
Submitted:
16 July 2024
Posted:
17 July 2024
You are already at the latest version
Abstract
Dysbiosis, as reflected by the altered composition of the microbiome favoring pathogenic species, is a major contributor to colorectal cancer (CRC) development, which could lead to bacterial translocation into the bloodstream. This study aimed to evaluate the presence of circulated bacterial DNA (cbDNA) in CRC patients, with or without surgical removal of the primary tumor. In total, 100 participants enrolled in this prospective clinicolaboratory study, equally divided into four groups; healthy controls (Group 1), patients with non-metastatic CRC with surgical removal of the primary tumor (Group 2), patients with metastatic CRC with surgically excised (Group 3) or non-excised (Group 4) primary tumor. DNA extracted from peripheral blood was analyzed using PCR with specific primers targeting 16S rRNA, Escherichia coli (E. coli), and Fusobacterium nucleatum (F. nucleatum). Only the detection of F. nucleatum in the blood was significantly higher in Group 4 compared to Group 1 (p < 0.001), Group 2 (p = 0.023) and Group 3 (p = 0.023). The association of cbDNA with other clinical parameters or co-morbidities was also evaluated. These results highlighted the importance of bacterial translocation in CRC patients and the role of F. nucleatum as an intratumoral oncomicrobe associated with metastatic CRC.
Keywords:
colorectal cancer
; dysbiosis
; bacterial translocation
; surgery
; intestinal microbiota
; circulating bacterial DNA
; metastasis
; Fusobacterium nucleatum
1. Introduction
Colorectal cancer (CRC) is the third most common cancer worldwide, a leading cause of cancer mortality, accounting for 9% of all malignancies in adult patients [1]. Gradual accumulation of genetic mutations results in the formation of pre-cancerous lesions, which subsequently evolve into colorectal adenoma, and ultimately invasive CRC [2]. Immune and inflammatory responses are crucial in all stages of colorectal tumorigenesis [3]. Earlier CRC detection has been made feasible through current innovations in screening endoscopy, imaging modalities, and therapeutic strategies (surgery, radiotherapy, and chemotherapy), leading to enhanced survival rates [4].
Up to 22% of CRC patients are identified with metastatic disease at diagnosis, and various studies indicate that approximately 70% of patients would inevitably exhibit metastatic disease or recurrence, with up to 50% of cases involving synchronous or metachronous distant metastases [5]. Stage IV CRC patients typically have a poor prognosis, with a 14% 5-year survival rate. Hence, the development of efficient prognostic or diagnostic biomarkers is still a necessity.
The correlation between CRC and intestinal microbiota has been extensively evidenced. The diverse microbial ecosystem of the human intestinal microbiota constantly interacts with the host maintaining homeostasis through constant synergistic interactions with the host. However, disruptions in microbiota composition could favor the proliferation of pathogenic bacteria leading to detrimental effects, collectively termed “dysbiosis” [6]. Apart from alterations in microbial composition, dysbiosis also encompasses shifts in bacterial distribution, and dysregulated metabolism promoting colorectal tumorigenesis [7]. Such effects could lead to DNA damage, modulation of immunity, and inflammatory response [8]. Interestingly, surgical stress subsequently to curative resection for CRC affects the host-microbiota interaction, further promoting dysbiosis [9]. Alterations in various bacterial genera, including Bacteroides, Bifidobacterium, Escherichia, Fusobacterium, and Lactobacillus, among others, have been demonstrated after CRC surgery [10]. Fusobacterium nucleatum (F. nucleatum), an invasive, pro-inflammatory pathogen, indigenous to the oral microbiota [11], is one of the most researched bacterial species in CRC. F. nucleatum could trigger carcinogenesis mainly by stimulating the β-catenin cascade following the binding of Fusobacterium adhesion A (FadA) protein to E-cadherin [12]. Studies associate the higher abundance of F. nucleatum with reduced overall or cancer-specific survival in CRC patients [13].
Circulating bacterial DNA (cbDNA) in human blood has become evident [14]. Detection of cbDNA, through various molecular techniques [15,16], has also been reported as a reliable, non-invasive method for CRC screening and prediction of long-term outcomes in CRC patients, possibly participating in CRC pathogenesis [17,18]. However, there is limited data regarding the detection of cbDNA in patients with metastatic CRC, especially concerning any differences regarding the absence of the primary tumor due to previous surgical resection, and focusing on the presence of F. nucleatum.
In this context, the present study aimed to investigate the detection of cbDNA in the blood of patients with CRC using a polymerase chain reaction (PCR)-based method, compare patients with or without surgical resection of the primary tumor, as well as non-metastatic and metastatic disease, and to evaluate any associations with the patients’ demographic and clinical parameters.
2. Results
2.1. Patients’ Characteristics
The epidemiological and clinical characteristics of patients and healthy individuals are summarized in Table 1 and Data S1. Patient groups with CRC were older (mean age range 57.8 – 62.9 years old), and had higher body mass index (BMI) (mean BMI range 27.7 – 28.4 kg/m2) compared to Group 1. Most of the total participants (63%) were males. By design, all patients in Groups 2 and 3 have undergone surgical resection of the primary tumor, whereas all patients in Groups 3 and 4 have metastatic disease (stage IV CRC). The majority of Group 2 patients (80%) had locally advanced (stage III) CRC. In Group 4 patients the tumor location was evenly distributed between the left colon and rectum (40% each), whereas the commonest CRC site regarding non-metastatic groups was the right colon (40% in Group 2 and 3 respectively). Pathologic staging of CRC was mostly intermediate (58.7% of all patients), while tumors presented microsatellite instability (MSI) in only 9.3% of CRC patients. The majority of patients were non-active smokers (64%) and reported no or infrequent alcohol consumption (89.3%) as determined by a negative Alcohol Use Disorders Identification Test-Concise (AUDIT-C) score (< 3 in females and < 4 in males). Regarding co-morbidities, most of the patients had cardiovascular disease (CVD) (45.3%), whereas diabetes and dyslipidemia were more common in Group 3 (44%) and Group 2 (36%), respectively. None of the Group 1 individuals had any co-morbidity.
2.2. Detection of cbDNA
16S rRNA was detected in 84% of Group 1, 96% of Group 2, and 100% of Groups 3 and 4. The β-galactosidase gene of Escherichia coli (E. coli) was detected in 80% of Groups 1 and 2, 84% of Group 3 and 68% of Group 4. Finally, the NusG gene of F. nucleatum was detected in 12% of Group 1, 32% of Groups 2 and 3, and 68% in Group 4 (Table 2 and Figure 1).
