google Scholar] [CrossRef]
- Dolle, P.; Ruberte, E.; Leroy, P.; Morriss-Kay, G.; Chambon, P. Retinoic acid receptors and cellular retinoid binding proteins. I. A systematic study of their differential pattern of transcription during mouse organogenesis. Development 1990, 110, 1133–1151. [Google Scholar] [CrossRef] [PubMed]
- Dolle, P. Developmental expression of retinoic acid receptors (RARs). Nucl Recept Signal 2009, 7, e006. [Google Scholar] [CrossRef] [PubMed]
- Crowe, D.L.; Hu, L.; Gudas, L.J.; Rheinwald, J.G. Variable expression of retinoic acid receptor (RAR beta) mRNA in human oral and epidermal keratinocytes; relation to keratin 19 expression and keratinization potential. Differentiation 1991, 48, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Hale, L.A.; Tallafuss, A.; Yan, Y.L.; Dudley, L.; Eisen, J.S.; Postlethwait, J.H. Characterization of the retinoic acid receptor genes raraa, rarab and rarg during zebrafish development. Gene Expr Patterns 2006, 6, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Ruberte, E.; Dolle, P.; Krust, A.; Zelent, A.; Morriss-Kay, G.; Chambon, P. Specific spatial and temporal distribution of retinoic acid receptor gamma transcripts during mouse embryogenesis. Development 1990, 108, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Heyworth, C.M.; Glasow, A.; Huang, Q.H.; Petrie, K.; Lanotte, M.; Benoit, G.; Gallagher, R.; Waxman, S.; Enver, T.; et al. Lineage restriction of the RARalpha gene expression in myeloid differentiation. Blood 2001, 98, 2563–2567. [Google Scholar] [CrossRef]
- Purton, L.E.; Dworkin, S.; Olsen, G.H.; Walkley, C.R.; Fabb, S.A.; Collins, S.J.; Chambon, P. RARgamma is critical for maintaining a balance between hematopoietic stem cell self-renewal and differentiation. J Exp Med 2006, 203, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Breitman, T.R.; Selonick, S.E.; Collins, S.J. Induction of differentiation of the human promyelocytic leukemia cell line (HL-60) by retinoic acid. Proc Natl Acad Sci U S A 1980, 77, 2936–2940. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.; Marchwicka, A.; Cunningham, A.; Toellner, K.M.; Marcinkowska, E. Antagonizing Retinoic Acid Receptors Increases Myeloid Cell Production by Cultured Human Hematopoietic Stem Cells. Arch Immunol Ther Exp (Warsz) 2017, 65, 69–81. [Google Scholar] [CrossRef]
- Gratas, C.; Menot, M.L.; Dresch, C.; Chomienne, C. Retinoid acid supports granulocytic but not erythroid differentiation of myeloid progenitors in normal bone marrow cells. Leukemia 1993, 7, 1156–1162. [Google Scholar]
- Wang, Z.Y.; Chen, Z. Acute promyelocytic leukemia: from highly fatal to highly curable. Blood 2008, 111, 2505–2515. [Google Scholar] [CrossRef] [PubMed]
- Kastner, P.; Chan, S. Function of RARalpha during the maturation of neutrophils. Oncogene 2001, 20, 7178–7185. [Google Scholar] [CrossRef] [PubMed]
- Wai, H.A.; Kawakami, K.; Wada, H.; Muller, F.; Vernallis, A.B.; Brown, G.; Johnson, W.E. The development and growth of tissues derived from cranial neural crest and primitive mesoderm is dependent on the ligation status of retinoic acid receptor gamma: evidence that retinoic acid receptor gamma functions to maintain stem/progenitor cells in the absence of retinoic acid. Stem Cells Dev 2015, 24, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, V.; Laursen, K.B.; Brenet, F.; Viale, A.J.; Scandura, J.M.; Gudas, L.J. RARgamma is essential for retinoic acid induced chromatin remodeling and transcriptional activation in embryonic stem cells. J Cell Sci 2013, 126, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, V.; Gudas, L.J.; Brenet, F.; Funk, P.; Viale, A.; Scandura, J.M. Epigenomic reorganization of the clustered Hox genes in embryonic stem cells induced by retinoic acid. J Biol Chem 2011, 286, 3250–3260. [Google Scholar] [CrossRef] [PubMed]
- Handberg-Thorsager, M.; Fernandez, E.; Salo, E. Stem cells and regeneration in planarians. Front Biosci 2008, 13, 6374–6394. [Google Scholar] [CrossRef] [PubMed]
- Cruz, F.D.; Matushansky, I. Solid tumor differentiation therapy - is it possible? Oncotarget 2012, 3, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Richter, F.; Joyce, A.; Fromowitz, F.; Wang, S.; Watson, J.; Watson, R.; Irwin, R.J., Jr.; Huang, H.F. Immunohistochemical localization of the retinoic Acid receptors in human prostate. J Androl 2002, 23, 830–838. [Google Scholar] [CrossRef] [PubMed]
