
Submitted:
15 July 2024
Posted:
16 July 2024
You are already at the latest version
Abstract
The relationship of Amyotrophic Lateral Sclerosis, Parkinson Disease and other age-related neurodegenerative diseases associated with mitochondrial dysfunction has led to study of the mitochondrial fission gene Drp1 in Drosophila. Drp1 can interact with mitochondrial proteins including parkin and the Drosophila Bcl-2 proteins. Mutations in Drp1 can lead to mitochondrial dysfunction and neuronal loss. Ddc-Gal4 was exploited to direct the expression of Drp1 and the inhibition of Drp1 in neurons. Here, the inhibition of Drp1 seems to compromise locomotor function throughout life but does not alter longevity. The consequences of Drp1 overexpression can be suppressed through overexpression of pro-survival Bcl-2 gene Buffy or by inhibition of the pro-cell death Bcl-2 homologue Debcl. The expression of Buffy suppresses the poor climbing induced by loss of Drp1 function. Interestingly, the loss of parkin function is rescued by the directed inhibition of Drp1. Alteration of the expression of Drp1 acts to phenocopy neurodegenerative disease phenotypes in Drosophila, while overexpression of Buffy can counteract these phenotypes to improve overall health. The pro-survival effect of Buffy can rescue the phenotypic effect due to the Drp1-induced apoptosis. The diminished health due to either overexpression or inhibition of Drp1 has produced a novel model to investigate mechanisms underlying neurodegenerative disease.
Keywords:
Drp1
; Buffy
; Debcl
; parkin
; Drosophila melanogaster
; Amyotrophic Lateral Sclerosis
; Parkinson Disease
; mitochondrial fission
; mitochondrial dysfunction
1. Introduction
The mitochondrial network is critical for many aspects of the subcellular survival mechanisms of organisms. Known to be the powerhouse of the cell, mitochondria are responsible for various aspects of energy homeostasis, oxidative stress, calcium handling, cell signalling, and, thus, cell survival [1]. The dynamic nature of the mitochondria population is critical to the integrity of the subcellular network structures that these organelles maintain and to the control of the quality of mitochondrial proteins and other components [2,3,4]. For example, the early dynamic events that occur during apoptosis include cristae remodeling, mitochondrial fragmentation, and membrane “blebbing” or zeiosis [5]. Inhibition of these processes, either through downregulation directed by RNA-interference or by expression of a dominant-negative mutant form of Drp1, can slow the rate of mitochondrial fragmentation and, in turn, the cascade of apoptotic events [6,7,8]. The activity of the Drp1 protein promotes caspase-independent mitochondrial fission and cristae remodeling to amplify the process of apoptosis whether cell death is instigated by either the specific activity of the pro-apoptotic protein BID or by the general consequences of oxidative stress [9]. Overall, the mechanics of mitochondrial fission plays a crucial role in the amplification of aspects of the essential cellular process of apoptosis.
Modifications to the mitochondrial network seem to differentially influence a number of signalling pathways. In response to a series of molecular cues [10,11,12], the mitochondrial network participates in a delicate balance between continuous division and fusion processes [13,14]. Mitochondrial fusion helps compromised mitochondria, with likely bearing highly damaged DNA and proteins, to actively exchange components with other more healthy mitochondria to decrease the severity of heteroplasmy, and help with functional complementation [15,16]. Mitochondrial fission allows for the segregation of irreversibly damaged portions of the mitochondrial network and subsequent degradation [17]. Mitochondrial fission necessarily requires dynamin-related protein 1 (Drp1) and FIS1 [18]. Nevertheless, an explicit understanding of the factors promoting fission and fusion remains limited.
The Bcl-2 family proteins interact with Drp1, and Drp1 can promote apoptosis in Bcl-2 protein-dependent and independent manners [9,19].The two Bcl-2 family homologues in Drosophila melanogaster are Buffy (anti-apoptotic) and Debcl (pro-apoptotic). The Debcl protein can interacts with Drp1 in Drosophila to activate apoptosis via the JNK pathway [20]. Drp1 is required for a standard rate of Cyt-c release and caspase activation during programmed cell death [21]. Drp1 increases the mitochondrial fragmentation under nitrosative stress in primary cerebrocortical neuron culture [22]. The Drp1 protein interacts with other proteins involved in a number of mitochondrial processes, such as protein product of Bax [10,12,22], the products of Pink1 (PTEN-induced putative kinase 1) and park in Cos-7 cells [23]. Mutation of the Pink1 and park genes are among the most prominent causes of early onset of PD [24]. The roles of Pink1 and park are vital to ubiquitin-dependent mitophagy [25]. The Pink and parkin loss of function mutation increase the mitochondrial localization of Drp1 in flies [26]. The parkin protein ubiquitinates the Drp1 for proteasomal degradation, incriminate the Drp1 for dysregulated mitochondrial dynamics in parkin loss of function induced mitochondrial morphology in HeLa and SH-SY5Y cells [27]. Number of stresses can increase the mitochondrial translocation of Drp1 in neurons and initiate apoptosis or mitophagy [28]. In a number of models of ALS, dephosphorylation of Drp1 through the activity of protein phosphatase 1, has been identified as causative of the disease-associated phenotypes [29]. As well, Drp1-mediated mitochondrial fragmentation caused by the administration of rotenone has been identified in a rat model of PD-like changes to the olfactory bulb [30]. As adjustment of Drp1 activity may be key to the inhibition of the pathology of ALS and PD, investigation of such alterations is essential to knowledge required for the development of therapies. Excessive activity of Drp1 increase mitochondrial fission and consequently promote cell death.
Here we propose that alteration of Drp1 expression results in a degenerative phenotype. We utilize Drosophila to model neurodegenerative disease because it is an excellent model system to study the genes and proteins affected in ALS and PD [31]. The anticipated role of mitochondria in PD pathogenesis has made the study of the interactions of Drp1 gene important to modelling this disease in Drosophila. In these experiments, we exploited the UAS-Gal4 system to direct the overexpression and inhibition of the genes of interest in selected neuronal tissues using the Ddc-Gal4 transgene [32,33]. The proposal is that the Drp1 overexpression phenotype is due to excessive activities related to apoptosis and can be rescued by the appropriate regulation by anti-apoptotic Bcl-2 gene, Buffy. In an established park-RNAi model of PD [34], we directed and inhibited the expression of the Drp1 gene. The ND-like phenotypes of Ddc-Gal4 park-RNAi were rescued by the expression of Drp1-RNAi transgenes. The phenotype produced in response to the expression of Drp1-RNAi is due to the diminishment of mitochondrial integrity and maybe rescued through modification of the responsible signalling pathway. The careful regulation of mitochondrial dynamics is important to control mitochondrial induced defects. The strategy is to identify the basic mechanisms in the fly model to encourage further validation in mammalian model organisms.
2. Materials and Methods
Bioinformatic Analysis
Protein sequences were obtained from the National Center of Biotechnology Information (NCBI) database (https://www.ncbi.nlm.nih.gov/protein/). The conserved domains were identified using NCBI Conserved Domain Database (https://www.ncbi.nlm.nih.gov/cdd/), and Eukaryotic Linear Motif (http://elm.eu.org/). Multiple sequence alignment was done using Clustal Omega (https://www.ebi.ac.uk/Tools/msa/clustalo/) to reveal the conservation of domains. The Homo sapiens Dynamin-1 like protein (DLP-1/Drp1) structure (PDB ID 4BEJ) was obtained from NCBI structure database (https://www.ncbi.nlm.nih.gov/structure/) and Drosophila melanogaster Drp1 protein structure was developed using Phyre2 (http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index) modeling tool. The final models were edited with the PyMOL software (https://pymol.org/2/) to highlight the N-terminus, C-terminus and LIR regions.
Drosophila Stocks and Media
All stocks were maintained on a standard media prepared from cornmeal/molasses/yeast/agar medium treated with propionic acid and methylparaben to resist fungal growth. Aliquots of media were poured into plastic vials, allowed to solidify, and refrigerated at 4°C until used. Stocks are kept at room temperature while crosses and experiments were carried out at 25°C. The UAS-Drp1 (y[1] w[*]; P{w[+mC]=FLAG-FlAsH-HA-Drp1}3, Ki[1]); the UAS-Drp1-RNAi1JF02762 (y[1] v[1]; P{y[+t7.7] v[+t1.8]=TRiP.JF02762}attP2}), UAS-Drp1-RNAi2HMC03230 (y[1] v[1]; P{y[+t7.7]v[+t1.8]=TRiP.HMC03230}attP40), the UAS-Buffy (w[*]; P{w[+mC]=UAS-Buffy.S}E1); UAS-Buffy-RNAi (w[*]; P{w[+mC]=UAS-Buffy.RNAi}3), UAS-DebclEY05743 (y[1] w[67c23]; P{y[+mDint2] w[+mC]=EPgy2}Debcl[EY05743]), Ddc-Gal44.3D(w[1118]; P{w[+mC]=Ddc-GAL4.L}4.3D) and UAS lacZ4-1-2; stocks were obtained from Bloomington Drosophila Stock Center at Indiana University, Bloomington, Indiana, USA. The UAS-Debcl-RNAiv47515 (w1118; P{GD 1637}v47515) was obtained from Vienna Drosophila Resource Center. The UAS-park-RNAi line was obtained from Dr. B. Lu [51,52]. The Ddc-Gal4 park-RNAi line was produced through standard methods [53]. The Ddc-Gal4/CyO; UAS-Drp1/TM3, Ddc-Gal4/CyO; UAS-Drp1-RNAi /TM3, derivative lines were generated using standard recombination methods and used to overexpress or inhibit Drp1 in the selected DA neurons using the Ddc-Gal44.3D transgene.
Ageing Assay
Crosses of select virgin females and males were made, and a cohort of critical class males collected upon eclosion. At least 250 flies were aged per genotype in the cohorts of 25 or less per vial on fresh media, replenished every two to five days to avoid crowding. Flies were observed and scored every two days for the presence of deceased adults. As a rule, flies were considered dead when they did not display movement upon agitation [54]. Longevity data were analyzed using GraphPad Prism version 8 statistical software (graphpad.com), and survival curves were compared by the Mantel-Cox test. Significance were determined at a 95% confidence level (P ≤ 0.05) with Bonferroni correction.
Climbing Assay
A cohort of 70 critical class male flies were collected within 24 hours and maintained as a maximum of ten flies per vial at low density. The food was changed twice every week. Every week 50 males of each genotype were assayed, in groups of 10 or less, for their ability to climb a glass tube divided into five levels of 2 cm each according to standard protocol [54]. The climbing index was calculated for each week using GraphPad prism version 8 statistical software. The climbing curve was fitted using non-linear regression and determined at a 95% confidence interval (P ≤ 0.05).
