
Submitted:
15 July 2024
Posted:
16 July 2024
You are already at the latest version
Abstract
Climate change and the increasing frequency and severity of drought events pose significant challenges for sustainable agriculture worldwide. Soil microorganisms, both beneficial and pathogenic, play a crucial role in mediating plant-environment interactions and shaping the overall functioning of agroecosystems. This review synthesizes the current knowledge on the contrasting adaptive mechanisms utilized by different groups of plant-soil microorganisms focusing on beneficial and pathogenic bacterial and fungal communities in response to drought and desiccation stresses. The review examines the common survival strategies employed by microbes specifically rhizobacteria and arbuscular mycorrhizal fungi, such as the production of osmoprotectants, altered gene expression, and biofilm formation. It also highlights the distinct adaptive mechanisms of pathogenic versus mutualistic microbes, with pathogens tending to prioritize virulence factors and suppress plant growth, while beneficial microbes enhance plant growth and stress tolerance. Genetic exchange such as horizontal gene transfer (HGT) is identified as a key adaptive mechanism, allowing both pathogenic and non-pathogenic microbes to acquire beneficial traits like stress tolerance and virulence factors. Environmental stressors like drought can promote increased genetic exchange and the spread of pathogenic traits within the soil microbiome. The complex interplay between drought-adapted microbes and their interactions with plants is discussed, emphasizing the need for a deeper understanding of soil microbiome dynamics under climate change. This knowledge can be utilized in sustainable agricultural practices to mitigate the impacts of drought on plant health and productivity. This review provides insights into the divergent survival strategies of soil microorganisms in response to drought and desiccation, for managing the resilience of agroecosystems to climate change.
Keywords:
soil
; plant
; microorganisam
; gene transfer
; drought
; desiccation
1. Introduction
Soil is an important living ecosystem that supports all forms of terrestrial life [1]. In one g of soil, there are thousands of microbial taxa: viruses, bacteria, archaea, protists, and fungi, which form stable microbial communities that participate in nutrient cycling, transformation of organic matter, modification of biochemical and biophysical properties of soil, bioaccumulation of inorganic compounds, bioremediation of contaminated soils, and other activities or functions? [2]. The escalating prevalence of climate-induced drought and desiccation poses formidable challenges to soil microbiomes and the intricate ecosystems they support. The remarkable diversity of microbial communities within soil ecosystems is fundamental to the sustainable functioning of soil ecosystems, thereby facilitating efficient plant growth, development, and ultimately, crop yields [3]. Within these microbial communities, interactions among members manifest in diverse forms, including mutualism, competition, antagonism, commensalism, amensalism, or neutralism [4]. Moreover, their distribution within the soil volume is heterogeneous; notably, around plant roots in the rhizosphere, microbial cell concentrations, particularly comprising rhizobacteria and mycorrhizal fungi, exhibit significantly higher densities [5]. This heightened presence of rhizosphere microorganisms around plant roots plays a pivotal role in assisting plants to withstand various biotic and abiotic stresses, including diseases and drought.
Mankind is accustomed to the idea that drought is a common natural phenomenon that occurs periodically in almost all climatic zones. There are many places on Earth where water availability is limited and in the absence of precipitation for several weeks there is a so-called meteorological drought, and for a longer period - agricultural drought is associated with crop losses [6]. According to various estimates, arid regions occupy between 26.3% and 36.3% of the land area, occurring on all seven continents [7, 8]. In these regions, not only plants but also soil microorganisms face the possibility of desiccation during which the water content of cells can decrease to 0.1 g H2O g-1 dry mass, corresponding to no free water in the cytoplasm [9].
The projected climate change towards an increase in the average daily temperature on the planet by several degrees, will inevitably lead to an increase in the area of drylands and will negatively affect agriculture [10]. In this situation, it is particularly important to study the effects of drought and desiccation stress on soil microorganisms, both plant beneficials and pathogens, as well as on their relationships within microbial communities, and interactions with plants. Physiological, structural and genetic adaptation mechanisms of soil microorganisms allow them to adapt and survive under unfavorable drought conditions [11]. Microorganisms detect the slightest changes in environmental parameters, which are signals for them to act, and quickly respond by regulating the expression of their genes [12]. However, the adaptive responses of pathogenic and beneficial soil microbes during drought are different.
Prokaryotic bacteria, eukaryotic fungi and oomycetes are common members of soil-borne plant pathogen communities and can survive for many years in the absence of host plants [13]. However, we have little understanding of how drought affects the survival and reproduction of pathogenic microorganisms. Pathogens are known to survive the abiotic stress of drought in different ways. Some representatives increase their pathogenic activity in wet conditions and reduce it in dry soils, while others, on the contrary, are active during dry periods [14]. To cope with drought, all pathogens develop their adaptation strategies associated with the formation of a large number of stable structures e.g., microsclerotia, sclerotia, chlamydospores, and oospores, the production of pathogenicity factors, and a wide range of enzymes.
In most soils, there is a certain level of pathogen suppression (general soil suppression) due to competitive interactions between antagonist microorganisms [15]. Therefore, under normal conditions, plant infection is inhibited in soils [16]. Plant disease outcome depends on complex ecological interactions and communications between plants and pathogens, between plant microbiomes and pathogens, and between plants and their microbiomes [17]. The plant microbiome controls pathogens through a range of mechanisms including modulation of immune responses, competition with pathogens for resources and space, production of antifungal effectors, lytic enzymes and secondary metabolites (antibiotics, bacteriocins, toxins and siderophores) [18]. However, abiotic factors such as drought led to a reduced immune response of agricultural plants, making them less resistant to attack by soil pathogens [13]. In this situation, plants are assisted by beneficial soil microorganisms inhabiting the rhizosphere region - rhizobacteria and arbuscular mycorrhizal fungi. In addition to protecting microbial cells during drought due to changes in cell wall structure, production of protective substances, etc., they help plants to survive dry periods through numerous mechanisms [11, 19,20]. Despite the large amount of information accumulated in this field, some aspects of it are poorly understood. Many specific mechanisms of communication molecules in complex ecological interactions between plants and all members of the microbiome under drought conditions remain unclear [17].
New data on the survival mechanisms of rhizobacteria and AMF during drought are constantly emerging, but there has been no comparative analysis of these mechanisms. Also lacking is a comparison of adaptive drought survival mechanisms in beneficial and pathogenic soil microorganisms, which will help to provide new insights into sustainable agronomy. This paper addresses several critical aspects: (1) Investigating the complex interactions between plant roots and soil microbes under climate-induced drought and desiccation stresses, (2) Analyzing the distinct adaptive strategies of plant pathogenic and mutualistic bacterial and fungal communities in response to drought, (3) Examining how soil microbiomes mediate plant defence mechanism under drought and desiccation stresses, and (4) Exploring the role of horizontal gene transfer in the adaptation of both pathogenic and non-pathogenic microbes to drought stress.
2. Complex Interplay between Plant Roots and Soil Microbes under Drought and Desiccation Stresses
2.1. Microbial Community Structure Affected by Drought
As climate change escalates, marked by phenomena like drought and desiccation primarily caused by water deficits, the intricate dynamics within soil microbiomes are experiencing significant shifts. Scientists have long studied the effects of these phenomena on natural ecosystems, and most often on soil microbiota, as it is directly related to crop drought tolerance in particular and successful agronomy in general [21]. And currently, with global warming threatening to transform ecosystems, interest in this issue is increasing [22]. Scientists still do not know the answer to the question of which has a greater impact on soil processes and microbial activity: the drying process or subsequent moistening [23]. It is also unknown what the long-term effects on microbial communities will be from changing climatic conditions [24].
Notable progress has been made in the understanding of the influence of dry periods on the number and composition of soil microbial communities and their interactions with plants [22,25]. Increasing aridity has been shown to reduce soil microbial diversity and abundance [26], as well as microbial community structure and function [27]. It has also been noted in many studies that in most cases, it is not the drought period itself, but the constant change in drought-wetting regimes that has a greater impact on the microbial soil community [22,27].
Microbial cells must adapt when soils dry out, and reduce energy consumption, synthesize osmoprotectants (trehalose, glycine-betaene, amino compounds, etc.), structurally change the composition of phospholipids and fatty acids of the cell membrane, etc. [28]. In doing so, those species, and even strains that can do it better, will dominate the communities subjected to desiccation [29]. Most studies point to the advantages of gram-positive bacteria (apparently due to a thick layer of cell wall peptidoglycan) and their increase during drought periods in both soil and plant rhizosphere [30]. For example, Polish scientists showed that drought and rewetting stress changed the structure of the soil bacterial community in favor of Gram-positive bacteria of taxonomic categories Actinobacteria and Firmicutes, while the relative abundance of Gram-negative bacteria in most cases decreased, especially affecting representatives of Gammaproteobacteria and Bacteroidetes [31]. A study of Australian soil communities showed that during prolonged drought, an increase in the relative abundance of microorganisms belonging to oligotrophic taxa such as Actinobacteria, Alphaproteobacteria, Planctomycetes, Ascomycota and Basidiomycota occurred [32]. Whereas in wet periods, very different taxa such as Acidobacteria, TM7, Gemmatimonadates and Chytridiomycota dominated [32]. During dry periods, soils of Austrian mountain meadows were also dominated by specialized members of Actinobacteriota, especially of the genus Streptomyces. [33]. At the same time, >90% of bacterial and archaeal taxa stopped dividing altogether or divided at significantly reduced growth rates [33]. Water limitation during drought and the associated increase in soil C/N ratio in Swiss forest soils promoted the spread of desiccation-tolerant oligotrophic taxa and caused a shift in the potential lifestyle of microorganisms from symbiotic to saprotrophic [34]. Sometimes, scientists observed unusual facts, for example, in the soils of the hyper-arid Atacama Desert on the west coast of South America (territory of Chile), one of the most life-hostile habitats on the planet, evidence of a metabolically active community of archaea was found during periods of wetting [35]. In a study by Chilakala et al. a drought-induced increase in phototroph populations in both the soil and root microbiome was observed [36]. This indicates the complex ways in which drought affects microbial composition and dynamics in the plant-soil system [37].
Metagenomic studies by Chilakala et al. showed that in the rhizosphere zone, drought enhances the fungal microbiota by suppressing virus and algal populations [36]. In addition, prokaryotic communities were found to be less resistant to water limitation than fungal communities in most other studies [34]. Compared to bacteria, fungi have unique survival skills and physiological structures that enhance their drought tolerance [38]. However, in some cases, the opposite situation was also observed, apparently related to the predominance of drought-tolerant bacterial species such as Pseudomonas spp. and Bacillus spp. in the soil microbiome. These bacteria appeared to adapt to dehydration through their ability to form biofilms, produce stress-sensitive proteins, and utilize diverse nutrient sources even under challenging conditions [39].
The adaptive response of the soil microbiome to drought is affected not only by the lack of moisture but also by other parameters: soil composition (nitrogen and carbon content and ratio), pH, presence of heavy metals, etc. [2,39]. Dehydration stress has been shown to affect microorganisms more strongly in soils with low organic matter content [40]. Ambient air temperature and atmospheric CO2 content are also relevant [41]. For example, Metze et al. experimentally modelled what would happen to the soil microbial community under drought conditions during a six-year climate warming period involving a 3 °C increase in temperature and a 300-ppm increase in atmospheric CO2 concentration [33]. It was demonstrated that in this scenario, there would be a 2.3-fold increase in the number of taxa undergoing active division (from 4% to 9%, compared to 35% under normal conditions), indicating the adaptation of numerous species to adverse conditions.
Plants and multicellular fungi can regulate the composition of the bacterial community of the rhizosphere by root exudation of various substances into the soil during drought [42, 43]. For example, it was shown that the cactus Echinocactus platyacanthus in Mexico during drought periods changes the profile of amino acids secreted by its roots, which in turn restructures the microbial community in its rhizosphere in favor of bacteria belonging to the type Armatimonadota, classes Actinobacteria and Dehalococcoidia, families Gemmataceae and Nitrosococcaceae [44]. The fungi Rhizophagus and Serendipita have a protective effect on the bacterial communities of their hyphosphere, which are subject to water limitation [45].
Soil microorganisms can increase plant resistance to drought through the production of substances (polysaccharides, indoleacetic acid, proline, etc.) that improve soil structure and its water-holding capacity [46]. However, their effect is insufficient during prolonged periods of dehydration, which is especially evident in the yield reduction of legume crops dependent on nitrogen fixation processes associated rhizobacteria [47]. Drying severely limits atmospheric nitrogen fixation by these bacteria and results in yield loss [48]. In this case, additional inoculation of plant roots with different species of rhizobacteria belonging to the genera Pseudomonas, Bacillus, Rhizobium, Enterobacter, Azospirillum, Azotobacter, Arthrobacter, Mesorhizobium, Xanthomonas, Acinetobacter, Alcaligenes, Burkholderia, Erwinia and Flavobacterium help plants cope with the effects of drought by enhancing their growth, root elongation, production or inhibition of phytohormones, and production of volatile organic compounds [49].
Currently, drought stands out as one of the most perilous abiotic stresses affecting soil microorganisms, diminishing their activity and population while disrupting community composition. The adverse impacts of drought on soil microbiota result in decreased soil fertility and crop productivity. With the anticipation of more frequent droughts due to climate warming, research on innovative strategies to enhance the drought tolerance of microorganisms, thereby aiding plants, is of paramount importance. Mechanisms enabling microbes to endure and proliferate amidst diminishing moisture levels could introduce new functional groups into the ecosystem, offering substantial advantages to agriculture [43].
2.2. Rhizobacteria-Plants Interactions Affected by Drought
The rhizosphere, a small zone around roots influenced by exudates and root secretions, contains 10-100 times more microorganisms than the rest of the soil (over 1000 1011 microbial cells per gram of root) [5,50]. These are mainly rhizobacteria, of which there are more than 30,000 species [50], and mycorrhizal fungi (mainly arbuscular fungi (AMF), about 250 species [51], which are obligate symbionts of 70-90% of plant species [52]. Rhizobacteria and mycorrhizal fungi inhabit root tissue as endophytes or on the surface of plant roots (rhizoplane) and represent an important part of the soil microbiota [53]. Microorganisms participate in biological nitrogen fixation, convert high molecular weight soil organic polymers into organic, inorganic and amino acids, sugars, vitamins, purines, nucleosides, etc., produce phytohormones, redistribute the flow of nutrients throughout the soil-plant system, which helps plant growth [54]. Also, rhizobial microorganisms exert biocontrol and increase plant resistance to biotic (pathogens) and abiotic (drought) stresses. [55,56]. In addition, they participate in other biophysical and biochemical soil processes of ecological importance and interact with the rest of the soil microbiota [57,58]. This group of microorganisms is very important in terms of efficient agronomy (confined agriculture without chemicals).
The response of rhizobia microorganisms to desiccation stress began to be studied about 100 years ago. This was due to their use as seed inoculum in agriculture [59]. At the same time, the attention of scientists was directed to investigate the effects of growth medium, pH and temperature on the survival of cells in the dry state and to study the conditions for the effective use of dry cell preparations in agronomy [60]. The increasing need for the use of inoculants in agriculture has led to the discovery of a large number of soil microorganisms (bacteria and fungi) that somehow contribute to plant growth and stress tolerance [61]. Particular attention has been focused on rhizosphere-dwelling bacteria, which in the late 1970s came to be called “plant growth-promoting rhizobacteria” or PGPR for short [62]. This group of bacteria includes several genera such as Pseudomonas, Enterobacter, Bacillus, Variovax, Klebsiella, Paraburkholderia, Azospirillum, Herbaspirillum, Gluconacetobacter, Serratia, Azotobacter and others. They are widely distributed in soils throughout the globe, phylogenetically diverse and often not species-specific to the host plant [54,63]. Among rhizobial microorganisms, it is the bacteria that are believed to play a crucial role in helping the host plant under drought conditions [64]. Whereas, in the case of arbuscular mycorrhizal fungi (AMF), their dependence on host plants and susceptibility to changes in water availability may limit their protective potential during drought periods [65].