Pairwise comparisons between groups revealed significantly higher detection of F. nucleatum in Group 4 compared to Group 1 (p < 0.001, adj. p < 0.001), as well as compared to Group 2 or 3 (p = 0.023, adj. p = 0.045). However, there were no statistically significant differences regarding the detection of 16S rRNA or E. coli among all groups (Table 3).
Further correlation of E. coli and F. nucleatum presence was performed after grouping the patients according to their clinical parameters. The detection of E. coli and F. nucleatum in blood was not significantly associated with differences in sex, metastasis, tumor location, grade, Mismatch Repair (MMR) status, smoking, alcohol consumption, dyslipidemia, or CVD (Table 4). Patients without surgical resection of the primary tumor presented significantly higher detection of F. nucleatum compared to those with surgical resection of the primary tumor (68% vs. 32%, p = 0.006). However, no such difference was observed concerning the detection of E. coli (68% vs. 82%, p = 0.242). Additionally, the detection of E. coli was significantly higher in patients with diabetes compared to those without (100% vs. 69.6%, p = 0.004), whereas such association was not evident with the presence of F. nucleatum (31.6% vs. 48.2%, p = 0.286).
3. Discussion
Lately, research groups have become increasingly interested in peripheral blood as a novel valuable source of cbDNA. PCR-based methods have been developed in earlier studies enabling the detection of cbDNA in CRC patients [19,20]. However, small cohorts were enrolled, and there was no discrimination between CRC patients’ characteristics or proper integration of control subjects. More recent studies relying on advanced PCR-based methods [17,18,21,22] or next-generation sequencing (NGS) methods [23,24] shed light on the elusive subject of cbDNA detection in CRC patients.
In brief, the present study reveals that cbDNA of 16S rRNA, E. coli, and F. nucleatum is present in the blood of healthy subjects and CRC patients. However, the origin of the cbDNA remains elusive. The presence of microbes in the blood may be attributed to occasional dissemination from various body reservoirs into the circulation, known as microbial translocation [25]. The main proposed mechanisms for this phenomenon are intestinal dysbiosis, dysfunction of the intestinal epithelial barrier, and increased permeability (“leaky-gut”). Notably, microbial components, including endotoxins, lipopeptides, and nucleic acids, among others, could also be present in the blood [26]. Additional mechanisms for bacterial translocation into the blood include the interaction of the microbiota with immune system cells, affecting multiple host functions [27], promoting chronic local and systemic inflammation [28], and utilizing dendritic cells or micro-fold cells [29]. Studies have shown that cbDNA was predominantly related to intestinal dysbiosis, although oral or skin microbiota could also serve as potential sources of cbDNA [23]. Studies have revealed great similarity of the F. nucleatum subtypes between saliva and tumor tissue samples in CRC [30], enhancing the hypothesis of orally-mediated intestinal dysbiosis. It is evident that in periodontitis several oral pathogens, including F. nucleatum, are incorporated into complex oral biofilms, facilitating the translocation of oral pathogens into the intestinal ecosystem by invading the bloodstream or swallowing saliva [31]. Residing into the colonic microbiota they further promote dysbiosis. In our cohort, it is reasonable to suggest that the majority of the cbDNA could probably have originated from the intestinal or oral microbial community. Nevertheless, the primary factor, between inflammatory response, alterations in microbiota composition, or increased intestinal permeability, leading to bacterial translocation remains unknown [32,33].
Our study demonstrated a high detection of 16S rRNA (84%) and E. coli (80%) in the blood of healthy subjects, which was non-significantly different compared to CRC patients (96%-100% and 68%-84%, respectively). Similar to our study Giacconi et al. [21], by using real-time qPCR identified the presence of 16S rRNA in all 40 control subjects and 50 CRC patients, although the bacterial load was higher in CRC patients compared to healthy subjects. Mutignani et al. [24], using NGS detected 16S rRNA in all healthy controls and subjects with colorectal adenomas; CRC patients again presented higher cbDNA load. The abundance of E. coli did not differentiate between non-CRC and CRC subjects. Another study by Xiao et al. [23], also using NGS, analyzed the cbDNA between healthy controls, and patients with colorectal adenoma or CRC. A prominent and distinctive circulating cbDNA profile was identified between CRC patients and healthy subjects, highlighting 28 species deriving from intestinal or oral microbiota, which did not include E. coli. Messaritakis et al. [17], however, reported a higher PCR detection of 16S rRNA in a larger cohort of 397 CRC patients (64.5%) compared to 32 healthy controls (15.6%). Notably, although there was no association concerning E. coli detection between these groups (p = 0.186) in accordance with our results, the percentages of positivity were significantly lower compared to our study (15.6% in the control group, and 26.2% in the CRC group).
The biological causes underlying these discrepancies are largely unknown. Concerning 16S rRNA our study demonstrates a potentially substantial impairment of the intestinal permeability in healthy individuals leading to bacterial circulation, in accordance with previous reports [34]. Moreover, E. coli is an almost exclusively nonpathogenic commensal species of the intestinal microbiota, having been detected as a member of the intestinal microbiome of over 90% of healthy individuals [35]. The current literature regarding E. coli and CRC is largely ambiguous regarding its over- or under-representation in CRC-related microbiota compared to controls. This phenomenon is possibly due to the different abundances of the various phylotypes of E. coli (A, B1, B2, D), where E. coli strains belonging to phylotype A are mostly commensal, while strains of B2 phylotype are mainly considered as virulent species [36]. It is difficult to explain the lack of difference of 16S rRNA or E. coli between CRC patients and healthy subjects since inflammatory responses in the CRC microenvironment could also affect bacterial dissemination [37]. This outcome could be partially attributed to a variety in size or shape inclinations in gut-blood bacterial translocation. It should further be emphasized that the present study compared healthy controls with stage IV CRC with or without surgical resection of the primary tumor and stage III CRC with surgical resection of the primary tumor, whereas in the aforementioned studies, the pool of CRC patients included patients with intestinal adenomas or stage I-III CRC without surgical resection of the primary tumor. Hence, our results are not directly comparable to these studies and should be carefully interpreted. The detection of bacterial by-products (metabolites or toxins) in the bloodstream may further aid the differentiation between tumor-free individuals and CRC patients. Future multi-omic studies integrating the analysis of cbDNA with microbiota profiling and metabolome could unveil the molecular mechanism of cbDNA alteration in CRC tumorigenesis.
In this study, we also report a significantly higher detection of F. nucleatum cbDNA in CRC patients (32%-68%) compared to healthy subjects (12%). This is in accordance with the higher identification of F. nucleatum in mucosal and fecal samples from CRC patients promoting intestinal dysbiosis [38,39,40]. To date, only the study by Xiao et al. [23] compared the F. nucleatum in the blood but found no significant difference between CRC patients and controls. However, this difference may be due to the relatively small sample size of their study in addition to the inclusion of only earlier stage CRC (II/III), and the different demographic or environmental variables.
