
Submitted:
12 July 2024
Posted:
15 July 2024
You are already at the latest version
Abstract
One of the most important agrotechnical factors that influence the growth conditions of crops is plant density. Alterations in habitat conditions result in the emergence of phytophagous organisms, whose development is closely associated with host plants. The objective of this study was to assess the abundance and species composition of thrips (Thysanoptera) collected in soybean plants of different varieties cultivated under different plant arrangements (row spacing, number of seeds sown differed). The research was conducted at two locations in southwestern Poland between 2015 and 2018. The herbivores that inhabit plants were caught using an entomological net. The experiments were carried out using the complete randomised block design, with four replications. Regardless of location and treatment, thrips were the most numerous group of pests. Thrips tabaci and T. fuscipes were the dominant species within the collected material. A total of 24 species of thrips were identified and their dietary specialisation was determined. The assemblages of studied insects were also analysed due to the density of the plants and the stage of development. The increase in the sowing density of soybean (90 seeds per m2) resulted in a greater total number of thrips compared to a lower sowing density (50 seeds per m2). The number of thrips significantly increased during the flowering period and shortly after the flowering time, which made them a significant threat to the plants. Therefore, it is crucial to develop innovative pest management strategies tailored to effectively grow soybean to mitigate potential pest-related challenges.
Keywords:
crop
; Thysanoptera
; herbivores
; plant density
; row spacing
; number of seeds sown
1. Introduction
Soybean is one of the most important crops in many parts of the world, along with wheat, rice and maize. Most of the production is located in the United States, Brazil and Argentina [1]. The seeds of this crop contain approximately 40% protein and 20% oil, making it a high-quality food for humans and animals. According to FAOSTAT data, the area under soybean in the world exceeds 132 million hectares, of which about 4% is in the European Union [2]. The area of soybean increased also in Poland, and the demand for its seeds is increasing every year. New varieties are being developed that can adapt to European climatic conditions and day length. There are currently 40 varieties in the National Register of Agricultural Crops in Poland, and the number of varieties has increased significantly in recent years. This indicates a significant increase in interest in non-GM soybean cultivation. In 2022, the area under soybean was 48,194 hectares [3], however this crop is still consider as a minor crop due to low and unstable yields [4].
Both plant spacing and plant density have a major influence on the environmental conditions of the field. It is also directly related to the presence of entomofauna in the field [5,6,7]. A higher plant density prevents the soil from heating up and keeps the air humidity relatively constant. The microclimate of a given field affects not only the size and quality of the crop, but also the presence and development of insects [8]. When considering the effect of plant density on the presence of harmful and beneficial entomofauna, it must also be taken into account that density affects not only the amount of food available in the form of host plants, but also the quality of this food, because a sufficiently high density causes excessive competition for light and microelements contained in the soil, a change in the nutrient content of the plant and consequently a change in the attractiveness of plants to herbivores [9].
Thrips (Thysanoptera) are one of the most important herbivores feeding on soybean plants. Neves et al. [10] found that soybean yield losses can be as high as 17%. More than 6,500 thrips species have been identified worldwide, of which about 60 are considered crop pests and only about a dozen are capable of transmitting viruses [11,12]. Thrips have piercing sucking mouthparts, but unlike Hemiptera, the mouthparts of Thysanoptera are asymmetrical (strongly reduced right lower mandibles) [13]. These insects with incomplete metamorphosis damage various parts of the plant (leaves, flowers and rarely fruits), suck the contents of the mesophyll cells and inject toxins into the plants, causing the typical discolouration in the form of silver-white spots and visible pieces of dark faeces. Later feeding causes deformation and death of young leaves, their premature fall and the death of healthy plants. Thrips appear to be particularly dangerous during flowering, when soybean is often attacked. They spend most of their time hiding under leaves, which is made possible by their small size (most species reach 1-2 mm in body size) [14,15]. Their hidden lifestyle makes them difficult to control with insecticides [16].
One of the most common pests in the order Thysanoptera is the tobacco thrips (Thrips tabaci). It is a polyphagous species and is widespread in field and greenhouse crops throughout the world. Its relatively short developmental cycle and ability to undergo parthenogenesis make it an extremely dangerous pest of many crops [17]. In agricultural and horticultural practice, thrips play a major negative role on cereals (Haplothrips aculeatus, H. tritici, Limothrips cerealium, L. denticornis, Kakothrips robustus), legumes (Thrips major, T. fuscipennis, T. flavus, Frankliniella intosa, Taeniothrips atratus, Odontothrips loti), flax and vegetables (K. robustus, T. tabaci), greenhouse plants (F. occidentalis, T. tabaci, Echinothrips americanus) [16,18,19,20,21,22,23]. Crops also often contain beneficial Thysanoptera species. A common species in Europe is Aeolothrips intermedius, which is mainly a natural enemy of other thrips; both adults and larvae are predators [24,25].
To date, no specific studies have been conducted on thrips feeding on soybean plants in Europe. The main objective of this study was to analyse the abundance and species composition of thrips in soybean crops with different plant arrangements.
2. Materials and Methods
2.1. Study Area
The field research was carried out in 2015-2018 in two locations in south-western Poland: the Agricultural Experimental Station in Wrocław-Pawłowice (51°10’36.5 N, 17°06’24. 9 E) (Location 1), which is part of the University of Environment and Life Sciences in Wroclaw (Lower Silesia), and in the experimental fields of the Opole Agricultural Advisory Centre (OODR) in Łosiów (50°80’14 N, 17°55’22 E) (Location 2) (Opole Voivodeship).
In Location 1, the experiment was carried out in 2015-2017 on a soybean plantation of the Merlin variety, growing on fawn soil of light clay on medium soil, classified as a good agricultural suitability complex, evaluation class III b [26]. In each year of the study, the soybean was grown under the same habitat conditions, next to the place of the previous year’s cultivation. A slight change in the location of the research plantation was caused by the principles of crop rotation. The forecrop was always winter wheat of the Ostroga variety. The wheat was harvested with a combine harvester in the first ten days of August and the crop residues were left in the field. Autumn tillage treatments included pre-winter ploughing and treatments with a tillage unit (spring cultivator + spring roller). In spring, the field was pre-fertilised with the following doses (kg ha-1) 60 P2O5, 120 K2O and 30 N. In each year of the study, soybean seeds were sown with a plot seeder in the third decade of April. The sowing depth was 3-4 cm. Before soybean emergence, herbicide treatments were applied to reduce the occurrence of dicotyledonous weeds (Afalon 450 SC with linuron as active ingredient) at a dose of 1.5 dm³ ha-1. In the second decade of May, after emergence, a treatment against dicotyledonous and some monocotyledonous weeds was applied (Corum 502.4 SL, bentazon and imazamox as active ingredients) at a dose of 1.25 dm³ ha-1, combined with an adjuvant containing methyl oleate and fatty alcohol (Dash HC) at a dose of 0.6 dm³ ha-1. No insecticide or fungicide treatments were applied in the field trial. Soybean was harvested in the middle of September each year when the pods were fully ripe.