3. Results
2.1. Drp1 Is Highly Conserved between Homo Sapiens and Drosophila Melanogaster
The D. melanogaster Drp1 protein sequence was sourced from NCBI protein, and the conserved sequences were identified using NCBI CDD. NCBI protein Blast of Drp1 protein of D. melanogaster (NP_608694.2) with the H. sapiens, identified dynamin-1-like protein (isoform 4) (NP_001265392.1), it is 65% identical with a bit score of 957. The multiple sequence alignment of the two proteins derived by Clustal Omega (Figure 1A) shows a highly conserved dynamin-like protein family domain, a dynamin central domain, and a dynamin GTPase effector domain. Two well-documented phosphorylation sites are identified; S606 and S627 in dynamin-1-like protein isoform 4 of H. sapiens; and S616 and T637 in Drp1 of D. melanogaster. A template-based modeling of D. melanogaster Drp1 protein by use of a combination of empirically derived energy functions and physics-based simulated folding was produced using Phyre2. The modeled D. melanogaster Drp1 protein (i) and the H. sapiens Dynamin-1 like protein (ii) from the NCBI database share a near identical structure (Figure 1B). The amino-terminus region of the Drp1 protein is highly conserved and has a consensus LC3-interacting region (LIR) sequence for binding to the ATG8/LC3 protein as determined by the Eukaryotic Linear Motif (ELM) resource. As this protein structure is so highly conserved, it seems very likely that the functions are highly conserved.
2.2. The Overexpression and Inhibition of Drp1 with Ddc-Gal44.3D
In these experiments, the control Ddc-Gal44.3D; UAS-lacZ critical class males were determined to have a median lifespan of 68 days (n=340). The overexpression of Drp1 by the Ddc-Gal4 transgene results in a decreased lifespan of 56 days in 314 flies, much lower compared to the control as determined by log-rank (Mantel-Cox) test with a P-value at <0.0001 (Figure 2A). Inhibition of Drp1 by two distinct RNAi transgenes, via the UAS-Drp1-RNAi1 and UAS-Drp1-RNAi2 directed by the Ddc-Gal4 transgene, results in median lifespans of 70 (n=377) and 72 days (n=323), respectively; very similar to the control (Figure 2A) as determined by log-rank (Mantel-Cox) test with P-value 0.0566 and 0.0213. The non-linear fitting of the climbing curve shows that altering the Drp1 expression have compromised the climbing ability phenotype compared to control at 95% CI (P- value <0.0001) (Figure 2B) (n=50).
2.3. Alteration of the Expression of Drp1 and Drp1-RNAi Directed by Ddc-Gal44.3D
In these experiments, the control Ddc-Gal44.3D; UAS-lacZ critical class males were determined to have a median lifespan of 62 days (n = 308). The directed inhibition of Drp1 by the Ddc-Gal4 transgene results in a greater median lifespan of 70 days in 321 flies compared to the control as determined by log-rank (Mantel-Cox) test with P-value <0.0001 (Figure 3). In contrast, the directed expression of Drp1 by the Ddc-Gal4 transgene results in a reduced median lifespan of 56 days in 255 flies compared to the control as determined by log-rank (Mantel-Cox) test with P-value <0.0001 (Figure 3A). Furthermore, the Ddc-Gal4 UAS-Drp1-RNAi UAS-lacZ critical class males have a median lifespan of 70 days in 310 flies. The overexpression of Drp1 along with UAS-Drp1-RNAi under the direction of the Ddc-Gal4 transgene (Ddc-Gal4; UAS-Drp1-RNAi; UAS-Drp1) has a median lifespan of 64 days, similar to control (Ddc-Gal4; UAS-lacZ) with a P value of 0.0633 as determined by the Log-rank Mantel-Cox test with a Bonferroni correction (Figure 3A). The Ddc-Gal4 UAS-Drp1 UAS-lacZ critical class males have a median lifespan of 58 days in 294 flies. The inhibition of Drp1 along with UAS-Drp1 under the direction of the Ddc-Gal4 transgene (Ddc-Gal4; UAS-Drp1; UAS-Drp1-RNAi) has a median lifespan of 64 days (n=327), similar to control (Ddc-Gal4; UAS-lacZ) with a P value of 0.0582 as determined by the Log-rank Mantel-Cox test with a Bonferroni correction (Figure 3C). The non-linear fitting of the climbing ability curve shows the Drp1 expression and inhibition both have compromised the climbing ability phenotype compared to control at 95% CI (p <0.0001) (Figure 3B & D). The climbing ability curve of Ddc-Gal4 UAS-Drp1-RNAi UAS-Drp1 and Ddc-Gal4 UAS-Drp1 UAS-Drp1-RNAi is very close to the control (Ddc-Gal4; UAS-lacZ) as determined by the non-linear fitting of the climbing curve at a 95% CI at P-value 0.2752 and 0.0589, respectively. The co-expression of Drp1-RNAi along with Drp1 overexpression in flies has resulted in the phenotypes that are similar to the control and suggests that these phenotypes are primarily due the changes in expression of Drp1.
2.4. Alteration of the Expression of Buffy and Debcl in Combination with Drp1 Directed by the Ddc-Gal44.3D Transgene
The control Ddc-Gal44.3D; UAS-Drp1; UAS-lacZ critical class males were determined to have a median lifespan of 58 days (n=282). The overexpression of Buffy along with UAS-Drp1 under the direction of the Ddc-Gal4 transgene (Ddc-Gal4; UAS-Drp1; UAS-Buffy) has a median lifespan of 68 days (n=255), much higher compared to control with a P value of <0.0001 as determined by log-rank (Mantel-Cox) test with a Bonferroni correction. The inhibition of Buffy along with UAS-Drp1 under the direction of the Ddc-Gal4 transgene (Ddc-Gal4; UAS-Drp1; UAS-Buffy-RNAi) has a median lifespan of 52 days (n=274), much less compared to the control (Figure 4A) with a P value 0.0125 as determined by log-rank Mantel-Cox test with a Bonferroni correction. The overexpression of Buffy in neurons rescued the early onset of impairment in the climbing ability of Ddc-Gal4; UAS-Drp1 flies. The non-linear fitting of the climbing curve shows Buffy overexpression has rescued the climbing ability defect compared to control at 95% CI (p <0.0001) (Figure 4B). The inhibition of Buffy by Ddc-Gal4 UAS-Drp1; UAS-Buffy-RNAi further contributes to loss of the climbing ability throughout the life of critical class flies compared to control at 95% CI at a P-value 0.0125 (n=50) (Figure 4B).
The overexpression of Debcl along with UAS-Drp1 under the direction of Ddc-Gal4 transgene (Ddc-Gal4; UAS-Drp1; UAS-Debcl EY05743) has a median lifespan of 60 days (n=331 flies), similar to the control (Figure 4A) with a P value at 0.0114 as determined by log-rank Mantel-Cox test with Bonferroni correction. The inhibition of Debcl, along with UAS-Drp1 under the direction of the Ddc-Gal4 transgene (Ddc-Gal4; UAS-Drp1; UAS-Debcl-RNAi v47515) has a median lifespan of 66 days (n=303), much higher compared to the control (Figure 4A) with a P value at 0.0004 as determined by log-rank Mantel-Cox test with Bonferroni correction. The non-linear fitting of the climbing curve shows Debcl overexpression has no change in the climbing ability defect compared to control at 95% CI (p=0.3293) (Figure 1.4B). The inhibition of Debcl by Ddc-Gal4 UAS-Drp1; UAS-Debcl-RNAi v47515 has rescued the climbing ability throughout the life of critical class flies compared to control at 95% CI at a P-value 0.0057 (n=50) (Figure 1.4B).
2.5. Altering the Expression of Buffy and Debcl along with Drp1-RNAi by Ddc-Gal44.3D Transgene
The control Ddc-Gal44.3D; UAS-Drp1-RNAi; UAS-lacZ critical class males were determined to have a median lifespan of 70 days (n=323). The overexpression of Buffy along with UAS-Drp1-RNAi under the direction of the Ddc-Gal4 transgene (Ddc-Gal4; UAS-Drp1-RNAi; UAS-Buffy) has a median lifespan of 64 days (n=308), much lower compared to control with a P value of <0.0001 as determined by log-rank (Mantel-Cox) test with a Bonferroni correction. The co-inhibition of Buffy and Drp1 under the direction of the Ddc-Gal4 transgene (Ddc-Gal4; UAS-Drp1-RNAi; UAS-Buffy-RNAi) has a median lifespan of 62 days (n=273), much less compared to the control (Figure 5A) with a P value at <0.0001 as determined by log-rank Mantel-Cox test with a Bonferroni correction. The non-linear fitting of the climbing curve shows Buffy overexpression has rescued the climbing ability defect compared to control at 95% CI (p <0.0001) (Figure 5B). The inhibition of Buffy by Ddc-Gal4 UAS-Drp1-RNAi; UAS-Buffy-RNAi further contributes to loss of the climbing ability throughout the life of critical class flies compared to control at 95% CI at a P-value <0.0001 (n=50) (Figure 5B).
The overexpression of Debcl along with UAS-Drp1-RNAi under the direction of Ddc-Gal4 transgene (Ddc-Gal4; UAS-Drp1-RNAi; UAS-Debcl EY05743) has a median lifespan of 68 days (n=156 flies), similar to the control (Figure 1.5A) with a P value at 0.0003 as determined by log-rank Mantel-Cox test with Bonferroni correction. The inhibition of Debcl, along with UAS-Drp1-RNAi under the direction of the Ddc-Gal4 transgene (Ddc-Gal4; UAS-Drp1-RNAi; UAS-Debcl-RNAi v47515) results in a median lifespan of 72 days (n=321), higher compared to the control (Figure 5A) with a P value at 0.0211 as determined by log-rank Mantel-Cox test with Bonferroni correction. The non-linear fitting of the climbing curve shows Debcl overexpression has further increased the climbing ability defect compared to control at 95% CI (p=0.0004) (Figure 5B). The inhibition of Debcl by Ddc-Gal4 UAS-Drp1-RNAi; UAS-Debcl-RNAi v47515 has rescued the climbing ability throughout the life of critical class flies compared to control at 95% CI at a P-value 0.0211 (n=50) (Figure 5B).