- Hammond, L.A.; Van Krinks, C.H.; Durham, J.; Tomkins, S.E.; Burnett, R.D.; Jones, E.L.; Chandraratna, R.A.; Brown, G. Antagonists of retinoic acid receptors (RARs) are potent growth inhibitors of prostate carcinoma cells. Br J Cancer 2001, 85, 453–462. [Google Scholar] [CrossRef]
- Gao, M.; Ossowski, L.; Ferrari, A.C. Activation of Rb and decline in androgen receptor protein precede retinoic acid-induced apoptosis in androgen-dependent LNCaP cells and their androgen-independent derivative. J Cell Physiol 1999, 179, 336–346. [Google Scholar] [CrossRef]
- Gao, T.; He, B.; Pan, Y.; Li, R.; Xu, Y.; Chen, L.; Nie, Z.; Gu, L.; Wang, S. The association of retinoic acid receptor beta2(RARbeta2) methylation status and prostate cancer risk: a systematic review and meta-analysis. PLoS One 2013, 8, e62950. [Google Scholar] [CrossRef]
- Dou, M.; Zhou, X.; Fan, Z.; Ding, X.; Li, L.; Wang, S.; Xue, W.; Wang, H.; Suo, Z.; Deng, X. Clinical Significance of Retinoic Acid Receptor Beta Promoter Methylation in Prostate Cancer: A Meta-Analysis. Cell Physiol Biochem 2018, 45, 2497–2505. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.J.; Park, S.; Uskokovic, M.R.; Dawson, M.I.; Koeffler, H.P. Expression of retinoic acid receptor-beta sensitizes prostate cancer cells to growth inhibition mediated by combinations of retinoids and a 19-nor hexafluoride vitamin D3 analog. Endocrinology 1998, 139, 1972–1980. [Google Scholar] [CrossRef]
- Pasquali, D.; Thaller, C.; Eichele, G. Abnormal level of retinoic acid in prostate cancer tissues. J Clin Endocrinol Metab 1996, 81, 2186–2191. [Google Scholar] [CrossRef] [PubMed]
- Idres, N.; Marill, J.; Flexor, M.A.; Chabot, G.G. Activation of retinoic acid receptor-dependent transcription by all-trans-retinoic acid metabolites and isomers. J Biol Chem 2002, 277, 31491–31498. [Google Scholar] [CrossRef]
- Petrie, K.; Urban-Wojciuk, Z.; Sbirkov, Y.; Graham, A.; Hamann, A.; Brown, G. Retinoic acid receptor gamma is a therapeutically targetable driver of growth and survival in prostate cancer. Cancer Rep (Hoboken) 2020, 3, e1284. [Google Scholar] [CrossRef]
- Huang, G.L.; Luo, Q.; Rui, G.; Zhang, W.; Zhang, Q.Y.; Chen, Q.X.; Shen, D.Y. Oncogenic activity of retinoic acid receptor gamma is exhibited through activation of the Akt/NF-kappaB and Wnt/beta-catenin pathways in cholangiocarcinoma. Mol Cell Biol 2013, 33, 3416–3425. [Google Scholar] [CrossRef]
- Yamakawa, K.; Koyanagi-Aoi, M.; Machinaga, A.; Kakiuchi, N.; Hirano, T.; Kodama, Y.; Aoi, T. Blockage of retinoic acid signaling via RARgamma suppressed the proliferation of pancreatic cancer cells by arresting the cell cycle progression of the G1-S phase. Cancer Cell Int 2023, 23, 94. [Google Scholar] [CrossRef]
- Keedwell, R.G.; Zhao, Y.; Hammond, L.A.; Wen, K.; Qin, S.; Atangan, L.I.; Shurland, D.L.; Wallace, D.M.; Bird, R.; Reitmair, A.; et al. An antagonist of retinoic acid receptors more effectively inhibits growth of human prostate cancer cells than normal prostate epithelium. Br J Cancer 2004, 91, 580–588. [Google Scholar] [CrossRef]
- Colucci, M.; Zumerle, S.; Bressan, S.; Gianfanti, F.; Troiani, M.; Valdata, A.; D'Ambrosio, M.; Pasquini, E.; Varesi, A.; Cogo, F.; et al. Retinoic acid receptor activation reprograms senescence response and enhances anti-tumor activity of natural killer cells. Cancer Cell 2024, 42, 646–661. [Google Scholar] [CrossRef]
- Brown, G. Deregulation of All-Trans Retinoic Acid Signaling and Development in Cancer. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.S.J.Y.H., L.; Sbirkov, Y.; Brown, G.; Zelent, A.; Petrie, K. Expression of retinoic acid receptor gamma is regulated by miR-30a. Klinische Pädiatrie 2023, 235. [Google Scholar]
- Zeng, W.; Zhang, C.; Cheng, H.; Wu, Y.L.; Liu, J.; Chen, Z.; Huang, J.G.; Ericksen, R.E.; Chen, L.; Zhang, H.; et al. Targeting to the non-genomic activity of retinoic acid receptor-gamma by acacetin in hepatocellular carcinoma. Sci Rep 2017, 7, 348. [Google Scholar] [CrossRef]
- Quintana, E.; Shackleton, M.; Sabel, M.S.; Fullen, D.R.; Johnson, T.M.; Morrison, S.J. Efficient tumour formation by single human melanoma cells. Nature 2008, 456, 593–598. [Google Scholar] [CrossRef]
