
Submitted:
12 July 2024
Posted:
15 July 2024
You are already at the latest version
Abstract
Lifecycle assessment (LCA) quantified changes in environmental impact categories (global warming, eutrophication, etc.) from 2021 to 2030 due to genetic trends in (re)production traits in pig lines of breeding company Genus-PIC. The 2030 levels were projected with se-lection index theory based on weightings of traits in the breeding goals and genetic covari-ances among them. Projected improvement was 0.9% annually for most impact categories. Another LCA compared impacts of 2021 North American pig production based on PIC ge-netics versus the industry average. Software openLCA converted material and energy flows to impact categories of frameworks ReCiPe-2016, PEF-3.1 and IPCC-2021. Flows came from data recorded by customers (1.1/4.7 million sows/finishing pigs) and by subscribers to a third-party data-aggregator (1.3/9.1 million). PIC genetics have a 7-8% better impact than industry average for 13/18 categories of ReCiPe-2016, 19/25 of PEF-3.1, and all categories of IPCC-2001. Pig breeding delivers positive environmental outcomes as correlated respons-es to selection for profitability-oriented breeding goals. This trend is additive; technology development will increase it. Different investment levels in breeding population structure and technology, and different operational efficiencies of breeding companies, cause substan-tial differences in the environmental impact of pig production.
Keywords:
LCA
; animal breeding
; pig
; environmental impact
; climate change
; global warming
1. Introduction
Pig production has long been guided by efforts to increase productivity and decrease production costs – this also leads to an indirect reduction of environmental impacts. Consumers and businesses are increasingly aware of sustainability issues in our food systems, which have increased the pressure for producers to become more efficient, more focused on environmental impact, and more strategic about resource allocation and investment. The livestock sector uses resources such as land, water and energy to produce animal products, and these activities impact air, water and soil quality (De Vries and De Boer, 2010). An important component of sustainability is supported by this focus on resource efficiency, so that the global availability of high-quality, affordable, safe animal protein can be increased, and its overall environmental impact can be reduced at the same time. This challenge is intensified by global climate change and environmental degradation, by the risks to food security that follow from these issues, and by the increasing demand for animal-sourced protein by the growing global middle class.
The challenges of sustainability go beyond regulatory compliance and include identifying innovations to increase production efficiency, safety, and resilience. Sustainability is good for business: it addresses risks and creates opportunities to increase efficiency and decrease negative impacts (Dyllick and Hockerts, 2002). There remain gaps in our understanding of the impacts of production systems and in our ability to make informed decisions to guide changes in breeding, feeding, waste management, and other practices. A better understanding of these systems and their interactions will support development and implementation of technology and management practices to maintain or increase productivity and improve sustainability at the same time. System-scale models integrate scientific knowledge, management practices, and environmental and economic impacts of agroecosystems to support a rational basis for decision making.
The basic principle of Life Cycle Assessment (LCA) is to follow a product through its life cycle, by defining a boundary between its production system and the surrounding environment, and tracking the inputs and outputs of all the activities involved. LCA is commonly used in sustainability studies that quantify the potential environmental impacts of agricultural production systems and the tradeoffs between them, providing an overall framework for measuring sustainability characteristics. A brief overview of the methodology from the point of view of livestock production is given by Van Middelaar et al. (2019).
The environmental impact of pig production depends on many factors; the dominating factors are feed production and its utilization (feed conversion), and manure management. The animal as such plays a pivotal role here because its intrinsic capacities for lean tissue growth, feed efficiency, survivability and reproductive performance influence these factors. Those performance characteristics are partly under genetic control. Livestock breeding increases farm animal productivity so that more animal protein can be produced by fewer animals, using less land, water and other resources; it also increases the efficiency of that production which reduces the emission intensity of undesirable outputs such as greenhouse gases. An example of focus on sustainability specifically related to livestock genetics and breeding is the Code EFABAR initiative (http://www.responsiblebreeding.eu) where breeding and sustainability intersect multiple metrics: animal health and welfare, environment, resource use efficiency, and biodiversity.
It would then be relevant to quantify the contribution of genetic improvement to positive environmental and sustainable outcomes for the pig meat production system. In this study, we first present the genetic trends of the abovementioned traits (lean tissue growth, feed efficiency, survivability, and reproductive performance) as they were realized in PIC’s pig breeding populations, from 2010 to 2023. We then apply Life Cycle Assessment to transform such trends into trends of environmental impact factors, and we present a forecast to 2030. Finally, we compare the environmental impacts of PIC’s populations to the North American industry average of 2021 (a baseline comparison).
2. Material and Methods, or: Goal and Scope
Feed production causes much of the greenhouse gas (GHG) emission from pig production; the 84 studies cited in the first paragraph of section 4.2 report proportions between 22 and 95 % (59 ± 18 %). In line with this, the GHG emissions from a pig farm are strongly correlated to the whole-enterprise feed conversion ratio (FCRwe); FCRwe can be quantified in terms of feed intake, growth rate and survival rate of the growing-finishing pig, and feed intake and lifetime reproductive performance of the sow. Appendix A in the Supplementary Material gives more detail. Commercial pig breeding creates genetic change in all these traits, so the contribution of genetic change to FCRwe can be quantified and from there the change in GHG emission due to genetic improvement can be predicted. Section 2.4 gives more detail.
2.1. Functional Unit, System Boundary, Cut-Off Criteria and Multi-Functionality
The functional unit of this study is the delivery of 1000 kg pig live weight at the farmgate; this includes culled sows from the upstream breeding herd. It does not include any animals not fit for slaughter; these do not enter the food chain (no live weight delivered, no desirable output) but their resource consumption, and their emissions and other undesirable outputs, are included in the calculations. The systems being compared are (i) the average slaughter pig produced from the North American breeding program of a particular breeding company (Genus-PIC of Hendersonville TN, USA; see www.pic.com) versus (ii) North American industry average genetics, assuming identical animal husbandry practices, farm infrastructure, post-farm activities and ancillary activities such as accounting and travel. Our system boundary (Figure 1) begins with the extraction of raw materials and includes all animal husbandry activities, feed production (production of fertilizer, pest and disease control chemicals, seed, fuel, and energy) and manure management, and veterinary and genetic research and development (R&D) activities.
No simplifying cut-off criteria (ISO, 2006a) are implemented.
The primary multi-functional processes are the parent, grandparent (GP) and great-grandparent (GGP) stages (collectively known as the genetic pyramid; the “parent” is the parent of the slaughter pig) from where culled sows (a coproduct) are sent to slaughter; all environmental impact from these stages is assigned to the progeny, and an internal credit for the displaced growing-finishing pigs is assigned to the GP and GGP sow barns (similar to a consequential LCA approach to avoid allocation). Likewise, male piglets produced in the GP stage form another coproduct; they are sent to finishing where they displace piglets from the parent sow barn. Thus, the live weight at the farmgate accounts for all the animals leaving the system to slaughter (Figure 1).
An inventory of inputs and emissions is created for each activity and constructs the cradle-to-farmgate lifecycle inventory (LCI) model, linked from the GGP stage to the finishing stage.
2.2. Unit Processes and Their Data Requirements
The foreground system includes the main activities that deliver the functional unit of section 2.1, i.e. a typical North American commercial pig production operation from gestation and farrowing facilities to finishing and slaughter. To simulate this operation and estimate the utilities consumption we use the Pig Production Environmental Calculator of the University of Arkansas Resiliency Center (Sandefur et al., 2015). The primary data required are listed in Table S1 in the Supplementary Material. We use the market group process for USA electricity (Ecoinvent V 3.9.1 cutoff system model), and the I/O model by Carnegie Mellon University (see Hendrickson et al., 2006) for veterinary and genetic R&D services. Environmental impacts associated with infrastructure (buildings, machinery, etc.) are not included for foreground processes except when an existing Ecoinvent (see the next paragraph) unit process includes infrastructure. Emissions associated with manure management (ammonia, methane, non-methane volatile organic compounds, and nitrous oxide) are not commonly measured and are therefore modeled. This requires estimates of manure excretion and composition; we follow the LEAP recommendations (https://www.fao.org/partnerships/leap/en) to treat manure as a residual, consistent with the treatment of similar products in the background. The most relevant data gap in this study is the industry-wide distribution of systems for manure management and subsequent application; we model a uniform use of deep pit systems, as used in a significant fraction of North American production systems.
The background system includes all upstream activities. Purchased inputs to the main production processes (electricity, fuel, transportation, etc.) are sourced from the Ecoinvent v3.9.1 cut-off database (Wernet et al., 2016; https://ecoinvent.org/the-ecoinvent-association); some feed ingredients are sourced from the Agrifootprint 5.0 database (Van Paassen et al., 2019ab). In North America, activities downstream of the abattoir are not affected by genetic factors, so we exclude them from the system boundary.
2.3. Realized Genetic Trends in Performance Traits
The genetic trend of a trait is commonly calculated by relating the trait’s estimated breeding values (EBVs) of a group of animals to the animals’ date of birth. In this case the group included animals born and performance tested in PIC’s genetic nucleus farms during the recent 11 years, of the four genetic lines behind PIC’s main types of slaughter pig. The EBVs are routinely calculated, within genetic line and on a weekly basis, for all animals with a record in PIC’s worldwide database. The methodology follows Searle (1997), extended with the use of genomic information following Legarra et al. (2014); this same methodology is in use by all serious livestock breeding organizations, worldwide. For this LCA study we focused on the traits that directly influence a farm’s whole-enterprise feed conversion ratio (FCRwe i.e. the amount of feed required to produce the functional unit of section 2.1): the sow’s litter size (number born alive) and the lactation mortality rate of her piglets; and the growing-finishing pig’s feed intake, growth rate, and wean-to-finish mortality rate.
2.4. Scenario 1: Forecasting to 2030
For this forecast we assume that the composition of the various pig diets remains constant over time, and that the gross sustainability of feed production is not changed due to progress in animal genetics or management.
The genetic improvement in PIC’s routine selection traits is transformed into genetic improvement in the environmental impact factors of the LCIA. This is a semi-empirical process, based on the currently active breeding goals of the four genetic lines behind PIC’s main types of slaughter pig. Each line’s breeding goal includes about 30 traits. For each animal in the breeding program, the routine EBVs (see section 2.3) for each trait are weighted into a selection index. Selection of the next generation of breeding pigs on that index will result in genetic change (ΔG) in the underlying traits. This change can be predicted from the regression coefficients of the trait EBVs on the index; in vector form, , where i is the selection intensity on the index, b is the vector of index weighting factors, and E is the covariance matrix among the trait EBVs (Schneeberger et al., 1992). The weighting factors are constants that quantify the breeding goal, set to reflect each trait’s economic impact on farm profitability; for the forecasting in this study, E held the covariances among the trait EBVs of animals of each genetic line that were born in PIC’s nucleus farms the previous 18 months. In that same period, the genetic improvement in the index was 20.2 points per year (Genus, 2022; p.14) which is close to one standard deviation, so i can be set to 1.
As mentioned in section 2.3, the combined genetic trends of some of these traits lead to an annual change in FCRwe; Van der Peet-Schwering et al. (1999), Shirali et al. (2012), Saintilan et al. (2013), Soleimani and Gilbert (2020), Kasper et al. (2021), and Ewaoluwagbemiga et al. (2023) have shown that FCR is strongly correlated with nitrogen excretion, which is the main contributor to the GHG emissions from monogastric animals. We may then expect a profitability-oriented pig breeding program to result in a correlated genetic reduction of those emissions: “mitigation as a co-benefit to improved production efficiency” to quote Reisinger et al. (2021). Such effects can also be expected for other environmental impact factors such as eutrophication, acidification, and land use.
2.5. Scenario 2: Baseline Comparison
We model a generic pig production system with a sow barn (gestation and lactation) followed by a wean-to-finish barn with 5000 pigs (50:50 females and castrated males), with finished animals transferred to slaughter as the functional unit.
2.5.1. Data
The baseline comparison data provide the same key performance indicators (KPIs: feed conversion, mortality, etc.) from two sources.
First, PIC customers across North America with a wide range of production and nutrition practices and systems, health statuses and PIC crosses, report performance records directly to PIC. This includes data recorded in 2021 on 1.1 million parent sows and 4.7 million growing-finishing pigs from 53 and 34 multi-farm production companies, respectively.