In Location 2, soybean was grown under the same habitat conditions in each year of the study (2016-2018), next to the previous year’s field location. A slight shift in field location was necessary due to crop rotation principles. In each year, soybean was grown on soil belonging to the brown soil type, class III a, and its soil suitability complex was defined as very good rye. In each year, the forecrop was winter wheat of the Dakotana variety. In spring, pre-sowing fertilisation was applied in the following doses P2O5 - 20 (kg ha-1), K2O - 40 (kg ha-1), MgO - 31 (kg ha-1), S - 26 (kg ha-1) and N - 28 (kg ha-1). Soybean was sown in the third decade of April. The sowing depth was 4 cm. Pre-emergence herbicide treatments were applied throughout the plantation to reduce dicotyledonous weeds (Sencor Liquid 600 SC + Command 480 EC (metribuzin and clomazone as active ingredients) at doses of 0.6 and 0.2 dm³ ha-1). The post-emergence treatment against dicotyledonous and some monocotyledonous weeds (Corum 502.4 SL, bentazon and imazamox) was applied at a dose of 1.25 dm³ ha-1 together with an adjuvant containing methyl oleate and fatty alcohol (Dash HC at a dose of 0.6 dm³ ha-1 and Select Super 120 EC (cletodymium) at a dose of 0.8 dm³ ha-1). Seeds were harvested when the soybean plants were fully mature, in the middle of September.
2.2. Experiment Layout
The experimental factors in Location 1 were different row spacing (15 cm and 30 cm) and the number of soybean seeds sown per square metre (50 and 90) (Table 1). The experiment was designed as a complete randomised block design with four replications. A total of 16 plots were used for the entomological study. Each experimental plot was 10 metres long and 3 metres (30 m2) wide.
At the second site (Location 2), the field experiment was conducted in a randomised block design with three replications. The experimental factors were row spacing and soybean varieties. The research was carried out in the following variants: row spacing of 12 cm and 45 cm and soybean varieties: Abeline (variety 1) and Lissabon (variety 2) (12 research plots in total) (Table 1). Each plot was 5 metres long and 4 metres wide (20 m2).
2.3. Insect Sampling
Arthropod collection on soybean plants was carried out at two sites using the same methodology. The collection was carried out using an entomological net with a diameter of 40 cm and a telescopic handle of 80 cm. The collection was carried out in three stages of the soybean, i.e., before flowering (BBCH 12-49 stage), during flowering (BBCH 51-60) and after flowering (BBCH 61-71). The collection was carried out by moving along the longer side of the plot, in its central part. Twenty movements with the entomological net were made in each plot. Caught arthropods were immediately killed by poisoning with ethyl acetate. The entomological material was taken to the laboratory of the Department of Plant Protection and preserved in a 75% ethanol solution. All captured thrips were counted and identified to species.
The following keys were used to identify species on the basis of morphological characteristics: ‘Thysanoptera. Handbooks for the Identification of British Insects’ [15], ‘Pictorial key to the economically important species of Thysanoptera in Central Europe’ [27], ‘Thysanoptera: an identification guide’ [28], ‘Die terebranten Thysanopteren Europas und des Mittelmeer-Gebietes’ [29], ‘Comparative morphology of the second larval instar of Thrips genus species (Thysanoptera: Thripidae) occurring in Poland’ [30].
2.4. Data Analysis
Total thrips abundance was analysed using a three-way ANOVA. For Location 1 the independent variables were sowing density, row spacing and sampling date, and for Location 2 variety, row spacing and sampling date. For both localities three sampling dates were compared: before flowering, during flowering and after flowering. Only the significant results are presented in the paper. Analyses were carried out using Origin Pro software.
The thrips species communities in each site and sampling year were correlated with sampling date, row spacing, sampling density (Location 1) and variety (Location 2) using redundancy analysis. The contribution of the explanatory variables to the explained variance was determined using factorial analysis. The analyses were carried out in Canoco 5. The species community in Location 1 and Location 2 was compared using PERMANOVA analysis. In addition, rarefaction curves were constructed for the species community in both localities. The analyses were carried out using PAST software.
3. Results
Thrips communities were relatively abundant, with mean counts ranging from 20 to 60 individuals per plot in Location 1 and from 5 to 20 individuals per plot in Location 2 across different sampling dates (see Figure 1A,D). In both locations, sampling date had a significant effect on total thrips abundance (p<0.0001) (Table 2). Furthermore, in Location 1, sowing density had a significant effect on the abundance of thrips. Specifically, thrips abundance increased during soybean flowering in Location 1 (Figure 1A), whereas it increased after flowering in Location 2 (Figure 1D). In particular, a reduction in seed density from 90 to 50 seeds per m2 resulted in a decrease in thrips abundance at Location 1 (p=0.031) (Figure 1B). Especially during flowering, a higher number of thrips was observed in the 90 seeds per m2 compared to 50 seeds per m2 (Figure 1C). However, neither row spacing (in both locations 1 and 2) nor cultivar (in location 2) had a significant effect on thrips abundance (Table 2).
In the soybean crop, a total of 24 thrips species were identified in two different localities, with 20 species observed in each locality. The predominant species in both localities was Thrips tabaci, followed by Aeolothrips intermedius and Thrips fuscipes (Figure 2A-B). In particular, T. tabaci and T. fuscipes are phytophagous, whereas A. intermedius is a predatory species (Table 4). Apart from these three species, the composition of dominant species within the communities varied between the two locations. Statistical analysis using PERMANOVA revealed significant dissimilarities in the thrips species communities between Location 1 and Location 2 (p=0.0001) (Table 3).
Most of the identified species exhibit phytophagous behaviour and have a wide range of host plants that are not exclusively associated with soybean cultivation, but are able to survive and reproduce in soybean plantations (Table 3). Species that are particularly associated with soybean include Odontothrips loti, T. tabaci and T. fuscipennis. The prevalence of polyphagous species suggests a lack of specialised thrips species closely associated with soybean cultivation at both sites. Oligophagous species such as Neohydatothrips gracilicornis and Thrips atratus were present in smaller numbers, but were trapped throughout the growing season, indicating potential feeding on soybeans beyond their primary trophic spectrum.
The predatory A. intermedius was prominently represented, along with the less abundant A. fasciatus, both of which prey on other thrips species, including adults and larvae. After flowering, a number of species associated with monocotyledonous plants appeared, including Limothrips cerealium, Haplothrips aculeatus, H. leucanthemi, Anapothrips obscurus and Chirothrips manicatus. Interestingly, infestations of these species coincided with the harvest period, suggesting that thrips migrate to soybean fields due to a lack of preferred food plants.