Interestingly, in our setting, the detection of F. nucleatum cbDNA was significantly higher in the stage IV CRC patients without surgical resection of the primary tumor compared to stage II/III CRC or stage IV CRC patients with surgical resection of the primary tumor. This finding is in line with the current evidence that F. nucleatum constitutes a predominantly intratumoral oncomicrobe affecting the tumor microenvironment in promoting CRC pathogenesis [41], and its presence has also been extensively correlated with advanced CRC stages [42]. Nevertheless, it has been demonstrated that the surgical resection of the primary tumor in CRC patients could enhance or reduce the abundance of F. nucleatum in the gut [43,44,45]. Thus, our results indicate that the resection of the primary tumor could disrupt the active oral-gut axis in stage IV CRC reducing the circulation of F. nucleatum.
E. coli cbDNA was not significantly different between CRC patients with or without surgical resection of the primary tumor in our study. Only one study by Koulouridi et al. [18] investigated the detection of E. coli in the blood of stage III CRC patients with surgical resection of the primary tumor, reporting lower frequency (21.5%) in comparison with our results (82%). Studies in the gut microbiota of CRC patients reveal reduced populations of E. coli in cancerous tissue compared to adjacent healthy mucosa [46], while others report inconsistent alterations of E. coli abundance in surgically-treated CRC patients [45,47,48], which could merely explain these results. Research has revealed that in CRC, E. coli could invade the weakened intestinal vascular barrier, and be released into the portal circulation, colonizing the liver and promoting liver metastasis [49]. This fact, in combination with the observation that E. coli is not a strictly intratumoral microbe [47,50], could explain the similarity in the detection of E. coli cbDNA between stage IV CRC patients with or without surgical resection of the primary tumor. Notably, we further observed that neither E. coli nor F. nucleatum cbDNA was significantly altered between non-metastatic (stage II/III) and metastatic (stage IV) CRC patients. F. nucleatum has been thoroughly correlated with increased tumor invasion and lymph node or distant metastases [13,42], and E. coli has also been implicated in metastatic colorectal disease in combination with circulating tumor cells [51]. Similar to our results Giacconi et al. [21] revealed no correlation between increased cbDNA levels and the tumor stage or the presence of distant metastases. This finding might support the concept that cbDNA primarily plays a role in the early development of CRC. The study by Messaritakis et al. [17] reported a significantly higher presence of E. coli in stage IV CRC compared to stage II/III; however, cases with or without surgical resection of the primary tumor were pooled together not allowing an accurate comparison with our data. Our study did not include stage II/III CRC cases without surgical resection of the primary tumor, thus presenting a further limitation in elucidating the aforementioned discrepancies regarding cbDNA association with CRC stage or metastases.
Tumor location was not associated with cbDNA detection in our setting similar to previous PCR-based reports [17]. The utilization of NGS by Mutignani et al. [24] displayed enhanced bacterial transition from the intestinal environment to blood circulation in right colon cancer compared to rectal cancer. Regional discrepancies in genetic expression and immunological characteristics have also been emphasized [52]. Furthermore, intestinal microbiota composition-related tumorigenic mechanisms also vary between the left colon, right colon, or rectum [53]. These data indicate the potential usefulness of sensitive NGS-based methods in discerning bacterial translocation according to the CRC site.
Regarding other demographic or clinical parameters of CRC patients, our study is unique in investigating any possible associations with cbDNA. Gut microbiota studies reveal a distinct microbiota profile in correlation with deficient mismatch repair [54], or high-graded CRC [55]. However, cbDNA detection in our cohorts revealed no significant differences in these characteristics. Several risk factors can contribute to intestinal dysbiosis, such as alcohol consumption [56], smoking, as well as obesity [57]. Studies have also shown the definite presence of cbDNA, in physiological conditions and various systematic diseases, including diabetes, metabolic, or cardiovascular diseases [29,58]. The CRC patients in this study display several clinical characteristics, together with the aforementioned diseases linked to dysbiosis. Between all these factors our study only revealed a strong correlation between higher E. coli cbDNA presence in CRC patients with diabetes. The abundance of E. coli in the gut microbiota of patients with diabetes is increased, serving as an opportunistic pathogen [59]. Nevertheless, the association of cbDNA detection with all the above clinical factors or co-morbidities may be influenced by the impact of the small sample size involved in our research.
Apart from the aforementioned limitations, it should be noted that our study, by design, is a non-randomized prospective study. We were not able to adjust for age between CRC and controls, with the latter belonging to a significantly lower age range, removing potential factors that could influence the results of cbDNA by controlling for these covariates. However, to mitigate the effect of the small sample size the gender and geographical distribution were equivalent between patient groups. The lack of thorough examination of detailed blood microbiota profiling among participants is another limitation, as we used primers targeting specific species. The primary focus of the present study was on the cbDNA, with no investigation conducted on the composition of other microorganisms, such as viruses or fungi, which have been recently discovered to be disrupted in fecal samples of CRC patients. Due to these factors, it is advisable to approach the outcomes cautiously, primarily to generate new hypotheses.
4. Materials and Methods
4.1. Patients Enrollment and Study Design
From September 2021 to April 2024, 75 patients aged > 18 years old with previously diagnosed and histologically confirmed CRC were included in the present, prospective, multi-centered study. For reference, 25 healthy individuals aged > 18 years old were also included, without any benign or malignant neoplastic colorectal disease (Group 1). The patients were equally divided into three groups. Group 2 (n = 25) included patients with non-metastatic CRC who have undergone surgical resection. Group 3 (n = 25) included patients with metastatic CRC who have undergone surgical resection of the primary tumor. Group 4 (n = 25) included patients with metastatic CRC without surgical resection of the primary tumor.
Further exclusion criteria, which applied to all groups included the administration of specific medications (antiemetic, antidiarrheal, antiparasitic, antiviral, antibiotic, probiotic, laxatives, non-steroidal anti-inflammatory, or corticosteroid drugs) within 4 weeks prior to sampling, radiographic studies with barium enema within 1 week prior sampling, active rectal bleeding, individuals with creatinine clearance (measured by Cockcroft-Gault equation) < 30 mL/min, pregnancy, presence of any active bacterial or viral infection, synchronous neoplastic disease elsewhere in the body, genetic disease, and CRC due to familial adenomatous polyposis or Lynch syndrome.
All patients were treated at the University Hospital of Heraklion (Department of Medical Oncology, Department of General Surgery, and Department of Surgical Oncology), at the Venizeleio General Hospital of Heraklion (Department of Oncology, Department of Surgery), and the Creta InterClinic Hospital (Department of Surgery).