- Lohnes, D.; Kastner, P.; Dierich, A.; Mark, M.; LeMeur, M.; Chambon, P. Function of retinoic acid receptor gamma in the mouse. Cell 1993, 73, 643–658. [Google Scholar] [CrossRef]
- Chung, S.S.; Wang, X.; Roberts, S.S.; Griffey, S.M.; Reczek, P.R.; Wolgemuth, D.J. Oral administration of a retinoic Acid receptor antagonist reversibly inhibits spermatogenesis in mice. Endocrinology 2011, 152, 2492–2502. [Google Scholar] [CrossRef]
- Schulze, G.E.; Clay, R.J.; Mezza, L.E.; Bregman, C.L.; Buroker, R.A.; Frantz, J.D. BMS-189453, a novel retinoid receptor antagonist, is a potent testicular toxin. Toxicol Sci 2001, 59, 297–308. [Google Scholar] [CrossRef]
- Long, M.D.; Singh, P.K.; Russell, J.R.; Llimos, G.; Rosario, S.; Rizvi, A.; van den Berg, P.R.; Kirk, J.; Sucheston-Campbell, L.E.; Smiraglia, D.J.; et al. The miR-96 and RARgamma signaling axis governs androgen signaling and prostate cancer progression. Oncogene 2019, 38, 421–444. [Google Scholar] [CrossRef]
- Nicholson, R.C.; Mader, S.; Nagpal, S.; Leid, M.; Rochette-Egly, C.; Chambon, P. Negative regulation of the rat stromelysin gene promoter by retinoic acid is mediated by an AP1 binding site. EMBO J 1990, 9, 4443–4454. [Google Scholar] [CrossRef] [PubMed]
- Yasuhara, R.; Yuasa, T.; Williams, J.A.; Byers, S.W.; Shah, S.; Pacifici, M.; Iwamoto, M.; Enomoto-Iwamoto, M. Wnt/beta-catenin and retinoic acid receptor signaling pathways interact to regulate chondrocyte function and matrix turnover. J Biol Chem 2010, 285, 317–327. [Google Scholar] [CrossRef]
- Konsavage, W.M., Jr.; Kyler, S.L.; Rennoll, S.A.; Jin, G.; Yochum, G.S. Wnt/beta-catenin signaling regulates Yes-associated protein (YAP) gene expression in colorectal carcinoma cells. J Biol Chem 2012, 287, 11730–11739. [Google Scholar] [CrossRef]
- Bauzone, M.; Souidi, M.; Dessein, A.F.; Wisztorski, M.; Vincent, A.; Gimeno, J.P.; Monte, D.; Van Seuningen, I.; Gespach, C.; Huet, G. Cross-talk between YAP and RAR-RXR Drives Expression of Stemness Genes to Promote 5-FU Resistance and Self-Renewal in Colorectal Cancer Cells. Mol Cancer Res 2021, 19, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Ou, C.; Sun, Z.; Li, S.; Li, G.; Li, X.; Ma, J. Dual roles of yes-associated protein (YAP) in colorectal cancer. Oncotarget 2017, 8, 75727–75741. [Google Scholar] [CrossRef] [PubMed]
- Hindley, C.J.; Condurat, A.L.; Menon, V.; Thomas, R.; Azmitia, L.M.; Davis, J.A.; Pruszak, J. The Hippo pathway member YAP enhances human neural crest cell fate and migration. Sci Rep 2016, 6, 23208. [Google Scholar] [CrossRef] [PubMed]
- Marchwicka, A.; Cebrat, M.; Laszkiewicz, A.; Sniezewski, L.; Brown, G.; Marcinkowska, E. Regulation of vitamin D receptor expression by retinoic acid receptor alpha in acute myeloid leukemia cells. J Steroid Biochem Mol Biol 2016, 159, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Schrader, M.; Muller, K.M.; Becker-Andre, M.; Carlberg, C. Response element selectivity for heterodimerization of vitamin D receptors with retinoic acid and retinoid X receptors. J Mol Endocrinol 1994, 12, 327–339. [Google Scholar] [CrossRef]
- Holick, M.F.; Uskokovic, M.; Henley, J.W.; MacLaughlin, J.; Holick, S.A.; Potts, J.T., Jr. The photoproduction of 1 alpha,25-dihydroxyvitamin D3 in skin: an approach to the therapy of vitamin-D-resistant syndromes. N Engl J Med 1980, 303, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.R.; McDonnell, D.P.; Hughes, M.; Crisp, T.M.; Mangelsdorf, D.J.; Haussler, M.R.; Pike, J.W.; Shine, J.; O'Malley, B.W. Cloning and expression of full-length cDNA encoding human vitamin D receptor. Proc Natl Acad Sci U S A 1988, 85, 3294–3298. [Google Scholar] [CrossRef]
- Crofts, L.A.; Hancock, M.S.; Morrison, N.A.; Eisman, J.A. Multiple promoters direct the tissue-specific expression of novel N-terminal variant human vitamin D receptor gene transcripts. Proc Natl Acad Sci U S A 1998, 95, 10529–10534. [Google Scholar] [CrossRef]
- Zella, L.A.; Meyer, M.B.; Nerenz, R.D.; Lee, S.M.; Martowicz, M.L.; Pike, J.W. Multifunctional enhancers regulate mouse and human vitamin D receptor gene transcription. Mol Endocrinol 2010, 24, 128–147. [Google Scholar] [CrossRef]
