
Submitted:
16 July 2024
Posted:
16 July 2024
You are already at the latest version
Abstract
With the development of social economy, the quality of national life has been rapidly improved, along with the accelerated aging of the population and the continued prevalence of metabolic risk factors. Cardiometabolic diseases (CMDs) have become a major disease burden for both urban and rural residents in China, and most of them are related to lifestyle changes in modern society, especially circadian rhythm disorders. Circadian rhythm regulating the activity of metabolism, such as carbohydrates, lipids, and other nutrients, is the most important pathway modulating the health. Nowadays, studies have shown that the interaction between circadian rhythmicity and gut microbiota counts for the homeostasis of metabolic health, while circadian rhythm disruption is associated with gut microbiota dysbiosis and metabolic diseases. However, there is a lack of research on the topic of gut microbiota mechanisms regulating circadian rhythm disruption leading to CMDs. With increased burden of CMDs prevention and treatment, as well as the demand of healthcare resource allocation, it prompts us to put forward a new strategy to solve this problem. This review focuses on the interactions between circadian rhythm and gut microbiota to elucidate the potential mechanisms of circadian rhythm disruption induced CMDs, providing a solid support for further prevention and treatment.
Keywords:
Cardiometabolic diseases
; Circadian rhythm disruption
; Gut microbiota
1. Introduction
CMDs encompass a range of disorders that initiate with insulin resistance, progress to metabolic syndrome and prediabetes, and ultimately culminate in cardiovascular disease and type 2 diabetes mellitus (T2DM)[1]. CMDs mainly include coronary obesity, diabetes mellitus, hyperlipidemia, heart disease, and non-alcoholic fatty liver disease. They are associated with high mortality and disability rates, posing a serious threat to human health. With the improvement of living standards and the increasing prevalence of unhealthy lifestyles, the prevalence of obesity, disorders of glucose-lipid metabolism, and T2DM has significantly increased, making these cardiometabolic diseases driven by metabolic risk factors more prevalent[2]. As risk factors for cardiovascular disease such as metabolic syndrome, diabetes, and obesity, there is a need to explore better strategies for preventing and modifing the course of these CMDs. This has led to a growing interest in studying the pathogenesis of CMDs.
Circadian rhythms are variations in vital activities that occur on a 24-hour cycle. The circadian system is a hierarchical oscillatory network with the main pacemaker located in the suprachiasmatic nucleus (SCN) of the hypothalamus. This pacemaker coordinates the peripheral organization of the oscillatory network, leading to circadian rhythms in the body from gene expression and cellular metabolism at the microscopic level to macroscopic biological behavior[3,4]. Circadian clocks control many aspects of an organism’s physiology and are influenced by various external stimuli, including environmental factors (e.g., light, food, and temperature), genetic polymorphisms, or behavior[5,6]. The most common causes of circadian disruption in our current society are jet lag, shift work, socialization, light or food at night, inconsistent eating times, and the high-fat diet[7,8,9,10,11,12,13,14,15,16,17]. The primary function of the circadian rhythm clock is to anticipate daily changes in the external environment in order to maintain homeostasis and provide an adaptive physiological response to a changing environment[18,19]. It has been demonstrated that genetic or environmental disorders of the circadian clock can lead to a high incidence of disease or exacerbation of conditions[20,21].
Gut microbiota exhibits circadian rhythmicity in both structural composition and function, influencing brain neurobiochemistry and behavioural phenotypes via the brain-gut axis. The composition and function of the gut microbiota are modulated by a variety of factors including diet, ethnicity, past or current substance use, smoking, and gender[22,23,24,25]. Several human studies have shown that manipulating the gut microbiota can modify the risk of cardiovascular diseases by affecting the production of microbial metabolites [26,27,28,29].
It has been found that disruption of the circadian system alters microbial communities, as well as host metabolism, energy homeostasis, and inflammatory pathways[30]. An in-depth investigation into the mechanisms by which both circadian rhythm and gut microbiota contribute to the development of CMDs is necessary. At the same time, it is important to find out whether the gut microbiota has an impact in the process of circadian rhythm disruption leading to CMDs, in order to define the preventive and therapeutic approaches to these diseases.
Current treatment options for CMDs are costly, ineffective, and associated with numerous adverse effects[31,32]. With cardiovascular disease remaining a leading cause of mortality worldwide, there is an urgent need for a more affordable and effective prevention and treatment strategies. Several studies have indicated a close relationship between biorhythms, gut microbiota, and CMDs development, making them promising therapeutic targets[33]. The contributory role of these two factors in the development of CMDs has recently been realized, thus providing a new therapeutic target for drug development. This review summarizes the potential roles of circadian rhythms, gut microbiota and their metabolites in the development of CMDs[34]. By exploring how these two factors can be utilized as therapeutic strategies, it aims to shed light on potential treatment and prevention avenues for CMDs, as well as the pathophysiological pathways by which the two factors interact in the development of CMDs.
2. Relationship between Circadian Rhythm Disruption and CMDs
Many studies have focused on the interactions between clock genes and metabolism including glucose, lipids, and amino acids/proteins, as well as CMDs[35,36,37,38,39,40]. For example, RORα knockout (KO) mice have shown decreased obesity rates, lower serum HDL cholesterol levels, and reduced serum and liver triglyceride levels [41]. However, these mice were more susceptible to atherosclerosis than wild-type mice, indicating a potential protective role of RORα in atherosclerotic lesions [41].
2.1. Obesity
The development of CMDs is often accompanied by obesity, which is an important contributor to CMDs. One of the biological areas of current interest in obesity research is circadian rhythm disruption [37]. The main causes of obesity are the abnormal accumulation of white adipose tissue due to excessive food intake, physical inactivity and chronic insulin resistance [42]. Studies have shown that circadian rhythm disruption (nocturnal light, sleep disorders, social jet lag, and late-night snacking), whether genetic or acquired environmental factors, impacts host lipid metabolism and accelerates the development of obesity [43,44,45]. For example, Clock mutant mice and Bmal1 KO mice exhibit obese phenotypes: glucose intolerance, reduced insulin secretion, increased sensitivity to high-fat diets, hyperphagia, and weight gain [46,47]. Furthermore, glucose metabolism is impaired in a peripheral tissue-specific Bmal1 KO model [48]. Inducing disruption of central circadian rhythms through modulation of circadian rhythms results in an obese phenotype in rodents [49,50]. These findings in rodents have been largely replicated in humans [51]. Environmental disruptions to circadian rhythms, such as prolonged exposure to artificial light at night, can also lead to disruptions in lipid metabolism and circadian rhythms, ultimately contributing to obesity [52]. Overall, the available date suggest that genetic and/or environmental disruptions to circadian rhythms impair the interaction between circadian rhythms and lipid metabolism, leading to an increased risk of obesity. There is a close relationship exists between circadian rhythms and metabolism, as circadian rhythms regulate metabolic functions; on the other hand, many metabolic functions exhibit circadian rhythmicity, including enzymes (expression and function) important for the regulation of cholesterol, other lipids, glucose and amino acids [53]. Moerover, various hormones (melatonin [54,55,56,57,58,59], leptin[60,61,62,63,64] and glucocorticoids[65,66,67]) and their associated receptors are involved in the regulation of feeding, lipid metabolism and fat accumulation [54,68].
2.2. Diabetics
A study has shown that even just one night of disturbed sleep can impair glucose metabolism in humans [50]. Sleep disruption is a major cause of circadian rhythm disruptions and metabolic dysfunctions. Individuals who fall asleep later and for shorter periods of time are more likely to be obese than those who regularly get adequate sleep [70,71]. Additionally, poor quality and insufficient duration of sleep increases the risk of T2DM [71], which may be the result of the interaction of multiple genetic and environmental risk factors [34,72]. Sleep disruption enhances sympathetic nervous system activity, leading to elevated gluconeogenesis, glucose intolerance, and insulin resistance [73]. An interesting factor that has been shown to play a role in the prevalence of T2DM is genetic and environmental variation in circadian rhythms. For example, T2DM is more common in people who work shifts, such as night and evening shifts [74].
2.3. Atherosclerosis
Atherosclerosis, the underlying pathology of most CMDs, is an inflammatory condition characterized by the accumulation of lipids and leukocytes in the arterial wall [75]. Despite recent advancements in treatment and management, atherosclerosis and its complications remain the leading cause of death worldwide [76,77]. Recent evidence also suggests that the pathogenesis of atherosclerosis progression may be controlled by circadian rhythms. Both systemic factors (hematopoietic and dyslipidemia) and local cellular events (endothelial cell activation, macrophage behavior, inflammation, and vascular remodeling) in atherosclerosis exhibit circadian patterns[78]. (Figure 1)
3. Circadian Rhythm Disruptions Lead to Gut Microbiota Dysbiosis
Genetic variation in the molecular clock mechanism of circadian rhythms has been linked to metabolic disorders in humans[79]. The single nucleotide polymorphisms in Clock and Bmal1 alter an individual’s risk for T2DM, obesity, and dyslipidemia, as well as disrupted sleep/wake patterns [80]. Genetic variation in the molecular clock mechanism of circadian rhythms has been linked between genetic variation and disease risk. Studies have reported that Bmal1 and Per1/2 KO mice have significantly reduced or absent circadian oscillations of gut microbiota, increased susceptibility to intestinal inflammation leading to inflammatory bowel disease, and abnormal microbial circadian rhythms due to disturbances in feeding rhythms, along with glucose intolerance and obesity, as compared to wild-type mice [81]. Oral administration of short-chain fatty acids (SCFA) and lactic acid to mice receiving antibiotics temporarily alters PER2 rhythms [13]. The addition of SCFA such as butyric or acetic acid resulted in an increase in the phase shift and amplitude of PER2 and BMAL1 rhythms [14]. Unconjugated bile acids(BAs) have been shown to both modulate circadian genes in an in vitro cellular model and alter circadian gene expression and the regulators of these genes in the mouse ileum, colon and liver[82].
3.1. Food Intake and Circadian Rhythm
It has been shown that both scheduled and restricted feeding cause dramatic changes in gut microbiota rhythms. Ad libitum-fed Per1/2 KO mice lose the circadian rhythm of gut microbes, but the circadian oscillations of their gut microbes are restored when they are fed at a certain regularity [81]. Normal mice fed freely and regularly have a very significant circadian rhythm of gut microbes, which disappears when changed to a high-fat diet. However, when mice are fed a high-fat diet restrictively, they still maintain some circadian rhythm of gut microbes and a normal body phenotype [83]. When mice fed a high-fat diet were subjected to intermittent fasting, the gut microbiota was altered and the chances of obesity were reduced, a situation that did not occur in germ-free mice [84]. (Figure 2)
3.2. Light and Circadian Rhythm
Many studies manipulate the light-dark cycle to investigate its effect on circadian rhythmicity, using methods such as phase shift, a 24 h light exposure, or a 24 h darkness. Circadian rhythm disruption has been shown to alter the composition of gut microbiome taxa and functional genes [9,68,81,85]. Animals kept under constant 24-hour light or dark conditions, along with regular feeding schedules, exhibited a loss of diurnal rhythmicity in their gut microbiota compared to mice housed under normal light-dark cycles. Mice housed in a 24-hour darkness also showed an increase in abundance of Clostridia [68,85]. Both jet lag and shift work have been associated with a wide variety of CMDs, inflammatory, and stress-related diseases [7,68].
3.3. Shift Work and Circadian Rhythm
Standard-chow-fed mice that underwent a phase-shift paradigm representative of jet lag (8 h shift) incurred a loss of circadian rhythm in their gut microbiota. Furthermore, the jet lag paradigm also exacerbated the effect of a high-fat diet in mice, namely increased weight gain and glucose intolerance. These results were corroborated in a human study during which two subjects underwent 8 to 10 h flights to incur jet lag[81]. Their stool, 24 h post-flight, encompassed a significantly altered microbiota composition characterized by a relative increase in Firmicutes compared to baseline and at recovery 2 weeks later. It is becoming apparent that alterations in host circadian rhythmicity have a profound effect on gut microbiota, potentially leading to a feedback mechanism of gut microbial modulation of further circadian rhythm-dependent activity. This communication pathway requires further investigation at both the preclinical and clinical levels to help elucidate potential therapeutic interventional strategies. (Figure 3)
4. Relationship between Gut Microbiota Dysbiosis and CMDs
Early studies have shown that most gut microbiota exhibit circadian oscillations with the same period of approximately 24 hours as the human central circadian clock[81,86]. Circadian oscillations of the gut microbiota and metabolome further control host circadian rhythms and circadian fluctuations in host physiology and disease[87]. Without gut microbes, germ-free mice fed low or high-fat diets exhibit entirely different central and hepatic circadian clock gene expression and metabolic pathways compared to conventionally raised counterparts[88].