4.2. Ethics Approval
The study has been approved by the Research Ethics Committee of Creta InterClinic Hospital (9/7/2021), of the Venizeleio General Hospital of Heraklion (23/14-07-2023), of the University Hospital of Heraklion (23743/02-10-2023), and of the University of Crete (119/24.10.2023). Informed consent was provided to and signed by all participants. The conducted procedures adhered to the ethical criteria set by the aforementioned committees.
4.3. Blood Sampling and DNA Extraction
Peripheral blood (3 mL in EDTA) was collected from all healthy subjects and CRC patients. Samples were collected just before the initiation of any adjuvant or first-line treatment. NucleoSpin© Blood DNA kit (Macherey-Nagel, Düren, Germany) was used for whole-blood genomic DNA extraction, according to the manufacturer’s protocol. DNA quantification was performed by utilizing the NanoDrop ND-1000 v3.3 (Thermo Fisher Scientific, Waltham, MA, USA) spectrophotometer.
4.4. PCR Amplification of Microbial DNA
Two oligonucleotide primer pairs were used to detect cbDNA via PCR; the β-galactosidase gene of E. coli [17,18], and the NusG gene of F. nucleatum [60]. DNA integrity of the samples was performed by using glyceraldehyde phospho-dehydrogenase (GAPDH) as reference primers [17,18]. For reference marker in the detection of cbDNA, 16S rRNA primers were also used in accordance with previous studies [17,18]. The PCR conditions, primer sequences, and fragment size of amplicons for each target gene are reported in Table S1.
4.5. Statistical Analysis
Descriptive and inferential statistics were performed regarding the patients’ and healthy individuals’ baseline variables (demographic characteristics, clinical parameters). Comparisons between categorical variables were calculated with Pearson’s Chi-square test. Adjusted (adj.) p values through Bonferroni correction are also provided in cases of multiple comparisons. Statistical analysis was performed via SPSS v. 26 (IBM Corp. Armonk, New York, USA). Statistical significance was indicated at the conventional p < 0.05 threshold.
5. Conclusions
In conclusion, our study confirms the presence of cbDNA in healthy individuals and patients with CRC. High presence of 16S rRNA and E. coli was revealed in all participants, providing strong evidence for bacterial translocation. Moreover, the significantly higher detection of F. nucleatum in the blood of patients with metastatic CRC without surgical resection of the primary tumor, confirms the role of this bacterium as an intratumoral pathogen associated with advanced stages of CRC. Future studies should focus on correlating these findings with patients’ outcomes and survival, possibly elucidating the role of F. nucleatum as a prognostic biomarker in metastatic CRC. For the further investigation of the sources of cbDNA, fecal and oral samples should be analyzed in the future, along with blood microbiota, using more advanced NGS-based methods, assessing the relationship between CRC and microbiota dysbiosis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Data S1: Raw data material; Table S1: PCR primers and conditions for DNA amplification.
Author Contributions
Conceptualization, J.T.; methodology, I.K., I.L., K.V., E.I., I.M. and J.T.; software, I.L.; validation, I.K. and I.L.; formal analysis, I.K., I.L. and E.I.; investigation, K.V. and I.T.; resources, K.V. and I.T.; data curation, I.K., I.L. and I.M.; writing—original draft preparation, I.K.; writing—review and editing, I.M., J.S. and J.T.; visualization, I.K. and K.V.; supervision, J.T.; project administration, J.S. and J.T.; funding acquisition, J.S. and J.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by GastoIntestinal Cancer Study Group (GIC-SG).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Research Ethics Committee of Creta InterClinic Hospital (9/7/2021), of the Venizeleio General Hospital of Heraklion (23/14-07-2023), of the University Hospital of Heraklion (23743/02-10-2023), and of the University of Crete (119/24.10.2023).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
All data used for the analysis of the current study are provided as Supplementary Data S1.
Acknowledgments
The authors acknowledge the contribution of Andreas Strehle and Michail I Giakoumakis from the General Surgery Department of the Venizeleio General Hospital of Heraklion in the collection of the patients’ samples.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Vogelstein, B.; Papadopoulos, N.; Velculescu, V.E.; Zhou, S.; Diaz, L.A.; Kinzler, K.W. Cancer Genome Landscapes. Science 2013, 339, 1546–1558. [Google Scholar] [CrossRef] [PubMed]
- Terzić, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and Colon Cancer. Gastroenterology 2010, 138, 2101–2114.e5. [Google Scholar] [CrossRef] [PubMed]