The number of studies on the effects of drought on rhizobial microorganisms is steadily increasing [66]. Rhizobacteria and AMF have been shown to have biochemical and physiological defence mechanisms that are activated during periods of drought and allow them not only to cope with its effects but also to protect the host plant [67,68]. Microbiomes have been found to make critical contributions to plant adaptation and induce plant adaptability to combined biotic (pathogens) and abiotic (drought) stresses, especially in arid climates [69,70].
3. Adaptive Strategies Employed by Pathogenic and Beneficial Microbes under Drought
3.1. Adaptive Mechanisms of Rhizobacteria under Drought
Adaptive strategies of rhizobacteria in response to climatic stressors include cellular, genetic and enzymatic changes. Under drought conditions, changes in the cellular structure and metabolic activity of rhizobacteria are observed, affecting their survival and function under water-limited conditions [71,72]. Moreover, the main mechanism that allows rhizobacteria to survive under stress conditions is the alteration of gene expression toward the synthesis of protective substances such as trehalose, proline, glycine-betaine, glutamate, mannitol, hydroxylation, exopolysaccharides, heat shock proteins, and proteins involved in cell membrane protection, DNA damage repair, and response to oxidative stress [73]. A metagenomic study by Bulgarelli et al. identified specific genes involved in mechanisms of responses to various abiotic stresses? [74].
Cytryn et al. showed that desiccation of cells of Bradyrhizobium japonicum, a gram-negative soil bacterium that converts atmospheric nitrogen to ammonia by forming a symbiotic association through soybean root nodules, results in differential expression of 15 to 20% of 8453 genes within 3 days [75]. The induction of trehalose-6-phosphate synthase (otsA), trehalose-6-phosphate phosphatase (otsB), and trehalose synthase (treS) genes, which encode two of the three trehalose synthesis pathways, is more than 2-fold increased, correlating with an increase in its intracellular concentration. The authors also showed that the expression of genes encoding isocitrate lyase, responses to oxidative stress, exopolysaccharide synthesis and transport, heat shock proteins, enzymes for nucleic acid modification and repair, and the synthesis of pili and flagella increased under desiccation conditions [75]. Zhu et al. also demonstrated that different types of genes are differentially expressed in Bradyrhizobium japonicum cells during drought at10% relative humidity and subsequent rehydration. [76]. Significant activation of genes of trehalose biosynthesis (otsAB, treS and treYZ), ligD, oprB and sigma factor rpoH, as well as genes involved in signal transduction and transport of inorganic ions was observed [76].
The accumulation of trehalose in cells in response to desiccation stress appears to be a universal mechanism of both rhizobacteria and other soil bacteria [76-78]. Trehalose accumulation in the cytoplasm rather than in the periplasmic space was found to be more important for cell survival during dehydration. Also, the chances of bacterial survival increase when trehalose is added to the medium [78]. Double mutants of otsA treY, which cannot accumulate trehalose, are more sensitive to the effects of desiccation and are also less competitive for nodule occupancy [79].
Trehalose is a non-reducing disaccharide (α- D -glucopyranosyl-1 and 1-α- D -glucopyranoside) containing two α-glucose molecules. The involvement of trehalose in cellular defence against stress is attributed to its stable structure: the low energy (1 kcal/mol) of the glycosidic bond connecting the two hexose rings [80]; and several unique physical properties: high hydrophilicity and chemical stability, non-hygroscopic glass formation, and lack of internal hydrogen bond formation [81]. There are three mutually exclusive mechanisms of biomolecule protection during drying with the participation of trehalose: 1) substitution of water molecules by trehalose molecules in the hydrate layer of biopolymers to stabilize their structure and prevent denaturation; 2) the formation by trehalose of amorphous, non-hygroscopic crystals causing a vitreous effect in the cell, which helps to keep all biopolymers intact for a long time; 3) the interaction of a very flexible glycosidic bond between two D -glucose residues of trehalose with the polar groups of various biomacromolecules, ensuring their chemical stability [80,81]. These mechanisms play a vital role in the survival of rhizobia under both free-living and symbiosis conditions and also help plants mitigating abiotic and biotic stresses [82].
3.2. Sensitivity of Rhizobacterial Species to Desiccation
Rhizobacteria tolerate full or partial desiccation stress in different ways [83]. In experiments where aliquots of PGPR bacterial suspensions were subjected to drying under different conditions to a certain humidity or complete drying and then re-watered, the titer of surviving cells varied greatly [84]. For example, after complete drying and then re-watering, (3.23 %±0.2 %) cells of Acinetobacter Calcoaceticus strain PADD68; (1.5 %±0.41 %) Arthrobacter phenanthrenivorans DSM 18606T and (31.58 %±6.9 %) Arthrobacter siccitolerans 4J27T [85] remained viable. Two strains of Bradyrhizobium japonicum, TXVA and TXEA, isolated from soybean root nodules in Texas showed (87.1% ± 0.61%) and (93.9% ± 0.63%) survival rates, respectively, under 27% relative humidity conditions, while their counterpart, the wild-type strain B. japonicum USDA110, was also very resistant to desiccation, with 80.00% ± 0.98% of cells surviving the experiments [86]. Some PGPRs are particularly sensitive to desiccation. For example, the titer of viable Pseudomonas putida KT2440T cells was not detected at all after undergoing desiccation stress [83,84]. Although, there is a suggestion that this may be due, not to the death of all cells, but their transition to a non-culturable state, which is one of the defense mechanisms against this type of stress [87]. For example, an experiment with PGPR Bradyrhizobium japonicum 5038 used fluorescent staining of stressed cells with Live/Dead cry-site and showed that several viable but unculturable cells were present in the culture medium after drying and rehydration [76]. The authors [88] analyzed 28 strains of rhizosphere bacteria able to withstand 18 days of air-drying stress (at 30°C and 50% relative humidity) and proposed to divide all rhizobacteria into 5 categories: highly tolerant, tolerant, medium tolerant, low tolerant and very low tolerant. In doing so, they showed that under desiccation conditions, only the highly tolerant strains could adhere to maize seeds, colonize its rhizosphere and stimulate plant growth. However, bacterial strains with low tolerance levels, unlike highly tolerant strains, were unable to do so [88]. A lot of work has to be done to isolate drought-resistant strains with beneficial properties. For example, Indian researchers analyzed 164 isolates of rhizobacteria, out of which only five of them belonging to Enterobacter cloacae, Bacillus cereus and Bacillus megaterium showed resistance to desiccation and yet were able to stimulate the growth performance of wheat during drought under desiccation stress [89].
3.3. Adaptive Mechanisms of AMFs under Drought
AMFs have been able to develop adaptive mechanisms and adjust to drought conditions, primarily by producing more spores [90,91]. However, water deficit affects the growth and development of AMF, which is manifested by reduced growth of extra-radical hyphae, and reduced ability to colonize plant roots, and to carry out the sporulation process [92]. All this leads to a decrease in the ability of AMF to form symbiotic associations with plant roots, which disturbs the balance of the soil microbiome [93]. Moreover, under dry conditions, the total abundance of all native AMF species in plant roots decreased, leading to a reduction in the amount of extraradical mycelium and a partial decrease in plant biomass [94]. In contrast, conventional flooding or excess water depressed AMF colonization in rice roots [95]. This indicates that extreme water regimes can significantly alter the structure of a native AMF assemblage, consequently impacting the entire symbiotic mycorrhizal relationship [94-95].
Adaptive responses of AMF to dehydration and drought stress occur at cellular, genetic, and enzymatic levels (Figure 1). Cellular adaptations in response to drought include changes in hyphal morphology and spore production, as well as adjustments in aquaporin expression to regulate water uptake [96,97]. At the genetic level (based on transcriptome analyses), drought exposure causes significant shifts in stress gene expression [98]. During dry periods, there are strategic changes in the activity of key enzymes: phosphatases (involved in enhanced mobilization of phosphate) and nitrogenases (contribute to improved nitrogen assimilation) [99]. Also, AMF cells show increased activity of antioxidant enzymes such as superoxide dismutase and peroxidase, which play a vital role in mitigating oxidative stress [100]. This generalized response of AMPs at cellular, genetic and enzymatic levels highlights their adaptability to adverse environmental conditions.
3.4. Alteration of Taxonomic Structure of AMFs under Drought
Drought and water stress significantly affect AMF, which live within and on the root surfaces of most terrestrial plants [101]. Many AMF species are believed to be widespread as they are quite resistant to any abiotic influences and can tolerate drought stress well [102]. However, the direct effects of abiotic stress, including drought, on AMF remain poorly understood [90]. Few studies have reported that mycorrhizal fungi respond to abiotic stress by reducing abundance and species biodiversity [91]. It has been shown that AMF content is lower in arid soils than in soils with normal moisture content, with representatives of the family Glomeraceae being the most common [103]. Within this family, different genera and species respond to water stresses in unique ways, affecting plant health and ecosystem function. For example, Augé et al. in 2015 identified significant changes in taxonomic diversity and community structure of the genus Glomus spp. in response to drought, emphasizing the sensitivity of symbiotic associations of these AMFs to environmental stress [104]. Similar studies of AMF communities in Mediterranean ecosystems by Veresoglou et al. showed changes in the taxonomic composition of Funneliformis spp. and other AMF genera in response to water stress [105]. Somewhat adaptive responses to drought stress have been shown for the genera Diversispora spp. [106], Rhizophagus spp. Archaeospora and Paraglomus [107]. In Lozano et al. the effect of drought stress on fungi belonging to the Basidiomycota and Ascomycota was noted. The relative abundance of Basidiomycota representatives decreased and Ascomycota representatives increased, presumably due to the formation of ascospores adapted to the drought environment [108].
Each study emphasizes the flexibility of AMF communities in adapting to water stress and discusses the potential effects of drought on plant-mycorrhizal associations under changing climatic conditions. Changes in AMF communities, especially those involved in nitrogen (N) and phosphorus (P) cycling, can negatively affect other beneficial microbial taxa, which may ultimately lead to the degradation of some ecosystems.
3.5. Distinct Adaptation Strategies in Rhizobacteria and AMFs to Drought
Rhizobia bacteria and arbuscular mycorrhizal fungi (AMF) exhibit active resistance to drought through evolutionary adaptive mechanisms involving cellular, genetic, and enzymatic alterations (Figure 1). These strategies in rhizobacteria and AMF share several similarities. For instance, when moisture levels in the environment decrease, both bacterial and fungal cells undergo transformation in cellular structure and metabolic activity primarily through changes in gene expression, while effectively regulating the osmotic state of cells. Rhizobacterial cells actively produce desiccation-resistant compounds, primarily osmolytes, while fungal cells exhibit strategic changes in the activity of key enzymes such as phosphatases, nitrogenases, superoxide dismutase, and peroxidases, along with active secretion of enzymes into the external environment. The formation of stable cellular structures is another common mechanism observed in both bacteria and fungi. Under severe drought conditions, bacteria can slow down cell metabolism and enter a dormant state, forming dormant cellular forms, whereas fungi produce drought-resistant cells with thicker cell walls, such as sclerotia.
A third important mechanism observed during drought, shared by rhizobacteria and AMF, involves increased interactions with plant roots to establish symbiotic relationships. Bacteria utilize biofilm-forming mechanisms, aiding their survival outside of symbiotic relationships, while fungi incorporate processes of considerable hyphae outgrowth to better colonize roots. Thus, both rhizobacteria and AMF, through active adaptation to drought, not only protect themselves but also assist plants in survival, especially when acting in concert.
3.6. Distinct Adaptive Strategies of Pathogenic and Beneficial Microbes to Drought
Pathogens, as well as mutualistic microorganisms, defend themselves against drought stress, with most of their adaptive mechanisms overlapping (Figure 2). One of the main defense mechanisms of pathogenic fungi and bacteria during drought and other unfavorable conditions, as well as AMF and some spore-forming species of rhizobacteria, is the formation of large amounts of stable structures, in the form of which they can persist in the soil for a very long time. For example, the main crop pathogens of fungi (belonging to the genera Fusarium or Verticillium) and oomycetes form stable structures: microsclerotia, sclerotia, chlamydospores, oospores, etc., to survive in arid conditions. [14].
Like mutualistic microorganisms, pathogens produce osmoprotectants such as proline, glycine-betaene, and other amino acids in their cells when water is scarce during drought, for example, as the oomycete Phytophthora infestans does [111]. In a study by El-Abyad et al. it was shown that the sugar beet root rot pathogens Fusarium solani and Sclerotium rolfsii under stress conditions produce alanine, proline and histidine, which play an important role in the osmoregulation of their cells [112]. In response to the changing osmotic conditions that accompany drought, pathogens not only produce osmoprotectants but also trigger other osmosensing mechanisms that help them survive. A study by Rossier and Vorhölter [113] showed how this occurs in the pathogenic bacterium Xanthomonas campestris, which causes a multitude of diseases in monocotyledonous and dicotyledonous crops worldwide [113]. The same signal transduction cascades under osmotic stress conditions are also triggered in mutualistic soil microbes such as Bacillus subtilis [114].
3.7. Adaptation of Phytopathogenic Fungi and Bacteria to Drought
Seventy to 80% of plant diseases resulting in enormous losses in agriculture are caused by pathogenic fungi belonging to the genera Alternaria, Aspergillus, Botrytis, Cladosporium, Verticillium, Pythium, Fusarium and Rhizoctonia [115]. Several hundred species of bacteria belonging to the Proteobacteria, Mollicutes and Actinomycetes are also plant pathogens. Entering the plant through natural openings such as stomata and lenticels, or wounds, the bacteria cause necrosis, maceration of tissues, wilting and hyperplasia [116].
During drought, the spread of pathogens increases and the likelihood of plant infection and disease severity increases. Modeling coupled with experimental data has shown that the prevalence of major soil fungal pathogens belonging to the genera Alternaria, Fusarium, Venturia, and Phoma is likely to increase under projected global warming [117]. However, phytopathogenic bacteria and fungi, as well as beneficial ones, tolerate drought in different ways. For some, drought is an additional factor that promotes their activity, while for others, reduced moisture becomes a hardship and they reduce their pathogenic claims on plants [118]. For example, chickpea production worldwide is significantly constrained by a disease called dry root rot (DRR) caused by the phytopathogenic fungus Macrophomina phaseolina. Studies conducted by multiple groups of scientists have shown that drought and high temperatures exacerbate the impact of pathogens on various crop plants, including chickpea [119,120]. These pathogens encompass pea root rot (Aphanomyces euteiches), white rot of onion (Sclerotium cepivorum), wheat spot (Gaeumannomyces graminis var. tritici), wheat crown rot (Fusarium spp.), black foot of Brassica (Leptosphaeria maculans), and black foot of grape (Ilyonectria/Dactylonectria spp.) [118]. Botryosphaeria doithidea becomes more active during dry periods, colonizing the above-ground parts of host plants through wounds and causing sores (lesions?) on a wide variety of tree and shrub species [14]. And climate change is projected to contribute to more frequent and intense outbreaks of diseases caused by Botryosphaeria dothidea as well as Neofusicoccum parvum due to their expanding range [121]. Also, according to a global study by Guerra, C. and colleagues, increased drought will favor the spread of soil fungi Penicillium spp. that cause fruit and vegetable spoilage.