- Miyamoto, K.; Kesterson, R.A.; Yamamoto, H.; Taketani, Y.; Nishiwaki, E.; Tatsumi, S.; Inoue, Y.; Morita, K.; Takeda, E.; Pike, J.W. Structural organization of the human vitamin D receptor chromosomal gene and its promoter. Mol Endocrinol 1997, 11, 1165–1179. [Google Scholar] [CrossRef] [PubMed]
- Sunn, K.L.; Cock, T.A.; Crofts, L.A.; Eisman, J.A.; Gardiner, E.M. Novel N-terminal variant of human VDR. Mol Endocrinol 2001, 15, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Fleet, J.C. The role of vitamin D in the endocrinology controlling calcium homeostasis. Mol Cell Endocrinol 2017, 453, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Aranda, A.; Pascual, A. Nuclear hormone receptors and gene expression. Physiol Rev 2001, 81, 1269–1304. [Google Scholar] [CrossRef] [PubMed]
- Carlberg, C. Vitamin D and Its Target Genes. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Fleet, J.C.; DeSmet, M.; Johnson, R.; Li, Y. Vitamin D and cancer: a review of molecular mechanisms. Biochem J 2012, 441, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Cheskis, B.J.; Freedman, L.P.; Nagpal, S. Vitamin D receptor ligands for osteoporosis. Curr Opin Investig Drugs 2006, 7, 906–911. [Google Scholar] [PubMed]
- Fendler, A.; Stephan, C.; Ralla, B.; Jung, K. Discordant Health Implications and Molecular Mechanisms of Vitamin D in Clinical and Preclinical Studies of Prostate Cancer: A Critical Appraisal of the Literature Data. Int J Mol Sci 2024, 25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Liu, M.D.; Yao, K.; Xu, S.; Yu, D.X.; Xie, D.D.; Xu, D.X. Vitamin D deficiency aggravates growth and metastasis of prostate cancer through promoting EMT in two beta-catenin-related mechanisms. J Nutr Biochem 2023, 111, 109177. [Google Scholar] [CrossRef]
- Muindi, J.R.; Yu, W.D.; Ma, Y.; Engler, K.L.; Kong, R.X.; Trump, D.L.; Johnson, C.S. CYP24A1 inhibition enhances the antitumor activity of calcitriol. Endocrinology 2010, 151, 4301–4312. [Google Scholar] [CrossRef]
- Blutt, S.E.; McDonnell, T.J.; Polek, T.C.; Weigel, N.L. Calcitriol-induced apoptosis in LNCaP cells is blocked by overexpression of Bcl-2. Endocrinology 2000, 141, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.V.; Shinghal, R.; Raghavachari, N.; Brooks, J.D.; Peehl, D.M.; Feldman, D. Analysis of vitamin D-regulated gene expression in LNCaP human prostate cancer cells using cDNA microarrays. Prostate 2004, 59, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Erzurumlu, Y.; Aydogdu, E.; Dogan, H.K.; Catakli, D.; Muhammed, M.T.; Buyuksandic, B. 1,25(OH)(2) D(3) induced vitamin D receptor signaling negatively regulates endoplasmic reticulum-associated degradation (ERAD) and androgen receptor signaling in human prostate cancer cells. Cell Signal 2023, 103, 110577. [Google Scholar] [CrossRef] [PubMed]
- Bao, B.Y.; Yeh, S.D.; Lee, Y.F. 1alpha,25-dihydroxyvitamin D3 inhibits prostate cancer cell invasion via modulation of selective proteases. Carcinogenesis 2006, 27, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Abu El Maaty, M.A.; Alborzinia, H.; Khan, S.J.; Buttner, M.; Wolfl, S. 1,25(OH)(2)D(3) disrupts glucose metabolism in prostate cancer cells leading to a truncation of the TCA cycle and inhibition of TXNIP expression. Biochim Biophys Acta Mol Cell Res 2017, 1864, 1618–1630. [Google Scholar] [CrossRef] [PubMed]
- Giangreco, A.A.; Vaishnav, A.; Wagner, D.; Finelli, A.; Fleshner, N.; Van der Kwast, T.; Vieth, R.; Nonn, L. Tumor suppressor microRNAs, miR-100 and -125b, are regulated by 1,25-dihydroxyvitamin D in primary prostate cells and in patient tissue. Cancer Prev Res (Phila) 2013, 6, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Bao, B.Y.; Ting, H.J.; Hsu, J.W.; Lee, Y.F. Protective role of 1 alpha, 25-dihydroxyvitamin D3 against oxidative stress in nonmalignant human prostate epithelial cells. Int J Cancer 2008, 122, 2699–2706. [Google Scholar] [CrossRef] [PubMed]
- Kovalenko, P.L.; Zhang, Z.; Cui, M.; Clinton, S.K.; Fleet, J.C. 1,25 dihydroxyvitamin D-mediated orchestration of anticancer, transcript-level effects in the immortalized, non-transformed prostate epithelial cell line, RWPE1. BMC Genomics 2010, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- McCray, T.; Pacheco, J.V.; Loitz, C.C.; Garcia, J.; Baumann, B.; Schlicht, M.J.; Valyi-Nagy, K.; Abern, M.R.; Nonn, L. Vitamin D sufficiency enhances differentiation of patient-derived prostate epithelial organoids. iScience 2021, 24, 101974. [Google Scholar] [CrossRef]