Second, North American industry average production data as provided in aggregated form by a prominent industry benchmark with similar data quality control protocols as used by PIC. This dataset represents a similarly wide range of production practices, nutrition programs, and health statuses. The aggregator does not differentiate data collection based on genetic products, and we believe the (undocumented) contribution from PIC genetics to this data to be between 30 and 50 %.
Table 1 summarizes the data available from both sources. These sow performance data were recorded on crossbred parent sows; to properly cover the GGP and GP stages in the model (i.e. purebred sows, producing purebred and crossbred litters, respectively), their reproduction traits were adjusted for maternal and direct heterosis based on Table 10.2 of Bidanel (2011). Similarly, growth rate and feed efficiency in gilt rearing at these stages were heterosis-adjusted based on Clutter (2011, p. 347). Costs of 3.5 USD per finished pig and 2.5 USD per piglet were adopted based on enterprise production budgets (https://www.extension.iastate.edu/agdm/decisiontools.html#livestock). Veterinary and genetic R&D costs were estimated from financial data from Genus (2021) coupled with estimates of the genetic pyramid herd size; this led to an expenditure of about 22,000 USD per GGP sow. Emissions were found to be insensitive to this parameter; therefore, any uncertainty in this estimate will not affect the study conclusions.
2.5.2. Diet Composition
Diets for sows and for growing-finishing pigs were formulated according to the PIC (2021) nutrition and feeding guidelines, based on ingredients typically used in North America.
The amounts of sow feed consumed in the gestation and lactation phases were estimated with PIC’s Dynamic Sow Feeding Tool (http://dynamicfemalefeeding.pic.com/en). For the industry average evaluation, the same diets and computational procedures were used, averaging the feeding guidelines of PIC and of other breeding companies in North America because the industry average data does not specify the genetic background of its pigs. Table S2 in the Supplementary Material gives the diet compositions.
For growing-finishing pigs, diet composition was derived from the growth rate and feed conversion performance reported in the baseline comparison data of section 2.5.1. The allocation per phase was determined by the weaning body weight, market body weight and overall feed conversion using the PIC Feed Budgeting tool (https://www.pic.com/resources/nutrition-links-and-tools; overall growth rate 0.77 kg/d, overall FCR 2.55 kg/kg). Similar procedures were used for the industry average to adjust for the differences in growth rate and feed conversion (0.75 kg/d, 2.71 kg/kg). Table S3 in the Supplementary Material gives the diet compositions.
2.6. Lifecycle Impact Assessment
LCIA frameworks consist of a series of category-specific characterization factors used to convert the cumulative inventory results of an LCI model to the cumulative impact in multiple environmentally relevant categories for the functional unit of the system under study. For example, the climate change impact is calculated by taking the product of each greenhouse gas emission and its respective characterization factor to convert the cumulative, relative radiative forcing into carbon dioxide equivalents, CO2eq. We use three LCIA frameworks to characterize the environmental impact of the functional unit, as follows.
ReCiPe-2016 includes 18 impact categories covering the areas of protection identified under human health, ecosystem health, and resource conservation. It includes a 2010 world-based normalization method that can aid in defining which categories are relatively larger in the context of the annual average global per capita emissions. We apply all three the cultural perspectives of ReCiPe 2016: Egalitarian, Hierarchist and Individualist.
Environmental Footprint-3.1 is the framework required for the Product Environmental Footprint (PEF) recommended by the European Commission. It includes 25 impact categories, 14 of which are qualitatively the same as the ReCiPe 2016 categories, but may have different quantitative reporting units.
IPCC-2021 climate change includes twelve climate-related impact categories. Its category of primary interest for this study is the 100-year global warming potential (“GWP-100”), as it coincides with the climate change categories in ReCiPe and PEF. Other categories include global warming potential for different time horizons, and global temperature change potential.
The lifecycle impact assessment (LCIA) calculations are performed using openLCA v2.02 to link the individual stages of production, creating a supply chain model to convert the material and energy flows of pig production to the impact categories of the three frameworks. Background data are separately licensed from the Ecoinvent v3.9.1 cutoff and APOS system models and the Agri-footprint 5.0 databases; this includes the probability distributions for emissions of ammonia, nitrous oxide, etc. from crop production. We adopt the background database’s uncertainty distributions without revision.
2.7. Contribution Analysis
The contributions of the various processes to each impact category are quantified for scenario 1 (section 2.4). The inventory projection focuses only on the KPIs of the production systems – it does not include projected changes in efficiency or other contributing sectors such as electricity, crop yield, or transportation. Hence the contributions for scenario 2 (section 2.5) are very similar to scenario 1, and we do not perform a separate contribution analysis.
2.8. Methodological Quality Assessment
LCA has several sources of uncertainty that can influence the interpretation of the results and limit the conclusions (Ciroth et al. 2016; Mendoza Beltran et al. 2018). These include (i) natural variability in input parameters such as fertilizer or fuel use, (ii) estimated values obtained through proxy sources, substituting a similar product for one that does not exist in available databases, and (iii) results from mathematical models that include multi-year simulations of parameters associated with variable factors such as weather or soil conditions. There are also uncertainties in impact assessment characterization factors, but the studied systems are quite similar; therefore we expect these to be uniform across all compared systems and to not affect the study conclusions. Nonetheless, the selection of an LCIA framework is a value choice and should be tested for sensitivity. Comparison of the results from the ReCiPe and the PEF frameworks (section 2.6) evaluates the robustness of the results.
We quantify the uncertainty in the LCA results by Monte Carlo simulation (MCS), available in the openLCA software. This process is rules-based and incorporates prior probability distributions for uncertain or variable input data that reflect the knowledge and process uncertainty associated with the variables, and describe the range of expected or permissible parameter values. We apply log-Normal prior probability distributions to avoid sampling of negative input values – most of the reference flows in LCA are positive definite, so negative values are not permissible. MCS produces posterior probability distributions for its predicted output variables (i.e. the impact categories) which can be used for statistical significance testing; we perform two-sided unpaired ANOVA on sets of 100 MCS results for each impact category to test the significance of the contrasts (i) between PIC-2021 and the 2021 industry average, and (ii) between PIC-2021 and PIC-2030.
Characterization factors are used to translate the inventory to the impact categories. The magnitude of an observed contrast is not necessarily indicative of its statistical significance: a factor of two for climate change may be significant, while a factor of 1000 for toxicity effects may be not, partially due to the much larger range in characterization factors for toxicity. An additional explanatory factor arises in the process of uncertainty propagation in the MCS when the uncertainty of some of the contributing flows is higher, resulting in a wider frequency distribution in the MCS and therefore less differentiating power in the statistical tests. The flow uncertainty for foreground processes is quantified via the Ecoinvent pedigree matrix: five characteristics of the data (reliability, i.e. how was it measured; completeness; temporal correlation, i.e. how recent is it; geographical correlation, i.e. where is it from; technological correlation, i.e. which technology generated it) are qualitatively assessed in terms of five indicator scores (scored 1 for the highest quality e.g. “verified data based on measurements” to 5 for the lowest quality e.g. “non-qualified estimate”; Table 10.4 of Weidema et al., 2013). These are converted into five default variance values (their Table 10.5), which increase from zero to higher values for more unfavorable indicator scores. Each of these is combined with its corresponding basic uncertainty (estimated from aggregate statistics for the KPIs). A default variance value (based on expert judgement) is applied when no sampled data was available. This defines the final prior variance of the contributing flow as it goes into the MCS. Appendix B in the Supplementary Material gives the elements of our inventory with indicator scores different from [1 1 1 1 1].
Our uncertainty assessment does not include uncertainty associated with the characterization factors. Because the systems being compared are very similar, this omission of a source of variability in the results will not introduce bias in the conclusions: any single MCS would have selected the same characterization factor for both systems.
3.Results
3.1. Realized Genetic Trends of Performance Traits
Realized genetic trends of the FCRwe-relevant traits (see section 2.3) are shown in Figure 2. The white trendlines in each graph represent the pureline breeding populations; the black trendline is the average of the white ones and hence represents the typical PIC-derived slaughter pig or parent sow. The pre-2022 time trends are very stable (for feed intake since 2017), and we predict them to be continued into the future; this would lead to a forecast such as our Scenario 1 which is reported in section 3.2.1.
3.2. Scenario Analysis
For both scenarios studied here (sections 2.3 and 2.4), the estimates of the impact categories are presented not as absolute values but proportionally: for the forecast of section 2.4 we report the PIC-2030 predicted values as proportions of the PIC-2021 results, and for the baseline comparison of section 2.5 we report the PIC results as proportions of the industry average results. Fourteen of the 18 and 25 impact categories of the ReCiPe-2016 and PEF-3 frameworks (see section 2.6) are functionally similar across these frameworks, and this proportional approach makes them quantitatively comparable. For example, acidification is quantified in terms of SO2eq (in kg) by ReCiPe-2016 but in terms of H+eq (in mol) by PEF-3; this is resolved when both are transformed to a percentage of a similar associated reference value. In all these cases, a lower environmental impact is more favourable, but to stay conservative we apply two-sided significance tests.
3.2.1. Scenario 1: Forecast to 2030
As mentioned in section 2.4, this forecast was not derived from an extrapolation of the historical trends in Figure 2, but from multivariate regression exploiting the covariance matrix among the trait EBVs, based on the currently active breeding goals of these breeding populations. The predicted 2030 levels of the performance traits are in Figure 2. The resulting projected 2030 performance of the LCIA environmental impact categories (section 2.6) can be compared to the 2021 performance, and this suggests a predicted improvement of 7 to 9 % (i.e. about 0.9 % per year) for most of the impact categories in both the ReCiPe-2016 and PEF-3.1 frameworks; specifically, the predicted improvement of the global warming potential is 7.5 % (i.e. 0.83 % per year). The exceptions are for ionizing radiation (10.6 % in ReCiPe-2016 and 12.3 % in PEF-3.1); these are exceptions in the favourable direction, but the estimates are statistically non-significant (see the third paragraph of section 2.8 for the paradox). All these results are presented in Appendix A in the Supplementary Material. Figure 3 shows the results for the 14 functionally overlapping categories across the two frameworks.
3.2.1.1. Contribution Analysis
Figure 4 presents the relative contribution analysis for the ReCiPe-2016 Hierarchist and PEF-3.1 frameworks, both under the cutoff system model. The category axis has been sorted to emphasize commonalities across the contributing activities.
For ReCiPe-2016, the animal-rearing phases (finisher and piglet) are major contributors only to terrestrial acidification and fine particulate matter formation; both are dominated by ammonia emissions. Corn production is the dominant contributor across many impact categories; its primary contributing factor is emissions associated with fertilizer application. Metals, extracted and consumed in the supply chain, and fossil fuel consumption are the obvious dominant contributors to mineral resource and fossil fuel resource scarcity, respectively. The ionizing radiation category is dominated by waste management, with formaldehyde production as the primary contributing factor.
PEF-3.1 largely repeats the ReCiPe-2016 patterns, with interesting exceptions in the categories ozone depletion, land use, water use (where corn production is the dominant contributor in ReCiPe but is very minor in PEF), and in carcinogenic toxicity (where the opposite holds). In ReCiPe, land use is an indicator of potential impact on biodiversity associated with different land use classifications; in PEF the main contributor is “other” processes, dominated by infrastructure such as buildings and roads, with some contribution from forestry. The primary “waste management” contributing factor to ionizing radiation in PEF-3.1 is polystyrene.
3.2.2. Scenario 2: Baseline Comparison
Our comparison of the 2021 performance of PIC genetics to the industry average reveals that PIC genetics have a significantly lower environmental impact for 13 of the 18 impact categories in the ReCiPe-2016 framework, for 19 of the 25 categories in the PEF-3.1 framework, and all twelve climate change categories in the IPCC-2001 framework. All these results are presented in Appendix A in the Supplementary Material, and Figure 5 shows the results for the 14 categories that are functionally overlapping across ReCiPe-2016 and PEF-3.1, and for the GWP-100 category of IPCC-2021. For most of the impact categories, the PIC benchmark performance is 7 to 8 % better than the industry average; because the industry average includes an undocumented proportion of PIC genetics (see sections 2.5.1 and 4.3), these estimates are conservative.