Organisms typically feed during active periods and fast during inactive periods. Meals consumed at different times result in varying levels of glucose at different times. Dinner produces higher peak plasma glucose than breakfast [89,90]. The timing of our meals also affects the microbes active in the gut [89]. Both humans and rodents exhibit diurnal fluctuations in their intestinal microbiota in response to different diets and feeding times. Various bacterial genera and species in the gut, along with the overall microbial community, exhibit oscillatory behavior characterized by population fluctuations with specific patterns [14,15,87]. For example, mice fed a high-fat diet showed a decrease in Lactococcus spp. after the initiation of time-restricted feeding, particularly during inactive periods. Moreover, the bacterial populations of mice (most notably the asexual phylum) peak at 11 p.m. (the active period for nocturnal organisms); the bacterial populations (most notably the thick-walled phylum) reach their lowest values at 7 a.m. (the resting period) [14,15]. The alpha diversity (local species diversity) of the gut microbiota increases with feeding and decreases with fasting [83].
4.1. Gut Microbiota Dysbiosis Leads to CMDs
Gut microbiota dysbiosis has been implicated in the pathogenesis of many diseases, including metabolic disorders such as obesity, diabetes, and CMDs [35,91,92,93,94,95]. CMDs have been associated with changes in the gut microbiota and its metabolites [96,97,98]. In recent years, particular attention has been paid to the interactions between cardiovascular diseases such as atherosclerosis, hypertension and heart failure, and dysbiosis of the gut microbiota[99,100,101,102]. Circadian rhythms of the gut microbiota are primarily driven by circadian signals from the host, diet composition, and food intake [103]. For example, elimination of the host’s molecular clock component (Per1/2 KO) or induction of jet lag leads to abnormal circadian fluctuations in the microbiota, resulting in impaired feeding rhythms, glucose intolerance and obesity [104].
4.2. Pathogenesis of CMDs Caused by Gut Microbiota Dysbiosis
4.2.1. Metabolism-Independent Pathway
Metabolism-independent pathways involve translocation of bacteria and their structural components to the circulatory system in response to inflammatory activation [105]. Gut bacteria can be directly translocated, contributing to the development and progression of CMDs. And gut bacterial genes can be detected in atherosclerotic plaques [106,107,108]. Gut bacteria can also influence the onset and progression of CMDs through immunoinflammatory metabolic pathways. For example, visceral congestion caused by some CMDs (e.g., heart failure) can lead to intestinal edema and leakage of bacterial components (e.g., LPS) into the circulatory, thereby compromising the intestinal barrier [109]. Alternatively, intestinal dysbiosis may increased intestinal permeability leading to the release of bacterial LPS into the circulation, up-regulating the expression of Toll-like receptor 4 (TLR4), which generates downstream signals that promote inflammation and foam cell formation [110]. Several studies have confirmed the association between inflammatory processes and CMDs risk [111,112]. In addition, IL-22 has been used in the treatment of CMDs. studies have found that IL-22 reduces inflammation and endotoxemia, protects the intestinal barrier, enhances insulin sensitivity and endocrine effects, and controls lipid metabolism [113,114,115]. A better understanding of the impact of the gut microbiota in triggering inflammation could provide therapeutic strategies for CMDs [108]. Therefore, identification of the gut microbiota responsible for the immune response leading to CMDs could be a therapeutic target for the prevention and treatment of inflammation-related CMDs. Although this approach is prone to opportunistic infections, immunomodulators have emerged as an optimistic therapy for CMDs.
4.2.2. Metabolism-Dependent Pathway
The gut microbiota can directly influence cardiovascular risk factors and the progression of CMDs through the production of metabolites such as Bas, short-chain fatty acids, trimethylamine-N-oxides and uremic toxins [116,117]. These metabolites enter the circulation, or localize intestinal function and can act either directly or after additional metabolism by the host [101,118,119,120].
SCFAs are products of degradation of undigested dietary fiber by intestinal microbes and are predominantly acetate, propionate, and butyrate in the body [109,121,122]. SCFA may improve intestinal and systemic immune responses and modulate CMDs [123]. The results of several animal studies support the role of SCFA released by the gut microbiota in the regulation of cardiovascular functions and their cardioprotective effects [86]. The number of bacteria responsible for the release of SCFA is significantly reduced in patients suffering from hypertension, heart failure and other CMDs [86,124,125]. In addition, butyric acid prevents obesity and CMDs [126,127].
Choline, phosphatidylcholine, lecithin, and L-carnitine may be interpreted as releasing trimethylamine (TMA) in the presence of microbial-specific TMA lyases in the gut [118,128,129]. Once absorbed in the intestines and transported to the liver, hepatic flavin monooxygenases (FMOs) convert TMA to TMAO[130]. Increased expression of FMO3 in mice affects intestinal cholesterol absorption, reverse cholesterol transport (RCT), and bile acid formation in mice thereby increases the risk of CMDs by increasing plaque formation [131]. FMO3 was found to be up-regulated in obese insulin-resistant mice and resulted in elevated TMAO levels [132]. Two major determinants of circulating TMAO are high TMA-producing gut microbiota composition [168] and dietary intake of TMAO substrates [33,133]. TMAO is a risk factor for cardiac and renal diseases [134]. It has various atherogenic and/or thrombogenic and inflammatory effects in experimental animals [135]. Studies have also shown that TMAO plays a regulatory role in inflammatory responses, oxidative stress, and vascular dysfunction [136]. Studies in germ-free mice and humans have demonstrated a strong association between elevated TMAO levels and an increased risk of CMDs [137,138]. Elevated levels of TMAO are also predictive of a high risk of peripheral arterial disease, coronary artery disease, myocardial infarction (MI), stroke and HF [137,138,139,140,141]. Normal gut microbiota mice fed a high choline diet showed elevated blood TMAO levels, leading to foam cell formation and the development of atherosclerotic plaques [129]. Furthermore, elevated TMAO levels are associated with adverse myocardial fibrosis and ventricular remodeling [33]. (Figure 3)
Figure 4.
Pathogenesis of CMDs Caused by Gut Microbiota Dysbiosis。.
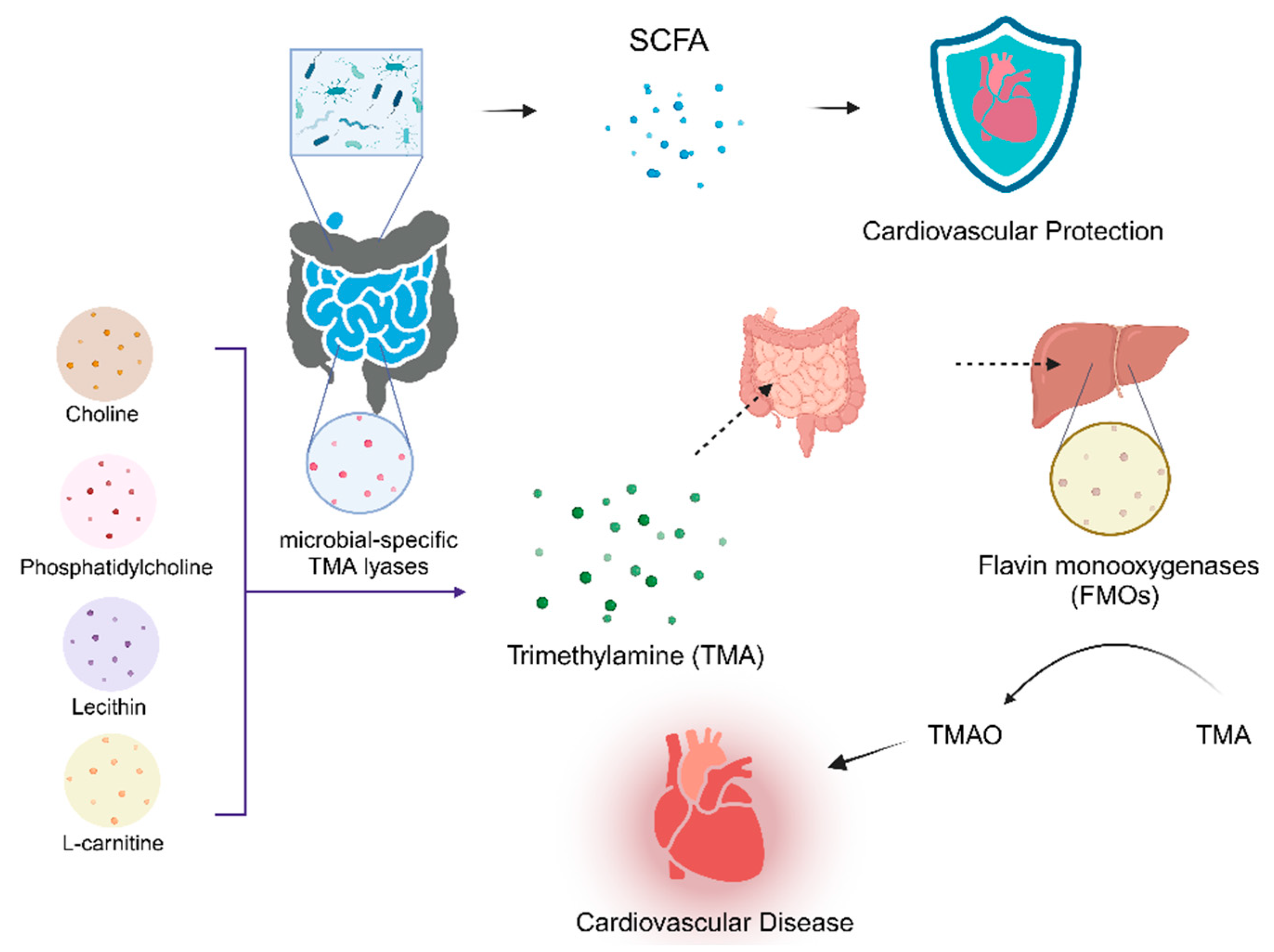
In addition to the previously discussed gut microbiota-associated metabolites, other microbial metabolites with potential modulatory effects on the host are being recognized, such as BAs and uremic toxin [121]. The gut microbiota metabolite BAs affect hepatic cholesterol metabolism and thus modulate the onset and progression of atherosclerosis[142,143]. Uremic toxin stimulates cardiac fibroblasts and collagen synthesis through activation of the p38 mitogen-activated protein kinase (MAPK), p42/44 MAPK, and NF-kB pathways, thereby acting directly on cardiomyocytes and leading to adverse cardiac remodeling [144,145]. Tryptophan (Trp) is an essential amino acid and a precursor of the neurotransmitter serotonin. Metabolites of intestinal microbiota tyrosine and phenylalanine have been shown to be associated with the severity of myocardial infarction in rats [146]. Patients with atherosclerosis have significantly lower levels of tryptophan metabolites produced by circulating microorganisms. [147].
5. Crosstalk between Circadian Rhythms and Gut Microbiota:the Potential Role in CMDs
It has been found that host and microbial biological clocks can co-evolve based on their diet and availability. Disruption of the host’s biological clock can lead to alterations in the microbial biological clock, which in turn affects the composition of the gut microbiota. Animals with mutations in circadian rhythm-related genes often develop diet-related obesity and metabolic syndrome with hypoinsulinemia. Conversely, high-fat diets in turn disrupt normal sleep/wake patterns, indicating a reciprocal regulatory relationship between the circadian clock and organismal metabolism. This bidirectional interaction between the host’s circadian clock and oscillations of gut microbes is crucial for maintaining homeostasis in the host. Therefore, it is imperative that we focus on how this interaction affects key physiological and pathological processes in the host.