- Maestro, M.A.; Molnar, F.; Carlberg, C. Vitamin D and Its Synthetic Analogs. J Med Chem 2019, 62, 6854–6875. [Google Scholar] [CrossRef]
- Maestro, M.A.; Seoane, S. The Centennial Collection of VDR Ligands: Metabolites, Analogs, Hybrids and Non-Secosteroidal Ligands. Nutrients 2022, 14. [Google Scholar] [CrossRef]
- Baurska, H.; Klopot, A.; Kielbinski, M.; Chrobak, A.; Wijas, E.; Kutner, A.; Marcinkowska, E. Structure-function analysis of vitamin D(2) analogs as potential inducers of leukemia differentiation and inhibitors of prostate cancer proliferation. J Steroid Biochem Mol Biol 2011, 126, 46–54. [Google Scholar] [CrossRef]
- Trynda, J.; Turlej, E.; Milczarek, M.; Pietraszek, A.; Chodynski, M.; Kutner, A.; Wietrzyk, J. Antiproliferative Activity and in Vivo Toxicity of Double-Point Modified Analogs of 1,25-Dihydroxyergocalciferol. Int J Mol Sci 2015, 16, 24873–24894. [Google Scholar] [CrossRef]
- Wietrzyk, J.; Nevozhay, D.; Milczarek, M.; Filip, B.; Kutner, A. Toxicity and antitumor activity of the vitamin D analogs PRI-1906 and PRI-1907 in combined treatment with cyclophosphamide in a mouse mammary cancer model. Cancer Chemother Pharmacol 2008, 62, 787–797. [Google Scholar] [CrossRef]
- Milczarek, M.; Chodynski, M.; Pietraszek, A.; Stachowicz-Suhs, M.; Yasuda, K.; Sakaki, T.; Wietrzyk, J.; Kutner, A. Synthesis, CYP24A1-Dependent Metabolism and Antiproliferative Potential against Colorectal Cancer Cells of 1,25-Dihydroxyvitamin D(2) Derivatives Modified at the Side Chain and the A-Ring. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Banwell, C.M.; Singh, R.; Stewart, P.M.; Uskokovic, M.R.; Campbell, M.J. Antiproliferative signalling by 1,25(OH)2D3 in prostate and breast cancer is suppressed by a mechanism involving histone deacetylation. Recent Results Cancer Res 2003, 164, 83–98. [Google Scholar] [CrossRef]
- Trump, D.L.; Aragon-Ching, J.B. Vitamin D in prostate cancer. Asian J Androl 2018, 20, 244–252. [Google Scholar] [CrossRef]
- Xu, Y.; Shao, X.; Yao, Y.; Xu, L.; Chang, L.; Jiang, Z.; Lin, Z. Positive association between circulating 25-hydroxyvitamin D levels and prostate cancer risk: new findings from an updated meta-analysis. J Cancer Res Clin Oncol 2014, 140, 1465–1477. [Google Scholar] [CrossRef]
- Gilbert, R.; Martin, R.M.; Beynon, R.; Harris, R.; Savovic, J.; Zuccolo, L.; Bekkering, G.E.; Fraser, W.D.; Sterne, J.A.; Metcalfe, C. Associations of circulating and dietary vitamin D with prostate cancer risk: a systematic review and dose-response meta-analysis. Cancer Causes Control 2011, 22, 319–340. [Google Scholar] [CrossRef]
- Manson, J.E.; Cook, N.R.; Lee, I.M.; Christen, W.; Bassuk, S.S.; Mora, S.; Gibson, H.; Gordon, D.; Copeland, T.; D'Agostino, D.; et al. Vitamin D Supplements and Prevention of Cancer and Cardiovascular Disease. N Engl J Med 2019, 380, 33–44. [Google Scholar] [CrossRef]
- Manson, J.E.; Bassuk, S.S.; Buring, J.E.; Group, V.R. Principal results of the VITamin D and OmegA-3 TriaL (VITAL) and updated meta-analyses of relevant vitamin D trials. J Steroid Biochem Mol Biol 2020, 198, 105522. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Principles of type I and type II NR-driven gene regulation. AR, Androgen recptor; DHT, dihydrotestosterone; TR, thyroid receptor; T3, 3,5,3’-triiodo-L-thyronine; RAR, retinoic acid receptor; ATRA, all-trans retinoic acid; VDR, vitamin D receptor; 1,25 D3, 1α,25-dihydroxyvitamin D3; RXR, retinoid X recptor; CoA, coactivator; TM, transcription machinery, ARE, androgen response element; TRE, thyroid response element; RARE, retinoic acid response element; VDRE, vitamin D response element. Created with BioRender.com.
Figure 1.
Principles of type I and type II NR-driven gene regulation. AR, Androgen recptor; DHT, dihydrotestosterone; TR, thyroid receptor; T3, 3,5,3’-triiodo-L-thyronine; RAR, retinoic acid receptor; ATRA, all-trans retinoic acid; VDR, vitamin D receptor; 1,25 D3, 1α,25-dihydroxyvitamin D3; RXR, retinoid X recptor; CoA, coactivator; TM, transcription machinery, ARE, androgen response element; TRE, thyroid response element; RARE, retinoic acid response element; VDRE, vitamin D response element. Created with BioRender.com.