4. Discussion
4.1. Boundaries within the Current LCA Environment
This study is an environmental lifecycle assessment (an E-LCA); it focuses on the undesirable biophysical impacts of its functional unit. As a complementary activity, UNEP (2009) formalized the socio-economic lifecycle assessment (the S-LCA), focusing on impacts on the well-being of stakeholders such as workers, consumers and the local community, among others. UNEP notices that such impacts may be desirable (for example when they lead to employment opportunity or to affordable food security) as well as undesirable. A complete lifecycle sustainability assessment of a production system would then involve an E-LCA, an S-LCA, and also a lifecycle costing exercise to quantify “the direct costs and benefits from economic activities”. Following Elkington (1998), the triple bottom line of a production system must quantify its impacts on the three pillars of sustainability: People, Planet and Profit. The latter pillar was euphemized to Prosperity in the 2002 World Summit on Sustainable Development (https://tinyurl.com/8ppnbe35); UNEP (2009) adopted this terminology. S-LCA focuses on the People and Prosperity pillars, E-LCA on the Planet pillar. As we argued previously (Knap, 2012, p. 7974), a complete assessment of a livestock production system should involve a fourth pillar in its quadruple bottom line, focusing on the interests of the animals; for pig or poultry production this would conveniently lead to a fourth P-labeled pillar, such as Pigs (we could also think of gimmicks such as Puminants or Phish). In terms of the present study, this would call for quantification of the impacts of pig breeding on pig welfare. Zira et al. (2022) modelled this in an elegantly integrated way by introducing pigs as an additional stakeholder in the One Health approach (One Health Commission, 2021) to assess the impacts of changes to (i) animal housing conditions and to (ii) the breeding goal on (iii) the environment, on (iv) people and on (v) pigs. A similar approach was followed by Tallentire et al. (2019) in an S-LCA of broiler chicken production. Zira assessed the impact of (ii) on (v) in terms of piglet mortality rate, tail biting incidence, and sow longevity. Unfortunately, changes to his items (i) and (ii) were confounded, so Zira’s study could not produce unequivocal conclusions in terms of (ii) breeding goals. Reflecting on his S-LCA-related work, Zira (2023, p. 86) concluded that this is “a daunting task if not herculean, requiring immense time. Trying to assess future livestock systems based on forecasts using scant current and historic data is also difficult”. Hence “more research is required to develop an integrated assessment of the social, economic and environmental impacts of food production from farmed animals. Such an evaluation should also include the farm’s sensitivity to economic changes and competition for arable land for feed or food”, and “many methods of sustainability assessment have so far focused on negative environmental effects because they are the easiest to measure” (Zira, 2023, p. 107). So, pending such “research to develop an integrated assessment”, we chose to restrict ourselves to the current E-LCA, noticing that measuring its effects credibly and defensibly is difficult enough.
4.2. Comparison to other Studies
The scientific literature holds many LCA studies of livestock production systems; reviews of such studies include McAuliffe et al. (2016), McClelland et al. (2018), and Pereira Silva et al. (2023). Pig-specific LCAs include the 74 studies cited by Gislason et al. (2023) plus, recently, Reckmann and Krieter (2015; Germany), Rougoor et al. (2015; Netherlands), Watson et al. (2018; Australia), Bonesmo and Gjerlaug-Enger (2021; Norway), Monteiro et al. (2021; France), Zira et al. (2021; Sweden), CIEL (2022; UK), Lamnatou et al. (2022; Spain), Shurson et al. (2022; USA), Savian et al. (2023; Brazil), and Yang et al. (2023; China). Andretta et al. (2021) and Pexas & Kyriazakis (2023) provide other reviews.
LCAs like our current study, specifically focused on the impact of livestock breeding, are much scarcer. A nice example outside pig breeding is Farrell et al. (2022) who conducted high-low sampling of sheep in Ireland, based on their estimated breeding values (EBVs) for the production traits that feature in the local profitability-oriented breeding goals. This was a top 20 % / bottom 20 % sampling scheme; hence assuming Normality, the sample means must have been 2.8 standard deviations apart. The LCA-based GHG emission intensities were 22.1 (high production EBVs) and 23.7 (low production EBVs) kg CO2eq per kg lamb carcass. This comes down to a correlated GHG reduction of 2.4 % per standard deviation of the breeding goals. Berry et al. (2022; their Figure 7) show that the 2008-2018 genetic improvement in these breeding goals was on average 0.048 standard deviations per year; hence the realized correlated genetic reduction of GHG emission in that period must have been around 0.12 % per year. An important detail here is that these sheep breeding goals do not include feed intake or feed efficiency – the traits with the strongest connection to GHG production.
De Haas et al. (2021) present case studies of the reduction of GHG emission through genetic change in poultry, pigs and cattle in the Netherlands. Their pig-specific case study contrasts two yearly cohorts (2014 and 2016) of growing pigs of a widespread commercial cross kept on a single experimental facility. The 2016 cohort showed a 1.2 to 1.5 % lower GHG emission than the 2014 cohort; given the uniform housing and nutritional conditions, this would suggest a correlated annual genetic reduction of GHG emission of 0.7 % of the mean level. This mean level was 180 kg CO2eq from 22 to 120 kg live weight, so the absolute annual reduction must have been 1.26 kg CO2eq per slaughter pig. Notice that this was not an LCA study but a biological trial.
Apart from that, we are aware of the following LCA studies that attempt to quantify such effects arising from pig breeding.
Bonesmo and Gjerlaug-Enger (2021) and Gjerlaug-Enger et al. (2022) conducted an LCA of the pig production sector in Norway. Based on the genetic improvement in feed efficiency, postweaning mortality and sow reproductive output that was realized during the year 2021, the correlated genetic reduction of GHG emission intensity in that year was reported as 1.4 % and 1.9 % of the mean level for the Norwegian Landrace and Duroc populations, respectively. This mean level was 186 kg CO2eq for a pig with 80 kg carcass weight, so the absolute annual reduction must have been 2.61-3.54 kg CO2eq per slaughter pig.
Knap et al. (2023) used the conversion factors described in our Appendix A in the Supplementary Material to derive a genetic reduction of GHG emission from the genetic improvement of the traits described there, as realized between 2012 and 2023 in the same commercial pig crosses as covered in the current study. The correlated annual reduction was estimated at 2.93 kg CO2eq per slaughter pig.
Alfonso (2019) derived conversion factors for sow productivity traits (mainly litter size and piglet mortality rate) with respect to GHG emission, similar to the ones described in our Appendix A in the Supplementary Material. As suggested by Amer et al. (2018), he added these to the trait weighting factors of his profitability-oriented breeding goal, assuming a carbon emission shadow price of 0.04 EUR per kg CO2eq. The impact of this addition was then quantified, and it was concluded “that no relevant changes are produced in the relative [weightings] of sow efficiency traits after the inclusion of GHG costs”. In other words, at that level of the carbon shadow price (which corresponds to the EU Emissions Trading System price of early 2021, see e.g. https://www.investing.com/commodities/carbon-emissions), inclusion of explicit GHG-related elements to a breeding goal that was designed without GHG emission in mind does not add much to the predicted GHG output of the production system.
As in the abovementioned sheep study of Farrell et al. (2022), Alfonso’s breeding goal did not include feed intake or feed efficiency traits, so his results are not surprising. By contrast, Ali et al. (2018) conducted a similar study with that same carbon shadow price but with a pig breeding goal based on litter size, piglet mortality, growth rate and, notably, feed efficiency. One simulated generation of selection on this profitability-oriented breeding goal produced a correlated reduction of the GHG emission per slaughter pig of 0.510 %; the addition of the carbon shadow price changed this to a reduction of 0.514 %. So again, at that level of the carbon shadow price, an addition of explicit GHG-related elements to a profitability-oriented breeding goal (designed without GHG emission in mind but this time including feed efficiency) does not add much to the predicted GHG output of the production system.
Ottosen (2021; his Chapter 5) calculated sets of trait weighting factors for breeding goals aimed at reducing the cost of pig production (i.e. profitability-oriented) or at reducing each of the ReCiPe-2016 impact categories (see our section 2.6 and our Figure 3). Each of these breeding goals was used in a deterministically simulated 10-generation pig breeding program with full specification of the genetic covariances among the traits, integrated into an LCA study to predict its impact on those same categories; in other words, the impact on each category was quantified for each category-oriented breeding goal, with the logical hypothesis that “a breeding goal designed to reduce one impact category will be substantially better at doing so with relatively smaller reductions in other impact categories”. However, their overall conclusion was that the various breeding goals “led to very similar reductions for each targeted impact category, with only minimal differences”. In more detail (derived from Ottosen’s Figure 5.12c), these 10-generation (i.e. about 15 years) reductions ranged from 17.9-18.3 % (for impact category fossil fuel depletion, across all the breeding goals) to 24.2-24.5 % (for impact category acidification, across all the breeding goals): a 6.2 % range across impact categories, and a 0.3-0.4 % range across breeding goals. Hence “the present [profitability-oriented] breeding programme performs well in reducing all investigated environmental impacts”.
In summary (see Table 2), and consistent with our forecast of section 3.2.1, current profitability-oriented pig breeding goals have been (and are still) reducing the GHG emission of pig production by about 1 % per year, dependent on local conditions and on the body weight trajectory studied.
Specific orientation of breeding goals to environmental impact categories does not significantly improve this reduction, at least not with the current carbon shadow prices. The 0.04 EUR per kg CO2eq shadow price used by Ali et al. (2018) and Alfonso (2019) can be usefully compared to the Emission-weighted Carbon Prices as realized in 2020 for the Agriculture & Forestry sectors in nine European countries, Argentina, Mexico and South Africa (Dolphin, 2022); these ranged more or less uniformly from 0.00032 USD per kg (Ukraine) to 0.034 USD per kg (Sweden) with a single extreme value at 0.07 USD per kg (Finland). From that point of view, Ali’s and Alfonso’s value seems very realistic. Those shadow prices were either explicitly set by a government through carbon taxation, or implicitly influenced by a government through the quota of CO2-equivalents that it makes available in an emissions trading system. A very different macro-economic approach was described by Boussemart et al. (2017) who essentially regressed the gross domestic product (GDP: national income) of 119 countries on their annual CO2 emission, annually from 1990 to 2011. The CO2 shadow price is then proportional to that regression coefficient, quantifying how much additional GDP is generated per additional unit of CO2 emitted: a higher regression coefficient indicates a “greener” country that manages to increase its GDP with a lower increase of its GHG emission. The results of that analysis are very variable across regions, ranging from 0.113 (China) to 10.2 (Africa) USD per kg in 2011; the important point here is that they are much higher than today’s politically imposed shadow prices that were shown above to be ineffective for pig breeding purposes.
The abovementioned percent-wise reductions of GHG emission also depend strongly on the calculation methods used: percentage values obviously depend on the base value that the genetic change is a percentage of , and in the literature quoted at the top of this section (ignoring suspicious outliers) these base values range from 2 to 5 kg CO2eq per kg liveweight. Many of these reported base values represent simulated averages across many farms, but Ruckli et al. (2021, their Table 4) give such results for 27 individual farms across six European countries, ranging from 1.9 to 5.1 kg CO2eq per kg liveweight. It follows that in this field, percent-wise differences or changes must be interpreted with considerable caution. Apart from that, the LCA methodology that was applied in those studies, and the way the results were reported, is by no means uniform and standardized; in their abovementioned review, Gislason et al. (2023) recommend that “the reader should refrain from making direct comparisons across different studies. This is due to differences in systems boundary settings, assumptions, methodologies, and varying transparency across the studies […], emission models, LCIA methods and background databases”. Indeed, in our study the CO2eq emission of a slaughter pig was estimated 48 % higher under ReCiPe-2016-H than under PEF-3.1.
4.3. Scenario 2: Baseline Comparison
Averaged across the LCIA frameworks of Figure 4, the global warming performance of PIC-2021 is 7.5 % lower than the 2021 North American industry average level. Considering that the industry average data contain an undocumented proportion of PIC animals which we believe to be between 30 and 50 % (see section 2.5.1), this result can be extrapolated to a 2021 global warming performance of PIC genetics that is 10 to 14 % lower than that of the non-PIC-derived part of the North American pig sector.