2.6. The Altered Co-Expression of Drp1 along with Park-RNAi with Ddc-Gal44.3D
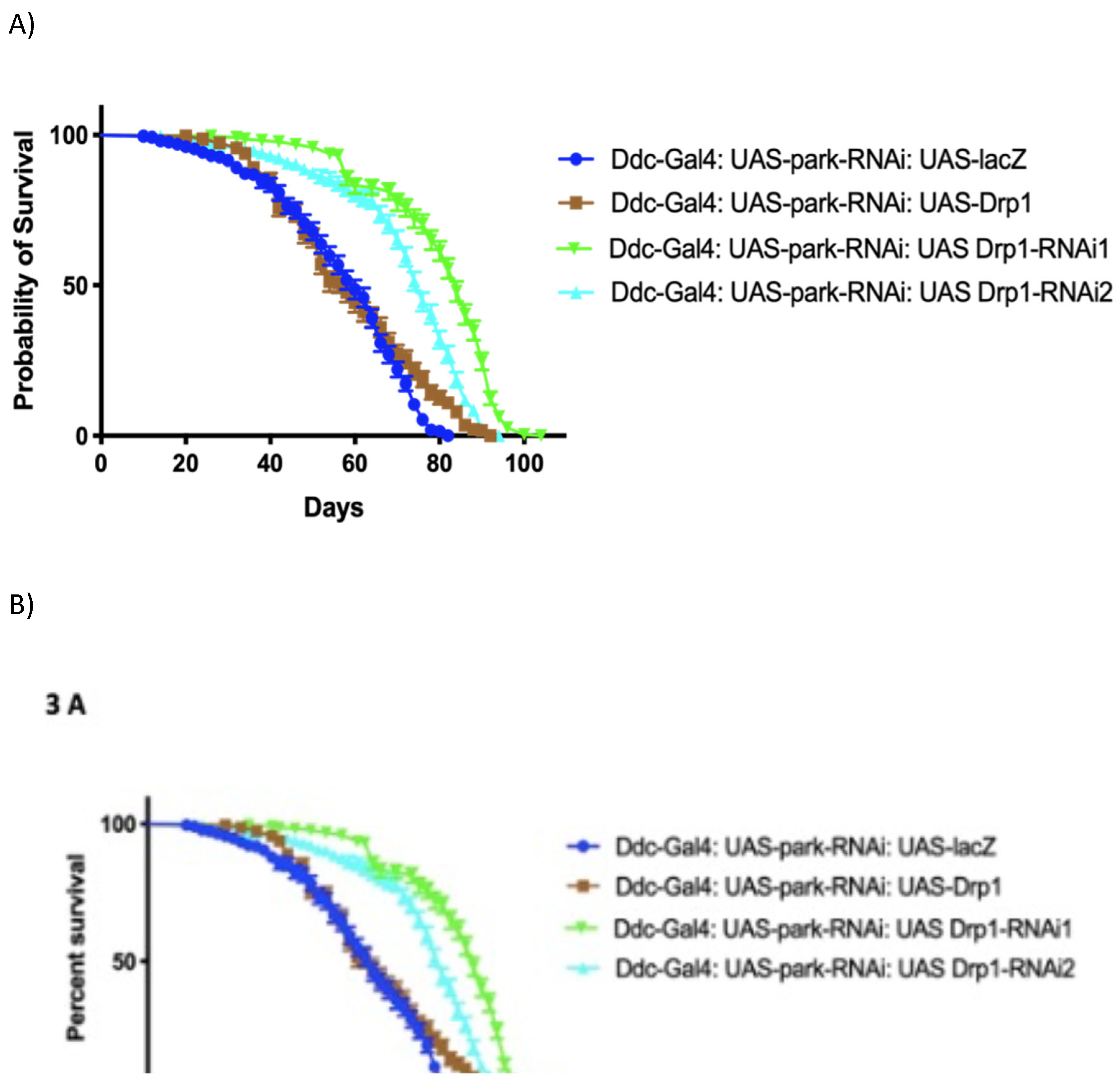
The loss of function of park has led to the establishment of a number of Drosophila models of PD. The Ddc-Gal44.3D park-RNAi UAS-lacZ critical males were determined to have a median lifespan of 60 days (n=259). The overexpression of UAS-Drp1 along with UAS-park-RNAi under the direction of the Ddc-Gal4 transgene has a median life span of 58 days (n=258), similar to the control with a P-value of 0.0026. The inhibition of Drp1 by the two RNAi transgenes, UAS-Drp1-RNAi1JF02762 and UAS-Drp1-RNAi2HMC03230, results in much longer median lifespans of 84 (n=283) and 76 days (n=277) respectively, compared to the control (Figure 6A) as determined by log-rank (Mantel-Cox) test with a P-value at <0.0001. The overexpression of Drp1 by Ddc-Gal4 along with park-RNAi slightly increases the climbing ability over time. However, the locomotor activity of the critical classes with the directed expression of the Drp1-RNAi transgenes are rescued compared to control (Figure 6B).
4. Discussion
The protein product of the Drp1 gene, along with the participation of other mitochondrial protection proteins, is involved in the processes of mitochondrial fission, apoptosis, and mitophagy. Excessive mitochondrial fragmentation can be associated with dysfunctional metabolic diseases whereas a “hyper-fused” mitochondrial network can serve to protect cells from metabolic insult and autophagy [35]. In the skeletal muscle of mice, Drp1 overexpression can cause a severe impairment of post-natal muscle growth as the production of protein may become attenuated and growth hormone pathways may be down regulated [36]. Conditions of high fat and/or high glucose levels can cause excessive oxidative stress along with mitochondrial fragmentation as mediated by the Drp1 protein [37,38]. These phenotypes are similar to the increased activity of Drp1 as observed with Cos and PC12 cells [39]. In humans, protein kinase A (or PKA) can phosphorylate and inactivate the pro-apoptotic Bcl-2 family member protein Bad [40] and the Drp1 protein [41] in a complex effort to promote cell survival. The effect of Drp1 overexpression and consequently excessive mitochondrial fragmentation can be toxic to many physiological processes.
The Bcl-2 family proteins assist the pro-fission activity of the Drp1 protein during apoptosis in nematodes and mammals [42]. However, in non-apoptotic cells of mammals, the Bcl-2 family proteins have both pro-fission and pro-fusion activities. The overexpression of Drp1 in selected neurons along with the overexpression of Buffy or the inhibition of Debcl has resulted in an increase in median lifespan and of the ability to climb over the increased lifespan. In complementary experiments, Buffy inhibition and Debcl overexpression have resulted in reduced lifespans accompanied with impaired climbing abilities, consistent with the conclusion that Buffy can function as the antithesis of Debcl [43]. The rescue of the Drp1 expression phenotype is in accordance with the role of the Buffy protein as “the guardian of mitochondria”. As proteins, Buffy can interact with Debcl to inhibit Debcl-induced cell death. As a mechanism, this process could be due to decreased activity of the Debcl protein to influence cooperation with Drp1 in the promotion of cell death [20]. The pro-apoptotic Debcl protein acts to induces apoptosis through a caspase-independent mechanism that triggers the release of Cytochrome C [44] in an activity that resembles the loss of Drp1 [9]. The overexpression of Debcl and Drp1 together in selected neurons does not alter the phenotype generated by overexpression of Drp1 without Debcl. This may not be surprising as Drp1 and Debcl functions seem to cooperate to promote apoptosis [20]. Indeed, this and earlier studies have demonstrated that Drp1 can play various roles in mitochondrial fragmentation and apoptosis, to act in concert with anti- and pro-survival proteins, dependent upon the stimuli.
The directed inhibition in Drosophila melanogaster of Drp1 in a subset of neurons results in an age-dependent loss in climbing ability, a phenotype strongly associated with the modelling of ND in flies. The overexpression of Buffy in neurons that co-express Drp1-RNAi led to a decrease in the median lifespan accompanied with a rescue of the impaired locomotor ability. The recovery in age-dependent climbing ability over time may be evidence of a complicated regulatory relationship. A study shows Drp1 inhibition reduces the total accumulation of pro-apoptotic Bcl-2 protein, Bax, on mitochondria outer membrane in HeLa cell lines [12]. This intermediate phenotype was not expected but may be important in the determination of the pathology of neurological diseases [45,46]. The inhibition of anti-apoptotic Buffy or overexpression of pro-apoptotic Debcl enhanced the loss of Drp1-induced phenotype. The interaction of Bax with Drp1 in mammals seems to evolve from the Debcl and Drp1 protein interactions during the course of evolution. Drp1 protein interacts directly and indirectly with Bcl-2 family protein to facilitate MOMP in apoptotic cells [42]. Overall, we believe that we have established that Buffy confers survival advantage to flies overexpressing Drp1 and provides a partially rescued intermediate phenotype in flies with a loss of Drp1 function.
The park gene is crucial to the function of Pink1-dependent mitochondrial mitophagy. The loss of the parkin protein is a cause of great cellular stress as a major mechanism that controls mitophagy is compromised. Under normal circumstances, the park-encoded ubiquitin ligase recruits the Drp1 protein to mediate mitochondrial fragmentation during mitophagy [23,47]. In mouse embryonic fibroblasts, loss of park does not produce visible effects upon the net number of mitochondria [48]. Furthermore, the loss of park along with the loss of Drp1 increases the number of mitochondria by threefold, which can be interpreted that park controls mitochondria fragmentation in a Drp1 knockout background. Therefore, park may regulate, negatively, the Drp1-independent mitochondrial division. Alternatively, the parkin protein may direct the ubiquitination of the mitochondrial localised Drp1 protein to lead its proteasomal mediated degradation [26] and, hence, the activity of Drp1 protein on mitochondria is higher in the Drp1-parkin co-inhibition state as compared to the inhibition of Drp1 alone. In these experiments, the critical class flies that have the directed co-expression of Drp1-RNAi and park-RNAi inhibitory transgenes live longer than those that express park-RNAi and Drp1-RNAi under the Ddc-Gal4 transgene individually. This support the hypothesis that the basic mechanism of PD-like phenotypes may be conserved among mammals [49,50] and the Diptera. This could be due to the establishment of an altered mitochondrial network to enhance homeostasis and benefit cellular health. The inhibition of Drp1 has been shown to suppress parkin mutant phenotypes which suggests a role for this protein in 1) in a pathway that regulates mitochondrial health and integrity and 2) in interactions with mitochondrial proteins. Regardless of the underlying mechanism, this work provides strong evidence of Drp1 as an important therapeutic target for maintaining mitochondrial health.
5. Conclusions
Closely associated with cell death pathways in neurons, recently Drp1 has been associated with ALS, Parkinson disease and age-related disease. Our studies demonstrate that the overexpression and inhibition of Drp1 activity in selected neurons can phenocopy some neurodegenerative-like symptoms in Drosophila and, therefore, may represent a novel model of disease. Importantly, the decrease in lifespan and age-dependent loss in climbing ability observed with overexpression of Drp1 in flies is “rescued” to near controls either by overexpression of Buffy or by Debcl inhibition. The age-dependent loss of climbing ability in flies expressing Drp1-RNAi can be rescued by Buffy overexpression or Debcl-RNAi-directed inhibition. The co-inhibition of the parkin with Drp1 results in the rescue of the phenotypes observed, thus it is possible that Drp1 and parkin can participate in cellular pathways that promote cell death. Future studies of these interactions will be required to chart out a pathway for Drp1, Bcl-2 and parkin interaction in Drosophila and, importantly, the molecular changes associated with the loss-of-function of these genes in the development, function and aging of neurons and of the organism.
Author Contributions
All experiments were initiated, conceived, and designed by AH with experimental design, consultation and critical review provided by BES.