- Compton, C.; Fenoglio-Preiser, C.M.; Pettigrew, N.; Fielding, L.P. American Joint Committee on Cancer Prognostic Factors Consensus Conference: Colorectal Working Group. Cancer 2000, 88, 1739–1757. [Google Scholar] [CrossRef]
- Ilyas, M.I.M. Epidemiology of Stage IV Colorectal Cancer: Trends in the Incidence, Prevalence, Age Distribution, and Impact on Life Span. Clin Colon Rectal Surg 2023, 37, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Bien, J.; Palagani, V.; Bozko, P. The Intestinal Microbiota Dysbiosis and Clostridium Difficile Infection: Is There a Relationship with Inflammatory Bowel Disease? Therap Adv Gastroenterol 2013, 6, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Abreu, M.T.; Peek, R.M. Gastrointestinal Malignancy and the Microbiome. Gastroenterology 2014, 146, 1534–1546.e3. [Google Scholar] [CrossRef] [PubMed]
- Irrazábal, T.; Belcheva, A.; Girardin, S.E.; Martin, A.; Philpott, D.J. The Multifaceted Role of the Intestinal Microbiota in Colon Cancer. Mol Cell 2014, 54, 309–320. [Google Scholar] [CrossRef]
- Shimizu, K.; Ogura, H.; Goto, M.; Asahara, T.; Nomoto, K.; Morotomi, M.; Yoshiya, K.; Matsushima, A.; Sumi, Y.; Kuwagata, Y.; et al. Altered Gut Flora and Environment in Patients with Severe SIRS. J Trauma 2006, 60, 126–133. [Google Scholar] [CrossRef]
- Koliarakis, I.; Athanasakis, E.; Sgantzos, M.; Mariolis-Sapsakos, T.; Xynos, E.; Chrysos, E.; Souglakos, J.; Tsiaoussis, J. Intestinal Microbiota in Colorectal Cancer Surgery. Cancers (Basel) 2020, 12, 3011. [Google Scholar] [CrossRef]
- Signat, B.; Roques, C.; Poulet, P.; Duffaut, D. Fusobacterium Nucleatum in Periodontal Health and Disease. Curr Issues Mol Biol 2011, 13, 25–36. [Google Scholar]
- Shang, F.-M.; Liu, H.-L. Fusobacterium Nucleatum and Colorectal Cancer: A Review. World J Gastrointest Oncol 2018, 10, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Mima, K.; Nishihara, R.; Qian, Z.R.; Cao, Y.; Sukawa, Y.; Nowak, J.A.; Yang, J.; Dou, R.; Masugi, Y.; Song, M.; et al. Fusobacterium Nucleatum in Colorectal Carcinoma Tissue and Patient Prognosis. Gut 2016, 65, 1973–1980. [Google Scholar] [CrossRef]
- Lelouvier, B.; Servant, F.; Païssé, S.; Brunet, A.-C.; Benyahya, S.; Serino, M.; Valle, C.; Ortiz, M.R.; Puig, J.; Courtney, M.; et al. Changes in Blood Microbiota Profiles Associated with Liver Fibrosis in Obese Patients: A Pilot Analysis. Hepatology 2016, 64, 2015–2027. [Google Scholar] [CrossRef]
- Bruns, T.; Reuken, P.A.; Stengel, S.; Gerber, L.; Appenrodt, B.; Schade, J.H.; Lammert, F.; Zeuzem, S.; Stallmach, A. The Prognostic Significance of Bacterial DNA in Patients with Decompensated Cirrhosis and Suspected Infection. Liver Int 2016, 36, 1133–1142. [Google Scholar] [CrossRef]
- Merlini, E.; Bellistri, G.M.; Tincati, C.; d’Arminio Monforte, A.; Marchetti, G. Sequencing of Bacterial Microflora in Peripheral Blood: Our Experience with HIV-Infected Patients. J Vis Exp 2011, 2830. [Google Scholar] [CrossRef]
- Messaritakis, I.; Vogiatzoglou, K.; Tsantaki, K.; Ntretaki, A.; Sfakianaki, M.; Koulouridi, A.; Tsiaoussis, J.; Mavroudis, D.; Souglakos, J. The Prognostic Value of the Detection of Microbial Translocation in the Blood of Colorectal Cancer Patients. Cancers (Basel) 2020, 12, 1058. [Google Scholar] [CrossRef]
- Koulouridi, A.; Messaritakis, I.; Theodorakis, E.; Chondrozoumaki, M.; Sfakianaki, M.; Gouvas, N.; Tsiaoussis, J.; Mavroudis, D.; Tzardi, M.; Souglakos, J. Detection of Circulating Tumor Cells and Microbial DNA Fragments in Stage III Colorectal Cancer Patients under Three versus Six Months of Adjuvant Treatment. Cancers (Basel) 2021, 13, 3552. [Google Scholar] [CrossRef] [PubMed]
- Lescut, D.; Colombel, J.F.; Vincent, P.; Cortot, A.; Fournier, L.; Quandalle, P.; Vankemmel, M.; Triboulet, J.P.; Wurtz, A.; Paris, J.C. Bacterial Translocation in Colorectal Cancers. Gastroenterol Clin Biol 1990, 14, 811–814. [Google Scholar]
- Ono, S.; Tsujimoto, H.; Yamauchi, A.; Hiraki, S.; Takayama, E.; Mochizuki, H. Detection of Microbial DNA in the Blood of Surgical Patients for Diagnosing Bacterial Translocation. World J Surg 2005, 29, 535–539. [Google Scholar] [CrossRef]
- Giacconi, R.; Donghia, R.; Arborea, G.; Savino, M.T.; Provinciali, M.; Lattanzio, F.; Caponio, G.R.; Coletta, S.; Bianco, A.; Notarnicola, M.; et al. Plasma Bacterial DNA Load as a Potential Biomarker for the Early Detection of Colorectal Cancer: A Case-Control Study. Microorganisms 2023, 11, 2360. [Google Scholar] [CrossRef]
- Messaritakis, I.; Koulouris, A.; Boukla, E.; Vogiatzoglou, K.; Lagkouvardos, I.; Intze, E.; Sfakianaki, M.; Chondrozoumaki, M.; Karagianni, M.; Athanasakis, E.; et al. Exploring Gut Microbiome Composition and Circulating Microbial DNA Fragments in Patients with Stage II/III Colorectal Cancer: A Comprehensive Analysis. Cancers (Basel) 2024, 16, 1923. [Google Scholar] [CrossRef]