Given the way our results were calculated, the lower environmental impact of PIC genetics as compared to the North American industry average is due to differences in production traits such as feed intake, growth rate and survival rate of the growing-finishing pig, and feed intake and reproductive performance of the sow. Such differences between genetic populations are due to differences in genetic levels of the founder populations and in the subsequent rate of genetic change, which is determined by (i) the weighting of traits in the breeding goal and its associated selection index, (ii) the intensity of selection on that index at the GGP, GP and Parent levels, (iii) the accuracies of the estimated breeding values of the various traits, and (iv) the length of the generation interval at the GGP and GP levels (e.g. Bichard, 1971). Most pig breeding companies have productivity- and/or profitability-oriented selection programs (e.g. Knap, 2014), so element (i) should not differ much between companies. Element (ii) is largely determined by population size, element (iii) by population size, by the quantity and quality of data recording, and by the quality of statistical data processing, and element (iv) by farm logistics. Generally, configurations with levels of elements (ii) to (iv) that lead to a faster rate of genetic change are more costly. Differences in the realized progress between companies will then be mostly due to different levels of investment in technology and population structure, and to different operational efficiencies.
4.4. Extrapolation to 2030
Averaged across the LCIA frameworks of Figure 3, the predicted PIC-2030 global warming performance is 7.5 % lower than the PIC-2021 level. An obvious question is then how the predicted PIC-2030 levels would compare to predicted 2030 industry average values – not only for global warming but also for the other impact categories. The 2030 industry average values cannot be predicted with the semi-empirical approach of section 2.4 because the selection index weighting factors and the genetic covariances of breeding programs other than PIC’s are not available to us. Instead we fitted a logarithmic regression to the pre-2021 North American time trends of the relevant KPI traits (i.e. growth rate, feed intake, mortality rate, litter size etc.) as reported by Interpig (2021), MetaFarms (2021), PigCHAMP (2023) and AgriStats, and we extrapolated the fitted patterns to 2030. Background data and feed composition were kept unchanged. Appendix D in the Supplementary Material gives the KPI data used and the projections.
Figure 6 shows the results for the 14 functionally overlapping categories across the ReCiPe and PEF frameworks; the predicted PIC-2030 levels are 13 to 16 % more favorable than the predicted 2030 industry average values (14 % for the global warming performance). Notice that this comparison carries a high uncertainty level, for two reasons.
First, the prediction methods are different: the PIC-2030 performance is based on semi-deterministic prediction making use of known trait covariances and known trait weighting factors, whereas the 2030 industry average was predicted by extrapolation of historical trends.
Second, future selection strategies (with their differential weighting of traits) are likely to differ from past (for the industry average) and current (for PIC) strategies. This may be in response to changing costs or prices, or to externally imposed structural changes, e.g. in housing conditions as recently triggered in USA by the California ballot proposition 12 (https://en.wikipedia.org/wiki/2018_California_Proposition_12#) and by similar trends in Europe (e.g. EC, 2021). As we argued in section 4.3, most pig breeding companies have productivity- and/or profitability-oriented selection programs, and similar changes in market or production pressure should lead to similar decisions around future selection directions. Future differences in the realized progress between companies will then be mostly due, again, to different levels of investment in technology and population structure, and to different operational efficiencies. Also, novel technologies such as digital phenotyping (e.g. Neethirajan & Kemp, 2021; Tripodi et al., 2022; Liu et al., 2023), genome editing (e.g. Proudfoot et al., 2020) and semen sexing may change the dynamics substantially over and above the conventional effects of selection.
In the LCIA, this increased uncertainty is accounted for using the Ecoinvent pedigree matrix, as described in section 2.8.
4.5. Livestock Genetic Improvement
As we argued in Section 1, livestock breeding (i) increases farm animal productivity so that more animal protein (the desirable output) can be produced by fewer animals, using less land, water and other resources; it also (ii) increases the efficiency of that production which reduces the emission intensity of undesirable outputs such as greenhouse gases. Our results in sections 3.2.1 and 3.2.2, and the literature reviewed in section 4.2, provide sufficient illustration of point (ii). With regard to point (i), Figure 7 shows how increased cattle and pig productivity has reduced the worldwide standing populations of these species as they were required to cater for the production of a certain amount of animal protein. Across both species worldwide in 2020, this value was 46 % of the 1960 level.
It follows that increased livestock productivity and efficiency has been reducing the footprint intensity of animal protein for many decades; this is fortunate, for the worldwide demand for (and, hopefully, the associated production of) animal-sourced food increased fourfold from 1960 to 2020 and has been projected to increase fivefold from 1960 to 2040 (Bodirsky et al., 2018; their Figure 7). Hence the current discussion about the livestock footprint would have been much more pessimistic if the intensity reduction would not have taken place. Significantly, the contribution to this trend made by livestock breeding is cumulative and permanent, in contrast to interventions such as feed additives.
4.6. Future Work
The model developed here can be (and will be) used to assess such trends in other regions than North America and can also be adapted to cater for other production systems such as nursery-finisher (rather than farrow-to-finish as in the current study) or, in a different dimension, Parma or Ibérico (Italy, Spain).
5.Conclusions
Current pig breeding programs have been delivering positive environmental outcomes as a correlated response to selection for the (re)production traits that feature in the conventional profitability-oriented breeding goals. This trend will continue and is likely to become stronger due to technology development.
Our LCA study is the first one to use long-term genetic trends in pig breeding, and to forecast the mitigation of environmental impacts. The forecast predicts, for PIC-derived slaughter pigs in North America, a statistically significant genetic reduction of twelve impact factors including the global warming potential, each by 0.7 to 1 % per year up to 2030.
Such trends may also become stronger due to shifts of the profitability-oriented breeding goals towards more focus on the traits that are most strongly related to environmental impact categories, but this will require considerably more focus than what current carbon shadow prices would dictate. This is largely a political issue.
Due to different levels of investment in technology and in the structure of breeding populations, and due to different operational efficiencies of North American pig breeding companies, the current (2022) status quo is that PIC genetics have a 7 to 8 % lower impact on most environmental impact categories than the North American industry average.
Supplementary Material
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: primary data required. Table S2: sow diets. Table S3: G/F diets. Appendix A: FCRwe. Appendix B: inventory and pedigree matrixes. Appendix C: scenario 1 and 2 results. Appendix D: extrapolation to 2030.
Author contributions
Greg J Thoma: Methodology, Software, Formal analysis, Validation, Writing - Review & Editing. Banks Baker: Conceptualization, Supervision, Funding acquisition, Resources, Writing - Review & Editing. Pieter W Knap: Validation, Visualization, Writing - Original Draft
Funding
This study was supported by Genus-PIC, Hendersonville TN, USA
Data availability
The datasets presented in this article are not readily available because they are commercially sensitive. Requests to access the datasets should be directed to pieter.knap@genusplc.com.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgements
Sarah Frank, Garth Boyd and Matt Sutton-Vermeulen of the Context Network llc ensured the coordinated progress of this study. Our PIC colleagues Lindsay Case, Steve Dritz, Dan Hamilton, Craig Lewis and Lucina Galina-Pantoja provided data, discussion and critical reviews.
Conflicts of interest
Greg J. Thoma was contracted by Genus-PIC (Hendersonville TN, USA) to carry out this research. The other authors are employees of Genus-PIC.
Abbreviations
Abbreviations used in this text. ANOVA: analysis of variance. APOS: allocation at point of substitution. CO2: carbon dioxide. EBV: estimated breeding value. …-eq: … equivalents. E-LCA: environmental LCA. EUR: Euro (the currency). FCR: feed conversion ratio (kg feed / kg gain). FCRwe: whole-enterprise FCR. GDP: gross domestic product. GHG: greenhouse gas. GGP: great-grandparent. GP: grandparent. GWP: global warming potential. H+: hydrogen ion. IPCC: Intergovernmental Panel on Climate Change. ISO: International Organization for Standardization. KPI: key performance indicator. LCA: lifecycle assessment. LCI: lifecycle inventory. LCIA: lifecycle impact assessment. LEAP: livestock environmental assessment and performance. MCS: Monte Carlo simulation. PEF: product environmental footprint. PIC: Pig Improvement Company. R&D: research and development. S-LCA: social LCA. SO2: sulphur dioxide. USD: United States Dollar.
References
- Alfonso, L. Impact of incorporating greenhouse gas emission intensities in selection indexes for sow productivity traits. Livest. Sci. 2019, 219, 57–61. [Google Scholar] [CrossRef]
- Ali BM, Bastiaansen JWM, De Mey Y, Oude Lansink AGJM (2018) Effects of incorporating environmental cost and risk aversion on economic values of pig breeding goal traits. J. Anim. Breed. Genet. 135:194-207.
- Amer, P.; Hely, F.; Quinton, C.; Cromie, A. A methodology framework for weighting genetic traits that impact greenhouse gas emission intensities in selection indexes. Animal 2018, 12, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Andretta, I.; Hickmann, F.M.W.; Remus, A.; Franceschi, C.H.; Mariani, A.B.; Orso, C.; Kipper, M.; Létourneau-Montminy, M.-P.; Pomar, C. Environmental Impacts of Pig and Poultry Production: Insights From a Systematic Review. Front. Veter- Sci. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.; Dunne, F.; McHugh, N.; McParland, S.; O’brien, A.; Twomey, A. The development of effective ruminant breeding programmes in Ireland from science to practice. Ir. J. Agric. Food Res. 2022, 61, 38–54. [Google Scholar] [CrossRef]
- Bichard, M. Dissemination of genetic improvement through a livestock industry. Anim. Sci. 1971, 13, 401–411. [Google Scholar] [CrossRef]
- Bidanel JP (2011) Biology and genetics of reproduction. In: The genetics of the pig, 2nd edition (eds. Rothschild MF, Ruvinsky A), pp. 218-241. CABI, Wallingford, UK. ISBN 9781845937560.
- Bodirsky, B.L.; Rolinski, S.; Biewald, A.; Weindl, I.; Popp, A.; Lotze-Campen, H. Global Food Demand Scenarios for the 21st Century. PLOS ONE 2015, 10, e0139201. [Google Scholar] [CrossRef]
- Bonesmo, H.; Enger, E.G. The effects of progress in genetics and management on intensities of greenhouse gas emissions from Norwegian pork production. Livest. Sci. 2021, 254, 104746. [Google Scholar] [CrossRef]
- Boussemart, J.-P.; Leleu, H.; Shen, Z. Worldwide carbon shadow prices during 1990–2011. Energy Policy 2017, 109, 288–296. [Google Scholar] [CrossRef]
- CIEL (2022) Net Zero and livestock: how farmers can reduce emissions. Centre for Innovation & Excellence in Livestock, York, UK. https://cielivestock.co.uk/wp-content/uploads/2022/02/CIEL-LR-220405.pdf.
- Ciroth, A.; Muller, S.; Weidema, B.; Lesage, P. Empirically based uncertainty factors for the pedigree matrix in ecoinvent. Int. J. Life Cycle Assess. 2016, 21, 1338–1348. [Google Scholar] [CrossRef]
- Clutter AC (2011) Genetics of performance traits. In: The genetics of the pig, 2nd edition (eds. Rothschild MF, Ruvinsky A), pp. 325-354. CABI, Wallingford, UK. ISBN 9781845937560.
- De Haas Y, Bink MCAM, Borg R, Koenen EPC, Verschuren LMG, Mollenhorst H (2021) The contribution of animal breeding to reducing the environmental impact of livestock production. Wageningen Livestock Research Report 1156, Wageningen, Netherlands. [CrossRef]
- de Vries, M.; de Boer, I. Comparing environmental impacts for livestock products: A review of life cycle assessments. Livest. Sci. 2010, 128, 1–11. [Google Scholar] [CrossRef]
- Dolphin, G (2022) World carbon pricing database. Resources for the Future, Washington DC, USA. https://www.rff.org/publications/data-tools/world-carbon-pricing-database.
- Dyllick, T.; Hockerts, K. Beyond the business case for corporate sustainability. Bus. Strategy Environ. 2002, 11, 130–141. [Google Scholar] [CrossRef]
- EC (2021) Communication from the Commission on the European Citizens' Initiative (ECI) "End the Cage Age". European Commission, Brussels, Belgium. https://ec.europa.eu/transparency/documents-register/detail?ref=C(2021)4747&lang=en.