On the other hand, soil pathogens belonging to the taxa Oomycota, Phytophthora spp. and Pythium spp. are more sensitive to drought and their distribution will decrease during dry periods [122]. For example, Phytophthora, which destroys the fine root system of plants, has been shown to increase its pathogenic activity in wet conditions and decrease in dry soils. Drought has also been found to alleviate the severity of sclerotinia rot of kiwifruit (S. sclerotiorum) and red needle leaf blight of radiata pine (Phytophthora pluvialis) [118]. In dry periods, the activity of the bacterial pathogen Xylella fastidiosa, which affects grapevines, also decreases. During drought, necrotrophic pathogens will be more active, depleting plant resources through regeneration and compartmentalization processes. In contrast, diseases caused by biotrophic pathogens will be less severe due to the close correlation between pathogen effectiveness and plant nutritional status, but only until the plant is on the verge of destruction [123].
4. Soil Microbiomes Mediate Plant Defence Mechanism under Drought and Desiccation Stresses
4.1. Rhizobacteria-Mediated Plant Defense Mechanisms under Drought Conditions
In soil, the relationship between rhizobia and plants is crucial for the maintenance of plant productivity. Therefore, the main objective of effective agronomy - to increase plant resistance to various types of stresses, including drought - has been successfully addressed by the application of rhizobacteria and AMF [124]. Recently, researchers have focused their attention on the unique abilities of rhizobacteria [125], mycorrhizal fungi [90,126,127] or their compositions [68,128] to help plants overcome various abiotic and biotic stresses.
PGPR induces immunity to abiotic stresses in plants, which is called induced systemic tolerance (IST) [129]. In addition, through physicochemical modifications, rhizobacteria induce increased plant resistance to drought, which is termed rhizobacterial-induced drought tolerance and resistance (RIDER) [125,130]. Numerous studies have demonstrated how PGPRs help crops combat drought and other types of abiotic stress by positively affecting growth, yield, and nutrient uptake [125,131,132]. Rubin et al. (2017), conducted a meta-analysis of 52 publications on the effects of rhizobacteria (mainly Pseudomonas, Azospirillum, Azotobacter and Bacillus) on plant growth (mainly maize, wheat, sunflower and lettuce) [67]. The authors systematized the data and summarized that under standard moisture conditions, rhizobacteria contributed 35%, 28% and 19% to the increase in root weight, shoot weight and seed yield, respectively, whereas under drought conditions rhizobacteria contributed 43%, 45% and 40%, respectively [67]. Recently, research in this area has continued with even greater intensity. To effectively help crops to cope with drought, researchers propose the use of increasingly effective PGPR strains [133] or combinations of bacterial cultures [131,134] as well as mixed combinations of rhizobacteria with mycorrhizal fungi [134,135]. Attention is also paid to selecting the stage of plant development at which the application of rhizobacteria will be most effective [136]. The most interesting works are those in which researchers propose to apply a combination of rhizobacteria and various additives: fertilizers, metabolites, hormones, nanoparticles, etc. to help plants during drought. For example, Khan showed that the combined application of rhizobial cultures Planomicrobium chinense and Bacillus cereus and the plant growth hormone, salicylic acid, effectively mitigates the adverse effects of water stress on wheat by increasing protein, sugar and chlorophyll content in leaves [137]. The same positive synergism increasing yield and physiological parameters of wheat under drought conditions was observed with rhizobacteria and cytokinins applied at different stages of plant development (tillering, flowering, grain filling) [138]. Muhammad et al. showed that the combined application of rhizobacteria Azospirillum brasilense and ZnO nanoparticles (NP) has a synergistic effect on wheat, providing growth stimulation, reducing drought stress and increasing yield [139].
There is no single mechanism by which rhizobacteria protect plants from desiccation stress. Typically, PGPBs use a wide range of adaptive measures that may vary depending on the species and strain of rhizobacteria [140]. According to the type of effects on plants, they can be divided into direct and indirect, and according to the organizational level at which changes occur, into physical, molecular and biochemical [141]. Thus, direct mechanisms (and at the same time biochemical) include how PGPR directly affects plants during drought: enhancing nitrogen fixation; inducing the production of abscisic acid, which leads to the partial closure of the stomata and reduces water evaporation through transpiration; influencing the levels of phytohormones (auxins, gibberellins and cytokinins); influencing the production of antioxidants to reduce reactive oxygen species (ROS) [142]; effects on increasing the production of osmolytes, carbohydrates, amino acids, polyamines and volatile organic compounds [143]; production of ACC deaminase, which reduces the concentration of ethylene (a plant hormone responsible for slowing down root and shoot growth under drought conditions) in plants by cleaving its precursor, which supports normal plant growth [144]; and others. The study of these mechanisms has been systematized in detail in numerous reviews published in recent years [125,145].
Indirect mechanisms may include PGPR-mediated actions that also contribute to plant survival during drought. For example, increasing the availability of soil nutrients and minerals by solubilizing phosphate or producing siderophores that help iron uptake [146]. Also, indirect methods include preventing the harmful effects of phytopathogens by producing PGPR antibiotics, antagonistic substances (phenazine, diacetylphloroglucinol, hydrogen cyanide, 2-3-butanediol, acetoin), siderophores, lytic enzymes (chitinase and glucanase that destroy the cell wall of fungal pathogens) [147,148]. Molecular mechanisms are related to the effect of rhizobacteria on plant gene expression [149]. For example, it was shown that PGPR inoculation caused up-regulation of drought stress-related genes (early dehydration response genes, heat shock proteins, various enzymes, etc.) in wheat, pepper, mungbean and Arabidopsis [150]. Physical mechanisms include the ability of PGPR to produce exopolysaccharides in the soil, which fulfill several functions [145]. On the one hand, oligo- and polysaccharides are absorbed by the surface of soil particles, and through cationic, hydrogen and Van der Waals interactions with soil chemical compounds, increase its stability and moisture retention capacity [151]. The water-holding capacity of such exometabolism can reach seventy grams of water per gram of polysaccharide [152]. On the other hand, EPSs promote the formation of hydrophilic biofilms (in the form of capsules) on the surface of plant roots, further protecting them from drying [153,154].
4.2. Arbuscular Mycorrhizal Fungal-Mediated Plant Defense Mechanisms under Drought
AMFs significantly increase host plant resistance to abiotic and biotic stresses [155,156], especially, as previously mentioned, they are symbionts with most terrestrial plants [157]. They manage to do so thanks to the developed network of their mycelium, the production of glomalin [158], as well as a number of other mechanisms acting at three levels: physiological, morphological and molecular [159]. Studies investigating the phenomenon and mechanisms of AMF-mediated enhancement of plant resistance during drought are plentiful and interest continues to grow [126,160]. Yu Wang [159] analyzed the publication activity over several years on this issue. The authors write that in 2011 there were 27 articles related to this topic, in 2021 there are already 124 articles, and in 2022 there are 131 articles, with the focus increasingly shifting to studying the molecular mechanisms that explain how arbuscular mycorrhizae enhance plant tolerance to drought [159].
AMFs alter host plant physiology under drought conditions: they adjust water potential and gas exchange, increase water and nutrient uptake, and improve stomatal regulation [92,161,162]. AM extraradical hyphae can create shared mycorrhizal networks between neighboring plants, helping to transmit signals and exchange nutrients [160]. At the biochemical level, AMFs modulate levels of hormones (such as strigolactones, jasmonic acid, abscisic acid, etc.), activate antioxidant defense systems, and help plant cells maintain osmotic pressure [126, 157-163]. For example, Zou [164] analyzed changes in root metabolites of walnut (Juglans regia) after inoculation with the arbuscular mycorrhizal fungus Diversispora spurca under drought conditions using ultra-high performance liquid chromatography. It was found that inoculation of plants with AMF under drought stress increased the activity of 49 metabolites (including 2,3,5-trihydroxy-5-7-dimethoxyflavanone) and decreased the activity of 116 metabolites [164]. In a study by Sheteiwy [165] it was shown that inoculation of soybean (Giza 111) with AMF together with rhizobacteria under stress conditions increased the content of primary metabolites and mitigated the drought-induced decrease in soluble sugars, lipids, protein, and oil, and resulted in decreased levels of stress hormones (abscisic acid) and increased gibberellin, trans-zeatin-riboside and indolylacetic acid in seeds of inoculated plants [165]. In another work on soybean, it was shown that inoculation of plants with mycorrhizal-rhizobial consortium during drought increased the relative water content of leaves and the amount of osmoprotectant proline in beans, as well as pod number, pod fresh weight and seed dry weight [68].
All these changes are based on molecular mechanisms associated with changes in gene expression and regulation of transcription factors [159]. For example, in [165] it was shown that AMF inoculation of plants induced the expression of genes involved in lipid and protein biosynthesis and blocked the expression of genes involved in their degradation. Studies of mycorrhizal fungi and soybean interactions have demonstrated that under soil drought conditions, AMP can trigger the expression of catalase and peroxidase genes in plants [166]. AMF have also been shown to significantly up-regulate the expression of key genes of ABA signaling pathway TFT2 and TFT3 in tomatoes under drought conditions [167]. Recently, mycorrhizal fungi were found to modulate plant drought tolerance through the regulation of transcription factors (GRAS, MYB, and AP2/ERF) [159]. Thus, the study of the molecular mechanisms of AMF assistance to plants during drought is ongoing. The findings contribute to the promising development of efficient agronomy.
5. Horizontal Gene Transfer in the Adaptation of Plant Pathogenic and Non-Pathogenic Beneficial Microbes under Drought
5.1. Genetic Exchange Employed by Pathogens through Horizontal Gene Transfer (HGT)
Drought and water deficit exert selective pressure on soil microbial communities, promoting genetic exchange as a survival strategy. Both non-pathogens and pathogens undergo horizontal gene transfer (HGT) to adapt to changing environmental conditions [168]. For example, in response to drought stress, nonpathogenic bacteria such as Pseudomonas putida can acquire stress tolerance genes [169], while pathogens such as Xylella fastidiosa can exchange genes related to virulence and host specificity [170].
Some genomic adaptation mechanisms to drought in pathogenic and nonpathogenic microorganisms, although having a common basis, differ in the direction of action (Figure 3). Such mechanisms include changes in gene expression and production of antioxidant enzymes. Both pathogenic and non-pathogenic microorganisms in response to drought regulate their gene expression toward the production of essential metabolites involved in defence mechanisms. However, non-pathogenic soil microbes, especially plant-growth promoting species, express genes associated with stimulating plant growth under stress conditions, for example Azospirillum brasilense produces large amounts of phytohormones [171]. Whereas pathogens during drought stimulate the expression of pathogenicity, virulence and host colonization factors (including adhesins, polysaccharides, LPS and degradative enzymes) as in Fusarium oxysporum [172] or Botrytis cinerea [173]. One of the key pathogenicity factors is the type III secretion system, which introduces effector proteins into the host cell cytosol to manipulate plant cellular processes, such as basal defense, to the benefit of the pathogen [174]. In addition, both microbial groups can produce antioxidant enzymes, but their main focus is different: non-pathogens use them to increase their own stress tolerance, such as Trichoderma harzianum [175], while pathogens use them to manipulate host invasion, such as the pathogen Botrytis cinerea [173]. Environmental stressors, including drought, can promote biofilm formation in both non-pathogenic and pathogenic microbes. Biofilms serve as foci of genetic exchange, facilitating interactions between different microbial species. For example, soil microbes responding to drought can engage in biofilm-mediated interactions, affecting the transfer of genetic material and contributing to the evolution of pathogenic and non-pathogenic traits [176]. However, rhizobacteria defend themselves against drought stress by forming biofilms that encompass plant roots and reorganize soil structure. Pathogens do not use this strategy as they need to invade rather than protect the plant. But some distinguish the adaptive strategy of pathogens from mutualistic microorganisms. These include the ability of pathogens to synthesize phytohormones that help suppress plant growth, as well as pathogenicity factors (absent in mutualistic microorganisms) that help them infect the host [115,177].
Drought and water deficit exert selective pressure on soil microbial communities, promoting genetic exchange as a survival strategy. Both pathogenic and non-pathogenic microorganisms undergo adaptive changes in response to drought stress [178,179]. Some of the common adaptation mechanisms include: production of osmo-protectants like trehalose, proline, glycine-betaine to maintain cellular osmotic balance [81]. Altered gene expression, production of antioxidant enzymes, and biofilm formation were facilitated during genetic exchange and interactions between microbial species [180]. However, there are also mechanisms that distinguish the adaptive strategies of pathogens vs. mutualistic microorganisms. Pathogens tend to focus more on virulence factors and suppressing plant growth, while mutualistic microbes enhance plant growth and stress tolerance [181,182]. Horizontal gene transfer (HGT) is an important adaptive mechanism, allowing both pathogenic and non-pathogenic microbes to acquire beneficial traits like stress tolerance and virulence factors [183]. Environmental stressors like drought can promote increased genetic exchange and the spread of pathogenic traits [184,185]. In summary, drought drives complex adaptive responses in soil microbial communities, involving both common and distinct mechanisms between pathogenic and beneficial microbes, with HGT playing a key role in their evolution and survival.
5.2. Virulence and Pathogenicity Gene Exchange under Drought
Horizontal gene transfer (HGT) (a mechanism by which DNA is transferred between unrelated organisms in a non-Mendelian manner) is fairly considered to be an important adaptation to unfavorable environmental conditions such as drought and desiccation [186,187]. Interspecific gene transfer was initially shown for bacteria, in which it occurs in more than 75% of all protein families [187,188]. Interspecies transfer of virulence genes from pathogenic to nonpathogenic cells was first studied, contributing to the virulence of(towards?) the new host and, as a consequence, the rapid spread of bacterial antibiotic resistance in hospitals [189]. In 2000, Walton suggested the possibility of HGT of toxin genes in fungal pathogens [190]. Friesen et al. (2006) [191] provided evidence that the gene encoding the critical virulence factor ToxA was transferred from one species of fungal phytopathogen Stagonospora nodorum, the causal agent of brown (or yellow) spot disease of wheat, to another fungus Pyrenophora tritici-repentis [191]. More recently, the wheat virulence protein encoding ToxA was shown to be horizontally transferred between three fungal pathogens of wheat (Parastagonospora nodorum, Pyrenophora tritici-repentis, and Bipolaris sorokiniana) as part of a conserved ∼14 kb element that contains coding and noncoding regions [192]. Knowledge in this area continues to expand. Recently, Japanese scientists proved that the genome of the wheat-infecting strain P. oryzae Br48 (isolated in Brazil in 1990 and belonging to an ancient MoT population that originated in South America in the 1980s) contains many genes derived from HGT [192]. A pathogenicity chromosome from a Fusarium pathogen to a non-pathogenic Fusarium species, making it pathogenic on tomato (3), which can have significant implications for agricultural and environmental health. Full-genome sequencing projects have made significant contributions to the study of the importance of HGT in the genome evolution of prokaryotes [193] and unicellular eukaryotes [186,194].