A large number of studies have been carried out to explore the effects of gut microbiota and its metabolites on host circadian rhythms from various perspectives[14,82,88]. The subsequent discussion focuses on how alterations in gut microbiota affect the peripheral circadian clock, as well as the influence of microbiota-derived metabolites or diet-induced microbiota changes on the circadian clock. For instance, The intestinal microbiota influences circulatory signaling, enteric and vagal afferent neurons, participates in circadian regulation of food intake and feeding rhythms, and mediates host lipid metabolism [81].
The cardioprotective factor short-chain fatty acids (SCFA), a major end product produced by anaerobic bacteria fermenting dietary fiber or polysaccharides in the colon, is an important manifestation of the mutually beneficial symbiosis between humans and bacterial symbionts [148,149]. SCFA have now been shown to reduce the risk of CMDs. In healthy individuals, intestinal homeostasis and lipid metabolism benefit from an enriched intestinal microbiota produced from SCFA, including acetic, butyric and propionic acids [13,87]. The main SCFA-producing bacteria in the human gut belong to the phylum Thick-walled Bacteria, in particular Faecalibacterium prausnitzii, Roseburia intestinalis, and Eubacterium hallii [150]. SCFA play a protective role in the gut by regulating energy homeostasis, lipid and carbohydrate metabolism, and the composition of the host immune system. host immune system composition to protect cardiometabolism. It has been shown that SCFA-stimulated secretion of glucagon-like peptide 1 (GLP1) from intestinal L-cells regulates food intake and thus controls glucose homeostasis [151].
The main receptors for SCFA are GPR41, GPR43, and GPR109A, which are expressed in the gastrointestinal tract epithelium in colonic epithelial cells, sympathetic ganglia, and adipose tissue lymphocytes [151]. Among them, GPR43 can affect glucose and fatty acid uptake through a signaling mechanism and promote white fat browning, thereby reducing the incidence of obesity [54]. SCFA can directly convert acetic acid or oxidized propionic and butyric acids to acetyl coenzyme A (CoA), which is a substrate for histone acetyltransferase (HAT), thereby affecting host gene expression. It also directly regulates the expression of the clock genes Per2 and Bmal1. Oral administration of short-chain fatty acids (SCFA) to mice had a significant effect on host peripheral tissue rhythms. This suggests a potential mechanism for the interaction of the gut microbiota and the circadian clock in lipid metabolism, and targeting the gut microbiota and its metabolites could be used for the prevention and treatment of obesity and CMDs.
6. CMDs Treatment and Prevention
6.1. Treatment Associated with the Gut Microbiota
The relationship between susceptibility to CMDs and the composition and metabolites of gut microbiota suggests that the gut microbiota may act as a regulator of CMDs, offering a new target for CMDs therapy [108,152]. By understanding the mechanisms involved, it is possible to develop drugs that can inhibit known gut microbial enzymes [31]. Interventions that target the host by altering the gut microbiota have fewer adverse effects on the host compared to interventions that target the host’s metabolism [121]. Therefore, there is an urgent need to explore therapeutic strategies focusing on the gut microbiota for the treatment of CMDs. Current therapeutic approaches targeting the gut microbiota focus on microbiota composition, metabolic pathways, and mucosal barrier protection [86].
Gut microbiota can be used as an in vivo microbial therapy for the treatment of diseases, in addition to being a target for drug therapy [121]. In vivo microbial therapies most commonly involve the use of fecal microbiota transplantation (FMT), probiotics [108]. FMT is the process of restoring normal gut microbiota composition and function by transferring healthy microbiota directly from an individual donor to the gut of a dysbiotic recipient [153,154]. FMT, a pioneering medical procedure, has gained attention in recent years for its potential to impact cardiometabolic disease. The effects of FMT on CMDs have begun to be demonstrated, and future preclinical studies are expected to examine its effectiveness in lipid metabolism, hypertension, and atherosclerosis. Numerous studies support the use of FMT in both experimental and clinical settings in humans and animals (Table 1). FMT has been successfully used for the treatment of CMDs. difficile infections and is now gaining attention in managing CMDs[155]. Because of the generalized gut microbial abundance resulting from this transplantation, the therapeutic application of fecal preparations became an early Chinese practice for the treatment of metabolic diseases [156], highlighting the potential of gut microbiota as a valuable source of beneficial bacteria for CMDs treatment. However, it has been reported that FMT may transfer infectious organisms or endotoxins simultaneously with the transfer of microbiota, resulting in the development of infections and other diseases [157]. To address this issue, transplantion of specific classes of bacteria instead of whole fecal material has been proposed[158]. This innovative approach of transferring healthy gut bacteria offers promising prospects for preventing and managing CMDs.
The utilization of probiotic therapy can alter the assortment of gut microorganisms, boost the production of beneficial microbial byproducts, and decrease levels of trimethylamine oxide and atherogenic amino acids. These changes are believed to contribute to the enhancement of quality of life and the observed anti-inflammatory and lipid-lowering effects. The researchers mentioned that probiotic treatment changed the composition of intestinal microorganisms, increased bioactive microbial metabolites, and reduced TMAO and atherogenic amino acids, which may be the reason for the observed improvement in quality of life and the reported anti-inflammatory and lipid-lowering effects [159]. Probiotics, defined as live microorganisms, can offer health benefits when consumed in appropriate quantities. Numerous animal studies have demonstrated their positive impacts on heart health. Furthermore, research has highlighted the heart-healthy effects of a diet high in fiber. Firstly, increased fiber intake can produce more SCFA through bacterial fermentation, potentially lowering blood pressure [160,161]. Secondly, a fiber rich diet may also lead to a decrease in TMAO levels, which is beneficial for cardiovascular health [162].
6.2. Treatment Associated with Dysfunction of Both Circadian Rhythms and the Gut Microbiota
Stimuli such as light and food can act as signals for the phase of the clock and regulate the circadian system. Gut microbiota-derived metabolites, including SCFAs (propionic acid, butyric acid, acetic acid) and BAs, also alter circadian rhythms. Bacterial metabolites oscillate in composition and content. It has been shown that metabolites produced by germ-free and antibiotic-treated mice do not oscillate diurnally [87]. It has also been reported in the literature that removal of polyamines from the diet of mice leads to abnormalities in the expression of hundreds of circadian genes [81]. This indicates an intricate interaction between diet, gut microbiota, and biological rhythmicity, highlighting the importance of polyamines for regulating circadian gene expression patterns.
Recently, intermittent fasting (IF) has been increasingly recognized as a promising treatment for CMDs. IF involves abstaining from food for a set period of time, usually achieved by restricting food for 12 hours to a few days per week[174]. There are various types of IF, including alternate-day fasting (ADF), fasting-mimicking diet (FMD), and time-restricted feeding (TRF). TRF, considered one of the more extreme approaches, involves regulating or pausing eating every day. The flexibility in fasting frequency and duration allows people to choose their preferred eating pattern, while affecting the gut microbiota and the host’s circadian rhythm. Within the TRF framework, the specific time of day for eating is important. Calorie intake during active periods is often associated with improved cardiometabolic health compared with resting periods, potentially influencing circadian rhythms[175]. Understanding the molecular mechanisms by which host circadian microbiota interactions influence host metabolism and immunity may provide an important dimension to the effective implementation of IF diets used for CMDs therapy.
It has been shown that restricting eating to the active phase in obese mice consuming a high-fat diet results in decreased body weight and fat mass, improved glycemic control, reduced hepatic steatosis, and improved running endurance compared with mice consuming the same number of calories and eating ad libitum[176]. Therefore, it is recommended that calories be consumed during the active phase of the TRF diet. Additionally, clinical trials have shown that eating breakfast may be associated with weight loss, improved glycemic control, and better cardiovascular outcomes in humans[177]. On the other hand, several randomized controlled trials have demonstrated that eating breakfast significantly affects body fat composition or cardiometabolic parameters in humans more than skipping breakfast, even when the total calorie intake remains the same throughout the day[148]. TRF provides a variety of benefits, including improvements in obesity, hypertension, T2DM, and HF[149].
7. Discussion
Circadian rhythms have an important influence on the individual metabolism of the host. And circadian fluctuations in the gut microbiota are also important components in the metabolic process. The interaction between host and gut microbiota is important for maintaining host metabolic homeostasis. The loss of regulation of such rhythms may lead to obesity, metabolic disorders, and CMDs. Therefore, understanding the interaction between host rhythms and gut microbiota is an important advancement in the field of metabolic diseases, offering new insights.. Such interactions have been found to be able to impact on aspects such as immunity and nutrient metabolism.
It is widely recognized that diet is a primary factor in modulating gut microbiota. Consequently, it is not surprising that diet would alter and interact with the gut microbiota, particularly in a cyclic manner due to natural feeding patterns. While much is known about the factors regulating intestinal microbiota, the underlying mechanisms and their effects are not fully understood, prompting further exploration into potential influencing factors. Investigating the circadian rhythm of gut microbiota and its interaction with host metabolism can open up new avenues for studying metabolic diseases. However, understanding the mechanisms underlying changes in the gut microbiota rhythm in the host’s pathological state resulting from dysbiosis remains a direction for future research. Undoubtedly, the study of circadian rhythm of intestinal microbiota is a very important entry point for investigating the pathogenesis of CMDs in both animals and humans.
Funding
This work was supported by the National Natural Science Foundation of China (No 32100905, No 82270467, Key projects of Military Logistics Science Foundation (No BHJ22C028), and National Postdoctoral Science Foundation of China (No 2022M713841).