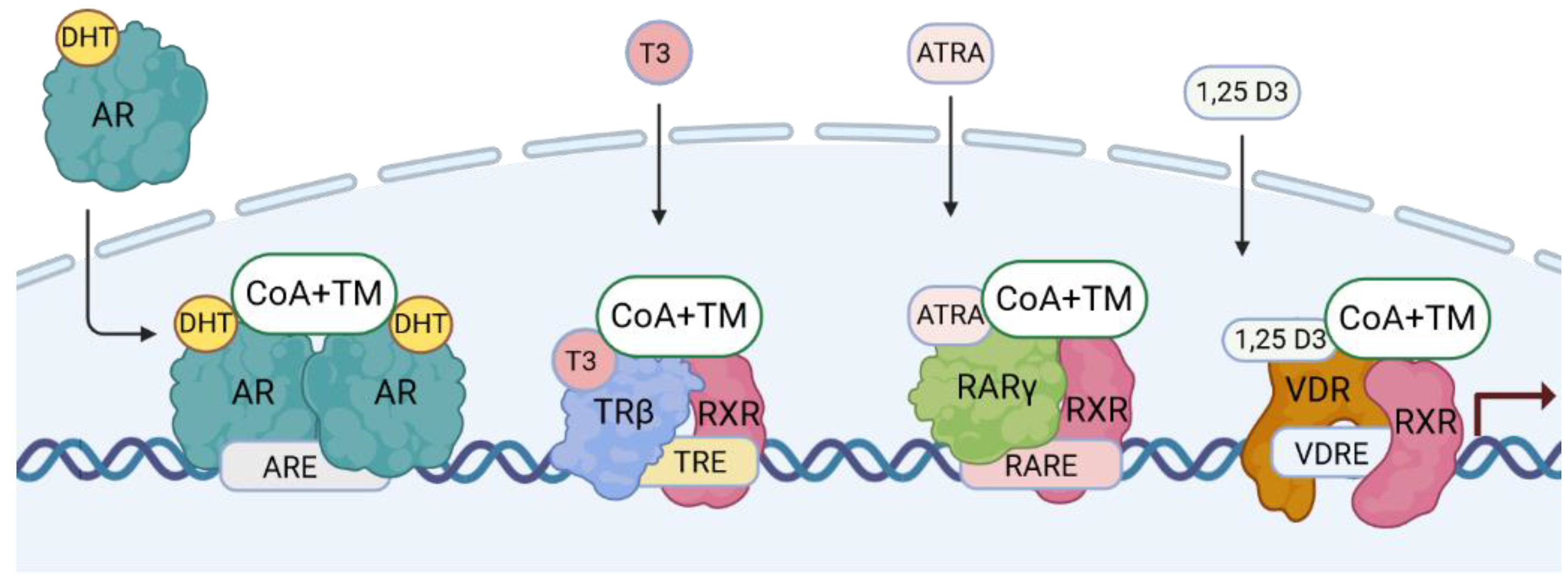
Figure 2.
Structure of the androgen (AR), thyroid (TR), retinoic acid (RAR), and vitamin D receptor (VDR) genes and proteins. NTD, N-terminal region; DBD, DNA-binding domain; HR, hinge region; LBD, ligand binding domain; AF, activation domain. Created with BioRender.com.
Figure 2.
Structure of the androgen (AR), thyroid (TR), retinoic acid (RAR), and vitamin D receptor (VDR) genes and proteins. NTD, N-terminal region; DBD, DNA-binding domain; HR, hinge region; LBD, ligand binding domain; AF, activation domain. Created with BioRender.com.
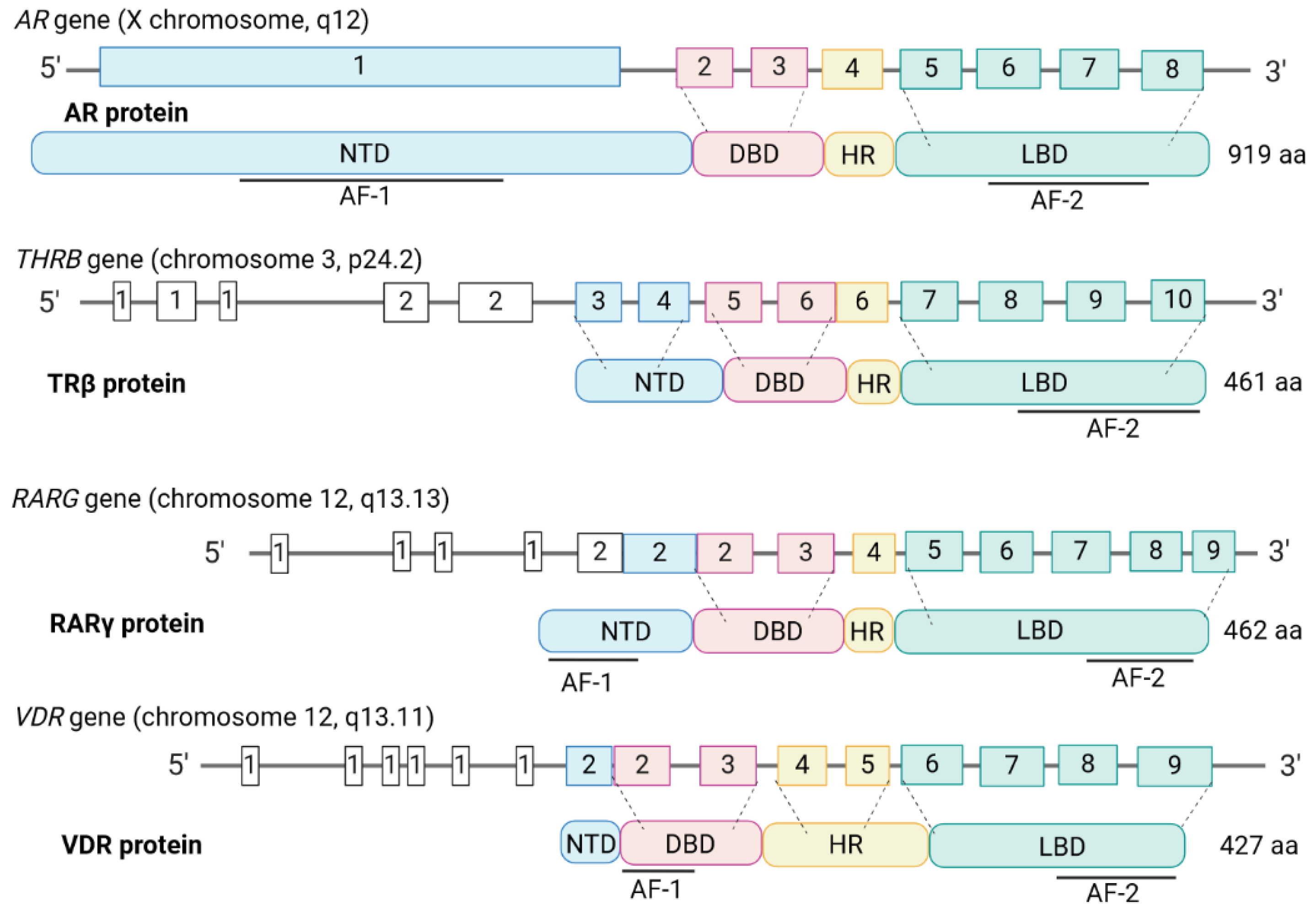
Figure 3.
Androgen influences on prostate cancer (PCa). PCa is primarily an androgen-dependent disease. Estrogen receptors (ERs) play a role due to aromatase conversion of testosterone to estradiol. There is increased expression of peroxisome proliferator-activated receptor γ (PPARγ) during PCa and AR inhibits the expression of PPARγ in healthy prostate. DHT, dihydrotestosterone. Created with BioRender.com.
Figure 3.
Androgen influences on prostate cancer (PCa). PCa is primarily an androgen-dependent disease. Estrogen receptors (ERs) play a role due to aromatase conversion of testosterone to estradiol. There is increased expression of peroxisome proliferator-activated receptor γ (PPARγ) during PCa and AR inhibits the expression of PPARγ in healthy prostate. DHT, dihydrotestosterone. Created with BioRender.com.
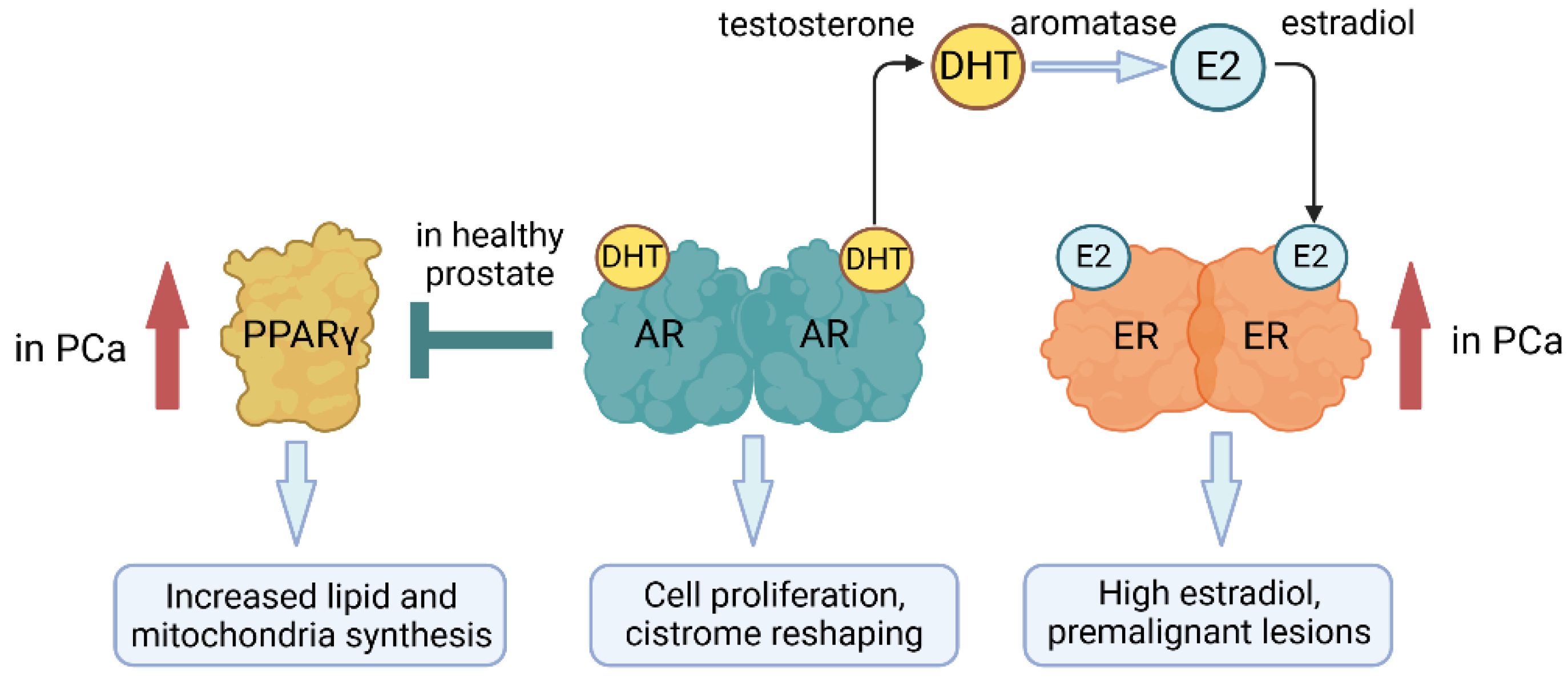
Figure 4.