- Ewaoluwagbemiga, E.O.; Bee, G.; Kasper, C. Genetic analysis of protein efficiency and its association with performance and meat quality traits under a protein-restricted diet. Genet. Sel. Evol. 2023, 55, 1–16. [Google Scholar] [CrossRef]
- Farrell, L.; McHugh, N.; Pabiou, T.; Bohan, A.; McDermott, K.; Shalloo, L.; Herron, J. 716. Modelling of productivity, profitability, Rotterdam, Netherlands, and greenhouse gas emissions of Irish sheep flocks divergent in genetic merit. In Proceedings of 12th World Congress on Genetics Applied to Livestock Production; pp. 2952–2955.
- Vivanco, D.F. The role of services and capital in footprint modelling. Int. J. Life Cycle Assess. 2019, 25, 280–293. [Google Scholar] [CrossRef]
- Genus (2021) Annual Report 2021. Genus plc, Basingstoke, UK. https://www.genusplc.com/media/1875/genus-plc-annual-report-2021.pdf.
- Genus (2022) Preliminary results year ended 30 June 2022. Genus plc, Basingstoke, UK. https://www.genusplc.com/media/2011/genus_fy22-full-results-final.pdf.
- Gislason, S.; Birkved, M.; Maresca, A. A systematic literature review of life cycle assessments on primary pig production: Impacts, comparisons, and mitigation areas. Sustain. Prod. Consum. 2023, 42, 44–62. [Google Scholar] [CrossRef]
- Enger, E.G.; Bonesmo, H.; Martinsen, K.; Andersen-Ranberg, I.; Olsen, D.; Grindflek, E. 17. The genetic progress has effect on greenhouse gas emission intensities from Norwegian pork production. In Proceedings of the World Congress on Genetics Applied to Livestock Production, Rotterdam, Netherlands; pp. 115–118.
- Hendrickson CT, Lave LB, Matthews HS (2006) Environmental Life Cycle Assessment of goods and services: an Input-Output approach. RFF Press, Washington DC, USA. ISBN 978-1933115246.
- Huijbregts, M.A.J.; Steinmann, Z.J.N.; Elshout, P.M.F.; Stam, G.; Verones, F.; Vieira, M.D.M.; Hollander, A.; Zijp, M.; Van Zelm, R. (2016) ReCiPe 2016 v1.1 A Harmonized Life Cycle Impact Assessment Method at Midpoint and Endpoint Level Report I: Characterization; Available online: https://www.rivm.nl/bibliotheek/rapporten/2016-0104.pdf; National Institute for Public Health and the Environment: Bilthoven, The Netherlands, 2017. [Google Scholar]
- Huijbregts, M.A.J.; Steinmann, Z.J.N.; Elshout, P.M.F.; Stam, G.; Verones, F.; Vieira, M.; Zijp, M.; Hollander, A.; van Zelm, R. ReCiPe2016: A harmonised life cycle impact assessment method at midpoint and endpoint level. Int. J. Life Cycle Assess. 2017, 22, 138–147. [Google Scholar] [CrossRef]
- Interpig (2021) 2021 pig cost of production in selected countries. AHDB, Kenilworth, UK. https://projectblue.blob.core.windows.net/media/Default/Pork/Pork%20MI%20files/CostPigProduction2021_221117_WEB.pdf.
- ISO (2006a) ISO 14040: environmental management life cycle assessment: principles and framework. International Organization for Standardization, Geneva, Switzerland. https://www.iso.org/obp/ui/en/#iso:std:iso:14040:ed-2:v1:en.
- ISO (2006b) ISO 14044: environmental management life cycle assessment: requirements and guidelines. International Organization for Standardization, Geneva, Switzerland. https://www.iso.org/obp/ui/en/#iso:std:iso:14044:ed-1:v1:en.
- Kasper C (2021) Genetic basis of protein efficiency in pigs. Visite étudiantes AgroCampus Ouest à Agroscope-Agridea, Posieux, Switzerland. http://tinyurl.com/355k58fk.
- Knap PW (2012) Pig breeding for increased sustainability. In: Encyclopedia of sustainability science and technology, 1st edition (ed. Meyers RA), volume 11, pp. 7972-8012. Springer Science+Business Media LLC, New York NY, USA. ISBN 978-0387894690.
- Knap PW (2014) Pig breeding goals in competitive markets. 10th WCGALP, Vancouver, Canada; 007. http://www.wcgalp.org/proceedings/2014/pig-breeding-goals-competitive-markets.
- Knap PW, Olson KM, Cleveland MA (2023) Breeding livestock for sustainable systems. Abstracts of 74th annual meeting of the European Federation of Animal Science, Lyon, France; p. 157. https://members.eaap.org/wp-content/uploads/2023/theater/S02T06.pdf.
- Lamnatou, C.; Ezcurra-Ciaurriz, X.; Chemisana, D.; Plà-Aragonés, L. Life Cycle Assessment (LCA) of a food-production system in Spain: Iberian ham based on an extensive system. Sci. Total. Environ. 2022, 808, 151900. [Google Scholar] [CrossRef]
- Legarra, A.; Christensen, O.F.; Aguilar, I.; Misztal, I. Single Step, a general approach for genomic selection. Livest. Sci. 2014, 166, 54–65. [Google Scholar] [CrossRef]
- Liu, D.; Parmiggiani, A.; Psota, E.; Fitzgerald, R.; Norton, T. Where's your head at? Detecting the orientation and position of pigs with rotated bounding boxes. Comput. Electron. Agric. 2023, 212. [Google Scholar] [CrossRef]
- MacLeod M, Leinonen I, Wall E, Houdijk J, Eory V, Burns J, Ahmadi BV, Gómez-Barbero M (2019) Impact of animal breeding on GHG emissions and farm economics. Publications Office of the European Union, Luxembourg, report EUR-29844-EN. [CrossRef]
- McAuliffe, G.A.; Chapman, D.V.; Sage, C.L. A thematic review of life cycle assessment (LCA) applied to pig production. Environ. Impact Assess. Rev. 2016, 56, 12–22. [Google Scholar] [CrossRef]
- McClelland, S.; Arndt, C.; Gordon, D.; Thoma, G. Type and number of environmental impact categories used in livestock life cycle assessment: A systematic review. Livest. Sci. 2018, 209, 39–45. [Google Scholar] [CrossRef]
- Beltran, A.M.; Prado, V.; Vivanco, D.F.; Henriksson, P.J.G.; Guinée, J.B.; Heijungs, R. Quantified Uncertainties in Comparative Life Cycle Assessment: What Can Be Concluded? Environ. Sci. Technol. 2018, 52, 2152–2161. [Google Scholar] [CrossRef] [PubMed]
- MetaFarms (2021) Production analysis summary for, U.S. pork industry: 2017-2021. National Pork Board, Des Moines IO, USA. https://porkcheckoff.org/wp-content/uploads/2022/05/MetaFarms-2021-Production-Analysis-for-NPB.pdf.
- Monteiro, A.N.T.R.; Brossard, L.; Gilbert, H.; Dourmad, J.-Y. Environmental Impacts and Their Association With Performance and Excretion Traits in Growing Pigs. Front. Veter- Sci. 2021, 8. [Google Scholar] [CrossRef]
- Myhre, G.; Shindell, D.; Bréon, F.-M.; Collins, W.; Fuglestvedt, J.; Huang, J.; Koch, D.; Lamarque, J.-F.; Lee, D.; Mendoza, B.; et al. Anthropogenic and Natural Radiative Forcing. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 659–740. [Google Scholar]
- Neethirajan, S.; Kemp, B. Digital Phenotyping in Livestock Farming. Animals 2021, 11, 2009. [Google Scholar] [CrossRef]
- One Health Commission (2021) World health through collaboration. https://www.onehealthcommission.org.
- Ottosen M (2021) Integrating Life Cycle Assessment with genetic selection to reduce the environmental impacts of pig production. PhD thesis, Newcastle University, UK. http://theses.ncl.ac.uk/jspui/handle/10443/5514.
- Silva, J.P.; Giehl, C.J.; Cardinal, K.M.; Andretta, I.; Ribeiro, A.M.L. Different life cycle assessment methods and causes of variation in estimates of global warming in chicken and pork production systems: A critical systematic review. Livest. Sci. 2023, 276. [Google Scholar] [CrossRef]
- Pexas, G.; Kyriazakis, I. Hotspots and bottlenecks for the enhancement of the environmental sustainability of pig systems, with emphasis on European pig systems. Porc. Heal. Manag. 2023, 9, 1–23. [Google Scholar] [CrossRef] [PubMed]
- PigCHAMP (2023) Benchmarking summaries. PigCHAMP, Ames IO, USA. https://www.pigchamp.com/benchmarking/benchmarking-summaries.
- PIC (2021) PIC nutrition and feeding guidelines. Pig Improvement Company, Hendersonville TN, USA. www.pic.com/resources/pic-nutrition-and-feeding-guidelines-metric.
- Proudfoot, C.; McFarlane, G.; Whitelaw, B.; Lillico, S. Livestock breeding for the 21st century: the promise of the editing revolution. Front. Agric. Sci. Eng. 2020, 7, 129–135. [Google Scholar] [CrossRef]
- Reckmann, K.; Krieter, J. Environmental impacts of the pork supply chain with regard to farm performance. J. Agric. Sci. 2014, 153, 411–421. [Google Scholar] [CrossRef]
- Reisinger, A.; Clark, H.; Cowie, A.L.; Emmet-Booth, J.; Fischer, C.G.; Herrero, M.; Howden, M.; Leahy, S. How necessary and feasible are reductions of methane emissions from livestock to support stringent temperature goals? Philos. Trans. R. Soc. A: Math. Phys. Eng. Sci. 2021, 379, 20200452. [Google Scholar] [CrossRef]
- Rougoor C, Elfrink E, Lap T, Balkema A (2015) LCA of Dutch pork: assessment of three pork production systems in the Netherlands. CLM Onderzoek en Advies, Culemborg, Netherlands. https://www.clm.nl/wp-content/uploads/2021/09/glamur-wp3-dutch-pork.pdf.
- Ruckli, A.K.; Dippel, S.; Durec, N.; Gebska, M.; Guy, J.; Helmerichs, J.; Leeb, C.; Vermeer, H.; Hörtenhuber, S. Environmental Sustainability Assessment of Pig Farms in Selected European Countries: Combining LCA and Key Performance Indicators for Biodiversity Assessment. Sustainability 2021, 13, 11230. [Google Scholar] [CrossRef]
- Saintilan R, Mérour I, Brossard L, Tribout T, Dourmad JY, Sellier P, Bidanel J, Van Milgen J, Gilbert H (2013) Genetics of residual feed intake in growing pigs: relationships with production traits, and nitrogen and phosphorus excretion traits. Journal of Animal Science 91:2542-2554. [CrossRef]
- Sandefur HN, Burek J, Matlock M, Thoma G, Boles EC (2015) Development of Life Cycle Inventory data for U.S. swine production scenarios: dataset documentation and user’s guide version 2. Center for Agricultural and Rural Sustainability, University of Arkansas, Little Rock AR, USA. https://tinyurl.com/mrycpdyk.