How HGT-derived genes are integrated into the epigenetic regulation system of the genome and how environmental conditions affect HGT, especially concerning virulence genes, are now on the agenda. During environmental stress, an increased potential for genetic exchange is created among plant-soil microorganisms (Figure 3). Stress-induced genetic exchange enables nonpathogenic microorganisms to acquire beneficial traits, increasing their ability to survive and contribute to plant health under unfavorable conditions [195]. On the other hand, there is a threat of greater spread of pathogenic traits and the emergence of new pathogenic strains [196]. However, the reverse can also occur with phytopathogenic microorganisms. It has been shown that in response to increased ambient temperature, phytopathogens can undergo genetic changes to increase heat tolerance, which affects (reduces) their ability to infect and survive in the soil [197]. Thus, in a study by Cubeta et al. [198], it was shown that to cope with water shortage and desiccation stress the pathogen Rhizoctonia spp. alters cellular and enzymatic functions, potentially changing cell virulence and pathogenicity. The effects extend to cellular, genetic, and enzymatic levels, affecting the overall adaptability and virulence of Rhizoctonia spp. [198]. That is, under drought conditions, Rhizoctonia spp. become non-pathogenic.
The soil and plant rhizosphere environments can promote HGT due to factors such as the presence of diverse microbial communities, the availability of nutrients, and environmental stressors that can select for the retention of transferred genes [199]. HGT is a common mechanism that allows the rapid spread of virulence and pathogenicity genes across microbial communities in soil and plant environments. Virulence genes frequently transferred horizontally include those involved in bacterial adherence, secretion systems, toxin production, and iron acquisition [200]. Examples include the virulence factors of Bacillus anthracis and Fusarium oxysporum causing severe diseases in plant root systems.
Therefore, drought, along with environmental fluctuations and water scarcity, profoundly impacts the dynamics of soil microbiomes, inducing alterations in the virulence and pathogenicity of microorganisms through processes like horizontal gene transfer and genetic exchange. The intricate interplay among environmental stressors, microbial community dynamics, and genetic exchange underscores the necessity for a thorough understanding of these mechanisms to effectively manage soil health and counteract the emergence of new pathogenic strains [201,202]. Further investigation is imperative to elucidate the precise mechanisms through which drought stress influences genetic exchange and its role in shaping the virulence and pathogenicity of soil microbiomes. By examining the dynamics of soil microbial communities under varied stress conditions, we can gain insight into how genetic exchange contributes to the adaptability and resilience of both non-pathogenic and pathogenic microbes.
5.3. Implication of Genomic Changes in the Plant-Soil System under Drought
Research indicates that soil microbes play a significant role in shaping plant responses to both biotic and abiotic stresses, such as drought. This involvement is multifaceted, encompassing changes in the architecture of the host plant’s root system through hormonal regulation, osmoregulation, signaling via reactive oxygen species (ROS), induction of systemic tolerance (IST), production of large-chain extracellular polysaccharides (EPS), and transcriptional regulation of the host’s stress response genes [142,203]. An integrated approach to managing the rhizosphere is proposed as a strategy for mitigating drought stress in plants, with a specific emphasis on rhizosphere management. This holistic method may involve rhizosphere manipulation through the introduction of drought-tolerant bacteria, nanoparticles, liquid nano clay (LNC), nutrients, and organic matter, in conjunction with plant modification using advanced genome editing tools like CRISPR/Cas9, aimed at promptly addressing evolving agricultural challenges. Several mechanisms facilitate genomic changes including horizontal gene transfer (HGT) in soil and plant-associated microbial communities, including conjugation, which is a common mode of HGT, is where bacteria can transfer genetic material via cell-to-cell contact and specialized transfer machinery, such as sex pili [204]. Secondly, there is transformation, in which bacteria can also take up free DNA from the environment through natural competence and incorporate it into their genomes, a process known as transformation. Thirdly, transduction is where bacteriophages (viruses that infect bacteria) can package and transfer bacterial DNA between host cells, a process called transduction (3) and lastly, conjugative plasmids, which carry genes for their transfer, can facilitate the spread of virulence and antibiotic-resistance genes between bacterial communities [205]. Bacteria, horizontal gene transfer (HGT) and horizontal chromosome transfer (HCT) have also been observed in plant pathogenic fungi, allowing them to broaden their host range [202]. Examples include the transfer of virulence genes in Alternaria alternata and Fusarium oxysporum. The origin and mechanisms of how DNA exchange occurs after microbial anastomosis or fusion events, which can facilitate unusual HGT or HCT, are still largely unknown and remain a major challenge to understand.
5.4. Shift of Pathogens and Non-Pathogens through HGT under Drought
Drought stress can have significant impacts on soil-borne plant pathogens and beneficial microbes, including effects on their genetic exchange. Drought stress can alter the composition and dynamics of the soil microbial community, making bacteria more dominant over fungi [206]. This shift in the microbial balance can lead to a decline in the quality of organic substances in the rhizosphere, which can negatively impact plant health. Drought can also directly reduce the activity and diversity of beneficial microbes like plant growth-promoting rhizobacteria (PGPR) and mycorrhizal fungi. This can diminish their ability to protect plants against soil-borne pathogens and provide nutrients. The key evidence supporting this claim is that drought conditions can trigger abrupt and persistent shifts in the structural and functional attributes of the soil microbial community, including reduced microbial functional capacity and modified taxonomic composition [207]. This drought-induced stress on the soil microbiome may increase the likelihood of HGT and HCT events, as microbes allocate more resources towards survival mechanisms than normal metabolic functions.
On the other hand, some soil-borne pathogens like Phytophthora can thrive better under drier conditions, leading to increased disease severity [208]. Drought-stressed plants also become more susceptible to infection by these pathogens (4). Drought can further impact genetic exchange between microbes in the rhizosphere. Reduced soil moisture and nutrient availability can limit horizontal gene transfer between bacteria, fungi, and oomycetes. This can constrain the spread of beneficial traits like antibiotic production or drought tolerance. In summary, drought disrupts the delicate balance between soil-borne pathogens and beneficial microbes. It can selectively favour certain pathogens, diminish the protective effects of PGPR and mycorrhizae, and limit genetic exchange that could otherwise improve microbial adaptations to stress [206,208]. These results explain that under drought stress, soil microorganisms have to divert more energy towards adjusting to the water deficit, such as producing osmolytes and increasing enzyme synthesis. This shift in resource allocation under drought conditions may increase the likelihood of HGT and HCT events between soil microbes. This is because microbes under stress are more likely to engage in mechanisms like conjugation, transduction, and transformation that facilitate the horizontal transfer of genetic material, rather than focusing on their regular cellular processes. There are several ways that soil microbes adapt to drought depending on the nature of microbes either pathogenetic or mutualistic fungi or bacteria. Adaptions such as through tolerance strategies like osmolyte production and dormancy, as well as changes in community composition with drought-tolerant taxa becoming more dominant. However, the role of HGT in shifting processes between pathogens and non-pathogens is not clearly understood and new hypotheses and predictions are evolving [209].
6. Summary
The review has provided a detailed examination of the contrasting adaptive strategies employed by beneficial and pathogenic soil bacterial and fungal communities in response to climate-induced drought and desiccation stresses. Key results are: 1. Both pathogenic and non-pathogenic microorganisms undergo a range of cellular, genetic, and enzymatic changes to survive under drought conditions. Common adaptation mechanisms include the production of osmoprotectants, altered gene expression, and biofilm formation. 2. There are distinct differences in the focus of adaptive strategies between pathogens and mutualistic (beneficial) microbes. Pathogens tend to prioritize virulence factors and suppress plant growth, while beneficial microbes enhance plant growth and stress tolerance. 3. Horizontal gene transfer (HGT) is a crucial adaptive mechanism, allowing both pathogenic and non-pathogenic microbes to acquire beneficial traits like stress tolerance and virulence factors. Environmental stressors like drought can promote increased genetic exchange and the spread of pathogenic traits. 4. The complex interplay between drought-adapted microbes, both beneficial and pathogenic, and their interactions with plants, highlights the need for a deeper understanding of soil microbiome dynamics under climate change. In conclusion, this review highlights the various survival strategies of soil microorganisms during climate induced drought and desiccation, shedding light on the adaptive mechanisms and evolutionary trajectories of both beneficial and pathogenic soil microbiome members. These insights are crucial for enhancing sustainable agriculture, aiding experts in agronomy, pathology, microbiology, ecology, and policy. With this knowledge, researchers can lead pioneering studies and develop effective strategies to combat challenges posed by global warming. Utilizing innovative methods to harness plant-soil microbial resources, these efforts aim to significantly benefit crop producers globally.
Author Contributions
Both authors, M.N.I. and N.L., have contributed equally to this work in the following aspects: conceptualization, methodology, software, validation, formal analysis, investigation, resources, data curation, original draft preparation, review and editing, visualization, and funding acquisition. Authors have read and approved the final version of the manuscript for publication.
Funding
This work was supported by the Ministry of Science and Higher Education of the Russian Federation № 122040800164-6 and this manuscript is also supported by grants for invited featured manuscripts from the MDPI Agronomy Journal editorials.
Acknowledgements
The authors gratefully acknowledge and thank Dr. Brent McCallum and Dr. James Menzies, Senior Phytopathology Research Scientists at AAFC MRDC, as well as Dr. Randy Kutcher, Professor at the Department of Plant Sciences, University of Saskatchewan, for their invaluable contributions in reviewing the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bach, E.M.; Ramirez, K.S.; Fraser, T.D.; Wall, D.H. Soil biodiversity integrates solutions for a sustainable future. Sustainability 2020, 12(7), 2662. [CrossRef]
- Philippot, L.; Chenu, C.; Kappler, A.; Rillig, M.C.; Fierer, N. The interplay between microbial communities and soil properties. Nat Rev Microbiol 2023, 22, 226-239. [CrossRef]
- Sahu, P.K.; Singh, D.P.; Prabha, R.; Meena, K.K.; Abhilash, P.C. Connecting microbial capabilities with the soil and plant health: Options for agricultural sustainability. Ecological Indicators 2019, 105, 601-612. [CrossRef]
- Sarsan, S.; Pandiyan, A.; Rodhe, A.V.; Jagavati, S. Synergistic interactions among microbial communities. Microbes in Microbial Communities: Ecological and Applied Perspectives 2022, 1-37. [CrossRef]
- Kumar, I.; Mondal, M.; Gurusamy, R.; Balakrishnan, S.; Natarajan S. Plant-microbiome interaction and the effects of biotic and abiotic components in agroecosystem. Microb. Interv. Agric. Environ. 2019, 2, 517-546. [CrossRef]
- Ault, T. R. On the essentials of drought in a changing climate. Science 2020, 368(6488), 256-260.
- Vásquez-Dean, J.; Maza, F.; Morel, I.; Pulgar R.; González M. Microbial communities from arid environments on a global scale. A systematic review. Biol Res 2020, 53(1), 29. [CrossRef]
- Naorem, A.; Jayaraman, S.; Dang, Y.P.; Dalal, R.C.; Sinha, N.K.; Rao, C.S.; Patra, A.K. Soil Constraints in an Arid Environment—Challenges, Prospects, and Implications. Agronomy 2023, 13, 220. [CrossRef]
- Alpert, P. The limits and frontiers of desiccation-tolerant life. Integrative and comparative biology 2005, 45(5), 685-695. [CrossRef]
- Vicente-Serrano, S.M.; Quiring, S.M.; Peña-Gallardo, M.; Yuan, S.; Domínguez-Castro, F. A review of environmental droughts: Increased risk under global warming? Earth-Science Reviews 2020, 201, 102953. [CrossRef]
- Esbelin, J.; Santos, T.; Hébraud, M. Desiccation: An environmental and food industry stress that bacteria commonly face. Food Microbiology 2018, 69, 82–88. [CrossRef]
- Brettner, L.; Ho W.C.; Schmidlin K.; Apodaca, S.; Eder, R.; Geiler-Samerotte, K. Challenges and potential solutions for studying the genetic and phenotypic architecture of adaptation in microbes. Curr Opin Genet Dev. 2022, 75, 101951. [CrossRef]