Data Availability Statement
No new data were created.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Guo, F.; Moellering, D.R.; Garvey, W.T. The Progression of Cardiometabolic Disease: Validation of a New Cardiometabolic Disease Staging System Applicable to Obesity. Obesity (Silver Spring) 2014, 22, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.; Das, S.R.; de Ferranti, S.; Després, J.-P.; et al.; Writing Group Members Heart Disease and Stroke Statistics-2016 Update: A Report From the American Heart Association. Circulation 2016, 133, e38–360. [Google Scholar] [CrossRef] [PubMed]
- Koronowski, K.B.; Kinouchi, K.; Welz, P.-S.; Smith, J.G.; Zinna, V.M.; Shi, J.; Samad, M.; Chen, S.; Magnan, C.N.; Kinchen, J.M.; et al. Defining the Independence of the Liver Circadian Clock. Cell 2019, 177, 1448–1462. [Google Scholar] [CrossRef] [PubMed]
- Patke, A.; Young, M.W.; Axelrod, S. Molecular Mechanisms and Physiological Importance of Circadian Rhythms. Nat Rev Mol Cell Biol 2020, 21, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Feng, M.; Zhang, S.; Dong, Z.; Wang, Y.; Zhang, W.; Liu, C. Angptl8 Mediates Food-Driven Resetting of Hepatic Circadian Clock in Mice. Nat Commun 2019, 10, 3518. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Lamaze, A.; De, J.; Mena, W.; Chélot, E.; Martin, B.; Hardin, P.; Kadener, S.; Emery, P.; Rouyer, F. Reconfiguration of a Multi-Oscillator Network by Light in the Drosophila Circadian Clock. Curr Biol 2018, 28, 2007–2017. [Google Scholar] [CrossRef] [PubMed]
- Inder, M.L.; Crowe, M.T.; Porter, R. Effect of Transmeridian Travel and Jetlag on Mood Disorders: Evidence and Implications. Aust N Z J Psychiatry 2016, 50, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Reid, K.J.; Abbott, S.M. Jet Lag and Shift Work Disorder. Sleep Medicine Clinics 2015, 10, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Schilperoort, M.; van den Berg, R.; Dollé, M.E.T.; van Oostrom, C.T.M.; Wagner, K.; Tambyrajah, L.L.; Wackers, P.; Deboer, T.; Hulsegge, G.; Proper, K.I.; et al. Time-Restricted Feeding Improves Adaptation to Chronically Alternating Light-Dark Cycles. Sci Rep 2019, 9, 7874. [Google Scholar] [CrossRef]
- Roenneberg, T.; Pilz, L.K.; Zerbini, G.; Winnebeck, E.C. Chronotype and Social Jetlag: A (Self-) Critical Review. Biology (Basel) 2019, 8, 54. [Google Scholar] [CrossRef]
- Asher, G.; Sassone-Corsi, P. Time for Food: The Intimate Interplay between Nutrition, Metabolism, and the Circadian Clock. Cell 2015, 161, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Burgess, H.J.; Molina, T.A. Home Lighting before Usual Bedtime Impacts Circadian Timing: A Field Study. Photochem Photobiol 2014, 90, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Tahara, Y.; Yamazaki, M.; Sukigara, H.; Motohashi, H.; Sasaki, H.; Miyakawa, H.; Haraguchi, A.; Ikeda, Y.; Fukuda, S.; Shibata, S. Gut Microbiota-Derived Short Chain Fatty Acids Induce Circadian Clock Entrainment in Mouse Peripheral Tissue. Sci Rep 2018, 8, 1395. [Google Scholar] [CrossRef] [PubMed]
- Leone, V.; Gibbons, S.M.; Martinez, K.; Hutchison, A.L.; Huang, E.Y.; Cham, C.M.; Pierre, J.F.; Heneghan, A.F.; Nadimpalli, A.; Hubert, N.; et al. Effects of Diurnal Variation of Gut Microbes and High-Fat Feeding on Host Circadian Clock Function and Metabolism. Cell Host Microbe 2015, 17, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Bushman, F.D.; FitzGerald, G.A. Rhythmicity of the Intestinal Microbiota Is Regulated by Gender and the Host Circadian Clock. Proc Natl Acad Sci U S A 2015, 112, 10479–10484. [Google Scholar] [CrossRef] [PubMed]
- Voigt, R.M.; Summa, K.C.; Forsyth, C.B.; Green, S.J.; Engen, P.; Naqib, A.; Vitaterna, M.H.; Turek, F.W.; Keshavarzian, A. The Circadian Clock Mutation Promotes Intestinal Dysbiosis. Alcohol Clin Exp Res 2016, 40, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Patke, A.; Murphy, P.J.; Onat, O.E.; Krieger, A.C.; Özçelik, T.; Campbell, S.S.; Young, M.W. Mutation of the Human Circadian Clock Gene CRY1 in Familial Delayed Sleep Phase Disorder. Cell 2017, 169, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Daut, R.A.; Hartsock, M.J.; Tomczik, A.C.; Watkins, L.R.; Spencer, R.L.; Maier, S.F.; Fonken, L.K. Circadian Misalignment Has Differential Effects on Affective Behavior Following Exposure to Controllable or Uncontrollable Stress. Behav Brain Res 2019, 359, 440–445. [Google Scholar] [CrossRef] [PubMed]
- Hor, C.N.; Yeung, J.; Jan, M.; Emmenegger, Y.; Hubbard, J.; Xenarios, I.; Naef, F.; Franken, P. Sleep-Wake-Driven and Circadian Contributions to Daily Rhythms in Gene Expression and Chromatin Accessibility in the Murine Cortex. Proc Natl Acad Sci U S A 2019, 116, 25773–25783. [Google Scholar] [CrossRef]
- Kuehn, B.M. Resetting the Circadian Clock Might Boost Metabolic Health. JAMA 2017, 317, 1303–1305. [Google Scholar] [CrossRef]
- Allen, N.C.; Philip, N.H.; Hui, L.; Zhou, X.; Franklin, R.A.; Kong, Y.; Medzhitov, R. Desynchronization of the Molecular Clock Contributes to the Heterogeneity of the Inflammatory Response. Sci Signal 2019, 12, eaau1851. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Deschasaux, M.; Bouter, K.E.; Prodan, A.; Levin, E.; Groen, A.K.; Herrema, H.; Tremaroli, V.; Bakker, G.J.; Attaye, I.; Pinto-Sietsma, S.-J.; et al. Depicting the Composition of Gut Microbiota in a Population with Varied Ethnic Origins but Shared Geography. Nat Med 2018, 24, 1526–1531. [Google Scholar] [CrossRef] [PubMed]
- Maier, L.; Pruteanu, M.; Kuhn, M.; Zeller, G.; Telzerow, A.; Anderson, E.E.; Brochado, A.R.; Fernandez, K.C.; Dose, H.; Mori, H.; et al. Extensive Impact of Non-Antibiotic Drugs on Human Gut Bacteria. Nature 2018, 555, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Santos-Marcos, J.A.; Haro, C.; Vega-Rojas, A.; Alcala-Diaz, J.F.; Molina-Abril, H.; Leon-Acuña, A.; Lopez-Moreno, J.; Landa, B.B.; Tena-Sempere, M.; Perez-Martinez, P.; et al. Sex Differences in the Gut Microbiota as Potential Determinants of Gender Predisposition to Disease. Mol Nutr Food Res 2019, 63, e1800870. [Google Scholar] [CrossRef]
- Cotillard, A.; Kennedy, S.P.; Kong, L.C.; Prifti, E.; Pons, N.; Le Chatelier, E.; Almeida, M.; Quinquis, B.; Levenez, F.; Galleron, N.; et al. Dietary Intervention Impact on Gut Microbial Gene Richness. Nature 2013, 500, 585–588. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut Bacteria Selectively Promoted by Dietary Fibers Alleviate Type 2 Diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Quercia, S.; Turroni, S.; Fiori, J.; Soverini, M.; Rampelli, S.; Biagi, E.; Castagnetti, A.; Consolandi, C.; Severgnini, M.; Pianesi, M.; et al. Gut Microbiome Response to Short-Term Dietary Interventions in Reactive Hypoglycemia Subjects. Diabetes Metab Res Rev 2017, 33. [Google Scholar] [CrossRef]
- Erickson, M.L.; Malin, S.K.; Wang, Z.; Brown, J.M.; Hazen, S.L.; Kirwan, J.P. Effects of Lifestyle Intervention on Plasma Trimethylamine N-Oxide in Obese Adults. Nutrients 2019, 11, 179. [Google Scholar] [CrossRef]
- Moreira, G.C.; Cipullo, J.P.; Ciorlia, L.A.S.; Cesarino, C.B.; Vilela-Martin, J.F. Prevalence of Metabolic Syndrome: Association with Risk Factors and Cardiovascular Complications in an Urban Population. PLoS One 2014, 9, e105056. [Google Scholar] [CrossRef]
- Tuteja, S.; Ferguson, J.F. Gut Microbiome and Response to Cardiovascular Drugs. Circ Genom Precis Med 2019, 12, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Schuster, H. Improving Lipid Management--to Titrate, Combine or Switch. Int J Clin Pract 2004, 58, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Li, H.; Zhou, H.; Zhang, X.; Zhang, A.; Xie, Y.; Li, Y.; Lv, S.; Zhang, J. Role and Effective Therapeutic Target of Gut Microbiota in Heart Failure. Cardiovasc Ther 2019, 2019, 5164298. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Sun, B.; Yu, D.; Zhu, C. Gut Microbiota: An Important Player in Type 2 Diabetes Mellitus. Front Cell Infect Microbiol 2022, 12, 834485. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Jia, H.; Feng, Q.; Wang, D.; Liang, D.; Wu, X.; Li, J.; Tang, L.; Li, Y.; et al. The Oral and Gut Microbiomes Are Perturbed in Rheumatoid Arthritis and Partly Normalized after Treatment. Nat Med 2015, 21, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Z.; Wang, Y.; Li, Y.; Ye, C.; Ruhn, K.A.; Behrendt, C.L.; Olson, E.N.; Hooper, L.V. The Intestinal Microbiota Programs Diurnal Rhythms in Host Metabolism through Histone Deacetylase 3. Science 2019, 365, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Adamovich, Y.; Rousso-Noori, L.; Zwighaft, Z.; Neufeld-Cohen, A.; Golik, M.; Kraut-Cohen, J.; Wang, M.; Han, X.; Asher, G. Circadian Clocks and Feeding Time Regulate the Oscillations and Levels of Hepatic Triglycerides. Cell Metab 2014, 19, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Wang, J.; Klein, P.S.; Lazar, M.A. Nuclear Receptor Rev-Erbalpha Is a Critical Lithium-Sensitive Component of the Circadian Clock. Science 2006, 311, 1002–1005. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Chen, J.; Wang, J.; Yao, J.; Huang, Y.; Zhang, G.; Bao, Z. Circadian Clock Genes in the Metabolism of Non-Alcoholic Fatty Liver Disease. Front Physiol 2019, 10, 423. [Google Scholar] [CrossRef]
- Dang, F.; Sun, X.; Ma, X.; Wu, R.; Zhang, D.; Chen, Y.; Xu, Q.; Wu, Y.; Liu, Y. Insulin Post-Transcriptionally Modulates Bmal1 Protein to Affect the Hepatic Circadian Clock. Nat Commun 2016, 7, 12696. [Google Scholar] [CrossRef]
- Lau, P.; Fitzsimmons, R.L.; Raichur, S.; Wang, S.-C.M.; Lechtken, A.; Muscat, G.E.O. The Orphan Nuclear Receptor, RORalpha, Regulates Gene Expression That Controls Lipid Metabolism: Staggerer (SG/SG) Mice Are Resistant to Diet-Induced Obesity. J Biol Chem 2008, 283, 18411–18421. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Huang, X.; Song, B.-L.; Yang, H. The Biogenesis of Lipid Droplets: Lipids Take Center Stage. Prog Lipid Res 2019, 75, 100989. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, J.E.; Livelo, C.; Trujillo, A.S.; Chandran, S.; Woodworth, B.; Andrade, L.; Le, H.D.; Manor, U.; Panda, S.; Melkani, G.C. Time-Restricted Feeding Restores Muscle Function in Drosophila Models of Obesity and Circadian-Rhythm Disruption. Nat Commun 2019, 10, 2700. [Google Scholar] [CrossRef] [PubMed]
- Schäbler, S.; Amatobi, K.M.; Horn, M.; Rieger, D.; Helfrich-Förster, C.; Mueller, M.J.; Wegener, C.; Fekete, A. Loss of Function in the Drosophila Clock Gene Period Results in Altered Intermediary Lipid Metabolism and Increased Susceptibility to Starvation. Cell Mol Life Sci 2020, 77, 4939–4956. [Google Scholar] [CrossRef] [PubMed]