The ordination analysis of the thrips community highlighted the sampling date as the main factor influencing the dynamics of the thrips community within the soybean crop (Table 5 and Figure 2). Factorial analysis (also shown in Table 5) indicated significant effects of three different sampling dates on the distribution of thrips species. At Location 1, thrips species richness was highest during the flowering season, although the specific composition varied between years (Figure 3ACA). In contrast, at Location 2, species richness peaked after flowering, with notable representatives including L. cerealium, H. niger, H. leucanthemi, H. aculeatus (Figure 2D-E).
4. Discussion
The spatial structure of the canopy is determined by the spacing of plant rows, and the distance between plants in a row significantly alters the microclimate that shapes the habitat conditions of a given crop. The arrangement of plants and their density per unit area can, in turn, influence the number of herbivores inhabiting the crop and, consequently, their harmfulness [31]. In the case of soybean, there has been no comprehensive study of the influence of canopy plant density, seeding rate or row spacing on the occurrence of pests, including thrips. Only preliminary studies on herbivores and beneficial carabid beetles have been published [32]. Meanwhile, Santos et al. [33] mentioned that thrips are an emerging problem, leading to soybean yield losses of up to 15%.
In the experiment, the lower sowing density (50 seeds per m2) generally reduced the occurrence of thrips, consistent with the resource concentration hypothesis [34,35]. Based on this assumption, we would expect herbivores to be more abundant in areas with higher densities of host plants [36]. The resource concentration hypothesis predicts that the density of insect herbivores per plant will increase as the density of host plants increases [34]. Pobereżny et al. [37] captured more Oulema beetles and larvae gathered with an entomological net in spring wheat grown at a seed density of 600 seeds/m2 (the highest number of seeds used in the experiment described). In addition, dense plant assemblages provide a more stable habitat for herbivores and increase reproductive potential [38]. Research by Krobb et al. [39] suggests that the density of cotton plants has a significant effect on the occurrence of thrips and the extent of damage within plantations. Their study found that cotton plants sown at the highest density (52.6 seeds per square metre) were less susceptible to damage and had lower thrips populations than the lower sowing densities tested. Underwood and Halpern [40] show that increasing plant density, as well as plant size and canopy architecture, has a significant effect on insect density. In maize, Nowatzki et al. [41] investigated the damage potential of corn rootworm to the crop. It was shown that reducing the distance between maize rows from 76 to 38 cm should not increase the risk of root damage caused by Diabrotica virgifera larvae, although the number of adult corn rootworm individuals increased with lower plant density. Hurej and Twardowski [5] assessed the presence of herbivores in hybrid varieties of winter oilseed rape grown at different plant densities. In this case, some pests, such as Brassicogethes aeneus, were significant at lower plant densities. Nerlekar [42] states that the presence of herbivorous species is influenced by the amount and condition of vegetation. According to this author, phytophagous organisms are usually more likely to occur in less plant-dense environments, but this is not a rule as it depends on the biology of the pests.
Thysanoptera, an important group of phytophagous insects, are known to feed on various crops. However, in Poland there has been no assessment of their occurrence specifically on soybean plantations, nor has there been a comprehensive identification of Thysanoptera species in this context. Research has shown that the species composition and abundance of thrips changes with the development of the host plant. Most of the thrips species trapped in the study sites show polyphagous behaviour, the most abundant species being T. tabaci and T. fuscipennis, neither of which are directly associated with soybean cultivation. K. robustus, cited in the literature as a legume-associated species, was conspicuously absent from all surveyed locations [43,44]. Thrips species with stable numbers throughout the growing season are T. tabaci and T. fuscipennis, together with the predatory species A. intermedius. These species appear relatively early in the plantation cycle and persist until the end of the season, benefiting from a broad trophic spectrum that enables successful colonisation of soybean crops. The most notable population dynamics of these species occurs during the flowering period, which coincides with the increased availability of high-protein food sources such as soybean flower pollen. This period promotes significant population growth due to the abundant food resources available. During flower feeding, thrips can cause anthers to rupture, resulting in pollen shed, a common symptom that reduces flower pollination and ultimately reduces yield [45,46,47]. In addition, thrips damage to flowers and fruit buds often results in deformation, inhibition of fruit growth or even complete fruit mortality [46]. Similar trends for the occurrence of T. tabaci, T. fuscipennis and A. intermedius were observed in different Fabaceae cultures in Poland. The less abundant species F. intonsa occurred in our study before and during the flowering period, similar to studies in Poland [43], and Hungary [48]. Larvae of this species feed on pollen grains [43].
The second group of thrips, although less abundant, appears relatively early (BBCH 12-49) and survives feeding on soybeans until the end of the growing season. This group includes T. flavus, T. major, F. intonsa and H. aculeatus, which are ubiquitous species with a broad host spectrum and no specific habitat preferences [49]. A notable specialist species in this context is Odontothrips loti, which is exclusively associated with legume crops and was found in relatively low numbers at both study sites. O. loti is closely tied to legumes and can only complete its life cycle by supplementing itself with pollen from plants of this genus [50]. In Romania, O. loti, together with the related species O. confuses and O. phaleratus, has been identified as a significant pest of lucerne, alfalfa and white clover used for green fodder, causing substantial yield losses [51]. Another species, N. gracilicornis, known to be associated with legumes, especially Vicia species such as V. cracca (bird vetch), was present in low numbers at both locations.
Taking into account the expected expansion of soybean production in Poland and Europe due to factors such as European Union policies, subsidies for legume production and the development of new soybean varieties better adapted to the Polish climate, it is likely that thrips pressure will increase. The emergence of more specialised pests specific to soybean is also a possibility. Therefore, it is crucial to develop innovative pest management strategies tailored to effective soybean cultivation to mitigate potential pest-related challenges.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Pictures of selected thrips species (by co-author Marcin Cierpisz).
Author Contributions
Conceptualization, J.T. and M.K.; methodology, M.K., J.T. and I.G.; validation, J.T. and I.G; formal analysis, J.T. and I.G; investigation, J.T., M.C., K.T., J.M-D; resources, J.T., and M.K; data curation, J.T., M.C., K.T. and I.G..; writing—original draft preparation, J.T., M.C, I.G.; writing—review and editing, J.T.; visualization, M.C. and I.G.; supervision, J.T.; project administration, M.K.; funding acquisition, M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially financed from the multi-annual programme 2016–2020 by the Ministry of Agriculture and Rural Development titled “Increasing the use of domestic fodder protein for the production of high-quality animal products in the conditions of sustainable development”, Resolution No. 222/2015 of the Council of Ministers of 15 December 2015 (Poland).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data are contained within the article.