- Meisner, A.; Jacquiod, S.; Snoek, B.L.; Ten Hooven, F.C.; van der Putten, W.H. Drought legacy effects on the composition of soil fungal and prokaryote communities. Front Microbiol. 2018, 9, 294. [CrossRef]
- Thompson, S.; Levin, S.; Rodriguez-Iturbe, I. Linking plant disease risk and precipitation drivers: A dynamical systems framework. Am Nat. 2013, 181(1), E1-16. [CrossRef]
- Garbeva, P.; Hol, W.H.; Termorshuizen, A. J.; Kowalchuk, G.A.; de Boer, W. Fungistasis and general soil biostasis – A new synthesis. Soil Biology and Biochemistry 2011, 43, 469-477. [CrossRef]
- Durán, P.; Tortella, G.; Sadowsky, M.J.; Viscardi, S.; Barra, P.J.; Mora, M.L. Engineering Multigenerational Host-Modulated Microbiota against Soilborne Pathogens in Response to Global Climate Change. Biology 2021, 10(9), 865. [CrossRef]
- Leach, J.E.; Triplett, L.R.; Argueso, C.T.; Trivedi, P. Communication in the phytobiome. Cell 2017, 169, 587–596. [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [CrossRef]
- Manzanera, M. Dealing with water stress and microbial preservation. Environ Microbiol. 2021, 23(7), 3351-3359. [CrossRef]
- Loiko, N.; Tereshkina, K.; Kovalenko, V.; Moiseenko, A.; Tereshkin, E.; Sokolova, O.S.; Krupyanskii, Y. DNA-binding protein Dps protects Escherichia coli cells against multiple stresses during desiccation. Biology, 2023, 12(6), 853. [CrossRef]
- Bogati, K.; Walczak, M. The Impact of Drought Stress on Soil Microbial Community, Enzyme Activities and Plants. Agronomy 2022, 12, 189. [CrossRef]
- Matthews, K.E.; Facelli, J.M.; Cavagnaro, T.R. Response of soil microbial community structure, carbon and nitrogen cycling to drying and rewetting. Applied Soil Ecology 2023, 192, 105099. [CrossRef]
- Schimel, J.P. Life in dry soils: Effects of drought on soil microbial communities and processes. Annual Review of Ecology, Evolution, and Systematics 2018, 49(1), 409-4321. [CrossRef]
- Evans, S.E.; Allison, D.S.; Hawkes, V.C. Microbes, memory and moisture: Predicting microbial moisture responses and their impact on carbon cycling. Functional Ecology 2022, 36(6), 1430-1441. [CrossRef]
- Gillespie, L.M.; Prada-Salcedo, L.D.; Shihan, A.; Fromin, N.; Goldmann, K.; Milcu, A.; Buscot, F.; Buatois, B.; Hättenschwiler, S. Taxonomical and functional responses of microbial communities from forest soils of differing tree species diversity to drying-rewetting cycles. Pedobiologia 2023, 97–98, 150875. [CrossRef]
- Maestre, F.T., Delgado-Baquerizo, M., Jeffries, T.C., Eldridge, D.J., Ochoa, V., Gozalo, B., ... & Singh, B.K. Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proceedings of the National Academy of Sciences 2015, 112(51), 15684-15689. [CrossRef]
- Evans, S.E.; Wallenstein, M.D. Soil microbial community response to drying and rewetting stress: Does historical precipitation regime matter? Biogeochemistry 2012, 109, 101–116. [CrossRef]
- Lebre, P., De Maayer, P.; Cowan, D. Xerotolerant bacteria: Surviving through a dry spell. Nat Rev Microbiol. 2017, 15, 285–296. [CrossRef]
- Scales, N.C.; Huynh, K.T.; Weihe, C.; Martiny, J.B.H. Desiccation induces varied responses within a soil bacterial genus. Environ Microbiol. 2023, 25(12), 3075-3086. [CrossRef]
- Breitkreuz, C.; Herzig, L.; Buscot, F.; Reitz, T.; Tarkka, M. Interactions between soil properties, agricultural management and cultivar type drive structural and functional adaptations of the wheat rhizosphere microbiome to drought. Environ. Microbiol. 2021, 23, 5866–5882. [CrossRef]
- Chodak, M.; Gołębiewsk, M.; Morawska-Płoskonka, J.; Kuduk, K.; Niklińska, M. Soil chemical properties affect the reaction of forest soil bacteria to drought and rewetting stress. Ann Microbiol. 2015, 65(3),1627-1637. [CrossRef]
- Maisnam, P.; Jeffries, T.C.; Szejgis, J.; Bristol, D.; Singh, B.K.; Eldridge, D.J.; Horn, S.; Chieppa, J.; Nielsen, U.N. Severe prolonged drought favours stress-tolerant microbes in australian drylands. Microb Ecol. 2023, 86(4), 3097-3110. [CrossRef]
- Metze, D.; Schnecker, J.; Canarini, A.; Fuchslueger, L.; Koch, B.J.; Stone, B.W.; Hungate, B.A.; Hausmann, B.; Schmidt, H.; Schaumberger, A.; Bahn, M.; Kaiser, C.; Richter, A. Microbial growth under drought is confined to distinct taxa and modified by potential future climate conditions. Nat Commun. 2023,14(1), 5895. [CrossRef]
- Jaeger, A.C., Hartmann, M.; Six, J.; Solly, E.F. Contrasting sensitivity of soil bacterial and fungal community composition to one year of water limitation in Scots pine mesocosms. FEMS Microbiology Ecology 2023, 99(6), fiad051. [CrossRef]
- Rosinger, C.; Rousk, J.; Bonkowski, M.; Rethemeyer, J.; Jaeschke, A. Rewetting the hyper-arid Atacama Desert soil reactivates a carbon-starved microbial decomposer community and also triggers archaeal metabolism. Sci Total Environ. 2023, 892, 164785. [CrossRef]
- Chilakala, A. R.; Pandey, P.; Durgadevi, A.; Kandpal, M.; Patil, B. S.; Rangappa, K.; Reddy, P.C.O.; Ramegowda, V.; Senthil-Kumar, M. Drought attenuates plant responses to multiple rhizospheric pathogens: A study on a dry root rot-associated Disease complex in chickpea fields. Field Crops Research 2023, 298, 108965. [CrossRef]
- Sinha, R.; Irulappan, V.; Mohan-Raju, B.; Suganthi, A.; Senthil-Kumar, M. Impact of drought stress on simultaneously occurring pathogen infection in field-grown chickpea. Scientific Reports 2019, 9, 5577. [CrossRef]
- Kaisermann, A.; Maron, P.A.; Beaumelle, L.; Lata, J.C. Fungal communities are more sensitive indicators to non-extreme soil moisture variations than bacterial communities. Applied Soil Ecology 2015, 86, 158-164. [CrossRef]
- Marasco, R., Rolli, E., Ettoumi, B., Vigani, G., Mapelli, F., Borin, S., Abou-Hadid, A.F.; El-Behairy, U.A.; Sorlini, C.; Cherif, A.; Zocchi, G.; Daffonchio, D. A A drought resistance-promoting microbiome is selected by root system under desert farming. PLoS ONE 2012, 7(10), e48479. [CrossRef]
- Siebielec, S.; Siebielec, G.; Klimkowicz-Pawlas, A.; Gałązka, A.; Grządziel, J.; Stuczyński, T. Impact of Water Stress on Microbial Community and Activity in Sandy and Loamy Soils. Agronomy 2020, 10, 1429. [CrossRef]
- Barnard, R.L.; Blazewicz, S.J.; Firestone, M.K. Rewetting of soil: Revisiting the origin of soil CO2 emissions. Soil Biology and Biochemistry 2020, 147, 107819. [CrossRef]
- Liao, Z.; Junliang, F., Zhenlin, L.; Zhentao, B.; Haidong, W.; Minghui, C.; Fucang, Z.; Zhijun, L. Chapter Three - Response network and regulatory measures of plant-soil-rhizosphere environment to drought stress. Advances in Agronomy, 2023, 180, 93-196. [CrossRef]
- Xu, L.; Dong, Z.; Chiniquy, D.; Pierroz, G.; Deng, S.; Gao, C.; Diamond, S.; Simmons, T.; Wipf, H.M.; Caddell, D.; Varoquaux, N.; Madera, M.A.; Hutmacher, R.; Deutschbauer, A.; Dahlberg, J.A; Guerinot, M.L.; Purdom, E.; Banfield, J.F.; Taylor, J.W.; Lemaux, P.G.; Coleman-Derr, D. Genome-resolved metagenomics reveals role of iron metabolism in drought-induced rhizosphere microbiome dynamics. Nat Commun. 2021, 12(1), 3209. [CrossRef]
- Estrada-González, Á.J.; Medina-De la Rosa, G.; Bautista, E.; Flores, J.; López-Lozano, N.E. Physiological regulations of a highly tolerant cactus to dry season modify its rhizospheric microbial communities. Rhizosphere, 2023, 25, 100655. [CrossRef]
- Hestrin, R.; Kan, M.; Lafler, M.; Wollard, J.; Kimbrel, J.A.; Ray, P.; Blazewicz, S.J.; Stuart, R.; Craven, K.; Firestone, M.; Nuccio, E.E.; Pett-Ridge, J. Plant-associated fungi support bacterial resilience following water limitation. ISME J. 2022, 16(12), 2752-2762. [CrossRef]
- Milošević, N.A.; Marinković, J.B.; Tintor, B.B. Mitigating abiotic stress in crop plants by microorganisms. Proc. Nat. Sci. Matica Serpska Novi. Sad 2012, 123, 17–26. [CrossRef]
- Bhat, B.A.; Tariq, L.; Nissar, S.; Islam, S.T.; Islam, S.U.; Mangral, Z.; Ilyas, N.; Sayyed, R.Z.; Muthusamy, G.; Kim, W.; Dar, T.U.H. The role of plant-associated rhizobacteria in plant growth, biocontrol and abiotic stress management. J Appl Microbiol. 2022, 133(5), 2717-2741. [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front Plant Sci. 2018, 9, 1473. [CrossRef]
- Chieb, M.; Gachomo, E.W. The role of plant growth promoting rhizobacteria in plant drought stress responses. BMC Plant Biol. 2023, 23(1), 407. [CrossRef]
- Berendsen, Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478-486. [CrossRef]
- Öpik, M.; Zobel, M.; Cantero, J.J.; Davison, J.; Facelli, J.M.; Hiiesalu, I.; Jairus, T.; Kalwij, J.M.; Koorem, K.; Leal, M.E.; Liira, J.; Metsis, M.; Neshataeva, V.; Paal, J.; Phosri, C.; Põlme, S.; Reier, Ü.; Saks, Ü.; Schimann, H.; Thiéry, O.; Vasa, M.; Moora, M. Global sampling of plant roots expands the described molecular diversity of arbuscular mycorrhizal fungi. Mycorrhiza 2013, 23(5),411-30. [CrossRef]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbiosis. Nat. Rev. Microbiol. 2008, 6, 763-775. [CrossRef]
- Bennett, A.E.; Groten, K. The costs and benefits of plant–arbuscular mycorrhizal fungal interactions. Annual Review of Plant Biology 2022, 73, 649-672. [CrossRef]
- Bittencourt, P.P.; Alves, A.F.; Ferreira, M.B.; da Silva Irineu, L.E.S.; Pinto, V.B.; Olivares, F.L. Mechanisms and Applications of Bacterial Inoculants in Plant Drought Stress Tolerance. Microorganisms 2023, 11, 502. [CrossRef]
- Prasad, M.; Srinivasan, R.; Chaudhary, M.; Choudhary, M.; Jat, L. K. Plant growth promoting rhizobacteria (PGPR) for sustainable agriculture: Perspectives and challenges. PGPR amelioration in sustainable agriculture 2019, 129-157. [CrossRef]
- Wahab, A.; Muhammad, M.; Munir, A.; Abdi, G.; Zaman, W.; Ayaz, A.; Khizar, C.; Reddy, S. P. P. Role of arbuscular mycorrhizal fungi in regulating growth, enhancing productivity, and potentially influencing ecosystems under abiotic and biotic stresses. Plants 2023, 12(17), 3102. [CrossRef]
- Bao, X.; Zou, J.; Zhang, B.; Wu, L.; Yang, T.;Huang, Q. Arbuscular mycorrhizal fungi and microbes interaction in rice mycorrhizosphere. Agronomy 2022, 12(6), 1277. [CrossRef]
- Agudelo, M.G.; Ruiz, B., Capela, D.; Remigi P. The role of microbial interactions on rhizobial fitness. Front Plant Sci. 2023, 14,1277262. [CrossRef]
- Santos, M.S.; Nogueira, M.A.; Hungria, M. Microbial inoculants: Reviewing the past, discussing the present and previewing an outstanding future for the use of beneficial bacteria in agriculture. AMB Express 2019, 9(1), 205. [CrossRef]
- Fred, E.B.; Baldwin, I.L.; McCoy, E. Some factors which influence the growth and longevity of the nodule bacteria. In Root nodule bacteria and leguminous plants; Baldwin, I. L.; McCoy, E.; Fred, E. B.; Publisher: University of Wisconsin, Madison, WI, 1932, Volume 5, pp. 104-117.
- Massa, F.; Defez, R.; Bianco, C. Exploitation of Plant Growth Promoting Bacteria for Sustainable Agriculture: Hierarchical Approach to Link Laboratory and Field Experiments. Microorganisms 2022, 10(5), 865. [CrossRef]
- Kloepper, J.; Leong, J.; Teintze, M.; Milton N.S. Enhanced plant growth by siderophores produced by plant growth-promoting rhizobacteria. Nature 1980, 286, 885–886. [CrossRef]
- Amaresan, N.; Kumar, M.S.; Annapurna, K.; Kumar, K.; Sankaranaryanan, N. Beneficial Microbes in Agro-Ecology: Bacteria and Fungi; Academic Press: Cambridge, MA, USA, 2020.
- Santos L.F.; Olivares, F. L. Plant microbiome structure and benefits for sustainable agriculture. Current Plant Biology 2021, 26, 100198. ISSN 2214-6628. [CrossRef]
- Lehmann, J.; Bossio D.A.; Kögel-Knabner I., Rillig, M.C. The concept and future prospects of soil health. Nat. Rev. Earth Environ. 2020, 1, 544-553. [CrossRef]
- Hanaka, A.; Ozimek, E.; Reszczyńska, E.; Jaroszuk-Ściseł, J.; Stolarz, M. Plant Tolerance to Drought Stress in the Presence of Supporting Bacteria and Fungi: An Efficient Strategy in Horticulture. Horticulture 2021, 7, 390. [CrossRef]
- Rubin, R.L.; van Groenigen, K.J.; Hungate, B.A. Plant growth promoting rhizobacteria are more effective under drought: A meta-analysis. Plant Soil 2017, 416, 309–323. [CrossRef]
- Igiehon, O.N.; Babalola, O.O. Rhizobium and mycorrhizal fungal species improved soybean yield under drought stress conditions. Current Microbiology 2021, 78(4), 1615-1627. [CrossRef]
- Trivedi, P.; Batista, B.D.; Bazany, K.E.; Singh, B.K. Plant–microbiome interactions under a changing world: Responses, consequences and perspectives. N. Phytol. 2022, 234, 1951–1959. [CrossRef]
- Suryanarayanan, T.S.; Shaanker, R.U. Can fungal endophytes fast-track plant adaptations to climate change? Fungal Ecol. 2021, 50, 101039. [CrossRef]
- Marasco, H.I.; Rolli, E.; Vigani, G.; Borin, S.; Sorlini, C.; Ouzari, H.; Zocchi, G.; Daffonchio, D. Are drought-resistance promoting bacteria cross-compatible with different plant models? Plant and Soil 2016, 399(1-2), 219-229. [CrossRef]
- Naylor, D.; Coleman-Derr, D. Drought stress and root-associated bacterial communities. Frontiers in Plant Science 2018, 8, 2223. [CrossRef]
- Maryani, Y.; Dewi, W.S.; Yunus, A. Study on osmoprotectant rhizobacteria to improve mung bean growth under drought stress. In IOP Conference Series: Earth and Environmental Science 2018, 129(1), 012014. [CrossRef]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and function of the bacterial root microbiota in wild and domesticated barley. Cell Host Microbe 2015, 17(3), 392-403. [CrossRef]
- Cytryn, E.J.; Sangurdekar, D.P.; Streeter, J.G.; Franck, W.L.; Chang, W.S.; Stacey, G.; Emerich, D.W.; Joshi, T.; Xu, D.; Sadowsky, M.J. Transcriptional and physiological responses of Bradyrhizobium japonicum to desiccation-induced stress. J Bacteriol. 2007, 189(19), 6751-6762. [CrossRef]
- Zhu, J.; Jiang, X.; Guan, D.; Kang, Y.; Li, L.; Cao, F.; Zhao, B.; Ma, M.; Zhao, J.; Li, J. Effects of rehydration on physiological and transcriptional responses of a water-stressed rhizobium. J Microbiol. 2022, 60(1), 31-46. [CrossRef]
- Vílchez, J.I.; García-Fontana, C.; Román-Naranjo, D.; González-López, J.; Manzanera, M. Plant drought tol-erance enhancement by trehalose production of desiccation-tolerant microorganisms. Frontiers in Microbiology 2016, 7, 1577.