- Baron, K.G.; Reid, K.J.; Kim, T.; Van Horn, L.; Attarian, H.; Wolfe, L.; Siddique, J.; Santostasi, G.; Zee, P.C. Circadian Timing and Alignment in Healthy Adults: Associations with BMI, Body Fat, Caloric Intake and Physical Activity. Int J Obes (Lond) 2017, 41, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Kovac, J.; Kolbe, I.; Ehrhardt, L.; Leliavski, A.; Husse, J.; Salinas, G.; Lingner, T.; Tsang, A.H.; Barclay, J.L.; Oster, H. Hepatic Gene Therapy Rescues High-Fat Diet Responses in Circadian Clock Mutant Mice. Mol Metab 2017, 6, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Marcheva, B.; Ramsey, K.M.; Buhr, E.D.; Kobayashi, Y.; Su, H.; Ko, C.H.; Ivanova, G.; Omura, C.; Mo, S.; Vitaterna, M.H.; et al. Disruption of the Clock Components CLOCK and BMAL1 Leads to Hypoinsulinaemia and Diabetes. Nature 2010, 466, 627–631. [Google Scholar] [CrossRef]
- Perelis, M.; Marcheva, B.; Ramsey, K.M.; Schipma, M.J.; Hutchison, A.L.; Taguchi, A.; Peek, C.B.; Hong, H.; Huang, W.; Omura, C.; et al. Pancreatic β Cell Enhancers Regulate Rhythmic Transcription of Genes Controlling Insulin Secretion. Science 2015, 350, aac4250. [Google Scholar] [CrossRef]
- Qian, J.; Block, G.D.; Colwell, C.S.; Matveyenko, A.V. Consequences of Exposure to Light at Night on the Pancreatic Islet Circadian Clock and Function in Rats. Diabetes 2013, 62, 3469–3478. [Google Scholar] [CrossRef]
- Gale, J.E.; Cox, H.I.; Qian, J.; Block, G.D.; Colwell, C.S.; Matveyenko, A.V. Disruption of Circadian Rhythms Accelerates Development of Diabetes through Pancreatic Beta-Cell Loss and Dysfunction. J Biol Rhythms 2011, 26, 423–433. [Google Scholar] [CrossRef]
- Tahira, K.; Ueno, T.; Fukuda, N.; Aoyama, T.; Tsunemi, A.; Matsumoto, S.; Nagura, C.; Matsumoto, T.; Soma, M.; Shimba, S.; et al. Obesity Alters the Expression Profile of Clock Genes in Peripheral Blood Mononuclear Cells. Arch Med Sci 2011, 7, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Touitou, Y.; Reinberg, A.; Touitou, D. Association between Light at Night, Melatonin Secretion, Sleep Deprivation, and the Internal Clock: Health Impacts and Mechanisms of Circadian Disruption. Life Sci 2017, 173, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Panda, S. Circadian Physiology of Metabolism. Science 2016, 354, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Li, Y.; Han, H.; Chen, S.; Gao, J.; Liu, G.; Wu, X.; Deng, J.; Yu, Q.; Huang, X.; et al. Melatonin Reprogramming of Gut Microbiota Improves Lipid Dysmetabolism in High-Fat Diet-Fed Mice. J Pineal Res 2018, 65, e12524. [Google Scholar] [CrossRef] [PubMed]
- Cipolla-Neto, J.; Amaral, F.G. do Melatonin as a Hormone: New Physiological and Clinical Insights. Endocr Rev 2018, 39, 990–1028. [Google Scholar] [CrossRef] [PubMed]
- Bazwinsky-Wutschke, I.; Wolgast, S.; Mühlbauer, E.; Albrecht, E.; Peschke, E. Phosphorylation of Cyclic AMP-Response Element-Binding Protein (CREB) Is Influenced by Melatonin Treatment in Pancreatic Rat Insulinoma β-Cells (INS-1). J Pineal Res 2012, 53, 344–357. [Google Scholar] [CrossRef] [PubMed]
- Faria, J.A.; Kinote, A.; Ignacio-Souza, L.M.; de Araújo, T.M.; Razolli, D.S.; Doneda, D.L.; Paschoal, L.B.; Lellis-Santos, C.; Bertolini, G.L.; Velloso, L.A.; et al. Melatonin Acts through MT1/MT2 Receptors to Activate Hypothalamic Akt and Suppress Hepatic Gluconeogenesis in Rats. Am J Physiol Endocrinol Metab 2013, 305, E230–E242. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Li, Y.; Han, H.; Ma, J.; Liu, G.; Wu, X.; Huang, X.; Fang, R.; Baba, K.; Bin, P.; et al. Administration of Exogenous Melatonin Improves the Diurnal Rhythms of the Gut Microbiota in Mice Fed a High-Fat Diet. mSystems 2020, 5, e00002-20. [Google Scholar] [CrossRef] [PubMed]
- Ríos-Lugo, M.J.; Cano, P.; Jiménez-Ortega, V.; Fernández-Mateos, M.P.; Scacchi, P.A.; Cardinali, D.P.; Esquifino, A.I. Melatonin Effect on Plasma Adiponectin, Leptin, Insulin, Glucose, Triglycerides and Cholesterol in Normal and High Fat-Fed Rats. Journal of pineal research 2010, 49. [Google Scholar] [CrossRef]
- Downs, J.L.; Urbanski, H.F. Aging-Related Sex-Dependent Loss of the Circulating Leptin 24-h Rhythm in the Rhesus Monkey. J Endocrinol 2006, 190, 117–127. [Google Scholar] [CrossRef]
- Kettner, N.M.; Mayo, S.A.; Hua, J.; Lee, C.; Moore, D.D.; Fu, L. Circadian Dysfunction Induces Leptin Resistance in Mice. Cell Metab 2015, 22, 448–459. [Google Scholar] [CrossRef]
- Hackl, M.T.; Fürnsinn, C.; Schuh, C.M.; Krssak, M.; Carli, F.; Guerra, S.; Freudenthaler, A.; Baumgartner-Parzer, S.; Helbich, T.H.; Luger, A.; et al. Brain Leptin Reduces Liver Lipids by Increasing Hepatic Triglyceride Secretion and Lowering Lipogenesis. Nat Commun 2019, 10, 2717. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhu, Y.; Schultz, R.D.; Li, N.; He, Z.; Zhang, Z.; Caron, A.; Zhu, Q.; Sun, K.; Xiong, W.; et al. Partial Leptin Reduction as an Insulin Sensitization and Weight Loss Strategy. Cell Metab 2019, 30, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Ravussin, Y.; Edwin, E.; Gallop, M.; Xu, L.; Bartolomé, A.; Kraakman, M.J.; LeDuc, C.A.; Ferrante, A.W. Evidence for a Non-Leptin System That Defends against Weight Gain in Overfeeding. Cell Metab 2018, 28, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Caratti, G.; Iqbal, M.; Hunter, L.; Kim, D.; Wang, P.; Vonslow, R.M.; Begley, N.; Tetley, A.J.; Woodburn, J.L.; Pariollaud, M.; et al. REVERBa Couples the Circadian Clock to Hepatic Glucocorticoid Action. J Clin Invest 2018, 128, 4454–4471. [Google Scholar] [CrossRef]
- Shimba, A.; Cui, G.; Tani-Ichi, S.; Ogawa, M.; Abe, S.; Okazaki, F.; Kitano, S.; Miyachi, H.; Yamada, H.; Hara, T.; et al. Glucocorticoids Drive Diurnal Oscillations in T Cell Distribution and Responses by Inducing Interleukin-7 Receptor and CXCR4. Immunity 2018, 48, 286–298. [Google Scholar] [CrossRef]
- Quagliarini, F.; Mir, A.A.; Balazs, K.; Wierer, M.; Dyar, K.A.; Jouffe, C.; Makris, K.; Hawe, J.; Heinig, M.; Filipp, F.V.; et al. Cistromic Reprogramming of the Diurnal Glucocorticoid Hormone Response by High-Fat Diet. Mol Cell 2019, 76, 531–545. [Google Scholar] [CrossRef]
- Wu, T.; Yang, L.; Jiang, J.; Ni, Y.; Zhu, J.; Zheng, X.; Wang, Q.; Lu, X.; Fu, Z. Chronic Glucocorticoid Treatment Induced Circadian Clock Disorder Leads to Lipid Metabolism and Gut Microbiota Alterations in Rats. Life Sci 2018, 192, 173–182. [Google Scholar] [CrossRef]
- van den Berg, R.; Mook-Kanamori, D.O.; Donga, E.; van Dijk, M.; van Dijk, J.G.; Lammers, G.-J.; van Kralingen, K.W.; Prehn, C.; Adamski, J.; Romijn, J.A.; et al. A Single Night of Sleep Curtailment Increases Plasma Acylcarnitines: Novel Insights in the Relationship between Sleep and Insulin Resistance. Arch Biochem Biophys 2016, 589, 145–151. [Google Scholar] [CrossRef]
- Brum, M.C.B.; Dantas Filho, F.F.; Schnorr, C.C.; Bertoletti, O.A.; Bottega, G.B.; da Costa Rodrigues, T. Night Shift Work, Short Sleep and Obesity. Diabetol Metab Syndr 2020, 12, 13. [Google Scholar] [CrossRef]
- Pugliese, G.; Barrea, L.; Laudisio, D.; Salzano, C.; Aprano, S.; Colao, A.; Savastano, S.; Muscogiuri, G. Sleep Apnea, Obesity, and Disturbed Glucose Homeostasis: Epidemiologic Evidence, Biologic Insights, and Therapeutic Strategies. Curr Obes Rep 2020, 9, 30–38. [Google Scholar] [CrossRef]
- Ling, C.; Bacos, K.; Rönn, T. Epigenetics of Type 2 Diabetes Mellitus and Weight Change - a Tool for Precision Medicine? Nat Rev Endocrinol 2022, 18, 433–448. [Google Scholar] [CrossRef]
- Mullington, J.M.; Cunningham, T.J.; Haack, M.; Yang, H. Causes and Consequences of Chronic Sleep Deficiency and the Role of Orexin. Front Neurol Neurosci 2021, 45, 128–138. [Google Scholar] [CrossRef]
- Kalsbeek, A.; la Fleur, S.; Fliers, E. Circadian Control of Glucose Metabolism. Mol Metab 2014, 3, 372–383. [Google Scholar] [CrossRef]
- Swirski, F.K.; Nahrendorf, M. Leukocyte Behavior in Atherosclerosis, Myocardial Infarction, and Heart Failure. Science 2013, 339, 161–166. [Google Scholar] [CrossRef]
- Englund, A.; Kovanen, L.; Saarikoski, S.T.; Haukka, J.; Reunanen, A.; Aromaa, A.; Lönnqvist, J.; Partonen, T. NPAS2 and PER2 Are Linked to Risk Factors of the Metabolic Syndrome. J Circadian Rhythms 2009, 7, 5. [Google Scholar] [CrossRef]
- Yang, Y.-D.; Zeng, Y.; Li, J.; Zhou, J.-H.; He, Q.-Y.; Zheng, C.-J.; Reichetzeder, C.; Krämer, B.K.; Hocher, B. Association of BMAL1 Clock Gene Polymorphisms with Fasting Glucose in Children. Pediatr Res 2023, 94, 653–659. [Google Scholar] [CrossRef]
- McAlpine, C.S.; Swirski, F.K. Circadian Influence on Metabolism and Inflammation in Atherosclerosis. Circulation Research 2016, 119, 131–141. [Google Scholar] [CrossRef]
- Corella, D.; Asensio, E.M.; Coltell, O.; Sorlí, J.V.; Estruch, R.; Martínez-González, M.Á.; Salas-Salvadó, J.; Castañer, O.; Arós, F.; Lapetra, J.; et al. CLOCK Gene Variation Is Associated with Incidence of Type-2 Diabetes and Cardiovascular Diseases in Type-2 Diabetic Subjects: Dietary Modulation in the PREDIMED Randomized Trial. Cardiovasc Diabetol 2016, 15, 4. [Google Scholar] [CrossRef]
- Dupuis, J.; Langenberg, C.; Prokopenko, I.; Saxena, R.; Soranzo, N.; Jackson, A.U.; Wheeler, E.; Glazer, N.L.; Bouatia-Naji, N.; Gloyn, A.L.; et al. New Genetic Loci Implicated in Fasting Glucose Homeostasis and Their Impact on Type 2 Diabetes Risk. Nat Genet 2010, 42, 105–116. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Zeevi, D.; Levy, M.; Zilberman-Schapira, G.; Suez, J.; Tengeler, A.C.; Abramson, L.; Katz, M.N.; Korem, T.; Zmora, N.; et al. Transkingdom Control of Microbiota Diurnal Oscillations Promotes Metabolic Homeostasis. Cell 2014, 159, 514–529. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, K.; MacSharry, J.; Casey, P.G.; Shanahan, F.; Joyce, S.A.; Gahan, C.G.M. Unconjugated Bile Acids Influence Expression of Circadian Genes: A Potential Mechanism for Microbe-Host Crosstalk. PLoS One 2016, 11, e0167319. [Google Scholar] [CrossRef] [PubMed]
- Zarrinpar, A.; Chaix, A.; Yooseph, S.; Panda, S. Diet and Feeding Pattern Affect the Diurnal Dynamics of the Gut Microbiome. Cell Metab 2014, 20, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Xie, C.; Lu, S.; Nichols, R.G.; Tian, Y.; Li, L.; Patel, D.; Ma, Y.; Brocker, C.N.; Yan, T.; et al. Intermittent Fasting Promotes White Adipose Browning and Decreases Obesity by Shaping the Gut Microbiota. Cell Metab 2017, 26, 672–685. [Google Scholar] [CrossRef]
- Deaver, J.A.; Eum, S.Y.; Toborek, M. Circadian Disruption Changes Gut Microbiome Taxa and Functional Gene Composition. Front Microbiol 2018, 9, 737. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, H.-Y.; Hu, X.-M.; Zhang, Y.; Zhang, S.-Y. Current Understanding of Gut Microbiota Alterations and Related Therapeutic Intervention Strategies in Heart Failure. Chin Med J (Engl) 2019, 132, 1843–1855. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Levy, M.; Korem, T.; Dohnalová, L.; Shapiro, H.; Jaitin, D.A.; David, E.; Winter, D.R.; Gury-BenAri, M.; Tatirovsky, E.; et al. Microbiota Diurnal Rhythmicity Programs Host Transcriptome Oscillations. Cell 2016, 167, 1495–1510. [Google Scholar] [CrossRef] [PubMed]
- Montagner, A.; Korecka, A.; Polizzi, A.; Lippi, Y.; Blum, Y.; Canlet, C.; Tremblay-Franco, M.; Gautier-Stein, A.; Burcelin, R.; Yen, Y.-C.; et al. Hepatic Circadian Clock Oscillators and Nuclear Receptors Integrate Microbiome-Derived Signals. Sci Rep 2016, 6, 20127. [Google Scholar] [CrossRef] [PubMed]