Acknowledgments
The cooperation and support of Prof. Hanna Kucharczyk (Department of Zoology, Maria Curie-Sklodowska University in Lublin) in identification of thrips are greatly appreciated.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Pagano, M.C.; Miransari, M. The importance of soybean production worldwide. In book: Abiotic and Biotic Stresses in Soybean Production, Editor Mohammad Miransari; Publisher: Academic Press. 2016.
- Faostat. Food and Agriculture Organization of the United Nations (FAO). 2021. Available online: https://www.fao.org/faostat/en/#home (accessed on 10 May 2024).
- ARiMR. Agencja Restrukturyzacji i Modernizacji Rolnictwa (Poland). In English: The Agency for Restructuring and Modernisation of Agriculture (ARMA). 2024. Available online: https://www.gov.pl/web/arimr/agencja-restrukturyzacji-i-modernizacji-rolnictwa (accessed on 10 May 2024).
- Serafin-Andrzejewska, M.; Helios, W.; Białkowska, M.; Kotecki, A.; Kozak, M. Sowing Date as a Factor Affecting Soybean Yield – A Case Study in Poland. Agriculture. 2024, 14, 970. [Google Scholar] [CrossRef]
- Hurej, M.; Twardowski, J. Wpływ rozmieszczenia roślin w łanie na występowanie ważniejszych fitofagów rzepaku ozimego. Zesz. Nauk. UP we Wrocławiu, Seria Rolnictwo. 2007, 553, 67–74. [Google Scholar]
- Cierpisz, M.; Twardowski, J.; Kozak, M. The effect of plant arrangement in soybean crop on more important herbivores – preliminary results. J. Res.Appl. Agric. Eng. 2016, 61, 48–52. [Google Scholar]
- Twardowski, J.; Hurej, M.; Ścibior, R.; Kotecki, A. The effect of different seeding density of linseed (Linum usitatissimum L.) on flax flea beetles (Col., Chrysomelidae). J. Plant Prot. Res. 2017, 57, 158–166. [Google Scholar] [CrossRef]
- Hanh, P.G.; Orrock, J.L. Spatial arrangement of canopy structure and land use history alter the effect that herbivores have on plant growth. Ecosphere. 2015, 10, 1–16. [Google Scholar] [CrossRef]
- Mondal, M.M.A.; Puteh, A.B.; Malek, M.A.; Ismail, M.R. Determination of optimum seed rate for mungbean based on morpho-physiological criteria. Legume Research. 2012, 35, 126–131. [Google Scholar]
- Neves, D.V.C.; Lopes, M.C.; Sarmento, R.A.; Pereira, P.S.; Pires, W.S.; Peluzio, J.M.; Picanço, M.C. Economic injury levels for control decision-making of thrips in soybean crops (Glycine max (L.) Merrill). Res. Soc. Develop. 2022, 11, e52411932114. [Google Scholar] [CrossRef]
- Moritz, G.; Paulsen, M.; Delker, C.; Picl, S.; Kumm, S. Identification of thrips using ITS–RFLP analysis. Thrips and Tospoviruses: Proceedings of the 7th International Symposium on Thysanoptera. 2002, 365–367.
- Thekke-Veetil, T.; Lagos-Kutz, D.; McCoppin, N.K.; Hartman, G.L.; Ju, H.-K.; Lim, H.-S.; Domier, L.L. Soybean Thrips (Thysanoptera: Thripidae) Harbor Highly Diverse Populations of Arthropod, Fungal and Plant Viruses. Viruses 2020, 12, 1376. [Google Scholar] [CrossRef] [PubMed]
- Kucharczyk, H.; Kucharczyk, M. Characteristic and diagnostic features of the most frequently occurring species of the Thripidae family (Insecta, Thysanoptera) in crown canopies of Central European forests. Leśne Prace Badawcze. 2013, 74, 5–11. [Google Scholar] [CrossRef]
- Sierka, W. Wciornastki, czyli thripsy (Insecta, Thysanoptera). Pol. Pismo Entomol. 2004, 1, 1–12. [Google Scholar]
- Mound, L.A.; Morison, G.D.; Pitkin, B.R.; Palmer, J.M. Thysanoptera. Handbooks for the Identification of British Insects. Royal Entomological Society of London. 1976, 1, 1–79. [Google Scholar]
- Hurej, M.; Kucharczyk, H.; Twardowski, J.; Kozak, M. Thrips (Thysanoptera) associated with narrow-leafed lupin (Lupinus angustifolius L., 1753) intercropped with spring triticale (x Triticosecale Wittm. ex A. Camus, 1927). Rom. Agric. Res. 2014, 31, 337–345. [Google Scholar]
- Gill, H.K.; Garg, H.; Gill, A.K.; Gillett-Kaufman, J.L.; Nault, B.A. Onion thrips (Thysanoptera: Thripidae) biology, ecology, and management in onion production systems. J. Integr. Pest Manag. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Hurej, M.; Twardowski, J. Thrips (Thysanoptera) occuring in spring triticale intercropped with yellow lupine. Acta Sci. Pol. Agricultura. 2004, 3, 263–270. [Google Scholar]
- Hurej, M.; Twardowski, J.; Chrzanowska-Drożdż, B. Thrips (Thysanoptera) occurring in ears of Triticum durum Desf. in conditions of different protection level. Acta Sci. Pol. Agricultura. 2010, 9, 3–10. [Google Scholar]
- Hurej, M.; Kucharczyk, H.; Twardowski, J.; Kotecki, A. Thrips (Thysanoptera) associated with two morphological forms of Andean lupin (Lupinus mutabilis Sweet). Biologia. 2015, 70, 935–942. [Google Scholar] [CrossRef]
- Hurej, M.; Kucharczyk, H.; Twardowski, J.; Kotecki, A. Thrips (Thysanoptera) associated with two genetically modified types of linseed (Linum usitatissimum L.). J. Plant Dis. Prot. 2017, 124, 81–91. [Google Scholar] [CrossRef]
- Łuczak, I.; Gaborska, M.; Pobożniak, M.; Świderski, A.; Kruczek, M. Occurrence of phytophagous thrips (Thysanoptera) and harmfulness of Thrips tabaci Lind. in carrot (Daucus carota L. ssp. sativus) cultivation (Daucus carota L. ssp. sativus). Prog. Plant Prot. 2014, 54, 198–204. [Google Scholar] [CrossRef]
- Olczyk, M.; Pobożniak, M. Thrips (Thysanoptera) associated with onion (Allium cepa L.) and Welsh onion (Allium fistulosum L.). Folia Hort. 2020, 3, 1–17. [Google Scholar] [CrossRef]
- Trdan, S.; Andjus, L.; Raspudić, E.; Kač, M. Distribution of Aeolothrips intermedius Bagnall (Thysanoptera: Aeolothripidae) and its potential prey Thysanoptera species on different cultivated host plants. J. Pest Sci. 2005, 78, 217–226. [Google Scholar] [CrossRef]
- Gruss, I.; Twardowski, J.P.; Cierpisz, M. The Effects of Locality and Host Plant on the Body Size of Aeolothrips intermedius (Thysanoptera: Aeolothripidae) in the Southwest of Poland. Insects, 2019, 10, 266. [Google Scholar] [CrossRef]
- Włodarczyk, M. Effect of planting date of soybean (Glycine max (L.) Merill) on cultivars growth and yield. Ph.D. Thesis, Institute of Agroecology and Plant Production, Wroclaw University of Environmental and Life Sciences, Wrocław, Poland, 2020; 96p. (In Polish). [Google Scholar]
- Moritz, G.B. Pictorial key to the economically important species of Thysanoptera in Central Europe. Bulletin OEPP/EPPO 1994, 24, 181–208. [Google Scholar] [CrossRef]
- Mound, L.A.; Kibby, G. Thysanoptera an Identification guide. CAB Intl., ISBN: 0851992110. 1998, 70 pp.