Crosstalk between thyroid hormone β (TRβ) and other nuclear receptors. 3,5,3’-triiodo-L-thyronine (T3) is known to increase dihydrotestosterone (DHT) production, and thyroid hormones increase the expression of androgen receptor (AR) and androgen synthesis enzymes. T3 and DHT strongly stimulate TRβ-mediated gene expression. TR/RAR heterodimers can regulate T3-mediated gene expression, and TR might interact directly with VDR. RAR, retinoic acid receptor; VDR, vitamin D receptor. ATRA, all-trans retinoic acid; 1,25 D3, 1α,25-dihydroxyvitamin D3. Created with BioRender.com.
Figure 4.
Crosstalk between thyroid hormone β (TRβ) and other nuclear receptors. 3,5,3’-triiodo-L-thyronine (T3) is known to increase dihydrotestosterone (DHT) production, and thyroid hormones increase the expression of androgen receptor (AR) and androgen synthesis enzymes. T3 and DHT strongly stimulate TRβ-mediated gene expression. TR/RAR heterodimers can regulate T3-mediated gene expression, and TR might interact directly with VDR. RAR, retinoic acid receptor; VDR, vitamin D receptor. ATRA, all-trans retinoic acid; 1,25 D3, 1α,25-dihydroxyvitamin D3. Created with BioRender.com.
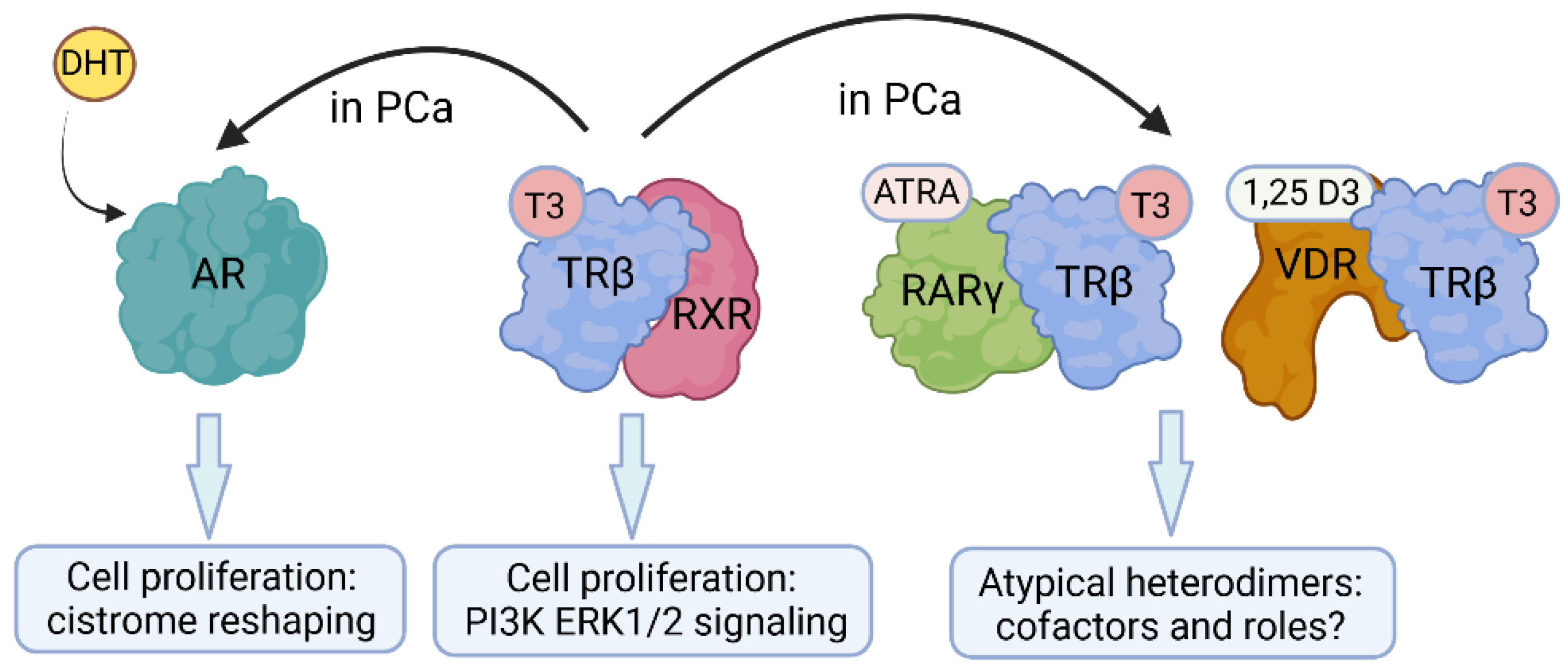
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