- Parkar, S.G.; Kalsbeek, A.; Cheeseman, J.F. Potential Role for the Gut Microbiota in Modulating Host Circadian Rhythms and Metabolic Health. Microorganisms 2019, 7, 41. [Google Scholar] [CrossRef]
- Van Cauter, E.; Polonsky, K.S.; Scheen, A.J. Roles of Circadian Rhythmicity and Sleep in Human Glucose Regulation. Endocr Rev 1997, 18, 716–738. [Google Scholar] [CrossRef]
- Chu, H.; Khosravi, A.; Kusumawardhani, I.P.; Kwon, A.H.K.; Vasconcelos, A.C.; Cunha, L.D.; Mayer, A.E.; Shen, Y.; Wu, W.-L.; Kambal, A.; et al. Gene-Microbiota Interactions Contribute to the Pathogenesis of Inflammatory Bowel Disease. Science 2016, 352, 1116–1120. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.W.; Adams, J.B.; Gregory, A.C.; Borody, T.; Chittick, L.; Fasano, A.; Khoruts, A.; Geis, E.; Maldonado, J.; McDonough-Means, S.; Pollard, E.L. Microbiota Transfer Therapy Alters Gut Ecosystem and Improves Gastrointestinal and Autism Symptoms: An Open-Label Study. Microbiome 2017, 5. [Google Scholar] [CrossRef]
- Tremaroli, V.; Bäckhed, F. Functional Interactions between the Gut Microbiota and Host Metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of Gut Microbiomes in Nonalcoholic Steatohepatitis (NASH) Patients: A Connection between Endogenous Alcohol and NASH. Hepatology 2013, 57, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Caro, F.; Robins, W.; Mekalanos, J.J. Antagonism toward the Intestinal Microbiota and Its Effect on Vibrio Cholerae Virulence. Science 2018, 359, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Liao, S.X.; He, Y.; Wang, S.; Xia, G.H.; Liu, F.T.; Zhu, J.J.; You, C.; Chen, Q.; Zhou, L.; Pan, S.Y. Dysbiosis of Gut Microbiota With Reduced Trimethylamine-N-Oxide Level in Patients With Large-Artery Atherosclerotic Stroke or Transient Ischemic Attack. Journal of the American Heart Association 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Karbach, S.H.; Schönfelder, T.; Brandão, I.; Wilms, E.; Hörmann, N.; Jäckel, S.; Schüler, R.; Finger, S.; Knorr, M.; Lagrange, J.; et al. Gut Microbiota Promote Angiotensin II-Induced Arterial Hypertension and Vascular Dysfunction. J Am Heart Assoc 2016, 5, e003698. [Google Scholar] [CrossRef]
- Kim, S.; Goel, R.; Kumar, A.; Qi, Y.; Lobaton, G.; Hosaka, K.; Mohammed, M.; Handberg, E.M.; Richards, E.M.; Pepine, C.J.; et al. Imbalance of Gut Microbiome and Intestinal Epithelial Barrier Dysfunction in Patients with High Blood Pressure. Clin Sci (Lond) 2018, 132, 701–718. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Ye, L.; Li, J.; Jin, L.; Wang, W.; Li, S.; Bao, M.; Wu, S.; Li, L.; Geng, B.; et al. Metagenomic and Metabolomic Analyses Unveil Dysbiosis of Gut Microbiota in Chronic Heart Failure Patients. Sci Rep 2018, 8, 635. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Dai, Z.; Wang, Z.; Deng, Z.; Zhang, J.; Pu, J.; Cao, W.; Pan, T.; Zhou, Y.; Yang, Z.; et al. Gut Microbiota Dysbiosis Contributes to the Development of Chronic Obstructive Pulmonary Disease. Respir Res 2021, 22, 274. [Google Scholar] [CrossRef]
- Zhu, W.; Gregory, J.C.; Org, E.; Buffa, J.A.; Gupta, N.; Wang, Z.; Li, L.; Fu, X.; Wu, Y.; Mehrabian, M.; et al. Gut Microbial Metabolite TMAO Enhances Platelet Hyperreactivity and Thrombosis Risk. Cell 2016, 165, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Roberts, A.B.; Buffa, J.A.; Levison, B.S.; Zhu, W.; Org, E.; Gu, X.; Huang, Y.; Zamanian-Daryoush, M.; Culley, M.K.; et al. Non-Lethal Inhibition of Gut Microbial Trimethylamine Production for the Treatment of Atherosclerosis. Cell 2015, 163, 1585–1595. [Google Scholar] [CrossRef]
- Wang, Y.; Kuang, Z.; Yu, X.; Ruhn, K.A.; Kubo, M.; Hooper, L.V. The Intestinal Microbiota Regulates Body Composition through NFIL3 and the Circadian Clock. Science 2017, 357, 912–916. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The Gut Microbiota as an Environmental Factor That Regulates Fat Storage. Proc Natl Acad Sci U S A 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed]
- Sandek, A.; Bauditz, J.; Swidsinski, A.; Buhner, S.; Weber-Eibel, J.; von Haehling, S.; Schroedl, W.; Karhausen, T.; Doehner, W.; Rauchhaus, M.; et al. Altered Intestinal Function in Patients with Chronic Heart Failure. J Am Coll Cardiol 2007, 50, 1561–1569. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, A.L.; Bäckhed, F. Role of Gut Microbiota in Atherosclerosis. Nat Rev Cardiol 2017, 14, 79–87. [Google Scholar] [CrossRef]
- Mitra, S.; Drautz-Moses, D.I.; Alhede, M.; Maw, M.T.; Liu, Y.; Purbojati, R.W.; Yap, Z.H.; Kushwaha, K.K.; Gheorghe, A.G.; Bjarnsholt, T.; et al. In Silico Analyses of Metagenomes from Human Atherosclerotic Plaque Samples. Microbiome 2015, 3, 38. [Google Scholar] [CrossRef]
- Xu, H.; Wang, X.; Feng, W.; Liu, Q.; Zhou, S.; Liu, Q.; Cai, L. The Gut Microbiota and Its Interactions with Cardiovascular Disease. Microb Biotechnol 2020, 13, 637–656. [Google Scholar] [CrossRef]
- Tang, W.H.W.; Bäckhed, F.; Landmesser, U.; Hazen, S.L. Intestinal Microbiota in Cardiovascular Health and Disease: JACC State-of-the-Art Review. J Am Coll Cardiol 2019, 73, 2089–2105. [Google Scholar] [CrossRef]
- Lau, K.; Srivatsav, V.; Rizwan, A.; Nashed, A.; Liu, R.; Shen, R.; Akhtar, M. Bridging the Gap between Gut Microbial Dysbiosis and Cardiovascular Diseases. Nutrients 2017, 9, 859. [Google Scholar] [CrossRef]
- Geovanini, G.R.; Libby, P. Atherosclerosis and Inflammation: Overview and Updates. Clin Sci (Lond) 2018, 132, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Lawler, P.R.; Bhatt, D.L.; Godoy, L.C.; Lüscher, T.F.; Bonow, R.O.; Verma, S.; Ridker, P.M. Targeting Cardiovascular Inflammation: Next Steps in Clinical Translation. Eur Heart J 2021, 42, 113–131. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, K.; Ogawa, A.; Mizoguchi, E.; Shimomura, Y.; Andoh, A.; Bhan, A.K.; Blumberg, R.S.; Xavier, R.J.; Mizoguchi, A. IL-22 Ameliorates Intestinal Inflammation in a Mouse Model of Ulcerative Colitis. J Clin Invest 2008, 118, 534–544. [Google Scholar] [CrossRef]
- Wang, X.; Ota, N.; Manzanillo, P.; Kates, L.; Zavala-Solorio, J.; Eidenschenk, C.; Zhang, J.; Lesch, J.; Lee, W.P.; Ross, J.; et al. Interleukin-22 Alleviates Metabolic Disorders and Restores Mucosal Immunity in Diabetes. Nature 2014, 514, 237–241. [Google Scholar] [CrossRef]
- Zheng, Y.; Valdez, P.A.; Danilenko, D.M.; Hu, Y.; Sa, S.M.; Gong, Q.; Abbas, A.R.; Modrusan, Z.; Ghilardi, N.; de Sauvage, F.J.; et al. Interleukin-22 Mediates Early Host Defense against Attaching and Effacing Bacterial Pathogens. Nat Med 2008, 14, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Avery, E.G.; Bartolomaeus, H.; Maifeld, A.; Marko, L.; Wiig, H.; Wilck, N.; Rosshart, S.P.; Forslund, S.K.; Müller, D.N. The Gut Microbiome in Hypertension: Recent Advances and Future Perspectives. Circ Res 2021, 128, 934–950. [Google Scholar] [CrossRef]
- Brown, J.M.; Hazen, S.L. The Gut Microbial Endocrine Organ: Bacterially Derived Signals Driving Cardiometabolic Diseases. Annu Rev Med 2015, 66, 343–359. [Google Scholar] [CrossRef]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal Microbiota Metabolism of L-Carnitine, a Nutrient in Red Meat, Promotes Atherosclerosis. Nat Med 2013, 19, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Seldin, M.M.; Meng, Y.; Qi, H.; Zhu, W.; Wang, Z.; Hazen, S.L.; Lusis, A.J.; Shih, D.M. Trimethylamine N-Oxide Promotes Vascular Inflammation Through Signaling of Mitogen-Activated Protein Kinase and Nuclear Factor-κB. J Am Heart Assoc 2016, 5, e002767. [Google Scholar] [CrossRef]
- Tang, W.H.W.; Kitai, T.; Hazen, S.L. Gut Microbiota in Cardiovascular Health and Disease. Circ Res 2017, 120, 1183–1196. [Google Scholar] [CrossRef]
- Du, Y.; Li, X.; Su, C.; Wang, L.; Jiang, J.; Hong, B. The Human Gut Microbiome - a New and Exciting Avenue in Cardiovascular Drug Discovery. Expert Opin Drug Discov 2019, 14, 1037–1052. [Google Scholar] [CrossRef] [PubMed]
- Blacher, E.; Levy, M.; Tatirovsky, E.; Elinav, E. Microbiome-Modulated Metabolites at the Interface of Host Immunity. J Immunol 2017, 198, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Shi, X.; Yang, J.; Zhao, Y.; Xue, L.; Xu, L.; Cai, J. Gut Microbes in Cardiovascular Diseases and Their Potential Therapeutic Applications. Protein Cell 2021, 12, 346–359. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, F.H.; Fåk, F.; Nookaew, I.; Tremaroli, V.; Fagerberg, B.; Petranovic, D.; Bäckhed, F.; Nielsen, J. Symptomatic Atherosclerosis Is Associated with an Altered Gut Metagenome. Nat Commun 2012, 3, 1245. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut Dysbiosis Is Linked to Hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- Berni Canani, R.; Di Costanzo, M.; Leone, L. The Epigenetic Effects of Butyrate: Potential Therapeutic Implications for Clinical Practice. Clin Epigenetics 2012, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.V.; Frassetto, A.; Kowalik, E.J.; Nawrocki, A.R.; Lu, M.M.; Kosinski, J.R.; Hubert, J.A.; Szeto, D.; Yao, X.; Forrest, G.; et al. Butyrate and Propionate Protect against Diet-Induced Obesity and Regulate Gut Hormones via Free Fatty Acid Receptor 3-Independent Mechanisms. PLoS One 2012, 7, e35240. [Google Scholar] [CrossRef] [PubMed]
- Rath, S.; Heidrich, B.; Pieper, D.H.; Vital, M. Uncovering the Trimethylamine-Producing Bacteria of the Human Gut Microbiota. Microbiome 2017, 5, 54. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; DuGar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; Wu, Y. Gut Flora Metabolism of Phosphatidylcholine Promotes Cardiovascular Disease. Nature 2011, 472. [Google Scholar] [CrossRef]
- Bennett, B.J.; de Aguiar Vallim, T.Q.; Wang, Z.; Shih, D.M.; Meng, Y.; Gregory, J.; Allayee, H.; Lee, R.; Graham, M.; Crooke, R.; et al. Trimethylamine-N-Oxide, a Metabolite Associated with Atherosclerosis, Exhibits Complex Genetic and Dietary Regulation. Cell Metab 2013, 17, 49–60. [Google Scholar] [CrossRef]
- Warrier, M.; Shih, D.M.; Burrows, A.C.; Ferguson, D.; Gromovsky, A.D.; Brown, A.L.; Marshall, S.; McDaniel, A.; Schugar, R.C.; Wang, Z.; et al. The TMAO-Generating Enzyme Flavin Monooxygenase 3 Is a Central Regulator of Cholesterol Balance. Cell Rep 2015, 10, 326–338. [Google Scholar] [CrossRef]
- Randrianarisoa, E.; Lehn-Stefan, A.; Wang, X.; Hoene, M.; Peter, A.; Heinzmann, S.S.; Zhao, X.; Königsrainer, I.; Königsrainer, A.; Balletshofer, B.; et al. Relationship of Serum Trimethylamine N-Oxide (TMAO) Levels with Early Atherosclerosis in Humans. Sci Rep 2016, 6, 26745. [Google Scholar] [CrossRef]
- Martin, F.-P.J.; Wang, Y.; Sprenger, N.; Yap, I.K.S.; Lundstedt, T.; Lek, P.; Rezzi, S.; Ramadan, Z.; van Bladeren, P.; Fay, L.B.; et al. Probiotic Modulation of Symbiotic Gut Microbial-Host Metabolic Interactions in a Humanized Microbiome Mouse Model. Mol Syst Biol 2008, 4, 157. [Google Scholar] [CrossRef]