- Zur Strassen, R. Die Terebranten Thysanopteren Europas. Keltern, Germany: Die Tierwelt Deutschlands, 74 Teils. Goecke and Evers. 2003.
- Kucharczyk, H. Comparative morphology of the second larval instar of the Thrips genus species (Thysanoptera: Thripidae) occurring in Poland, Mantis, Olsztyn, ISBN: 8392999770, 2010, 152 pp.
- Nuss, H. Effect of plant density and plant architecture on the abundance and within–plant distribution of stem borers in winter oilseed rape. Ph.D. Thesis, University of Göttingen, in Deutsch, 2004. [Google Scholar]
- Cierpisz, M.; Twardowski, J.; Gruss, I.; Kozak, M. Different soybean plant arrangements affect ground beetle assemblages. J. Plant Prot. Res. 2019, 59, 441–450. [Google Scholar] [CrossRef]
- Santos, J.L.; Pereira, P.S.; Reis, K.H.B.; Freitas, D.R.; Picanço Filho, M.C.; Peluzio, J.M.; Sarmento, R.A.; Guedes, R.N.C.; Picanço, M.C. Decision-making for thrips control in soybean fields using precision agriculture principles. J. Appl. Entomol. 2024, 148, 140–149. [Google Scholar] [CrossRef]
- Root, R.B. Organization of a plant-arthropod association in simple and diverse habitats: the fauna of collards (Brassica oleracea). Ecol. Monogr. 1973, 43, 95–124. [Google Scholar] [CrossRef]
- Grez, A.A.; González, R.H. Resource concentration hypothesis: effect of host plant patch size on density of herbivorous insects. Oecologia. 1995, 103, 471–474. [Google Scholar] [CrossRef]
- Stephens, A.E.A.; Myers, J.H. Resource concentration by insects and implications for plant populations. J. Ecol. 2012, 100, 923–931. [Google Scholar] [CrossRef]
- Pobereżny, J.; Wszelaczyńska, E.; Lamparski, R.; Lemanowicz, J.; Bartkowiak, A.; Szczepanek, M.; Gościnna, K. The impact of spring wheat species and sowing density on soil biochemical properties, content of secondary plant metabolites and the presence of Oulema ssp. PeerJ, 2023, 11, e14916. [Google Scholar] [CrossRef] [PubMed]
- Maron, J.L.; Crone, E. Herbivory: effects on plant abundance, distribution and population growth. Proc. Royal Soc. B. 2006, 273, 2575–2584. [Google Scholar] [CrossRef]
- Krobb, J.L.; Stewart, S.D.; Brown, S.A. Effects of plant density, seed spacing, and seed treatment on thrips injury to cotton. Crop Prot. 2022, 161, 106059. [Google Scholar] [CrossRef]
- Underwood, N.; Halpern, S. Insect herbivores, density dependence, and the performance of the perennial herb Solanum carolinense. Ecology. 2012, 93, 1026–1035. [Google Scholar] [CrossRef] [PubMed]
- Nowatzki, T.M.; Tollefson, J.J.; Bailey, T.B. Effects of row spacing and plant density on corn rootworm (Coleoptera: Chrysomelidae) emergence and damage potential to corn. J. Econ. Entomol. 2002, 95, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Nerlekar, A. Seasonally dependent relationship between insect herbivores and host plant density in Jatropha nana, a tropical perennial herb. Biology Open. 2018, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pobożniak, M. The occurrence of thrips (Thysanoptera) on food legumes (Fabaceae). J. Plant Dis. Prot. 2011, 118, 185–193. [Google Scholar] [CrossRef]
- Tratwal, A.; Strażyński, P.; Mrówczyński, M. Poradnik sygnalizatora ochrony bobowatych grubonasiennych. Instytut Ochrony Roślin – Państwowy Instytut Badawczy. 2017, 57–121.
- Przybysz, A.; Fiedler, Z.; Obrępalska–Stęplowska, A. Klucze do diagnostyki morfologicznej i molekularnej Frankliniella occidentalis (Pergande) i Thrips palmi (Karny). Instytut Ochrony Roślin – Państwowy Instytut Badawczy. 2015, 5–24.
- Kumar, V.; Kakkar, G.; Palmer, C.L.; McKenzie, C.L.; Osborne, S.L. Thrips Management Program for Horticulture. University of Florida IFAS Extension. 2016; 1–7. [Google Scholar]
- Shimat, J.; Braman, K.; Hudson, W.; Nair, S. Biology and Management of Thrips Affecting the Production Nursery and Landscape. University of Georgia Extension. 2019; 1–5. [Google Scholar]
- Ábrahám, R. Thrips species associated with soybean in Hungary. Acta Phyt. Ent. Hung. 2008, 43, 211–218. [Google Scholar] [CrossRef]
- Kucharczyk, H. Przylżeńce (Thysanoptera) Roztocza. Frag. Faun. 1994, 37, 168–180. [Google Scholar] [CrossRef]
- Pitkin, B.R. A revision of the flower–living genus Odontothrips Amyot & Serville (Thysanoptera: Thripidae). Bulletin of the British Museum (Natural History) Entomology. 1972, 26, 371–402. [Google Scholar]
- Virteiu, A.M.; Steff, R.; Carabet, A.; Molnar, R.; Grozea, I. Revision of the genus Odontothrips Amyot & Serville (Thysanoptera, Thripidae) with the redescription of Odontothrips loti (Haliday, 1852) species on Lotus corniculatus crops. Res. J. Agric. Sci. 2021, 53, 255–261. [Google Scholar]
Figure 1.