Funding
This research is funded by Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant Program, grant number 2016-04828, and the Aging Research Centre of Newfoundland and Labrador (ARC-NL) Research Grant Program, grant number to Dr. Brian E. Staveley, Memorial University of Newfoundland School of Graduate Studies Baseline Funding and Memorial University of Newfoundland Department of Biology Teaching Assistantships.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Galluzzi, L.; Kepp, O.; Kroemer, G. Mitochondria: Master regulators of danger signalling. Nat. Rev. Mol. Cell. Biol. 2012, 13, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Wikstrom, J.D.; Israeli, T.; Bachar-Wikstrom, E.; Swisa, A.; Ariav, Y.; Waiss, M.; Kaganovich, D.; Dor, Y.; Cerasi, E.; Leibowitz, G. AMPK regulates ER morphology and function in stressed pancreatic β-Cells via phosphorylation of DRP1. Mol. Endocrinol. 2013, 27, 1706–1723. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Shirakabe, A.; Maejima, Y.; Zhai, P.; Sciarretta, S.; Toli, J.; Nomura, M.; Mihara, K.; Egashira, K.; Ohishi, M.; Abdellatif, M.; Sadoshima, J. Endogenous Drp1 mediates mitochondrial autophagy and protects the heart against energy stress. Circ. Res. 2015, 116, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, K.; Grimm, A.; Dallmann, R.; Oettinghaus, B.; Restelli, L.M.; Witzig, M.; Ishihara, N.; Mihara, K.; Ripperger, J.A.; Albrecht, U.; Frank, S.; Brown, S.A.; Eckert, A. Circadian control of DRP1 activity regulates mitochondrial dynamics and bioenergetics. Cell Metab. 2018, 27, 657–666. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Lu, N.; Zhou, Z. Cellular and nuclear degradation during apoptosis. Curr. Opin. Cell Biol. 2009, 21, 900–912. [Google Scholar] [CrossRef] [PubMed]
- Estaquier, J.; Arnoult, D. Inhibiting Drp1-mediated mitochondrial fission selectively prevents the release of cytochrome c during apoptosis. Cell Death Differ. 2007, 14, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhou, F.; Zhang, Z.; Xing, D. Bax is essential for Drp1-mediated mitochondrial fission but not for mitochondrial outer membrane permeabilization caused by photodynamic therapy. J. Cell. Physiol. 2011, 226, 530–541. [Google Scholar] [CrossRef]
- Wang, P.; Wang, P.; Liu, B.; Zhao, J.; Pang, Q.; Agrawal, S.G.; Jia, L.; Liu, F.-T. Dynamin-related protein Drp1 is required for Bax translocation to mitochondria in response to irradiation-induced apoptosis. Oncotarget 2015, 6, 22598–22612. [Google Scholar] [CrossRef] [PubMed]
- Oettinghaus, B.; D’Alonzo, D.; Barbieri, E.; Restelli, L.M.; Savoia, C.; Licci, M.; Tolnay, M.; Frank, S.; Scorrano, L. DRP1-dependent apoptotic mitochondrial fission occurs independently of BAX, BAK and APAF1 to amplify cell death by BID and oxidative stress. Biochim. Biophys. Acta. 2016, 1857, 1267–1276. [Google Scholar] [CrossRef]
- Karbowski, M.; Lee, Y.-J.; Gaume, B.; Jeong, S.-Y.; Frank, S.; Nechushtan, A.; Santel, A.; Fuller, M.; Smith, C.L.; Youle, R.J. Spatial and temporal association of Bax with mitochondrial fission sites, Drp1, and Mfn2 during apoptosis. J. Cell Biol. 2002, 159, 931–938. [Google Scholar] [CrossRef]
- Hoitzing, H.; Johnston, I.G.; Jones, N.S. What is the function of mitochondrial networks? A theoretical assessment of hypotheses and proposal for future research. BioEssays 2015, 37, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.E.; Grosser, J.A.; Fehrman, R.L.; Schlamp, C.L.; Nickells, R.W. Completion of BAX recruitment correlates with mitochondrial fission during apoptosis. Sci. Rep. 2019, 9, 16565. [Google Scholar] [CrossRef]
- Lee, S.; Jeong, S.Y.; Lim, W.C.; Kim, S.; Park, Y.Y.; Sun, X.; Youle, R.J.; Cho, H. Mitochondrial fission and fusion mediators, hFis1 and OPA1, modulate cellular senescence. J. Biol. Chem. 2007, 282, 22977–22983. [Google Scholar] [CrossRef]
- Scott, I.; Youle, R.J. Mitochondrial fission and fusion. Essays Biochem. 2010, 47, 85–98. [Google Scholar] [CrossRef]
- Nakada, K.; Sato, A.; Hayashi, J.I. Mitochondrial functional complementation in mitochondrial DNA-based diseases. Int. J Biochem. Cell. Biol. 2009, 41, 1907–1913. [Google Scholar] [CrossRef]
- Chan, D.C. Fusion and fission: interlinked processes critical for mitochondrial health. Annu. Rev. Genet. 2012, 46, 265–287. [Google Scholar] [CrossRef]
- Twig, G.; Elorza, A.; Molina, A.J.A.; Mohamed, H.; Wikstrom, J.D.; Walzer, G.; Stiles, L.; Haigh, S.E.; Katz, S.; Las, G.; Alroy, J.; Wu, M.; Py, B.F.; Yuan, J.; Deeney, J.T.; Corkey, B.E.; Shirihai, O.S. Fission and selective fusion govern mitochondrial segregation and elimination by autophagy. EMBO J. 2008, 27, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Sebastián, D.; Palacín, M.; Zorzano, A. Mitochondrial dynamics: coupling mitochondrial fitness with healthy aging. Trends Mol. Med. 2017, 23, 201–215. [Google Scholar] [CrossRef]
- Cassidy-Stone, A.; Chipuk, J.E.; Ingerman, E.; Song, C.; Yoo, C.; Kuwana, T.; Kurth, M.J.; Shaw, J.T.; Hinshaw, J.E.; Green, D.R.; Nunnari, J. Chemical inhibition of the mitochondrial division dynamin reveals its role in Bax/Bak-dependent mitochondrial outer membrane permeabilization. Dev. Cell 2008, 14, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Clavier, A.; Ruby, V.; Rincheval-Arnold, A.; Mignotte, B.; Guénal, I. The Drosophila retinoblastoma protein, Rbf1, induces a Debcl- and Drp1-dependent mitochondrial apoptosis. J. Cell Sci. 2015, 128, 3239–3249. [Google Scholar] [CrossRef]
- Breckenridge, D.G.; Kang, B.-H.; Kokel, D.; Mitani, S.; Staehelin, L.A.; Xue, D. Caenorhabditis elegans drp-1 and fis-2 regulate distinct cell-death execution pathways downstream of ced-3 and independent of ced-9. Mol. Cell 2008, 31, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Gerencser, A.A.; Liot, G.; Lipton, S.A.; Ellisman, M.; Perkins, G.A.; Bossy-Wetzel, E. Mitochondrial fission is an upstream and required event for bax foci formation in response to nitric oxide in cortical neurons. Cell Death Differ. 2007, 14, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Buhlman, L.; Damiano, M.; Bertolin, G.; Ferrando-Miguel, R.; Lombès, A.; Brice, A.; Corti, O. Functional interplay between Parkin and Drp1 in mitochondrial fission and clearance. Biochim. Biophys. Acta 2014, 1843, 2012–2026. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Westenberger, A. Genetics of Parkinson’s disease. Cold Spring Harb. Perspect. Med. 2012, 2, a008888. [Google Scholar] [CrossRef] [PubMed]
- Pickrell, A.M.; Youle, R.J. The roles of PINK1, Parkin, and mitochondrial fidelity in parkinson’s disease. Neuron 2015, 85, 257–273. [Google Scholar] [CrossRef]
- Poole, A.C.; Thomas, R.E.; Yu, S.; Vincow, E.S.; Pallanck, L. The mitochondrial fusion-promoting factor mitofusin is a substrate of the PINK1/Parkin pathway. PLoS One 2010, 5, e10054. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Song, P.; Du, L.; Tian, W.; Yue, W.; Liu, M.; Li, D.; Wang, B.; Zhu, Y.; Cao, C.; Zhou, J.; Chen, Q. Parkin ubiquitinates Drp1 for proteasome-dependent degradation. J. Biol. Chem. 2011, 286, 11649–11658. [Google Scholar] [CrossRef]
- Pradeep, H.; Sharma, B.; Rajanikant; G. K. Drp1 in ischemic neuronal death: an unusual suspect. Curr. Med. Chem. 2014, 21, 2183–2189. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Lee, J.-H.; Chung, A.-Y.; Jo, Y.; Shin, J.-H.; Park, H.-C.; Kim, H.; Lopez-Gonzalez; J. R., Ryu; R., Sun; W. Prevention of mitochondrial impairment by inhibition of protein phosphatase 1 activity in amyotrophic lateral sclerosis. Cell Death Dis. 2020, 11, 888. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, W.; Shao, Q.; Yang, Y.; Xu, Z.; Chen, J.; Zhang, X.; Xiaoqun, G. Drp1, a potential therapeutic target for Parkinson's disease, is involved in olfactory bulb pathological alteration in the Rotenone-induced rat model. Toxicol Lett. 2020, 325, 1–13. [Google Scholar] [CrossRef]
- Xiong, Y.; Yu, J. Modeling Parkinson’s disease in Drosophila: What have we learned for dominant traits? Front. Neurol. 2018, 9, 228. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chaney, S.; Forte, M.; Hirsh, J. Ectopic g-protein expression in dopamine and serotonin neurons blocks cocaine sensitization in Drosophila melanogaster. Curr. Biol. 2000, 10, 211–214. [Google Scholar] [CrossRef]
- Riemensperger, T.; Issa, A.R.; Pech, U.; Coulom, H.; Nguyễn, M.V.; Cassar, M.; Jacquet, M.; Fiala, A.; Birman, S. A single dopamine pathway underlies progressive locomotor deficits in a Drosophila model of Parkinson Disease. Cell Rep. 2013, 5, 952–960. [Google Scholar] [CrossRef]
- M'Angale, P.G.; Staveley, B.E. Overexpression of Buffy enhances the loss of parkin and suppresses the loss of Pink1 phenotypes in Drosophila. Genome 2017, 60, 241–247. [Google Scholar] [CrossRef]