- Bergeron, N.; Williams, P.T.; Lamendella, R.; Faghihnia, N.; Grube, A.; Li, X.; Wang, Z.; Knight, R.; Jansson, J.K.; Hazen, S.L.; Krauss, R.M. Diets High in Resistant Starch Increase Plasma Levels of Trimethylamine-N-Oxide, a Gut Microbiome Metabolite Associated with CVD Risk. The British journal of nutrition 2016, 116. [Google Scholar] [CrossRef] [PubMed]
- Ea, G.; Ja, S. The Human Microbiome: Our Second Genome. Annual review of genomics and human genetics 2012, 13. [Google Scholar] [CrossRef]
- Ve, B.; Ra, G.-R.; Ag, C.; Ns, V.; Bp, Z.; Zj, S.; Jj, R.; Mc, Z.; Ap, N.; Kp, D.; et al. Trimethylamine-N-Oxide Promotes Age-Related Vascular Oxidative Stress and Endothelial Dysfunction in Mice and Healthy Humans. Hypertension (Dallas, Tex. : 1979) 2020, 76. [Google Scholar] [CrossRef]
- Wang, Z.; Tang, W.H.W.; Buffa, J.A.; Fu, X.; Britt, E.B.; Koeth, R.A.; Levison, B.S.; Fan, Y.; Wu, Y.; Hazen, S.L. Prognostic Value of Choline and Betaine Depends on Intestinal Microbiota-Generated Metabolite Trimethylamine-N-Oxide. Eur Heart J 2014, 35, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.W.; Wang, Z.; Shrestha, K.; Borowski, A.G.; Wu, Y.; Troughton, R.W.; Klein, A.L.; Hazen, S.L. Intestinal Microbiota-Dependent Phosphatidylcholine Metabolites, Diastolic Dysfunction, and Adverse Clinical Outcomes in Chronic Systolic Heart Failure. Journal of cardiac failure 2015, 21. [Google Scholar] [CrossRef] [PubMed]
- Li, X.S.; Obeid, S.; Klingenberg, R.; Gencer, B.; Mach, F.; Räber, L.; Windecker, S.; Rodondi, N.; Nanchen, D.; Muller, O.; Miranda, M.X. Gut Microbiota-Dependent Trimethylamine N-Oxide in Acute Coronary Syndromes: A Prognostic Marker for Incident Cardiovascular Events beyond Traditional Risk Factors. European heart journal 2017, 38. [Google Scholar] [CrossRef]
- Tang, W.H.W.; Wang, Z.; Fan, Y.; Levison, B.; Hazen, J.E.; Donahue, L.M.; Wu, Y.; Hazen, S.L. Prognostic Value of Elevated Levels of Intestinal Microbe-Generated Metabolite Trimethylamine-N-Oxide in Patients with Heart Failure: Refining the Gut Hypothesis. J Am Coll Cardiol 2014, 64, 1908–1914. [Google Scholar] [CrossRef]
- Senthong, V.; Wang, Z.; Fan, Y.; Wu, Y.; Hazen, S.L.; Tang, W.H.W. Trimethylamine N-Oxide and Mortality Risk in Patients With Peripheral Artery Disease. J Am Heart Assoc 2016, 5, e004237. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.L.; Martoni, C.J.; Ganopolsky, J.G.; Labbé, A.; Prakash, S. The Human Microbiome and Bile Acid Metabolism: Dysbiosis, Dysmetabolism, Disease and Intervention. Expert Opin Biol Ther 2014, 14, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.L.; Martoni, C.J.; Parent, M.; Prakash, S. Cholesterol-Lowering Efficacy of a Microencapsulated Bile Salt Hydrolase-Active Lactobacillus Reuteri NCIMB 30242 Yoghurt Formulation in Hypercholesterolaemic Adults. Br J Nutr 2012, 107, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Lekawanvijit, S.; Adrahtas, A.; Kelly, D.J.; Kompa, A.R.; Wang, B.H.; Krum, H. Does Indoxyl Sulfate, a Uraemic Toxin, Have Direct Effects on Cardiac Fibroblasts and Myocytes? Eur Heart J 2010, 31, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Nagatomo, Y.; Tang, W.H.W. Intersections Between Microbiome and Heart Failure: Revisiting the Gut Hypothesis. J Card Fail 2015, 21, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Lam, V.; Su, J.; Hsu, A.; Gross, G.J.; Salzman, N.H.; Baker, J.E. Intestinal Microbial Metabolites Are Linked to Severity of Myocardial Infarction in Rats. PLoS One 2016, 11, e0160840. [Google Scholar] [CrossRef] [PubMed]
- Cason, C.A.; Dolan, K.T.; Sharma, G.; Tao, M.; Kulkarni, R.; Helenowski, I.B.; Doane, B.M.; Avram, M.J.; McDermott, M.M.; Chang, E.B.; et al. Plasma Microbiome-Modulated Indole- and Phenyl-Derived Metabolites Associate with Advanced Atherosclerosis and Postoperative Outcomes. J Vasc Surg 2018, 68, 1552–1562. [Google Scholar] [CrossRef]
- Wicherski, J.; Schlesinger, S.; Fischer, F. Association between Breakfast Skipping and Body Weight-A Systematic Review and Meta-Analysis of Observational Longitudinal Studies. Nutrients 2021, 13, 272. [Google Scholar] [CrossRef] [PubMed]
- Effects of Intermittent Fasting on Health, Aging, and Disease. N Engl J Med 2020, 382, 978. [CrossRef] [PubMed]
- Deleu, S.; Machiels, K.; Raes, J.; Verbeke, K.; Vermeire, S. Short Chain Fatty Acids and Its Producing Organisms: An Overlooked Therapy for IBD? EBioMedicine 2021, 66, 103293. [Google Scholar] [CrossRef]
- Sun, J.; Furio, L.; Mecheri, R.; van der Does, A.M.; Lundeberg, E.; Saveanu, L.; Chen, Y.; van Endert, P.; Agerberth, B.; Diana, J. Pancreatic β-Cells Limit Autoimmune Diabetes via an Immunoregulatory Antimicrobial Peptide Expressed under the Influence of the Gut Microbiota. Immunity 2015, 43, 304–317. [Google Scholar] [CrossRef]
- Witkowski, M.; Weeks, T.L.; Hazen, S.L. Gut Microbiota and Cardiovascular Disease. Circ Res 2020, 127, 553–570. [Google Scholar] [CrossRef]
- Barnes, D.; Park, K.T. Donor Considerations in Fecal Microbiota Transplantation. Curr Gastroenterol Rep 2017, 19, 10. [Google Scholar] [CrossRef]
- Fuentes, S.; de Vos, W.M. How to Manipulate the Microbiota: Fecal Microbiota Transplantation. Adv Exp Med Biol 2016, 902, 143–153. [Google Scholar] [CrossRef]
- Gregory, J.C.; Buffa, J.A.; Org, E.; Wang, Z.; Levison, B.S.; Zhu, W.; Wagner, M.A.; Bennett, B.J.; Li, L.; DiDonato, J.A.; et al. Transmission of Atherosclerosis Susceptibility with Gut Microbial Transplantation. J Biol Chem 2015, 290, 5647–5660. [Google Scholar] [CrossRef]
- Drew, L. Microbiota: Reseeding the Gut. Nature 2016, 540, S109–S112. [Google Scholar] [CrossRef]
- Chehoud, C.; Dryga, A.; Hwang, Y.; Nagy-Szakal, D.; Hollister, E.B.; Luna, R.A.; Versalovic, J.; Kellermayer, R.; Bushman, F.D. Transfer of Viral Communities between Human Individuals during Fecal Microbiota Transplantation. mBio 2016, 7, e00322. [Google Scholar] [CrossRef]
- Tamburini, S.; Shen, N.; Wu, H.C.; Clemente, J.C. The Microbiome in Early Life: Implications for Health Outcomes. Nat Med 2016, 22, 713–722. [Google Scholar] [CrossRef]
- Sun, B.; Ma, T.; Li, Y.; Yang, N.; Li, B.; Zhou, X.; Guo, S.; Zhang, S.; Kwok, L.-Y.; Sun, Z.; et al. Bifidobacterium Lactis Probio-M8 Adjuvant Treatment Confers Added Benefits to Patients with Coronary Artery Disease via Target Modulation of the Gut-Heart/-Brain Axes. mSystems 2022, 7, e0010022. [Google Scholar] [CrossRef]
- Hutchison, E.R.; Kasahara, K.; Zhang, Q.; Vivas, E.I.; Cross, T.-W.L.; Rey, F.E. Dissecting the Impact of Dietary Fiber Type on Atherosclerosis in Mice Colonized with Different Gut Microbial Communities. NPJ Biofilms Microbiomes 2023, 9, 31. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Marques, F.Z. How Dietary Fibre, Acting via the Gut Microbiome, Lowers Blood Pressure. Curr Hypertens Rep 2022, 24, 509–521. [Google Scholar] [CrossRef]
- Evans, M.; Dai, L.; Avesani, C.M.; Kublickiene, K.; Stenvinkel, P. The Dietary Source of Trimethylamine N-Oxide and Clinical Outcomes: An Unexpected Liaison. Clin Kidney J 2023, 16, 1804–1812. [Google Scholar] [CrossRef]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.W.M.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of Intestinal Microbiota from Lean Donors Increases Insulin Sensitivity in Individuals with Metabolic Syndrome. Gastroenterology 2012, 143, 913–916. [Google Scholar] [CrossRef]
- Hu, X.-F.; Zhang, W.-Y.; Wen, Q.; Chen, W.-J.; Wang, Z.-M.; Chen, J.; Zhu, F.; Liu, K.; Cheng, L.-X.; Yang, J.; et al. Fecal Microbiota Transplantation Alleviates Myocardial Damage in Myocarditis by Restoring the Microbiota Composition. Pharmacol Res 2019, 139, 412–421. [Google Scholar] [CrossRef]
- Bastos, R.M.C.; Simplício-Filho, A.; Sávio-Silva, C.; Oliveira, L.F.V.; Cruz, G.N.F.; Sousa, E.H.; Noronha, I.L.; Mangueira, C.L.P.; Quaglierini-Ribeiro, H.; Josefi-Rocha, G.R.; et al. Fecal Microbiota Transplant in a Pre-Clinical Model of Type 2 Diabetes Mellitus, Obesity and Diabetic Kidney Disease. Int J Mol Sci 2022, 23, 3842. [Google Scholar] [CrossRef]
- Chen, L.; Guo, L.; Feng, S.; Wang, C.; Cui, Z.; Wang, S.; Lu, Q.; Chang, H.; Hang, B.; Snijders, A.M.; et al. Fecal Microbiota Transplantation Ameliorates Type 2 Diabetes via Metabolic Remodeling of the Gut Microbiota in Db/Db Mice. BMJ Open Diabetes Res Care 2023, 11, e003282. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.D.T.; Kang, J.H.; Lee, M.S. Characterization of Lactobacillus Plantarum PH04, a Potential Probiotic Bacterium with Cholesterol-Lowering Effects. Int J Food Microbiol 2007, 113, 358–361. [Google Scholar] [CrossRef]
- Yadav, H.; Jain, S.; Sinha, P.R. Antidiabetic Effect of Probiotic Dahi Containing Lactobacillus Acidophilus and Lactobacillus Casei in High Fructose Fed Rats. Nutrition 2007, 23, 62–68. [Google Scholar] [CrossRef]
- Asemi, Z.; Zare, Z.; Shakeri, H.; Sabihi, S.-S.; Esmaillzadeh, A. Effect of Multispecies Probiotic Supplements on Metabolic Profiles, Hs-CRP, and Oxidative Stress in Patients with Type 2 Diabetes. Ann Nutr Metab 2013, 63, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Moludi, J.; Kafil, H.S.; Qaisar, S.A.; Gholizadeh, P.; Alizadeh, M.; Vayghyan, H.J. Effect of Probiotic Supplementation along with Calorie Restriction on Metabolic Endotoxemia, and Inflammation Markers in Coronary Artery Disease Patients: A Double Blind Placebo Controlled Randomized Clinical Trial. Nutr J 2021, 20, 47. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhao, D.; Liu, X. Nutritional Supplements Improve Cardiovascular Risk Factors in Overweight and Obese Patients: A Bayesian Network Meta-Analysis. Front Nutr 2023, 10, 1140019. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, J.; Quan, G.; Wang, X.; Yang, L.; Zhong, L. Lactobacillus Acidophilus ATCC 4356 Prevents Atherosclerosis via Inhibition of Intestinal Cholesterol Absorption in Apolipoprotein E-Knockout Mice. Appl Environ Microbiol 2014, 80, 7496–7504. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, S.; Zhao, Z.; Song, X.; Qu, H.; Liu, H. Phenylacetylglutamine Is Associated with the Degree of Coronary Atherosclerotic Severity Assessed by Coronary Computed Tomographic Angiography in Patients with Suspected Coronary Artery Disease. Atherosclerosis 2021, 333, 75–82. [Google Scholar] [CrossRef]
- Di Francesco, A.; Di Germanio, C.; Bernier, M.; de Cabo, R. A Time to Fast. Science 2018, 362, 770–775. [Google Scholar] [CrossRef] [PubMed]
- Regmi, P.; Chaudhary, R.; Page, A.J.; Hutchison, A.T.; Vincent, A.D.; Liu, B.; Heilbronn, L. Early or Delayed Time-Restricted Feeding Prevents Metabolic Impact of Obesity in Mice. J Endocrinol 2021, 248, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.J.; et al. Time-Restricted Feeding without Reducing Caloric Intake Prevents Metabolic Diseases in Mice Fed a High-Fat Diet. Cell Metab 2012, 15, 848–860. [Google Scholar] [CrossRef]
- Takahashi, M.; Ozaki, M.; Kang, M.-I.; Sasaki, H.; Fukazawa, M.; Iwakami, T.; Lim, P.J.; Kim, H.-K.; Aoyama, S.; Shibata, S. Effects of Meal Timing on Postprandial Glucose Metabolism and Blood Metabolites in Healthy Adults. Nutrients 2018, 10, 1763. [Google Scholar] [CrossRef]
Figure 1.