- Xiao, Q.; Lu, W.; Kong, X.; Shao, Y.W.; Hu, Y.; Wang, A.; Bao, H.; Cao, R.; Liu, K.; Wang, X.; et al. Alterations of Circulating Bacterial DNA in Colorectal Cancer and Adenoma: A Proof-of-Concept Study. Cancer Lett 2021, 499, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Mutignani, M.; Penagini, R.; Gargari, G.; Guglielmetti, S.; Cintolo, M.; Airoldi, A.; Leone, P.; Carnevali, P.; Ciafardini, C.; Petrocelli, G.; et al. Blood Bacterial DNA Load and Profiling Differ in Colorectal Cancer Patients Compared to Tumor-Free Controls. Cancers (Basel) 2021, 13, 6363. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.C.S.; Ko, K.K.K.; Chen, H.; Liu, J.; Loh, M.; SG10K_Health Consortium; Chia, M; Nagarajan, N. No Evidence for a Common Blood Microbiome Based on a Population Study of 9,770 Healthy Humans. Nat Microbiol 2023, 8, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, V.; Moreau, R.; Kamath, P.S.; Jalan, R.; Ginès, P.; Nevens, F.; Fernández, J.; To, U.; García-Tsao, G.; Schnabl, B. Acute-on-Chronic Liver Failure in Cirrhosis. Nat Rev Dis Primers 2016, 2, 16041. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between Microbiota and Immunity in Health and Disease. Cell Res 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Kouzu, K.; Tsujimoto, H.; Kishi, Y.; Ueno, H.; Shinomiya, N. Bacterial Translocation in Gastrointestinal Cancers and Cancer Treatment. Biomedicines 2022, 10, 380. [Google Scholar] [CrossRef]
- Potgieter, M.; Bester, J.; Kell, D.B.; Pretorius, E. The Dormant Blood Microbiome in Chronic, Inflammatory Diseases. FEMS Microbiol Rev 2015, 39, 567–591. [Google Scholar] [CrossRef]
- Komiya, Y.; Shimomura, Y.; Higurashi, T.; Sugi, Y.; Arimoto, J.; Umezawa, S.; Uchiyama, S.; Matsumoto, M.; Nakajima, A. Patients with Colorectal Cancer Have Identical Strains of Fusobacterium Nucleatum in Their Colorectal Cancer and Oral Cavity. Gut 2019, 68, 1335–1337. [Google Scholar] [CrossRef] [PubMed]
- Koliarakis, I.; Messaritakis, I.; Nikolouzakis, T.K.; Hamilos, G.; Souglakos, J.; Tsiaoussis, J. Oral Bacteria and Intestinal Dysbiosis in Colorectal Cancer. Int J Mol Sci 2019, 20, 4146. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms Regulating Intestinal Barrier Integrity and Its Pathological Implications. Exp Mol Med 2018, 50, 1–9. [Google Scholar] [CrossRef]
- Wasinger, V.C.; Lu, K.; Yau, Y.Y.; Nash, J.; Lee, J.; Chang, J.; Paramsothy, S.; Kaakoush, N.O.; Mitchell, H.M.; Leong, R.W.L. Spp24 Is Associated with Endocytic Signalling, Lipid Metabolism, and Discrimination of Tissue Integrity for “leaky-Gut” in Inflammatory Bowel Disease. Sci Rep 2020, 10, 12932. [Google Scholar] [CrossRef]
- Païssé, S.; Valle, C.; Servant, F.; Courtney, M.; Burcelin, R.; Amar, J.; Lelouvier, B. Comprehensive Description of Blood Microbiome from Healthy Donors Assessed by 16S Targeted Metagenomic Sequencing. Transfusion 2016, 56, 1138–1147. [Google Scholar] [CrossRef] [PubMed]
- Martinson, J.N.V.; Walk, S.T. Escherichia Coli Residency in the Gut of Healthy Human Adults. EcoSal Plus 2020, 9, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Wassenaar, T.M. E. Coli and Colorectal Cancer: A Complex Relationship That Deserves a Critical Mindset. Crit Rev Microbiol 2018, 44, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Genua, F.; Raghunathan, V.; Jenab, M.; Gallagher, W.M.; Hughes, D.J. The Role of Gut Barrier Dysfunction and Microbiome Dysbiosis in Colorectal Cancer Development. Front Oncol 2021, 11, 626349. [Google Scholar] [CrossRef]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium Nucleatum Infection Is Prevalent in Human Colorectal Carcinoma. Genome Res 2012, 22, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Duke, F.; Earl, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; et al. Genomic Analysis Identifies Association of Fusobacterium with Colorectal Carcinoma. Genome Res 2012, 22, 292–298. [Google Scholar] [CrossRef]
- Flanagan, L.; Schmid, J.; Ebert, M.; Soucek, P.; Kunicka, T.; Liska, V.; Bruha, J.; Neary, P.; Dezeeuw, N.; Tommasino, M.; et al. Fusobacterium Nucleatum Associates with Stages of Colorectal Neoplasia Development, Colorectal Cancer and Disease Outcome. Eur J Clin Microbiol Infect Dis 2014, 33, 1381–1390. [Google Scholar] [CrossRef]
- Joo, J.E.; Chu, Y.L.; Georgeson, P.; Walker, R.; Mahmood, K.; Clendenning, M.; Meyers, A.L.; Como, J.; Joseland, S.; Preston, S.G.; et al. Intratumoral Presence of the Genotoxic Gut Bacteria Pks+ E. Coli, Enterotoxigenic Bacteroides Fragilis, and Fusobacterium Nucleatum and Their Association with Clinicopathological and Molecular Features of Colorectal Cancer. Br J Cancer 2024, 130, 728–740. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Liu, L.; Li, H.; Qin, H.; Sun, Z. Clinical Significance of Fusobacterium Nucleatum, Epithelial-Mesenchymal Transition, and Cancer Stem Cell Markers in Stage III/IV Colorectal Cancer Patients. Onco Targets Ther 2017, 10, 5031–5046. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Coker, O.O.; Nakatsu, G.; Wu, W.K.K.; Zhao, L.; Chen, Z.; Chan, F.K.L.; Kristiansen, K.; Sung, J.J.Y.; Wong, S.H.; et al. Multi-Cohort Analysis of Colorectal Cancer Metagenome Identified Altered Bacteria across Populations and Universal Bacterial Markers. Microbiome 2018, 6, 70. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.-H.; Jiang, J.-K.; Luo, J.-C.; Lin, C.-C.; Ting, P.-H.; Yang, U.-C.; Lan, Y.-T.; Huang, Y.-H.; Hou, M.-C.; Lee, F.-Y. The Long Term Microbiota and Metabolic Status in Patients with Colorectal Cancer after Curative Colon Surgery. PLoS One 2019, 14, e0218436. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.; Gao, R.; Yan, X.; Huang, L.; He, J.; Li, H.; You, J.; Qin, H. Alterations in Intestinal Microbiota of Colorectal Cancer Patients Receiving Radical Surgery Combined with Adjuvant CapeOx Therapy. Sci China Life Sci 2019, 62, 1178–1193. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Guo, B.; Gao, R.; Zhu, Q.; Qin, H. Microbiota Disbiosis Is Associated with Colorectal Cancer. Front Microbiol 2015, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-J.; Zhang, Y.-L.; Shang, Y.; Wu, B.; Yang, E.; Luo, Y.-Y.; Li, X.-R. Intestinal Bacteria Detected in Cancer and Adjacent Tissue from Patients with Colorectal Cancer. Oncol Lett 2019, 17, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Cai, G.; Qiu, Y.; Fei, N.; Zhang, M.; Pang, X.; Jia, W.; Cai, S.; Zhao, L. Structural Segregation of Gut Microbiota between Colorectal Cancer Patients and Healthy Volunteers. ISME J 2012, 6, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Cass, S.; White, M.G. The Influence of the Microbiome on Metastatic Colorectal Cancer. Clin Colon Rectal Surg 2023, 36, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, P.; Wang, J.; Wei, X.; Wang, M. Unveiling Intratumoral Microbiota: An Emerging Force for Colorectal Cancer Diagnosis and Therapy. Pharmacological Research 2024, 203, 107185. [Google Scholar] [CrossRef]
- Arthur, J.C.; Perez-Chanona, E.; Mühlbauer, M.; Tomkovich, S.; Uronis, J.M.; Fan, T.-J.; Campbell, B.J.; Abujamel, T.; Dogan, B.; Rogers, A.B.; et al. Intestinal Inflammation Targets Cancer-Inducing Activity of the Microbiota. Science 2012, 338, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Glebov, O.K.; Rodriguez, L.M.; Nakahara, K.; Jenkins, J.; Cliatt, J.; Humbyrd, C.-J.; DeNobile, J.; Soballe, P.; Simon, R.; Wright, G.; et al. Distinguishing Right from Left Colon by the Pattern of Gene Expression. Cancer Epidemiol Biomarkers Prev 2003, 12, 755–762. [Google Scholar]
- Tamas, K.; Walenkamp, A.M.E.; de Vries, E.G.E.; van Vugt, M. a. T.M.; Beets-Tan, R.G.; van Etten, B.; de Groot, D.J.A.; Hospers, G. a. P. Rectal and Colon Cancer: Not Just a Different Anatomic Site. Cancer Treat Rev 2015, 41, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Wu, J.; Shi, L.; Zhou, B.; Shang, F.; Chang, X.; Dong, X.; Deng, S.; Liu, L.; Cai, K.; et al. Gut Microbiota Distinct between Colorectal Cancers with Deficient and Proficient Mismatch Repair: A Study of 230 CRC Patients. Front Microbiol 2022, 13, 993285. [Google Scholar] [CrossRef]
- Qi, Z.; Zhibo, Z.; Jing, Z.; Zhanbo, Q.; Shugao, H.; Weili, J.; Jiang, L.; Shuwen, H. Prediction Model of Poorly Differentiated Colorectal Cancer (CRC) Based on Gut Bacteria. BMC Microbiol 2022, 22, 312. [Google Scholar] [CrossRef] [PubMed]
- Carelli, L.L.; D’Aquila, P.; Rango, F.D.; Incorvaia, A.; Sena, G.; Passarino, G.; Bellizzi, D. Modulation of Gut Microbiota through Low-Calorie and Two-Phase Diets in Obese Individuals. Nutrients 2023, 15, 1841. [Google Scholar] [CrossRef]
- Wilkins, L.J.; Monga, M.; Miller, A.W. Defining Dysbiosis for a Cluster of Chronic Diseases. Sci Rep 2019, 9, 12918. [Google Scholar] [CrossRef]
- Amar, J.; Lelouvier, B.; Servant, F.; Lluch, J.; Burcelin, R.; Bongard, V.; Elbaz, M. Blood Microbiota Modification After Myocardial Infarction Depends Upon Low-Density Lipoprotein Cholesterol Levels. J Am Heart Assoc 2019, 8, e011797. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chu, J.; Hao, W.; Zhang, J.; Li, H.; Yang, C.; Yang, J.; Chen, X.; Wang, H. Gut Microbiota and Type 2 Diabetes Mellitus: Association, Mechanism, and Translational Applications. Mediators Inflamm 2021, 2021, 5110276. [Google Scholar] [CrossRef] [PubMed]
- Proença, M.A.; Biselli, J.M.; Succi, M.; Severino, F.E.; Berardinelli, G.N.; Caetano, A.; Reis, R.M.; Hughes, D.J.; Silva, A.E. Relationship between Fusobacterium Nucleatum, Inflammatory Mediators and microRNAs in Colorectal Carcinogenesis. World J Gastroenterol 2018, 24, 5351–5365. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Frequencies of 16S rRNA, Escherichia coli (E. coli), and Fusobacterium nucleatum (F. nucleatum) detection in blood among control and patient groups.
Figure 1.
Frequencies of 16S rRNA, Escherichia coli (E. coli), and Fusobacterium nucleatum (F. nucleatum) detection in blood among control and patient groups.
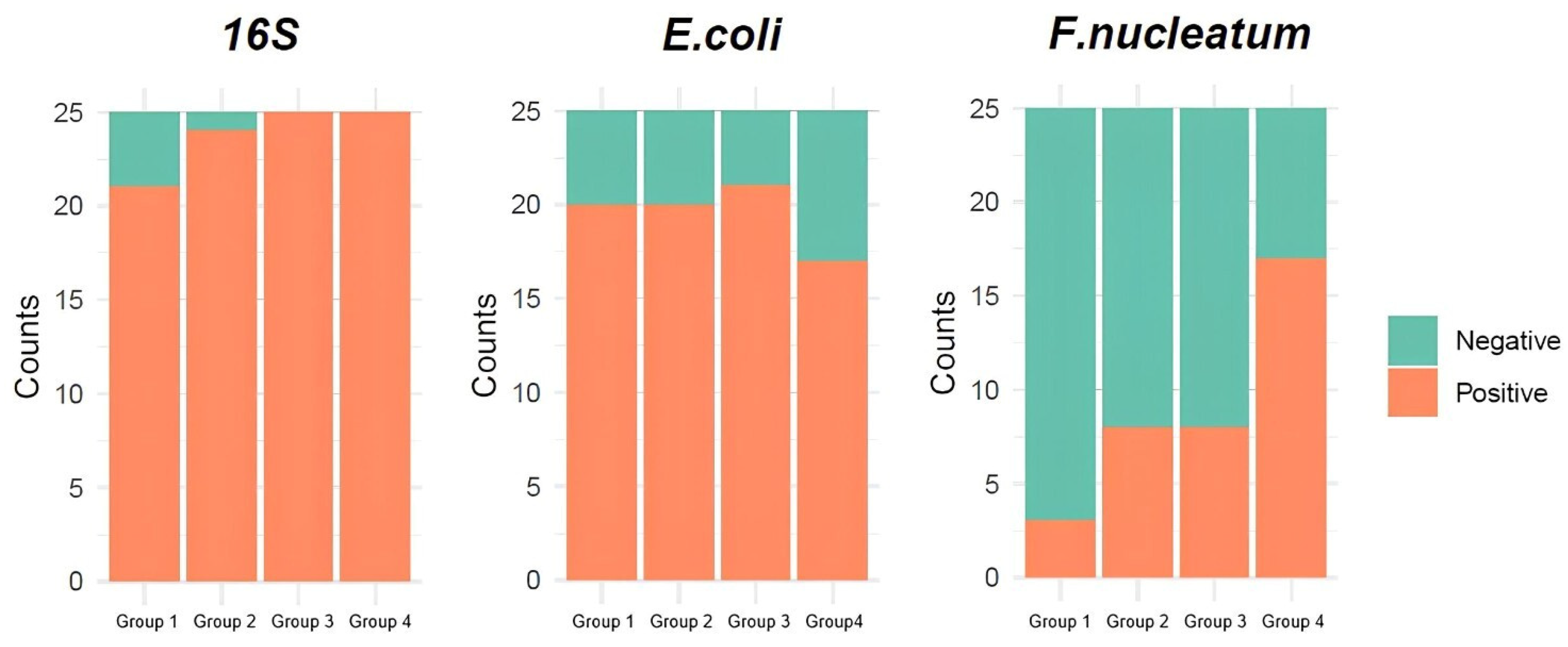

Table 1.