- Sharma, M.P.; Grover, M.; Chourasiya, D., Bharti, A., Agnihotri, R., Maheshwari, H.S.; Pareek, A.; Buyer, J.S.; Sharma, S.K.; Schütz, L.; Mathhimaran, N.; Singla-Pareek, S.L.; Grossman, J.M.; Bagyaraj, D. J. Deciphering the role of trehalose in tripartite symbiosis among rhizobia, arbuscular mycorrhizal fungi, and legumes for enhancing abiotic stress tolerance in crop plants. Frontiers in microbiology 2020, 11, 509919. [CrossRef]
- McIntyre, H.J.; Davies, H.; Hore, T.A.; Miller, S.H.; Dufour, J.P.; Ronson, C.W. Trehalose biosynthesis in Rhizobium leguminosarum bv. trifolii and its role in desiccation tolerance. Appl Environ Microbiol. 2007, 73(12), 3984-3992. [CrossRef]
- Iturriaga, G.; Suárez, R.; Nova-Franco, B. Trehalose metabolism: From osmoprotection to signaling. Int. J. Mol. Sci. 2009, 10(9), 3793-3810. [CrossRef]
- Nawaz, M.; Hassan, M.U.; Chattha, M.U.; Mahmood, A.; Shah, A.N.; Hashem, M.; Alamri, S.; Batool, M.; Rasheed, A.; Thabit, M.A.; Alhaithloul, H.A.S.; Qari, S.H. Trehalose: A promising osmo-protectant against salinity stress-physiological and molecular mechanisms and future prospective. Mol. Biol. Rep. 2022, 49(12), 11255-11271. [CrossRef]
- Dukare, A.; Mhatre, P.; Maheshwari, H.S.; Delineation of mechanistic approaches of rhizosphere microorganisms facilitated plant health and resilience under challenging conditions. 3 Biotech 2022, 12, 57. [CrossRef]
- Manzanera, M.; Garcia de Castro, A.; Tondervik, A.; Rayner-Brandes,M.; Strom, A.R.; Tunnacliffe, A. Hydroxyectoine is superior to trehalose for anhydrobiotic engineering of Pseudomonas putida KT2440. Appl. Environ. Microbiol. 2002, 68, 4328-4333. [CrossRef]
- Narváez-Reinaldo, J.J.; Barba, I.; González-López J.; Tunnacliffe, A.; Manzanera, M. Rapid method for isolation of desiccation-tolerant strains and xeroprotectants. Appl Environ Microbiol. 2010, 76(15), 5254-62. [CrossRef]
- SantaCruz-Calvo, L.; González-López, J.; Manzanera, M. Arthrobacter siccitolerans sp. nov., a highly desiccation-tolerant, xeroprotectant-producing strain isolated from dry soil. Int J Syst Evol Microbiol. 2013, 63(Pt11), 4174-4180. [CrossRef]
- Peterson, C.; Niraula, S.; Parks, D.; Chang, W.S. Draft Genome Sequences of Two Desiccation-Tolerant Strains, Bradyrhizobium japonicum TXVA and TXEA, Isolated from the Root Nodules of Soybean Grown in Texas. Microbiol Resour Announc. 2022, 11(8), e0046722. [CrossRef]
- Pazos-Rojas, L.A.; Cuellar-Sánchez, A.; Romero-Cerón, A.L.; Rivera-Urbalejo, A.; Van Dillewijn, P.; Luna-Vital, D.A.; Muñoz-Rojas, J.; Morales-García, Y.E.;Bustillos-Cristales,M.D.R. The Viable but Non-Culturable (VBNC) State, a Poorly Explored Aspect of Beneficial Bacteria. Microorganisms 2023, 12(1), 39. [CrossRef]
- Muñoz-Rojas, J. Desiccation-tolerant rhizobacteria maintain their plant growth-promoting capability after experiencing extreme water stress. SciFed J. Appl. Microbiol 2018, 1, 15-17.
- Shankar, A.; Prasad, V. Potential of desiccation-tolerant plant growth-promoting rhizobacteria in growth augmentation of wheat (Triticum aestivum L.) under drought stress. Front Microbiol. 2023, 14, 1017167. [CrossRef]
- Bahadur, A.; Batool, A.; Nasir, F.; Jiang, S.; Mingsen, Q.; Zhang, Q.; Pan, J.; Liu, Y.; Feng, H.; Mechanistic Insights into Arbuscular Mycorrhizal Fungi-Mediated Drought Stress Tolerance in Plants. Int. J. Mol. Sci. 2019, 20(17), 4199. [CrossRef]
- Millar, N.S.; Bennett, A.E. Stressed out symbiotes: Hypotheses for the influence of abiotic stress on arbuscular mycorrhizal fungi. Oecologia 2016, 182, 625–641. [CrossRef]
- Zhang, F.; Ying-Ning, Z.; Wu, Q. Quantitative estimation of water uptake by mycorrhizal extraradical hyphae in citrus under drought stress. Scientia Horticulturae 2018, 229, 132-136. [CrossRef]
- Liu, X. J. A.; Han, S.; Frey, S. D.; Melillo, J. M.; Zhou, J.; DeAngelis, K. M. Microbial responses to long-term warming differ across soil microenvironments. ISME Commun 2024, 4(1), ycae051. [CrossRef]
- Symanczik, S.; Courty, P.E.; Boller, T.; Wiemken, A.; Al-Yahya’ei, M.N. Impact of water regimes on an experimental community of four desert arbuscular mycorrhizal fungal (AMF) species, as affected by the intro-duction of a non-native AMF species. Mycorrhiza 2015, 25, 639-647. [CrossRef]
- Lumini, E.; Vallino, M.; Alguacil, M.M.; Romani, M.; Bianciotto, V. Different farming and water regimes in Italian rice fields affect arbuscular mycorrhizal fungal soil communities. Ecological Applications 2011, 21(5), 1696-1707. [CrossRef]
- Ruiz-Lozano, J.M.; Porcel, R.; Azcón, C.; Aroca, R. Regulation by arbuscular mycorrhizae of the integrated physiological response to salinity in plants: New challenges in physiological and molecular studies. J. Exp. Bot. 2012, 63(11), 4033-44. [CrossRef]
- Auge, R. M. Mycorrhiza and drought resistance. In Mycorrhiza - Function, Diversity, State of the Art, 2020. 207-226. IntechOpen.
- Balestrini, R.; Nerva, L.; Sillo, F.; Girlanda, M.; Perotto, S.; Plant and fungal gene expression in mycorrhizal protocorms of the orchid Serapias vomeracea colonized by Tulasnella calospora. Plant Signal Behav 2014, 9(11), e977707. [CrossRef]
- Miransari, M.; Interactions between arbuscular mycorrhizal fungi and soil bacteria. Appl Microbiol Biotechnol. 2011, 89(4), 917-930. [CrossRef]
- Zadworny, M., Górska, A., Politycka, B. Arbuscular mycorrhizal fungi alter enzymatic activities in phosphorus-transforming soils. Mycorrhiza, 2015, 25(4), 243-251.
- Chourasiya, D.; Gupta, M.M.; Sahni, S.; Oehl, F.; Agnihotri, R.; Buade, R.; Maheshwari, H.S.; Prakash, A.; Sharma, M. P. Unraveling the AM fungal community for understanding its ecosystem resilience to changed climate in agroecosystems. Symbiosis 2021, 84(3), 295-310. [CrossRef]
- Stahl, P.D.; Christensen, M. Population variation in the mycorrhizal fungus Glomus mosseae: Breadth of environmental tolerance. Mycological Research 1991, 95(3), 300-307. [CrossRef]
- Mahmoudi, N.; Cruz, C.; Mahdhi, M.; Mars, M.; Caeiro, M.F. Arbuscular mycorrhizal fungi in soil, roots and rhizosphere of Medicago truncatula: Diversity and heterogeneity under semi-arid conditions. PeerJ 2019, 7, e6401. [CrossRef]
- Augé, R.M.; Toler, H.D.; Saxton, A.M. Arbuscular mycorrhizal symbiosis alters stomatal conductance of host plants more under drought than under amply watered conditions: A meta-analysis. Mycorrhiza 2015, 25(1), 13-24. [CrossRef]
- Veresoglou, S.D.; Chen, B.; Rillig, M.C. Arbuscular mycorrhiza and soil nitrogen cycling. Soil Biology and Biochemistry 2012, 46, 53-62. [CrossRef]
- Berruti, A.; Lumini, E.; Balestrini, R.; Bianciotto, V. Arbuscular Mycorrhizal Fungi as Natural Biofertilizers: Let’s Benefit from Past Successes. Frontiers in Microbiology 2016, 6, 1559. [CrossRef]
- Moreira-Souza, M.; Trufem, S. F. B.; Gomes-Da-Costa, S. M.; Cardoso, E. J. B. N. Arbuscular mycorrhizal fungi associated with Araucaria angustifolia (Bert.) O. Ktze. Mycorrhiza 2003, 13(4), 211-5. [CrossRef]
- Lozano, Y. M.; Aguilar-Trigueros, C.A.; Roy, J.; Rillig, M.C. Drought induces shifts in soil fungal communities that can be linked to root traits across 24 plant species. New Phytologist 2021, 232, 1917-1929. [CrossRef]
- Kuyper, T.W.; Jansa J. Arbuscular mycorrhiza: Advances and retreats in our understanding of the ecological functioning of the mother of all root symbioses. Plant and Soil 2023, 489(1), 41-88. [CrossRef]
- Safari, M.M.R.; Farokhzad, M.; Kaviani, B.; Kulus, D. Endophytic Fungi as Potential Biocontrol Agents against Sclerotium rolfsii Sacc. - The Causal Agent of Peanut White Stem Rot Disease. Cells 2022, 11(17), 2643. [CrossRef]
- Grenville-Briggs, L.J.; Avrova, A.O.; Bruce, C. R.; Williams, A.; Whisson, S.C.; Birch, P.R.J.; West, P.V. Elevated temperature and CO2 levels affect the interactions between potato and the root pathogen Phytophthora infestans. Global Change Biology 2005, 11(9), 1714–1722. [CrossRef]
- El-Abyad, M.S.; Attaby, H.; Abu-Taleb, A.M. Impact of salinity stress on the free amino acid pools of some phytopathogenic fungi. Microbiological research 1994, 149(3), 309-315. [CrossRef]
- Rossier, O.; Vorhölter, F. J. New Insights into the Extracellular Secretion of Xanthomonads. Trends in Microbiology 2019, 27(7), 605–614.
- Bohnert, H. J.; Jensen, R. G.; Strategies for engineering water-stress tolerance in plants. Trends in Biotechnology 1996, 14, 89-97. [CrossRef]
- Peng, Y.; Li, S. J.; Yan, J.; Tang, Y.; Cheng, J. P.; Gao, A. J.; Yao, X.; Ruan, J. J.; Xu, B. L. Research Progress on Phytopathogenic Fungi and Their Role as Biocontrol Agents. Frontiers in microbiology 2021, 12, 670135. [CrossRef]
- van der Wolf, J., De Boer, S.H. Phytopathogenic bacteria. In Principles of plant-microbe interactions: Microbes for Sustainable Agriculture. 2014. pp. 65-77. Cham: Springer International Publishing.
- Delgado-Baquerizo, M.; Guerra, C. A.; Cano-Díaz, C.; Egidi, E.; Wang, J. T.; Eisenhauer, N.; Singh, B.; Maestre, F. The proportion of soil-borne pathogens increases with warming at the global scale. Nature Climate Change 2020. 10, 550-554. https://www.nature.com/articles/s41558-020-0759-3.
- Wakelin, S. A.; Gomez-Gallego, M.; Jones, E.; Smaill, S.; Lear, G.; Lambie, S. Climate change induced drought impacts on plant diseases in New Zealand. Australasian Plant Pathology 2018, 47, 101-114. [CrossRef]
- Rai, A.; Irulappan, V.; Muthappa, S. K. Dry root rot of chickpea: A disease favored by drought. Plant Disease 2021. [CrossRef]
- Sharath Chandran, U.S.; Tarafdar, A.; Mahesha, H. S.; Sharma, M. Temperature and Soil Moisture Stress Modulate the Host Defense Response in Chickpea During Dry Root Rot Incidence. Frontiers in plant science 2021. 12, 653265. [CrossRef]
- Chilakala, A.R.; Mali, K. V.; Irulappan, V.; Patil, B. S.; Pandey, P.; Rangappa, K.; Ramegowda, V.; Kumar, M. N.; Puli, C.O.R.; Mohan-Raju, B.; Senthil-Kumar, M. Combined Drought and Heat Stress Influences the Root Water Relation and Determine the Dry Root Rot Disease Development Under Field Conditions: A Study Using Contrasting Chickpea Genotypes. Frontiers in plant science 2022, 13, 890551. [CrossRef]
- Batista, E.; Lopes, A.; Miranda, P.; Alves, A. Can species distribution models be used for risk assessment analyses of fungal plant pathogens? A case study with three Botryosphaeriaceae species. European Journal of Plant Pathology 2023, 165(1), 41-56. [CrossRef]
- Oliva, J.; Stenlid, J.; Martínez-Vilalta, J. The effect of fungal pathogens on the water and carbon economy of trees: Implications for drought-induced mortality. New Phytologist 2014, 203: 1028-1035. [CrossRef]
- Al-Turki, A.; Murali, M.; Omar, A. F.; Rehan, M.; Sayyed, R.Z.; Recent advances in PGPR-mediated resilience toward interactive effects of drought and salt stress in plants. Front Microbiol. 2023, 27(14), 1214845. [CrossRef]
- Khan, N.; Ali, S.; Tariq, H.; Latif, S.; Yasmin, H.; Mehmood, A.; Shahid, M.A. Water Conservation and Plant Survival Strategies of Rhizobacteria under Drought Stress. Agronomy 2020, 10, 1683. [CrossRef]
- Jajoo, A.; Mathur, S. Role of arbuscular mycorrhizal fungi as an underground saviuor for protecting plants from abiotic stresses. Physiology and Molecular Biology of Plants 2021, 27(11), 2589-2603. [CrossRef]
- Newman, E.D.; Rowland, J.B.; Hammer, T.G. Frost, L.A.; Lumibao, C.Y.; Henning, J. A. Trade-Offs in Arbuscular Mycorrhizal Fungal Responses to Drought and Salinity Stress in Panicum amarum (United States Gulf Coast). Journal of Coastal Research 2024, 40(1), 51-63. [CrossRef]
- Igiehon, N.O.; Babalola, O.O.; Cheseto, X.; Torto, B. Effects of rhizobia and arbuscular mycorrhizal fungi on yield, size distribution and fatty acid of soybean seeds grown under drought stress. Microbiological Research 2021, 242, 126640. [CrossRef]
- Vaishnav, A., Kasotia, A., Choudhary, D.K. Role of Functional Bacterial Phylum Proteobacteria in Glycine max Growth Promotion Under Abiotic Stress: A Glimpse on Case Study. In Silico Approach for Sustainable Agriculture; Choudhary, D., Kumar, M., Prasad, R., Kumar, V.; Publisher: Springer, Singapore, 2018; pp. 17-49. [CrossRef]
- Nawaz, M.; Ishaq, S.; Ishaq, H.; Khan, N.; Iqbal, N.; Ali, S.; Rizwan, M.; Alsahli, A.A.; Alyemeni, M.N. Salicylic Acid Improves Boron Toxicity Tolerance by Modulating the Physio-Biochemical Characteristics of Maize (Zea mays L.) at an Early Growth Stage. Agronomy 2020, 10, 2013. [CrossRef]
- Lin, Y.; Watts, D.B.; Kloepper, J.W.; Feng, Y.; Torbert, H.A. Influence of plant growth-promoting rhizobacteria on corn growth under drought stress. Communications in Soil Science and Plant Analysis 2020, 51(2), 250-264. [CrossRef]
- Fadiji, A.E.; Santoyo, G.; Yadav, A.N.; Babalola, O.O. Efforts towards overcoming drought stress in crops: Revisiting the mechanisms employed by plant growth-promoting bacteria. Frontiers in Microbiology 2022, 13, 962427. [CrossRef]
- Admassie, M.; Woldehawariat, Y.; Alemu, T.; Gonzalez, E.; Jimenez, J.F. The role of plant growth-promoting bacteria in alleviating drought stress on pepper plants. Agricultural Water Management 2022, 272, 107831. [CrossRef]
- Kohler, J.; Hernández, J. A.; Caravaca, F.; Roldán, A.; Plant-growth-promoting rhizobacteria and arbuscular mycorrhizal fungi modify alleviation biochemical mechanisms in water-stressed plants. Funct Plant Biol. 2008, 35(2), 141-151. [CrossRef]
- Amini, R.; Zafarani-Moattar, P.; Shakiba, M.R.; Hasanfard, A. Inoculating moldavian balm (Dracocephalum moldavica L.) with mycorrhizal fungi and bacteria may mitigate the adverse effects of water stress. Scientific Reports 2023, 13(1), 16176. [CrossRef]
- Mondani, F.; Khani, K.; Honarmand, S.J.; Saeidi, M. Evaluating effects of plant growth-promoting rhizobacteria on the radiation use efficiency and yield of soybean (Glycine max) under water deficit stress condition. Agricultural water management 2019, 213, 707-713. [CrossRef]