Total abundance of thrips in Location 1 (A-C) in the three-year study period and Location 2 (D) in the two-year study period. A - Total abundance of thrips on 3 sampling dates in Location 1. B - Total abundance of thrips in 2 sowing densities in Location 1. C - Total abundance of thrips in 2 sowing densities in three sampling dates in Location 1. D - Total abundance of thrips on 3 sampling dates in Location 2. a, b, ab, c - indicate significant differences between variants. Before flowering - BBCH 40-53. During flowering- BBCH 54-69. After flowering- BBCH 70-84.
Figure 1.
Total abundance of thrips in Location 1 (A-C) in the three-year study period and Location 2 (D) in the two-year study period. A - Total abundance of thrips on 3 sampling dates in Location 1. B - Total abundance of thrips in 2 sowing densities in Location 1. C - Total abundance of thrips in 2 sowing densities in three sampling dates in Location 1. D - Total abundance of thrips on 3 sampling dates in Location 2. a, b, ab, c - indicate significant differences between variants. Before flowering - BBCH 40-53. During flowering- BBCH 54-69. After flowering- BBCH 70-84.
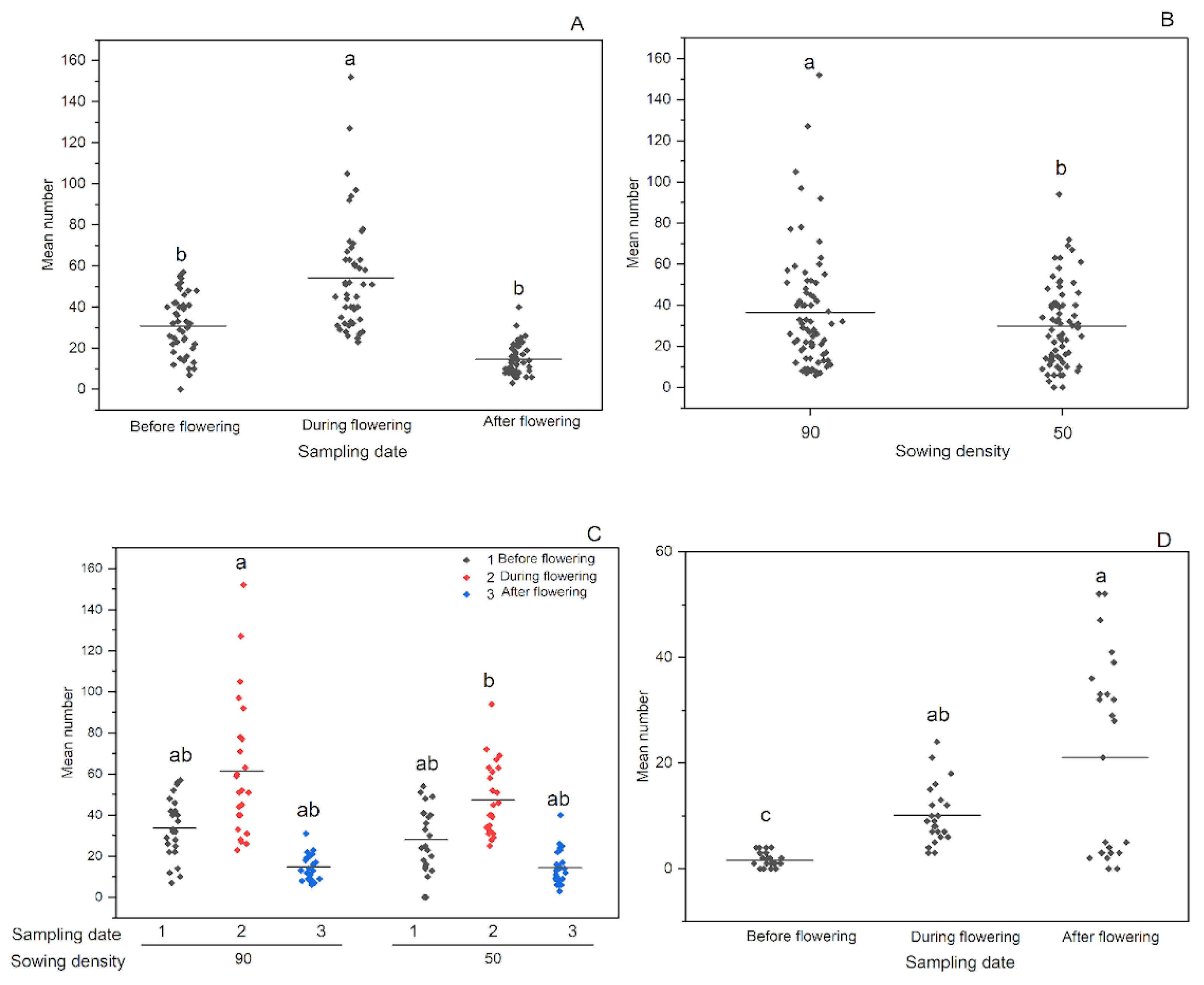
Figure 2.
Rarefaction curves showing the dominance structure of the species community in Locations 1 and 2.
Figure 2.
Rarefaction curves showing the dominance structure of the species community in Locations 1 and 2.
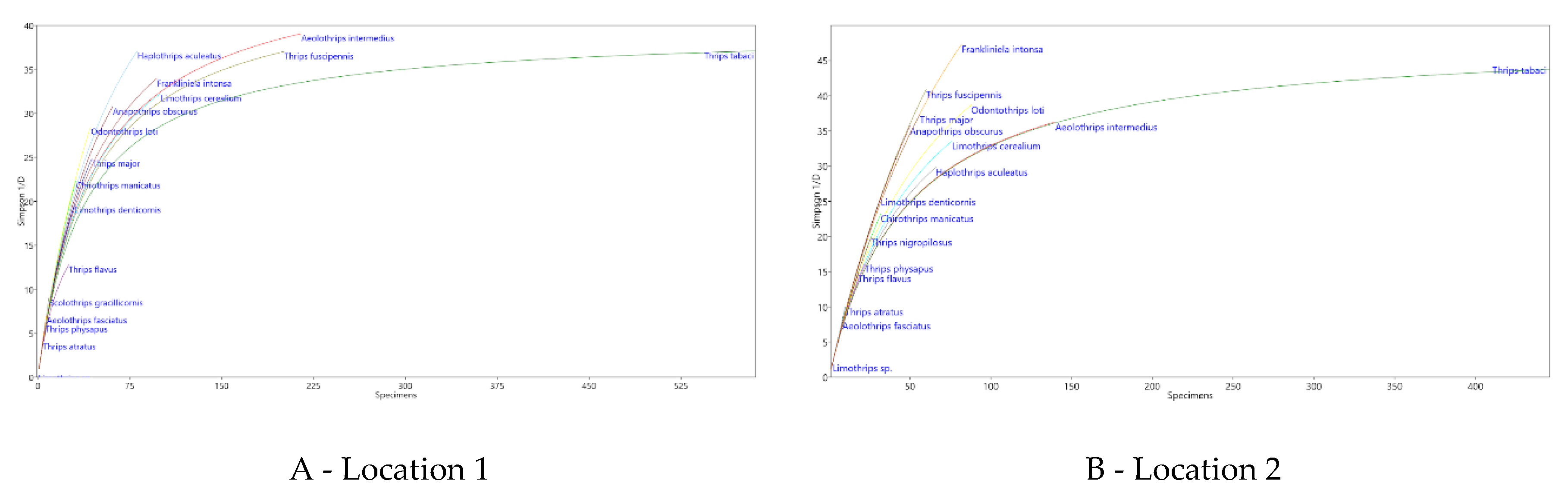
Figure 3.