Patients’ and healthy individuals’ demographic characteristics and clinical parameters.
| Characteristics | Group 1 (n = 25) | Group 2 (n = 25) | Group 3 (n = 25) | Group 4 (n = 25) |
|---|---|---|---|---|
| Mean (range) | ||||
| Age | 20.5 (20-21) | 61.5 (25-86) | 62.9 (39-80) | 57.8 (26-78) |
| BMI (kg/m2) | 23.1 (20.3-24.8) | 28.4 (22.1-46.9) | 27.7 (20.2-37.2) | 27.9 (17.1-52.5) |
| Frequency (percentage) | ||||
| Sex | ||||
| Male | 18 (72%) | 11 (44%) | 18 (72%) | 16 (64%) |
| Female | 7 (28%) | 14 (56%) | 7 (28%) | 9 (36%) |
| Surgery | ||||
| Yes | n/a * | 25 (100%) | 25 (100%) | 0 (0%) |
| No | n/a | 0 (0%) | 0 (0%) | 25 (100%) |
| Metastasis | ||||
| Yes | n/a | 0 (0%) | 25 (100%) | 25 (100%) |
| No | n/a | 25 (100%) | 0 (0%) | 0 (0%) |
| Tumor Location | ||||
| Right Colon | n/a | 10 (40%) | 10 (40%) | 5 (20%) |
| Left Colon | n/a | 9 (36%) | 8 (32%) | 10 (40%) |
| Rectum | n/a | 6 (24%) | 7 (28%) | 10 (40%) |
| Stage | ||||
| II | n/a | 5 (20%) | 0 (0%) | 0 (0%) |
| III | n/a | 20 (80%) | 0 (0 %) | 0 (0%) |
| IV | n/a | 0 (0%) | 25 (100%) | 25 (100%) |
| Grade | ||||
| Low | n/a | 6 (24%) | 2 (8%) | 2 (8%) |
| Intermediate | n/a | 13 (52%) | 15 (60%) | 16 (64%) |
| High | n/a | 6 (24%) | 8 (32%) | 7 (28%) |
| Mismatch Repair (MMR) status | ||||
| Microsatellite Stability (MSS) | n/a | 19 (76%) | 23 (92%) | 21 (84%) |
| Microsatellite Instability (MSI) | n/a | 3 (12%) | 2 (8%) | 2 (8%) |
| Unreported | n/a | 3 (12%) | 0 (0%) | 2 (8%) |
| Smoking | ||||
| Yes | 0 (0%) | 7 (28%) | 12 (48%) | 8 (32%) |
| No | 25 (100%) | 18 (72%) | 13 (52%) | 17 (68%) |
| Alcohol | ||||
| Yes | 0 (0%) | 1 (4%) | 3 (12%) | 4 (16%) |
| No | 25 (100%) | 24 (96%) | 22 (88%) | 21 (84%) |
| Diabetes | ||||
| Yes | 0 (0%) | 5 (20%) | 11 (44%) | 3 (12%) |
| No | 25 (100%) | 20 (80%) | 14 (56%) | 22 (88%) |
| Dyslipidemia | ||||
| Yes | 0 (0%) | 9 (36%) | 5 (20%) | 6 (24%) |
| No | 25 (100%) | 16 (64%) | 20 (80%) | 19 (76%) |
| Cardiovascular Disease (CVD) | ||||
| Yes | 0 (0%) | 12 (48%) | 13 (52%) | 9 (36%) |
| No | 25 (100%) | 13 (52%) | 12 (48%) | 16 (64%) |
* n/a, not applicable.
Table 2.
Frequency and percentages of circulating bacterial DNA (cbDNA) presence among control and patient groups.
Table 2.
Frequency and percentages of circulating bacterial DNA (cbDNA) presence among control and patient groups.
| Gene Target | Detection | Group 1 (n = 25) |
Group 2 (n = 25) |
Group3 (n = 25) |
Group 4 (n = 25) |
|---|---|---|---|---|---|
| 16S rRNA | Positive | 21 (84%) | 24 (96%) | 25 (100%) | 25 (100%) |
| Negative | 4 (16%) | 1 (4%) | 0 (0%) | 0 (0%) | |
| β-galactosidase gene of Escherichia coli | Positive | 20 (80%) | 20 (80%) | 21 (84%) | 17 (68%) |
| Negative | 5 (20%) | 5 (20%) | 4 (16%) | 8 (32%) | |
| NusG gene of Fusobacterium nucleatum | Positive | 3 (12%) | 8 (32%) | 8 (32%) | 17 (68%) |
| Negative | 22 (88%) | 17 (68%) | 17 (68%) | 8 (32%) |
Table 3.
Comparisons of cbDNA detection among control and patient groups.
| Pairwise comparison | 16S rRNA | E. coli | F. nucleatum | |||
|---|---|---|---|---|---|---|
| p-value | adj. p-value | p-value | adj. p-value | p-value | adj. p-value | |
| Group 1 – Group 2 | 0.349 | 0.697 | - * | - | 0.171 | 0.205 |
| Group 1 – Group 3 | 0.11 | 0.33 | - | - | 0.171 | 0.205 |
| Group 1 – Group 4 | 0.11 | 0.33 | 0.52 | - | <0.001 | <0.001 |
| Group 2 – Group 3 | - | - | - | - | - | - |
| Group 2 – Group 4 | - | - | 0.52 | - | 0.023 | 0.045 |
| Group 3 – Group 4 | - | - | 0.321 | - | 0.023 | 0.045 |
* p = 1.
Table 4.
Association of cbDNA detection with patients’ clinical parameters.
| Parameter / Pairwise comparison | E. coli | F. nucleatum | ||
|---|---|---|---|---|
| p-value | adj. p-value | p-value | adj. p-value | |
| Sex | ||||
| Female – Male | 0.403 | n/a 1 | 0.348 | n/a |
| Surgery | ||||
| No – Yes | 0.242 | n/a | 0.006 | n/a |
| Metastasis | ||||
| No (stage II/III) – Yes (stage IV) | 0.777 | n/a | 0.217 | n/a |
| Tumor Location | ||||
| Left Colon – Rectum | 0.308 | 0.462 | 0.252 | 0.596 |
| Right Colon – Rectum | 0.292 | 0.462 | - 2 | - |
| Left Colon – Right Colon | - | - | 0.397 | 0.596 |
| Grade | ||||
| High – Low | 0.634 | 0.951 | 0.701 | - |
| Intermediate – Low | 0.427 | 0.951 | - | - |
| High – Intermediate | - | - | 0.605 | - |
| MMR status | ||||
| MSI – MSS | - | n/a | - | n/a |
| Smoking | ||||
| No – Yes | 0.15 | n/a | 0.634 | n/a |
| Alcohol | ||||
| No – Yes | - | n/a | 0.725 | n/a |
| Diabetes | ||||
| No – Yes | 0.004 | n/a | 0.286 | n/a |
| Dyslipidemia | ||||
| No – Yes | - | n/a | - | n/a |
| CVD | ||||
| No – Yes | 0.414 | n/a | 0.816 | n/a |
1 n/a, not applicable; 2 p = 1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