- Savian, M.; Simon, C.d.P.; Holden, N.M. Evaluating environmental, economic, and social aspects of an intensive pig production farm in the south of Brazil: a case study. Int. J. Life Cycle Assess. 2023, 28, 1544–1560. [Google Scholar] [CrossRef]
- Schneeberger, M.; Barwick, S.A.; Crow, G.H.; Hammond, K. Economic indices using breeding values predicted by BLUP. J. Anim. Breed. Genet. 1992, 109, 180–187. [Google Scholar] [CrossRef]
- Searle, S.R. The matrix Handling of BLUE and BLUP in the mixed linear model. Linear Algebra its Appl. 1997, 264, 291–311. [Google Scholar] [CrossRef]
- Shirali, M.; Doeschl-Wilson, A.; Knap, P.; Duthie, C.; Kanis, E.; van Arendonk, J.; Roehe, R. Nitrogen excretion at different stages of growth and its association with production traits in growing pigs1. J. Anim. Sci. 2012, 90, 1756–1765. [Google Scholar] [CrossRef] [PubMed]
- Shurson, G.C.; E O Pelton, R.; Yang, Z.; E Urriola, P.; Schmitt, J. Environmental impacts of eco-nutrition swine feeding programs in spatially explicit geographic regions of the United States. J. Anim. Sci. 2022, 100. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, T.; Gilbert, H. Evaluating environmental impacts of selection for residual feed intake in pigs. Animal 2020, 14, 2598–2608. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, T.; Gilbert, H. An approach to achieve overall farm feed efficiency in pig production: environmental evaluation through individual life cycle assessment. Int. J. Life Cycle Assess. 2021, 26, 455–469. [Google Scholar] [CrossRef]
- Tallentire, C.W.; Edwards, S.A.; Van Limbergen, T.; Kyriazakis, I. The challenge of incorporating animal welfare in a social life cycle assessment model of European chicken production. Int. J. Life Cycle Assess. 2019, 24, 1093–1104. [Google Scholar] [CrossRef]
- Thoma GJ (2023) Environmental sustainability evaluation of PIC genetics vs. industry average: North America. Resilience Services pllc, Denver CO, USA. https://tinyurl.com/wtrczjm7.
- Tripodi, P.; Nicastro, N.; Pane, C. Digital applications and artificial intelligence in agriculture toward next-generation plant phenotyping. Crop. Pasture Sci. 2022, 74, 597–614. [Google Scholar] [CrossRef]
- UNEP (2009) Guidelines for social life cycle assessment of products. United Nations Environment Programme, Paris, France. https://www.unep.org/resources/report/guidelines-social-life-cycle-assessment-products.
- van der Peet-Schwering, C.; Jongbloed, A.; Aarnink, A. Nitrogen and phosphorus consumption, utilisation and losses in pig production: The Netherlands. Livest. Prod. Sci. 1999, 58, 213–224. [Google Scholar] [CrossRef]
- van der Werf, H.M.G.; Knudsen, M.T.; Cederberg, C. Towards better representation of organic agriculture in life cycle assessment. Nat. Sustain. 2020, 3, 419–425. [Google Scholar] [CrossRef]
- Van Middelaar CE, Van Zanten HHE, De Boer IJM (2019) Future of animal nutrition: the role of life cycle assessment. In: Poultry and pig nutrition: challenges of the 21st century (eds. Hendriks WH, Verstegen MWA, Babinszky L), pp 307-314. WAP, Wageningen, Netherlands. https://www.wageningenacademic.com/doi/pdf/10.3920/978-90-8686-884-1_14.
- Van Paassen M, Braconi N, Kuling L, Durlinger B, Gual P (2019a) Agri-Footprint 5.0 part 1: methodology and basic principles. Blonk Consultants, Gouda, Netherlands. https://tinyurl.com/47h4a6zc.
- Van Paassen M, Braconi N, Kuling L, Durlinger B, Gual P (2019b) Agri-Footprint 5.0 part 2: description of data. Blonk Consultants, Gouda, Netherlands. https://tinyurl.com/y76cssry.
- Watson K, Wiedemann S, Biggs L, McGahan E (2018) Trends in environmental impacts from the pork industry. Integrity Ag and Environment, Highfields QLD, Australia. https://australianpork.com.au/sites/default/files/2021-07/2017-2212.pdf.
- Weidema BP, Bauer C, Hischier R, Mutel C, Nemecek T, Reinhard J, Vadenbo CO, Wernet G (2013) Overview and methodology: data quality guideline for the ecoinvent database version 3. Ecoinvent Report 1(v3), The Ecoinvent Centre, St. Gallen, Switzerland. https://tinyurl.com/5bhe48ak.
- Wernet, G.; Bauer, C.; Steubing, B.; Reinhard, J.; Moreno-Ruiz, E.; Weidema, B. The ecoinvent database version 3 (part I): overview and methodology. Int. J. Life Cycle Assess. 2016, 21, 1218–1230. [Google Scholar] [CrossRef]
- Yang, P.; Yu, M.; Ma, X.; Deng, D. Carbon Footprint of the Pork Product Chain and Recent Advancements in Mitigation Strategies. Foods 2023, 12, 4203. [Google Scholar] [CrossRef] [PubMed]
- Zira S (2023) Towards sustainable livestock systems: developing and applying methods for broad sustainability assessment of pig and cattle systems. PhD thesis, Swedish University of Agricultural Sciences, Uppsala. https://pub.epsilon.slu.se/28531/1/zira-s-20220824.pdf.
- Zira, S.; Röös, E.; Ivarsson, E.; Friman, J.; Møller, H.; Samsonstuen, S.; Olsen, H.; Rydhmer, L. An assessment of scenarios for future pig production using a One Health approach. Livest. Sci. 2022, 260, 104929. [Google Scholar] [CrossRef]
Figure 1.
Top: the cradle-to-farmgate system boundary. “GGP & GP” sows are the great-grandparents and grandparents of the slaughter pigs, respectively. Bottom: the foreground system, showing animal flows (blue solid lines: the main product; red solid lines: coproducts; blue dashed lines: losses) and accounting flows for the allocation avoidance algorithm (green dashed lines).
Figure 1.
Top: the cradle-to-farmgate system boundary. “GGP & GP” sows are the great-grandparents and grandparents of the slaughter pigs, respectively. Bottom: the foreground system, showing animal flows (blue solid lines: the main product; red solid lines: coproducts; blue dashed lines: losses) and accounting flows for the allocation avoidance algorithm (green dashed lines).
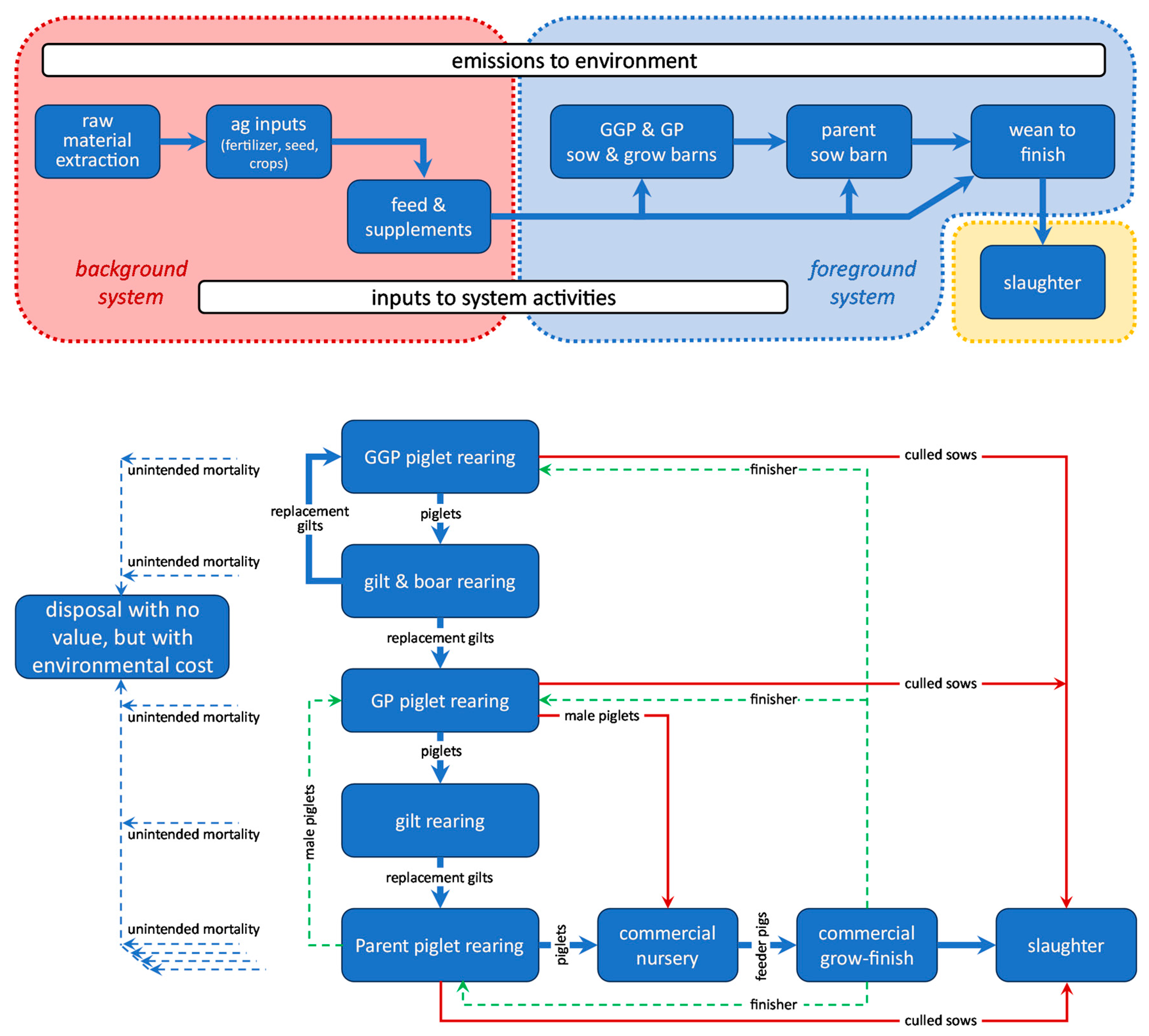
Figure 2.
Realized genetic trends of the traits that influence a farm’s whole-enterprise feed conversion ration FCRwe. The datapoints are annual within-line averages of the trait EBVs in two purebred sire lines (solid white trendlines) and two purebred dam lines (dashed white trendlines). Up to 2022 (vertical reference line), the black datapoints and trendlines represent the average realized performance of the resulting cross, i.e. the typical PIC-derived slaughter pig or parent sow. After 2022, the black trendlines represent the future performance as predicted by selection index theory (see section 2.4; for postweaning mortality, this prediction reaches unrealistic values after 2025 and these were capped). The trendlines are LOESS interpolations. The EBVs on the y-axes are deviations from arbitrary means that were forced to zero for 2022.
Figure 2.
Realized genetic trends of the traits that influence a farm’s whole-enterprise feed conversion ration FCRwe. The datapoints are annual within-line averages of the trait EBVs in two purebred sire lines (solid white trendlines) and two purebred dam lines (dashed white trendlines). Up to 2022 (vertical reference line), the black datapoints and trendlines represent the average realized performance of the resulting cross, i.e. the typical PIC-derived slaughter pig or parent sow. After 2022, the black trendlines represent the future performance as predicted by selection index theory (see section 2.4; for postweaning mortality, this prediction reaches unrealistic values after 2025 and these were capped). The trendlines are LOESS interpolations. The EBVs on the y-axes are deviations from arbitrary means that were forced to zero for 2022.