RDA biplot of the thrips community in relation to the sampling date (before flowering, during flowering and after flowering); sowing density (A, B, C), variety (D, E) and row spacing (A-D). Before flowering - BBCH 40-53, During flowering- BBCH 54-69. After flowering- BBCH 70-84. The full names of the species are given in Table 4.
Figure 3.
RDA biplot of the thrips community in relation to the sampling date (before flowering, during flowering and after flowering); sowing density (A, B, C), variety (D, E) and row spacing (A-D). Before flowering - BBCH 40-53, During flowering- BBCH 54-69. After flowering- BBCH 70-84. The full names of the species are given in Table 4.
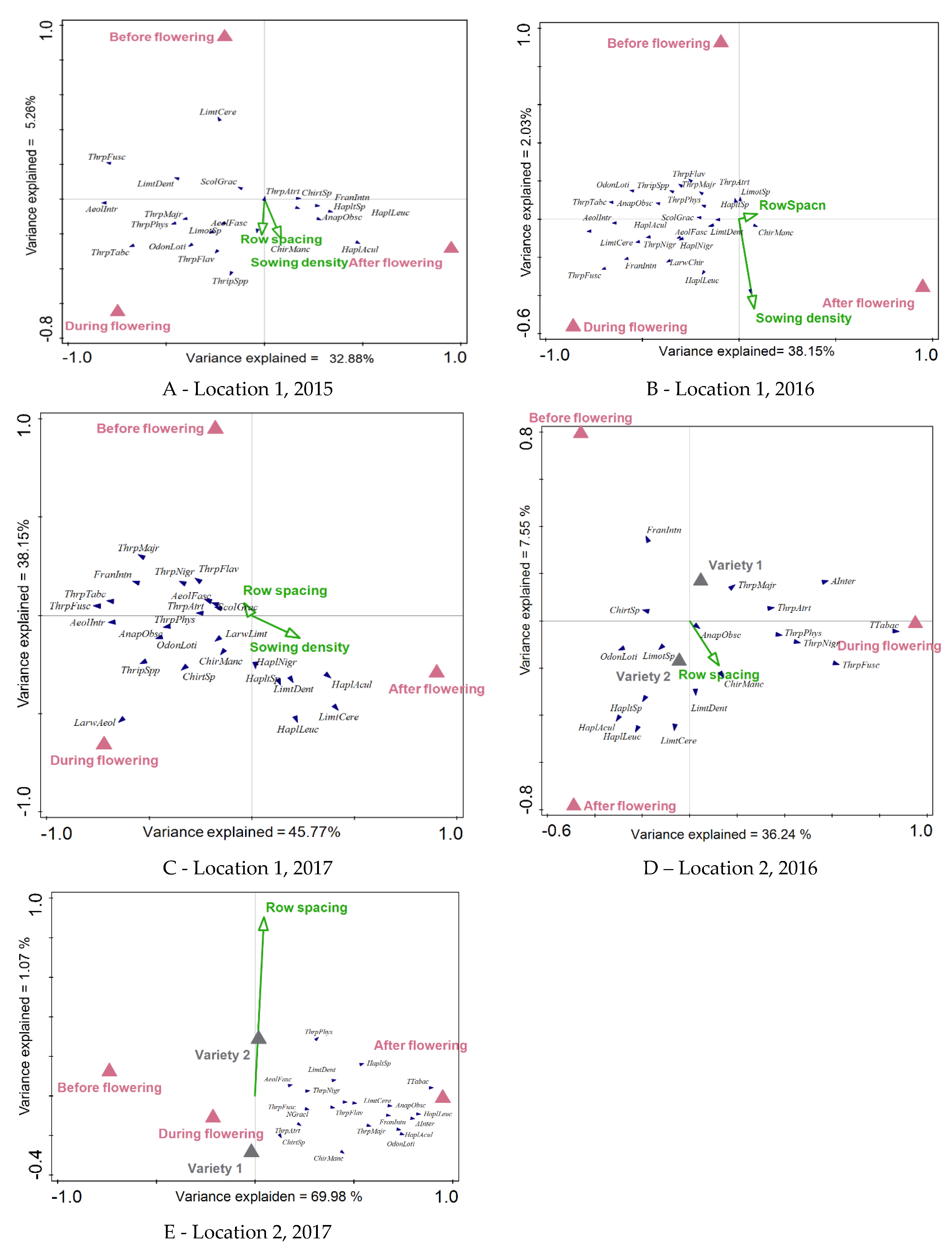

Table 1.
Experiment treatments at soybean fields in two research locations.
| Wrocław-Pawłowice (Location 1) | ||
| Research variant | Row spacing (cm) | Number of seeds sown on 1 m2 |
| A1 | 15 | 50 |
| A2 | 15 | 50 |
| A3 | 30 | 90 |
| A4 | 30 | 90 |
| Łosiów (Location 2) | ||
| Research variant | Row spacing (cm) | Soybean variety |
| B1 | 12 | Abeline (variety 1) |
| B2 | 12 | Lissabon (variety 2) |
| B3 | 45 | Abeline (variety 1) |
| B4 | 45 | Lissabon (variety 2) |
Table 2.
Results of the three-way ANOVA with row spacing, sowing density (Location 1) date of sampling and variety (Location 2) as the explanatory variables and thrips total abundance as response variable.
Table 2.