- Wai, T.; Langer, T. Mitochondrial dynamics and metabolic regulation. Trends Endocrinol. Metab. 2016, 27, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Touvier, T.; De Palma, C.; Rigamonti, E.; Scagliola, A.; Incerti, E.; Mazelin, L.; Thomas, J.-L.; D’Antonio, M.; Politi, L.; Schaeffer, L.; Clementi, E.; Brunelli, S. Muscle-specific Drp1 overexpression impairs skeletal muscle growth via translational attenuation. Cell Death Dis. 2015, 6, e1663. [Google Scholar] [CrossRef]
- Yu, T.; Robotham, J.L.; Yoon, Y. Increased production of reactive oxygen species in hyperglycemic conditions requires dynamic change of mitochondrial morphology. Proc. Natl. Acad. Sci. USA 2006, 103, 2653–2658. [Google Scholar] [CrossRef]
- Sun, Y.; Ge, X.; Li, X.; He, J.; Wei, X.; Du, J.; Sun, J.; Li, X.; Xun, Z.; Liu, W.; Zhang, H.; Wang, Z.Y.; Li, Y.C. High-fat diet promotes renal injury by inducing oxidative stress and mitochondrial dysfunction. Cell Death Dis. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cribbs, J.T.; Strack, S. Reversible phosphorylation of Drp1 by cyclic AMP-dependent protein kinase and calcineurin regulates mitochondrial fission and cell death. EMBO Rep. 2007, 8, 939–944. [Google Scholar] [CrossRef]
- Virdee, K.; Parone, P.A.; Tolkovsky, A.M. Phosphorylation of the pro-apoptotic protein BAD on serine 155, a novel site, contributes to cell survival. Curr. Biol. 2000, 10, 1151–1154. [Google Scholar] [CrossRef]
- Cereghetti, G.M.; Stangherlin, A.; Martins De Brito, O.; Chang, C.R.; Blackstone, C.; Bernardi, P.; Scorrano, L. Dephosphorylation by calcineurin regulates translocation of Drp1 to mitochondria. Proc. Natl. Acad. Sci. USA 2008, 105, 15803–15808. [Google Scholar] [CrossRef] [PubMed]
- Rolland, S.G.; Conradt, B. New role of the BCL2 family of proteins in the regulation of mitochondrial dynamics. Curr. Opin. Cell Biol. 2010, 22, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Quinn, L.; Coombe, M.; Mills, K.; Daish, T.; Colussi, P.; Kumar, S.; Richardson, H. Buffy, a Drosophila Bcl-2 protein, has anti-apoptotic and cell cycle inhibitory functions. EMBO J. 2003, 22, 3568–3579. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Huang, Q.; Ke, N.; Matsuyama, S.; Hammock, B.; Godzik, A.; Reed, J.C. Drosophila pro-apoptotic Bcl-2/Bax homologue reveals evolutionary conservation of cell death mechanisms. J. Biol. Chem. 2000, 275, 27303–27306. [Google Scholar] [CrossRef] [PubMed]
- Leuchter, A.F.; Hunter, A.M.; Krantz, D.E.; Cook, I.A. Intermediate phenotypes and biomarkers of treatment outcome in major depressive disorder. Dialogues Clin. Neurosci. 2014, 16, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Civelek, M.; Lusis, A.J. Systems genetics approaches to understand complex traits. Nat. Rev. Genet. 2014; 15, 34–48. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, C.; Cai, J.; Chen, G.; Zhang, D.; Zhang, Z.; Dong, Z. PINK1/Parkin-mediated mitophagy is activated in cisplatin nephrotoxicity to protect against kidney injury. Cell Death Dis. 2018, 9, 1113. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Itoh, K.; Iijima, M.; Sesaki, H. Parkin suppresses Drp1-independent mitochondrial division. Biochem. Biophys. Res. Commun. 2016, 475, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Rappold, P.M.; Cui, M.; Grima, J.C.; Fan, R.Z.; De Mesy-Bentley, K.L.; Chen, L.; Zhuang, X.; Bowers, W.J.; Tieu, K. Drp1 inhibition attenuates neurotoxicity and dopamine release deficits in vivo. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Filichia, E.; Hoffer, B.; Qi, X.; Luo, Y. Inhibition of Drp1 mitochondrial translocation provides neural protection in dopaminergic system in a Parkinson’s disease model induced by MPTP. Sci. Rep. 2016, 6, 32656. [Google Scholar] [CrossRef]
- Yang, Y.; Nishimura, I.; Imai, Y.; Takahashi, R.; Lu, B. Parkin suppresses dopaminergic neuron-selective neurotoxicity induced by Pael-R in Drosophila. Neuron 2003, 37, 911–924. [Google Scholar] [CrossRef]
- Yang, Y.; Gehrke, S.; Imai, Y.; Huang, Z.; Ouyang, Y.; Wang, J.W.; Yang, L.; Beal, M.F.; Vogel, H.; Lu, B. Mitochondrial pathology and muscle and dopaminergic neuron degeneration caused by inactivation of Drosophila Pink1 is rescued by Parkin. Proc. Natl. Acad. Sci. USA 2006, 103, 10793–10798. [Google Scholar] [CrossRef] [PubMed]
- M'Angale, P.G.; Staveley, B.E. Bcl-2 homologue Debcl enhances a-synuclein-induced phenotypes in Drosophila. PeerJ 2016, 4, e2461. [Google Scholar] [CrossRef] [PubMed]
- Todd, A.; Staveley, B. Expression of Pink1 with α-synuclein in the dopaminergic neurons of Drosophila leads to increases in both lifespan and healthspan. Genet. Mol. Res. 2012, 11, 1497–1502. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Drp1 is evolutionarily conserved between Drosophila and humans. (a) Clustal Omega multiple sequence alignment of D. melanogaster Drp1 (NP_608694.2) protein with the H. sapiens (NP_001265392.1) shows evolutionarily conserved domains identified using the NCBI Conserved Domain Database (CDD) and further confirmed by the Eukaryotic Linear Motif (ELM) resource. The two well documented phosphorylation sites are identified, S606 and S627 in dynamin-1-like protein (DLP-1) isoform 4 of H. sapiens; and S616 and T637 in Drp1 of D. melanogaster. The asterisks indicate the residues that are identical; the colons indicate the conserved substitutions; and the dots indicates the semi-conserved substitutions. Colour differences indicate the chemical nature of amino acids: red indicates small hydrophobic (includes aromatic) residues; blue indicates acidic; magenta indicates basic; and green indicates basic with hydroxyl or amine groups. (bi) The original Dynamin-1 like protein (DLP-1) structure of H. sapiens (NP_001265392.1) from the NCBI structure database. (bii) The Phyre2 web portal for protein modelling, prediction and analysis mediated the development of a model of the Drp1 protein of D. melanogaster (NP_608694.2) from a 76% identical protein with a confidence of 100%. The N terminus is coloured in Magenta; C terminus is coloured in Red and a consensus ATG8 binding region at N terminus is coloured in orange.
Figure 1.
Drp1 is evolutionarily conserved between Drosophila and humans. (a) Clustal Omega multiple sequence alignment of D. melanogaster Drp1 (NP_608694.2) protein with the H. sapiens (NP_001265392.1) shows evolutionarily conserved domains identified using the NCBI Conserved Domain Database (CDD) and further confirmed by the Eukaryotic Linear Motif (ELM) resource. The two well documented phosphorylation sites are identified, S606 and S627 in dynamin-1-like protein (DLP-1) isoform 4 of H. sapiens; and S616 and T637 in Drp1 of D. melanogaster. The asterisks indicate the residues that are identical; the colons indicate the conserved substitutions; and the dots indicates the semi-conserved substitutions. Colour differences indicate the chemical nature of amino acids: red indicates small hydrophobic (includes aromatic) residues; blue indicates acidic; magenta indicates basic; and green indicates basic with hydroxyl or amine groups. (bi) The original Dynamin-1 like protein (DLP-1) structure of H. sapiens (NP_001265392.1) from the NCBI structure database. (bii) The Phyre2 web portal for protein modelling, prediction and analysis mediated the development of a model of the Drp1 protein of D. melanogaster (NP_608694.2) from a 76% identical protein with a confidence of 100%. The N terminus is coloured in Magenta; C terminus is coloured in Red and a consensus ATG8 binding region at N terminus is coloured in orange.

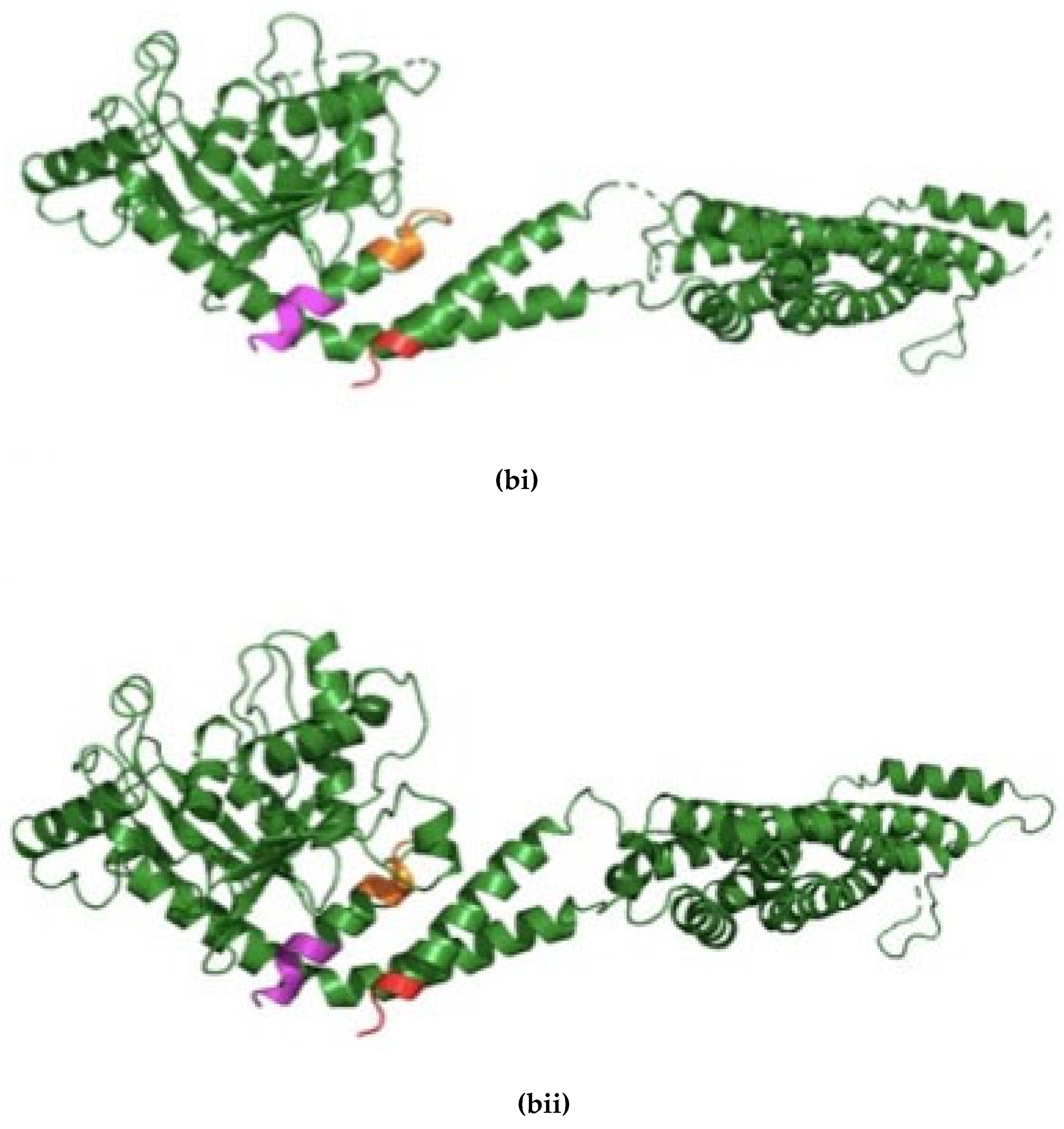
Figure 2.
Altered Drp1 expression under the control of Ddc-Gal44.3D influences the survival and climbing ability of flies. (a) The GraphPad prism8 generated graph of the longevity assay for the expression of Drp1, Drp1 RNAi’s under the control of Ddc-Gal4 transgene. The directed expression results in decreased median lifespan of 56 days compare to 68 days of control calculated by Log-rank Mantel Cox test, with Bonferroni correction. The inhibition of Drp1 under the control of Ddc-Gal4 transgene results in lifespan of 70 days with UAS-Drp1-RNAi1 and 72 days with UAS-Drp-RNAi2 compared to 68 days of control done by Log-rank Mantel Cox test, with Bonferroni correction. (b) The GraphPad prism8 generated graph of the climbing abilities of flies with overexpression of Drp1, Drp1 RNAi’s and control. The climbing ability of Drp1 overexpression and Drp1 RNAi’s flies have decreased compared to control as determined in nonlinear fitting of the climbing curve by 95% confidence interval (p-value <0.0001).
Figure 2.