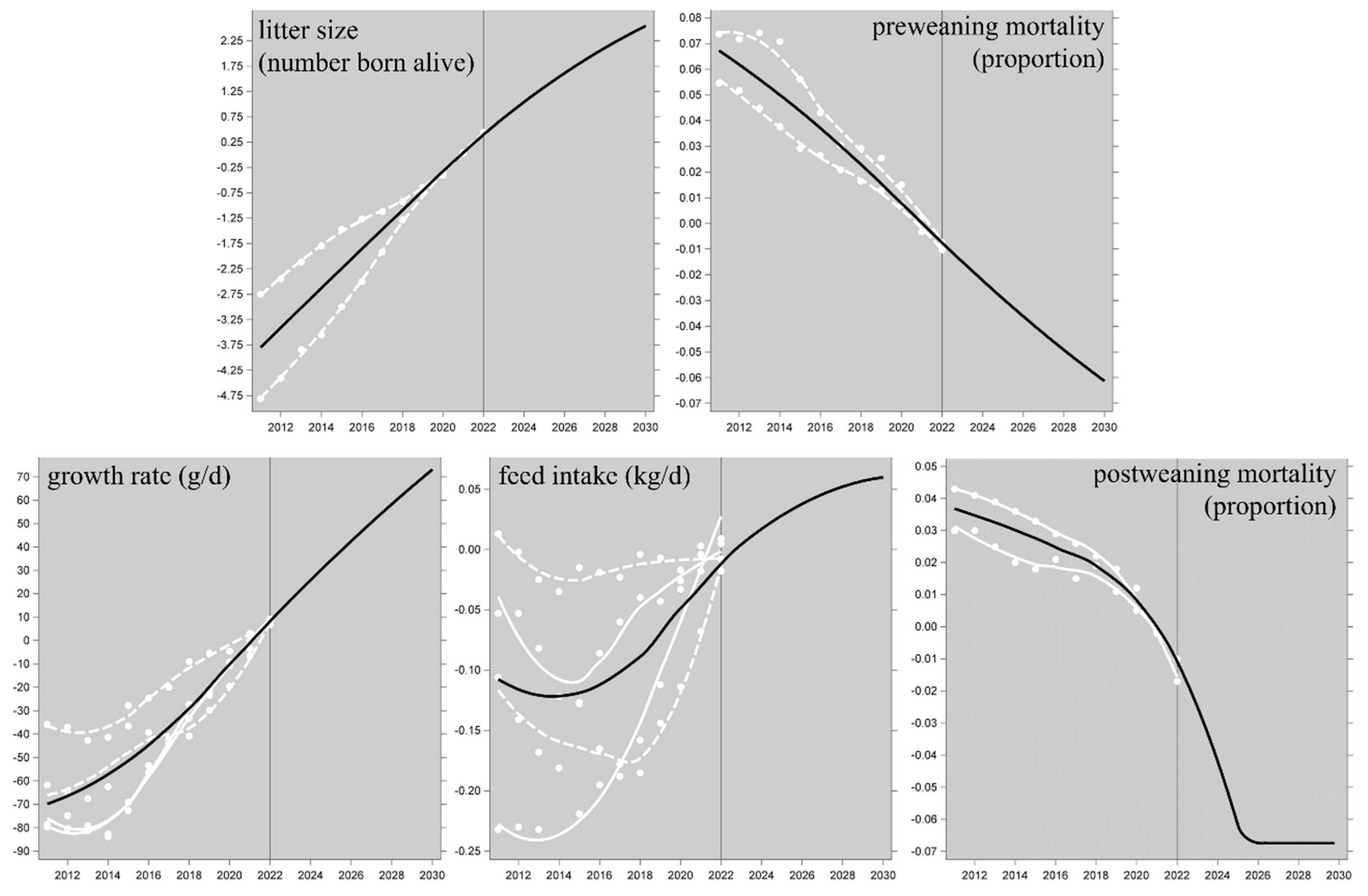
Figure 3.
Results for scenario 1: predicted performance of PIC genetics in 2030 as a proportion of the 2021 performance for 14 environmental impact categories, as modelled in ten LCIA frameworks (ReCiPe-2016 Egalitarian, Hierarchist and Individualist, cutoff and APOS; PEF-3.1 cutoff; IPCC-2021 cutoff and APOS). The IPCC cases for global warming (blue bars) are IPCC’s GWP-100 category; IPCC does not consider the other impact categories. Lower values are more favourable; the asterisks below the bars indicate cases where the predicted 2030 proportions are significantly (P<0.05) different from 1 (i.e. from the 2021 level).
Figure 3.
Results for scenario 1: predicted performance of PIC genetics in 2030 as a proportion of the 2021 performance for 14 environmental impact categories, as modelled in ten LCIA frameworks (ReCiPe-2016 Egalitarian, Hierarchist and Individualist, cutoff and APOS; PEF-3.1 cutoff; IPCC-2021 cutoff and APOS). The IPCC cases for global warming (blue bars) are IPCC’s GWP-100 category; IPCC does not consider the other impact categories. Lower values are more favourable; the asterisks below the bars indicate cases where the predicted 2030 proportions are significantly (P<0.05) different from 1 (i.e. from the 2021 level).
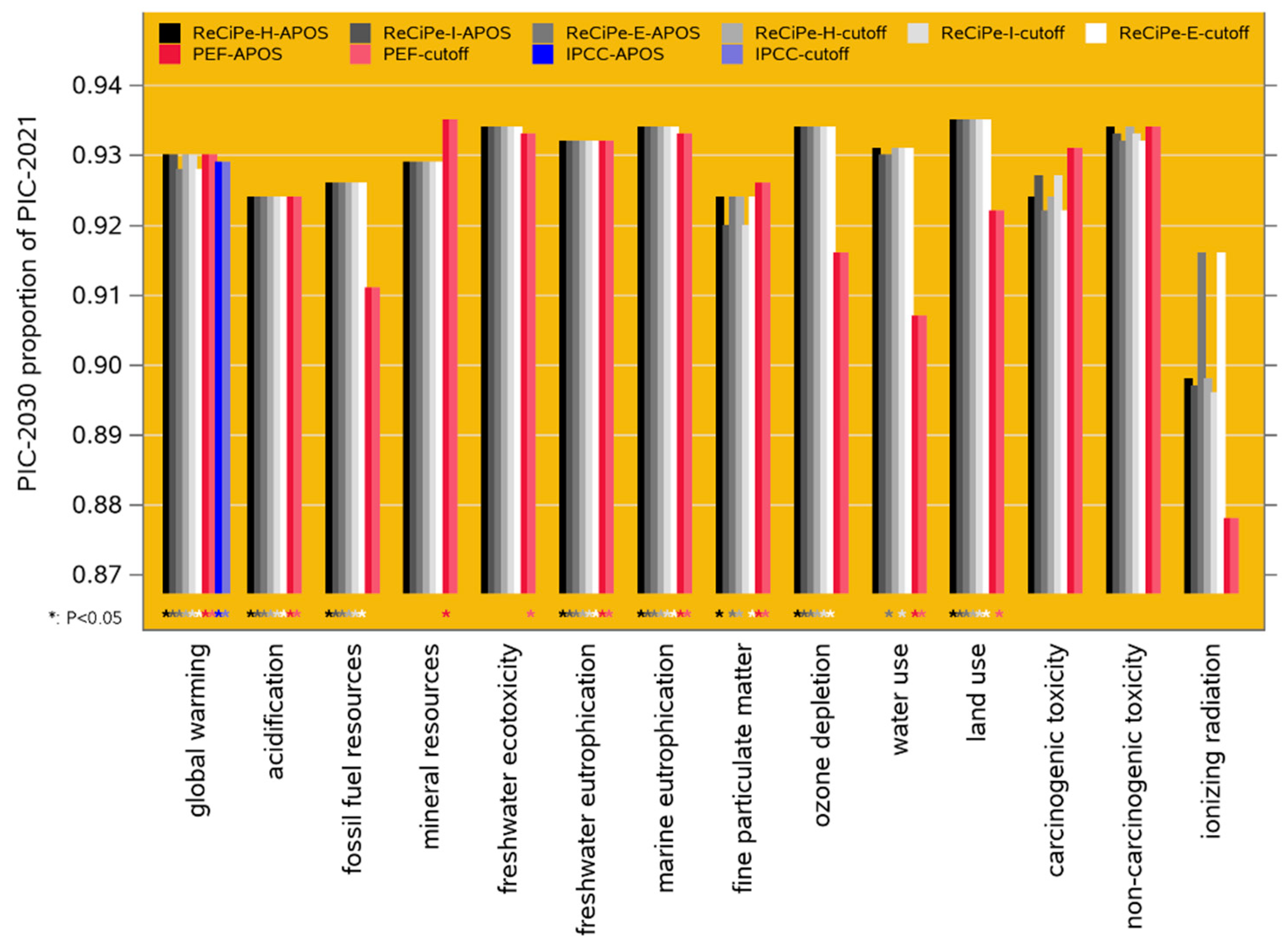
Figure 4.
Contribution analysis for the ReCiPe-2016 Hierarchist cutoff and PEF-3.1 cutoff LCIA frameworks. Note that the category axis is sorted differently than in Figure 3.
Figure 4.
Contribution analysis for the ReCiPe-2016 Hierarchist cutoff and PEF-3.1 cutoff LCIA frameworks. Note that the category axis is sorted differently than in Figure 3.
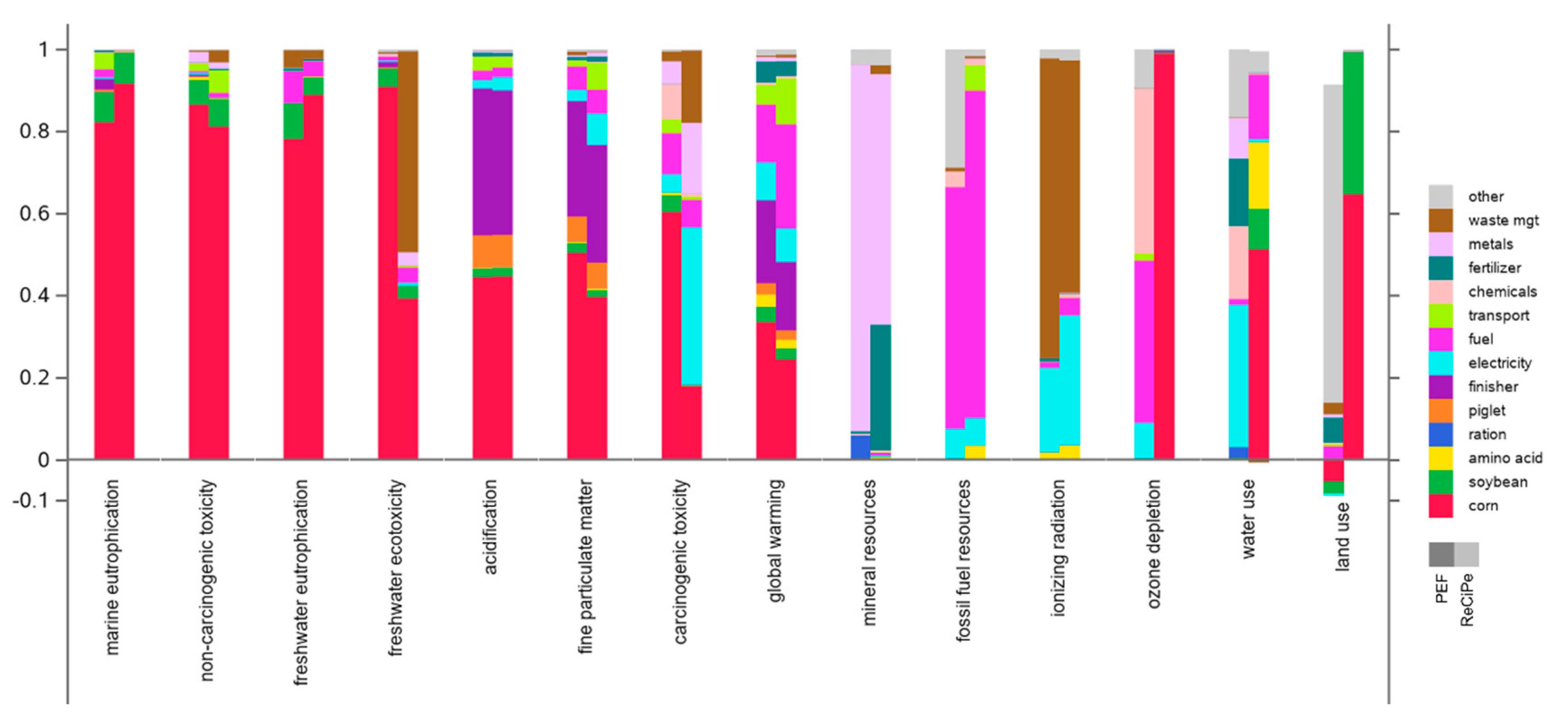
Figure 5.
Results for scenario 2: performance of PIC genetics as a proportion of the North American industry average (both in 2021) for 14 environmental impact categories, as modelled in ten LCIA frameworks. Lower values are more favorable; the asterisks below the bars indicate cases where the PIC proportions are significantly (P<0.05) different from 1 (i.e. from the industry average). The color coding and further formatting is the same as in Figure 3.
Figure 5.
Results for scenario 2: performance of PIC genetics as a proportion of the North American industry average (both in 2021) for 14 environmental impact categories, as modelled in ten LCIA frameworks. Lower values are more favorable; the asterisks below the bars indicate cases where the PIC proportions are significantly (P<0.05) different from 1 (i.e. from the industry average). The color coding and further formatting is the same as in Figure 3.