Results of the three-way ANOVA with row spacing, sowing density (Location 1) date of sampling and variety (Location 2) as the explanatory variables and thrips total abundance as response variable.
| DF | Sum of squares | Mean square | F value | P value | |
|---|---|---|---|---|---|
| Location 1 | |||||
| Row spacing | 1 | 152.11 | 152.11 | 0.44 | 0.51 |
| Sowing density | 1 | 1586.69 | 1586.69 | 4.6 | 0.031 |
| Date of sampling | 2 | 38421.54 | 19210.77 | 55.68 | <0.0001 |
| Sowing density x Date of sampling | 2 | 1127.1 | 563.55 | 1.63 | 0.20 |
| Model | 11 | 41852.83 | 3804.8 | 11.03 | <0.0001 |
| Error | 132 | 45538.17 | 344.991 | ||
| Corrected Total | 143 | 87391 | |||
| Location 2 | |||||
| Variety | 1 | 3.13 | 3.13 | 0.02 | 0.88 |
| Row spacing | 1 | 2.35 | 2.35 | 0.02 | 0.90 |
| Date | 2 | 4547.44 | 2273.72 | 16.16 | <0.0001 |
| Model | 11 | 4726.15 | 429.65 | 3.06 | 0.003 |
| Error | 60 | 8444.5 | 140.74 | ||
| Corrected Total | 71 | 13170.66 | |||
Table 3.
The results of the PERMANOVA analysis comparing the thrips species communities in Locations 1 and 2.
Table 3.
The results of the PERMANOVA analysis comparing the thrips species communities in Locations 1 and 2.
| Permutation N | Total sum of squares | Within-group sum of squares | F | p |
|---|---|---|---|---|
| 9999 | 3.6 | 3.119 | 32.87 | 0.0001 |
Table 4.
Ecological characteristics of the thrips species identified in soybean in two locations.
| Species | Abreviation | Trophic group | Food specialization | Host (if applicable) |
|---|---|---|---|---|
| Aeolothrips fasciatus (Linnaeus, 1758) | AelFasc | predator | mainly other thrips, adult stages and larvae | supplements the diet with plant pollen |
| Aeolothrips intermedius (Bagnall, 1934) | AeolIntr | predator | mainly other thrips, adult stages and larvae | supplements the diet with plant pollen |
| Chirothrips manicatus (Haliday, 1836) | ChirManc | phytophage | oligophage | associated with monocotyledonous plants |
| Frankliniella intonsa (Trybom, 1895) | FranInt | phytophage | phytophage | wide host spectrum, feeds on flowers and leaves |
| Haplothrips aculeatus (Fabricius, 1803) | HaplAcul | phytophage | oligophage | associated with monocotyledonous plants |
| Haplothrips leucanthemi (Schrank, 1781) | HaplLeuc | phytophage | oligophage | associated with monocotyledonous plants |
| Haplothrips niger (Osborn, 1883) | HaplNigr | phytophage | oligophage | associated with monocotyledonous plants |
| Limothrips cerealium (Haliday, 1836) | LimtCere | phytophage | oligophage | associated with monocotyledonous plants |
| Limothrips denticornis (Haliday, 1836) | LimtDent | phytophage | oligophage | associated with monocotyledonous plants |
| Odontothrips loti (Haliday, 1852) | OdonLoti | phytophage | oligophage | associated with Fabaceae |
| Neohydatothrips gracilicornis (Williams, 1916) | NGracl | phytophage | oligophage | mainly associated with related to Fabaceae |
| Thrips atratus (Haliday, 1836) | ThripAtr | phytophage | oligophage | associated mainly with carnation and Laminaceae |
| Thrips flavus (Schrank, 1776) | ThripFlav | phytophage | phytophage | feeds mainly on flowers, a wide range of host plants |
| Thrips fuscipennis (Hailday, 1836) | ThripFusc | phytophage | phytophage | feeds on flowers and leaves, wide range of hosts, most often Rosaceae |
| Thrips major (Uzel, 1895) | ThripMajr | phytophage | phytophage | feeds on flowers and leaves, wide range of hosts, most often Rosaceae |
| Thrips nigropilosus Uzel, 1985 | ThripNigr | phytophage | phytophage | feeds on flowers and leaves, a wide range of host plants |
| Thrips tabaci (Lindemann, 1889) | ThripTabc | phytophage | phytophage | feeds on flowers and leaves, a wide range of host plants |
Table 5.
Results of the factorial analysis with row spacing, sowing density (Location 1) date of sampling and variety (Location 2) as explanatory variables.
Table 5.
Results of the factorial analysis with row spacing, sowing density (Location 1) date of sampling and variety (Location 2) as explanatory variables.
| Factor | Year | Location | Explain % | Contribution % | pseudo-F | P |
|---|---|---|---|---|---|---|
| Sampling date | ||||||
| Before flowering | 2015 | Location 1 | 7.7 | 18.8 | 5.5 | 0.001 |
| 2016 | Location 1 | 8.1 | 19.8 | 6 | 0.002 | |
| 2017 | Location 1 | 9.4 | 18.1 | 8.2 | unknown | |
| 2016 | Location 2 | 7 | 18 | 3.5 | 0.001 | |
| 2017 | Location 2 | 6.7 | 9.3 | 7.4 | 0.002 | |
| During flowering | 2015 | Location 1 | 7.7 | 18.8 | 5.5 | 0.001 |
| 2016 | Location 1 | 8.1 | 19.8 | 6 | 0.006 | |
| 2017 | Location 1 | 9.4 | 18.1 | 8.2 | unknown | |
| 2016 | Location 2 | 28.1 | 72.8 | 13.3 | 0.001 | |
| 2017 | Location 2 | 6.7 | 9.3 | 7.4 | 0.001 | |
| After flowering | 2015 | Location 1 | 29.9 | 73.4 | 19.6 | 0.001 |
| 2016 | Location 1 | 31.2 | 76.3 | 20.8 | 0.001 | |
| 2017 | Location 1 | 39 | 74.9 | 29.5 | 0.001 | |
| 2016 | Location 2 | 7 | 18 | 3.5 | 0.001 | |
| 2017 | Location 2 | 63.4 | 88 | 58.8 | 0.001 | |
| Sowing density | 2015 | Location 1 | 2.2 | 5.3 | 1.6 | 0.11 |
| 2016 | Location 1 | 1 | 2.5 | 0.7 | unknown | |
| 2017 | Location 1 | 3.2 | 6.1 | 2.9 | 0.073 | |
| Variety | 2016 | Location 2 | 1.7 | 4.4 | 0.9 | 0.6 |
| 2017 | Location 2 | 0.8 | 1 | 0.8 | 0.48 | |
| Row spacing | 2015 | Location 1 | 1 | 2.5 | 0.7 | 0.71 |
| 2016 | Location 1 | 0.6 | 1.5 | 0.4 | 0.8 | |
| 2017 | Location 1 | 0.5 | 0.9 | 0.4 | 0.69 | |
| 2016 | Location 2 | 1.9 | 4.8 | 0.9 | 0.52 | |
| 2017 | Location 2 | 1.1 | 1.6 | 1.3 | unknown | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