Altered Drp1 expression under the control of Ddc-Gal44.3D influences the survival and climbing ability of flies. (a) The GraphPad prism8 generated graph of the longevity assay for the expression of Drp1, Drp1 RNAi’s under the control of Ddc-Gal4 transgene. The directed expression results in decreased median lifespan of 56 days compare to 68 days of control calculated by Log-rank Mantel Cox test, with Bonferroni correction. The inhibition of Drp1 under the control of Ddc-Gal4 transgene results in lifespan of 70 days with UAS-Drp1-RNAi1 and 72 days with UAS-Drp-RNAi2 compared to 68 days of control done by Log-rank Mantel Cox test, with Bonferroni correction. (b) The GraphPad prism8 generated graph of the climbing abilities of flies with overexpression of Drp1, Drp1 RNAi’s and control. The climbing ability of Drp1 overexpression and Drp1 RNAi’s flies have decreased compared to control as determined in nonlinear fitting of the climbing curve by 95% confidence interval (p-value <0.0001).
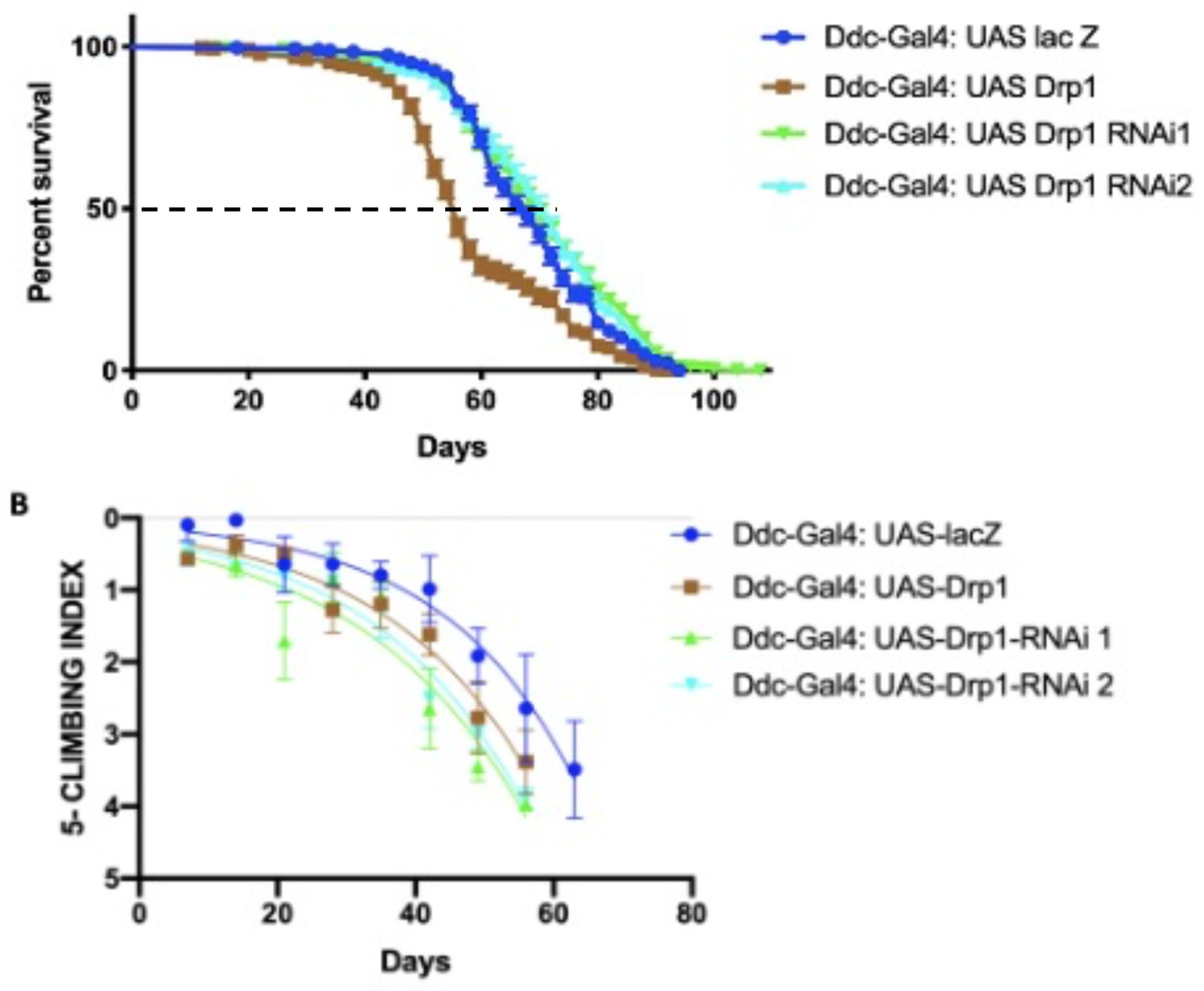
Figure 3.
The expression of Drp1-RNAi, directed by Ddc-Gal44.3D, can increase median lifespan and decrease climbing. (a) In control, Ddc-Gal44.3D UAS-lacZ critical class males resulted in a median life span of 62 days (n=308). Expression of Drp1 in Ddc-Gal44.3D resulted in a median life span of 56 days (n=310), much lower than the lacZ-expressing control; expression of Drp1 in Ddc-Gal44.3D UAS-Drp1-RNAi resulted in a median life span of 64 days (n=250) very similar to control (Ddc/lacZ) as determined by the Log-rank Mantel-Cox test (p value=0.0633) with Bonferroni correction. The graph of the longevity assay was generated by GraphPad prism8. (b) The Ddc-Gal44.3D flies express UAS-lacZ in control flies. The climbing abilities of Ddc-Gal44.3D UAS-Drp1 expressing flies have decreased compared to control as determined in the non-linear fitting of the climbing curve by a 95% confidence interval (p<0.0001). The flies' climbing ability expressing Drp1 in Ddc-Gal44.3D UAS-Drp1-RNAi transgene is similar to control as determined in the non-linear fitting of the climbing curve by a 95% confidence interval at P value=1.309. The graph of longevity assay was generated by GraphPad prism8 non-linear regression curve. (c) In control, Ddc-Gal44.3D UAS-lacZ critical class males resulted in a median life span of 62 days (n=308). Expression of Drp1-RNAi in Ddc-Gal44.3D resulted in a median life span of 70 days (n=321), much higher compared to the control; expression of Drp1-RNAi in Ddc-Gal44.3D UAS-Drp1 resulted in a median life span of 64 days (n=327), very similar to control (Ddc/lacZ) as determined by the Log-rank Mantel-Cox test (p value=0.0582) with Bonferroni correction. The graph of the longevity assay was generated by GraphPad prism8. (d) The Ddc-Gal44.3D flies express UAS-lacZ in control flies. The climbing abilities of Ddc-Gal44.3D UAS-Drp1-RNAi expressing flies have decreased compared to control as determined in the non-linear fitting of the climbing curve by a 95% confidence interval (p<0.0001). The flies' climbing ability expressing Drp1-RNAi in Ddc-Gal44.3D UAS-Drp1 transgene is similar to control (Ddc/lacZ) as determined in the non-linear fitting of the climbing curve by a 95% confidence interval at p value=0.0027. The graph of longevity assay was generated by GraphPad prism8 non-linear regression curve.
Figure 3.
The expression of Drp1-RNAi, directed by Ddc-Gal44.3D, can increase median lifespan and decrease climbing. (a) In control, Ddc-Gal44.3D UAS-lacZ critical class males resulted in a median life span of 62 days (n=308). Expression of Drp1 in Ddc-Gal44.3D resulted in a median life span of 56 days (n=310), much lower than the lacZ-expressing control; expression of Drp1 in Ddc-Gal44.3D UAS-Drp1-RNAi resulted in a median life span of 64 days (n=250) very similar to control (Ddc/lacZ) as determined by the Log-rank Mantel-Cox test (p value=0.0633) with Bonferroni correction. The graph of the longevity assay was generated by GraphPad prism8. (b) The Ddc-Gal44.3D flies express UAS-lacZ in control flies. The climbing abilities of Ddc-Gal44.3D UAS-Drp1 expressing flies have decreased compared to control as determined in the non-linear fitting of the climbing curve by a 95% confidence interval (p<0.0001). The flies' climbing ability expressing Drp1 in Ddc-Gal44.3D UAS-Drp1-RNAi transgene is similar to control as determined in the non-linear fitting of the climbing curve by a 95% confidence interval at P value=1.309. The graph of longevity assay was generated by GraphPad prism8 non-linear regression curve. (c) In control, Ddc-Gal44.3D UAS-lacZ critical class males resulted in a median life span of 62 days (n=308). Expression of Drp1-RNAi in Ddc-Gal44.3D resulted in a median life span of 70 days (n=321), much higher compared to the control; expression of Drp1-RNAi in Ddc-Gal44.3D UAS-Drp1 resulted in a median life span of 64 days (n=327), very similar to control (Ddc/lacZ) as determined by the Log-rank Mantel-Cox test (p value=0.0582) with Bonferroni correction. The graph of the longevity assay was generated by GraphPad prism8. (d) The Ddc-Gal44.3D flies express UAS-lacZ in control flies. The climbing abilities of Ddc-Gal44.3D UAS-Drp1-RNAi expressing flies have decreased compared to control as determined in the non-linear fitting of the climbing curve by a 95% confidence interval (p<0.0001). The flies' climbing ability expressing Drp1-RNAi in Ddc-Gal44.3D UAS-Drp1 transgene is similar to control (Ddc/lacZ) as determined in the non-linear fitting of the climbing curve by a 95% confidence interval at p value=0.0027. The graph of longevity assay was generated by GraphPad prism8 non-linear regression curve.