Relationship between Circadian Rhythm Disruption and CMDs.
Figure 2.
Food intake and circadian rhythm.
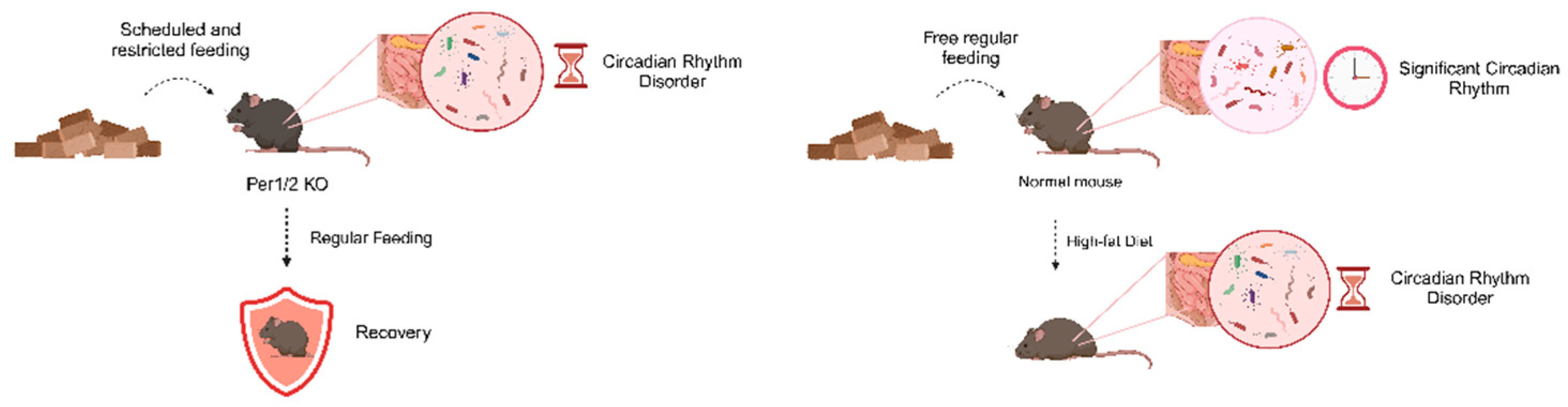
Figure 3.
Circadian Rhythm Disruptions Lead to Gut Microbiota Dysbiosis.
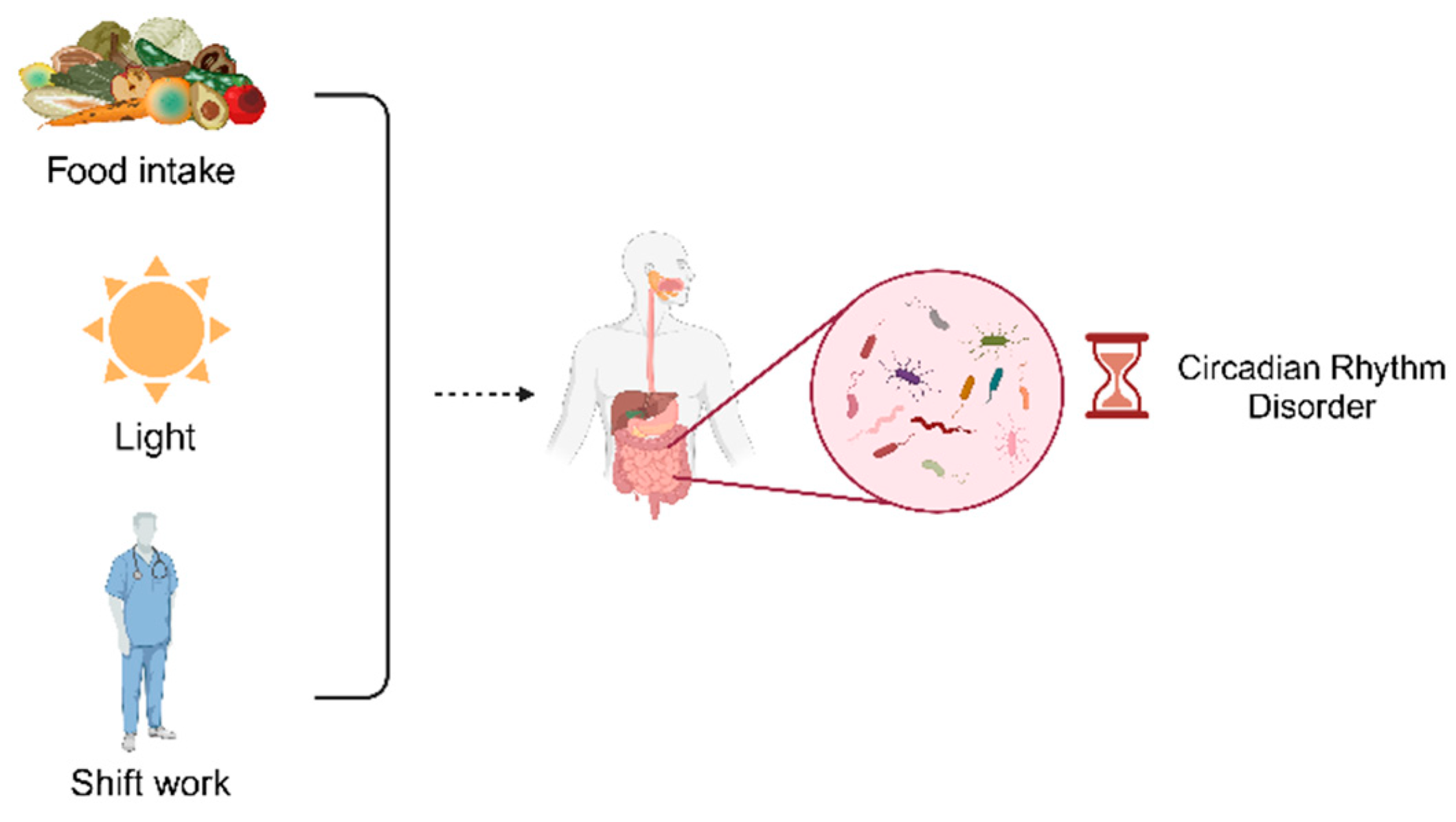

Table 1.
CMDs Treatment Associated with the Gut Microbiota
| Treatment | Species/Strain | Effect | Reference |
|---|---|---|---|
| FMT | Human | Transplantation of feces from lean healthy individuals into patients with metabolic syndrome for 6 weeks resulted in a significant enhancement of peripheral and hepatic insulin sensitivity respectively, and this enhancement was independent of differences in body weight | [163] |
| With regard to FMT in men with metabolic syndrome, shifting the gut microbiota from a lean donor improved insulin sensitivity while increasing butyrate-producing gut bacteria | [163] | ||
| Treatment of Clostridium difficile infection | [153] | ||
| C57BL/6J mouse | FMT restored gut microbial homeostasis in mice and prevented cardiac cell damage in a mouse model of myocarditis | [164] | |
| Administration of FMT in BTBRob/ob mice, a model of diabetic nephropathy, attenuated weight gain, inflammation, and insulin resistance, a finding that was accompanied by an increase in the number of Odoribacteraceae bacteria | [165] | ||
| FMT in diabetic db/db mice increased the abundance of Ruminococcaceae and Porphyromonas, restored the integrity of the intestinal barrier, and ameliorated inflammation | [166] | ||
| Human | Patients consuming Bifidobacterium bifidum and probiotics containing Lactobacillus acidophilus have shown significant reductions in lipid and/or blood glucose levels | [167,168] | |
| Probiotics | improve metabolic status in diabetic patients | [169] | |
| After 12 weeks of supplementation with Lactobacillus rhamnosus and calorie intake restriction in patients with Coronary Artery Disease(CAD), the patients lost significant weight and showed anti-inflammatory effects that were superior to those of calorie intake restriction alone | [170] | ||
| CAD patients treated with the probiotic strain Bifidobacterium lactis Probio-M8 in combination with conventional therapy showed significant improvements in angina, anxiety and depressive symptoms, as well as reductions in interleukin 6 and LDL-C levels, when compared to the control group. | [159] | ||
| Improvement of glycemic status (fasting glucose, fasting insulin levels, insulin resistance) in overweight or obese patients | [171] | ||
| C57BL/6J mouse | Treatment with Lactobacillus acidophilus ATCC 4358 attenuated atherosclerosis in ApoE-/- mice | [172] | |
| Probiotics can effectively reduce the burden of atherosclerotic plaque and have an anti-atherosclerotic effect | [173] | ||
| Rats | Administration of a single cholesterol-lowering probiotic strain to hypercholesterolemic rats affects weight gain, lipid markers, and hepatic steatosis | [30] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