- Khan, N.; Bano, A. Exopolysaccharide producing rhizobacteria and their impact on growth and drought tolerance of wheat grown under rainfed conditions. PLoS ONE 2019, 14(9), e0222302. [CrossRef]
- Zaheer, M.S.; Raza, M.A.S.; Saleem, M. F.; Erinle, K.O.; Iqbal, R.; Ahmad, S. Effect of rhizobacteria and cytokinins application on wheat growth and yield under normal vs drought conditions. Communications in Soil Science and Plant Analysis 2019, 50(20), 2521-2533. [CrossRef]
- Muhammad, F.; Raza, M. A. S;. Iqbal, R.; Zulfiqar, F.; Aslam, M.U.; Yong, J.W.H.; Altaf, M.A.; Zulfiqar, F.; Amin, J.; Ibrahim, M. A. Ameliorating drought effects in wheat using an exclusive or co-applied rhizobacteria and ZnO nanoparticles. Biology 2022, 11(11), 1564. [CrossRef]
- de Souza, R.; Ambrosini, A.; Passaglia, L. M. P. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet Mol Biol. 2015, 38(4), 401-19. [CrossRef]
- Swarnalakshmi, K.; Yadav, V.; Tyagi, D.; Dhar, D. W.; Kannepalli, A.; Kumar, S.; Significance of Plant Growth Promoting Rhizobacteria in Grain Legumes: Growth Promotion and Crop Production. Plants 2020, 9(11), 1596. [CrossRef]
- Zandi, P.; Schnug, E.; Reactive Oxygen Species, Antioxidant Responses and Implications from a Microbial Modulation Perspective. Biology 2022, 11(2), 155. [CrossRef]
- Tiepo, A. N.; Hertel, M. F.; Rocha, S. S.; Calzavara, A. K.; Oliveira, A. L. M. D.; Pimenta, J. A.; Oliveira, H. C.; Bianchini, E.; Stolf-Moreira, R.; Enhanced drought tolerance in seedlings of Neotropical tree species inoculated with plant growth-promoting bacteria. Plant Physiol Biochem 2018, 130, 277-288. [CrossRef]
- Siddikee, M. A.; Glick, B. R.; Chauhan, P. S.; jong Yim, W.; Sa, T. Enhancement of growth and salt tolerance of red pepper seedlings (Capsicum annuum L.) by regulating stress ethylene synthesis with halotolerant bacteria containing 1-aminocyclopropane-1-carboxylic acid deaminase activity. Plant Physiol Biochem 2011, 49(4), 427-434. [CrossRef]
- Carezzano, M.E.; Alvarez Strazzi, F.B.; Pérez, V.; Bogino, P.; Giordano, W. Exopolysaccharides Synthesized by Rhizospheric Bacteria: A Review Focused on Their Roles in Protecting Plants against Stress. Appl. Microbiol. 2023, 3, 1249-1261. [CrossRef]
- Bhargavi, G.; Arya, M.; Jambhulkar, P.P.; Singh, A.; Rout, A. K.; Behera, B.K.; Chaturvedi, S.K.; Singh, A.K. Evaluation of biocontrol efficacy of rhizosphere dwelling bacteria for management of Fusarium wilt and Botrytis gray mold of chickpea. BMC Genom Data 2024, 25(1), 7. [CrossRef]
- Huang, T.; Zhang, Y.; Yu Z.; Zhuang, W.; Zeng, Z. Bacillus velezensis BV01 Has Broad-Spectrum Biocontrol Potential and the Ability to Promote Plant Growth. Microorganisms 2023, 11(11), 2627. [CrossRef]
- Xie, J.; Singh, P.; Qi, Y.; Singh, R.K.; Qin, Q.; Jin, C.; Wang, B.; Fang, W. Pseudomonas aeruginosa Strain 91: A Multifaceted Biocontrol Agent against Banana Fusarium Wilt. Fungi 2023, 9(11), 1047. [CrossRef]
- Bouremani, N.; Cherif-Silini, H.; Silini, A.; Bouket, A.C.; Luptakova, L.; Alenezi, F.N., Baranov, O.; Belbahri, L. Plant growth-promoting rhizobacteria (PGPR): A rampart against the adverse effects of drought stress. Water 2023, 15(3), 418. [CrossRef]
- Kaushal, M.; Wani, S.P. Rhizobacterial-plant interactions: Strategies ensuring plant growth promotion under drought and salinity stress. Agriculture, Ecosystems & Environment, 2016, 231, 68-78. [CrossRef]
- Etesami, H.; Maheshwari, D. K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol Environ Saf 2018, 30(156), 225-246. [CrossRef]
- Vu, B.; Chen, M.; Crawford, R. J.; Ivanova, E. P.; Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 2009, 14(7), 2535-54. [CrossRef]
- Sato, Y.; Miwa, T.; Inaba, T.; Akachi, T.; Tanaka, E.; Hori, T.; Murofushi, K.; Takagi, H.; Futamata, H.; Aoyagi, T.; Habe, H. Microbially produced fertilizer provides rhizobacteria to hydroponic tomato roots by forming beneficial biofilms. Appl Microbiol Biotechnol. 2023, 107(23), 7365-7374. [CrossRef]
- Fadiji, A.E.; Orozco-Mosqueda, M.d.C.; Santos-Villalobos, S.d.l.; Santoyo, G.; Babalola, O.O. Recent Developments in the Application of Plant Growth-Promoting Drought Adaptive Rhizobacteria for Drought Mitigation. Plants 2022, 11, 3090. [CrossRef]
- Mitra, D.; Djebaili, R.; Pellegrini, M.; Mahakur, B.; Sarker, A.; Chaudhary, P.; Khoshru, B.; Gallo, M. D.; Kitouni, M.; Barik, D.P.; Panneerselvam, P.; Mohapatra, P.K.D. Arbuscular mycorrhizal symbiosis: Plant growth improvement and induction of resistance under stressful conditions. Journal of Plant Nutrition 2021, 44(13), 1993-2028. [CrossRef]
- Liu, R.C.; Ding, Y.E.; Wu, Q.S.; Zou, Y.N. Mycorrhizae enhance drought tolerance of trifoliate orange by regulating circadian clock response patterns. Scientia Horticulturae 2022, 305, 111426. [CrossRef]
- Dowarah, B.; Gill, S. S.; Agarwala, N. Arbuscular mycorrhizal fungi in conferring tolerance to biotic stresses in plants. Journal of Plant Growth Regulation 2022, 41(4), 1429-1444. [CrossRef]
- Latef, A.A.H.A.; Hashem, A.; Rasool, S.; Abd_Allah, E.F.; Alqarawi, A.A.; Egamberdieva, D.; Jan, S.; Anjum, N.A.; Ahmad, P. Arbuscular mycorrhizal symbiosis and abiotic stress in plants: A review. Journal of plant biology 2016, 59, 407-426. [CrossRef]
- Wang, Y.; Zou, Y.N.; Shu, B.; Wu, Q.S. Deciphering molecular mechanisms regarding enhanced drought tolerance in plants by arbuscular mycorrhizal fungi. Scientia Horticulturae 2022, 308, 111591. [CrossRef]
- Zou, Y.; Wu, Q.; Kuča, K. Unravelling the role of arbuscular mycorrhizal fungi in mitigating the oxidative burst of plants under drought stress. Plant Biology 2021, 23, 50-57. [CrossRef]
- Gholamhoseini, M.; Ghalavand, A.; Dolatabadian, A.; Jamshidi, E.; Khodaei-Joghan, A. Effects of arbuscular mycorrhizal inoculation on growth, yield, nutrient uptake and irrigation water productivity of sunflowers grown under drought stress. Agricultural Water Management 2013, 117, 106-114. [CrossRef]
- Zhang, X.; Yan, J.; Khashi u Rahman, M.; Wu, F. The impact of root exudates, volatile organic compounds, and common mycorrhizal networks on root system architecture in root-root interactions. Journal of Plant Interactions 2022, 17(1), 685-694. [CrossRef]
- Cheng, H.Q.; Zou, Y. N.; Wu, Q.S.; Kuča, K. Arbuscular mycorrhizal fungi alleviate drought stress in trifoliate orange by regulating H+-ATPase activity and gene expression. Frontiers in Plant Science 2021, 12, 659694. [CrossRef]
- Zou, Y.; Qin, Q.; Ma, W.; Zhou, L.; Wu, Q.; Xu, Y.; Kuča, K.; Hashem, A.; Al-Arjani, A. F.; Almutairi, K. F.; Abd-Allah, E. F. Metabolomics reveals arbuscular mycorrhizal fungi-mediated tolerance of walnut to soil drought. BMC Plant Biol. 2023, 23(1), 118. [CrossRef]
- Sheteiwy, M.S.; Elgawad, H.A.; Xiong, Y.; Macovei, A.; Brestic, M.; Skalicky, M.; Shaghaleh, H.; Hamoud, Y. A.; El-Sawah, A.M. Inoculation with Bacillus amyloliquefaciens and mycorrhiza confers tolerance to drought stress and improve seed yield and quality of soybean plant. Physiol Plant. 2021, 172(4), 2153-2169. [CrossRef]
- Sheteiwy, M.S.; Ali, D. F. I.; Xiong, Y.C.; Brestic, M.; Skalicky, M.; Hamoud, Y.A.; Ulhassan, Z.; Shaghaleh, H.; AbdElgawad, H.; Farooq, M.; Sharma, A.; El-Sawah, A. M. Physiological and biochemical responses of soybean plants inoculated with Arbuscular mycorrhizal fungi and Bradyrhizobium under drought stress. BMC plant biology 2021, 21(1), 195. [CrossRef]
- Xu, L.; Li, T.; Wu, Z.; Feng, H.; Yu, M.; Zhang, X.; Chen, B. Arbuscular mycorrhiza enhances drought tolerance of tomato plants by regulating the 14-3-3 genes in the ABA signaling pathway. Applied soil ecology 2018, 125, 213-221. [CrossRef]
- Drogue, B.; Doré, H.; Borland, S.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Which specificity in cooperation between phytostimulating rhizobacteria and plants? Research in microbiology 2012, 163(8), 500–510. [CrossRef]
- Matilla, M. A.; Ramos, J. L. Biosurfactant produced by a Pseudomonas strain growing on polycyclic aromatic hydrocarbons. Applied and Environmental Microbiology 2007, 73(4), 1423–1429. [CrossRef]
- Newman, K. L.; Almeida, R. P. P.; Purcell, A. H.; Lindow, S. E. Cell-cell signaling controls Xylella fastidiosa interactions with both insects and plants. Proc Natl Acad Sci USA 2004, 101(6), 1737-42. [CrossRef]
- Drogue, B.; Sanguin, H.; Chamam, A.; Mozar, M.; Llauro, C.; Panaud, O.; Prigent-Combaret, C.; Picault, N.; Wisniewski-Dyé, F. Plant root transcriptome profiling reveals a strain-dependent response during Azospirillum–rice cooperation. Frontiers in Plant Science 2014, 5, 607. [CrossRef]
- Michielse, C.B.; Rep, M. Pathogen profile update: Fusarium oxysporum. Mol Plant Pathol 2009, 10(3), 311-24. [CrossRef]
- Elad, Y.; Yunis, H.; Katan, J. Multiple resistance mechanisms to benzimidazole fungicides in Botrytis cinerea field isolates. Phytopathology 2007, 97(6), 686–695. [CrossRef]
- Büttner, D.; Bonas, U. Regulation and secretion of Xanthomonas virulence factors. FEMS Microbiol Rev 2010, 34(2), 107-33. [CrossRef]
- Shoresh, M.; Harman, G. E.; Mastouri, F. Induced systemic resistance and plant responses to fungal biocontrol agents. Annu Rev Phytopathol 2010, 48, 21-43. [CrossRef]
- Flemming, H.C.; Wingender, J.; The biofilm matrix. Nature Reviews Microbiology 2010, 8(9), 623–633. [CrossRef]
- de Pontes, J.G.M.; Fernandes, L.S.; Santos, R.V.D.; Tasic, L.; Fill, T.P. Virulence Factors in the Phytopathogen-Host Interactions: An Overview J Agric Food Chem. 2020, 68(29), 7555-7570. [CrossRef]
- Garbelotto, M. Host and environmental feed-backs on invasive plant pathogens: Ecological and evolutionary consequences on novel plant pathogen interactions. Journal of Plant Pathology 2012, 94(4), S41-S84.
- Yuen, J., Collinge, D.B., Djurle, A. and Tronsmo, A.M. Plant Pathology in a Changing World 23. In Plant Pathology and Plant Diseases; Tronsmo, A.M.; Collinge, D.B.; Djurle, A.; Munk, L.; Yuen, J.; Tronsmo A., 2020, pp. 379-386. [CrossRef]
- Yao, S.; Hao, L.; Zhou, R.; Jin, Y.; Huang, J.; Wu, C. Multispecies biofilms in fermentation: Biofilm for-mation, microbial interactions, and communication. Comprehensive reviews in food science and food safety 2022, 21(4), 3346-3375. [CrossRef]
- Plett, J.M.; Martin, F.M. Know your enemy, embrace your friend: Using omics to understand how plants respond differently to pathogenic and mutualistic microorganisms. The Plant Journal 2018, 93(4), 729-746. [CrossRef]
- Brader, G.; Compant, S.; Vescio, K.; Mitter, B.; Trognitz, F.; Ma, L.J.; Sessitsch, A. Ecology and genomic insights into plant-pathogenic and plant-nonpathogenic endophytes. Annual Review of Phytopathology 2017, 55(1), 61-83. [CrossRef]
- Saini, A.; Mani, I.; Rawal, M.K.; Verma, C.; Singh, V.; Mishra, S.K. An introduction to microbial genomic islands for evolutionary adaptation and pathogenicity. In Microbial Genomic Islands in Adaptation and Pathogenicity; Mani, I., Singh, V., Alzahrani, K.J., Chu, D.T., Eds.; Springer Nature, Singapore, 2023; pp. 1-15. [CrossRef]
- Bailey-Serres, J.; Parker, J. E.; Ainsworth, E. A.; Oldroyd, G. E.; Schroeder, J. I. Genetic strategies for improving crop yields. Nature 2019, 575(7781), 109-118. [CrossRef]
- Poudel, M.; Mendes, R.; Costa, L.A.; Bueno, C.G.; Meng, Y.; Folimonova, S.Y.; Garrett, K.A.; Martins, S.J. The role of plant-associated bacteria, fungi, and viruses in drought stress mitigation. Frontiers in microbiology 2021, 12, 743512. [CrossRef]
- Keeling, P.J; Palmer, J.D.; Horizontal gene transfer in eukaryotic evolution. Nature Reviews Genetics 2008 9, 605–618. [CrossRef]
- Kloesges, T.; Popa, O.; Martin, W.; Dagan, T. Networks of gene sharing among 329 proteobacterial genomes reveal differences in lateral gene transfer frequency at different phylogenetic depths. Mol Biol Evol 2011, 28(2), 1057-1074. [CrossRef]
- McDonald, M.C.; Taranto, A.P.; Hill, E.; Schwessinger, B.; Liu, Z.; Simpfendorfer, S.; Milgate, A.; Solomon, P. S.; Transposon-Mediated Horizontal Transfer of the Host-Specific Virulence Protein ToxA between Three Fungal Wheat Pathogens. mBio 2019, 10(5), e01515-19. [CrossRef]
- Wintersdorff, C.J. H.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P. H.M.; Wolffs P.F.G. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front Microbiol 2016, 19(7),173. [CrossRef]
- Walton, J.D. Horizontal gene transfer and the evolution of secondary metabolite gene clusters in fungi: Anhypothesis. Fungal Genet. Biol. 2000, 30(3), 167–171. [CrossRef]
- Friesen, T.; Stukenbrock, E.; Liu, Z.; Meinhardt, S.; Ling, H.; Faris, J. D.; Rasmussen J. B.; Solomon, P. S.; McDonald, B. A.; Oliver, R. P. Emergence of a new disease as a result of interspecific virulence gene transfer. Nat Genet 2006, 38(8), 953-6. [CrossRef]
- Kobayashi, N.; Dang, T.A.; Pham, K.T.M.; Gómez Luciano, L.B.; Van, V.B.; Izumitsu, K.; Shimizu, M.; Ikeda, K.I.; Li, W.H.; Nakayashiki H. Horizontally Transferred DNA in the Genome of the Fungus Pyricularia oryzae is Associated with Repressive Histone Modifications. Mol Biol Evol. 2023, 40(9), msad186. [CrossRef]
- Koonin, E.V.; Makarova, K.S.; Aravind L. Horizontal gene transfer in prokaryotes: Quantification and classification. Ann Rev Microbiol 2001, 55, 709–742. [CrossRef]
- Fitzpatrick, D.A. Horizontal gene transfer in fungi. FEMS Microbiol Lett 2012, 329(1), 1–8. [CrossRef]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiological Research 2016, 184, 13–24. [CrossRef]
- Singh, B.K.; Delgado-Baquerizo, M.; Egidi, E.; Guirado, E.; Leach, J.E.; Liu, H.; Trivedi, P. Climate change impacts on plant pathogens, food security and paths forward. Nat Rev Microbiol 2023, 21(10), 640-656. [CrossRef]
- Chen, L.; Huang, X.; Zhang, F.; Zhao, D.). The effect of climate change on soilborne diseases: Trends, methods, and challenges. Phytopathology, 2020, 110(7), 1120-1131.