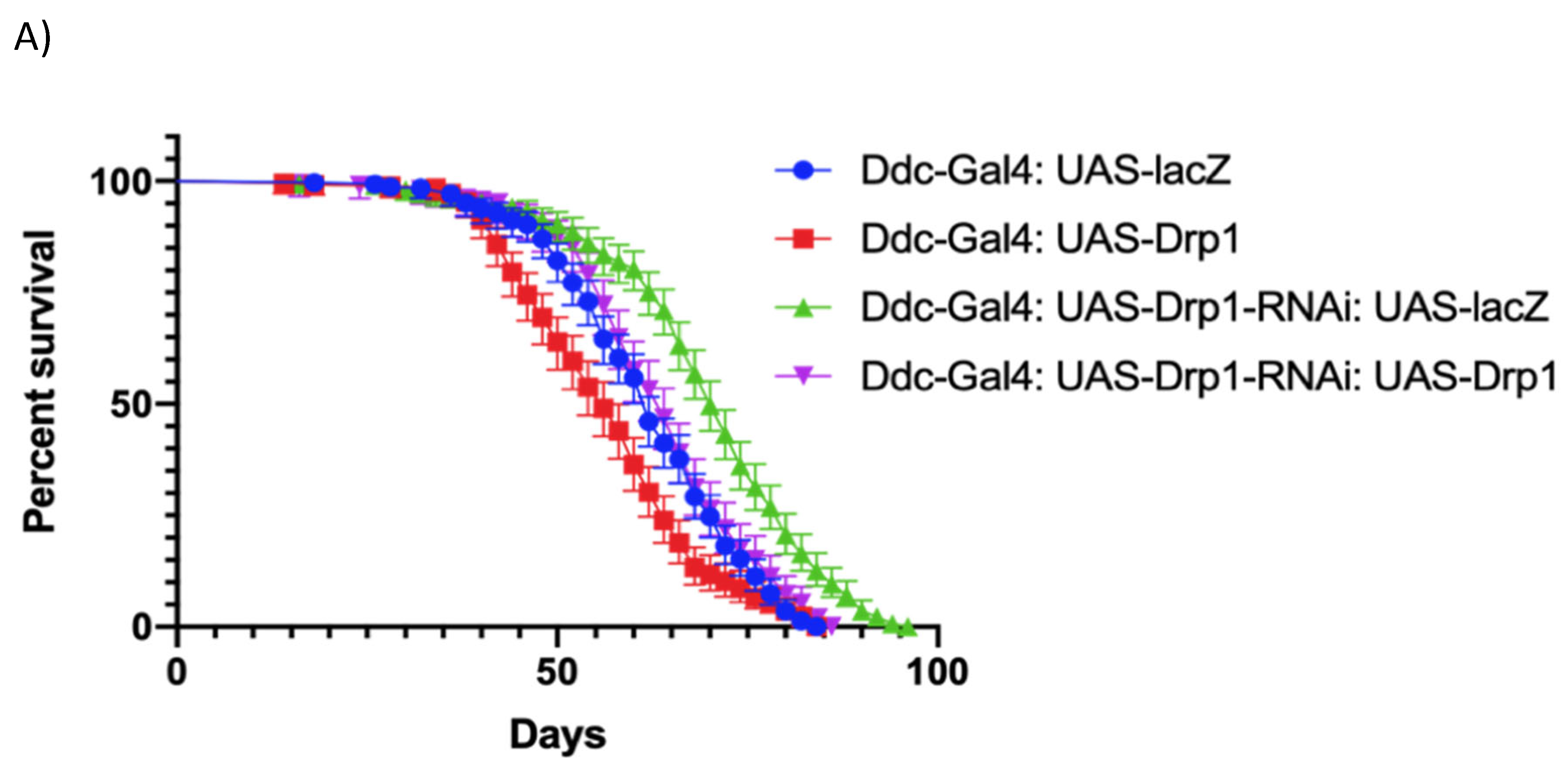
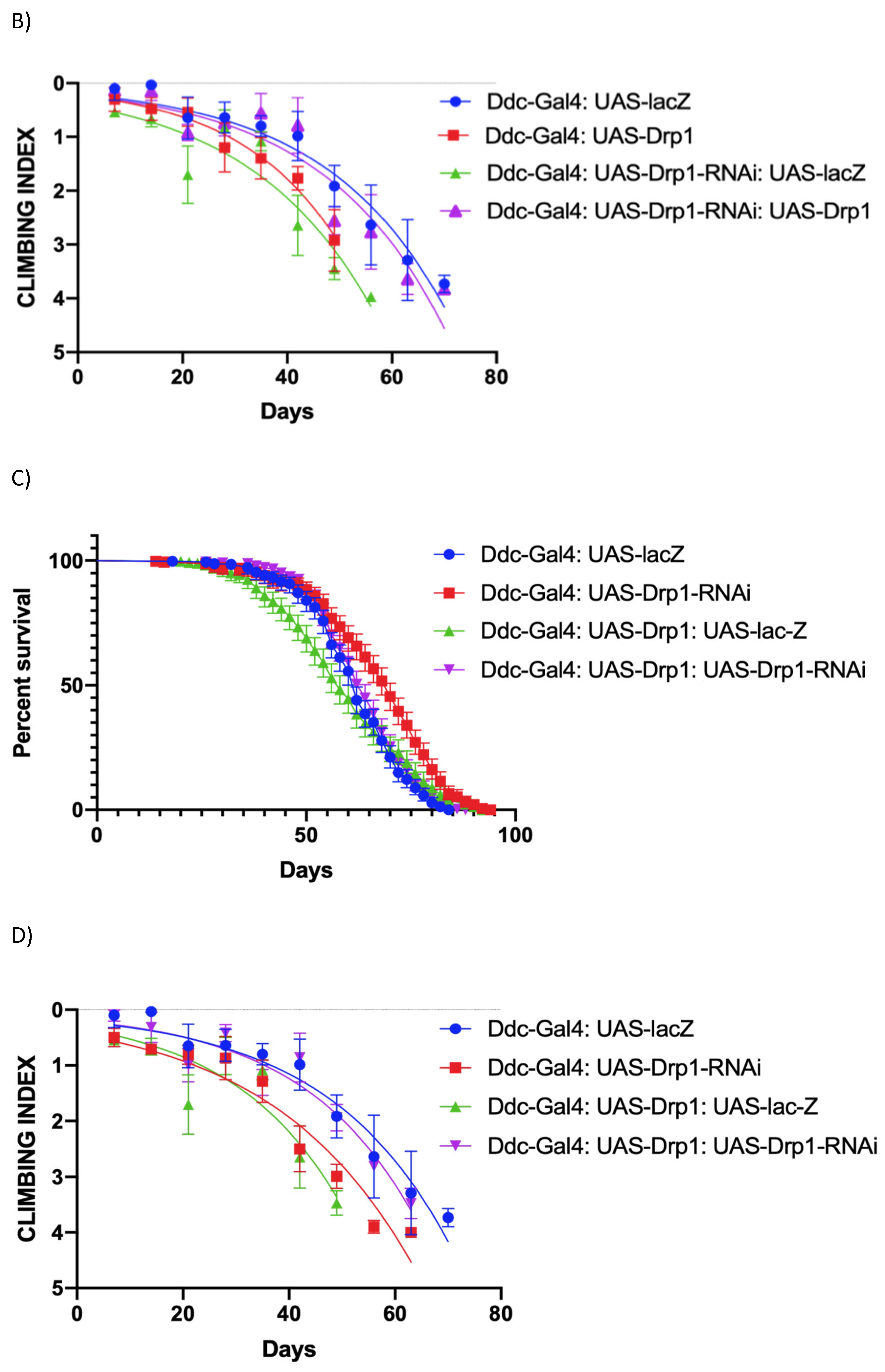
Figure 4.
Altered expression of Buffy and Debcl can enhance and suppress climbing ability in Drp1 over-expression flies. (a) In control, Ddc-Gal44.3D; UAS-Drp1 UAS-lacZ critical class males resulted in a median life span of 58 days (n=282). The overexpression of Buffy results in a median lifespan of 68 days (n=375) compares to 58 days of control (P value=0.0002); the inhibition of Buffy directed by the Ddc-Gal44.3D UAS-Drp1 transgene results in the median lifespan of 52 (n=274), much less compared to control, determined by Log-rank Mantel-Cox test at P-value <0.0001, with Bonferroni correction. The overexpression of DebclEY05743 results in a median lifespan of 60 days (n=331) similar to 58 days of control determined by Log-rank Mantel-Cox test at P-value 0.3293; the inhibition of Debcl directed by the Ddc-Gal4 UAS-Drp1 transgene result in the median lifespan of 66 (n=303); much higher than control, determined by Log-rank Mantel-Cox test at P value 0.0057, with Bonferroni correction. (b) The GraphPad prism8 generated graph of the climbing abilities of Ddc-Gal4 -Drp1 flies with the expression of Buffy, Buffy-RNAi, DebclEY05743, Debcl-RNAiv47515 and control. The climbing abilities of flies overexpressing Buffy have rescued compared to control as determined in the climbing curve's non-linear fitting by a 95% confidence interval (p<0.0001). The climbing ability of the flies was further weakened by the expression of UAS-Buffy-RNAi as determined in the non-linear fitting of the climbing curve by a 95% confidence interval at a p-value 0.0125 and 0.03293 respectively (n=50). The climbing abilities of flies expressing Debcl-RNAiv47515has rescued compared to control as determined by the non-linear fitting of the climbing curve by a 95% confidence interval (p value=0.0057). The graph of longevity assay was generated by GraphPad prism8 non-linear regression curve.
Figure 4.
Altered expression of Buffy and Debcl can enhance and suppress climbing ability in Drp1 over-expression flies. (a) In control, Ddc-Gal44.3D; UAS-Drp1 UAS-lacZ critical class males resulted in a median life span of 58 days (n=282). The overexpression of Buffy results in a median lifespan of 68 days (n=375) compares to 58 days of control (P value=0.0002); the inhibition of Buffy directed by the Ddc-Gal44.3D UAS-Drp1 transgene results in the median lifespan of 52 (n=274), much less compared to control, determined by Log-rank Mantel-Cox test at P-value <0.0001, with Bonferroni correction. The overexpression of DebclEY05743 results in a median lifespan of 60 days (n=331) similar to 58 days of control determined by Log-rank Mantel-Cox test at P-value 0.3293; the inhibition of Debcl directed by the Ddc-Gal4 UAS-Drp1 transgene result in the median lifespan of 66 (n=303); much higher than control, determined by Log-rank Mantel-Cox test at P value 0.0057, with Bonferroni correction. (b) The GraphPad prism8 generated graph of the climbing abilities of Ddc-Gal4 -Drp1 flies with the expression of Buffy, Buffy-RNAi, DebclEY05743, Debcl-RNAiv47515 and control. The climbing abilities of flies overexpressing Buffy have rescued compared to control as determined in the climbing curve's non-linear fitting by a 95% confidence interval (p<0.0001). The climbing ability of the flies was further weakened by the expression of UAS-Buffy-RNAi as determined in the non-linear fitting of the climbing curve by a 95% confidence interval at a p-value 0.0125 and 0.03293 respectively (n=50). The climbing abilities of flies expressing Debcl-RNAiv47515has rescued compared to control as determined by the non-linear fitting of the climbing curve by a 95% confidence interval (p value=0.0057). The graph of longevity assay was generated by GraphPad prism8 non-linear regression curve.