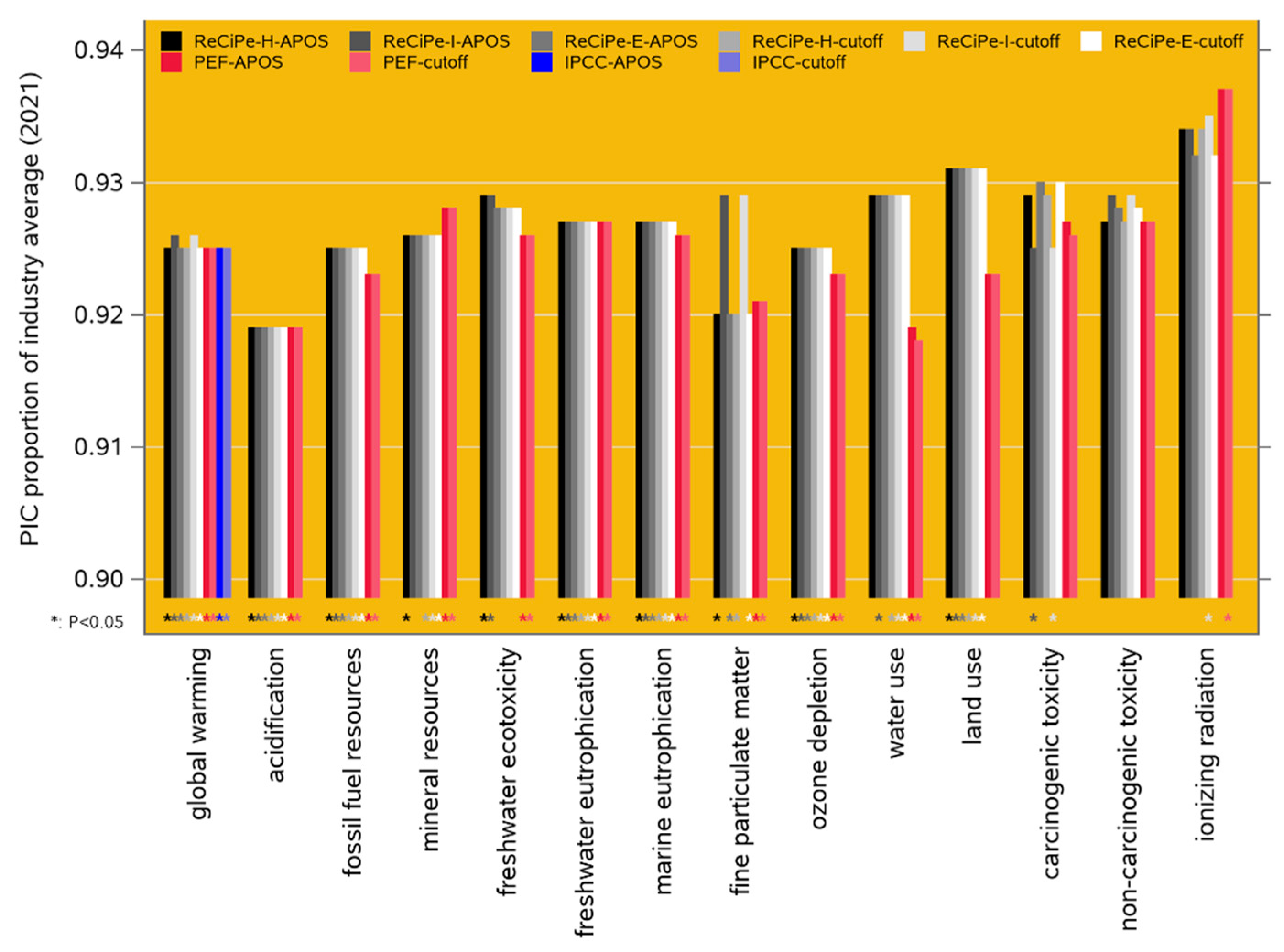
Figure 6.
Predicted performance of PIC genetics in 2030 as a proportion of the predicted North American industry average in 2030. See Figure 3 for details.
Figure 6.
Predicted performance of PIC genetics in 2030 as a proportion of the predicted North American industry average in 2030. See Figure 3 for details.

Figure 7.
Time trends of the standing population size of cattle and pigs associated with the production of 1000 kg animal protein, by continent. The percentages in the continent labels represent the proportion of the 2020 global populations. Data from https://www.fao.org/faostat/en/#data, assuming 3.5 % protein in milk and 16 % protein in meat; note that this will underestimate the earlier y-values for beef and pork due to higher fat content, especially in Asia.
Figure 7.
Time trends of the standing population size of cattle and pigs associated with the production of 1000 kg animal protein, by continent. The percentages in the continent labels represent the proportion of the 2020 global populations. Data from https://www.fao.org/faostat/en/#data, assuming 3.5 % protein in milk and 16 % protein in meat; note that this will underestimate the earlier y-values for beef and pork due to higher fat content, especially in Asia.
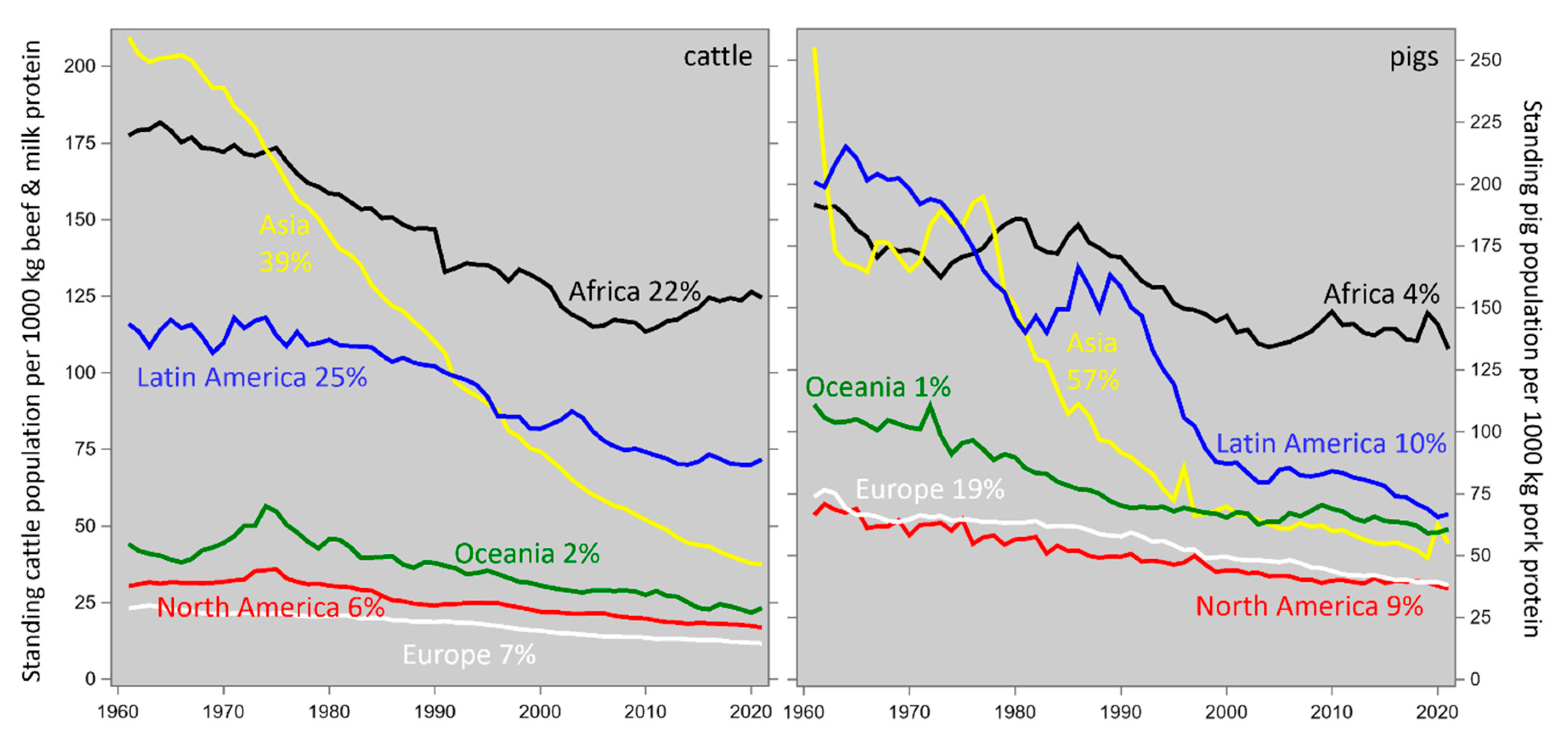

Table 1.
Performance KPI data from the PIC internal benchmark and the prominent industry benchmark, recorded in 2021. Means ± standard deviations.
Table 1.
Performance KPI data from the PIC internal benchmark and the prominent industry benchmark, recorded in 2021. Means ± standard deviations.
| PIC-USA internal benchmark |
North American prominent industry benchmark |
|
|---|---|---|
| Sow performance | ||
| Number of sows | 1,093,952 | 1,273,698 |
| Farrowing rate (%) | 85.7 ± 4.12 | 91.6 ± 4.74 |
| Total number born | 15.2 ± 0.68 | 15.1 ± 0.62 |
| Number born alive | 13.7 ± 0.62 | 13.7 ± 0.53 |
| Lactation mortality rate (%) | 15.8 ± 3.69 | 16.2 ± 2.63 |
| Weaning age (days) | 21.4 ± 2.01 | 21.0 ± 1.42 |
| Number weaned | 11.6 ± 0.59 | 11.5 ± 0.62 |
| PWMFY1 | 26.5 ± 4.17 | 27.0 ± 1.99 |
| Sow mortality rate (%) | 15.0 ± 4.70 | 14.1 ± 3.69 |
| Wean-to-finish performance | ||
| Number of pigs | 4,741,133 | 9,134,940 |
| Start weight (kg) | 5.98 ± 0.59 | 5.53 ± 0.39 |
| End weight (kg) | 129.0 ± 5.31 | 129.1 ± 2.09 |
| Days on feed | 158.7 ± 14.7 | 167.1 ± 5.15 |
| Growth rate (kg/d) | 0.77 ± 0.060 | 0.74 ± 0.023 |
| Feed intake (kg/d) | 1.96 ± 0.15 | 1.94 ± 0.11 |
| Feed conversion ratio | 2.55 ± 0.17 | 2.63 ± 0.12 |
| Mortality rate (%) | 6.30 ± 4.40 | 9.88 ± 3.48 |
| 1 PWMFY: piglets weaned per mated female per year | ||
Table 2.
Reported reductions of GHG emission from livestock production systems, due to genetic selection.
Table 2.
Reported reductions of GHG emission from livestock production systems, due to genetic selection.
| Species | Location | Annual reductiondue to genetics | Source | Comments | |||
| kg CO2eq per animal | % ofbase level | ||||||
| Sheep | Ireland | 0.12 % | Farrell et al. (2022)Berry et al. (2022) | Correlated GHG reduction due to selection for breeding goals that do not include feed intake or feed efficiency (the traits with the strongest connection to GHG). | |||
| Pigs | Netherlands | 1.26 | 0.7 % | De Haas et al. (2021) | 2014 and 2016 cohorts grown on an experimental farm with unchanged housing and nutrition. The 2016 cohort showed a 1.2 to 1.5 % lower GHG emission, at a base level of 180 kg CO2eq (22-120 kg liveweight). | ||
| Pigs | Norway | 2.61 - 3.54 | 1.4 - 1.9 % | Bonesmo & Gjerlaug-Enger (2021)Gjerlaug-Enger et al. (2022) | Correlated GHG reduction due to realized genetic change in feed efficiency, postweaning mortality and sow reproductive output (2021), at a base level of 186 kg CO2eq (80 kg carcass weight). | ||
| Pigs | North Americaand Europe | 2.93 | Knap et al. (2023) | Realized genetic change in (re)production traits (2012-2023) was converted to a change in whole-enterprise FCR, and from there to a change in GHG emission. GHG base level unknown. | |||
| Pigs | Western world | 2.4 | 1.2 % | Overall trend | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