- Cubeta, M.A.; Thomas, E.; Dean, R.A. Neotyphodium/Epichloë species endophytes of grasses: Tapping into a rich source of biodiversity. In Advances in endophytic research; Publisher: Springer, New York, USA, 2014; pp. 205-227.
- Maheshwari, M.; Abulreesh, H.H.; Khan, M.S.; Ahmad, I.; Pichtel, J. Horizontal gene transfer in soil and the rhizosphere: Impact on ecological fitness of bacteria. In Agriculturally Important Microbes for Sustainable Agriculture: Volume I: Plant-Soil-Microbe Nexus; Meena, V.S., Mishra, P.K., Bisht, J.K., Pattanayak, A., Eds.; Springer: Singapore, 2017; pp. 111–130. ISBN 978-981-10-5589-8.
- Jang, H.; Gopinath, G.R.; Eshwar, A.; Srikumar, S.; Nguyen, S.; Gangiredla, J.; Patel, I.R.; Finkelstein, S.B.; Negrete, F.; Woo, J.; Lee, Y. The secretion of toxins and other exoproteins of Cronobacter: Role in virulence, adaption, and persistence. Microorganisms 2020, 8(2), 229. [CrossRef]
- Aminov, R.I. Horizontal gene exchange in environmental microbiota. Frontiers in microbiology 2011, 2, 158. [CrossRef]
- Mehrabi, R.; Bahkali, A.H.; Abd-Elsalam, K.A.; Moslem, M.; Ben M’Barek, S.; Gohari, A.M.; Jashni, M.K.; Stergiopoulos, I.; Kema, G.H.; de Wit, P.J. Horizontal gene and chromosome transfer in plant pathogenic fungi affecting host range. FEMS microbiology reviews 2011, 35(3), 542-554. [CrossRef]
- Zia, R.; Nawaz, M.S.; Siddique, M.J.; Hakim, S.; Imran, A. Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiological research 2021, 242, 126626. [CrossRef]
- Nielsen, K.M.; Van Elsas, J.D. Horizontal gene transfer and microevolution in soil. In Modern Soil Microbiology, 3rd Ed., CRC Press, 2019, pp. 105-123.
- Zibo, L.; Yuan, T.; Zhou, L.; Sen, C.; Qu, X.; Lu, P.; Qiyan, F. Impact factors of the accumulation, migration and spread of antibiotic resistance in the environment. Environmental Geochemistry and Health 2021, 43(5), 1741-1758. [CrossRef]
- de Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.M.; Kretzschmar, M.; Lemanceau, P. Soil bacterial networks are less stable under drought than fungal networks. Nature communications 2018, 9(1), 3033. [CrossRef]
- Pritchard, S.G. Soil organisms and global climate change. Plant Pathology 2011, 60(1), 82-99. [CrossRef]
- Ruiz-Gómez, F.J.; Pérez-de-Luque, A.; Navarro-Cerrillo, R.M. The involvement of Phytophthora root rot and drought stress in holm oak decline: From ecophysiology to microbiome influence. Current Forestry Reports, 2019, 5(4), 251-266. [CrossRef]
- Zeilinger, S.; Gupta, V.K.; Dahms, T.E.; Silva, R.N.; Singh, H.B.; Upadhyay, R.S.; Gomes, E.V.; Tsui, C.K.M.; Nayak S.C. Friends or foes? Emerging insights from fungal interactions with plants. FEMS microbiology reviews 2016, 40(2), 182-207. [CrossRef]
Figure 1.
The adaptive strategies of soil and root-associated bacteria vs. fungi during environmental stress (e.g., drought). The part of the images used in this figure are modified from [109,110]. Website accessed in March 2024 in www.wikipedia.org, www.shutterstock.com, https://en.wikipedia.org/wiki/Biofilm, www.shutterstock.com/search/soybean-root and www.voanews.com/a/drought-tightens-its-grip-on-morocco/6700790.html.
Figure 1.
The adaptive strategies of soil and root-associated bacteria vs. fungi during environmental stress (e.g., drought). The part of the images used in this figure are modified from [109,110]. Website accessed in March 2024 in www.wikipedia.org, www.shutterstock.com, https://en.wikipedia.org/wiki/Biofilm, www.shutterstock.com/search/soybean-root and www.voanews.com/a/drought-tightens-its-grip-on-morocco/6700790.html.
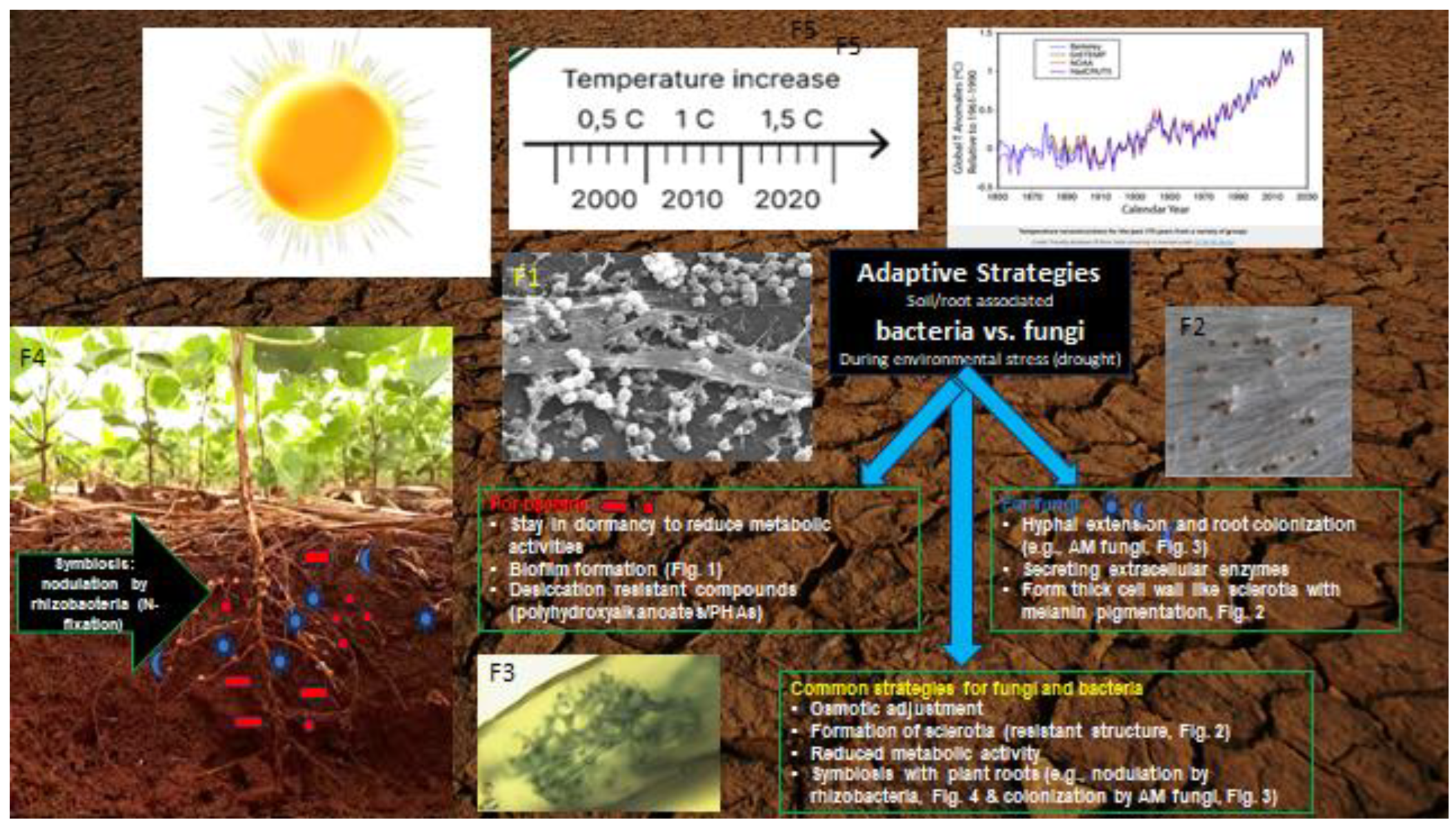
Figure 2.
The adaptive strategies of soil and root-associated pathogen vs. non-pathogen during environmental stress (e.g., drought). Credits for the partial photos and access on March 5, https://niwa.co.nz/natural-hazards/hazards/droughts , https://botanix.com/en/blogs/experts/seedlings-101-follow-the-instructions.
Figure 2.
The adaptive strategies of soil and root-associated pathogen vs. non-pathogen during environmental stress (e.g., drought). Credits for the partial photos and access on March 5, https://niwa.co.nz/natural-hazards/hazards/droughts , https://botanix.com/en/blogs/experts/seedlings-101-follow-the-instructions.

Figure 3.
Horizontal Gene Transfer (HGT) within pathogenic and non-pathogenic/beneficial soil microbes results in the loss or acquisition of virulence or stress tolerance. Photo credit goes to the following websites. References: https://cropwatch.unl.edu/2016/stalk-rot-diseases-including-anthracnose-top-dieback-developing-some-fields,, https://www.nidwater.com/water-conservation-in-agriculture.
Figure 3.
Horizontal Gene Transfer (HGT) within pathogenic and non-pathogenic/beneficial soil microbes results in the loss or acquisition of virulence or stress tolerance. Photo credit goes to the following websites. References: https://cropwatch.unl.edu/2016/stalk-rot-diseases-including-anthracnose-top-dieback-developing-some-fields,, https://www.nidwater.com/water-conservation-in-agriculture.
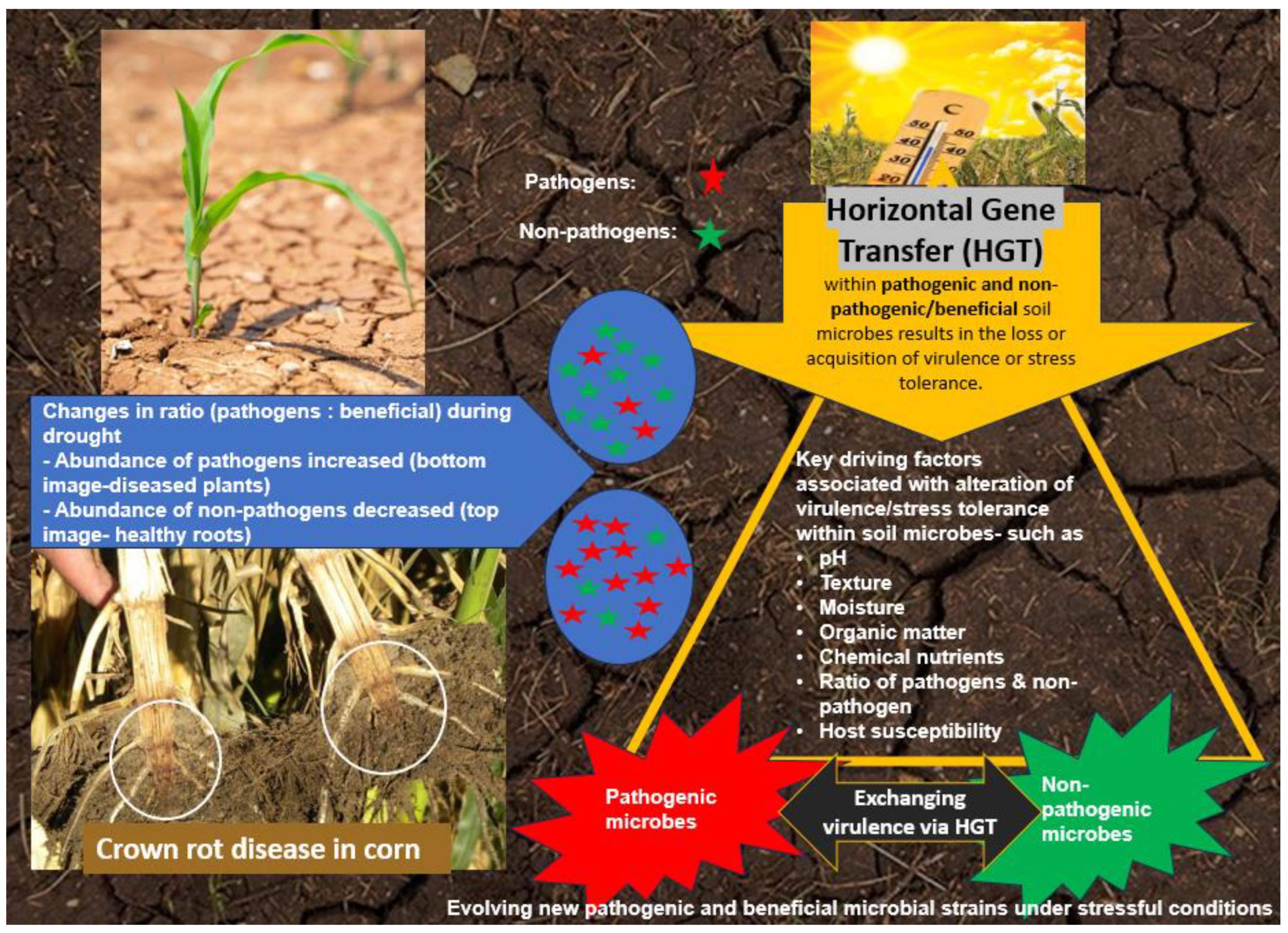
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