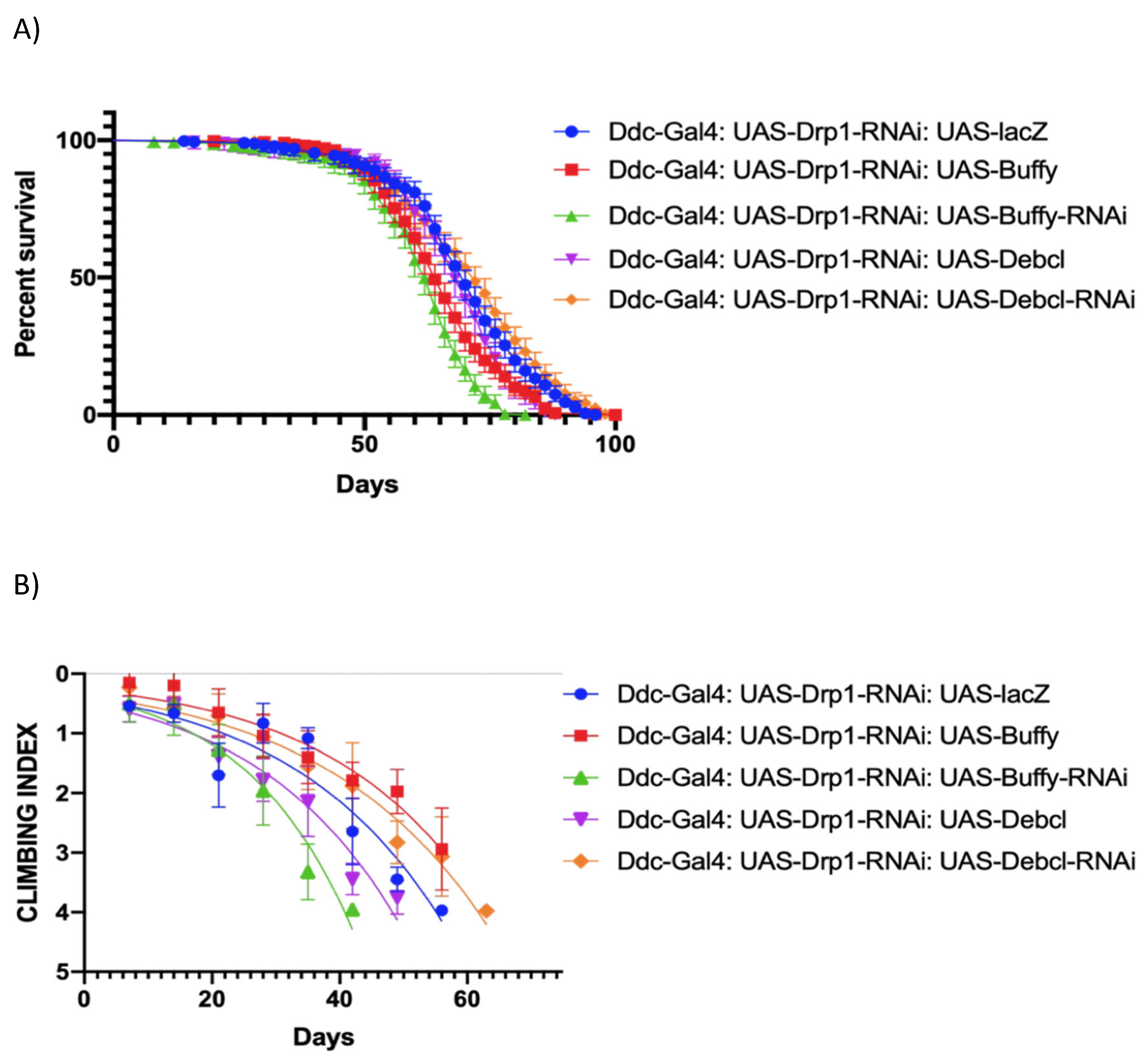
Figure 5.
Altered expression of Buffy and Debcl can enhance and suppress climbing ability in Drp1 loss of function flies. (a) In control, Ddc-Gal44.3D; UAS-Drp1 transgene results in the median lifespan of 62 (n=273), determined by Log-rank Mantel-Cox test at P-value <0.0001, with Bonferroni correction. The overexpression of DebclEY05743 results in a median lifespan of 68 days (n=331) much higher compared to control as determined by Log-rank Mantel-Cox test at P-value 0.0003; the inhibition of Debcl directed by the Ddc-Gal4; UAS-Drp1 transgene result in the median lifespan of 72 (n=303); similar to control, determined by Log-rank Mantel-Cox test at p-value 0.021, with Bonferroni correction. (b) The GraphPad prism8 generated graph of the climbing abilities of Ddc-Gal4 UAS-Drp1 flies with the expression of Buffy, Buffy-RNAi, DebclEY05743, Debcl-RNAiv47515 and control. The climbing abilities of flies overexpressing Buffy have rescued compared to control as determined in the climbing curve's non-linear fitting by a 95% confidence interval (p<0.0001). The climbing ability of the flies has further diminished through the expression of UAS-Buffy-RNAi and UAS-DebclEY05743 as determined in the non-linear fitting of the climbing curve by a 95% confidence interval at a p-value 0.0004 and 0.0002 respectively (n=50). The climbing abilities of flies expressing Debcl-RNAiv47515has rescued compared to control as determined by the non-linear fitting of the climbing curve by a 95% confidence interval (p value<0.0001). The graph of longevity assay was generated by GraphPad prism8 non-linear regression curve.
Figure 5.
Altered expression of Buffy and Debcl can enhance and suppress climbing ability in Drp1 loss of function flies. (a) In control, Ddc-Gal44.3D; UAS-Drp1 transgene results in the median lifespan of 62 (n=273), determined by Log-rank Mantel-Cox test at P-value <0.0001, with Bonferroni correction. The overexpression of DebclEY05743 results in a median lifespan of 68 days (n=331) much higher compared to control as determined by Log-rank Mantel-Cox test at P-value 0.0003; the inhibition of Debcl directed by the Ddc-Gal4; UAS-Drp1 transgene result in the median lifespan of 72 (n=303); similar to control, determined by Log-rank Mantel-Cox test at p-value 0.021, with Bonferroni correction. (b) The GraphPad prism8 generated graph of the climbing abilities of Ddc-Gal4 UAS-Drp1 flies with the expression of Buffy, Buffy-RNAi, DebclEY05743, Debcl-RNAiv47515 and control. The climbing abilities of flies overexpressing Buffy have rescued compared to control as determined in the climbing curve's non-linear fitting by a 95% confidence interval (p<0.0001). The climbing ability of the flies has further diminished through the expression of UAS-Buffy-RNAi and UAS-DebclEY05743 as determined in the non-linear fitting of the climbing curve by a 95% confidence interval at a p-value 0.0004 and 0.0002 respectively (n=50). The climbing abilities of flies expressing Debcl-RNAiv47515has rescued compared to control as determined by the non-linear fitting of the climbing curve by a 95% confidence interval (p value<0.0001). The graph of longevity assay was generated by GraphPad prism8 non-linear regression curve.
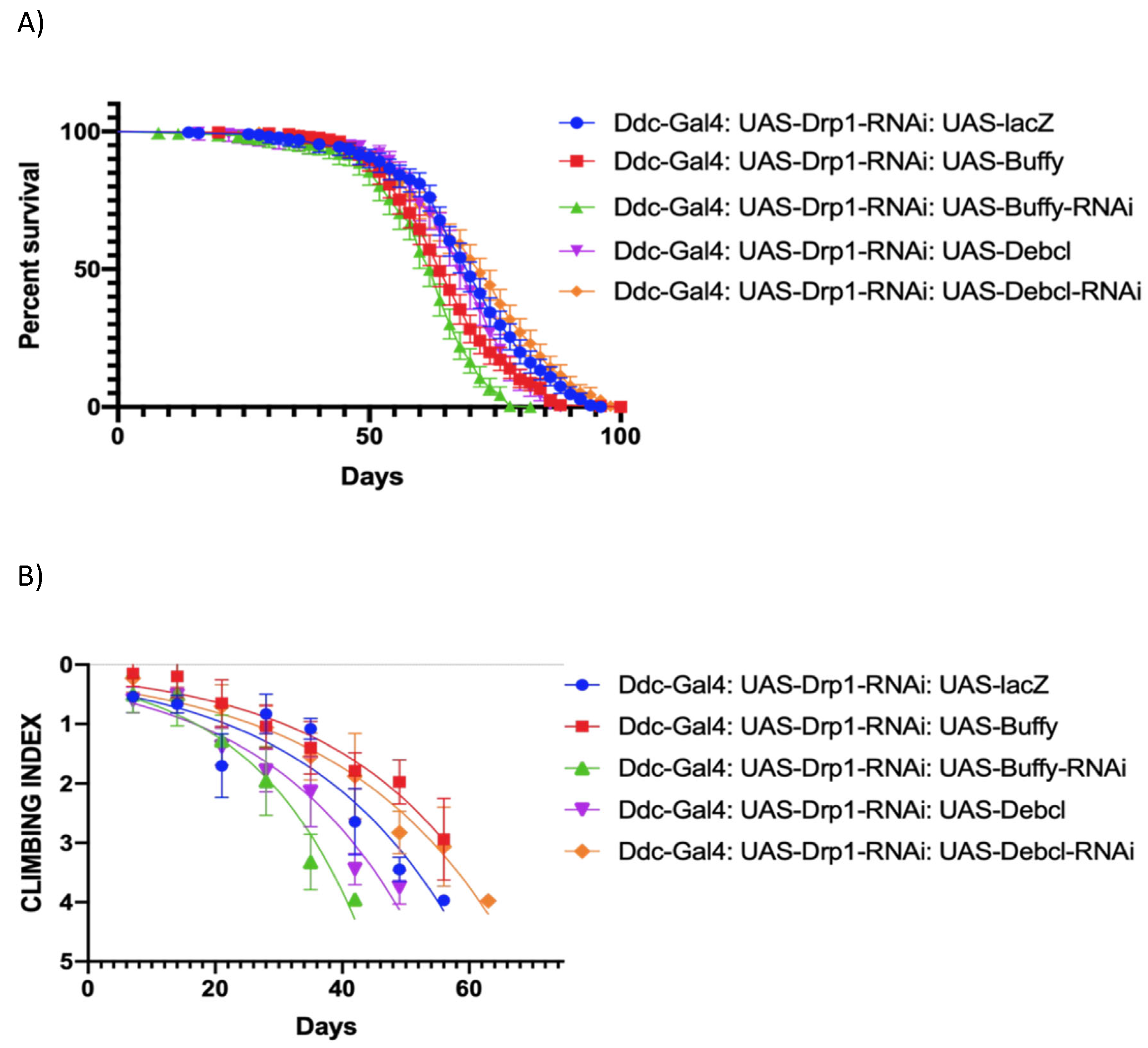
Figure 6.
Inhibition of Drp1 expression enhances lifespan and climbing ability of parkin loss of function in Ddc-Gal44.3D park-RNAi flies. (a) The graph of longevity assay generated by GraphPad prism8 with altered Drp1 expression in Ddc-Gal4 park-RNAi expressing flies. The overexpression results in median lifespan of 58 days similar to 60 days of control (lacZ/park-RNAi) determined by Log-rank Mantel-Cox test, with Bonferroni correction. The inhibition of Drp1 in neurons using Ddc-Gal4 transgene along with park-RNAi results in increased lifespan of 84 days with UAS-Drp1-RNAi1 and lifespan of 76 days with UAS-Drp1-RNAi2 compare to 60 days of control done by Log-rank Mantel Cox test, with Bonferroni correction. (b) The GraphPad prism8 generated graph of the climbing abilities of flies with overexpression of Drp1, Drp1 RNAi and control. The climbing abilities of and Drp1 RNAi flies has rescued compared to control as determined in non-linear fitting of the climbing curve by 95% confidence interval.
Figure 6.
Inhibition of Drp1 expression enhances lifespan and climbing ability of parkin loss of function in Ddc-Gal44.3D park-RNAi flies. (a) The graph of longevity assay generated by GraphPad prism8 with altered Drp1 expression in Ddc-Gal4 park-RNAi expressing flies. The overexpression results in median lifespan of 58 days similar to 60 days of control (lacZ/park-RNAi) determined by Log-rank Mantel-Cox test, with Bonferroni correction. The inhibition of Drp1 in neurons using Ddc-Gal4 transgene along with park-RNAi results in increased lifespan of 84 days with UAS-Drp1-RNAi1 and lifespan of 76 days with UAS-Drp1-RNAi2 compare to 60 days of control done by Log-rank Mantel Cox test, with Bonferroni correction. (b) The GraphPad prism8 generated graph of the climbing abilities of flies with overexpression of Drp1, Drp1 RNAi and control. The climbing abilities of and Drp1 RNAi flies has rescued compared to control as determined in non-linear fitting of the climbing curve by 95% confidence interval.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



